195 Studi Trent. Sci. Nat., 94 (2014): 195-207 1. INTRODUCTION Knowledge about bryozoans from the Mediterra- nean Sea is based on nearly two hundred years of investiga- tions. However, most works essentially deal with taxonomy (Rosso et al. 2010a and references therein), whereas rela- tively few works have been published on bryozoan com- munities, their composition and structure. Exceptions relate to selected localities and special habitats mostly concerning hard bottom communities and particularly submarine caves (Harmelin 1969; 1980; 1985; 1986; 1997, 2000; Balduzzi & Rosso 2003; Rosso et al. 2012 and references therein) as well as the bioconstructions produced by the “Corallig- enous Bioceonosis” (Gautier 1959; Laubier 1966; Rosso & Sanfilippo 2009 and references therein) and by the cold- water corals (Zabala et al. 1993; Mastrototaro et al. 2010; Rosso et al. 2010b). In contrast, soft bottom communities have gener- ally received less attention, probably also following the general assumption that bryozoans are mostly localised on hard surfaces (McKinney & Jackson 1989). Actually, only few papers focus on such communities, which, in the Medi- terranean region, include the Coastal Detritic Biocoenosis (Rosso 1989, 1996a, 1996b) and special bryozoan carpets in the Adriatic (McKinney & Jaklin 2000, 2001). Detritic environments from the open shelf have ne- arly completely been neglected and the unique knowledge derives from qualitative syntheses reported in the general, mostly taxonomic, monographies given on bryozoans from the Mediterranean Sea by Gautier (1962), Harmelin (1976) and Zabala (1986). However, these authors provide only some general information about the bryozoan communi- ties as a whole and, particularly, the bryozoans encrusting small, usually shelly, substrata on these bottoms (see for instance Harmelin 1976). The present paper aims to contribute preliminary ISSN 2035-7699 © Museo delle Scienze, Trento 2014 Open-shelf, soft-bottom bryozoans from the Ciclopi Marine Protected Area (E Sicily, Mediterranean) Antonietta ROSSO * , Rossana SANFILIPPO & Francesco SCIUTO Dipartimento di Scienze Biologiche, Geologiche e Ambientali, Sezione di Scienze della Terra, Laboratorio di Paleoe- cologia, University of Catania, Corso Italia 55, 95129, Catania, Italy Corresponding author e-mail: [email protected] SUMMARY - Open shelf, soft bottom bryozoans from the Ciclopi Marine Protected Area (E Sicily, Mediterranean) - The bryozoan bio- coenoses from five samples collected in open-shelf, soft-bottoms between 83 and 95 metres depth in the Ciclopi Marine Protected Area (north of Catania, Sicily) in the Ionian Sea (Mediterranean) have been studied. Communities are characterised by an apparently low total (44 species), and per station (4-24) species richness; an estimated colony abundance of 40-500 per square metre; the presence of a few number of shared species, and a large number of rare species that are often restricted to single samples, pointing to a patchiness that is possibly linked to substratum availability; the presence of some distinctive species hitherto not recorded from deep shelf soft bottoms; and morphological adaptations for the colonisation of ephemeral clasts mostly falling within the sand and granule sizes. Comparisons are made with scant literature data concerning bryozoans from similar environments from the Mediterranean area, and inferences are put forward regarding environmental factors driving community composition at species and zoarial growth habit level. RIASSUNTO - I briozoi dei fondi mobili della piattaforma esterna dell’Area Marina Protetta delle Isole Ciclopi (Sicilia orientale, Me- diterraneo) - Sono state studiate le associazioni a briozoi viventi di cinque campioni prelevati in fondi mobili della piattaforma esterna, fra 83 e 95 metri di profondità nell’Area Marina protetta delle Isole Ciclopi (nord di Catania, Sicilia) nel Mar Ionio (Mediterraneo). Le comunità sono caratterizzate da: una ricchezza specifica (complessiva: 44 specie, e per stazione: 4-24) apparentemente bassa; un numero di colonie per metro quadrato stimato fra 40 e 500; la presenza di un ridotto numero di specie comuni cui si accompagna un cospicuo numero di specie rare, spesso localizzate in un unico sito/campione, che indicherebbe una certa patchiness possibilmente connessa alla disponibilità di substrati adatti alla colonizzazione; la presenza di alcune specie caratterizzanti finora non segnalate da fondi mobili della piattaforma profonda; adattamenti morfologici funzionali alla colonizzazione di clasti effimeri prevalentemente ricadenti nelle dimensioni delle sabbie e dei granuli. Confronti sono stati fatti con i dati concernenti associazioni a briozoi di ambienti simili desumibili dalla scarsa letteratura disponibile per l’area mediterranea e inferenze sono state avanzate circa i fattori ambientali che condizionano la composizione delle associazioni riscontrate sia a livello di specie sia a livello di adattamenti zoariali. Keywords: Bryozoans, colony morphology, adaptations, lower circalittoral, Mediterranean, Recen Parole chiave: Briozoi, morfologia coloniale, adattamenti, circalitorale inferiore, Mediterraneo, Attuale Bryozoan Studies 2013 - Antonietta Rosso, Patrick N. Wyse Jackson & Joanne Porter (eds)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

195Studi Trent. Sci. Nat., 94 (2014): 195-207
1. IntroductIon
Knowledge about bryozoans from the Mediterra-nean Sea is based on nearly two hundred years of investiga-tions. However, most works essentially deal with taxonomy (rosso et al. 2010a and references therein), whereas rela-tively few works have been published on bryozoan com-munities, their composition and structure. Exceptions relate to selected localities and special habitats mostly concerning hard bottom communities and particularly submarine caves (Harmelin 1969; 1980; 1985; 1986; 1997, 2000; Balduzzi & rosso 2003; rosso et al. 2012 and references therein) as well as the bioconstructions produced by the “corallig-enous Bioceonosis” (Gautier 1959; Laubier 1966; rosso & Sanfilippo 2009 and references therein) and by the cold-water corals (Zabala et al. 1993; Mastrototaro et al. 2010; rosso et al. 2010b).
In contrast, soft bottom communities have gener-
ally received less attention, probably also following the general assumption that bryozoans are mostly localised on hard surfaces (McKinney & Jackson 1989). Actually, only few papers focus on such communities, which, in the Medi-terranean region, include the coastal detritic Biocoenosis (rosso 1989, 1996a, 1996b) and special bryozoan carpets in the Adriatic (McKinney & Jaklin 2000, 2001).
detritic environments from the open shelf have ne-arly completely been neglected and the unique knowledge derives from qualitative syntheses reported in the general, mostly taxonomic, monographies given on bryozoans from the Mediterranean Sea by Gautier (1962), Harmelin (1976) and Zabala (1986). However, these authors provide only some general information about the bryozoan communi-ties as a whole and, particularly, the bryozoans encrusting small, usually shelly, substrata on these bottoms (see for instance Harmelin 1976).
the present paper aims to contribute preliminary
ISSn 2035-7699© Museo delle Scienze, trento 2014
Open-shelf, soft-bottom bryozoans from the Ciclopi Marine Protected Area (E Sicily, Mediterranean)
Antonietta Rosso*, Rossana sAnfilippo & francesco sciuto
dipartimento di Scienze Biologiche, Geologiche e Ambientali, Sezione di Scienze della terra, Laboratorio di Paleoe-cologia, university of catania, corso Italia 55, 95129, catania, Italy corresponding author e-mail: [email protected]
SuMMArY - Open shelf, soft bottom bryozoans from the Ciclopi Marine Protected Area (E Sicily, Mediterranean) - the bryozoan bio-coenoses from five samples collected in open-shelf, soft-bottoms between 83 and 95 metres depth in the Ciclopi Marine Protected Area (north of catania, Sicily) in the Ionian Sea (Mediterranean) have been studied. communities are characterised by an apparently low total (44 species), and per station (4-24) species richness; an estimated colony abundance of 40-500 per square metre; the presence of a few number of shared species, and a large number of rare species that are often restricted to single samples, pointing to a patchiness that is possibly linked to substratum availability; the presence of some distinctive species hitherto not recorded from deep shelf soft bottoms; and morphological adaptations for the colonisation of ephemeral clasts mostly falling within the sand and granule sizes. comparisons are made with scant literature data concerning bryozoans from similar environments from the Mediterranean area, and inferences are put forward regarding environmental factors driving community composition at species and zoarial growth habit level.
rIASSunto - I briozoi dei fondi mobili della piattaforma esterna dell’Area Marina Protetta delle Isole Ciclopi (Sicilia orientale, Me-diterraneo) - Sono state studiate le associazioni a briozoi viventi di cinque campioni prelevati in fondi mobili della piattaforma esterna, fra 83 e 95 metri di profondità nell’Area Marina protetta delle Isole ciclopi (nord di catania, Sicilia) nel Mar Ionio (Mediterraneo). Le comunità sono caratterizzate da: una ricchezza specifica (complessiva: 44 specie, e per stazione: 4-24) apparentemente bassa; un numero di colonie per metro quadrato stimato fra 40 e 500; la presenza di un ridotto numero di specie comuni cui si accompagna un cospicuo numero di specie rare, spesso localizzate in un unico sito/campione, che indicherebbe una certa patchiness possibilmente connessa alla disponibilità di substrati adatti alla colonizzazione; la presenza di alcune specie caratterizzanti finora non segnalate da fondi mobili della piattaforma profonda; adattamenti morfologici funzionali alla colonizzazione di clasti effimeri prevalentemente ricadenti nelle dimensioni delle sabbie e dei granuli. confronti sono stati fatti con i dati concernenti associazioni a briozoi di ambienti simili desumibili dalla scarsa letteratura disponibile per l’area mediterranea e inferenze sono state avanzate circa i fattori ambientali che condizionano la composizione delle associazioni riscontrate sia a livello di specie sia a livello di adattamenti zoariali.
Keywords: Bryozoans, colony morphology, adaptations, lower circalittoral, Mediterranean, recenParole chiave: Briozoi, morfologia coloniale, adattamenti, circalitorale inferiore, Mediterraneo, Attuale
Bryozoan Studies 2013 - Antonietta rosso, Patrick n. Wyse Jackson & Joanne Porter (eds)

196 Rosso et al. Bryozoans from soft bottoms of the Ciclopi MPA
information about the species composition of bryozoan as-sociations from open-shelf soft-bottoms from the Mediter-ranean area of southern Italy, focusing also on the growth morphologies that colonies exploit for living on these fine-grained particulate bottoms.
