RESEARCH PAPER Rotifer species richness along an altitudinal gradient in the AlpsUlrike Obertegger 1 *, Bertha Thaler 2 and Giovanna Flaim 1 1 IASMA Research and Innovation Centre, FEM, Environment and Natural Resources Area, I-38010 San Michele all’Adige (TN), Italy, 2 Provincial Environmental Protection Agency, Bozen, I-39055 Leifers (BZ), Italy ABSTRACT Aim Biodiversity patterns along altitudinal gradients are less studied in aquatic than terrestrial systems, even though aquatic sites provide a more homogeneous environment independent of moisture constraints. We studied the altitudinal species richness pattern for planktonic rotifers in freshwater lakes and identified the environmental predictors for which altitude is a proxy. Location Two hundred and eighteen lakes of Trentino–South Tyrol (Italy) in the eastern Alps; lakes covered 98% (range 65–2960 m above sea level) of the altitudinal gradient in the Alps. Methods We performed: (1) linear regression between species richness and alti- tude to evaluate the general pattern, (2) multiple linear regression between species richness and environmental predictors excluding altitude to identify the most important predictors, and (3) linear regression between the residuals of the best model of step (2) and altitude to investigate any additional explanatory power of altitude. Selection of environmental predictors was based on limnological impor- tance and non-parametric Spearman correlations. We applied ordinary least squares regression, generalized linear, and generalized least squares modelling to select the most statistically appropriate model. Results Rotifer species richness showed a monotonic decrease with altitude inde- pendent of scale effects. Species richness could be explained (R 2 = 51%) by lake area as a proxy for habitat diversity, reactive silica and total phosphorus as proxies for productivity, water temperature as a proxy for energy, nitrate as a proxy for human influence and north–south and east–west directions as covariates. These predictors completely accounted for the species richness–altitude pattern, and altitude had no additional effect on species richness. Main conclusions The linear decrease of species richness along the altitudinal gradient was related to the interplay of habitat diversity, productivity, heat content and human influence. These factors are the same in terrestrial and aquatic habitats, but the greater environmental stability of aquatic systems seems to favour a linear pattern. Keywords Alps, biodiversity, habitat diversity, Italy, lakes, productivity, regression analysis, rotifers, temperature, zooplankton. *Correspondence: Ulrike Obertegger, IASMA, Research and Innovation Centre, FEM, Environment and Natural Resources Area, Via Edmund Mach, 1, I-38010 San Michele all’Adige (TN), Italy. E-mail: [email protected] INTRODUCTION Understanding the factors that govern species richness is a fun- damental issue for ecologists regardless of the habitat studied, especially as ecosystem functioning is related to biodiversity. Studies that address this topic are particularly important given current concerns about loss of biodiversity with global climate change (Nyman et al., 2005). To comprehend the mechanisms that determine biodiversity patterns, species richness along spatial gradients such as latitude and altitude are increasingly being studied (e.g. Nyman et al., 2005; Hessen et al., 2007). In particular, focus has been put on altitudinal gradients because Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2010) 19, 895–904 © 2010 Blackwell Publishing Ltd DOI: 10.1111/j.1466-8238.2010.00556.x www.blackwellpublishing.com/geb 895

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCHPAPER
Rotifer species richness along analtitudinal gradient in the Alpsgeb_556 895..904
Ulrike Obertegger1*, Bertha Thaler2 and Giovanna Flaim1
1IASMA Research and Innovation Centre,
FEM, Environment and Natural Resources
Area, I-38010 San Michele all’Adige (TN),
Italy, 2Provincial Environmental Protection
Agency, Bozen, I-39055 Leifers (BZ), Italy
ABSTRACT
Aim Biodiversity patterns along altitudinal gradients are less studied in aquaticthan terrestrial systems, even though aquatic sites provide a more homogeneousenvironment independent of moisture constraints. We studied the altitudinalspecies richness pattern for planktonic rotifers in freshwater lakes and identified theenvironmental predictors for which altitude is a proxy.
Location Two hundred and eighteen lakes of Trentino–South Tyrol (Italy) in theeastern Alps; lakes covered 98% (range 65–2960 m above sea level) of the altitudinalgradient in the Alps.
Methods We performed: (1) linear regression between species richness and alti-tude to evaluate the general pattern, (2) multiple linear regression between speciesrichness and environmental predictors excluding altitude to identify the mostimportant predictors, and (3) linear regression between the residuals of the bestmodel of step (2) and altitude to investigate any additional explanatory power ofaltitude. Selection of environmental predictors was based on limnological impor-tance and non-parametric Spearman correlations. We applied ordinary leastsquares regression, generalized linear, and generalized least squares modelling toselect the most statistically appropriate model.
Results Rotifer species richness showed a monotonic decrease with altitude inde-pendent of scale effects. Species richness could be explained (R2 = 51%) by lake areaas a proxy for habitat diversity, reactive silica and total phosphorus as proxies forproductivity, water temperature as a proxy for energy, nitrate as a proxy for humaninfluence and north–south and east–west directions as covariates. These predictorscompletely accounted for the species richness–altitude pattern, and altitude had noadditional effect on species richness.
Main conclusions The linear decrease of species richness along the altitudinalgradient was related to the interplay of habitat diversity, productivity, heat contentand human influence. These factors are the same in terrestrial and aquatic habitats,but the greater environmental stability of aquatic systems seems to favour a linearpattern.
KeywordsAlps, biodiversity, habitat diversity, Italy, lakes, productivity, regressionanalysis, rotifers, temperature, zooplankton.
*Correspondence: Ulrike Obertegger, IASMA,Research and Innovation Centre, FEM,Environment and Natural Resources Area, ViaEdmund Mach, 1, I-38010 San Micheleall’Adige (TN), Italy.E-mail: [email protected]
INTRODUCTION
Understanding the factors that govern species richness is a fun-
damental issue for ecologists regardless of the habitat studied,
especially as ecosystem functioning is related to biodiversity.
Studies that address this topic are particularly important given
current concerns about loss of biodiversity with global climate
change (Nyman et al., 2005). To comprehend the mechanisms
that determine biodiversity patterns, species richness along
spatial gradients such as latitude and altitude are increasingly
being studied (e.g. Nyman et al., 2005; Hessen et al., 2007). In
particular, focus has been put on altitudinal gradients because
Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2010) 19, 895–904
© 2010 Blackwell Publishing Ltd DOI: 10.1111/j.1466-8238.2010.00556.xwww.blackwellpublishing.com/geb 895

they offer more constant ecological conditions and history with
respect to other spatial gradients while at the same time are
linked to several environmental variables giving beneficial
insights for both theoretical and applied research on biodiversity
(Rowe, 2009). The species richness–altitude relationship gener-
ally shows two main patterns (i.e. decreasing or unimodal)
depending on the main attributes of scale (i.e. the unit of sam-
pling and the geographical space covered) (Rahbek, 2005). Spe-
cifically, the exclusion of altitudes lower than 500 m or higher
than 2000 m above sea level (a.s.l.) leads to a biased species
richness–altitude relationship (Rahbek, 2005; Nogués-Bravo
et al., 2008). Furthermore, the different extent of human impact
at different altitudes is decisive for observing different altitudi-
nal patterns of species richness in nature (Nogués-Bravo et al.,
2008).
