Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms Kim B. Barken, 1 Sünje J. Pamp, 1 Liang Yang, 1 Morten Gjermansen, 1 Jacob J. Bertrand, 2,3 Mikkel Klausen, 1 Michael Givskov, 1 Cynthia B. Whitchurch, 4 Joanne N. Engel 3,5,6 and Tim Tolker-Nielsen 1 * 1 Centre for Biomedical Microbiology, Technical University of Denmark, DK-2800, Lyngby, Denmark. 2 Department of Biochemistry and Biophysics, 5 Department of Medicine, 6 Department of Microbiology and Immunology, 3 Program in Microbial Pathogenesis and Host Defense, University of California, San Francisco, CA 94143-0654, USA. 4 Department of Microbiology, Monash University, Vic. 3800, Australia. Summary When grown as a biofilm in laboratory flow chambers Pseudomonas aeruginosa can develop mushroom- shaped multicellular structures consisting of distinct subpopulations in the cap and stalk portions. We have previously presented evidence that formation of the cap portion of the mushroom-shaped structures in P. aeruginosa biofilms occurs via bacterial migra- tion and depends on type IV pili (Mol Microbiol 50: 61–68). In the present study we examine additional factors involved in the formation of this multicellular substructure. While pilA mutants, lacking type IV pili, are deficient in mushroom cap formation, pilH and chpA mutants, which are inactivated in the type IV pili-linked chemosensory system, showed only minor defects in cap formation. On the contrary, fliM mutants, which are non-flagellated, and cheY mutants, which are inactivated in the flagellum-linked chemotaxis system, were largely deficient in cap formation. Experiments involving DNase treatment of developing biofilms provided evidence that extra- cellular DNA plays a role in cap formation. Moreover, mutants that are deficient in quorum sensing- controlled DNA release formed microcolonies upon which wild-type bacteria could not form caps. These results constitute evidence that type IV pili, flagellum- mediated motility and quorum sensing-controlled DNA release are involved in the formation of mature multicellular structures in P. aeruginosa biofilms. Introduction During the last 10 years there has been a rapidly increas- ing recognition of microbial biofilms as a highly significant topic in microbiology with relevance for a variety of areas in our society including the environment, industry and human health. It is generally believed that these struc- tured communities constitute the dominant mode of microbial life (Davey and O’Toole, 2000; Hall-Stoodley et al., 2004). Analysis of global gene expression by means of transcriptomics and proteomics has failed to provide evidence of a specific developmental biofilm programme (Hentzer et al., 2004). On the contrary, high- resolution experimental approaches provide increasing evidence that biofilms are complex and dynamic com- munities in which substantial phenotypic diversification allows microorganisms to adapt to different environments (Watnick and Kolter, 2000; Klausen et al., 2006). Under- standing the adaptive pathways to biofilm formation will eventually make it possible to manipulate the develop- ment of biofilms in nature and disease. The use of flow chamber in vitro set-ups and confocal laser scanning microscopy (CLSM) has provided knowl- edge about mechanisms involved in biofilm formation. Pseudomonas aeruginosa, a model organism in biofilm research, can, dependent on the conditions, form biofilms with distinct spatial structures. For example, flat biofilms are formed in flow chambers irrigated with citrate minimal medium (Klausen et al., 2003a), while heterogeneous biofilms with mushroom-shaped multicellular structures are formed in flow chambers irrigated with glucose minimal medium (Klausen et al., 2003b). The formation of the mushroom-shaped structures in glucose-grown P. aeruginosa biofilms evidently occurs in a sequential process involving a non-motile bacterial subpopulation which forms the initial microcolonies by growth in certain Received 26 January, 2008; accepted 9 April, 2008. *For correspondence. E-mail [email protected]; Tel. (+45) 45 25 27 93; Fax (+45) 45 88 73 28. Environmental Microbiology (2008) 10(9), 2331–2343 doi:10.1111/j.1462-2920.2008.01658.x © 2008 The Authors Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Roles of type IV pili, flagellum-mediated motilityand extracellular DNA in the formation ofmature multicellular structures in Pseudomonasaeruginosa biofilms
Kim B. Barken,1 Sünje J. Pamp,1 Liang Yang,1
Morten Gjermansen,1 Jacob J. Bertrand,2,3
Mikkel Klausen,1 Michael Givskov,1
Cynthia B. Whitchurch,4 Joanne N. Engel3,5,6 andTim Tolker-Nielsen1*1Centre for Biomedical Microbiology, TechnicalUniversity of Denmark, DK-2800, Lyngby, Denmark.2Department of Biochemistry and Biophysics,5Department of Medicine, 6Department of Microbiologyand Immunology, 3Program in Microbial Pathogenesisand Host Defense, University of California, SanFrancisco, CA 94143-0654, USA.4Department of Microbiology, Monash University, Vic.3800, Australia.
Summary
When grown as a biofilm in laboratory flow chambersPseudomonas aeruginosa can develop mushroom-shaped multicellular structures consisting of distinctsubpopulations in the cap and stalk portions. Wehave previously presented evidence that formation ofthe cap portion of the mushroom-shaped structuresin P. aeruginosa biofilms occurs via bacterial migra-tion and depends on type IV pili (Mol Microbiol 50:61–68). In the present study we examine additionalfactors involved in the formation of this multicellularsubstructure. While pilA mutants, lacking type IV pili,are deficient in mushroom cap formation, pilH andchpA mutants, which are inactivated in the typeIV pili-linked chemosensory system, showed onlyminor defects in cap formation. On the contrary,fliM mutants, which are non-flagellated, and cheYmutants, which are inactivated in the flagellum-linkedchemotaxis system, were largely deficient in capformation. Experiments involving DNase treatmentof developing biofilms provided evidence that extra-cellular DNA plays a role in cap formation. Moreover,mutants that are deficient in quorum sensing-
controlled DNA release formed microcolonies uponwhich wild-type bacteria could not form caps. Theseresults constitute evidence that type IV pili, flagellum-mediated motility and quorum sensing-controlledDNA release are involved in the formation of maturemulticellular structures in P. aeruginosa biofilms.
Introduction
During the last 10 years there has been a rapidly increas-ing recognition of microbial biofilms as a highly significanttopic in microbiology with relevance for a variety of areasin our society including the environment, industry andhuman health. It is generally believed that these struc-tured communities constitute the dominant mode ofmicrobial life (Davey and O’Toole, 2000; Hall-Stoodleyet al., 2004). Analysis of global gene expression bymeans of transcriptomics and proteomics has failed toprovide evidence of a specific developmental biofilmprogramme (Hentzer et al., 2004). On the contrary, high-resolution experimental approaches provide increasingevidence that biofilms are complex and dynamic com-munities in which substantial phenotypic diversificationallows microorganisms to adapt to different environments(Watnick and Kolter, 2000; Klausen et al., 2006). Under-standing the adaptive pathways to biofilm formation willeventually make it possible to manipulate the develop-ment of biofilms in nature and disease.
