~ """""""~""~"~-"~."'1f~;~"",~;:~".."", "'~:,-"-'W"~"1"~-~~~--"'" ~~~~'.!!"~~~W"" :~. E oorin1er1taJ ~ Researd1 @ Springer-Verlag 1982 Exp Brain Res (1982) 47: 417-427 Saccadic Responses Evoked by Presentationor Visual and Auditory Targets* D. ZambarbieriJ, R. Schmid1, G. Magenesl, and C. Prablan~ l Istituto di Infonnatica e Sistemistica, Università di Pavia, Strada Nuova106 C, 1-27100 Pavia, Italy 2 Laboratoire de Neuropsychologie Expérimentale, Unité 94 - INSERM, 16, Avenue du Doyen Lépine, F-69500 Bron, France Summary. Saccadiceye movements evoked by the presentation' of visual and auditory targets were examined arid compared. Differences were found either in ~he pattem of the saccadic responseand in the characteristics of single saccadesof the same amplitude. The longer latency and the higher percentage of multiple saccade responses in the auditory case were attributed to a more complex centrai processing,whereas the longer duration and the lower peak velocity of the saccades to auditory targets were attributed to reduced performances of the execution mechanism in the absence of vision. Key words: Saccades- Audio-ocular responses- Fixation - Sound localization Introduction Saccades afe fi.xation ere movements which need a position reference signal to be made available within the CNS. A reference signal can be reconstructed from sensory information about extemal targets or can be intemal1ygenerated without sensoryafference as far voluntary saccades in darkness. In the caseof saccades evoked by target presentation three differ- ent processes should be accomplished:acquisition of sensoryinformation, centraI reconstruction of target posjtion, and execution. Central processing is strictly related to the type of sensory information provided to the subject, and therefore to the sensory system involved in the acquisition processo Spatial information reaches sen- sory organs with a superimposednoise. The charac- teristics of this noise will depend on how information is coded with respect to the detecting mechanisms. Transductionwill aIsocontribute to noise in a specific way. The fiDaI result is that spatial information is made available within the CNS with some degree of uncertainty. The level of uncertainty is strictly related to the sensory channel through which infor- mation is received. Since spatiaI information can be presented in different ways, centraI processingwill also assumedifferent characteristics. Whatever the processing,the goal is always that of producing an appropriate oculomotor command to the execution mechanisms. To avoid errors causing the eyes to go back and forth wasting a lot of energy, an oculomotor commandis likely to be generatedonly when a given level of reliability is reached in the centraI recon- struction of target position. The neural mechanisms involved in the execution process seemto be the samefor alI types of saccadic ere movements (Ron et al. 1972). Nevertheless, their performances can vary in relation to the state of the subject (aIertness or drowsiness)and to the environ- mentaI situation (light or darkness) (Becker and Fuchs1969;Ron et al. 1972;Korner 1975; Jiirgens et aI. 1981). How much does the morphology of the oculo- motor response evoked by a target presentation and the characteristicsof individuaI saccades depend on the type, quality, and quantity of sensory information provided to the subject? A way to investigate this point is to compare the saccadic responses evoked by the presentation of visuaI and auditory targets in the sameexperirnental conditions. Such a comparison in man was first made by Zahn et al. (1978). Surprisingly, no multiple saccade responses (a prirnary saccade followed by corrective secondary saccades) were reported by these authors and comparison was based only on prirnary saccade characteristics (latency, accuracy,and peak velocity). The short duration oi target presentation considered . Supported by CNR (Italy) and by INSERM (France) Offprint requests IO: D. Zambarbieri (addresss~e above) 0014-4819/82/0047/0417/$ 2.20

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

~
"""""""~""~"~-"~."'1f~;~"",~;:~".."", "'~:,-"-'W"~"1"~-~~~--"'"~~~~'.!!"~~~W""
:~.
E oorin1er1taJ~ Researd1@ Springer-Verlag 1982
Exp Brain Res (1982) 47: 417-427
Saccadic Responses Evoked by Presentation or Visual and Auditory Targets*
D. ZambarbieriJ, R. Schmid1, G. Magenesl, and C. Prablan~l Istituto di Infonnatica e Sistemistica, Università di Pavia, Strada Nuova 106 C, 1-27100 Pavia, Italy2 Laboratoire de Neuropsychologie Expérimentale, Unité 94 - INSERM, 16, Avenue du Doyen Lépine, F-69500 Bron, France
Summary. Saccadic eye movements evoked by thepresentation' of visual and auditory targets wereexamined arid compared. Differences were foundeither in ~he pattem of the saccadic response and inthe characteristics of single saccades of the sameamplitude. The longer latency and the higherpercentage of multiple saccade responses in theauditory case were attributed to a more complexcentrai processing, whereas the longer duration andthe lower peak velocity of the saccades to auditorytargets were attributed to reduced performances ofthe execution mechanism in the absence of vision.
Key words: Saccades - Audio-ocular responses -Fixation - Sound localization
Introduction
Saccades afe fi.xation ere movements which need aposition reference signal to be made available withinthe CNS. A reference signal can be reconstructedfrom sensory information about extemal targets orcan be intemal1y generated without sensory afferenceas far voluntary saccades in darkness. In the case ofsaccades evoked by target presentation three differ-ent processes should be accomplished: acquisition ofsensory information, centraI reconstruction of targetposjtion, and execution.
Central processing is strictly related to the type ofsensory information provided to the subject, andtherefore to the sensory system involved in theacquisition processo Spatial information reaches sen-sory organs with a superimposed noise. The charac-teristics of this noise will depend on how information
is coded with respect to the detecting mechanisms.Transduction will aIso contribute to noise in a specificway. The fiDaI result is that spatial information ismade available within the CNS with some degree ofuncertainty. The level of uncertainty is strictlyrelated to the sensory channel through which infor-mation is received. Since spatiaI information can bepresented in different ways, centraI processing willalso assume different characteristics. Whatever theprocessing, the goal is always that of producing anappropriate oculomotor command to the executionmechanisms. To avoid errors causing the eyes to goback and forth wasting a lot of energy, an oculomotorcommand is likely to be generated only when a givenlevel of reliability is reached in the centraI recon-struction of target position.