Besides the need for a comprehensive altitudinal gradient,
terrestrial studies on altitudinal processes are often con-
founded by covariance between altitude and regional peculiari-
ties, moisture constraints (Körner, 2007) and temperature
fluctuations (Reynolds, 1998). On the contrary, the more
homogeneous character of aquatic ecosystems (e.g. marine,
Smith & Brown, 2002; freshwater, Hessen et al., 2007) offers
the possibility to investigate altitudinal gradients independent
of such terrestrial peculiarities. In contrast to the connectivity
of terrestrial and marine habitats, however, inland waters have
well-defined boundaries, are isolated and are surrounded by an
unsuitable landscape leading to a patchy environment (Rey-
nolds, 1998; Jankowski & Weyhenmeyer, 2006; Scheffer & van
Geest, 2006). Lakes are also unique because of their internal
physical gradients such as light, temperature and oxygen that
structure plankton communities. Aquatic organisms, because
of characteristics such as short life cycles, reduced size struc-
tures and efficient resource uptake, generally show faster
responses to environmental changes than terrestrial organisms,
corroborating the viewpoint that freshwater studies can con-
tribute much to the general understanding of ecosystem pro-
cesses (Reynolds, 1998).
However, most studies on altitudinal gradients focus on ter-
restrial species (Rahbek, 1995, 2005; Currie & Kerr, 2008;
Nogués-Bravo et al. 2008 and references therein), and four
main groups of hypothesis have been identified to explain
species richness–altitude patterns. The climatic hypothesis
considers temperature and humidity (e.g. Kluge et al., 2006;
McCain, 2007) and energy and ecosystem productivity (e.g.
Davies et al., 2007), the spatial hypothesis considers area (e.g.
Williams, 1964; MacArthur & Wilson, 1967), randomness and
neutral theories (e.g. Hubbell, 2001) and spatial constraints
(e.g. Colwell & Lees, 2000), the historical hypothesis considers
historical and evolutionary processes (e.g. Wiens & Donoghue,
2004) and the biotic hypothesis considers source–sink dynam-
ics (e.g. Grytnes et al., 2006) as the main drivers for the
observed patterns.
Less is known about altitudinal gradients in aquatic ecosys-
tems (Bêche & Statzner, 2009 and references therein); the species
richness–altitude relationship has been studied for only a few
aquatic taxa: phytoplankton (local species richness, Jankowski &
Weyhenmeyer, 2006), aquatic plants (Jones et al., 2003), crusta-
ceans (Hessen et al., 2007), stream macroinvertebrates (local
species richness, Jacobsen, 2004) and molluscs (Sturm, 2007)
show a linear decrease, while stream macroinvertebrates (altitu-
dinal bands, Jacobsen, 2004), chironomids (Nyman et al., 2005),
phytoplankton (altitudinal bands, Jankowski & Weyhenmeyer,
2006) and fish (Li et al., 2009) show a hump-shaped pattern
with altitude. Some of these nonlinear patterns can be attributed
to rescue effects that maintain the presence of species in unsuit-
able habitats by continuous colonization from suitable habitats
(Jacobsen, 2004) and investigating species richness in an
ecotonal transitional zone (Nyman et al., 2005) probably subject
to source–sink effects that result in species flow from optimal to
suboptimal sites. Both of these similar effects can inflate the
assessment of species richness (Jones et al., 2003; Grytnes &
McCain, 2007). In aquatic habitats, hypotheses on the species
richness–altitude pattern rely on area and climatic variables
(Jones et al., 2003; Li et al., 2009), geometric constraints, dis-
persal, environmental heterogeneity, productivity (Jankowski &
Weyhenmeyer, 2006; Hessen et al., 2007; Li et al., 2009) and
temperature (Nyman et al., 2005; Hessen et al., 2007).
In this paper, we focus on the mechanisms shaping species
richness in lakes over a complete altitudinal gradient in the
Alps. The Alps in particular are an especially interesting study
site because they provide broad ecological gradients, can serve
as a template for the many mountainous regions world-wide
and are a hotspot of biodiversity in Europe. We focused on
planktonic monogonont rotifers as a general model to study
biodiversity because they are: (1) widely distributed, (2) an
important component of the pelagic food web, and (3) readily
susceptible to changes in their environment (Reynolds, 1998;
Lampert & Sommer, 2007). In addition, a comprehensive study
on rotifer richness in relation to altitude is lacking, and only
ancillary information is available: monogonont rotifer richness
in high-altitude lakes decreases with altitude (Jersabek, 1995)
and Synchaeta species show a preference for lakes of specific
altitudinal belts (Obertegger et al., 2008). Our aims were
twofold: (1) investigate the pattern of the rotifer richness–
altitude relationship (linear or hump-shaped) and ascertain its
consistency across altitudinal bands, and (2) identify the envi-
ronmental predictors for which altitude is a proxy. We selected
variables related to habitat diversity, productivity, energy and
human influence covering the different factors outlined in ter-
restrial as well as freshwater studies to develop a holistic model
of the species richness–altitude relationship in lakes. While the
outcome of the first issue is of a more theoretical nature
necessary for the development of a unifying theory on
biodiversity, the outcome of the latter issue is relevant for
a mechanistic understanding of the environment and species
richness patterns sensu Gotelli et al. (2009). Our dataset, with
a wide range of predictors representing important environ-
mental gradients, offers a unique opportunity to study these
interconnected research questions. In addition, we provide a
better understanding of the processes behind the richness–
altitude relationship, an issue that is still controversial (Rowe,
2009).
U. Obertegger et al.
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd896

MATERIALS AND METHODS
Study sites and data acquisition
We studied rotifer species richness in relation to altitude in 218
lakes in the Trentino–South Tyrol region in the eastern Alps
(Italy), sampled during the summers of 1996–2008 (Fig. 1).
These lakes ranged from 65 to 2960 m a.s.l. with an average of
four lakes every 100 m, and covered 98.3% of the Alpine altitu-
dinal gradient (value according to Körner, 2007). The lakes
sampled spanned a wide range of environmental variables
(Table 1) and reflected the altitudinal distribution of lakes in the
study area (Fig. 2). The range of the north–south and east–west
direction was limited to less than 2°; therefore these spatial
parameters were not considered as gradients, but were impor-
tant for accounting for spatial autocorrelation (see statistical
modelling below). Species lists and environmental data were
either based on published data (45 lakes: IASMA, 1996–2000;
Salmaso & Naselli-Flores, 1999; Cantonati et al., 2002; Cantonati
& Lazzara, 2006) or provided directly by the authors. Zooplank-
ton samples were generally taken from the deepest part of the
lake with a plankton net (50 mm) and were preserved in 4%
formalin or 20% alcohol. The coordinates of the lakes’ centroids
were available on three different coordinate systems: the Univer-
sal Transverse Mercator (UTM) 32N/WGS84 (South Tyrol), the
Gauss Boaga/Rome40 W (Trentino), and the lat/long/WGS84
(Trentino); all the geographic positions were harmonized in the
lat/long/WGS84 system applying a datum-shift correction using
seven parameters specific to the study area.