The use of flow chamber in vitro set-ups and confocallaser scanning microscopy (CLSM) has provided knowl-edge about mechanisms involved in biofilm formation.Pseudomonas aeruginosa, a model organism in biofilmresearch, can, dependent on the conditions, form biofilmswith distinct spatial structures. For example, flat biofilmsare formed in flow chambers irrigated with citrate minimalmedium (Klausen et al., 2003a), while heterogeneousbiofilms with mushroom-shaped multicellular structuresare formed in flow chambers irrigated with glucoseminimal medium (Klausen et al., 2003b). The formationof the mushroom-shaped structures in glucose-grownP. aeruginosa biofilms evidently occurs in a sequentialprocess involving a non-motile bacterial subpopulationwhich forms the initial microcolonies by growth in certain
Received 26 January, 2008; accepted 9 April, 2008. *Forcorrespondence. E-mail [email protected]; Tel. (+45) 45 25 27 93;Fax (+45) 45 88 73 28.
Environmental Microbiology (2008) 10(9), 2331–2343 doi:10.1111/j.1462-2920.2008.01658.x
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd

foci of the biofilm, and a migrating bacterial subpopulationwhich initially forms a monolayer on the substratum, andsubsequently forms the mushroom caps by aggregatingon top of the initial microcolonies (which then becomemushroom stalks) (Klausen et al., 2003b; Haagensenet al., 2007). In glucose-grown biofilms containing amixture of Cfp-tagged and Yfp-tagged P. aeruginosa wild-type bacteria, mushroom-shaped structures were formedthat had single-colour stalks and two-colour caps, indicat-ing that the stalks were formed via growth of non-motilebacteria, and that formation of the caps involved aggre-gating motile bacteria (Klausen et al., 2003b). In glucose-grown biofilms containing a mixture of P. aeruginosa wildtype and P. aeruginosa pilA mutant (deficient in biogen-esis of type IV pili), the pilA mutants could only form stalkswhereas the wild-type bacteria formed all the caps, sug-gesting that type IV pili are necessary for cap formation(Klausen et al., 2003b). Because cap formation wasshown to occur via bacterial migration and require type IVpili, the process was suggested to involve type IV pili-mediated migration (Klausen et al., 2003b).
The extracellular matrix that holds the cells together inP. aeruginosa biofilms mainly consists of polysaccharide,protein and DNA (for a recent review see Pamp et al.,2007). Evidence has been presented that a basal level ofextracellular DNA in P. aeruginosa populations is gener-ated via a pathway which is not linked to quorum sensing,while a larger amount of extracellular DNA appears to begenerated via a pathway that depends on quorum sensingand results in lysis of a small subpopulation of the cells(Allesen-Holm et al., 2006). The P. aeruginosa quorum-sensing mutants, lasIrhlI and pqsA, formed biofilms thathad low extracellular DNA levels and increased suscepti-bility to treatment with SDS, indicating that the extra-cellular DNA in wild-type biofilms has a stabilizing effect(Allesen-Holm et al., 2006). In addition, mature wild-typebiofilms that had been treated with DNase for a short timewere more susceptible to SDS treatment than wild-typebiofilms that were not DNase treated (Allesen-Holm et al.,2006). The extracellular DNA in P. aeruginosa biofilmsappears to be organized in distinct patterns (Allesen-Holmet al., 2006). In young glucose-grown P. aeruginosa bio-films the extracellular DNA was shown to be presentin high concentrations specifically in the outer layer ofthe microcolonies and upon the microcolonies (whichsubsequently became stalks). In mature glucose-grownP. aeruginosa biofilms the extracellular DNA was shown tobe present in high concentrations specifically in the outerlayer of the stalks and between the stalk and cap portionof the mushroom-shaped structures (Allesen-Holm et al.,2006). In agreement with the spatial distribution of theextracellular DNA and a role of quorum sensing in DNArelease, experiments involving a P. aeruginosa pqsA-gfpreporter strain provided evidence that the pqs genes were
expressed specifically in the outer layer of the stalks thatsubsequently became capped by the migrating subpopu-lation (Yang et al., 2007).
Pseudomonas aeruginosa can swim in aqueous envi-ronments, propelled by its polar flagellum. In addition, twodistinct types of surface-associated motility have beendefined for P. aeruginosa grown on agar plates: twitchingmotility requires functional type IV pili (Henrichsen, 1972;Semmler et al., 1999; Mattick, 2002), whereas swarmingmotility requires functional flagella, biosurfactant produc-tion and under some conditions type IV pili (Köhler et al.,2000; Rashid and Kornberg, 2000; Déziel et al., 2003).Comparative sequence analysis has suggested thatP. aeruginosa encodes four chemotaxis-like signal trans-duction systems (Stover et al., 2000). The Pil-Chp system(PA0408-PA0417) is evidently involved in regulatingtwitching motility (Darzins, 1994; Whitchurch et al., 2004).The Che (PA1464-PA1456 and PA3348-PA3349) andChe2 (PA0173-PA0179) systems, both homologous to theEscherichia coli Che chemotaxis system (Parales et al.,2004), have been implicated in flagella-mediated chemo-taxis (Kato et al., 1999; Ferrandez et al., 2002; Honget al., 2004; 2005). The Wsp system (PA3702-3708)appears to control the expression of Cup fimbria(D’Argenio et al., 2002), and the PEL and PSL polysac-charides (Hickman et al., 2005), which are all compoundsthat have been implicated in P. aeruginosa biofilm forma-tion (Vallet et al., 2001; Friedman and Kolter, 2004;Jackson et al., 2004; Matsukawa and Greenberg, 2004).
The cap-forming and the stalk-forming subpopulation ofthe mushroom-shaped structures in P. aeruginosa bio-films in many cases display differential tolerance to anti-microbial compounds. For example evidence has beenprovided that the antibiotics tobramycin, ciprofloxacin andtetracycline preferentially kill bacteria located in the capportion of the mushroom-shaped structures, whereas theantibiotic colistin, the detergent SDS and the chelatorEDTA preferentially kill bacteria in the stalk portion ofthe mushroom-shaped structures (Bjarnsholt et al., 2005;Banin et al., 2006; Haagensen et al., 2007; Pamp et al.,2008). Pseudomonas aeruginosa biofilms that did notdevelop normally, either because they were formed bymutants or as a result of exposure to compounds thatinterfere with biofilm development, did not show the char-acteristic tolerance patterns after treatment with variousantimicrobials including tobramycin, ciprofloxacin, colistinand EDTA (Bjarnsholt et al., 2005; Haagensen et al.,2007; Yang et al., 2007). Because structure developmentin biofilms evidently plays a role in their antimicrobialtolerance properties, enhanced knowledge about struc-tural biofilm development may lead to novel strategies tocontrol biofilm formation.
The present study is focused on elucidating factorswhich are involved in the formation of the mushroom-
2332 K. B. Barken et al.
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343
shaped structures in glucose-grown P. aeruginosabiofilms. We provide evidence that type IV piliation,flagellum-mediated motility and quorum sensing-controlled DNA release are involved in the formation ofthe cap portion of the mushroom-shaped structures inP. aeruginosa biofilms.
