DATABASES rSNP_Guide: An Integrated Database-Tools System for Studying SNPs and Site-Directed Mutations in Transcription Factor Binding Sites Julia V. Ponomarenko, n Galina V. Orlova, Tatyana I. Merkulova, Elena V. Gorshkova, Oleg N. Fokin, Gennady V. Vasiliev, Anatoly S. Frolov, and Mikhail P. Ponomarenko Institute of Cytology and Genetics, Novosibirsk, Russia Communicated by Vladislav Baranov Since the human genome was sequenced in draft, single nucleotide polymorphism (SNP) analysis has become one of the keynote fields of bioinformatics. We have developed an integrated database-tools system, rSNP_Guide (http://wwwmgs.bionet.nsc.ru/mgs/systems/rsnp/), devoted to prediction of tran- scription factor (TF) binding sites, alterations of which could be associated with disease phenotype. By inputting data on alterations in DNA sequence and in DNA binding pattern of an unknown TF, rSNP_Guide searches for a known TF with alterations in the recognition score calculated on the basis of TF site’s sequence and consistent with the input alterations in DNA binding to the unknown TF. Our system has been tested on many relationships between known TF sites and diseases, as well as on site- directed mutagenesis data. Experimental verification of rSNP_Guide system was made on functionally important SNPs in human TDO2 and mouse K-ras genes. Additional examples of analysis are reported involving variants in the human cA-globin (HBG1), hsp70 (HSPA1A), and Factor IX (F9) gene promoters. Hum Mutat 20:239–248, 2002. r 2002 Wiley-Liss, Inc. KEY WORDS: SNP; transcription factor; DNA binding site; disease susceptibility; database; TDO2; F9; HBG1; HPFH; HSPA1A; K-ras; mutation analysis DATABASES: TDO2 – OMIM: 191070; GenBank: NM_005651; F9 – OMIM: 306900; GenBank: NM_000133; HBG1 – OMIM: 142200; GenBank: NM_000559; HSPA1A – OMIM: 140550; GenBank: NM_005345; http://wwwmgs.bionet.nsc.ru/mgs/systems/rsnp/ (rSNP_Guide); http://wwwmgs.bionet.nsc.ru/systems/Activ- ity/ (ACTIVITY); http://wwwmgs.bionet.nsc.ru/mgs/systems/selex/ (SELEX_DB) INTRODUCTION During the post-sequencing era in genomics, single nucleotide polymorphism (SNP) analysis has become one of the key research fields. Out of more than 2.84 million registered SNPs [Marth et al., 2001], at least 1.42 million have already been mapped [Sachidanan- dam et al., 2001]. Currently, the SNP-referred data are being accumulated in various databases, including the general databases, HGMD [Krawczak et al., 2000], dbSNP [Sherry et al., 2001], HGVbase [Brookes et al., 2000], ALFRED [Osier et al., 2001], OMIM [Hamosh et al., 2000], and the locus- specific ones, e.g., FANCONI [Gillio et al., 1997]. Questions regarding the functional role of SNPs in the coding regions and splice sites that cause alteration of protein structure and function now lend themselves to rather straightforward investigation. However, numerous SNPs that affect biological function are located outside the coding regions, i.e., in regulatory gene regions: promoters, enhancers, silencers, and introns. As known, gene transcription is launched by interaction of transcription factors (TFs) with relevant binding sites (TF-sites) in regulatory gene regions. Depending on localization, an SNP in a regulatory region may cause either complete elimina- tion of the natural TF site [Vasiliev et al., 1999; Boccia et al., 1996], formation of a novel spurious TF site [Knight et al., 1999; Piedrafita et al., 1996], or quantitative alteration in TF-binding efficiency Received 1 November 2001; accepted revised manuscript 10 June 2002. n Correspondence to: Julia V. Ponomarenko, Laboratory of Theoretical Genetics, Institute of Cytology and Genetics, 10 Lavrentyev Ave, Novosibirsk 630090, Russia. E-mail: jpon@ bionet.nsc.ru Contract grant sponsor: Russian Foundation for Basic Research; Contract grant number: RFBR-01-04-49860. DOI 10.1002/humu.10116 Published online in Wiley InterScience (www.interscience.wiley. com). r r2002 WILEY-LISS, INC. HUMAN MUTATION 20:239^248 (2002)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DATABASES
rSNP_Guide: An Integrated Database-ToolsSystem for Studying SNPs and Site-DirectedMutations in Transcription Factor Binding Sites
Julia V. Ponomarenko,nGalina V. Orlova, Tatyana I. Merkulova, Elena V. Gorshkova, Oleg N. Fokin,Gennady V. Vasiliev, Anatoly S. Frolov, and Mikhail P. Ponomarenko
Institute of Cytology and Genetics, Novosibirsk, Russia
Communicated by Vladislav Baranov
Since the human genome was sequenced in draft, single nucleotide polymorphism (SNP) analysis hasbecome one of the keynote fields of bioinformatics. We have developed an integrated database-toolssystem, rSNP_Guide (http://wwwmgs.bionet.nsc.ru/mgs/systems/rsnp/), devoted to prediction of tran-scription factor (TF) binding sites, alterations of which could be associated with disease phenotype. Byinputting data on alterations in DNA sequence and in DNA binding pattern of an unknown TF,rSNP_Guide searches for a known TF with alterations in the recognition score calculated on the basis ofTF site’s sequence and consistent with the input alterations in DNA binding to the unknown TF. Oursystem has been tested on many relationships between known TF sites and diseases, as well as on site-directed mutagenesis data. Experimental verification of rSNP_Guide system was made on functionallyimportant SNPs in human TDO2 and mouse K-ras genes. Additional examples of analysis are reportedinvolving variants in the human cA-globin (HBG1), hsp70 (HSPA1A), and Factor IX (F9) genepromoters. Hum Mutat 20:239–248, 2002. r 2002 Wiley-Liss, Inc.