Data treatment and statistical modelling
Species richness on a continuous altitudinal scale was calculated
as the number of rotifer taxa found in each lake. Species richness
at different altitudinal resolutions was calculated as the mean
Figure 1 Location of the 218 lakes sampled in Trentino–SouthTyrol (Italy). Continuous lines represent major rivers, filled circlesrepresent sampling sites.
Figure 2 Altitudinal distribution of lakes in Trentino–SouthTyrol for 100 m intervals. Black bars are mapped lakes (n = 667)and white bars are sampled lakes (n = 218).
Table 1 Minimum (min), maximum(max), median and mean values for themajor predictors of lakes sampled (n =218).
Parameter Min Max Median Mean
Altitude (m) 65 2960 2245 1954
Area (ha) < 1 36,800 2 185
Depth (m) 1 350 6 12
TP (mg l-1) 1 220 9 15
N-NO3 (mg l-1) 6 2165 174 232
N:P < 1 1017 40 68
Si (mg l-1) 0.1 10.7 1.3 1.7
SO4 (mg l-1) 0.2 94.9 3.6 8.7
Conductivity (mS cm-1) 4 504 45 98
Temperature (°C) 2.0 26.7 12.0 12.6
Latitude (N) 45°39′59″ 46°56′47″ 46°32′12″ 46°30′11″Longitude (E) 10°23′12″ 12°20′43″ 11°13′20″ 11°16′54″
Area is lake surface area, temperature is surface water temperature, TP is total phosphorus, and Si isreactive silica.
Rotifer species richness in the Alps
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd 897
value of species richness for lakes within bands of different
altitudinal widths (100 m, 200 m, 300 m and 400 m band
widths) over the 3000-m range. In terrestrial studies, standard-
ization of species richness by sampling area is recommended
(Rahbek, 1995), because area decreases with altitude. In this
study, however, sampling area was the pelagic zone and was
equally sampled in all lakes; therefore, we did not consider any
standardization.
Environmental predictors were log-transformed to account
for their non-Gaussian distribution; species richness data,
instead, were square-root transformed in the case of ordinary
least squares (OLS) modelling and generalized least squares
(GLS) modelling but not in the case of generalized linear mod-
elling (GLM). We performed a three-step analysis: first, we per-
formed an OLS regression to investigate the species richness–
altitude relationship. Because of spatial dependence of residuals,
we then applied a GLS regression by the inclusion of spatial
covariates and a specific term for autocorrelated residuals. Sec-
ondly, we performed a multiple linear regression (GLS model-
ling) to investigate species richness dependent on environmental
predictors excluding altitude. The pre-selection of environmen-
tal predictors out of all variables available for statistical model-
ling was based on limnological importance and non-parametric
Spearman correlations. We applied GLS modelling in the fol-
lowing way: spatial correlation of residuals was investigated and
quantified by a variogram analysis within the framework of
geostatistical models; we compared models of different spatial
correlation structure (linear, Gaussian, spherical, exponential
and rational quadratic) by the Akaike information criterion and
selected that with the lowest value. After including the appro-
priate correlation structure, we selected environmental predic-
tors to find the most parsimonious model; selection was based
on comparison of nested models by an ANOVA test. We also
performed a GLM with quasi-Poisson distribution and included
spatial covariates in the model because in the former GLS it was
difficult to find an appropriate correlation structure to model
spatial dependence. Thirdly, the residuals of the most parsimo-
nious model were regressed on altitude to investigate if altitude
had any additional explanatory power.
For every model, we reported: (1) the partial regression slope
as a measure for changes in a specific predictor while keeping the
other predictors constant, and (2) the relative influence of pre-
dictors according to the formula in Quinn & Keough (2002);
this parameter allowed us to investigate the hierarchy of influ-
ence on the response variable because its value is independent of
the magnitude of different measurement units of predictors.
Multicollinearity of predictors was investigated by the variance
inflation factor (VIF). For OLS models, we reported the stan-
dard R2, while for GLS models and GLM we reported the
pseudo-R2 according to the recommendations of Buse (1973).
All analyses were performed in R (R Development Core Team,
2005).
RESULTS
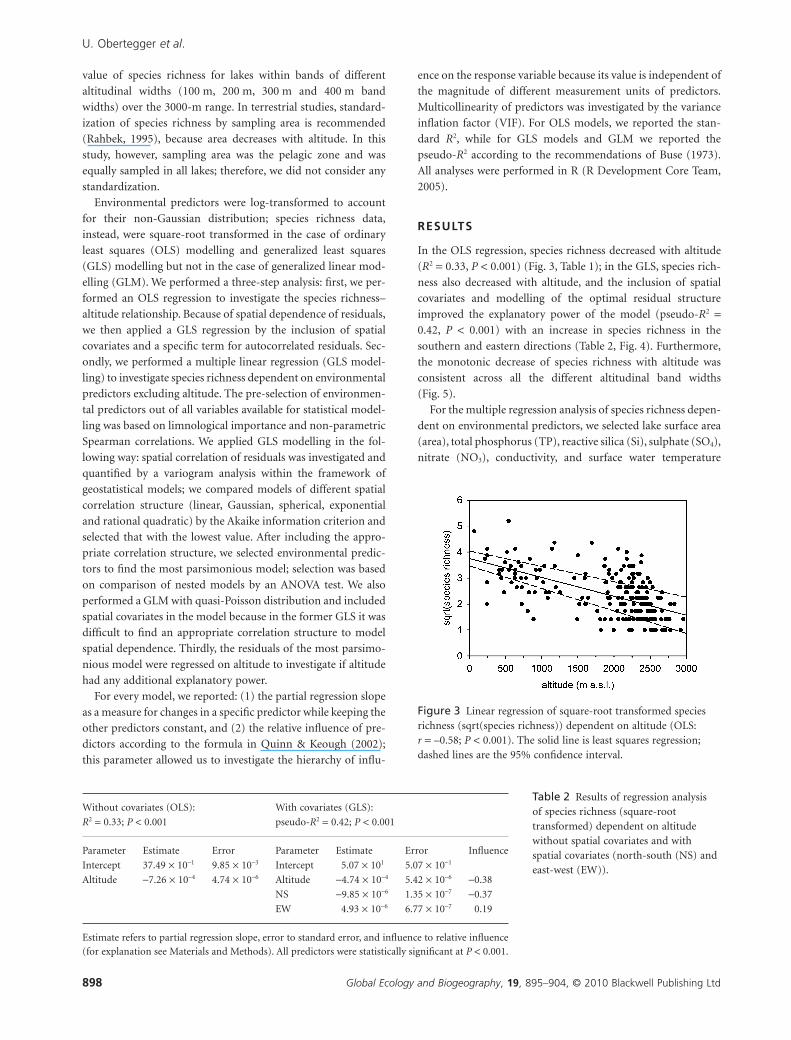
In the OLS regression, species richness decreased with altitude
(R2 = 0.33, P < 0.001) (Fig. 3, Table 1); in the GLS, species rich-
ness also decreased with altitude, and the inclusion of spatial
covariates and modelling of the optimal residual structure
improved the explanatory power of the model (pseudo-R2 =0.42, P < 0.001) with an increase in species richness in the
southern and eastern directions (Table 2, Fig. 4). Furthermore,
the monotonic decrease of species richness with altitude was
consistent across all the different altitudinal band widths
(Fig. 5).