Results
Involvement of type IV pili and the Pil/Chpchemosensory system in the structural development ofP. aeruginosa biofilms
Because local consumption may create nutrient gradientsin biofilms (DeBeer et al., 1994; Picioreanu et al., 1998),we speculated that the motile bacteria in glucose-grownP. aeruginosa biofilms might accumulate on top of theinitial microcolonies through the use of a type IV pili-coupled chemosensory system. In order to test thishypothesis we constructed an in-frame P. aeruginosapilH deletion mutant. The pilH gene encodes a predictedresponse regulator, which has been shown to be part ofthe type IV pili-coupled Pil/Chp chemosensory system inP. aeruginosa (Darzins, 1994; Whitchurch et al., 2004).The PAO1 pilH mutant constructed in the present studyshowed reduced twitching zones in an agar stab plateassay, and an increased level of surface piliation(Table 1). When grown as a biofilm in flow chambers thepilH mutant was able to form irregular mushroom-shapedstructures that were larger and less dense than themushroom-shaped structures formed by the wild type(Fig. 1A–D). Because cap formation can be difficult toevaluate in mono-strain biofilms, we investigated theability of the pilH mutant to form caps on top of stalksformed by a PAO1 pilA mutant in mixed-strain biofilms.When the pilH mutant was cultivated in mixed-strain bio-films together with the pilA mutant, the pilH mutant formedcaps on top of the pilA stalks that were irregular and largerand less dense than the caps formed by the wild type(Fig. 1E–H). Complementation of the pilH mutant restored
normal levels of surface pilin production and twitchingmotility, and the complemented pilH mutant formedmushroom-shaped structures in mono-strain biofilms, andcaps on top of pilA stalks in mixed-strain biofilms, similarto the wild type (data not shown).
To further examine roles of the Pil/Chp chemosensorysystem in P. aeruginosa biofilm development we alsostudied a PAO1 chpA in-frame deletion mutant. The chpAgene encodes a composite response regulator, and in astudy focusing mainly on the P. aeruginosa PAK strainevidence was provided that chpA mutants are deficient insignal sensing through the Pil/Chp chemosensory system(Whitchurch et al., 2004). While a PAK chpA in-framedeletion mutant was partially piliated and capable ofresidual twitching motility in an agar stab assay, the PAO1chpA in-frame deletion mutant showed a lower level ofsurface piliation and did not display twitching motility inthe agar stab assay (Table 1). Nevertheless, the PAO1chpA mutant was able to form irregular mushroom-shaped structures of normal size in mono-strain biofilms,and irregular caps of normal size in chpA/pilA mixed-strainbiofilms (Fig. 2).
We performed detailed studies of surface-associatedmotility of the wild type, pilH and chpA mutants in flowchambers using CLSM time-lapse microscopy. By thismethod it is possible to assess motility of cells close to thesubstratum in the early phase of biofilm formation, but dueto technical limitations of CLSM microscopy it is not pos-sible to assess motility of cells far from the substratum inthe later stages of biofilm development. In wild-type andpilH early-phase biofilms both motile and non-motile sub-populations were observed (data not shown). However, inchpA early-phase biofilms only non-motile cells wereobserved (data not shown). The CLSM time-lapse micros-copy therefore suggested that the chpA mutant is defi-cient in type IV pili-driven motility in the early phase ofbiofilm formation, but does not provide information regard-ing migration mechanisms used (far from the substratum)in cap formation in the later phase of biofilm formation.
Table 1. Piliation, twitching, swimming, swarming, chemotaxis and cap formation phenotypes.
Strain genotype Piliationa Twitchingb Swimmingc Swarmingd Chemotaxise Cap formationf
Wild type +++ +++ +++ +++ +++ +++pilA - - +++ +++ +++ -pilH ++++ + +++ +++ +++ +++chpA + - +++ +++ +++ ++fliM +++ +++ - - - -cheY +++ +++ +++ + - -pilAlasRrhlR - - +++ - +++ -
a. Cell-associated pilin was evaluated from immunoblotting and phage assays as described previously (Whitchurch et al., 2004).b. Twitching was evaluated from the diameter of the circular zone of motility formed at the bottom of 1.0% agar plates after 24 h incubation.c. Swimming was evaluated through phase-contrast microscopy of suspensions of bacteria from overnight cultures.d. Swarming was evaluated from the amount of coverage on top of 0.5% agar plates after 48 h incubation.e. Chemotaxis was evaluated from the diameter of the circular zone of motility formed in 0.3% agar plates after 24 h incubation.f. Cap formation was evaluated by CLSM analysis of flow chamber biofilms as described in the main text.
Factors involved in the formation of bacterial multicellular structures 2333
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

Fig. 1. Confocal laser scanning micrographsof 2-day-old (A, C, E and G) and 4-day-old(B, D, F and H) biofilms formed byP. aeruginosa pilH Gfp (A and B),P. aeruginosa wild-type Gfp (C and D), amixture of P. aeruginosa pilH Yfp andP. aeruginosa pilA Cfp (E and F), and amixture of P. aeruginosa wild-type Yfp andP. aeruginosa pilA Cfp (G and H). The centralimages show top-down views and the flankingimages show vertical optical sections. Thebars represent 30 mM.
2334 K. B. Barken et al.
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

From the experiments with the pilH and chpA mutantswe conclude that the Chp chemosensory system plays aminor role in the formation of the mushroom-shaped mul-ticellular structures in P. aeruginosa biofilms. The experi-ments with the chpA mutant appears to disprove ourprevious suggestion that cap formation in P. aeruginosabiofilms occurs via type IV pili-mediated motility (Klausenet al., 2003b). The finding that the non-piliated pilA mutantis cap formation deficient, while the (low level) piliatedchpA mutant is cap formation proficient, suggests that capformation in P. aeruginosa biofilms depends on the pres-ence of type IV pili on the cap-forming bacteria.
Involvement of flagella and the Che chemotaxis systemin the structural development of P. aeruginosa biofilms
The fact that a mixture of Cfp-tagged and Yfp-taggedP. aeruginosa wild-type bacteria forms mushroom-shapedbiofilm structures with single-colour stalks and two-colourcaps (Klausen et al., 2003b) clearly indicates a role ofcellular migration in mushroom cap formation in P. aerugi-nosa biofilms. Because the experiments described abovesuggested that a P. aeruginosa mutant deficient in type IVpili-driven motility can form (irregularly shaped) mushroomcaps, we investigated a role of flagellum-driven motilityin cap formation. The non-flagellated P. aeruginosa fliM
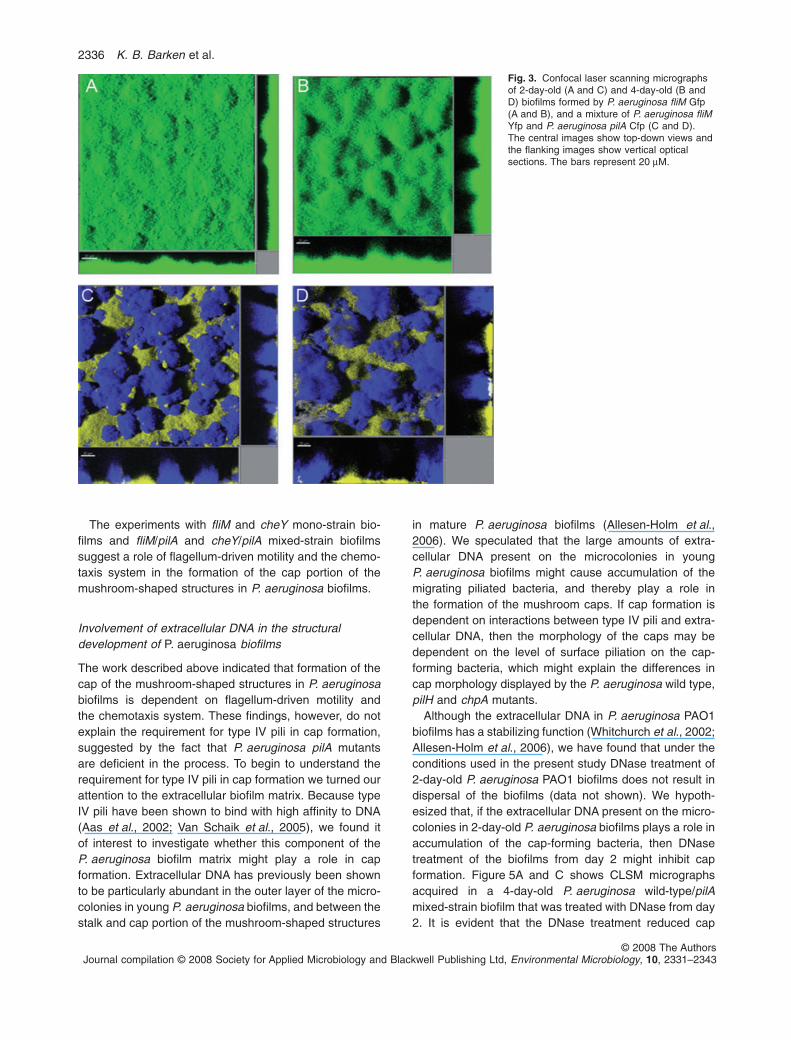
mutant could not form normal mushroom-shaped struc-tures in mono-strain biofilms (Fig. 3A and B), and did notform normal caps on top of pilA microcolonies in mixedfliM/pilA biofilms (Fig. 3C and D). Analysis of older biofilmsshowed that fliM mutants are not simply delayed in biofilmstructure formation (data not shown). The wild type andwild-type/pilA control biofilms developed as shown inFig. 1C, D, G and H (data not shown).