2. MAtErIALS And MEtHodS
Examined samples come from the ciclopi Marine Protected Area (cMPA), located a dozen km north of cata-nia along the eastern flanks of Mount Etna (Fig. 1).
All along the coast volcanic rocks crop out, largely constituting the bottoms down to 10-15 m, locally extend-ing down to 40 m near the isles (Fig. 1). Further isolated rocky outcrops are present between 60-80 m constituting local shoals.
Soft bottoms are coarse (mostly gravels and grav-elly coarse sands) down to 40-50 m in the south and even to 70 m in the north. In deeper zones mid to fine sands are common and, from 70-80 m downward, two narrow belts of muddy fine sands and muds develop, largely following isobaths and extending roughly parallel to the coastline. Interestingly, a large amount of the coarse sediment frac-tion consists of volcanic ashes ejected by the Etna Volcano.
Particularly, at sampling period (summer 2000, just a few months after the paroxysmal events of spring 2000) ashes accounted for about 30-40% of the bottom sediment, lo-cally even more.
According to rosso (2001), and following the scheme by Pérès & Picard (1964), surface rocky bottoms host the AP (Photophilic Algae) Biocoenosis, whereas the coralligenous Biocoenosis is present on deep rocks. A nar-row belt of Posidonia meadows (HP Biocoenosis) is locally present nearshore at the base of the cliff bordering shallow rocky outcrops, growing on coarse soft bottoms between interspersed blocks. Shallow soft bottoms are largely swept by moderate to strong currents leading to the establishment of the Biocoenosis of coarse Sands and fine Gravels under the influence of Bottom Currents (SGCF), or its ecotones with other impoverished infralittoral biocoenoses down to about 30 m, and with the detritic coastal Biocoenosis in the shallow circalittoral down to 40-55 m. A large belt of this latter biocoenosis extends down to 70-80 m where the decreasing water energy causes an increase of the muddy component and the development of the Muddy detritic (dE) and the offshore detritic (dL) Biocoenoses as well as their ecotones in between.
Samples were obtained in July 2000 during surveys to finalize the sedimentological and bionomic characteriza-
Fig. 1 - Bionomic map of soft-bottoms from the ciclopi Marine Protected Area with the locations of the studied sam-ples. the straight continuous lines mark the boundaries of the A, B and c zones of complete to general protection within the cMPA. a: rocky bottoms; b: Bio-coenosis of the Posidonia meadows; c: Biocoenosis of the Fine-grained, Well Sorted Sands (SFBC); d: SFBS-SGCF ecotone; e: Biocoenosis of coarse Sands and fine Gravels under the influence of Bottom Currents (SGCF); f: SGFC-DC ecotone; g: Biocoenosis of the coastal detritic (dc); h: dc-dE ecotone; i: Biocoenosis of detritic Muds (dE); j: dE-dL ecotone; k: Biocoenosis of the offshore detritic (dL).Fig. 1 - Carta bionomica dei fondi mo-bili dell’Area Marina Protetta Isole Ci-clopi con la localizzazione dei campio-ni studiati. Le linee continue segnano i limiti delle zone A, B e C di protezione integrale e generale dell’AMP. a: Fon-di rocciosi; b: Biocenosi della Prateria di Posidonia; c: Biocenosi delle Sabbie Fini Ben Classate (SFBC); d: ecotono SFBS-SGCF; e: Biocenosi delle Sabbie grossolane e Ghiaie fini sotto l’influen-za delle Correnti di Fondo (SGCF); f: ecotono SGFC-DC; g: Biocenosi del Detritico Costiero (DC); h: ecotono DC-DE; i: Biocenosi del Detritico Infangato (DE); j: ecotono DE-DL; k: Biocenosi del Detritico del Largo (DL).

197Studi Trent. Sci. Nat., 94 (2014): 195-207
tion (rosso 2001) of the ecosystems in the recently erected cMPA. Samples employed for the present analysis come from a narrow belt where an ecotone between the dE and dL biocoenoses has been detected. Particularly, 5 samples were randomly selected among those available, distributed along the outer boundary of the cMPA, but in proximity of the areas A and B of maximum protection, in the depth range of 83-95 m. Samples were collected through a small Van Veen grab with an opening of 25x35 centimetres, en-abling to sample a surface of about 1/11 square metre.
Each sample, of about 5 litres, was routinely treated, i.e. washed and sieved through a series of nets with de-creasing mesh width. Living bryozoan colonies were sepa-rated from the residue. All selected bryozoan colonies were counted and relative percentages calculated. this datum was used for bionomical purposes.
dead colonies and colony fragments belonging to the same species found alive were also picked from frac-tions larger than 500 mm. these specimens were used to add information on the relationships between colonies and colonised clasts. dead colonies were also often employed for photos owing to the absence of soft tissues.
Photographs were taken of some colonies of par-ticular interest either through a Zeiss discovery V8A ster-eomicroscope equipped with an Axiocam Mrc and Axio-vision acquisition system, or a low vacuum LMu tescan Vega Scanning Electron Microscope at the university of catania.
the material is deposited at the Paleontological Sec-tion of the Earth Science Museum of the university of ca-tania, under codes PMc.r.I.H. B-1 to PMc.r.I.H. B-2 and PMc.r.I.H. B-17 to PMc.r.I.H. B-26.
3. rESuLtS
3. 1. Sediments
Sediments consist of sandy silts and silty sands (tab. 1), with sands largely represented by their medium-to-fine sized components. the fraction <250 mm usually exceeds 60%, even reaching about 80% (sample 8H), except for a single sample (8I) where it comprises only 41%. In con-trast, the coarse fractions are always subordinate, with tho-
se larger than 2 mm accounting for no more than about 3% (samples 8I and 10G) and those larger than 1 mm ranging from about 3 to 22% with particularly low values in sam-ples 8H and 12G where coarse sands and gravel account for about 3 and 6%, respectively.
coarse clasts are mostly represented by biogenic material, primarily mollusc shells (mostly bivalves) and subordinate barnacle and echinoid plates. Fragments of ser-pulids and bryozoan colonies are also present. Lithic ele-ments widely fall within fractions finer than 1 mm, though very large lithic clasts can occasionally be present. they are essentially volcanics, above all ash deposits following Etna eruptions (very irregular surfaces pointing also to a recent production), or transported to the sea from the land (abraded surfaces). rarely, isolated small volcanic and me-tamorphic pebbles can be found.
3. 2. Bionomy
the living association is scant and entirely consists of infaunal burrowing organisms. Molluscs, and particu-larly bivalves, dominate, mostly with small sized filter-feeding species such as Timoclea ovata (Pennant, 1877), Saccella commutata (Philippi, 1844), Parvicardium mini-mum (Philippi, 1836), Cardiomya costellata (deshayes, 1833), Bathyarca pectunculoides (Scacchi, 1835) and Pan-dora inequivalvis (Linnaeus, 1758). the holothurian Thra-chytyone tergestina (M. Sars, 1857) has also been found. All these species are usually restricted to mid-shelf envi-ronments, some extending down to the bathyal. neverthe-less, all species have relatively large ecological distribu-tion, colonising granumetrically heterogeneous sediments but being tolerant to different amounts of mud. Furthermo-re, the bivalve Centrocardita aculeata (Poli 1795) and the scaphopod Dentalium panormum chenu 1843, indicative of the offshore detritic biocoenosis (dL) have been found. consequently the assemblage has been ascribed to an eco-tone between the dL and the dE (Muddy detritic) bioco-enoses.
3. 3. Bryozoans
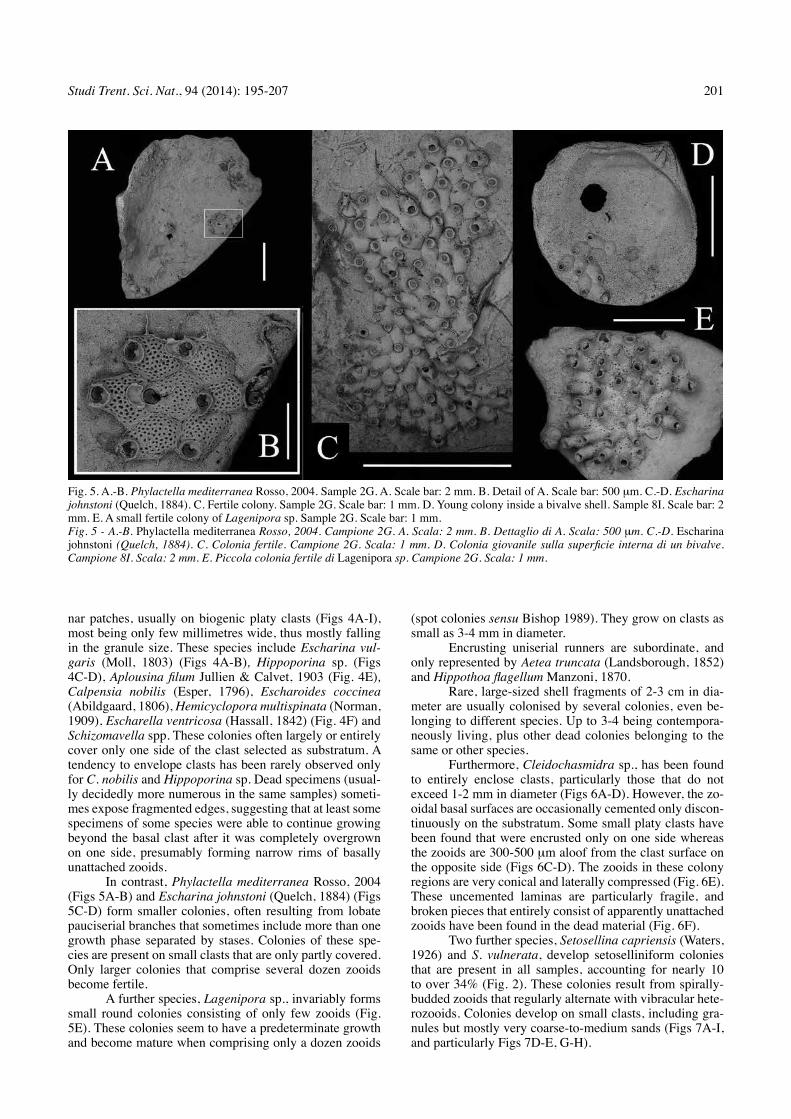
A total of 44 bryozoan species (tab. 2) have been detected forming the living community, mostly belonging
Tab. 1 - List of the studied samples from the Ciclopi Marine Protected Area. For each sample geographical, bathymetric and sedimento-logical information is given. c: coarse; m: medium; f: fine; Organ: bioclastic grains; Volc.: volcanic grains. Data on sediment texture and nature are reported as relative percentages.Tab. 1 - Lista dei campioni studiati, provenienti dall’Area Marina Protetta Isole Ciclopi. Per ciascun campione sono riportate informa-zioni circa la provenienza geografica e batimetrica e le caratteristiche tessiturali. c: grosse; m: medie; f: fini; Organ.: clasti organogeni; Volc.: clasti vulcanici. I dati su tessitura e natura dei sedimenti sono forniti in percentuale.