KEY WORDS: SNP; transcription factor; DNA binding site; disease susceptibility; database; TDO2; F9; HBG1;HPFH; HSPA1A; K-ras; mutation analysis
DATABASES: TDO2 – OMIM: 191070; GenBank: NM_005651; F9 – OMIM: 306900; GenBank: NM_000133;HBG1 – OMIM: 142200; GenBank: NM_000559; HSPA1A – OMIM: 140550; GenBank: NM_005345;http://wwwmgs.bionet.nsc.ru/mgs/systems/rsnp/ (rSNP_Guide); http://wwwmgs.bionet.nsc.ru/systems/Activ-ity/ (ACTIVITY); http://wwwmgs.bionet.nsc.ru/mgs/systems/selex/ (SELEX_DB)
INTRODUCTION
During the post-sequencing era in genomics, singlenucleotide polymorphism (SNP) analysis has becomeone of the key research fields. Out of more than 2.84million registered SNPs [Marth et al., 2001], at least1.42 million have already been mapped [Sachidanan-dam et al., 2001]. Currently, the SNP-referred dataare being accumulated in various databases, includingthe general databases, HGMD [Krawczak et al.,2000], dbSNP [Sherry et al., 2001], HGVbase[Brookes et al., 2000], ALFRED [Osier et al.,2001], OMIM [Hamosh et al., 2000], and the locus-specific ones, e.g., FANCONI [Gillio et al., 1997].
Questions regarding the functional role of SNPs inthe coding regions and splice sites that causealteration of protein structure and function now lendthemselves to rather straightforward investigation.However, numerous SNPs that affect biologicalfunction are located outside the coding regions, i.e.,in regulatory gene regions: promoters, enhancers,
silencers, and introns. As known, gene transcription islaunched by interaction of transcription factors (TFs)with relevant binding sites (TF-sites) in regulatorygene regions. Depending on localization, an SNP in aregulatory region may cause either complete elimina-tion of the natural TF site [Vasiliev et al., 1999;Boccia et al., 1996], formation of a novel spurious TFsite [Knight et al., 1999; Piedrafita et al., 1996], orquantitative alteration in TF-binding efficiency
Received 1 November 2001; accepted revised manuscript10 June 2002.
nCorrespondence to: Julia V. Ponomarenko, Laboratory ofTheoretical Genetics, Institute of Cytology and Genetics,10 Lavrentyev Ave, Novosibirsk 630090, Russia. E-mail: [email protected]
Contract grant sponsor: Russian Foundation for Basic Research;Contract grant number: RFBR-01-04-49860.
DOI10.1002/humu.10116Published online in Wiley InterScience (www.interscience.wiley.com).
rr2002 WILEY-LISS, INC.
HUMANMUTATION 20:239^248 (2002)

[O’Neill et al., 1990; Matsuda et al., 1992; Moi et al.,1992; Tsutsumi-Ishii et al., 1995; Langdon andKaufman, 1998]. Because gene expression ineukaryotes is mediated not only by individualtranscription factors, but also through sophisticatedtranscriptosome machinery [Gall et al., 1999] andgene network relationships, very few SNPs orgenome variations in regulatory gene regions mayultimately result in alteration of the expressionpatterns of many genes. All things considered,mutations in regulatory gene regions may be anunderlying cause of many phenomena, ranging fromdisease predisposition or drug susceptibility to severegenetic disorder.
The goal of our SNP-addressed system is to searchfor TFs that are potentially capable of binding to DNAregion with mutation. In practice, to decrease theexpensive and time-consuming procedure of choosingthe appropriate TFs binding to particular DNA,researchers use the software packages, e.g., MatIn-spector [Quandt et al., 1995] or TESS [Stoeckertet al., 1999]. As a rule, these programs implement TF-site recognition based on textual similarity of DNAunder study to that of known TF-sites. So, they yieldsatisfactory prediction only if the mutation leads toelimination or appearance of a TF site [Roulet et al.,1998, 1999], but not if the mutation quantitativelyalters TF-binding efficiency [Matsuda et al., 1992;Moi et al., 1992; Tsutsumi-Ishii et al., 1995]. Besides,a single mutation may cause misbalance betweenseveral overlapping TF-sites, thus causing diseasepenetration due to re-arrangement of gene networkfunctioning [O’Neill et al., 1990; Langdon andKaufman, 1998]. In fact, many experiments [Bienvenuet al., 1995; Cowell et al., 1996; NedelchevaKristensen et al., 1999; Vasiliev et al., 1999] demon-strate that a mutation-altered TF site cannot bereliably recognized by only its textual similarity to theknown TF site. All things considered, there is a needfor designing novel approaches aimed at SNP analysis.
In this paper, we introduce the integrated database-tools system rSNP_Guide devoted to search for TFscapable of binding to a DNA region with a mutation.The main idea of rSNP_Guide is to involve in thisprocess both sequences of known TF-sites andavailable data on alterations in binding of mutatedDNA to unknown TFs. As additional experimentalevidence, one may use gel mobility shift assay with thecell nuclear extracts, and expression of reporter geneconstructs carrying regulatory DNA of interest, etc.These data could be used as an additional round ofprediction procedure that will improve the accuracy ofTF site recognition. That is, all TF sites predictedcould be additionally tested for the requirement thatexpected alterations in their binding to DNA withmutation should be consistent with the experimentaldifference between the intact and mutated data sets.By applying this additional mutation-based round of
selection, the false-positive error rate, which is themain limitation of any site recognition approach,could be significantly reduced. The rSNP_Guidesystem implements this idea of performing an addi-tional round of selection of TF sites that werepreliminarily predicted by the MATRIX system[Ponomarenko et al., 1999].Currently, the rSNP_Guide tools system has been
tested on 33 control data sets representing bothnatural mutations associated with diseases (15 TF-sites with 21 mutations) and data on site-directedmutagenesis (18 TF-sites with 28 mutations). For twoSNPs in the sixth intron of the TDO2 gene (MIM#191070) associated with behavioral disorders [Ryanet al., 1987; Comings et al., 1996], the YY1-bindingsite altered by mutations was predicted by therSNP_Guide. The confirming experiments with anti-YY1 antibodies are also discussed below. Besides, themutations in the second intron of the mouse K-rasgene associated with lung tumor susceptibility weretreated. As a result, the GATA site specific to thetumor-sensitive allele was predicted and finallyconfirmed by the anti-GATA antibody assay.
DATABASE STRUCTURE ANDORGANIZATION
A scheme of the rSNP_Guide system is given inFigure 1. Three databases represent the depositorypart of the rSNP_Guide system: rSNP_DB, for storingthe experimental data; SYSTEM, for details ofexperimental design; and rSNP_BIB, for literaturesources. The data analysis part contains the samples ofexperimentally detected sequences of natural TF-sites(database SAMPLES) and their textual cores in theoligonucleotide-position frequency matrices (databaseMATRIX) [Ponomarenko et al., 1999]. Currently,rSNP_Guide provides a search for a potential TF-candidate among 41 types of known TF-sites. Thenumber of TF-sites to be predicted is restricted to 41,this being the number of representative samples ofnatural TF-sites documented in the databaseSAMPLES, and, consequently, the number of weightmatrices for potential TF-site recognition by theMATRIX tools. So, the current release of therSNP_Guide is based on more than 2,000 experimen-tally identified and multiply aligned DNA sequencescontaining 41 types of TF-sites. For these sites, morethan 500 frequency matrices at 32 oligonucleotidealphabets were optimized, documented, and used.The Java-script applet rSNP_Tools implementsthese frequency matrices for TF-site recognition.Finally, the rSNP_Report database accumulatesthe characteristic examples of successful TF-site’spredictions for both natural and artificial mutationsaltering DNA binding to TFs. The more detaileddescription of the rSNP_Guide system’s structure andorganization is given elsewhere [Ponomarenko et al.,2001b].