The neural mechanisms involved in the executionprocess seem to be the same for alI types of saccadicere movements (Ron et al. 1972). Nevertheless, theirperformances can vary in relation to the state of thesubject (aIertness or drowsiness) and to the environ-mentaI situation (light or darkness) (Becker andFuchs 1969; Ron et al. 1972; Korner 1975; Jiirgens etaI. 1981).
How much does the morphology of the oculo-motor response evoked by a target presentation andthe characteristics of individuaI saccades depend onthe type, quality, and quantity of sensory informationprovided to the subject? A way to investigate thispoint is to compare the saccadic responses evoked bythe presentation of visuaI and auditory targets in thesame experirnental conditions.
Such a comparison in man was first made byZahn et al. (1978). Surprisingly, no multiple saccaderesponses (a prirnary saccade followed by correctivesecondary saccades) were reported by these authorsand comparison was based only on prirnary saccadecharacteristics (latency, accuracy, and peak velocity).The short duration oi target presentation considered
. Supported by CNR (Italy) and by INSERM (France)
Offprint requests IO: D. Zambarbieri (address s~e above)
0014-4819/82/0047/0417/$ 2.20

418 D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
localized and flXed after 1 s from its presentation. As far conditionA, each subject participated in two sessions. In each sessionauditory targets were presented in a random sequence, and eachlateral position was considered three times. Subject's task in bothconditions A and B was to move bis eyes from the position of thecentrallight to the position of the lateral target (visual or auditory)and to fixate the target as accurately as possible.
in the study by Zahn et al. (1978) could bave beenODe of the reasons for the absence of correctivesaccades at least in the auditory case.
Since the pattern of the saccadic response is likelyto be much more related to centrai processing thansingle saccade parameters, it was worth repeating thestudy under experimental conditions giving secon-dary saccades the opportunity of occurring. In thisway the comparison between visually and acousti-cally evoked saccadic responses could bave beenextended by examining the frequency of single andmultiple saccade responses and by considering prim-ary and secondary saccades separately.
Condition C (Simple Reaction Times)
Material and Methods
Six subjects witb normal auditory, visual, and ocular motorfunctions were examined. Tests were perforrned in a quasi-anechoic room obtained by covering tbe walls by giass woolpanels. Subjects were seated in total darkness at tbe center of acircuiar frame 1.2 m in diameter. Tbeir head was restrained bybotb a chin rest and a bite board. Visual and auditory targets wereplaced on the frame at O, 5, lO, 15,20, and 30 deg right and left ofthe subject's saginal piane. Tbe centrai visual target was a greenlight emitting diode (LED) with an intensity of 64 cd/m2, whereasthe peripheral visual targets were red LEDs with an intensity of148 cdlm2. Tbe auditory targets were 5 cm diameter loudspeakersfed with a square wave signal at 7 Hz which produced pulsesinterspaced by about 66 ros. Sound pressure at the subject's earlevel was of 60 dB. Tbe parameters of the auditory stimulus werechosen after a previous study in which different auditory stimuli(pure tones, noise, pulses, and bursts) were compared (Zambar-bieri 1978). Comparison was based on the accuracy of tbe saccadicresponses foIlowing the presentation of the different types ofauditory targets. By using pulses at 15 Hz a fixation errar of lessthan 3 deg far target eccentricity betWeen O and 30 deg wasobserved. Intersubject variability was also found to be smaller withpulses at 15 Hz than with other types of auditory stimuli.
Tbree experimental conditions were considered in the presentstudy.
Condition A (Visually Evoked Responses)
The simple reaction Urne (RT) to the presentation of eccentricvisual and auditory targets was measured using a key-pressingdevice opening an electric circuit at tbe release of subject'spressure on a small sprung lever. Subjects were asked to releasethe key-pressing device as quickly as possible after stimuluspresentation. The modality of target presentation was the same asin conditions A and 8, with the only difference that the centraIlight was not switched off at the presentation of a lateral visual orauditory target. The preliminary fixation of the centrallight wasstilI necessary in arder to guarantee that visual targets appearedwith the desired eccentricities. On the other hand, care had to betaken to avoid that measured reaction times were those foUowingthe disappearance of the centrallight and not those related to theappearance of the lateral targets. The same use of the centrallightwas made also in the experiments with auditory targets to obtaincomparable experimental conditions.
AU the experiments were carried on under the contrai of aPDP 8 computer (Echallier et al. 1978). Eye movements wererecorded by means of an optoelectronic device. The left ere wasilluminated by infra-red sources on both nasal and temporal sides.The image of the ere was projected through a lens on two photo-transistors arrays mounted in a differential mode. This set-upallowed the measurement of the border position between iris andsclera without drift (Massé 1971). Accuracy in the measurement ofere position was of :t20' aver the fuIl range.
Eye movements were analyzed by means of an interactiveprogram implemented on a Laben 70 minicomputer (Cabiati andPastorrnerlo 1979). Latency, amplitude, duration, and peak veloc-ity were computed far primary and secondary saccades as shownschematicaUy in Fig. 1. Owing to their high predictability, recen-tering saccades were not considered in this analysis. In responsesevoked by auditory targets, only saccades executed within theperiod of sound presentation (1 s) were considered. Responseaccuracy was appreciated by computing the difference betweenere position at the end of sound presentation and ere positionduring the later fixation of the light presented at the same piace asthe previous sound source.
For the sake of brevity, we shall refer to the patterns of eremovement foUowing the presentation of visual and auditorytargets as "visual" or "auditory responses", respectively. In whatfoUows, we shall also use the subscripts "V" and "A" to distinguishwhether the parameters defined in Fig. 1 refer to a visual or to an
auditory response.