Ciclopi 2000 Survey Sediment texture Sediment natureStation lat. n long. E Depth Gravel c sand m-f sand silt clay organ. Volc. other2G 4158120 516110 87.5 1.63 10.47 33.87 50.25 3.78 43.75 55.75 0.508H 4156920 515750 92.5 1.09 2.01 44.16 50.29 2.45 47.50 49.80 2.708I 4156850 515740 95.0 2.95 19.37 48.03 28.01 1.64 58.10 41.90 0.0010G 4156630 515300 85.0 3.01 7.88 42.42 43.38 3.31 62.50 37.10 0.4012G 4156220 514920 83.0 1.27 4.33 52.44 39.41 2.55 45.81 53.44 0.75
198 Rosso et al. Bryozoans from soft bottoms of the Ciclopi MPA
to the cheilostomes, which account for 37 species. cyclo-stomes, with 7 species, are subordinate. Each species is usually present in few or even in a single sample, and nor-mally with a small number of colonies, and even a single colony.
the associations, in each sample and overall, appear to be poor and usually little diversified. The number of spe-cies ranges from 4 to 24 with a mean value of 13 species per sample whereas the number of colonies per sample ranges from 4 to 45, with a mean value of 25. taking into account that the sampled bottom surface corresponds to 875 cm2, a
total number of about 40 to more than 500 colonies can be estimated to thrive per square metre at the collection sites.
A few species are frequent (frequence indicating the number of samples where a given species is present out of the total number of samples) and only two species are present in all the investigated samples. these are Setosella vulnerata (Busk, 1860) and Hippoporina sp. Interestingly, these two species show the highest numbers of colonies (up to 9 in single samples) in the entire collection, though with relatively low mean values (4.2 and 3, respectively). In contrast, half of the species listed is present only in a single
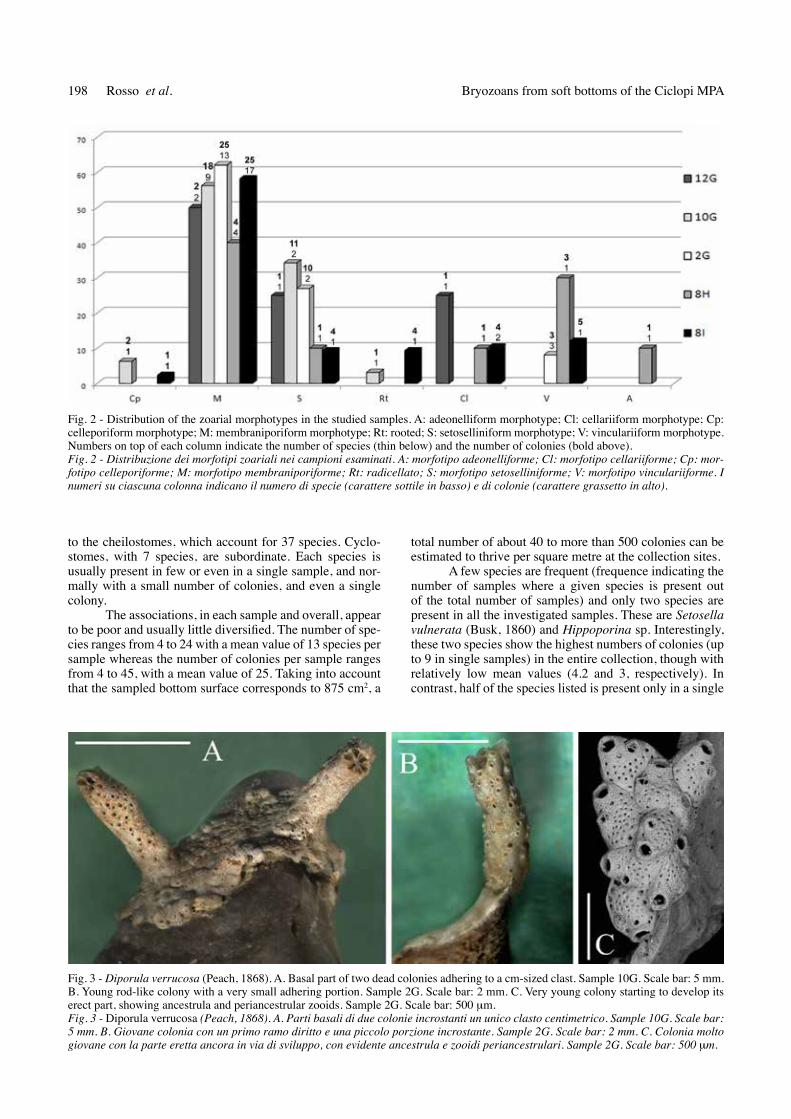
Fig. 2 - Distribution of the zoarial morphotypes in the studied samples. A: adeonelliform morphotype; Cl: cellariiform morphotype; Cp: celleporiform morphotype; M: membraniporiform morphotype; rt: rooted; S: setoselliniform morphotype; V: vinculariiform morphotype. numbers on top of each column indicate the number of species (thin below) and the number of colonies (bold above).Fig. 2 - Distribuzione dei morfotipi zoariali nei campioni esaminati. A: morfotipo adeonelliforme; Cl: morfotipo cellariiforme; Cp: mor-fotipo celleporiforme; M: morfotipo membraniporiforme; Rt: radicellato; S: morfotipo setoselliniforme; V: morfotipo vinculariiforme. I numeri su ciascuna colonna indicano il numero di specie (carattere sottile in basso) e di colonie (carattere grassetto in alto).
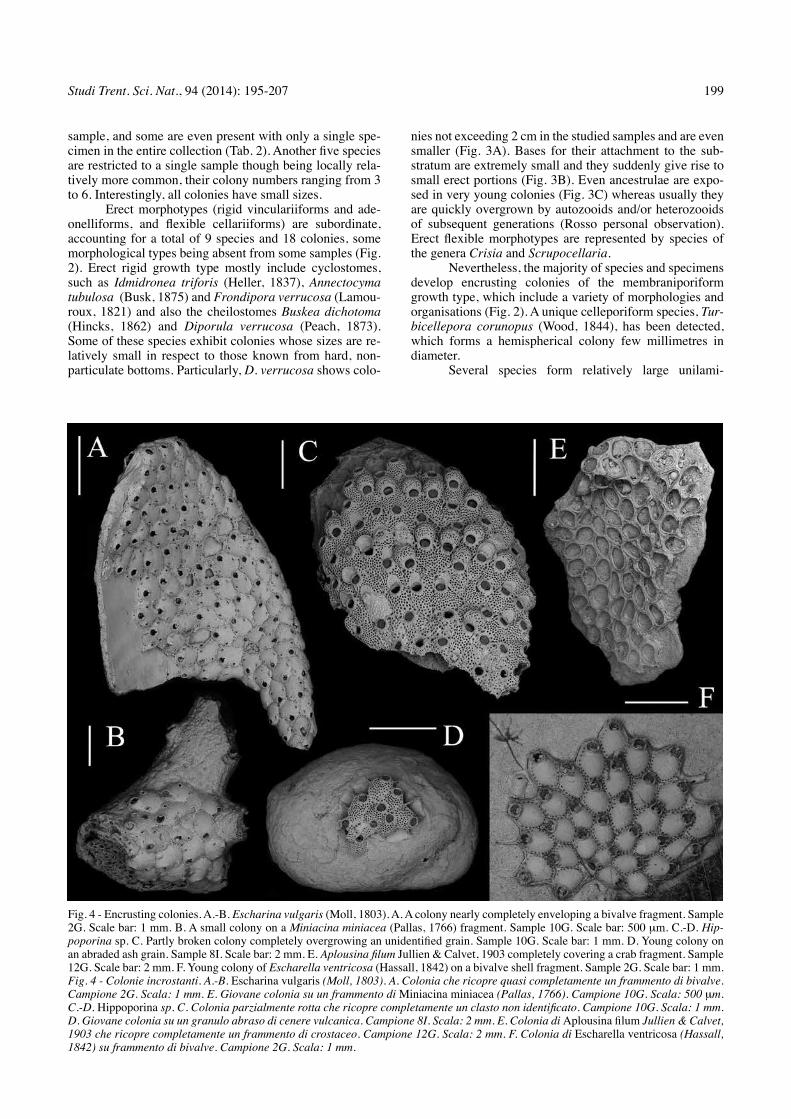
Fig. 3 - Diporula verrucosa (Peach, 1868). A. Basal part of two dead colonies adhering to a cm-sized clast. Sample 10G. Scale bar: 5 mm. B. Young rod-like colony with a very small adhering portion. Sample 2G. Scale bar: 2 mm. c. Very young colony starting to develop its erect part, showing ancestrula and periancestrular zooids. Sample 2G. Scale bar: 500 mm.Fig. 3 - diporula verrucosa (Peach, 1868). A. Parti basali di due colonie incrostanti un unico clasto centimetrico. Sample 10G. Scale bar: 5 mm. B. Giovane colonia con un primo ramo diritto e una piccolo porzione incrostante. Sample 2G. Scale bar: 2 mm. C. Colonia molto giovane con la parte eretta ancora in via di sviluppo, con evidente ancestrula e zooidi periancestrulari. Sample 2G. Scale bar: 500 mm.
199Studi Trent. Sci. Nat., 94 (2014): 195-207
sample, and some are even present with only a single spe-cimen in the entire collection (Tab. 2). Another five species are restricted to a single sample though being locally rela-tively more common, their colony numbers ranging from 3 to 6. Interestingly, all colonies have small sizes.