In light of the results described above we found it ofinterest to investigate if the flagellum-linked chemotaxissystem, Che, might play a role in the formation of mush-room caps in P. aeruginosa biofilms. The cheY geneencodes a response regulator of the Che chemotaxissystem in P. aeruginosa (Kato et al., 1999). In order tostudy a role of the chemotaxis system in P. aeruginosabiofilm development we constructed a P. aeruginosa cheYmutant, which proved deficient in chemotaxis but capableof swimming (Table 1). The P. aeruginosa cheY mutantwas unable to form normal mushroom-shaped structuresin mono-strain biofilms (Fig. 4A and B), and unable toform normal caps on top of pilA stalks in mixed cheY/pilAbiofilms (Fig. 4C and D). A complemented cheY mutantformed mushroom-shaped structures in mono-strainbiofilm, and caps on top of pilA stalks in mixed-strainbiofilms, similar to the wild type shown in Fig. 1C, D, Gand H (data not shown).
Fig. 2. Confocal laser scanning micrographsof 2-day-old (A and C) and 4-day-old (B andD) biofilms formed by P. aeruginosa chpA Gfp(A and B), and a mixture of P. aeruginosachpA Yfp and P. aeruginosa pilA Cfp (C andD). The central images show top-down viewsand the flanking images show vertical opticalsections. The bars represent 40 mM.
Factors involved in the formation of bacterial multicellular structures 2335
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343
The experiments with fliM and cheY mono-strain bio-films and fliM/pilA and cheY/pilA mixed-strain biofilmssuggest a role of flagellum-driven motility and the chemo-taxis system in the formation of the cap portion of themushroom-shaped structures in P. aeruginosa biofilms.
Involvement of extracellular DNA in the structuraldevelopment of P. aeruginosa biofilms
The work described above indicated that formation of thecap of the mushroom-shaped structures in P. aeruginosabiofilms is dependent on flagellum-driven motility andthe chemotaxis system. These findings, however, do notexplain the requirement for type IV pili in cap formation,suggested by the fact that P. aeruginosa pilA mutantsare deficient in the process. To begin to understand therequirement for type IV pili in cap formation we turned ourattention to the extracellular biofilm matrix. Because typeIV pili have been shown to bind with high affinity to DNA(Aas et al., 2002; Van Schaik et al., 2005), we found itof interest to investigate whether this component of theP. aeruginosa biofilm matrix might play a role in capformation. Extracellular DNA has previously been shownto be particularly abundant in the outer layer of the micro-colonies in young P. aeruginosa biofilms, and between thestalk and cap portion of the mushroom-shaped structures
in mature P. aeruginosa biofilms (Allesen-Holm et al.,2006). We speculated that the large amounts of extra-cellular DNA present on the microcolonies in youngP. aeruginosa biofilms might cause accumulation of themigrating piliated bacteria, and thereby play a role inthe formation of the mushroom caps. If cap formation isdependent on interactions between type IV pili and extra-cellular DNA, then the morphology of the caps may bedependent on the level of surface piliation on the cap-forming bacteria, which might explain the differences incap morphology displayed by the P. aeruginosa wild type,pilH and chpA mutants.
Although the extracellular DNA in P. aeruginosa PAO1biofilms has a stabilizing function (Whitchurch et al., 2002;Allesen-Holm et al., 2006), we have found that under theconditions used in the present study DNase treatment of2-day-old P. aeruginosa PAO1 biofilms does not result indispersal of the biofilms (data not shown). We hypoth-esized that, if the extracellular DNA present on the micro-colonies in 2-day-old P. aeruginosa biofilms plays a role inaccumulation of the cap-forming bacteria, then DNasetreatment of the biofilms from day 2 might inhibit capformation. Figure 5A and C shows CLSM micrographsacquired in a 4-day-old P. aeruginosa wild-type/pilAmixed-strain biofilm that was treated with DNase from day2. It is evident that the DNase treatment reduced cap
Fig. 3. Confocal laser scanning micrographsof 2-day-old (A and C) and 4-day-old (B andD) biofilms formed by P. aeruginosa fliM Gfp(A and B), and a mixture of P. aeruginosa fliMYfp and P. aeruginosa pilA Cfp (C and D).The central images show top-down views andthe flanking images show vertical opticalsections. The bars represent 20 mM.
2336 K. B. Barken et al.
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

formation compared with the P. aeruginosa wild-type/pilAmixed-strain biofilm that was not subjected to DNasetreatment (Fig. 5B and D). In addition the DNase treat-ment resulted in an unusual biofilm structure wherethe wild-type bacteria to a large extent formed bridgesbetween the pilA microcolonies (Fig. 5). Staining of theextracellular DNA in the biofilm, using our previously
described methods (Allesen-Holm et al., 2006), showedthat residual extracellular DNA was present after theDNase treatment (data not shown), which may explainwhy cap formation was reduced but not abolished.
We further studied the role of extracellular DNA inthe development of the mushroom-shaped P. aeruginosabiofilm structures via a genetic approach. We hypoth-
Fig. 4. Confocal laser scanning micrographsof 2-day-old (A and C) and 4-day-old (B andD) biofilms formed by P. aeruginosa cheY Gfp(A and B), and a mixture of P. aeruginosacheY Yfp and P. aeruginosa pilA Cfp (C andD). The central images show top-down viewsand the flanking images show vertical opticalsections. The bars represent 20 mM.
Fig. 5. Confocal laser scanning micrographsof 4-day-old biofilms formed by mixtures ofP. aeruginosa pilA Cfp and P. aeruginosawild-type Yfp with (A and C) or without (B andD) DNase treatment after 2 days of biofilmgrowth. The bars represent 20 mM.