After calibration of ere movement, the centrai iight was firstpresented to give the subject a reference point far keeping bis eyesin the centrai position. After a short period of rime varyingrandomly between 3 and 5 s, the centrai Iight was switched off anda new Iight was presented far 2 s in a iateral position. In eachsession tbis procedure was repeated in such a way that each iatera!Iight appeared three times within a random sequence. Eachsubject participated in two sessions separated by at ieast 1 dar
Results
Condilion B (Auditory Evoked Responses) Statistical analysis was first made separately far eachsubject. Since the trends of data were found to besimilar far alI subjects with the same statisticaldifferences between visual and auditory responses,subjects were then considered altogether. Whenuseful to appreciate intrasubject and intersubjectvariations, results referring to both one subject (the
As in condition A, after calibration of eye movement, the centrailight was first presented. When this light was turned off, aloudspeaker was switched on in a lateral position. After 1 s ofsound presentation, the light placed in the SarDe lateral position asthe active loudspeaker was also presented for an additionalsecondo This procedure aimed at provoking a visual1y elicitedcorrective saccade whenever the source of sound was not correctly
419D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
. Auditory responsesoVisual responses
"'Vi
E
W~I-
ZOl-O~wCI:
W-.JQ.~
CI)
l !- -L -.. - - -1 .
l r I j-" r
VP1
VP2
velocitythreshold
-~-- - - - - - --- . . - ., ,. .. '" .. ," ,, ." .. .", '" '. ,- -, - -,',- - -, - - -,
, ', T ' D ' T . D ': 1 . 1 ' 2 ' 2',
o lO 20 30
T ARGET POSITION (deg)
Fig. 2. Simple reaction time oi key releasing responses to visualand auditory stimuli versus target position. Vertical bars indicate1 SD. Dashed lines represent the average values oi visual andauditory RT (262 and 221 ms, respectively)
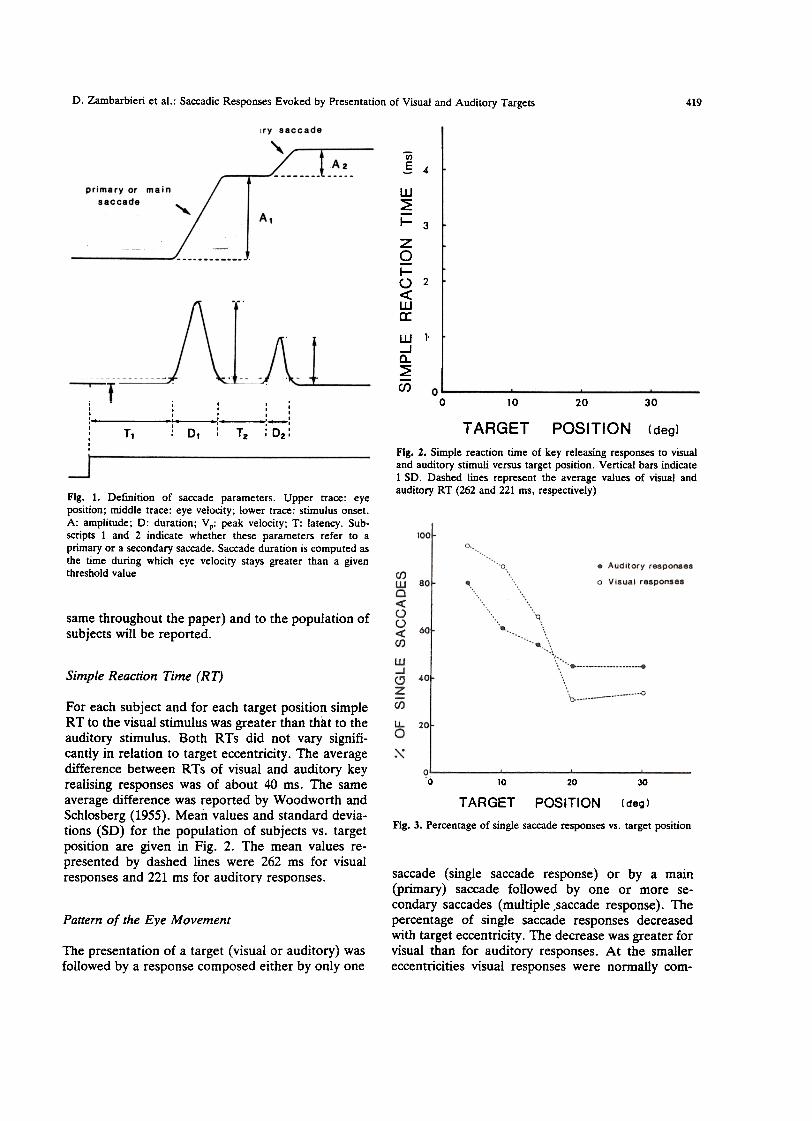
Fig. l. Definition of saccade parameters. Upper trace: eyeposition; middle trace: eye velocity; lower trace: stimulus onset.A: amplitude; D: duration; V p: peak velocity; T: latency. Sub-scripts l and 2 indicate whether these parameters refer to aprimary or a secondary saccade. Saccade duration is computed asthe rime during which eye velocity stays greater than a giventhreshold value
same throughout the paper) and to the popuiation oisubjects wiII be reported.
Simple Reaction Time (RT)
For each subject and for each target position simpleRT to the visual stimulus was greater than that to theauditory stimulus. Both RTs did not vary signifi-cantly in relation to target eccentricity. The averagedifference between RTs of visual and auditory keyrealising responses was of about 40 ms. The sameaverage difference was reported by Woodworth andSchlosberg (1955). Mean values and standard devia-tions (SD) for the population of subjects vs. targetposition afe given in Fig. 2. The mean values re-presented by dashed lines were 262 ms for visualresponses and 221 ms for auditorv responses.