240 PONOMARENKO ETAL.

MUTATION ANALYSIS
Each DNA variant of a user’s interest should becharacterized by the mutation-caused alteration inDNA binding to unknown TF, detected either byaffinity measurement, gel mobility assay, CAT-assay,dual-luciferase assay, or some other experiments.Because the data could be expressed not only asdichotic state, presence/absence, but also quantita-tively, the magnitude of DNA binding to unknown TFshould be expressed as the relative degree rangingfrom þ 1 to � 1. For this purpose, a user should followthree empirical rules: 1) the maximal degree, þ 1,should be attained only to the well-detectable DNA/TF complex; 2) if the DNA/TF-complex has not beendetected in appropriate experimental conditions, thenthe threshold degree, 0, should fit at best; 3) only ifadditional observations and/or other theoreticalreasoning support the absence of DNA/TF complex,then the minimal degree,�1, should be chosen. As aconsequence, each DNA variant of a user’s interestshould be assigned by the relative degree, Score(X), ofDNA binding to unknown TF denoted by X. As arule, there is no information, whether both chains ofdouble-stranded DNA or only a single one bind tounknown TF, so the rSNP_Guide system generatesand testifies all four possible cases, X’s, i.e., X++,both direct- and complement-chains bind to TF;X+�, only the direct-chain does; X�+, only thecomplement-chain does; and, in order to make self-verifying, X��, no binding.
Besides, for each DNA variant examined, twosequences corresponding to two strands of the double-stranded DNA, denoted as direct-chain and com-pliment-chain, are used, i.e., 2�N sequences,{Sn}1rnr2�N, for N variants of DNA. Upon eachsequence, Sn, for each TF of a user’s interest,the maximum rate of TF site recognition score,Scoren(TF), is calculated in compliance with themathematical description given previously [Ponomar-enko et al., 1999].
Thus, rSNP_Guide compares the input alterationsin DNA binding to unknown TF, Score(X), with thosecalculated for known TF’s, Score(TF)’s, in order toselect significantly similar pairs, X=TF. From a bulk of
similarity scales, the most clear and widely appliedEuclidean distance is chosen:
DTF;X ¼ ½SnðScorenðXÞ � ScorenðTFÞÞ2�1=2; ð1Þ
DXþ�;X� ¼ ½SnðScorenðXþ � Þ
� ScorenðX��Þ2�1=2; ð2Þ
where: X+�A{X++, X+�, X�+}, all threepossible cases of DNA binding to unknown TF.By the same reasoning, the t-test was preferred out
of available statistical tests:
dTF;Xþ� ¼ DX��;Xþ� �DTF;Xþ�
� ta;v�s:d:ðDTF;Xþ�Þ; ð3Þ
dTF;X�� ¼ ta;vxs:d:ðDTF;X��Þ �DTF:X��; ð4Þ
IF ½fdTF;Xþ�40gAND fdTF;X��o0g�THEN fX ¼ TFg;
ð5Þ
where ta;n, t-value at significance a and freedomdegree n; s.d., standard deviation.For mathematical and computing details of analysis
provided by the rSNP_Guide, see Ponomarenko et al.[2002a].
EXAMPLES OF rSNP GUIDEUSAGESNP in the Rat Angiotensin II Type1A ReceptorGene Promoter
Let us consider the rSNP_Tool’s usage for analysisof A/T-rich region in the rat angiotensin II type 1Areceptor gene promoter. As shown experimentally,two point-paired mutations in this region,-557(TtttaA)>(GtttaC), result in 20-fold decreaseof gene expression [Beason et al., 1999]. The direct-chain sequence of the wild-type DNA variant, WT, 50-aaatattttg tgccattttgttttggcagtctgtttttatTtttaAataaatg-cagctcgagcaacgtgtaaactagaaggaactg-30, was extractedfrom the EMBL databank (S67615, 19–104 bp). Fordirect-chain sequence of the mutant variant, MT, theunderlined TtttaA 6-bp fragment was replaced by theGtttaC sequence. Each direct-chain sequence wasautomatically converted to the opposite direction byusing the rSNP_Guide option "Converter" (Fig. 2d,arrow). With the sequences prepared, among 41 TFslisted in the rSNP_Guide user’s interface (Fig. 2a), aTF of user’s interest is selected and clicked on (arrow).The TF-site recognition tool is loaded (Fig. 2b). Eachsequence prepared is input into the box ‘‘Input DNAsequence: from screen,’’ and the bottom ‘‘Execute’’ isclicked on. The profile of the TF-site recognitionScore calculated upon the input sequence is gene-rated (in our example, Fig. 3a). The maximum valueof the profile, Score(TF), (in our example, 0.2) isinput into the proper box of the rSNP_Guide
SAMPLES
MATRIX
rSNP_Tools
Data analysis(natural TF sites)
How to use(character examples)
rSNP_Report
Depository(SNP & mutation)
rSNP_DB
SYSTEM
rSNP_BIB USER
FIGURE 1. Scheme of the rSNP Guide system.
DATABASE FOR SNPSAND SITE-DIRECTEDMUTATIONS 241

interface (Fig. 2a). Then this procedure is repeatedfor the next TF of interest (in our example, MEF-2).For our example, all Score(TF)s obtained are givenin Figure 2c and in Table 1. As seen, both TFs ofinterest, HNF1 and MEF-2, are characterized bythe positive degrees of their TF-site recognitionscores, because these TFs are specific to theA/T-rich DNA regions. Gel-shift assay using anti-MEF-2 antibody showed that MEF-2 is truly predicted[Beason et al., 1999], whereas the HNF1 predictionis a false-positive. For discarding the false-positiveprediction, additional data on alterations in DNAbinding to unknown TF were introduced into theanalysis.