For the multiple regression analysis of species richness depen-
dent on environmental predictors, we selected lake surface area
(area), total phosphorus (TP), reactive silica (Si), sulphate (SO4),
nitrate (NO3), conductivity, and surface water temperature
Figure 3 Linear regression of square-root transformed speciesrichness (sqrt(species richness)) dependent on altitude (OLS:r = –0.58; P < 0.001). The solid line is least squares regression;dashed lines are the 95% confidence interval.
Table 2 Results of regression analysisof species richness (square-roottransformed) dependent on altitudewithout spatial covariates and withspatial covariates (north-south (NS) andeast-west (EW)).
Without covariates (OLS):
R2 = 0.33; P < 0.001
With covariates (GLS):
pseudo-R2 = 0.42; P < 0.001
Parameter Estimate Error Parameter Estimate Error Influence
Intercept 37.49 ¥ 10-1 9.85 ¥ 10-3 Intercept 5.07 ¥ 101 5.07 ¥ 10-1
Altitude -7.26 ¥ 10-4 4.74 ¥ 10-6 Altitude -4.74 ¥ 10-4 5.42 ¥ 10-6 -0.38
NS -9.85 ¥ 10-6 1.35 ¥ 10-7 -0.37
EW 4.93 ¥ 10-6 6.77 ¥ 10-7 0.19
Estimate refers to partial regression slope, error to standard error, and influence to relative influence(for explanation see Materials and Methods). All predictors were statistically significant at P < 0.001.
U. Obertegger et al.
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd898

(temperature) based on limnological importance and non-
parametric Spearman correlations (Table 3). Area was a proxy
for habitat diversity, TP, Si and NO3 for productivity, SO4 for
human influence (atmospheric and terrestrial pollution), con-
ductivity for geology and temperature for the energy content of
water. In the multiple regression of species richness dependent
on environmental predictors, we first performed a GLS model
and considered autocorrelation of residuals; in this model, area,
Si, TP, NO3, SO4, temperature and conductivity were the most
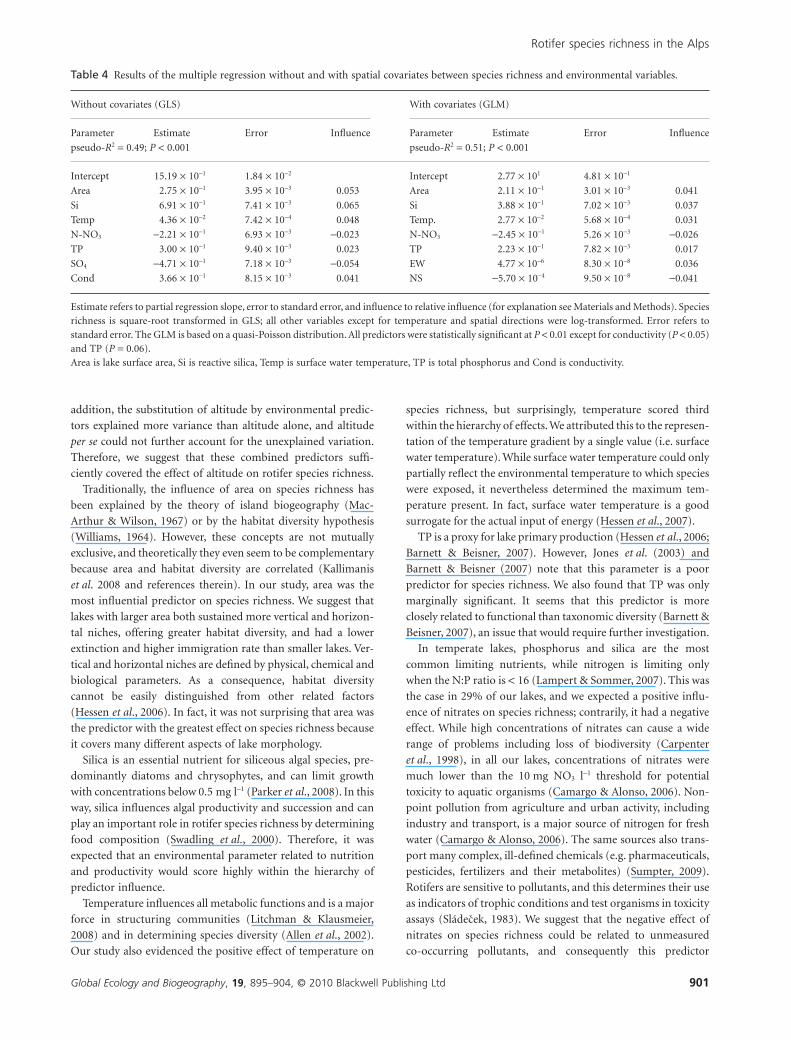
important factors (GLS: pseudo-R2 = 0.49, P < 0.001) (Table 4).
Because species richness as count data can also be modelled by a
Poisson distribution, we subsequently performed a GLM and
included spatial covariates in the model. This new model better
accounted for the spatial structure of data and explained slightly
more variance (GLM: pseudo-R2 = 0.51, P < 0.001), but conduc-
tivity and SO4 were no longer significant and TP was borderline
(Table 4). The VIFs for this model were less than 10 for all
predictors and did not indicate multicollinearity among
variables.
The regression of residuals of the multiple regression models
(with and without spatial covariates) on altitude gave no signifi-
cant relationship (OLS: P > 0.05).
DISCUSSION
The understanding of the species richness–altitude relationship
is crucial for the development of a general theory on species
diversity (Rowe, 2009). In terrestrial studies, a linear decrease of
species richness with altitude is less common than a hump-
shaped pattern (Rahbek, 2005; Grytnes & McCain, 2007). This
pattern is seen as independent of sampling bias, while the linear
richness–altitude relationship is attributed to a decreased sam-
pling effort along altitude or to a truncated altitudinal gradient
(Rahbek, 2005; Nogués-Bravo et al., 2008). In the few aquatic
studies, however, species richness–altitude relationships tend to
show a monotonic decrease (Jones et al., 2003; Jacobsen, 2004;
Fontaneto & Ricci, 2006; Jankowski & Weyhenmeyer, 2006;
Hessen et al., 2007). Besides source–sink and rescue effects that
can inflate the estimate of species richness (Jones et al., 2003;
Grytnes & McCain, 2007), aquatic studies that show nonlinear
species richness–altitude patterns (Nyman et al., 2005; Jan-
kowski & Weyhenmeyer, 2006; Li et al., 2009) may also be biased
by unconsidered broad latitudinal and longitudinal gradients. In
fact, Rahbek (2005) points out that in studies covering extensive
geographical ranges, species richness along altitudinal gradients
is influenced by the different impact of historical and ecological
mechanisms along large latitudinal and longitudinal gradients.