"o IO 20 30
TARGET POSITION (deg)
Fig. 3. Percentage of singie saccade responses vs. target position
saccade (single saccade response) or by a main(primary) saccade followed by oDe or more se-condary saccades (multiple ,saccade response). Thepercentage of singie saccade responses decreasedwith target eccentricity. The decrease was greater farvisual than far auditory responses. At the smallereccentricities visual responses were normally com-
Pattern oJ the Eye Movement
The presentation oi a target (visual or auditory) wasiollowed by a response composed either by only one
400
300
200
,
100
D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets420
-(/)E
~
>-()ZLUI-<{-.J
20 3030 o IOlO 20o
T ARGET POSITION (deg)
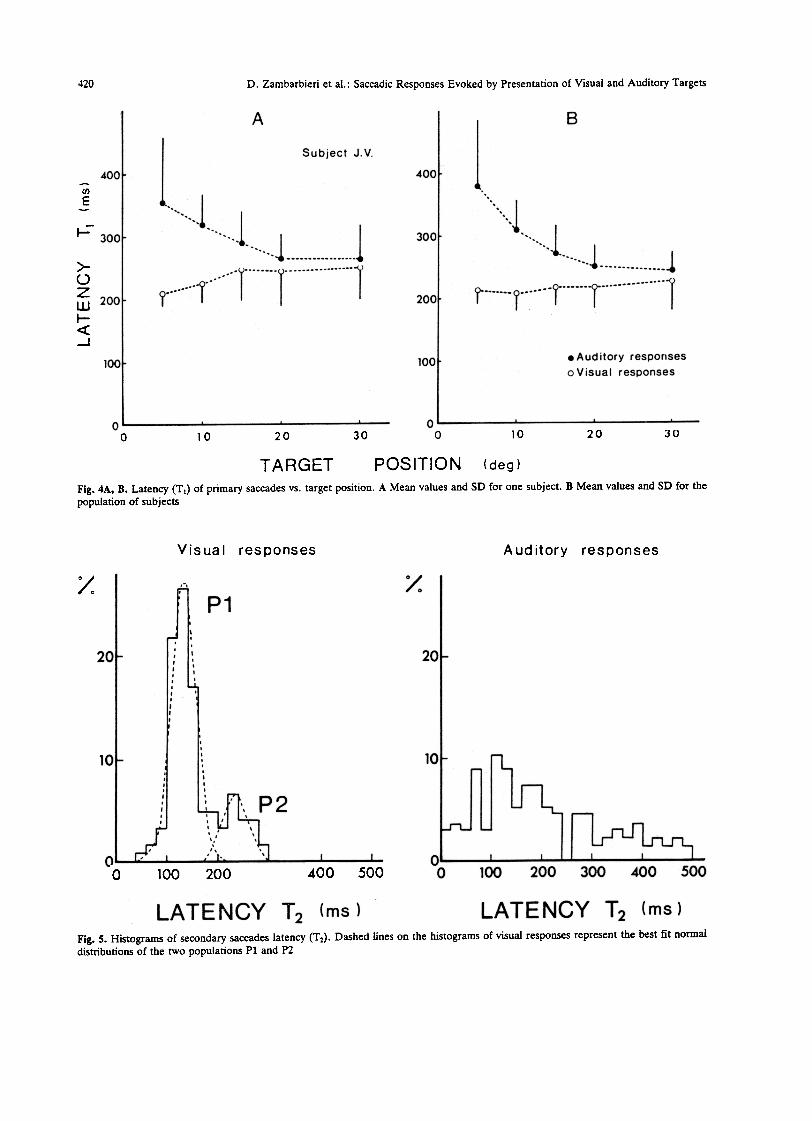
Fig. 4A, B. Latency (T J of primary saccades vs. target position. A Mean values and SD far one subject. B Mean values and SD far thepopulation of subjects
Visual responses A ud itory responses
%
400 500100 200o
Fig. S. Histograms oi secondary saccades latency (Tz). Dashed lines on the histograms oi visua! responses represent the best fit norma!
distributions oi the two populations Pl and P2

421D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
Table l. Statistica! parameters of secondary saccade latency. Twopopulations (Pl and PZ) of visual secondary saccades weredistinguished, whereas auditory saccades were considered asbelonging to only one population
Visual Auditory
Pl P2 onepopulation
posed by only one saccade, whereas auditoryresponses were often composed by at least twosaccades. At the larger eccentricities, the percentageof single saccade responses was greater in the audi-tory than in the visual case. Figure 3 gives thepercentage of single saccade responses vs. targetposition far the whole population of subjects.
The presence of a third saccade in multiple visualresponses was rare. At the end of these responses theeyes were right on the target. In contrast, auditoryresponses could be made of more than two saccadesand the fina! ere position generally underestimatedthat of the target. When the auditory target waspresented near the midline (i.e., 5 deg right or left)there could be no response at alI or a response withthe primary saccade in the wrong direction. Auditoryresponses were also observed with a primary saccadein one direction followed by a second saccade in theapposite direction and by a third saccade in thedirection of the first one. This pattem was neverfound in visual responses.
PercentageA verage latencyS.D.
77133.622.8
23237.529
100204.9131.4
"Oal
BWo::)t:-.Ja.2<{
wO<{()
~U)
>-CI:<{2CI:a..
"
/'b"
r
Latency/6
30
20
IO .6'. Auditory responsesoVisual responses
Results about latency afe reported separately farprimary and secondary saccades.
For alI subjects and far target eccentricities up to20 deg, the latency T lA of prirnary auditory saccadeswas significantly greater than the latency T lV ofprimary visual saccades. As shown in Fig. 4A, whichrefers to ODe subject, T lA decreased almost exponen-tiaUy with target eccentricity, whereas T lV slightlyincreased. This was a generaI trénd far aU subjects.Mean values and SD computed from a~i responsesafe given in Fig. 4B. The average value af T lAdecreased from 380 rns at 5 deg to 245 ms at 30 deg.T IV increased from 210 to 230 rns. Standard devia-tions decreased with target eccentricity in auditoryresponses and slightly increased in visual responses.For the same target position, the latency of primarysaccades in both visual and auditory responses wasfound to be unrelated to saccade amplitude.