The alterations in mutant DNA binding tounknown TF cause 20-fold decrease of the rAT1AR-C gene expression [Beason et al., 1999]. This meansthat both wild-type and mutant DNA variants bind tounknown TF, but the binding strength is lesser for themutant DNA. Thus, for wild-type DNA, put themaximal degree, +1, into the box ‘‘SNP#1,’’ the field‘‘DNA/Protein-Binding’’ (Fig. 2d), and for mutantDNA, put relatively lesser degree into the box
‘‘SNP#2.’’ Following this empirical reasoning, wemay input into the box ‘‘SNP#2’’ the values 0.5, or0.2, or even 0.0. Figure 2d shows how {X=MEF-2}was truly predicted by the rSNP_Guide with degree0.5 entered in the box ‘‘SNP#2.’’ In Table 1,the results obtained for all three possible degreesare presented. As seen, with each of threevariants, {X=MEF-2} is successfully predicted and{X=HNF1}, false-positive, is discarded. So, thisproves the evidence that the second round ofrecognition procedure enhances the accuracy of TFsite’s prediction.
Two SNPs in the Sixth Intron ofTDO2
As an example of the rSNP_Guide application inpractice, we consider here mutations in the humanTDO2 gene, which have been earlier associatedwith some behavioral disorders [Comings et al.,1996]. Tryptophan 2,3-dioxygenase (TDO2, MIM#191070; EC 1.13.11.11) is the rate-limiting enzymein the oxidative degradation of tryptophan, theserotonin precursor, which, in turn, controls serotonin
FIGURE 2. Example of rSNP Guide usage: a: User’s interface in the beginning. b: ATF-site recognition tool loaded. c: User’sinterfacewith the input data dot-framed. d:User’s interfacewith the rSNP Guide prediction. Arrows pinpoint details discussedin the section rSNP Guideusage.
242 PONOMARENKO ETAL.
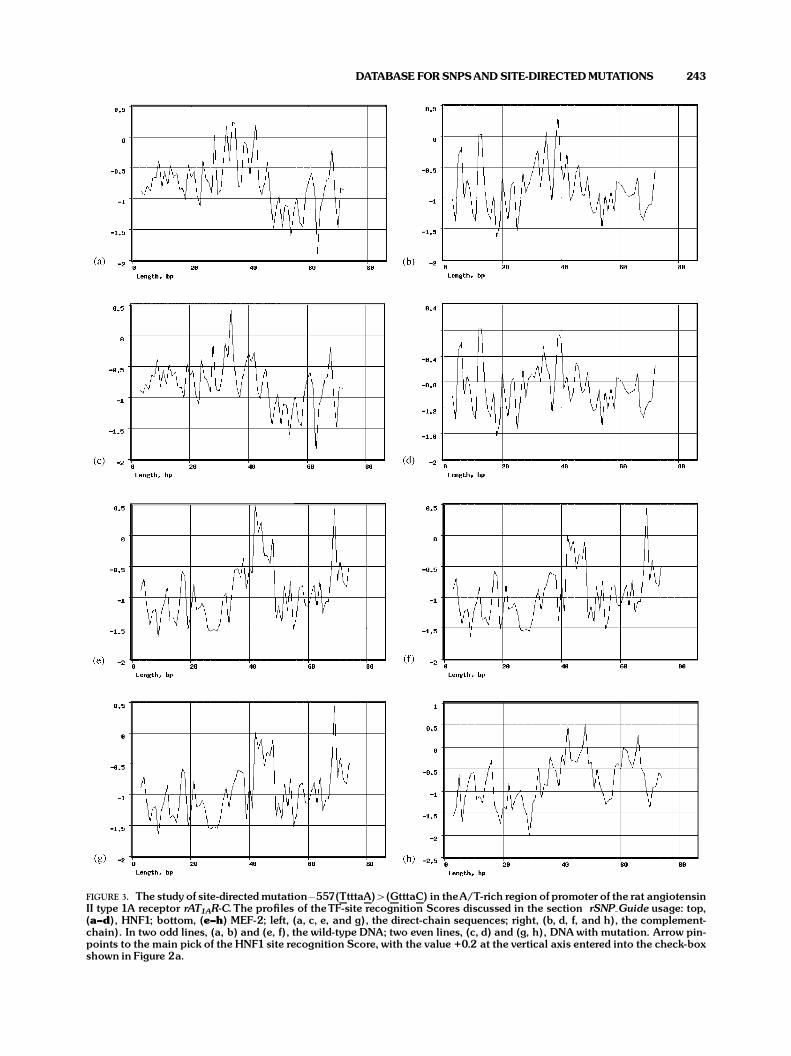
FIGURE 3. The study of site-directedmutation�557(TtttaA)4(GtttaC) in theA/T-rich region of promoter of the rat angiotensinII type 1A receptor rAT1AR-C.The pro¢les of theTF-site recognition Scores discussed in the section rSNP Guide usage: top,(a^d), HNF1; bottom, (e^h) MEF-2; left, (a, c, e, and g), the direct-chain sequences; right, (b, d, f, and h), the complement-chain). In two odd lines, (a, b) and (e, f), the wild-type DNA; two even lines, (c, d) and (g, h), DNAwith mutation. Arrow pin-points to the main pick of the HNF1 site recognition Score, with the value +0.2 at the vertical axis entered into the check-boxshown in Figure 2a.
DATABASE FOR SNPSAND SITE-DIRECTEDMUTATIONS 243
concentration in the organism. Defects in serotoninmetabolism and abnormal serotonin/tryptophan levelshave been reported for many behavioral disorders.This makes the TDO2 gene a potential candidategene in psychiatric genetics [Comings et al., 1996].Two SNPs, +663G>A and +666G>T in the sixthintron, have been associated with behavioral diseases[Comings et al., 1996]. However, how the mechan-isms of these mutations affect the TDO2 geneexpression remains unclear.