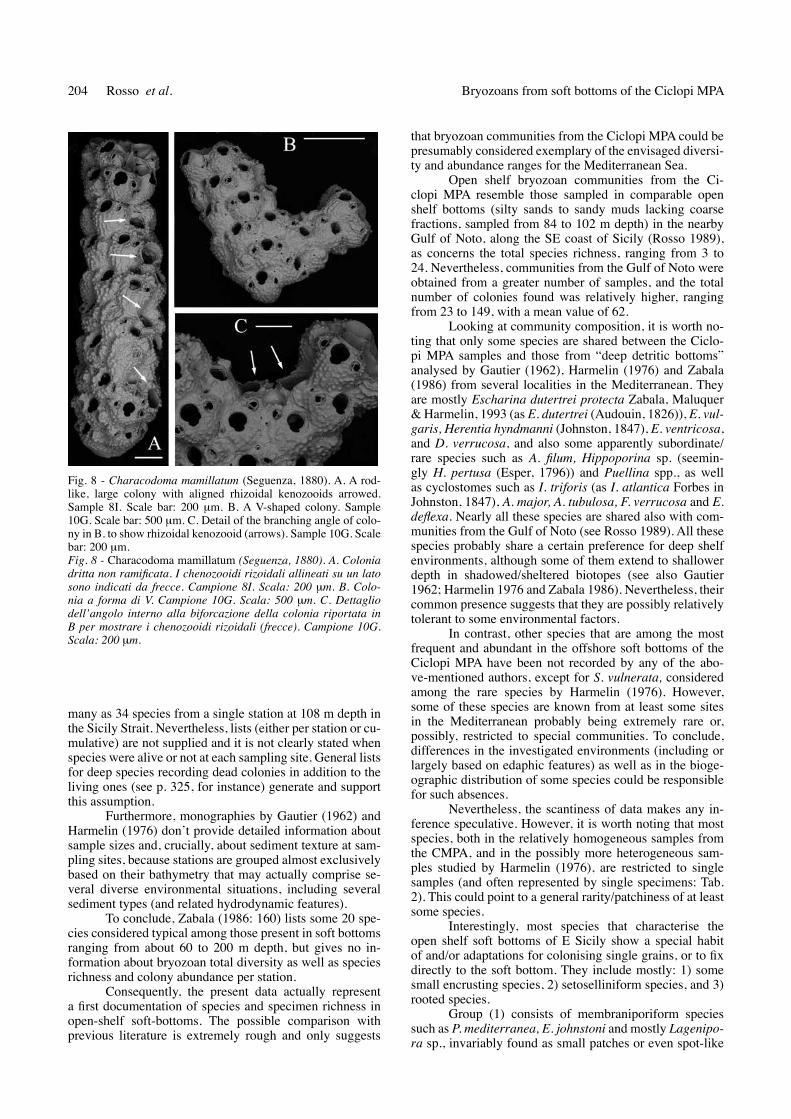
Erect morphotypes (rigid vinculariiforms and ade-onelliforms, and flexible cellariiforms) are subordinate, accounting for a total of 9 species and 18 colonies, some morphological types being absent from some samples (Fig. 2). Erect rigid growth type mostly include cyclostomes, such as Idmidronea triforis (Heller, 1837), Annectocyma tubulosa (Busk, 1875) and Frondipora verrucosa (Lamou-roux, 1821) and also the cheilostomes Buskea dichotoma (Hincks, 1862) and Diporula verrucosa (Peach, 1873). Some of these species exhibit colonies whose sizes are re-latively small in respect to those known from hard, non-particulate bottoms. Particularly, D. verrucosa shows colo-
nies not exceeding 2 cm in the studied samples and are even smaller (Fig. 3A). Bases for their attachment to the sub-stratum are extremely small and they suddenly give rise to small erect portions (Fig. 3B). Even ancestrulae are expo-sed in very young colonies (Fig. 3C) whereas usually they are quickly overgrown by autozooids and/or heterozooids of subsequent generations (rosso personal observation). Erect flexible morphotypes are represented by species of the genera Crisia and Scrupocellaria.
nevertheless, the majority of species and specimens develop encrusting colonies of the membraniporiform growth type, which include a variety of morphologies and organisations (Fig. 2). A unique celleporiform species, Tur-bicellepora corunopus (Wood, 1844), has been detected, which forms a hemispherical colony few millimetres in diameter.
Several species form relatively large unilami-
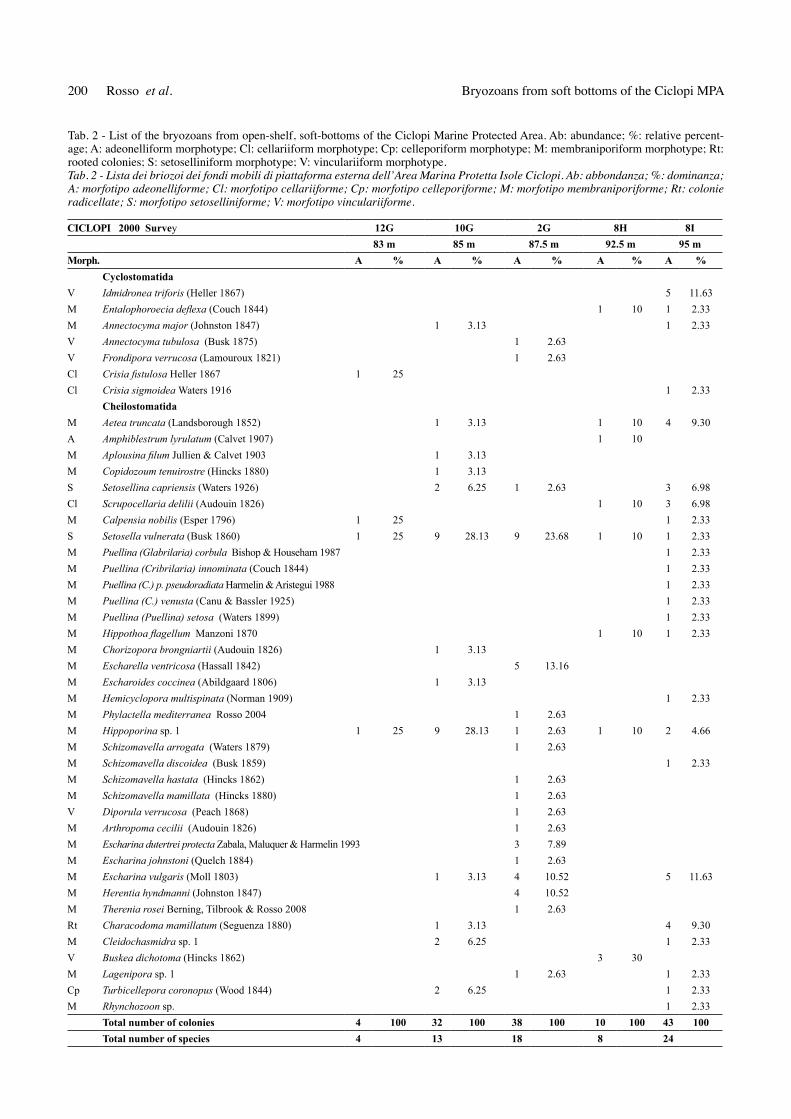
Fig. 4 - Encrusting colonies. A.-B. Escharina vulgaris (Moll, 1803). A. A colony nearly completely enveloping a bivalve fragment. Sample 2G. Scale bar: 1 mm. B. A small colony on a Miniacina miniacea (Pallas, 1766) fragment. Sample 10G. Scale bar: 500 mm. c.-d. Hip-poporina sp. C. Partly broken colony completely overgrowing an unidentified grain. Sample 10G. Scale bar: 1 mm. D. Young colony on an abraded ash grain. Sample 8I. Scale bar: 2 mm. E. Aplousina filum Jullien & calvet, 1903 completely covering a crab fragment. Sample 12G. Scale bar: 2 mm. F. Young colony of Escharella ventricosa (Hassall, 1842) on a bivalve shell fragment. Sample 2G. Scale bar: 1 mm.Fig. 4 - Colonie incrostanti. A.-B. Escharina vulgaris (Moll, 1803). A. Colonia che ricopre quasi completamente un frammento di bivalve. Campione 2G. Scala: 1 mm. E. Giovane colonia su un frammento di Miniacina miniacea (Pallas, 1766). Campione 10G. Scala: 500 mm. C.-D. Hippoporina sp. C. Colonia parzialmente rotta che ricopre completamente un clasto non identificato. Campione 10G. Scala: 1 mm. D. Giovane colonia su un granulo abraso di cenere vulcanica. Campione 8I. Scala: 2 mm. E. Colonia di Aplousina filum Jullien & Calvet, 1903 che ricopre completamente un frammento di crostaceo. Campione 12G. Scala: 2 mm. F. Colonia di Escharella ventricosa (Hassall, 1842) su frammento di bivalve. Campione 2G. Scala: 1 mm.
200 Rosso et al. Bryozoans from soft bottoms of the Ciclopi MPA
tab. 2 - List of the bryozoans from open-shelf, soft-bottoms of the ciclopi Marine Protected Area. Ab: abundance; %: relative percent-age; A: adeonelliform morphotype; cl: cellariiform morphotype; cp: celleporiform morphotype; M: membraniporiform morphotype; rt: rooted colonies; S: setoselliniform morphotype; V: vinculariiform morphotype.Tab. 2 - Lista dei briozoi dei fondi mobili di piattaforma esterna dell’Area Marina Protetta Isole Ciclopi. Ab: abbondanza; %: dominanza; A: morfotipo adeonelliforme; Cl: morfotipo cellariiforme; Cp: morfotipo celleporiforme; M: morfotipo membraniporiforme; Rt: colonie radicellate; S: morfotipo setoselliniforme; V: morfotipo vinculariiforme.