The latency histograms of secondary saccades afeshown in Fig. 5. Two populations of secondary sac-cades can be recognized in visual responses. Let Pldenote the population of saccades presenting shorterlatency, and P2 the remaining population. The sepa-ration between Pl and P2 could be obtained througha best fitting of the experimental data by two normaldistributions. The two populations did not corre-spond to two distinct groups of subjects. The statisti-cal parameters of Pl and P2 afe given in Table 1. Thelatency of secondary saccades in auditory responsespresented a more uniform distribution, and no dis-
.6"
O~o IO 20 30
TARGET POSITION (deg)
Fig. 6. Mean values and SD of primary saccade amplitude versustarget position for the population of subjects. Table inserted infigure gives the percentage of correct, hypometric, and hyper-metric primary saccades
tinct populations could be recognized. The averagevalue and SD of the latency of auditory secondarysaccades considered as belonging to only ODe popula-tion is given in the last column of Table 1. Theaverage latency of P2 visual saccades was almost thesame as that of primary visual saccades. In contrast,the average latency of Pl visual saccades was signifi-cantly shorter. Due to the broad distribution ofauditory secondary saccade latency, its average valueis not meaningful.
Precision
Precision was first examined in terms of primarysaccade amplitude vs. target position. The results are
D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets422
-01(lJ"O
zoI-U)oa..
L.U>-L.U
-.J«z
l.L..
POSITIONTARGET (deg)
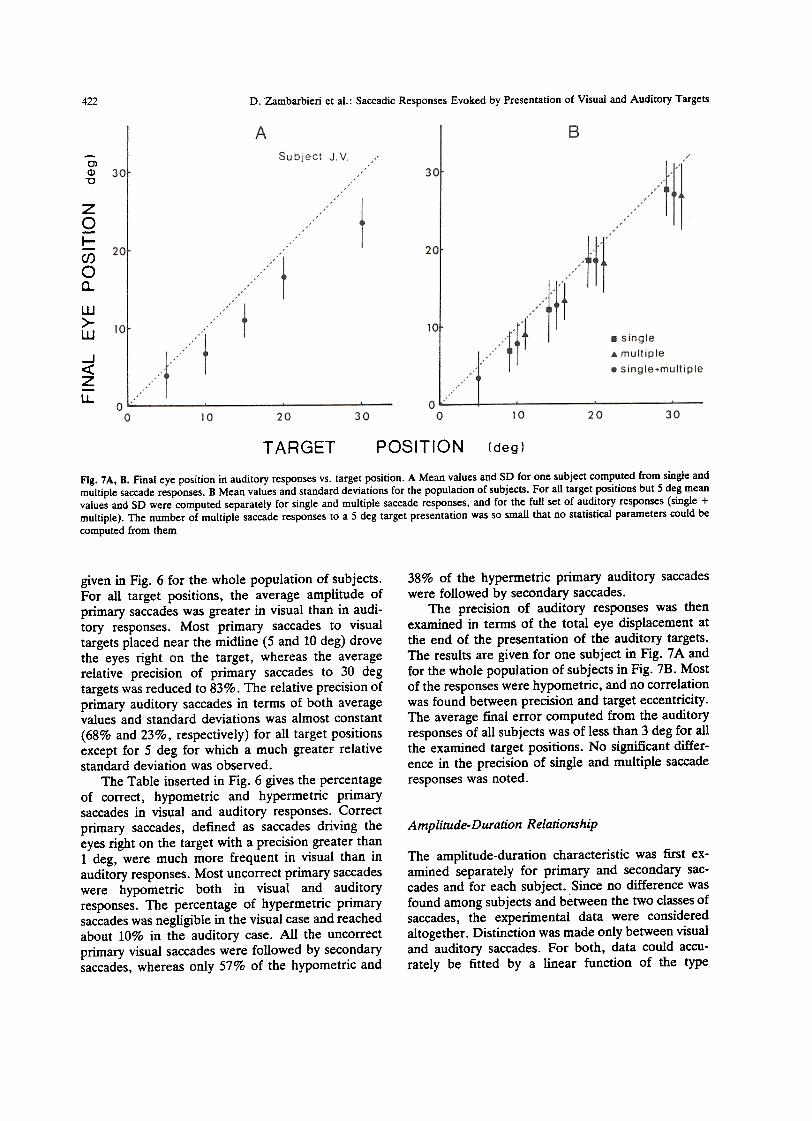
Fig. 7A, B. Final ere position in auditory responses vs. target position. A Mean values and SO far ODe subject computed from singie andmultiple saccade responses. B Mean values and standard deviations far the population of subjects. Far ali target positions but 5 deg meanvalues and SO were computed separately far single and multiple saccade responses, and far the full set of auditory responses (singie +multiple). The number of multiple saccade responses to a 5 deg target presentation was so small that no statistica! parameters could be
computed from them
38% of the hypermetric primary auditory saccadeswere followed by secondary saccades.
The precision of auditory responses was thenexamined in terms of the tota! eye displacement atthe end of the presentation of the auditory targets.The results afe given far ODe subject in Fig. 7 A andfar the whole population of subjects in Fig. 7B. Mostof the responses were hypometric, and no correlationwas found between precision and target eccentricity.The average fiDai errar computed from the auditoryresponses of ali subjects was of less than 3 deg far alithe examined target positions. No significant differ-ente in the precision of sÌngie and multiple saccaderesponses was noted.
given in Fig. 6 for the whole population of subjects.For all target positions, the average amplitude ofprimary saccades was greater in visual than in audi-tory responses. Most primary saccades to visualtargets placed near the midline (5 and lO deg) drovethe eyes right on the target, whereas the averagerelative precision of primary saccades to 30 degtargets was reduced to 83%. The relative precision ofprimary auditory saccades in terms of both averagevalues and standard deviations was almost constant(68% and 23%, respectively) for alI target positionsexcept for 5 deg for which a much greater relativestandard deviation was observed.