Our study, conducted over a narrow geographical extent,
demonstrated that rotifer species richness decreased linearly
with altitude on a continuous scale. While Rahbek (2005)
underlines that monotonic decreasing patterns are rare when
considering altitudinal bands, in our study a linear pattern also
prevailed within altitudinal bands of different widths. We
suggest that this consistency across different scales further cor-
roborates the goodness of fit of the linear shape. In addition, the
inclusion of spatial covariates in the species richness–altitude
relationship improved the fit of the model (R2 = 0.33 versus
pseudo-R2 = 0.42). In fact, models that include spatial autocor-
relation have a better predictive power than models without it
because autocorrelation accounts for variance in species rich-
ness data (Currie, 2007).
But what does the geographic variable ‘altitude’ actually stand
for in relation to rotifer species richness? Recent research has
shown that altitudinal gradients cannot be attributed to a simple
universal explanation (Rowe, 2009). Based on multivariate
regressions (GLS and GLM), we discussed the importance of
environmental predictors and focused on the GLM because it
was the most appropriate one with respect to spatial dependence
of data. When excluding altitude as an environmental predictor,
we could show that area, Si, temperature and TP had a positive
effect on species richness, while NO3 had a negative one. In
Figure 4 Rotifer species richness in sampled sites: the diameterof closed circles corresponds to different classes of speciesrichness.
Figure 5 Rotifer species richness at different altitudinalresolutions. The range of altitudinal bands differs: (a) 100 mrange; (b) 200 m range; (c) 300 m range; (d) 400 m range (forexplanations see Materials and Methods).
Rotifer species richness in the Alps
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd 899

Tab
le3
Non
para
met
ric
Spea
rman
corr
elat
ion
ofen
viro
nm
enta
lvar
iabl
es:t
he
upp
erse
ctio
ngi
ves
sign
ifica
nce
,th
elo
wer
sect
ion
give
sco
rrel
atio
n;m
issi
ng
valu
esre
fer
ton
on-s
ign
ifica
nt
corr
elat
ion
san
d/or
corr
elat
ion
s<
0.20
.
Alt
iA
rea
Dep
thSR
PT
PN
H4
NO
3D
INSi
SO4
Cl
Ca
Mg
Na
KpH
Alk
Con
dTe
mp
NS
EW
Alt
i**
***
***
***
***
***
***
***
***
***
***
***
***
***
***
***
***
***
Are
a-0
.49
***
***
***
***
***
***
***
***
***
****
***
***
***
***
***
Dep
th-0
.25
0.72
***
***
***
***
***
SRP
-0.4
8**
***
***
***
***
***
***
***
TP
-0.4
60.
50**
***
***
***
***
***
***
***
***
***
***
NH
4-0
.25
0.40
****
***
***
***
***
***
***
***
***
***
NO
3-0
.35
0.41
0.32
0.22
***
***
***
***
***
***
***
****
***
*
DIN
-0.3
60.
420.
330.
290.
99**
***
***
***
***
***
****
***
*
Si-0
.54
0.41
0.29
0.45
0.21
0.23
0.21
***
***
***
***
SO4
0.20
0.26
0.31
0.32
***
***
***
***
***
***
***
***
**
Cl
-0.5
90.
310.
350.
520.
470.
280.
310.
300.
39**
***
***
***
***
***
***
***
***
*
Ca
-0.5
00.
320.
200.
260.
240.
290.
290.
570.
48**
***
***
***
***
***
***
*
Mg
-0.4
10.
250.
280.
300.
280.
290.
580.
480.
88**
***
***
***
***
***
*
Na
-0.5
50.
260.
340.
530.
320.
520.
530.
640.
450.
38**
***
***
***
***
*
K-0
.20
0.21
0.36
0.33
0.26
0.26
0.68
0.57
0.44
0.45
0.55
***
***
***
**
pH-0
.33
0.20
0.35
0.31
0.83
0.78
0.33
***
***
***
***
Alk
-0.5
70.
310.
240.
300.
200.
210.
210.
340.
480.
920.
800.
390.
340.
87**
***
***
***
*
Con
d-0
.50
0.34
0.28
0.28
0.32
0.32
0.62
0.51
0.99
0.93
0.47
0.48
0.82
0.88
***
**
Tem
p-0
.59
0.22
0.33
0.45
0.31
0.23
0.53
0.44
0.45
0.45
0.22
0.33
0.47
0.44
***
NS
-0.2
4-0
.30
-0.2
10.
250.
25**
*
EW
0.54
-0.4
2-0
.32
-0.4
6-0
.24
-0.2
8-0
.30
-0.5
9-0
.33
-0.2
5-0
.26
-0.3
00.
32
Alt
iis
alti
tude
,SR
Pis
solu
ble
reac
tive
phos
phat
e,D
INis
tota
ldis
solv
edin
orga
nic
nit
roge
n,A
lkis
alka
linit
y,C
ond
isco
ndu
ctiv
ity,
and
Tem
pis
tem
per
atu
re.
**P
<0.
01;*
**P
<0.
001.
U. Obertegger et al.
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd900
addition, the substitution of altitude by environmental predic-
tors explained more variance than altitude alone, and altitude
per se could not further account for the unexplained variation.
Therefore, we suggest that these combined predictors suffi-
ciently covered the effect of altitude on rotifer species richness.
Traditionally, the influence of area on species richness has
been explained by the theory of island biogeography (Mac-
Arthur & Wilson, 1967) or by the habitat diversity hypothesis
(Williams, 1964). However, these concepts are not mutually
exclusive, and theoretically they even seem to be complementary
because area and habitat diversity are correlated (Kallimanis
et al. 2008 and references therein). In our study, area was the
most influential predictor on species richness. We suggest that
lakes with larger area both sustained more vertical and horizon-
tal niches, offering greater habitat diversity, and had a lower
extinction and higher immigration rate than smaller lakes. Ver-
tical and horizontal niches are defined by physical, chemical and
biological parameters. As a consequence, habitat diversity
cannot be easily distinguished from other related factors
(Hessen et al., 2006). In fact, it was not surprising that area was
the predictor with the greatest effect on species richness because
it covers many different aspects of lake morphology.
Silica is an essential nutrient for siliceous algal species, pre-
dominantly diatoms and chrysophytes, and can limit growth
with concentrations below 0.5 mg l-1 (Parker et al., 2008). In this
way, silica influences algal productivity and succession and can
play an important role in rotifer species richness by determining
food composition (Swadling et al., 2000). Therefore, it was
expected that an environmental parameter related to nutrition
and productivity would score highly within the hierarchy of
predictor influence.
Temperature influences all metabolic functions and is a major
force in structuring communities (Litchman & Klausmeier,
2008) and in determining species diversity (Allen et al., 2002).
Our study also evidenced the positive effect of temperature on
species richness, but surprisingly, temperature scored third
within the hierarchy of effects. We attributed this to the represen-
tation of the temperature gradient by a single value (i.e. surface
water temperature). While surface water temperature could only
partially reflect the environmental temperature to which species
were exposed, it nevertheless determined the maximum tem-
perature present. In fact, surface water temperature is a good
surrogate for the actual input of energy (Hessen et al., 2007).