CICLOPI 2000 Survey 12G 10G 2G 8H 8I83 m 85 m 87.5 m 92.5 m 95 m
Morph. A % A % A % A % A % Cyclostomatida V Idmidronea triforis (Heller 1867) 5 11.63M Entalophoroecia deflexa (couch 1844) 1 10 1 2.33M Annectocyma major (Johnston 1847) 1 3.13 1 2.33V Annectocyma tubulosa (Busk 1875) 1 2.63V Frondipora verrucosa (lamouroux 1821) 1 2.63cl Crisia fistulosa Heller 1867 1 25cl Crisia sigmoidea Waters 1916 1 2.33
CheilostomatidaM Aetea truncata (landsborough 1852) 1 3.13 1 10 4 9.30A Amphiblestrum lyrulatum (calvet 1907) 1 10M Aplousina filum Jullien & calvet 1903 1 3.13M Copidozoum tenuirostre (Hincks 1880) 1 3.13s Setosellina capriensis (Waters 1926) 2 6.25 1 2.63 3 6.98cl Scrupocellaria delilii (Audouin 1826) 1 10 3 6.98M Calpensia nobilis (Esper 1796) 1 25 1 2.33s Setosella vulnerata (Busk 1860) 1 25 9 28.13 9 23.68 1 10 1 2.33M Puellina (Glabrilaria) corbula Bishop & Househam 1987 1 2.33M Puellina (Cribrilaria) innominata (couch 1844) 1 2.33M Puellina (C.) p. pseudoradiata Harmelin & Aristegui 1988 1 2.33M Puellina (C.) venusta (canu & Bassler 1925) 1 2.33M Puellina (Puellina) setosa (Waters 1899) 1 2.33M Hippothoa flagellum Manzoni 1870 1 10 1 2.33M Chorizopora brongniartii (Audouin 1826) 1 3.13M Escharella ventricosa (Hassall 1842) 5 13.16M Escharoides coccinea (Abildgaard 1806) 1 3.13M Hemicyclopora multispinata (norman 1909) 1 2.33M Phylactella mediterranea Rosso 2004 1 2.63M Hippoporina sp. 1 1 25 9 28.13 1 2.63 1 10 2 4.66M Schizomavella arrogata (Waters 1879) 1 2.63M Schizomavella discoidea (Busk 1859) 1 2.33M Schizomavella hastata (Hincks 1862) 1 2.63M Schizomavella mamillata (Hincks 1880) 1 2.63V Diporula verrucosa (peach 1868) 1 2.63M Arthropoma cecilii (Audouin 1826) 1 2.63M Escharina dutertrei protecta Zabala, Maluquer & Harmelin 1993 3 7.89M Escharina johnstoni (Quelch 1884) 1 2.63M Escharina vulgaris (Moll 1803) 1 3.13 4 10.52 5 11.63M Herentia hyndmanni (Johnston 1847) 4 10.52M Therenia rosei Berning, tilbrook & Rosso 2008 1 2.63Rt Characodoma mamillatum (seguenza 1880) 1 3.13 4 9.30M Cleidochasmidra sp. 1 2 6.25 1 2.33V Buskea dichotoma (Hincks 1862) 3 30M Lagenipora sp. 1 1 2.63 1 2.33cp Turbicellepora coronopus (Wood 1844) 2 6.25 1 2.33M Rhynchozoon sp. 1 2.33 Total number of colonies 4 100 32 100 38 100 10 100 43 100 Total number of species 4 13 18 8 24
201Studi Trent. Sci. Nat., 94 (2014): 195-207
nar patches, usually on biogenic platy clasts (Figs 4A-I), most being only few millimetres wide, thus mostly falling in the granule size. these species include Escharina vul-garis (Moll, 1803) (Figs 4A-B), Hippoporina sp. (Figs 4c-d), Aplousina filum Jullien & calvet, 1903 (Fig. 4E), Calpensia nobilis (Esper, 1796), Escharoides coccinea (Abildgaard, 1806), Hemicyclopora multispinata (norman, 1909), Escharella ventricosa (Hassall, 1842) (Fig. 4F) and Schizomavella spp. these colonies often largely or entirely cover only one side of the clast selected as substratum. A tendency to envelope clasts has been rarely observed only for C. nobilis and Hippoporina sp. dead specimens (usual-ly decidedly more numerous in the same samples) someti-mes expose fragmented edges, suggesting that at least some specimens of some species were able to continue growing beyond the basal clast after it was completely overgrown on one side, presumably forming narrow rims of basally unattached zooids.
In contrast, Phylactella mediterranea rosso, 2004 (Figs 5A-B) and Escharina johnstoni (Quelch, 1884) (Figs 5c-d) form smaller colonies, often resulting from lobate pauciserial branches that sometimes include more than one growth phase separated by stases. colonies of these spe-cies are present on small clasts that are only partly covered. only larger colonies that comprise several dozen zooids become fertile.
A further species, Lagenipora sp., invariably forms small round colonies consisting of only few zooids (Fig. 5E). these colonies seem to have a predeterminate growth and become mature when comprising only a dozen zooids
(spot colonies sensu Bishop 1989). they grow on clasts as small as 3-4 mm in diameter.
Encrusting uniserial runners are subordinate, and only represented by Aetea truncata (Landsborough, 1852) and Hippothoa flagellum Manzoni, 1870.
rare, large-sized shell fragments of 2-3 cm in dia-meter are usually colonised by several colonies, even be-longing to different species. up to 3-4 being contempora-neously living, plus other dead colonies belonging to the same or other species.
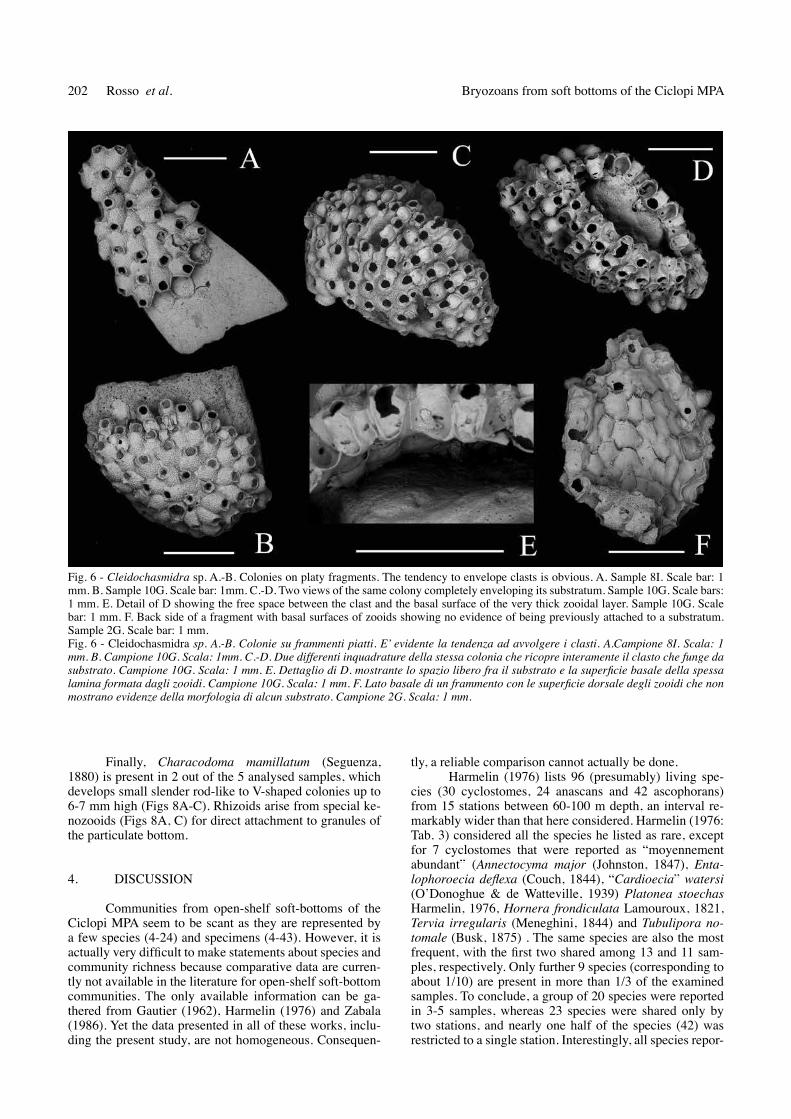
Furthermore, Cleidochasmidra sp., has been found to entirely enclose clasts, particularly those that do not exceed 1-2 mm in diameter (Figs 6A-D). However, the zo-oidal basal surfaces are occasionally cemented only discon-tinuously on the substratum. Some small platy clasts have been found that were encrusted only on one side whereas the zooids are 300-500 mm aloof from the clast surface on the opposite side (Figs 6C-D). The zooids in these colony regions are very conical and laterally compressed (Fig. 6E). these uncemented laminas are particularly fragile, and broken pieces that entirely consist of apparently unattached zooids have been found in the dead material (Fig. 6F).
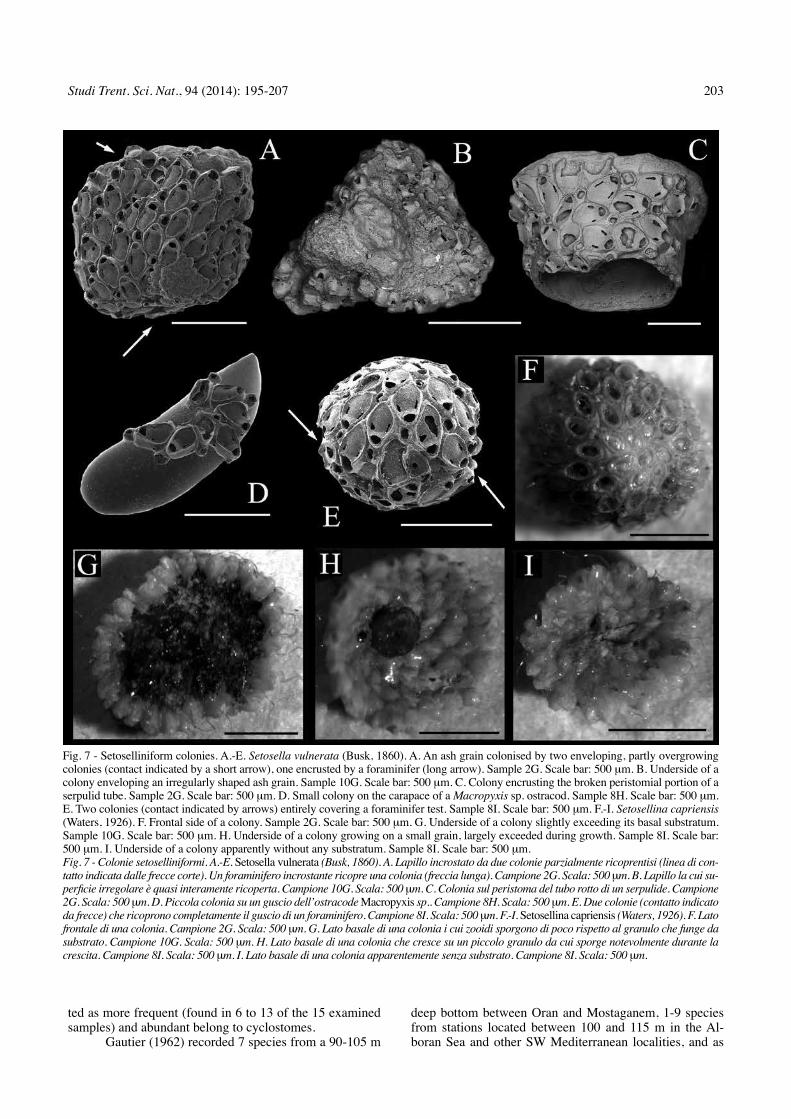
two further species, Setosellina capriensis (Waters, 1926) and S. vulnerata, develop setoselliniform colonies that are present in all samples, accounting for nearly 10 to over 34% (Fig. 2). These colonies result from spirally-budded zooids that regularly alternate with vibracular hete-rozooids. colonies develop on small clasts, including gra-nules but mostly very coarse-to-medium sands (Figs 7A-I, and particularly Figs 7D-E, G-H).