The Table inserted in Fig. 6 gives the percentageof correct, hypometric and hypermetric primarysaccades in visual and auditory responses. Correctprimary saccades, defined as saccades driving theeyes right on the target with a precision greater than1 deg, were much more frequent in visual than inauditory responses. Most uncorrect primary saccadeswere hypometric both in visual and auditoryresponses. The percentage of hypermetric primarysaccades was negligible in the visual case and reachedabout 10% in the auditory case. AlI the uncorrectprimary visual saccades were followed by secondarysaccades, whereas only 57% of the hypometric and
Amplitude-Duration Relationship
The amplitude-duration characteristic was first ex-amined separately for primary and secondary sac-cades and for each subject. Since no difference wasfound among subjects and between the two classes ofsaccades, the experimental data were consideredaltogether. Distinction was made only between visualand auditory saccades. For both, data could accu-rately be fitted by a linear function of the type
D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets 423
-CI)E
o
zo~«a::Jo
o lO 20 30 o lO 20 30
A MPLITUDE A (deg)
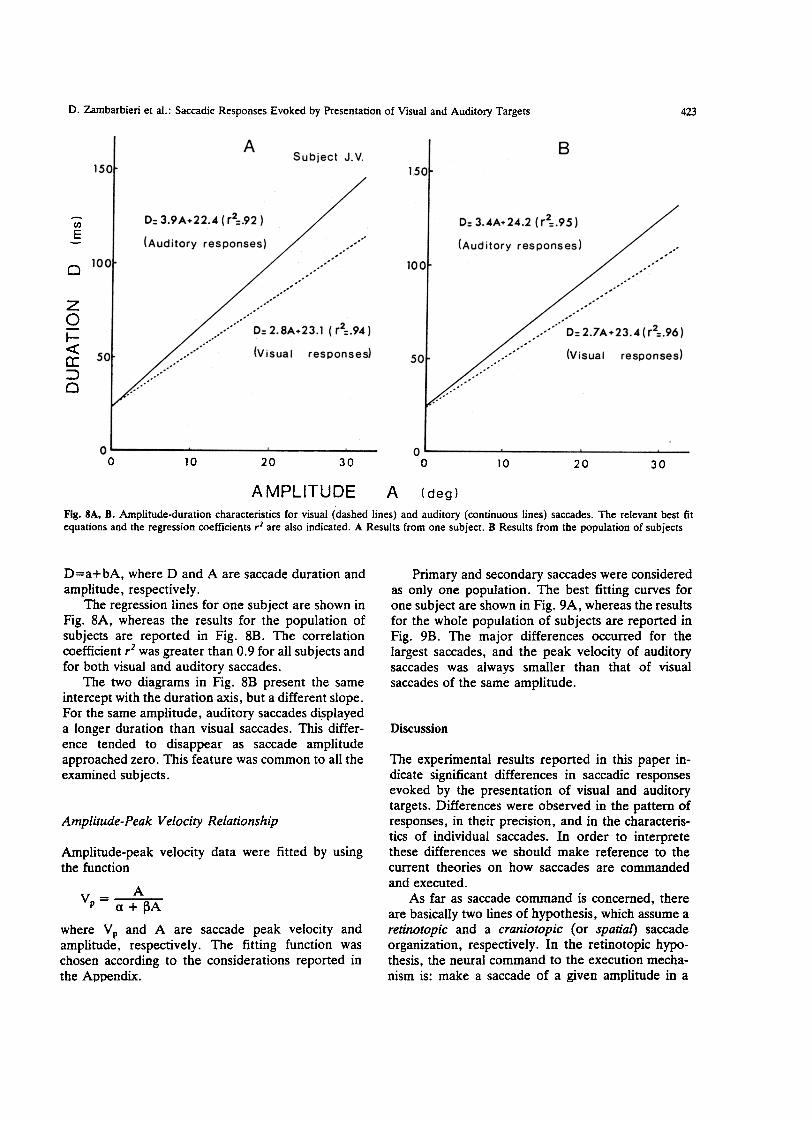
Fig. SA, B. Amplitude-duration characteristics far visual (dashed lines) and auditory (continuous lines) saccades. The relevant best fitequations and the regression coefficients r2 afe also indicated. A Results from one subject. B Results from the population of subjects
Primary and secondary saccades were consideredas only one population. The best fitting curves farone subject afe shown in Fig. 9A, whereas the resultsfar the whole population of subjects afe reported inFig. 9B. The major differences occurred far thelargest saccades, and the peak velocity of auditorysaccades was always smaller than that of visualsaccades of the same amplitude.
D=a+bA, where D and A afe saccade duration andamplitude, respectively.
The regression lines for ODe subject afe shown inFig. 8A, whereas the results for the population ofsubjects afe reported in Fig. 8B. The correlationcoefficient r2 was greater than 0.9 for alI subjects andfor both visual and auditory saccades.
The two diagrarns in Fig. 8B present the sameintercept with the duration axis, but a different slope.For the same amplitude, auditory saccades di.splayeda longer duration than visual saccades. This differ-ence tended to disappear as saccade amplitudeapproached zero. This feature was common to alI theexamined subjects.
Discussion
Amplitude-Peak Velocity Relationship
Amplitude-peak velocity data were fitted by usingthe function
The experimental results reported in this paper in-dicate significant differences in saccadic responsesevoked by the presentation of visual and auditorytargets. Differences were observed in the pattem ofresponses, in their precision, and in the characteris-tics of individuai saccades. In arder to interpretethese differences we should make reference to thecurrent theories on how saccades afe commandedand executed.
As far as saccade command is concemed, thereare basically two lines of hypothesis, which assume aretinotopic and a craniotopic (or spatia/) saccadeorganization, respectively. In the retinotopic hypo-thesis, the neural command to the execution mecha-nism is: make a saccade of a given amplitude in a
AV p = a + ~A
where Vp and A afe saccade peak velocity andamplitude, respectively. The fitting function waschosen according to the considerations reported inthe Appendix.