For studying +663G>A and +666G>T muta-tions, we have synthesized three double-stranded34 bp-oligoDNAs: 1) WT, direct-chain, 50-cagttgc-caaataatgGcaGataagaatagggag-30; 2) M1, direct-chain,50-cagttgccaaataatgAcaGataagaatagggag-30; and 3)M2, direct-chain, 50-cagttgccaaataatgGcaTataagaa-tagggag-30. Using a gel mobility shift assay, we haveobserved that binding of the rat liver cell nuclearextract to these oligoDNAs indicates at least fiveunknown TFs (Fig. 4a, arrows). As seen, two TFs(broken arrows) bind to all three oligoDNAs, regard-less of the type of mutation, and the binding patternof three other DNA/TFs complexes (solid arrows) isaltered by mutations. Let us analyze the unknownDNA/TF complex marked by bold-faced arrow inFigure 4a. Note that exactly this complex ischaracteristic only for the wild-type allele (WT),whereas it is undetectable (+663G>A) or weakened(+666G>T) in case of a mutation. As seen in Figure4b illustrating the usage of the rSNP_Guide, thebox ‘‘SNP#1’’ for WT data input, was assigned bythe maximal degree, +1; the box ‘‘SNP#2’’ for+663G>A data input, by the threshold degree, 0;and the box ‘‘SNP#3’’ for +666G>T, by intermediatedegree, 0.5. As shown in section ‘‘Prediction’’ in
Figure 4b, the YY1-binding site was predicted byrSNP_Guide.Figure 4c illustrates the experiment with the anti-
YY1 antibodies and wild-type oligoDNA used forverifying the rSNP_Guide prediction. As seen, addi-tion of specific anti-YY1 antibodies to the nuclear cellextract eliminates the DNA-protein complex understudy (lane 1, solid arrow) and produces the slowermigrating ternary complex (lane 2). The super-shiftassay proves that the DNA/TF-complex is formed byYY1 binding to the wild-type oligoDNA, as waspredicted by rSNP_Guide.As is known, YY1 is an ubiquitous multifunctional
nuclear protein, which either represses or stimulatesgene expression depending on the context of corre-sponding regulatory gene region. As a component ofthe nuclear matrix, YY1 may be also involved inchromatin organization, possibly by tethering DNA tothe nuclear matrix. Whichever it is, it influencestranscription efficacy. Thus, mutations damaging theYY1 site may change transcription of TDO2, which, inturn, may cause penetration of psychiatric disorders.
Analysis of SNP-Related Diseases
In Table 2, three additional examples of SNPsassociated with diseases and analyzed by therSNP_Guide system are listed.The first example deals with the K-ras gene, which
is widely used in different mouse strains as a geneticmarker of susceptibility to spontaneous and chemicallyinduced mutagenesis and tumor in lung [Ryan et al.,1987]. Three alleles of this gene, in its second intron,have the character nucleotides in positions +288 and+296, namely, the sensible alleles are +288C and
TABLE 1.The Illustrative Example of rSNP GuideUsage forAnalysis of A / T-RichRegion inPromoter of the Rat Angiotensin IIType1A ReceptorGene rAT1AR-Cn
DNA TFof interest Analysis, variant #1 Analysis, variant #2 Analysis, variant #3
(chain) HNF1 MEF-2 Rate X++ X+� X�+ X�� Rate X++ X+� X�+ X�� Rate X++ X+� X�+ X��
WT (+) 0.2 0.5 1.0 1.0 1.0 �0.1 �0.1 1.0 1.0 1.0 �0.1 �0.1 1.0 1.0 1.0 �0.1 �0.1(�) 0.4 1.0 1.0 �0.1 1.0 �0.1 1.0 �0.1 1.0 �0.1 1.0 �0.1 1.0 �0.1
MT (+) 0.4 0.0 0.5 0.5 0.5 �0.1 �0.1 0.2 0.2 0.2 �0.1 �0.1 0.0 0.0 0.0 �0.1 �0.1(�) �0.1 0.5 0.5 �0.1 0.5 �0.1 0.2 �0.1 0.2 �0.1 0.0 �0.1 0.0 �0.1
Euclidean DHNF1,X 1.2 0.9 1.0 0.8 1.1 1.0 0.9 0.8 1.1 1.0 0.8 0.8distance DMEF-2;X 0.7 1.4 0.6 1.4 0.6 1.4 0.7 1.4 0.7 1.3 0.8 1.4
(formulas1and 2) DX^,X 1.8 1.3 1.3 0.0 1.6 1.1 1.1 0.0 1.6 1.1 1.1 0.0s.d.(DFT,X) 0.3 0.4 0.3 0.4 0.3 0.3 0.1 0.4 0.3 0.2 0.04 0.4
t-value, tao0,05;n=2 2.92Similarity estimate dHNF1; X �0.4 �0.7 �0.6 0.5 �0.4 �0.6 �0.2 0.5 �0.3 �0.6 0.1 0.5(formulas 3 and 4) dMEF-2;X 0.1 �1.2 �0.2 �0.1 0.1 �1.0 0.0 �0.1 0.1 �0.9 0.2 �0.1
Prediction (formula 5) X=MEF-2 X=MEF-2 X=MEF-2
nNote: (+), direct-chain; (�), complement-chain; Rate, theempirical rates input in accordancewith causedbymutation 20-folddecrease inDNAbinding to unknownTF [Beason et al.,1999]; s.d., standard deviation; bold-faced, the rSNP_Guide prediction(X=MEF-2) consistent with the experiment with the anti-MEF-2 antibody [Beason et al., 1999]; underlined, false-positive(X=HNF1) prediction made by the frequency-matrix approach and discarded successfully by rSNP Guide due to the secondround of experimentally based recognition. As seen, both true (X=MEF-2) prediction and successful false-positive {X=HNF1}discard are reproducible for various empirical variants of mutation-related data.
244 PONOMARENKO ETAL.

+296A, the intermediate one is +296C, and resistantalleles are +288G and +296C [Chen et al., 1994].An unknown TF, specific only for the sensible allele,has been detected by the gel mobility shift assay in thelung cell nuclear extract with addition of three 30 bpoligoDNAs relevant to alleles [Ponomarenko et al.,2002a]. As follows from Table 2, the rSNP_guidesystem predicts that the GATA site in the secondintron of the K-ras gene is respective for the tumorsusceptibility, whereas disruption of this site bymutations causes tumor resistance. This predictionhas been confirmed by the anti-GATA antibody assay[Ponomarenko et al., 2002a].
The second example describes two natural muta-tions, �6G>A [Hirosawa et al., 1990] and +13A>G[Crossley and Brownlee, 1990], in the factor IX (F9;MIM# 306900) gene associated with hemophilia BLeyden in humans. Both mutations weaken theplasma factor IX concentrations less than 10% inchildren and between 40% and 80% in adults withrespect to its norm [Crossley and Brownlee, 1990]. Asseen in Table 2, this clinically observed decrease inbinding of the mutant promoter to unknown TFs canbe empirically described by the intermediate degree0.5 against the maximal +1 degree pre-defined innorm. Following these data, rSNP_Guide predicts that
FIGURE 4. Analysis of theTDO2 gene by rSNP Guide. A:Gel mobility shift assay with the liver nuclear cell extract and oligo-DNAs, WT, 50-cagttgccaaataatgGcaGataagaatagggag-30; M1, 50-cagttgccaaataatgAcaGataagaatagggag-30; M2, 50-cagttgc-caaataatgGcaTataagaatagggag-30. Arrow, ¢ve unknown DNA/TF-complexes; two broken arrows, two complexes insensitiveto mutations; three solid arrows, three complexes sensitive to mutation; bold-faced solid arrow, theWT-speci¢c complex. B:The rSNP Guide prediction, theYY1 site typical only for the wild-type oligoDNA. C:The supershift assay with the anti-YY1antibody, liver nuclear extract, and oligoDNAWT. Lane I, without anti-YY1 antibody, lane II, with addition of anti-YY1 anti-body. As seen, in consistencewith the rSNP Guideprediction, theDNA/YY1-complex (solid arrow) is visible only in the lane I,whereas the low-mobility complex of DNA/YY1-anti-YY1-antibody (dotted arrow) is seen only in the lane II.