TP is a proxy for lake primary production (Hessen et al., 2006;
Barnett & Beisner, 2007). However, Jones et al. (2003) and
Barnett & Beisner (2007) note that this parameter is a poor
predictor for species richness. We also found that TP was only
marginally significant. It seems that this predictor is more
closely related to functional than taxonomic diversity (Barnett &
Beisner, 2007), an issue that would require further investigation.
In temperate lakes, phosphorus and silica are the most
common limiting nutrients, while nitrogen is limiting only
when the N:P ratio is < 16 (Lampert & Sommer, 2007). This was
the case in 29% of our lakes, and we expected a positive influ-
ence of nitrates on species richness; contrarily, it had a negative
effect. While high concentrations of nitrates can cause a wide
range of problems including loss of biodiversity (Carpenter
et al., 1998), in all our lakes, concentrations of nitrates were
much lower than the 10 mg NO3 l-1 threshold for potential
toxicity to aquatic organisms (Camargo & Alonso, 2006). Non-
point pollution from agriculture and urban activity, including
industry and transport, is a major source of nitrogen for fresh
water (Camargo & Alonso, 2006). The same sources also trans-
port many complex, ill-defined chemicals (e.g. pharmaceuticals,
pesticides, fertilizers and their metabolites) (Sumpter, 2009).
Rotifers are sensitive to pollutants, and this determines their use
as indicators of trophic conditions and test organisms in toxicity
assays (Sládecek, 1983). We suggest that the negative effect of
nitrates on species richness could be related to unmeasured
co-occurring pollutants, and consequently this predictor
Table 4 Results of the multiple regression without and with spatial covariates between species richness and environmental variables.
Without covariates (GLS) With covariates (GLM)
Parameter Estimate Error Influence Parameter Estimate Error Influence
pseudo-R2 = 0.49; P < 0.001 pseudo-R2 = 0.51; P < 0.001
Intercept 15.19 ¥ 10-1 1.84 ¥ 10-2 Intercept 2.77 ¥ 101 4.81 ¥ 10-1
Area 2.75 ¥ 10-1 3.95 ¥ 10-3 0.053 Area 2.11 ¥ 10-1 3.01 ¥ 10-3 0.041
Si 6.91 ¥ 10-1 7.41 ¥ 10-3 0.065 Si 3.88 ¥ 10-1 7.02 ¥ 10-3 0.037
Temp 4.36 ¥ 10-2 7.42 ¥ 10-4 0.048 Temp. 2.77 ¥ 10-2 5.68 ¥ 10-4 0.031
N-NO3 -2.21 ¥ 10-1 6.93 ¥ 10-3 -0.023 N-NO3 -2.45 ¥ 10-1 5.26 ¥ 10-3 -0.026
TP 3.00 ¥ 10-1 9.40 ¥ 10-3 0.023 TP 2.23 ¥ 10-1 7.82 ¥ 10-3 0.017
SO4 -4.71 ¥ 10-1 7.18 ¥ 10-3 -0.054 EW 4.77 ¥ 10-6 8.30 ¥ 10-8 0.036
Cond 3.66 ¥ 10-1 8.15 ¥ 10-3 0.041 NS -5.70 ¥ 10-4 9.50 ¥ 10-8 -0.041
Estimate refers to partial regression slope, error to standard error, and influence to relative influence (for explanation see Materials and Methods). Speciesrichness is square-root transformed in GLS; all other variables except for temperature and spatial directions were log-transformed. Error refers tostandard error. The GLM is based on a quasi-Poisson distribution. All predictors were statistically significant at P < 0.01 except for conductivity (P < 0.05)and TP (P = 0.06).Area is lake surface area, Si is reactive silica, Temp is surface water temperature, TP is total phosphorus and Cond is conductivity.
Rotifer species richness in the Alps
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd 901

seemed to be related to human influence instead of
productivity.
Apart from environmental predictors, spatial covariates were
important factors for the species richness–environment rela-
tionship. In fact, the spatial structure of environmental data has
to be taken into account even if it is not an easy task to find the
most appropriate model (Beguería & Pueyo, 2009). In the
species richness–altitude model as well as in the species
richness–environment model, spatial covariates had the same
effect on species richness. We related the increase in species
richness along the easterly and southerly directions (Fig. 3) to
the geological heterogeneity of our study area, where very dif-
ferent geological formations can be found over short distances
(Fig. 6). While with the inclusion of covariates in the linear
regression the fit was markedly improved, in the multiple regres-
sion the variance increased only slightly (GLM: pseudo-R2 =0.49 versus 0.51) but led to the non-significance of conductivity
and sulphates. This comparison of different statistical ap-
proaches revealed the spatial structure of our data: we suggest
that conductivity and sulphates had a strong spatial component
that could be completely described by geographical directions.
In our study region, lake conductivity reflected geological origin
with metamorphic bedrock more common in the northern part
and easily weathered sedimentary bedrock more common in the
southern part of the region (Fig. 6). Sulphate was also related to
geographical directions even if it was regarded as a proxy for
human influence. However, we cannot exclude some overlap of
human influence within spatial dependence. Other parameters
linked to weathering of the bedrock, such as silica, might also
have a spatial component that could not, however, be completely
described by covariates.
Interestingly, five high-altitude lakes (Finailsee, Hunger-
schartensee, Langsee-Ulten, Kleiner Plombodensee, Schwarzsee-
Ulten) on metamorphic bedrock had higher than expected
sulphate (range 50–95 mg l-1 SO4) and conductivity (range 132–
233 mS cm-1) values (Fig. 6). While alpine lakes were once con-
sidered pristine environments, a recent review on European
mountain lakes shows them to be impacted by atmospheric
deposition and global warming (Battarbee et al., 2009). The
melting of ice and rock glaciers leads to an increase of conduc-
tivity due to the release of solutes and pollutants such as sul-
phate, nickel and manganese previously immobilized in the ice
(e.g. Thies et al., 2007). We suggest that these extraordinarily
high values of conductivity and sulphate for lakes located on
metamorphic bedrock could be an early warning sign of the
impact of melting ice on high-altitude aquatic systems.
By our model, we explained 51% of the variability of species
richness using environmental predictors and spatial covariates.
While environmental predictors of the species richness–altitude
pattern were related to specific key factors such as habitat diver-
sity, productivity, heat content and human influence, spatial
covariates were linked to the geologically heterogeneous terri-
tory. Our study underlined how including both environmental
and spatial predictors can enhance hypothesis-driven consider-
ations on species richness. Further unexplained variability of
species richness might be attributed to unconsidered factors
such as inter- and intraspecific competition for food sources
(Walz, 1995), hydrology (Soranno et al., 1999; Obertegger et al.,
2007) and dispersal (Swadling et al., 2000).