Factors involved in the formation of bacterial multicellular structures 2337
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

esized that if a pilA mutant carried additional mutations sothat it was unable to release extracellular DNA, it wouldform microcolonies that could not be capped by the wildtype in mixed biofilms. We have previously presentedevidence that a P. aeruginosa lasIrhlI quorum-sensingmutant releases considerably less DNA in biofilms thanthe wild type (Allesen-Holm et al., 2006). It was thereforeof interest to investigate whether a non-piliated quorum-sensing mutant would be capped by the wild type inmixed biofilms. Accordingly we constructed the non-piliated P. aeruginosa pilAlasRrhlR quorum-sensingmutant, which is deficient both in production of, andresponse to, homoserine lactone signal molecules.Assessment of extracellular DNA using our previouslydescribed assays (Allesen-Holm et al., 2006) confirmedthat the P. aeruginosa pilAlasRrhlR mutant is deficient inDNA release when it is grown in flow chambers (data notshown). In wild-type/pilAlasRrhlR mixed biofilms the wildtype was unable to form normal caps on top of the pilA-lasRrhlR microcolonies (Fig. 6). The mushroom-shapedstructures that were observed contained wild type in boththe stalk and the cap (Fig. 6B). The wild-type/pilA controlbiofilm developed as shown in Figs 1H and 5B (data notshown).
In order to obtain an objective measure of the role ofquorum sensing and DNA release in the developmentof mushroom-shaped structures, we acquired a largenumber of CLSM images at random positions in 4-day-oldbiofilms of wild type/pilAlasRrhlR and wild type/pilA, andassessed whether the pilAlasRrhlR and pilA microcolo-nies were colonized by the wild-type bacteria. In thewild-type/pilAlasRrhlR biofilm 6 out of 200 pilAlasRrhlRmicrocolonies were colonized by the wild type, whereas inthe wild-type/pilA biofilm 198 out of 200 pilA microcolonieswere capped by the wild type.
Together these experiments suggest that quorumsensing and DNA release in the initial microcoloniesin P. aeruginosa biofilms plays a role in the subsequentdevelopment of mushroom-shaped structures.
Discussion
In the present study, we present new findings thatenable us to further suggest steps involved in the for-mation of mushroom-shaped multicellular structures inP. aeruginosa biofilms. Our previous studies constituteevidence that the stalk portion of these multicellular struc-tures is formed by non-motile bacteria whereas the capportion is formed by migrating bacteria via a process thatdepends on type IV pili (Klausen et al., 2003a). In addi-tion, we have previously provided evidence that bacteriain the outer layer of the stalks release DNA (Allesen-Holmet al., 2006; Yang et al., 2007). In the current work wepresent evidence that cap formation depends on: (i) typeIV pili, but not type IV pili-mediated motility, (ii) flagellum-mediated motility and the chemotaxis system, and (iii)extracellular DNA released by the stalk subpopulation.
Because the P. aeruginosa chpA mutant investigated inthe present study was deficient in twitching motility in plateassays, and yet capable of forming (irregularly shaped)caps, type IV pili-mediated motility appears not to play arole in mushroom cap formation under the conditions usedin the present study. However the fact that the non-piliatedP. aeruginosa pilA mutant is cap formation deficient, andthe finding that the P. aeruginosa wild-type, chpA and pilHstrains, which differ in the level of surface piliation, formedcaps with different morphologies, suggest that type IV piliplay an important role in cap formation. We have previ-ously proposed that type IV pili are necessary for capformation because of a role in cellular migration (Klausenet al., 2003a). However the present study provides evi-dence that type IV pili instead function as niche-specificadhesins/matrix components in P. aeruginosa biofilms. Inagreement with our suggestion that type IV piliation of thecells is required for mushroom cap formation, whereastype IV pili-mediated motility is not necessary, Chiang andBurrows (2003) reported that a P. aeruginosa pilT mutant,which is hyper-piliated and twitching deficient, formed bio-films with large mushroom-shaped structures.
Fig. 6. Confocal laser scanning micrographsof a 2-day-old (A) and 4-day-old (B) biofilmformed by a mixture of P. aeruginosapilAlasRrhlR Cfp and P. aeruginosa wild-typeYfp. The central images show top-down viewsand the flanking images show vertical opticalsections. The bars represent 20 mM.
2338 K. B. Barken et al.
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

The finding that the P. aeruginosa fliM and cheYmutants did not form normal mushroom-shaped struc-tures in mono-strain biofilms, and did not form normalcaps on top of P. aeruginosa pilA microcolonies in mixedfliM/pilA and cheY/pilA biofilms, suggests that flagellum-driven motility plays the major role in cap formation underthe conditions used in the present study. Previous studiesshowing that chemotaxis mutants of E. coli and Salmo-nella typhimurium displayed conditional defects in swarm-ing motility (Burkart et al., 1998; Mariconda et al., 2006)indicate that the P. aeruginosa cheY and fliM mutantswere affected in cap formation in the present study dueto abolished flagellum-driven surface-associated motility(similar to swarming on agar surfaces). Besides flagellaactivity, swarming motility requires the presence of biosur-factants (Köhler et al., 2000; Rashid and Kornberg, 2000;Déziel et al., 2003), and we have previously shown that aP. aeruginosa rhlA mutant, which is deficient in biosurfac-tant production, displays reduced cap formation in mixedrhlA/pilA biofilms (Pamp and Tolker-Nielsen, 2007).
Although we favour the hypothesis that flagellum-drivensurface-associated motility (similar to swarming) isinvolved in cap formation in P. aeruginosa biofilms, theavailable evidence does not exclude that swimming motil-ity may be involved in the process. Cap formation could tosome extent involve bacteria that detach swim andreattach. Because rhamnolipids were shown to mediatedetachment of P. aeruginosa from biofilms (Boles et al.,2005), the finding that a P. aeruginosa rhlA mutant dis-plays reduced cap formation (Pamp and Tolker-Nielsen,2007) is not necessarily due to a defect in surface-associated motility. In addition, our finding thatP. aeruginosa cheY mutants are deficient in normal capformation could be consistent with a role of swimmingmotility in cap formation. As suggested in a previous study(Tolker-Nielsen et al., 2000), a substrate gradient maydirect the bacteria away from the lower part of the biofilm,whereas gradients of different metabolites leaking fromthe larger microcolonies could direct the swimming bac-teria towards these.
Our data, together with previously published studies,suggest that stalk formation may initiate in certain foci ofa P. aeruginosa biofilm as a consequence of downregu-lation of motility in a subpopulation of the cells. Lactoferrinwas shown to inhibit the formation of microcolonies inP. aeruginosa biofilms by preventing downregulation oftwitching motility (Singh et al., 2002). In addition, evi-dence has been presented that microcolony formation inP. aeruginosa biofilms was inhibited if the bacteria weregrown on a carbon source that stimulated either type IVpili-driven motility (Klausen et al., 2003b) or flagellum-driven motility (Shrout et al., 2006). However, it is alsopossible that microcolony formation initiates because thecells in certain foci of the biofilms produce matrix compo-
nents and adhere strongly to each other so that motilitybecomes arrested.