424 D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
o IO 20 30
AMPLITUDE A (deg)
Fig. 9A, B. Amplitude-peak velocity characteristics for visual (dashed lines) and auditory (continuous lines) saccades. The relevant best fitequations are also indicated. A Resuits from one subject. B Results from tbe popuiation of subjects
r ,. ,. .I 9 II -EH II III.I..,III
" I
9TH:
Sensoryinformetion
0EH
Efference
copy
8;9n815
. +., ',--+,
IIII -I
: EXECUTION MECHANISM~
Fig. lO. Model ofthe saccadicsystem. 8n1: estimated target position relative to the head; 9EH: eyeposition in the head; 9EH: efference copyof9EH; PG: pulse generator; NI: centrai neural integrator; K: gain
given direction. In the craniotopic hypothesis thecommand is: drive the eyes to a given position in theorbit. The retinotopic hypothesis is the simplest toexplain saccades evoked by the presentation of visualtargets since the retina directly provides the signalrequired by the execution mechanism (i.e., targetposition relative to the eyes). Nevertheless, there afe
experimental results which seem to indicate thatsaccades afe craniotopically and not retinotopicallyorganized. Some oi these results bave been reviewedby Robinson in 1975. More recently, further evi-dence was given by Mays and Sparks (1980)
As far as saccade execution is concerned, anaccurate description oi this process has been given by

425D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
Robinson (1975). Saccades afe executed through theaction of a neural pulse generator placed in thebrainstem. Since the accuracy of saccades of the sameamplitude was found to be largely independent oftheir duration and peak velocity (Zee et al. 1976;Jiirgens et al. 1981), the existence of a local feedbackhas been assumed (Zee et al. 1976; Berthoz 1981;Jiirgens et al. 1981). The pulse generator is switchedoff only when the intended fiDai ere position isreached.
Under the assumption that saccades afe spatiallyorganized, the model in Fig. 10 can be proposed. AcentraI reconstructor receives sensory informationand efference copy signals of ere, head, and bodypositions relative to their supporting platforms. Itreconstructs an absolute representation of externaland internaI space, and it sends appropriate refer-ence signals to the different motor systems. Inparticular, a reference signal representing the esti-mated target position relative to the head is sent tothe execution mechanism of the saccadic system. Acorollary discharge pathway tram the output of acentraI neural integrator (NI) provides the localfeedback required to switch off the pulse generator(PG).
Whatever the sensory system involved in theperception process, it is reasonable to assume thatinformation on target position reaches the. centraIreconstructor affected by some uncertainty. Thedegree of uncertainty will depend on the characteris-tics of both the incoming sensory signals and thedetecting and transducing mechanisms. The greaterthe uncertainty, the longer the processing timeneeded to obtain an accurate estimate of targetposition is.
The results on simple RT indicate that thedetection time is shorter far auditory than far visualstimuli. In both cases, it is almost independent ofstimulus position. Then, the observed greater latencyof auditory with respect to visual saccadic responsesin subjects asked to respond as accurately as possiblehas to be attributed to differences in the processingtime. It should also be inferred that the degree ofuncertainty associated to auditory spatial informationis not only greater than that affecting visual spatialinformation, but it is also dependent on stimulusposition with respect to the head. As a matter offact,when the source of sound is placed near the midline,both phase and amplitude differences at the level ofthe subject's ears afe very small and the estimate ofabsolute values could consequently be more difficult.
The longer time needed far processing auditoryspatial information would reverse the relationshipobserved between RTs of auditory and visualresponses. In monkeys trained to reach visual and
auditory targets only within :!:5 deg and therefore ina more "reflexive way", the latency of auditorysaccadic responses was found to be shorter than thatof visual responses (Whittington et al. 1981).
A greater uncertainty in the acquisition of spatialinformation by the auditory system does not neces-sarily imply that the accuracy of auditory saccadicresponses results much worse than that of visualresponses. Actually, it is noto An average fiDai errorof about 3 deg was observed at all target positions.Only the processing time would be different. Theexponential decrease of the latency of auditoryresponses with target eccentricity would be perfectlyconsistent with the hypothesis of an higher level ofuncertainty far the smaller eccentricities and with theobserved constant precision of the overall response.
The greater accuracy of primary saccades and thehigher percentage of single saccade responses in thevisual case can arso be related to the fact that theuncertainty in the acquisition of target position issmaller for visual than for auditory targets.
A more formai description of centrai processingin saccade generation has been developed in astochastic model that will be published elsewhere(Schmid et al. 1982). Results of computer simulationwill also be presented and compared with the resultsreported in this paper. A last comment is needed forthe amplitude-duration and amplitude-peak velocitycharacteristics of saccades to visual and auditorytargets. For the same amplitude, the latter displayeda longer duration and a smaller peak velocity. Sincethe execution mechanism is likely to be the same, theobserved difference can probably be related to alower Ì.;vel of the pulse generator activity. It has beenshown that the decrease of pulse amplitude producesa longer duration and a smaller peak velocity of sac-cades (Robinson 1975).
As far the anatomic localization of the centraireconstructor, it is likely that its functions are actu-ally distributed among several anatomical structures.The superior colliculus (SC) is very likely involved inthe centrai processing of sensory information forvisual and auditory saccade generation (Sparks andMays 1982). Both the visual and the auditory spaceafe represented in this structure (Cynader and Ber-man 1972; Gordon 1973; Drager and Hubel 1975,1976; Pollack and Hickey 1979). Moreover, electricalstimulation of SC has been proved to evoke saccadicresponses (Robinson 1972; Schiller and Stryker1972). Nevertheless, SC is certainly not the onlystructure in which centraI processing occurs asproved by the fact that saccades are preserved afterSC lesions (Pasik et al. 1966; Wurtz and Goldberg1972; Mohler and Wurtz 1977; Schiller et al. 1980).Visual saccades are abolished only if combined

426 D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
lesions of SC and visual cortex (Mohler and Wurtz1977) or of SC and frontal eye-field (Schiller et al.1980) are produced.
Appendix
The relationship between average (V m) and peak(V p) saccade velocity can be written as
Vm = yVp
~
where the coefficient y was reported to be the same(y = 0.6) for alI types of saccades and for alI saccadeamplitude (Baloh et al. 1975). Since
AV =1000-m D
where A is in deg, D in rns and V m in deg/s, and
D = a + bA,
the expected relationship between V p and A is
V = 1!!!J!1. ~ = ~p y a+bA a+~A ' .
y ywhere a = I@ a and ~ = 1000 b.