The linear decrease of species richness along the altitudinal
gradient was successfully captured by the interplay of habitat
diversity, productivity, heat content and human influence. But
why do terrestrial studies mainly show a hump-shaped pattern
of species richness while aquatic studies tend to show a linear
pattern? The inclusion of anthropogenically disturbed habitats
can cause a hump-shaped pattern (Nogués-Bravo et al., 2008),
and we argue that this may not completely apply to aquatic
systems. Recent improvements in water quality, especially in
temperate lowland lakes, while not limiting non-point sources,
have greatly reduced direct pollution (Søndergaard & Jeppesen,
2007). Moreover, precipitation (Gotelli et al., 2009) or water
availability (Grytnes & McCain, 2007) are very decisive in con-
junction with temperature for the unimodal species richness
pattern. However, aquatic systems – to be considered as such –
must retain their aquatic status, along with their main physical
properties related to water. Obviously, aquatic systems are also
subject to change, but in contrast to terrestrial ones, water pro-
vides a relatively stable set-up of environmental conditions in
which biotic interactions take place (Reynolds, 1998; Lampert
& Sommer, 2007).These characteristics of freshwater ecosys-
tems may ultimately distinguish them from their terrestrial
counterparts with regards to the species richness–altitude rela-
tionship. We argue that the factors determining species
richness–altitude patterns tend to be the same in terrestrial and
aquatic habitats, but the greater environmental stability of
Figure 6 Conductivity (mS cm-1) in sampled sites with theunderlying bedrock: the diameter of closed circles corresponds todifferent classes of conductivity. Lakes with unexpectedly highsulphate and conductivity values are circled.
U. Obertegger et al.
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd902

aquatic systems seems to lead to linear instead of hump-shaped
patterns.
ACKNOWLEDGEMENTS
U.O. is supported by post-doc grant CERCA (Province of
Trento, Italy). This work was carried out within the research
activity funded by IASMA and an ECOPLAN Research Grant
(Province of Trento, Italy). We thank Vigilio Pinamonti, Gino
Leonardi and Franz Obertegger for help with sampling, Gio-
vanna Pellegrini for access to some unpublished data and Fabio
Zottele for statistical advice and the site map. We also thank
David Currie, Patricia Soranno and an anonymous referee for
suggestions that substantially improved this manuscript.
REFERENCES
Allen, A.P., Brown, J.H. & Gillooly, J.F. (2002) Global biodiver-
sity, biochemical kinetics, and the energetic-equivalence rule.
Science, 297, 1545–1548.
Barnett, A. & Beisner, B.E. (2007) Zooplankton biodiversity and
lake trophic state: explanations invoking resource abundance
and distribution. Ecology, 88, 1675–1686.
Battarbee, R.W., Kernan, M. & Rose, N. (2009) Threatened and
stressed mountain lakes of Europe: assessment and progress.
Aquatic Ecosystem Health and Management, 12, 118–128.
Bêche, L.A. & Statzner, B. (2009) Richness gradients of stream
invertebrates across the USA: taxonomy- and trait-based
approaches. Biodiversity and Conservation, 18, 3909–3930.
Beguería, S. & Pueyo, Y. (2009) A comparison of simultaneous
autoregressive and generalized least squares models for
dealing with spatial autocorrelation. Global Ecology and Bio-
geography, 18, 273–279.
Buse, A. (1973) Goodness of fit in generalized least squares
estimation. The American Statistician, 27, 106–108.
Camargo, J.A. & Alonso, Á. (2006) Ecological and toxicological
effects of inorganic nitrogen pollution in aquatic ecosystems:
a global assessment. Environment International, 32, 831–849.
Cantonati, M. & Lazzara, M. (2006) High mountain lakes in the
Avisio River watershed (east Trentino). Natural History
Museum of Trento, Trento, Italy (in Italian).
Cantonati, M., Tolotti, M. & Lazzara, M. (2002) Lakes in the
Adamello-Brenta Natural Park. Natural History Museum of
Trento, Trento, Italy (in Italian).
Carpenter, S.R., Caraco, N.F., Correll, D.L., Howarth, R.W.,
Sharpley, A.N. & Smith, V.H. (1998) Nonpoint pollution of
surface waters with phosphorus and nitrogen. Ecological
Applications, 8, 559–568.
Colwell, R.K. & Lees, D.C. (2000) The mid-domain effect: geo-
metric constraints on the geography of species richness.
Trends in Ecology and Evolution, 15, 70–76.
Currie, D.J. (2007) Disentangling the roles of environment and
space in ecology. Journal of Biogeography, 34, 2009–2011.
Currie, D.J. & Kerr, J.T. (2008) Tests of the mid-domain hypoth-
esis: a review of the evidence. Ecological Monographs, 78, 3–18.
Davies, R.G., Orme, C.D.L., Webster, A.J., Jones, K.E., Blackburn,
T.M. & Gaston, K.J. (2007) Environmental predictors of global
parrot (Aves: Psittaciformes) species richness and phylogenetic
diversity. Global Ecology and Biogeography, 16, 220–233.
Fontaneto, D. & Ricci, C. (2006) Spatial gradients in species
diversity of microscopic animals: the case of bdelloid rotifers
at high altitude. Journal of Biogeography, 33, 1305–1313.
Gotelli, N.J., Anderson, M.J., Arita, H.T. et al. (2009) Patterns
and causes of species richness: a general simulation model for
macroecology. Ecology Letters, 12, 873–886.
Grytnes, J.A. & McCain, C.M. (2007) Elevational trends in
biodiversity. Encyclopedia of biodiversity (ed. by S.A. Levin),
pp. 1–8. Elsevier Inc., New York.
Grytnes, J.A., Heegaard, E. & Ihlen, P.G. (2006) Species richness
of vascular plants, bryophytes, and lichens along an altitudinal
gradient in western Norway. Acta Oecologica, 29, 241–246.
Hessen, D.O., Faafeng, B.A., Smith, V.H., Bakkestuen, V. &
Walseng, B. (2006) Extrinsic and intrinsic controls of zoop-
lankton diversity in lakes. Ecology, 87, 433–443.
Hessen, D.O., Bakkestuen, V. & Walseng, B. (2007) Energy input
and zooplankton species richness. Ecography, 30, 749–758.
Hubbell, S.P. (2001) The unified neutral theory of biodiversity and
biogeography. Princeton University Press, Princeton, NJ.
IASMA (1996–2000) Annual reports on the limnological charac-
teristics of Trentino lakes. Istituto Agrario di San Michele
all’Adige, Trento (in Italian).
Jacobsen, D. (2004) Contrasting patterns in local and zonal
family richness of stream invertebrates along an Andean alti-
tudinal gradient. Freshwater Biology, 49, 1293–1305.
Jankowski, T. & Weyhenmeyer, G.A. (2006) The role of spatial
scale and area in determining richness–altitude gradients in
Swedish lake phytoplankton communities. Oikos, 115, 433–
442.
Jersabek, C.D. (1995) Distribution and ecology of rotifer com-
munities in high-altitude alpine sites – a multivariate
approach. Hydrobiologia, 313, 75–89.
Jones, J.I., Li, W. & Maberly, S.C. (2003) Area, altitude and
aquatic plant diversity. Ecography, 26, 411–420.