The involvement of quorum sensing in P. aeruginosabiofilm development has been studied by monitoringbiofilm architectures, and appears to play a role onlyunder some conditions (Davies et al., 1998; Stoodleyet al., 1999; Heydorn et al., 2002; Purevdorj et al., 2002).In the present investigation the use of a model systememploying Cfp-tagged stalk formers and Yfp-tagged capformers provided evidence that quorum sensing in thestalk formers is necessary for the cap to be formed. Theavailable evidence suggests that the observed lack of capformation is due to the stalk formers deficiency in DNArelease, but it cannot be excluded that other quorumsensing-controlled factors in the stalk subpopulation mayaffect cap formation. Quorum sensing controls the pro-duction of biosurfactants in P. aeruginosa (Ochsner andReiser, 1995), and these amphipathic molecules appearsto have multiple roles in P. aeruginosa biofilm develop-ment, one of which is to facilitate bacterial migration andthereby the formation of mushroom caps (Pamp andTolker-Nielsen, 2007). Because the biosurfactants pro-duced via the rhl system (i.e. monorhamnolipid, dirham-nolipid and 3-(3-hydroxyalkanoyloxy)alkanoic acid)have been shown to modulate swarming motility ofP. aeruginosa (Tremblay et al., 2007), it is possible thatthe biosurfactant production defect in the P. aeruginosapilAlasRrhlR mutant may play a role in the lack of capformation on top of this mutant in the wild-type/pilAlasRrhlR mixed biofilm. In agreement with this hypoth-esis, a study employing fluorescent reporter genesindicated that biosurfactant production preferentially takesplace in the stalk portion of the mushroom-shaped struc-tures in mature wild-type P. aeruginosa biofilms (Lequetteand Greenberg, 2005).
The factors involved in coordinating the formation of themushroom-shaped structures in P. aeruginosa biofilmshas been investigated using computer-based modelling(Picioreanu et al., 2007). Because local consumptioncreates nutrient gradients in biofilms (DeBeer et al., 1994;Picioreanu et al., 1998), bacteria which are located inthe cap portion of the mushroom-shaped structures inP. aeruginosa biofilms will have more nutrients availablethan bacteria located in the stalk portion. However,according to the computer simulations, the formationof the mature mushroom-shaped structures cannotbe explained solely on the basis of random surface-associated migration of the motile subpopulation in com-bination with increased cell proliferation of the motile cellsthat settle on top of the stalk microcolonies. Furthermore,the computer simulations predicted that if motility andchemotaxis were the only factors in operation then verticalstrings of cells would be formed in the biofilms. A coordi-nating role of matrix components in the development of
Factors involved in the formation of bacterial multicellular structures 2339
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

the mushroom caps is therefore suggested by computer-based modelling.
In conclusion, the present work suggests that typeIV pili, flagellum-mediated motility and quorum sensing-controlled DNA release are involved in the formation ofthe cap portion of the mushroom-shaped structures inP. aeruginosa biofilms. Based on our present and previousfindings we propose that development of the mushroom-shaped structures in P. aeruginosa biofilms occurs ina sequential process that involves formation of initialmicrocolonies (stalks) by a non-motile subpopulation thatreleases extracellular DNA, and subsequently becomescapped by a motile subpopulation that uses flagellum-driven motility and the chemotaxis system for coordinatedmigration, and type IV pili for binding to the extracellularDNA. Although the proposed interaction between type IVpili and extracellular DNA can explain the requirement fortype IV pili in cap formation it is at present purely specu-lative, and subject to further study in our laboratory.
Experimental procedures
Bacterial strains and growth conditions
Pseudomonas aeruginosa PAO1 (Holloway and Morgan,1986) from John Mattick’s laboratory was used as the wild-type strain in the present study. The pilA and fliM mutantswere constructed by allelic displacement as described byKlausen and colleagues (2003a). The pilAlasRrhlR triplemutant was derived from the pilA mutant via allelic exchangeas described by Beatson and colleagues (2002). The chpAin-frame deletion mutant was constructed by allelic displace-ment as described by Whitchurch and colleagues (2004). ThepilH and cheY mutants were constructed as described below.The strains were fluorescently tagged at an intergenic neutralchromosomal locus with gfp, cfp or yfp in mini-Tn7 constructsas described by Klausen and colleagues (2003a). ModifiedFAB medium (Pamp and Tolker-Nielsen, 2007) was usedsupplemented with 30 mM glucose for batch overnight cul-tures, and with 0.3 mM glucose for biofilm cultivation. Biofilmsand batch cultures were grown at 30°C. Antibiotics were usedat the following concentrations: for P. aeruginosa, carbenicil-lin at 200 mg ml-1 and gentamicin at 30 mg ml-1; for E. coli,ampicillin at 100 mg ml-1, gentamicin at 15 mg ml-1 and kana-mycin at 50 mg ml-1.
Construction of the P. aeruginosa pilH mutant
5′ pilH and 3′ pilH fragments were amplified from PAO1genomic DNA using the pilH1/pilH2 and pilH3/pilH4 poly-merase chain reaction (PCR) primer pairs. PCR productswere A-tailed and ligated into pGEM-T to form pJB47 (5′deletion construct) and pJB48 (3′ deletion construct). The 5′deletion construct was excised from pJB47 as a KpnI/HindIIIfragment. The 3′ deletion construct was excised from pJB52as a HindIII/XbaI fragment. Excised 5′ and 3′ deletion con-struct fragments were concatamerized and cloned as a KpnI/XbaI fragment into pOK12 to form pJB62. The unmarked
deletion construct was excised from pJB64 as a SpeI frag-ment and ligated into the allelic exchange vector pJEN34 toform pJB89. pJB89 was transformed into E. coli S17.1 andtransformants were mated to PAO1 as previously described(Whitchurch et al., 2004) to create unmarked pilH mutants.This procedure deleted amino acids 5–81 of the PilH protein.Sequences of the primers used will be supplied uponrequest.
Complementation of pilH
The pilH gene and 1 kb of 5′ and 3′ flanking sequence wasamplified from PAO1 genomic DNA using the pilH1/pilH4primer pair, and cloned as a KpnI/XbaI fragment into pOK12to form pJB98. The complementation construct was excisedfrom pJB98 as a SpeI fragment and ligated into the allelicexchange vector pJB100 (a modified version of pEX100T inwhich the SmaI site has been replaced with a SpeI site) toform pJB101. pJB101 was transformed into E. coli S17.1 andtransformants were mated to the P. aeruginosa pilH mutantas previously described to create a P. aeruginosa pilHderivative complemented for pilH at its endogenous locus.Sequences of the primers used will be supplied upon request.
Construction of the P. aeruginosa cheY mutant
A knockout fragment of cheY containing a gentamicin resis-tance cassette was generated by PCR overlap extensionessentially as described by Choi and Schweizer (2005).Primers CheY-UpF-GW, CheY-UpR-Gm, CheY-DnF-Gm,CheY-DnR-GW were used to amplify chromosomal regions ofcheY, and primer Gm-F and Gm-R were used to amplify agentamicin resistance cassette from plasmid pPS856 (Hoanget al., 1998). The PCR fragments were fused together andamplified with primers GW-attB1 and GW-attB2 incorporatingthe attB1 and attB2 recombination sites at either end of theknockout cassette. Using the Gateway cloning system (Invit-rogen) the resulting knockout fragment was first transferredby the BP reaction into pDONR221 generating entry plasmidpDONR211cheY, and subsequently transferred by the LRreaction into pEX18ApGW generating the knockout plasmidpEX18ApcheY. The knockout plasmid pEX18ApcheY wastransferred into P. aeruginosa PAO1 by triparental matingusing helper strain E. coli HB101/pRK600 with selectionperformed on Pseudomonas Isolation Agar supplementedwith gentamicin. Resolution of single-cross-over events wasachieved by streaking on 5% sucrose plates via the counter-selectable sacB marker on the knockout plasmid. The result-ing double-cross-over in the P. aeruginosa cheY mutant wasconfirmed by PCR. Sequences of the primers used will besupplied upon request.