Fig. 5. A.-B. Phylactella mediterranea rosso, 2004. Sample 2G. A. Scale bar: 2 mm. B. detail of A. Scale bar: 500 mm. c.-d. Escharina johnstoni (Quelch, 1884). C. Fertile colony. Sample 2G. Scale bar: 1 mm. D. Young colony inside a bivalve shell. Sample 8I. Scale bar: 2 mm. E. A small fertile colony of Lagenipora sp. Sample 2G. Scale bar: 1 mm.Fig. 5 - A.-B. Phylactella mediterranea Rosso, 2004. Campione 2G. A. Scala: 2 mm. B. Dettaglio di A. Scala: 500 mm. C.-D. Escharina johnstoni (Quelch, 1884). C. Colonia fertile. Campione 2G. Scala: 1 mm. D. Colonia giovanile sulla superficie interna di un bivalve. Campione 8I. Scala: 2 mm. E. Piccola colonia fertile di Lagenipora sp. Campione 2G. Scala: 1 mm.
202 Rosso et al. Bryozoans from soft bottoms of the Ciclopi MPA
Finally, Characodoma mamillatum (Seguenza, 1880) is present in 2 out of the 5 analysed samples, which develops small slender rod-like to V-shaped colonies up to 6-7 mm high (Figs 8A-C). Rhizoids arise from special ke-nozooids (Figs 8A, C) for direct attachment to granules of the particulate bottom.
4. dIScuSSIon
communities from open-shelf soft-bottoms of the ciclopi MPA seem to be scant as they are represented by a few species (4-24) and specimens (4-43). However, it is actually very difficult to make statements about species and community richness because comparative data are curren-tly not available in the literature for open-shelf soft-bottom communities. the only available information can be ga-thered from Gautier (1962), Harmelin (1976) and Zabala (1986). Yet the data presented in all of these works, inclu-ding the present study, are not homogeneous. consequen-
tly, a reliable comparison cannot actually be done.Harmelin (1976) lists 96 (presumably) living spe-
cies (30 cyclostomes, 24 anascans and 42 ascophorans) from 15 stations between 60-100 m depth, an interval re-markably wider than that here considered. Harmelin (1976: tab. 3) considered all the species he listed as rare, except for 7 cyclostomes that were reported as “moyennement abundant” (Annectocyma major (Johnston, 1847), Enta-lophoroecia deflexa (couch, 1844), “Cardioecia” watersi (o’donoghue & de Watteville, 1939) Platonea stoechas Harmelin, 1976, Hornera frondiculata Lamouroux, 1821, Tervia irregularis (Meneghini, 1844) and Tubulipora no-tomale (Busk, 1875) . the same species are also the most frequent, with the first two shared among 13 and 11 sam-ples, respectively. only further 9 species (corresponding to about 1/10) are present in more than 1/3 of the examined samples. to conclude, a group of 20 species were reported in 3-5 samples, whereas 23 species were shared only by two stations, and nearly one half of the species (42) was restricted to a single station. Interestingly, all species repor-
Fig. 6 - Cleidochasmidra sp. A.-B. colonies on platy fragments. the tendency to envelope clasts is obvious. A. Sample 8I. Scale bar: 1 mm. B. Sample 10G. Scale bar: 1mm. c.-d. two views of the same colony completely enveloping its substratum. Sample 10G. Scale bars: 1 mm. E. detail of d showing the free space between the clast and the basal surface of the very thick zooidal layer. Sample 10G. Scale bar: 1 mm. F. Back side of a fragment with basal surfaces of zooids showing no evidence of being previously attached to a substratum. Sample 2G. Scale bar: 1 mm. Fig. 6 - Cleidochasmidra sp. A.-B. Colonie su frammenti piatti. E’ evidente la tendenza ad avvolgere i clasti. A.Campione 8I. Scala: 1 mm. B. Campione 10G. Scala: 1mm. C.-D. Due differenti inquadrature della stessa colonia che ricopre interamente il clasto che funge da substrato. Campione 10G. Scala: 1 mm. E. Dettaglio di D. mostrante lo spazio libero fra il substrato e la superficie basale della spessa lamina formata dagli zooidi. Campione 10G. Scala: 1 mm. F. Lato basale di un frammento con le superficie dorsale degli zooidi che non mostrano evidenze della morfologia di alcun substrato. Campione 2G. Scala: 1 mm.
203Studi Trent. Sci. Nat., 94 (2014): 195-207
ted as more frequent (found in 6 to 13 of the 15 examined samples) and abundant belong to cyclostomes.
Gautier (1962) recorded 7 species from a 90-105 m
deep bottom between oran and Mostaganem, 1-9 species from stations located between 100 and 115 m in the Al-boran Sea and other SW Mediterranean localities, and as
Fig. 7 - Setoselliniform colonies. A.-E. Setosella vulnerata (Busk, 1860). A. An ash grain colonised by two enveloping, partly overgrowing colonies (contact indicated by a short arrow), one encrusted by a foraminifer (long arrow). Sample 2G. Scale bar: 500 mm. B. underside of a colony enveloping an irregularly shaped ash grain. Sample 10G. Scale bar: 500 mm. c. colony encrusting the broken peristomial portion of a serpulid tube. Sample 2G. Scale bar: 500 mm. d. Small colony on the carapace of a Macropyxis sp. ostracod. Sample 8H. Scale bar: 500 mm. E. two colonies (contact indicated by arrows) entirely covering a foraminifer test. Sample 8I. Scale bar: 500 mm. F.-I. Setosellina capriensis (Waters, 1926). F. Frontal side of a colony. Sample 2G. Scale bar: 500 mm. G. underside of a colony slightly exceeding its basal substratum. Sample 10G. Scale bar: 500 mm. H. underside of a colony growing on a small grain, largely exceeded during growth. Sample 8I. Scale bar: 500 mm. I. underside of a colony apparently without any substratum. Sample 8I. Scale bar: 500 mm. Fig. 7 - Colonie setoselliniformi. A.-E. Setosella vulnerata (Busk, 1860). A. Lapillo incrostato da due colonie parzialmente ricoprentisi (linea di con-tatto indicata dalle frecce corte). Un foraminifero incrostante ricopre una colonia (freccia lunga). Campione 2G. Scala: 500 mm. B. Lapillo la cui su-perficie irregolare è quasi interamente ricoperta. Campione 10G. Scala: 500 mm. C. Colonia sul peristoma del tubo rotto di un serpulide. Campione 2G. Scala: 500 mm. D. Piccola colonia su un guscio dell’ostracode Macropyxis sp.. Campione 8H. Scala: 500 mm. E. Due colonie (contatto indicato da frecce) che ricoprono completamente il guscio di un foraminifero. Campione 8I. Scala: 500 mm. F.-I. Setosellina capriensis (Waters, 1926). F. Lato frontale di una colonia. Campione 2G. Scala: 500 mm. G. Lato basale di una colonia i cui zooidi sporgono di poco rispetto al granulo che funge da substrato. Campione 10G. Scala: 500 mm. H. Lato basale di una colonia che cresce su un piccolo granulo da cui sporge notevolmente durante la crescita. Campione 8I. Scala: 500 mm. I. Lato basale di una colonia apparentemente senza substrato. Campione 8I. Scala: 500 mm.
204 Rosso et al. Bryozoans from soft bottoms of the Ciclopi MPA
many as 34 species from a single station at 108 m depth in the Sicily Strait. nevertheless, lists (either per station or cu-mulative) are not supplied and it is not clearly stated when species were alive or not at each sampling site. General lists for deep species recording dead colonies in addition to the living ones (see p. 325, for instance) generate and support this assumption.
Furthermore, monographies by Gautier (1962) and Harmelin (1976) don’t provide detailed information about sample sizes and, crucially, about sediment texture at sam-pling sites, because stations are grouped almost exclusively based on their bathymetry that may actually comprise se-veral diverse environmental situations, including several sediment types (and related hydrodynamic features).
to conclude, Zabala (1986: 160) lists some 20 spe-cies considered typical among those present in soft bottoms ranging from about 60 to 200 m depth, but gives no in-formation about bryozoan total diversity as well as species richness and colony abundance per station.
consequently, the present data actually represent a first documentation of species and specimen richness in open-shelf soft-bottoms. the possible comparison with previous literature is extremely rough and only suggests
that bryozoan communities from the ciclopi MPA could be presumably considered exemplary of the envisaged diversi-ty and abundance ranges for the Mediterranean Sea.
open shelf bryozoan communities from the ci-clopi MPA resemble those sampled in comparable open shelf bottoms (silty sands to sandy muds lacking coarse fractions, sampled from 84 to 102 m depth) in the nearby Gulf of noto, along the SE coast of Sicily (rosso 1989), as concerns the total species richness, ranging from 3 to 24. nevertheless, communities from the Gulf of noto were obtained from a greater number of samples, and the total number of colonies found was relatively higher, ranging from 23 to 149, with a mean value of 62.