Kallimanis, A.S., Mazaris, A.D., Tzanopoulos, J., Halley, J.M.,
Pantis, J.D. & Sgardelis, S.P. (2008) How does habitat diversity
affect the species–area relationship? Global Ecology and Bioge-
ography, 17, 532–538.
Kluge, J., Kessler, M. & Dunn, R.R. (2006) What drives eleva-
tional patterns of biodiversity? A test of geometric constraints,
climate, and species pool effects for pteridophytes on an eleva-
tional gradient in Costa Rica. Global Ecology and Biogeogra-
phy, 15, 358–371.
Körner, C. (2007) The use of ‘altitude’ in ecological research.
Trends in Ecology and Evolution, 22, 569–574.
Lampert, W. & Sommer, U. (2007) Limnoecology. The ecology of
lakes and streams. Oxford University Press, Oxford.
Li, J., He, Q., Hua, X., Zhou, J., Xu, H., Chen, J. & Fu, C. (2009)
Climate and history explain the species richness peak at mid-
elevation for Schizothorax fishes (Cypriniformes: Cyprinidae)
distributed in the Tibetan Plateau and its adjacent regions.
Global Ecology and Biogeography, 18, 264–272.
Rotifer species richness in the Alps
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd 903

Litchman, E. & Klausmeier, C.A. (2008) Trait-based community
ecology of phytoplankton. Annual Review of Ecology, Evolu-
tion, and Systematics, 39, 615–639.
MacArthur, R.H. & Wilson, E.O. (1967) The theory of island
biogeography. Princeton University Press, Princeton, NJ.
McCain, C.M. (2007) Area and mammalian elevational diver-
sity. Ecology, 88, 76–86.
Nogués-Bravo, D., Araùjo, M.B., Romdal, T. & Rahbek, C.
(2008) Scale effects and human impact on the elevational
species richness gradients. Nature, 453, 216–220.
Nyman, M., Korhola, A. & Brooks, S.J. (2005) The distribution
and diversity of Chironomidae (Insecta: Diptera) in western
Finnish Lapland, with special emphasis on shallow lakes.
Global Ecology and Biogeography, 14, 137–153.
Obertegger, U., Flaim, G., Braioni, M.G., Sommaruga, R.,
Corradini, F. & Borsato, A. (2007) Water residence time as a
driving force of zooplankton structure and succession.
Aquatic Sciences, 69, 575–583.
Obertegger, U., Thaler, B. & Flaim, G. (2008) Habitat constraints
of Synchaeta (Rotifera) in north Italian lakes (Trentino–South
Tyrol). Verhandlungen Internationale Vereinigung für Theore-
tische und Angewandte Limnologie, 30, 302–306.
Parker, B.R., Vinebrooke, R.D. & Schindler, D.W. (2008) Recent
climate extremes alter alpine lake ecosystems. Proceedings of
the National Academy of Sciences USA, 105, 12927–12931.
Quinn, G.P. & Keough, M.J. (2002) Experimental design and
data analysis for biologists. Cambridge University Press,
Cambridge.
R Development Core Team (2005) R: a language and environ-
ment for statistical computing. R Foundation for Statistical
Computing, Vienna. Available at: http://www.r-project.org/
(accessed 26 June 2009).
Rahbek, C. (1995) The elevational gradient of species richness –
a uniform pattern. Ecography, 18, 200–205.
Rahbek, C. (2005) The role of spatial scale and the perception of
large-scale species-richness patterns. Ecological Letters, 8, 224–
239.
Reynolds, C.S. (1998) The state of freshwater ecology. Freshwa-
ter Biology, 39, 741–753.
Rowe, R.J. (2009) Environmental and geometric drivers of small
mammal diversity along elevational gradients in Utah.
Ecography, 32, 411–422.
Salmaso, N. & Naselli-Flores, L. (1999) Studies on the zooplank-
ton of the deep subalpine Lake Garda. Journal of Limnology,
58, 66–76.
Scheffer, M. & van Geest, G.J. (2006) Small habitat size and
isolation can promote species richness: second-order effects
on biodiversity in shallow lakes and ponds. Oikos, 112, 227–
231.
Sládecek, V. (1983) Rotifers as indicators of water-quality.
Hydrobiologia, 100, 169–201.
Smith, K.F. & Brown, J.H. (2002) Patterns of diversity, depth
range and body size among pelagic fishes along a gradient of
depth. Global Ecology and Biogeography, 11, 313–322.
Søndergaard, M. & Jeppesen, E. (2007) Anthropogenic impacts
on lake and stream ecosystems, and approaches to restoration.
Journal of Applied Ecology, 44, 1089–1094.
Soranno, P.A., Webster, K.E., Riera, J.L., Kratz, T.K., Baron, J.S.,
Bukaveckas, P.A., Kling, G.W., White, D.S., Caine, N., Lathrop,
R.C. & Leavitt, P.R. (1999) Spatial variation among lakes
within landscapes: ecological organization along lake chains.
Ecosystems, 2, 395–410.
Sturm, R. (2007) Freshwater molluscs in mountain lakes of the
eastern Alps (Austria): relationship between environmental
variables and lake colonization. Journal of Limnology, 66, 160–
169.
Sumpter, J.P. (2009) Protecting aquatic organisms from chemi-
cals: the harsh realities. Philosophical Transactions of the Royal
Society A: Mathematical Physical and Engineering Sciences,
367, 3877–3894.
Swadling, K.M., Pienitz, R. & Nogrady, T. (2000) Zooplankton
community composition of lakes in the Yukon and Northwest
Territories (Canada): relationship to physical and chemical
limnology. Hydrobiologia, 431, 211–224.
Thies, H.J., Nickus, U., Mair, V., Tessadri, R., Tait, D., Thaler, B.
& Psenner, R. (2007) Unexpected response of high alpine lake
waters to climate warming. Environmental Science and Tech-
nology, 41, 7424–7429.
Walz, N. (1995) Rotifer populations in plankton communities:
energetics and life history strategies. Experientia, 51, 437–453.
Wiens, J.J. & Donoghue, M.J. (2004) Historical biogeography,
ecology and species richness. Trends in Ecology and Evolution,
19, 639–644.
Williams, C.B. (1964) Patterns in the balance of nature. Academic
Press, London.
BIOSKETCHES
Ulrike Obertegger is a post-doc fellow at the IASMA
Research and Innovation Centre, FEM, Trento. Her
main research interests are rotifer taxonomy, ecology
and distribution patterns, and in particular the role of
environmental factors that drive zooplankton
biodiversity at different scales. She also is a
mountaineer with a passion for high-altitude lake
sampling.
Bertha Thaler is a limnologist at the provincial
Environmental Protection Agency, Bozen. Her main
interests are lake monitoring and zooplankton ecology,
with particular emphasis on alpine lakes.
Giovanna Flaim is a limnologist at the IASMA
Research and Innovation Centre, FEM, Trento. Her
main interests are in ecosystem functioning and the
relation of abiotic and biotic factors to plankton
autecology.
Editor: Tim Blackburn
U. Obertegger et al.
Global Ecology and Biogeography, 19, 895–904, © 2010 Blackwell Publishing Ltd904
Related Documents