TABLE 2. Relationships Between the SNP-referredAlterations ofGenome Sequence andDisease: rSNP GuideAnalysis
SNPs DNA/TF-complex Prediction
Disease Gene Allele DNA sequenceDisease
penetration Detected Score(X) TF-site dTF,XControl tests
experiment [reference]
Mouse K-ras, 288C,296A 50-aCtccacttAt-30 Susceptible Present 1.0 GATA 0,01 Anti-GATA antibodyLung tumor intron II 288C,296C 50-aCtccacttCt-30 Intermediate Undetected 0.0 [Ponomarenko et al., 2002a]
288G,296C 50-aGtccacttCt-30 Resistant Undetected 0.0
Human F9, �6G 50-taatcGacctt-30 Normal Present 1.0 C/EBPa 0,11 CotransfectionHemophilia B promoter �6A 50-taatcAacctt-30 Defect Weakened 0.5 [Hirosawa et al.,1990]Leyden form silencers +13A 50-tcacaAtctg-30 Normal Present 1.0 C/EBPb 0,04 Cotransfection
+13G 50-tcacaGtctg-30 Defect Weakened 0.5 [Crossley andBrownlee,1990]
Hereditary HumanHBG1 �175T 50-gcaaaTatctg-30 Normal Present 1.0 GATA 0,03 GATA-core competitorpersistence of (gA-globin), �175C 50-gcaaaCatctg-30 Defect Increased 1.5 [O’Neill et al.,1990]fetal haemog- promoter �175T 50-gcaaaTatctg-30 Normal Present 1.0 OCT 0,09 OCT-core competitorlobin (HPFH) (2 sites) �175C 50-gcaaaCatctg-30 Defect Weakened 0.5 [O’Neill et al.,1990]
DATABASE FOR SNPSANDSITE-DIRECTEDMUTATIONS 245

the C/EBPa site is damaged by mutation �6G>A,whereas the C/EBPb site is disrupted by +13A>Gmutation. Both these predictions are consistent withco-transfection tests [Hirosawa et al., 1990; Crossleyand Brownlee, 1990].
In the third example, we consider the hereditarypersistence of fetal hemoglobin (HPFH)-associatedmutation, �175T>C, in the human gA-globinpromoter (HBG1; MIM# 142200). This mutationhas altered two overlapping TF sites by increasing theGATA specificity and decreasing the OCT specificity,resulting in two- to three-fold enhancement of gA-globin gene expression [Mantovani et al., 1988;O’Neill et al., 1990]. Table 2 illustrates how thesedata were entered into the rSNP_Guide system. Thenormal �175T allele is assigned by the maximal +1degree. With respect to this norm, the mutant �175Callele is assigned by the intermediate 0.5 degree, andfor erythroid-specific TF, a novel abnormally high rate,+1.5 degree, was used. As can be seen in Table 2, for�175T>C mutation, rSNP_Guide predicts that theubiquitous OCT site is damaged and substituted bythe erythroid specific GATA site, these data being inaccordance with the control competitor assays[O’Neill et al., 1990].
Site-Directed Mutagenesis Analysis
In addition to disease-associated mutations, 18 datasets of site-directed mutagenesis were also treated byrSNP_Guide. Three examples of this analysis areshown in Table 3.
First, the �921TT>CC mutation in the mousenitric oxide synthase (iNOS) promoter makes thewild-type specific DNA/TF-complex undetectable bythe EMSA with RAW 264.7 cell nuclear extract[Martin et al., 1994]. This complex, lost due tomutation, was assigned by the threshold 0 degree withrespect to the maximum +1 degree typical for thewild-type DNA (Table 3). As seen in Table 3, therSNP_Guide predicts the presence of the wild-typeIRF-1 site, DNA/protein-binding of which decreases
with �921TT>CC mutation, this evidence beingconsistent with the experiment demonstrating elim-ination of this complex by the anti-IRF-1 immuneserum assay [Martin et al., 1994].For more complex design of mutagenesis, we
consider analysis of the p53 promoter by therSNP_Guide. The wild-type DNA/TF-complexesare detectable by EMSA with HeLa cell nuclearextract and a 20-bp fragment from +49 to 68 of p53sequence (see Table 3). Notably, these complexes areeliminated by the double {+56GGG>ATC;+64CCC>AAG} mutation [Wu and Lozano,1994]. By defining the wild-type expressionpattern as +1 degree and the mutant one as 0, wearrive at prediction that NF-kB is a potentialtranscription factor here. This prediction is inagreement with the anti-NF-kB antibody assay[Wu and Lozano, 1994].The final example given in Table 3 represents the
quantitative alterations in the human hsp70 genepromoter (HSPA1A; MIM# 140550) with singlenucleotide substitutions, �113A>G, �103A>G,�101T>A, and �98T>C. These mutations alterthe binding measured by the CAT-assay to unknownTF [Tsutsumi-Ishii et al., 1995]. Following theseexperimental data, rSNP_Guide predicts that the HSFsite suits the best in this case. This prediction isconsistent with the control test experiment on heatshock induction followed by the CAT-assay [Tsutsu-mi-Ishii et al., 1995].In total, the accuracy of prediction made by the
rSNP_Guide system has been verified on 33 controldata sets representing 15 TF sites with 21 mutationsassociated with diseases and 18 TF sites identified by28 site-directed artificial mutations. In comparison tothe MATRIX system that was previously described[Ponomarenko et al., 1999] and that gives almost50% of false-positive estimates, the applicationof the rSNP_Guide enables the user to discard thisoverestimation completely. This advantage makesthe rSNP_Guide system very useful for practicalapplication.