Looking at community composition, it is worth no-ting that only some species are shared between the ciclo-pi MPA samples and those from “deep detritic bottoms” analysed by Gautier (1962), Harmelin (1976) and Zabala (1986) from several localities in the Mediterranean. they are mostly Escharina dutertrei protecta Zabala, Maluquer & Harmelin, 1993 (as E. dutertrei (Audouin, 1826)), E. vul-garis, Herentia hyndmanni (Johnston, 1847), E. ventricosa, and D. verrucosa, and also some apparently subordinate/rare species such as A. filum, Hippoporina sp. (seemin-gly H. pertusa (Esper, 1796)) and Puellina spp., as well as cyclostomes such as I. triforis (as I. atlantica Forbes in Johnston, 1847), A. major, A. tubulosa, F. verrucosa and E. deflexa. nearly all these species are shared also with com-munities from the Gulf of noto (see rosso 1989). All these species probably share a certain preference for deep shelf environments, although some of them extend to shallower depth in shadowed/sheltered biotopes (see also Gautier 1962; Harmelin 1976 and Zabala 1986). nevertheless, their common presence suggests that they are possibly relatively tolerant to some environmental factors.
In contrast, other species that are among the most frequent and abundant in the offshore soft bottoms of the ciclopi MPA have been not recorded by any of the abo-ve-mentioned authors, except for S. vulnerata, considered among the rare species by Harmelin (1976). However, some of these species are known from at least some sites in the Mediterranean probably being extremely rare or, possibly, restricted to special communities. to conclude, differences in the investigated environments (including or largely based on edaphic features) as well as in the bioge-ographic distribution of some species could be responsible for such absences.
nevertheless, the scantiness of data makes any in-ference speculative. However, it is worth noting that most species, both in the relatively homogeneous samples from the cMPA, and in the possibly more heterogeneous sam-ples studied by Harmelin (1976), are restricted to single samples (and often represented by single specimens: tab. 2). this could point to a general rarity/patchiness of at least some species.
Interestingly, most species that characterise the open shelf soft bottoms of E Sicily show a special habit of and/or adaptations for colonising single grains, or to fix directly to the soft bottom. they include mostly: 1) some small encrusting species, 2) setoselliniform species, and 3) rooted species.
Group (1) consists of membraniporiform species such as P. mediterranea, E. johnstoni and mostly Lagenipo-ra sp., invariably found as small patches or even spot-like
Fig. 8 - Characodoma mamillatum (Seguenza, 1880). A. A rod-like, large colony with aligned rhizoidal kenozooids arrowed. Sample 8I. Scale bar: 200 mm. B. A V-shaped colony. Sample 10G. Scale bar: 500 mm. c. detail of the branching angle of colo-ny in B. to show rhizoidal kenozooid (arrows). Sample 10G. Scale bar: 200 mm.Fig. 8 - characodoma mamillatum (Seguenza, 1880). A. Colonia dritta non ramificata. I chenozooidi rizoidali allineati su un lato sono indicati da frecce. Campione 8I. Scala: 200 mm. B. Colo-nia a forma di V. Campione 10G. Scala: 500 mm. C. Dettaglio dell’angolo interno alla biforcazione della colonia riportata in B per mostrare i chenozooidi rizoidali (frecce). Campione 10G. Scala: 200 mm.

205Studi Trent. Sci. Nat., 94 (2014): 195-207
colonies. these taxa appear selectively adapted to colonise small clasts owing to the small sizes of their fertile colonies, as it has been already suggested for P. mediterranea (rosso 2004). Interestingly, even colonies of these species obser-ved on large hard substrata, mostly from the coralligenous Biocoenosis, invariably grow as small patches (Ar, perso-nal observations). the same adaptation is shared by some species listed also by Gautier (1962) and Harmelin (1976), such as E. dutertrei protecta and, mostly, H. hyndmanni, that commonly grow as very small lobate colonies (see Berning et al. 2008). In contrast, among other encrusting taxa in the studied samples most are able to develop exten-sive patches (E. vulgaris, E. coccinea, E. ventricosa) to very large sheet-like colonies (C. nobilis, Schizomavella spp.) when they have enough substratum space available or special bottom conditions (Ar personal observation). ne-vertheless, in the studied samples the growth of these spe-cies appears strongly affected by the size of available sedi-ment clasts, and consequently, their colonies don’t usually exceed one to several square millimetres and exceptionally one square centimetre. this different behaviour could in-dicate that open shelf soft bottoms characterised by sandy-silt to silty-sand texture, with a subordinate coarse fraction, represent the elective biotope for the former species but only a subordinate opportunity of colonisation for the lat-ter ones. Gravelly-sized, mostly bioclastic fraction actually supply short-lived/unstable substrata (sensu McKinney & Jackson 1989) for bryozoan colonisation.
Furthermore, the usual exploitation of only one side of the clasts (particularly of those showing platy morpho-logies) seems to limit colony expansion, although some co-lony edges were clearly broken at the clast boundary (see results), which point to the ability of at least some species/colonies to grow free beyond the encrusted grain, above the sediment, as reported also by McKinney & Jaklin (1993). Interestingly, it has been sometimes observed that the same colony covers a platy fragment on both sides but only along one border. only very few species, such as C. nobilis, Clei-dochasmidra sp. (elsewhere reported as C. canakkalense, but it is in need of revision), and E. vulgaris, were rarely able to largely cover, or completely envelop, some small and relatively spherical grains. this ability is relatively common and well known for at least the first species that is able not only to envelop its grain substratum but to form true bryoliths (or circumrotatory masses) through self over-growing laminas (Poluzzi & coppa 1991; McKinney & Jaklin 1993; Moissette et al. 2010; Ar personal observa-tions). the observed general polarity in the colonisation, above all for platy clasts, points to their position at the sea floor (with one exposed side/border and the other one in contact with, or partly buried by, the sediment), and to their stability for some time, at least corresponding to the colony life span. nevertheless, the presence of some completely enveloped clasts testifies to the possibility that smaller bot-tom elements can be at least periodically turned around.
Cleidochasmidra sp. is a relatively rare species that seems to be restricted to particulate bottoms from mid- to shallow open shelf environments from about 50 to 95 m (rosso 1996a; 1996b). It has the ability to entirely envelo-pe clasts but also to develop very elevated conical colonies on small grains, even to form partially free laminas (Fig. 6), a condition observed also in fossil Pliocene specimens (Pizzaferri & Berning 2007). the position that these colo-
nies take on the bottom is unknown but they could be able to rest free on the sediment.
Species from group (2) consist of only two species: S. vulnerata and S. capriensis, both specially adapted to co-lonise particulate bottoms (rosso 2008). Particularly, while the former species is less selective for the size of the sub-stratum grain, and some colonies even encrust relatively large, usually biogenic clasts (Ar personal observations), S. capriensis is more selective and prefers smaller, usual-ly sand-sized, clasts (rosso 2008). Interestingly, although both species could be considered as free-living because they live on single, potentially mobile grains, most S. ca-priensis colonies become cup-shaped and largely develop beyond the encrusted substratum, thus resembling lunuli-tiforms (rosso 2008). of those with a free mode of life, these species possibly represent the best adaptation to co-lonise the investigated bottoms. Again, the presence and abundance of these two species seems to be distinctive of the present communities, although they are not restricted to the investigated bottoms and their distribution largely over-laps with that of Cleidochasmidra sp. and extends down to about 130 m (rosso 2008).
Group (3) is exclusively represented by C. mamilla-tum, a species specifically adapted to thrive in muddy se-diments that has been reported from the mid- to deep shelf down to upper slope environments (rosso 1999). colonies fix through thin-walled but relatively thick tubular rhizoids, possibly rising only slightly above the sediment-water in-terface, at least in the adult stage (rosso 1999).
An elevation from the sediment water interface is also gained by the few erect species present, including most cyclostomes and the cheilostomes D. verrucosa and B. dichotoma. In the studied samples these species deve-lop colonies smaller than their normal sizes. this has been observed particularly for D. verrucosa, often represented by young colonies (Figs 3B-C), and whose adult colonies were presumably smaller than usually sampled in the Me-diterranean Sea, which often reach heights of 5-6 cm (Ar personal observations).
5. concLuSIonS
the main features of open shelf soft bottom bryozo-an communities of the ciclopi MPA seem to be: low total (and per station) species richness and presumable colony scantiness; possible patchiness presumably linked to the availability of suitable substrata; presence of some distin-ctive species so far not recorded from deep-shelf soft-bot-toms; special morphological adaptations for the colonisa-tion of sand- to gravel-sized, mostly biogenic clasts.
Despite difficulties to compare the bryozoan com-munities from the open-shelf soft-bottoms of the ciclopi MPA with others from deep shelf detritic bottoms repor-ted in the literature, a distinctive feature appears to be the abundant occurrence of encrusting species and colonies, to-gether with “free-living” morphotypes. In contrast, Zabala (1986) remarked that in deep detritic bottoms encrusting species are subordinate, as substrata suitable for colonisa-tion are scarce, whereas species developing erect colonies, as an adaptation to escape sediment burial, are dominant. Edaphic features are of paramount importance at depths where other parameters, such as light, become less signi-

206 Rosso et al. Bryozoans from soft bottoms of the Ciclopi MPA
ficant (Harmelin 1976). Particularly for the Ciclopi MPA bryozoan communities dominated by encrusting morphot-ypes, sediment texture (including the presence and abun-dance of the gravel fraction as the main source of colonisa-ble substrata) is strongly linked to near-bottom water-ener-gy, and drives bryozoan colonisation and the persistence of populations. High dominance of encrusting bryozoans from soft bottoms have been reported by Winston & Håkansson (1986) when analysing interstitial bryozoans from shallow water, well-sorted, medium to coarse sands from capron Shoal (Florida). These authors discussed the importance of the presence of a biogenic component in the sediment that allows colonisation by very small sized bryozoans. Irregularly shaped biogenic clasts would have enhanced the creation of large pore spaces in between particles as well as water circulation, and oxygen and food availability in interstices. this hypothesis was, in turn, questioned by Winston & Migotto (2005) as interstitial bryozoan associa-tions were also found in sediments with wider ranges of composition, texture and sorting. In the ciclopi MPA case, where bottoms consist of unsorted silty sediments with su-bordinate coarse fractions, the colony sizes of encrusting bryozoans are larger than those of true interstitial species colonising shallow water, unstable sandy sediments, as re-ported by Winston & Håkansson (1986) and by Winston & Vieira (2013). the composition and structure of the zoarial association, the comparably large colony size, and the po-sition and development of encrusters on single grains all point to an epibenthic life style for the bryozoans from the ciclopi MPA. consequently, the most important factors for bryozoan colonisation in the present instance seems to be the position of clasts (large enough to be colonised) on the bottom and their persistence at the sediment-water inter-face, which strongly depends on siltation and deposition rate. An important role in this respect is probably played by bottom currents sweeping near shelf-break bottoms, ham-pering deposition and/or partly removing fine particles. In this way clasts (supplied by local biogenic production, mo-stly by infaunal bivalves, and by the continuous input of volcanic ashes) persist at the sediment-water interface for a time span sufficient for larval settlement and growth of small colonies that often becoming fertile at early stages. Like for shallow-water interstitial faunas, the maintenance of these bryozoan associations on ephemeral substrata re-lies on larval recruitment, but in the former case the asso-ciation is driven by larval recruitment/abrasion (McKinney & Jackson 1989) whereas for deep-water, open-shelf, soft bottoms the major constrain could be larval recruitment/siltation-burying.