Complementation of cheY
The cheY gene was PCR amplified from PAO1 genomic DNAusing the cheY-F and cheY-R primer pair, and the PCRproduct was ligated to the shuttle vector pUCP22Not (Herreroet al., 1990) via BamHI and KpnI restriction sites. The con-struct was transformed into the P. aeruginosa cheY mutant byelectroporation. Sequences of the primers used will be sup-plied upon request.
2340 K. B. Barken et al.
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

Cultivation of biofilms
Biofilms were grown in flow chambers with individual chan-nel dimensions of 1 ¥ 4 ¥ 40 mm. The flow system wasassembled and prepared as described previously (Sternbergand Tolker-Nielsen, 2005). The flow chambers were inocu-lated by injecting 350 ml of overnight culture diluted to anOD600 of 0.01 into each flow channel with a small syringe.After inoculation flow channels were left without flow for 1 h,after which medium flow (0.2 mm s-1) was started using aWatson Marlow 205S peristaltic pump. In one experiment thebiofilm medium was supplemented with 100 mg ml-1 DNase I(Sigma) after 48 h of biofilm cultivation.
Microscopy and image acquisition
All microscopic observations and image acquisitions wereperformed using a Zeiss LSM 510 confocal laser scanningmicroscope (Carl Zeiss, Jena, Germany) equipped withdetectors and filter sets for monitoring of Gfp, Cfp and Yfpfluorescence. Images were obtained using a 63¥/1.4 objec-tive or a 40¥/1.3 objective. Simulated three-dimensionalimages and sections were generated using the IMARIS soft-ware package (Bitplane AG, Zürich, Switzerland).
Acknowledgements
Financial support from the Lundbeck Foundation and theDanish Research Agency is gratefully acknowledged.
References
Aas, F.E., Wolfgang, M., Frye, S., Dunham, S., Lovold, C.,and Koomey, M. (2002) Competence for natural transfor-mation in Neisseria gonorrhoeae: components of DNAbinding and uptake linked to type IV pilus expression. MolMicrobiol 46: 749–760.
Allesen-Holm, M., Barken, K.B., Yang, L., Klausen, M., Webb,J.S., Kjelleberg, S., et al. (2006) A characterization of DNArelease in Pseudomonas aeruginosa cultures and biofilms.Mol Microbiol 59: 1114–1128.
Banin, E., Brady, K.M., and Greenberg, E.P. (2006) Chelator-induced dispersal and killing of Pseudomonas aeruginosacells in a biofilm. Appl Environ Microbiol 72: 2064–2069.
Beatson, S.A., Whitchurch, C.B., Sargent, J.L., Levesque,R.C., and Mattick, J.S. (2002) Differential regulation oftwitching motility and elastase production by Vfr inPseudomonas aeruginosa. J Bacteriol 184: 3605–3613.
Bjarnsholt, T., Jensen, P.O., Burmolle, M., Hentzer, M.,Haagensen, J.A., Hougen, H.P., et al. (2005) Pseudomo-nas aeruginosa tolerance to tobramycin, hydrogen perox-ide and polymorphonuclear leukocytes is quorum-sensingdependent. Microbiology 151: 373–383.
Boles, B.R., Thoendel, M., and Singh, P.K. (2005) Rhamno-lipids mediate detachment of Pseudomonas aeruginosafrom biofilms. Mol Microbiol 57: 1210–1223.
Burkart, M., Toguchi, A., and Harshey, R.M. (1998) Thechemotaxis system, but not chemotaxis, is essential forswarming motility in Escherichia coli. Proc Natl Acad SciUSA 95: 2568–2573.
Chiang, P., and Burrows, L.L. (2003) Biofilm formationby hyperpilated mutants of Pseudomonas aeruginosa.J Bacteriol 185: 2374–2378.
Choi, K.H., and Schweizer, H.P. (2005) An improved methodfor rapid generation of unmarked Pseudomonas aerugi-nosa deletion mutants. BMC Microbiol 235: 30.
D’Argenio, D.A., Calfee, M.W., Rainey, P.B., and Pesci, E.C.(2002) Autolysis and autoaggregation in Pseudomonasaeruginosa colony morphology mutants. J Bacteriol 184:6481–6489.
Darzins, A. (1994) Characterization of a Pseudomonasaeruginosa gene cluster involved in pilus biosynthesis andtwitching motility: sequence similarity to the chemotaxisproteins of enterics and the gliding bacterium Myxococcusxanthus. Mol Microbiol 11: 137–153.
Davey, M.E., and O’Toole, G.A. (2000) Microbial biofilms:from ecology to molecular genetics. Microbiol Mol Biol 64:847–867.
Davies, D.G., Parsek, M.R., Pearson, J.P., Iglewski, B.H.,Costerton, J.W., and Greenberg, E.P. (1998) The involve-ment of cell-to-cell signals in the development of a bacterialbiofilm. Science 280: 295–298.
DeBeer, D., Stoodley, P., Roe, F., and Lewandowski, Z.(1994) Effects of biofilm structures on oxygen distributionand mass transport. Biotechnol Bioeng 43: 1131–1138.
Déziel, E., Lepine, F., Milot, S., and Villemur, R. (2003) rhlAis required for the production of a novel biosurfactant pro-moting swarming motility in Pseudomonas aeruginosa:3-(3-hydroxy-alkanoyloxy)alkanoic acids (HAAs), the pre-cursors of rhamnolipids. Microbiology 149: 2005–2013.
Ferrandez, A., Hawkins, A.C., Summerfield, D.T., andHarwood, C.S. (2002) Cluster II che genes fromPseudomonas aeruginosa are required for an optimalchemotactic response. J Bacteriol 184: 4374–4383.
Friedman, L., and Kolter, R. (2004) Two genetic loci producedistinct carbohydrate-rich structural components of thePseudomonas aeruginosa biofilm matrix. J Bacteriol 186:4457–4465.
Haagensen, J.A., Klausen, M., Ernst, R.K., Miller, S.I.,Folkesson, A., Tolker-Nielsen, T., and Molin, S. (2007)Differentiation and distribution of colistin- and sodiumdodecyl sulfate-tolerant cells in Pseudomonas aeruginosabiofilms. J Bacteriol 189: 28–37.
Hall-Stoodley, L., Costerton, J.W., and Stoodley, P. (2004)Bacterial biofilms: from the natural environment to infec-tious diseases. Nat Rev Microbiol 2: 95–108.
Henrichsen, J. (1972) Bacterial surface translocation: asurvey and a classification. Bacteriol Rev 36: 478–503.
Hentzer, M., Eberl, L., and Givskov, M. (2004) Quorumsensing in biofilms: gossip in the slime world? In MicrobialBiofilms. Ghannoum, M., and O’Toole, G. (eds). Washing-ton, DC, USA: American Society For Microbiology Press,pp. 118–140.
Herrero, M., de Lorenzo, V., and Timmis, K.N. (1990) Trans-poson vectors containing non-antibiotic resistance selec-tion markers for cloning and stable chromosomal insertionof foreign genes in gram-negative bacteria. J Bacteriol 172:6557–6567.