TABLE 3. Experimental Data on Site-directedMutagenesis Studied byUsing rSNP Guide
Site-directedmutagenesis DNA/TF-complex Prediction
Gene DNAvariant DNA sequence Detected Score(X) TF-site dTF,XControl experimental
tests [reference]
Mouse iNOS, Wild-type (EMBL: L09126) 50-caatatTTcactttc-30 Present 1.0 IRF-1 0.04 Anti-IRF-1antibodypromoter �921TT4CC 50-caatatCCcactttc-30 Undetected 0.0 [Martin et al.,1994]
Mouse p53, Wild-type (EMBL: M26862) 50-ttGGGactttCCCct-30 Present 1.0 NF-kB 0,12 Anti-NF-kB antibodypromoter +56GGG4ATC, +64CCC4AAG 50-ttATCactttAAGct-30 Undetected 0.0 [Wu and Lozano,1994]
Wild-type (EMBL: M11717) 50-gAaacccctggAaTatTc-30 Present 1.0 Heat-shock inductionHuman hsp70 �113A4G 50-gGaacccctggaatattc-30 Weakened 0.4 for the CAT-assay(HSPA1A), �103A4G 50-gaaacccctggGatattc-30 Weakened 0.1 HSF 0,08 [Tsutsumi-Ishii et al.,1995]promoter �101T4A 50-gaaacccctggaaAattc-30 Weakened 0.9
�98T4C 50-gaaacccctggaatatCc-30 Weakened 0.6
246 PONOMARENKO ETAL.

FUTUREGOALSAND PERSPECTIVES
Analysis of SNPs in regulatory gene regions by thecurrent release of rSNP_Guide is limited by thesearching for potential TF-candidates out of 41 knownTF-sites. With this limitation in mind, we intend toincrease the number of TFs used for analysis. In thefuture, we are going to enlarge the contents of theSAMPLES database and, hence, the number of weightmatrices for TF-site recognition implemented in theMATRIX database.
In our recent publication [Ponomarenko et al.,2002b], we have reported that the false-positive errorcould be essentially reduced by coordinated usage ofmultiple-tools aimed at TF site recognition. For thispurpose, we are going to involve some other database-tools systems: SELEX_DB, on in vitro-selected DNAsbinding to TF-targets [Ponomarenko et al., 2002b];and ACTIVITY, on quantitative sequence-activityrelationships [Ponomarenko et al., 2001a], intoanalysis of SNPs and mutagenesis.
Please contact to our database-tools system admin-istrator, Mrs. Julia Ponomarenko, regarding allrSNP_Guide applications, comments, corrections,requests, and contributions to the database (E-mail:[email protected]; Phone: +7(3832)333119; Fax:+7(3832)331278).
REFERENCES
Beason KB, Acuff CG, Steinhelper ME, Elton TS. 1999. AnA/T-rich cis-element is essential for rat angiotensin II type1A receptor transcription in vascular smooth muscle cells.Biochim Biophys Acta 1444:25–34.
Bienvenu T, Lacronique V, Raymondjean M, Cazeneuve C,Hubert D, Kaplan J, Beldjord C. 1995. Three novel sequencevariations in the 50 upstream region of the cystic fibrosistransmembrane conductance regulator (CFTR) gene: twopolymorphisms and one putative molecular defect. HumGenet 95:698–702.
Boccia LM, Lillicrap D, Newcombe K, Mueller CR. 1996.Binding of the Ets factor GA-binding protein to an upstreamsite in the factor 9 promoter is a critical event intransactivation. Mol Cell Biol 16:1929–1935.
Brookes A, Lehvaeslaiho H, Siegfried M, Boem J, Yuan Y,Sarkar C, Bork P, Ortigao F. 2000. HGBASE: a database ofSNPs and other variations in and around human genes.Nucleic Acids Res 28:356–360.
Chen B, Johanson L, Wiest J, Anderson M, You M. 1994. Thesecond intron of the K-ras gene contains regulatory elementsassociated with mouse lung tumor susceptibility. Proc NatlAcad Sci USA 91:1589–1593.
Comings DE, Gade R, Muhleman D, Chiu C, Wu S, To M,Spence M, Dietz G, Winn-Deen E, Rosenthal RJ, LesieurHR, Rugle L, Sverd J, Ferry L, Johnson JP, MacMurray JP.1996. Exon and intron variants in the human tryptophan2,3-dioxygenase gene: potential association with Tourettesyndrome, substance abuse and other disorders. Pharmaco-genetics 6:307–318.
Cowell JK, Bia B, Akoulitchev A. 1996. A novel mutation in
the promotor region in a family with a mild form ofretinoblastoma indicates the location of a new regulatory
domain for the RB1 gene. Oncogene 12:431–436.Crossley M, Brownlee GG. 1990. Disruption of a C/EBP
binding site in the factor IX promoter is associated with
haemophilia B. Nature. 345:444–446.Gall J, Bellini M, Wu Z, Murphy C. 1999. Assembly of the
nuclear transcription and processing machinery: Cajal bodies
(coiled bodies) and transcriptosomes. Mol Biol Cell 10:4385–4402.
Gillio AP, Verlander PC, Batish SD, Giampietro PF, Auerbach
AD. 1997. Phenotypic consequences of mutations in the
Fanconi anemia FAC gene: an international fanconi anemia
registry study. Blood 90:105–110.Hamosh A, Scott AF, Amberger J, Valle D, McKusick VA.
2000. Online Mendelian inheritance in man (OMIM). HumMutat 15:57–61.
Hirosawa S, Fahner JB, Salier JP, Wu CT, Lovrien EW,
Kurachi K. 1990. Structural and functional basis of the
developmental regulation of human coagulation factor IX
gene: factor IX Leyden. Proc Natl Acad Sci USA 87:4421–4425.
Knight JC, Udalova I, Hill AV, Greenwood BM, Peshu N,Marsh K, Kwiatkowski D. 1999. A polymorphism that affects
OCT-1 binding to the TNF promoter region is associated
with severe malaria. Nat Genet 22:145–150.Krawczak M, Ball E, Fenton I, Stenson PD, Abeysinghe S,
Thomas N, Cooper DN. 2000. Human gene mutationdatabase—a biomedical information and research resource.
Hum Mutat 15:45–51.Langdon S, Kaufman R. 1998. Gamma-globin gene promoter
elements required for interaction with globin enhancers.
Blood 91:309–318.Mantovani R, Malgaretti N, Nicolis S, Ronchi A, Giglioni B,
Ottolenghi S. 1988. The effects of HPFH mutations in thehuman gamma-globin promoter on binding of ubiquitous
and erythroid specific nuclear factors. Nucleic Acids Res
16:7783–7797.Marth G, Yeh R, Minton M, Donaldson R, Li Q, Duan S,
Davenport R, Miller R, Kwok P-Y. 2001. Single-nucleotidepolymorphisms in the public domain: how useful are they?