AcKnoWLEdGEMEntS
This paper was financially supported through PRA grants to Ar. Sampling was granted by the Ministero dell’Ambiente through capitaneria di Porto of catania. thanks are due to A. Viola for SEM assistance and to A. Viola and M. Vagliasindi (dip. Scienze Biologiche, Geo-logiche e Ambientali, university of catania) for textural analyses. I. di Geronimo (formerly at the university of catania) is kindly acknowledged for his help in the con-struction of the bionomic map. catania Palaeoecological research Group: contribution n. XXX.
REFEREnCES
Balduzzi A. & Rosso A., 2003 - Briozoi. In: Cicogna F., Bianchi n.C., Ferrari G. & Forti P. (eds), Grotte marine: cinquant’an-ni di ricerca in Italia, cLEM-onLuS Ministero Ambiente e tutela territorio: 195-202.
Berning B., tilbrook K.J. & rosso A., 2008 - revision of the north-eastern Atlantic and Mediterranean species of the gen-era Herentia and Therenia (Bryozoa: cheilostomata). Journ. Nat. Hist., 42/21-22: 1509-1547.
Bishop J.d.d., 1989 - colony form and the exploitation of spatial refuges by encrusting bryozoans. Biol. Rev., 64: 197-218.
Gautier Y.V., 1959 - Essai d’etude quantitative sur les Bryozoaires d’un fond coralligène à Gorgones. Rec. Trav. Stn. Mar. En-doume, 26/16: 137-142.
Gautier Y.V., 1962 - recherches ecologiques sur les Bryozoaires chilostomes en Méditerranée occidentale. Rec. Trav. Stat. Mar. Endoume, 38/24: 1-435.
Harmelin J.G., 1969 - Bryozoaires des grottes sous-marines obscures de la région Marseillaise: faunistique et écologie. Téthys, 1/3: 793-806.
Harmelin J.G., 1976 - Le sous-ordre des tubuliporina (Bryozoai-res cyclostomes) en Méditerranée: Ecologie et systémati-que. Mem. Inst. Oceanogr. Monaco, 10: 1-326.
Harmelin J.G., 1980 - Etablissement des communautés de subs-trats durs en milieu obscur. résultats préliminaires d’une expérience à long terme en Méditerranée. Mem. Biol. Mar. Oceanogr., 10: 29-52
Harmelin J.G., 1985 - organisation spatiale des communautés des grottes sous-marines de Méditerranée. Rapp. Comm. Int. Mer Médit., 29/5: 149-153.
Harmelin J.G., 1986 - Patterns in the distribution of bryozoans in the Mediterranean marine caves. Stygologia, 2/1-2: 10-25.
Harmelin J.G., 1997 - diversity of bryozoans in a Mediterranean sublittoral cave with bathyal-like conditions: role of disper-sal processes and local factors. Mar. Ecol. Progr. Ser., 153: 139-152.
Harmelin J.G., 2000 - Ecology of cave and cavity dwelling bryo-zoans. In: Herrera cubilla A. & Jackson J.B.c. (eds), Pro-ceedings of the 11th International Bryozoology Association Conference: 38-55.
Laubier L., 1966 - Le coralligène des Albères: monographie bio-cénotique. Ann. Inst. Oceanogr. Monaco, 43: 137-316.
Mastrototaro F., D’Onghia G., Corriero G., Matarrese A., Maio-rano P., Panetta P., Gherardi M., Longo c., rosso A., Sciuto F., Sanfilippo R., Gravili C., Boero F, Taviani M. & Tursi A., 2010 - Biodiversity of the white coral bank off cape Santa Maria di Leuca (Mediterranean Sea): An update. Deep Sea Research II, 57/5-6: 412-430.
McKinney F.K. & Jackson J.B.C., 1989 - Bryozoan Evolution. unwin Hyman, Boston, 238 pp.
McKinney F.K. & Jaklin A., 1993 - Living populations of free-living bryozoans: implications for post-Paleozoic decline of the growth habit. Lethaia, 26: 171-179.
McKinney F.K. & Jaklin A., 2000 - Spatial niche partitioning in the Cellaria meadow epibiont association, northern Adriatic Sea. Cah. Biol. Mar., 41: 1-17.
McKinney F.K. & Jaklin A., 2001 - Sediment accumulation in a shallow-water meadow carpeted by a small erect bryozoan. Sedim. Geol., 145: 397-410.
Moissette P., cornée J.-J. & Kosperidou E., 2010 - Pleistocene rolling stones or large bryozoan nodules in a mixed silici-clastic carbonate environmeny (rhodes, Greece). Palaios,

207Studi Trent. Sci. Nat., 94 (2014): 195-207
25/1: 24-39.Pérès J.M. & Picard J., 1964 - nouveau manuel de bionomie
benthique de la Mer Méditerranée. Rec. Trav. Stat. Mar. En-doume, 31: pp. 1-137.
Pizzaferri c. & Berning B., 2007 - taxonomic notes on some cheilostome Bryozoa from the Pliocene of the western Emil-ia region (n Italy). Riv. It. Paleont. Strat., 113/1: 97-108.
Poluzzi A. & coppa M.G., 1991 - Zoarial strategies to win sub-stratum space in Calpensia nobilis (Esper). In: Bigey F.P. & d’Hondt J.-L. (eds), Bryozoa: Living and Fossil. Soc. Sci. Nat. Ouest France. Mem. hors série, 337-360. nantes, France.
rosso A., 1989 - Contributo alla conoscenza di alcuni popola-menti, tanatocenosi e tafocenosi a briozoi di alcuni fondi mobili circalitorali. unpublished Phd thesis, Messina.
rosso A., 1996a - Valutazione della biodiversità in Mediterraneo: l’esempio dei popolamenti a briozoi della Biocenosi del de-tritico costiero. Biol. Mar. Medit., 3/1: 58-65.
rosso A., 1996b - Popolamenti e tanatocenosi a Briozoi di fondi mobili circalitorali del Golfo di noto (Sicilia SE). Natur. Si-ciliano, ser. 4, 20/3-4: 189-225.
rosso A., 1999 - recent and fossil species of Characodoma Ma-pleston, 1900 (Bryozoa) from the Mediterranean with de-scription of two new species. Journ. Nat. Hist., 33: 415-437.
rosso A., 2001 - Cartografia topografica e biocenotica dei fon-dali mobili della Riserva Marina “Isole Ciclopi”. Ministero dell’Ambiente - capitaneria di Porto - università degli Studi di catania, 119 pp., with annexed sedimentological and bio-nomical maps.
rosso A., 2004 - two new species of Phylactella (Bryozoa chei-lostomatida) from the Mediterranean area belonging to the P. labrosa (Busk) complex of species. Journ. Nat. Hist., 38/20: 2655-2668.
rosso A., 2008 - Mediterranean setoselliniforms and their exploi-tation of small-sized substrates. In: Winston J., Key M.M. Jr. & Hageman S.J. (eds), Bryozoan Research 2007: Pro-ceedings of the 14th International Bryozoology Association
conference, Boone, north carolina, July 1-8, 2007, Virginia Mus. Natural History Spec. Publ., 15: 261-268.
rosso A., chimenz Gusso c. & Balduzzi A., 2010. Bryozoa. In: Relini G. Checklist of flora and fauna from the Italian seas (part II). Biol. Mar. Medit., 17/suppl.: 589-615.
Rosso A., Di Martino E., Sanfilippo R. & Di Martino V., 2012 - Bryozoan communities and thanatocoenoses from Subma-rine caves in the Plemmirio Marine Protected Area (SE Sicily). In: Ernst A., Schäfer P., Scholz (eds), Bryozoan Studies 2010. Proceedings of the 15th IBA Conference, Kiel, Germany. Lecture Notes in Earth System Sciences, 143: 251-269, Springer, Berlin, Heidelberg.
Rosso A. & Sanfilippo R., 2009 - The contribution of bryozoans and serpuloideans to coralligenous concretions from SE Si-cily. In: unEP-MAP-rAc/SPA, Proceedings of the First Symposium on the Coralligenous and other calcareous bio-concretions of the Mediterranean Sea (Tabarka, 15-16 Ja-nuary, 2009): 123- 128.
Rosso A., Vertino A., Di Geronimo I., Sanfilippo R., Sciuto F., Di Geronimo r., Violanti d., corselli c., taviani M., Mastroto-taro F. & Tursi A., 2010 - Hard and soft-bottom thanatofacies from the Santa Maria di Leuca deep-water coral province, Mediterranean. Deep Sea Research II., 57/5-6: 360-379.
Winston J.E. & Håkansson E., 1986 - the interstitial bryozoan fauna from Capron Shoal, Florida. Am. Mus. Novit., 2865: 1-50.
Winston J.E. & Migotto A. E., 2005 - A new encrusting interstitial marine fauna from Brazil. Invert. Biology, 124/1: 79-87.
Winston J.E. & Vieira M.L., 2013 - Systematics of interstitial en-crusting bryozoans from southeastern Brazil. Zootaxa, 3710 (2):101-146.
Zabala M., 1986 - Fauna dels bryozous dels Països Catalans. Inst. Estudis catalans, Sec. ciences, Barcelona, 84: 1-833.
Zabala M., Maluquer P. & Harmelin J.-G., 1993 - Epibiotic Bryo-zoans on deep-water scleractinian corals from the catalonia slope (western Mediterranean, Spain, France). Sci. Mar., 57: 65-78.

Related Documents