When function (1) was used to fit the experimentaldata obtained tram the visual and the auditoryresponses of the whole population of subjects, acorrelation coefficient of 0.96 and 0.91, respectively,was found between experimental and theoreticalvalues under best fit conditions.
The ratios ala and ~/b, which afe both expected togive an estimate of the sa me parapeter y = V mN p'
were computed tram the best fit of amplitude-duration and amplitude-peak velocity data. lt wasfound to be ala = 0.75 10-3 and ~/b = 0.6 10-3 far bothvisual and auditory saccades. The observed differ-ence might indicate that y = 0.6 is only an averagevalue, and small variations of y in relation to saccadeamplitude can actually occur.
(1)
Acknowledgement. The authors wish to tank Prof. M. Jeannerodfor helpful suggestions and criticism.
References
Baloh RW, Konrad HR, Sills A W, Honrubia V (1975) Thesaccade velocity test. Neurology 25: 1071-1076
Becker W, Fuchs AF (1969) Further properties of the humansaccadic system: ere movements and correction saccades withan without fixation Doints. Vision Res 9: 1247-1258
Berthoz A (1981) Discussion Summary. In: Fuchs AF, Becker W(eds) Progress in oculomotor research. Elsevier, North Hol-land, pp 53-56
Cabiati C, Pastormerlo M (1979) Un sistema per l'analisi auto-matica dei movimenti saccadici degli occhi. Thesis, Universityof Pavia, ltaly
Cynader M, Berman N (1972) Receptive field organization ofmonkey superior colliculus. J Neurophysiol 35: 187-201
Drager UC, Hubel DH (1975) Responses to visual stimulation andrelationship between visual, auditory and somatosensoryinputs in mouse superior colliculus. J Neurophysiol 38:690-713
Drager UC, Hubel DH (1976) Topography of visual andsomatosensory projections to mouse superior colliculus. JNeurophysiol 39: 91-101
Echallier JF, Pernier J, Prablanc C (1978) Système de pilotage destimulations et de recueil de paramétres relatifs à des réponsesmatrices. Int J Bio Med Comput 9: 341-352
Gordon B (1973) Receptive fields in deep layers of cat superiorcolliculus. J Neurophysiol 36: 157-178
Jiirgens R, Becker W, Komhuber HH (1981) Natural and drug-induced .variations of velocity and duration of human saccadiceye movements: evidence for a control of the neural pulsegenerator by local feedback. Biol Cybem 39: 87-96
Komer FH (1975) Non-visual contrai of human saccadic eyemovements. In: Lennerstrand G, Bach-y-Rita P (eds) Basicmechanisms of ocular motility and their clinica! implications.Pergamon Press, Oxford, pp 565-569
Massé D (1971) Etude de l'asservissement en vitesse du gIobeoculaire. In: Colloque sur les applications au domaine médicaldes méthodes et technologies de l'electronique. CEA Ed,Grenoble, pp 105-109
Mays LE, Sparks DL (1980) Saccades afe spatiaI1y, not retinocen-trically, coded. Science 208: 1163-1165
MohIer CW, Wurtz RH (1977) Role of striate cortex and superiorcolliculus in visual guidance of saccadic eye movements inmonkeys. J Neurophysiol 40: 74-94
Pasik T, Pasik P, Bender MB (1966) The superior colliculi and eyemovements. Arch Neurol15: 420-436
Pollack JG, Hickey TL (1979) The distributions of retino-collicularaxon temlinals in rhesus monkey. J Comp Neurol 185:587-002
Robinson DA (1972) Eye movements evoked by collicular stimu-lation in the aIert monkey. Vision Res 12: 1795-1808
Robinson DA (1975) Oculomotor contrai signals. In: Lenner-strand G, Bach-y-Rita P (eds) Basic mechanisms of ocularmotility and their clinical implications. Pergamon Press,Oxford, pp 337-374
Ron S, Robinson DA, Skavensky AA (1972) Saccades and tbequick phase of nystagmus. Vision Res 12: 2015-2022
SchiIIer PH, Stryker M (1972) SingIe-unit recording and stimula-tion in superior colliculus of tbe aIert rhesus monkey. JNeurophysiol 35: 915-924
SchiIIer PH, True SD, Conway JL (1980) Deficits in eye move-ments following frontal eye-field and superior colliculusablations. J Neurophysiol 44: 1175-1189
Schmid R, Magenes G, Zambarbieri D (1982) A stochastic modelof centrai processing in the generation of fixation saccades.In: Roucoux A (ed) Physiologica! and patbologica! aspects ofeye movements. Junk Pubi., The Hague (in press)
Sparks DL, Mays LE (1982) The role of the monkey superiorcolliculus in tbe spatial localization of saccade targets. In:Hein A, Jeannerod M (eds) SpatiaI1y oriented behavior.Springer, New York, in press
Whittington DA, Hepp-Reymond MC, F100d W (1981) Eye andhead movements to auditory targets. Exp Brain Res 41:3~R-1ti:;

427D. Zambarbieri et al.: Saccadic Responses Evoked by Presentation of Visual and Auditory Targets
stimoli visivi, acustici e vestibolari. Thesis, Polytechnic orMilan, Italy
Zee DS, Optican LM, Cook JD, Robinson DA, King Engel W(1976) Slow saccades in spinocerebellar degeneration. ArchNeurol 33: 243-251
Woodworth RS, Schlosberg H (1955) Experimental psychology.Methuen, London
Wurtz RH, Goldberg ME (1972) Activity of the superior colliculusin behaving monkey. IV. Effects of lesions on eye movement.J Neurophysiol 35: 587-596
Zahn JR. Abel LA, Dell'Osso LF (1978) The audio-ocularresponse characteristics. Sens Process 2: 32-37 .
Zambarbieri D (1978) Studio dei movimenti oculari indotti da Received July 21, 1981
Related Documents