Nat Genet 27:371–372.Martin E, Nathan C, Xie QW. 1994. Role of interferon
regulatory factor 1 in induction of nitric oxide synthase.
J Exp Med 180:977–984.Matsuda M, Sakamoto N, Fukumaki Y. 1992. Delta-thalasse-
mia caused by disruption of the site for an erythroid-specifictranscription factor, GATA-1, in the delta-globin gene
promoter. Blood 80:1347–1351.Moi P, Loudianos G, Lavinha J, Murru S, Cossu P, Casu R,
Oggiano L, Longinotti M, Cao A, Pirastu M. 1992. Delta-
thalassemia due to a mutation in an erythroid-specificbinding protein sequence 30 to the delta-globin gene. Blood
79:512–516.Nedelcheva Kristensen V, Haraldsen EK, Anderson KB,
Lonning PE, Erikstein B, Karesen R, Gabrielsen OS,
Borresen-Dale AL. 1999. CYP17 and breast cancer risk:the polymorphism in the 50 flanking area of the gene does not
influence binding to Sp-1. Cancer Res 59:2825–2828.
DATABASE FOR SNPSANDSITE-DIRECTEDMUTATIONS 247

O’Neill D, Kaysen J, Donovan-Peluso M, Castle M, Bank A.1990. Protein-DNA interactions upstream from the humanA gamma globin gene. Nucleic Acids Res 18:1977–1982.
Osier M, Cheung K-H, Kidd J, Pakstis A, Miller P, Kidd K.2001. ALFRED: an allele frequency database for diversepopulations and DNA polymorphisms: an update. NucleicAcids Res 29:317–319.
Piedrafita FJ, Molander RB, Vansant G, Orlova EA, Pfahl M,Reynolds WF. 1996. An Alu element in the myeloperoxidasepromoter contains a composite SP1-thyroid hormone-retinoic acid response element. J Biol Chem 271:14412–14420.
Ponomarenko M, Ponomarenko J, Frolov A, Podkolodnaya O,Vorobyev DG, Kolchanov NA, Overton GC. 1999. Oligo-nucleotide frequency matrices addressed to recognizingfunctional DNA signals. Bioinformatics 15:631–643.
Ponomarenko J, Furman D, Frolov A, Podkolodny N, Orlova G,Ponomarenko M, Kolchanov N, Sarai A. 2001a. ACTIVITY:a database on DNA/RNA sites activity adapted to applysequence-activity relationships from one system to another.Nucleic Acids Res 29:284–287.
Ponomarenko J, Merkulova T, Vasiliev G, Levashova Z, OrlovaG, Lavryushev S, Fokin O, Ponomarenko M, Frolov A, SaraiA. 2001b. rSNP_Guide, a database system for analysis oftranscription factor binding to target sequences: applicationto SNPs and signal-directed mutations. Nucleic Acids Res29:312–316.
Ponomarenko J, Merkulova T, Orlova G, Fokin O, GorshkovaE, Ponomarenko M. 2002a. Mining DNA sequences topredict sites which mutations cause genetic diseases. Knowl-edge-Based Systems 15:225–233.
Ponomarenko J, Orlova G, Frolov A, Gelfand M, PonomarenkoM. 2002b. SELEX_DB: a database on in vitro selectedoligomers adapted for recognizing natural sites and foranalyzing both SNPs and site-directed mutagenesis data.Nucleic Acids Res 30:195–199.
Quandt K, Frech K, Karas H, Wingender E, Werner T. 1995.MatInd and MatInspector: new fast and sensitive tools fordetection of consensus matches in nucleotide sequence data.Nucleic Acids Res 23:4878–4884.
Roulet E, Fisch I, Bucher P, Mermod N. 1998. Evaluation ofcomputer tools for prediction of transcription factor bindingsites on genomic DNA. In Silico Biol 1:21–28.
Roulet E, Bucher P, Schneider R, Wingender E, Dusserre Y,Werner T, Mermod N. 1999. Experimental analysis andcomputer prediction of CTF/NFI transcription factor DNAbinding sites. J Mol Biol 297:833–848.
Ryan J, Barker P, Nesbitt M, Ruddle F. 1987. KRAS2 as agenetic marker for lung tumor susceptibility in inbred mice. JNatl Cancer Inst 79:1351–1357.
Sachidanandam R, Weissman D, Schmidt SC, Kakol JM, SteinLD, Marth G, Sherry S, Mullikin JC, Mortimore BJ, WilleyDL, Hunt SE, Cole CG, Coggill PC, Rice CM, Ning Z,Rogers J, Bentley DR, Kwok P-Y, Mardis ER, Yeh RT, SchultzB, Cook L, Davenport R, Dante M, Fulton L, Hillier L,Waterston RH, McPherson JD, Gilman B, Schaffner S, vanEtten WJ, Reich D, Higgins J, Daly MJ, Blumenstiel B,Baldwin J, Stange-Thomann N, Zody MC, Linton L, LanderES, Atshuler D, The International SNP Map Working Group.2001. A map of human genome sequence variation contain-ing 1,42 million single nucleotide polymorphisms. Nature409:928–933.
Sherry S, Ward M, Kholodov M, Baker J, Phan L, SmigielskiEM, Sirotkin K. 2001. dbSNP: the NCBI database of geneticvariation. Nucleic Acids Res 29:308–311.
Stoeckert C Jr, Salas F, Brunk B, Overton C. 1999. EpoDB: aprototype database for the analysis of genes expressed duringvertebrate erythropoiesis. Nucleic Acids Res 27:200–203.
Tsutsumi-Ishii Y, Tadokoro K, Hanaoka F, Tsuchida N.1995. Response of heat shock element within ofhuman HSP70 promoter of mutated p53 genes. Cell GrowthDiffer 6:1–8.
Vasiliev G, Merkulov V, Kobzev V, Merkulova T, PonomarenkoM, Kolchanov N. 1999. Point mutations within 663–666 bpof intron 6 of the human TDO2 gene, associated with anumber of psychiatric disorders, damage the YY-1 transcrip-tion factor binding signal. FEBS Lett 462:85–88.
Wu H, Lozano G. 1994. NF-kappa B activation of p53. Apotential mechanism for suppressing cell growth in responseto stress. J Biol Chem 269:20067–20074.
248 PONOMARENKO ETAL.
Related Documents