Heydorn, A., Ersbøll, B., Kato, J., Hentzer, M., Parsek, M.,Tolker-Nielsen, T., et al. (2002) Statistical analysis ofPseudomonas aeruginosa biofilm development: impact of
Factors involved in the formation of bacterial multicellular structures 2341
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

mutations in genes involved in twitching motility, cell-to-cellsignaling, and stationary-phase sigma factor expression.Appl Environ Microbiol 68: 2008–2017.
Hickman, J.W., Tifrea, D.F., and Harwood, C.S. (2005) Achemosensory system that regulates biofilm formationthrough modulation of cyclic diguanylate levels. Proc NatlAcad Sci USA 102: 14422–14427.
Hoang, T.T., Karkhoff-Schweizer, R.R., Kutchma, A.J., andSchweizer, H.P. (1998) A broad-host-range Flp-FRTrecombination system for site-specific excision ofchromosomally-located DNA sequences: application forisolation of unmarked Pseudomonas aeruginosa mutants.Gene 212: 77–86.
Holloway, B.W., and Morgan, A.F. (1986) Genome organiza-tion in Pseudomonas. Annu Rev Microbiol 40: 79–105.
Hong, C.S., Shitashiro, M., Kuroda, A., Ikeda, T., Takiguchi,N., Ohtake, H., and Kato, J. (2004) Chemotaxis proteinsand transducers for aerotaxis in Pseudomonas aerugi-nosa. FEMS Microbiol Lett 231: 247–252.
Hong, C.S., Kuroda, A., Takiguchi, N., Ohtake, H., and Kato,J. (2005) Expression of Pseudomonas aeruginosa aer-2,one of two aerotaxis transducer genes, is controlled byRpoS. J Bacteriol 187: 1533–1535.
Jackson, K.D., Starkey, M., Kremer, S., Parsek, M.R., andWozniak, D.J. (2004) Identification of psl, a locus encod-ing a potential exopolysaccharide that is essential forPseudomonas aeruginosa PAO1 biofilm formation.J Bacteriol 186: 4466–4475.
Kato, J., Nakamura, T., Kuroda, A., and Ohtake, H. (1999)Cloning and characterization of chemotaxis genes inPseudomonas aeruginosa. Biosci Biotechnol Biochem 63:155–161.
Klausen, M., Heydorn, A., Ragas, P., Lambertsen, L., Aaes-Jorgensen, A., Molin, S., and Tolker-Nielsen, T. (2003a)Biofilm formation by Pseudomonas aeruginosa wild type,flagella and type IV pili mutants. Mol Microbiol 48: 1511–1524.
Klausen, M., Aaes-Jorgensen, A., Molin, S., and Tolker-Nielsen, T. (2003b) Involvement of bacterial migration inthe development of complex multicellular structures inPseudomonas aeruginosa biofilms. Mol Microbiol 50:61–68.
Klausen, M., Gjermansen, M., Kreft, J.-U., and Tolker-Nielsen, T. (2006) Dynamics of development and dispersalin sessile microbial communities: examples from Pseu-domonas aeruginosa and Pseudomonas putida biofilms.FEMS Microbiol Lett 261: 1–11.
Köhler, T., Curty, L.K., Barja, F., van Delden, C., andPechere, J.C. (2000) Swarming of Pseudomonas aerugi-nosa is dependent on cell-to-cell signaling and requiresflagella and pili. J Bacteriol 182: 5990–5996.
Lequette, Y., and Greenberg, E.P. (2005) Timing and local-ization of rhamnolipid synthesis gene expression inPseudomonas aeruginosa biofilms. J Bacteriol 187: 37–44.
Mariconda, S., Wang, Q., and Harshey, R.M. (2006) Amechanical role for the chemotaxis system in swarmingmotility. Mol Microbiol 60: 1590–1602.
Matsukawa, M., and Greenberg, E.P. (2004) Putativeexopolysaccharide synthesis genes influence Pseudomo-nas aeruginosa biofilm development. J Bacteriol 186:4449–4456.
Mattick, J.S. (2002) Type IV pili and twitching motility. AnnuRev Microbiol 56: 289–314.
Ochsner, U.A., and Reiser, J. (1995) Autoinducer-mediatedregulation of rhamnolipid biosurfactant synthesis inPseudomonas aeruginosa. Proc Natl Acad Sci USA 92:6424–6428.
Pamp, S.J., and Tolker-Nielsen, T. (2007) Multiple rolesof biosurfactants in structural biofilm developmentby Pseudomonas aeruginosa. J Bacteriol 189: 2531–2539.
Pamp, S.J., Gjermansen, M., and Tolker-Nielsen, T. (2007)The biofilm matrix – a sticky framework. In Bacterial BiofilmFormation and Adaptation. Kjelleberg S., and Givskov, M.(eds). Wymondham, Norfolk, UK: Horizon BioScience, pp.37–69.
Pamp, S.J., Gjermansen, M., Johansen, H.K., and Tolker-Nielsen, T. (2008) Tolerance to the antimicrobial peptidecolistin in Pseudomonas aeruginosa biofilms is linked tometabolically active cells, and depends on the pmr andmexAB-oprM genes. Mol Microbiol 68: 223–240.
Parales, E.R., Ferrandez, A., and Harwood, C.S. (2004)Chemotaxis in pseudomonas. In Pseudomonas, Genom-ics, Life Style and Molecular Architecture, Vol. 1. Ramos,J.-L. (ed.). New York: Kluwer Academic/Plenum Publish-ers, pp. 793–815.
Picioreanu, C., van Loosdrecht, M.C.M., and Heijnen, J.J.(1998) Mathematical modeling of biofilm structure with ahybrid differential-discrete cellular automaton approach.Biotechnol Bioeng 58: 101–116.
Picioreanu, C., Kreft, J.-U., Klausen, M., Haagensen, J.A.,Tolker-Nielsen, T., and Molin, S. (2007) Microbial motilityinvolvement in biofilm structure formation – a 3D modelingstudy. Water Sci Technol 55: 337–343.
Purevdorj, B., Costerton, J.W., and Stoodley, P. (2002) Influ-ence of hydrodynamics and cell signaling on the structureand behavior of Pseudomonas aeruginosa biofilms. ApplEnviron Microbiol 68: 4457–4464.
Rashid, M.H., and Kornberg, A. (2000) Inorganic polyphos-phate is needed for swimming, swarming, and twitchingmotilities of Pseudomonas aeruginosa. Proc Natl Acad SciUSA 25: 4885–4890.
Semmler, A.B., Whitchurch, C.B., and Mattick, J.S. (1999)A re-examination of twitching motility in Pseudomonasaeruginosa. Microbiology 145: 2863–2873.
Shrout, J.D., Chopp, D.L., Just, C.L., Hentzer, M., Givskov,M., and Parsek, M.R. (2006) The impact of quorum sensingand swarming motility on Pseudomonas aeruginosa biofilmformation is nutritionally conditional. Mol Microbiol 62:1264–1277.
Singh, P.K., Parsek, M.R., Greenberg, E.P., and Welsh, M.J.(2002) A component of innate immunity prevents bacterialbiofilm development. Nature 417: 552–555.
Sternberg, C., and Tolker-Nielsen, T. (2005) Growing andanalyzing biofilms in flow cells. In Current Protocols inMicrobiology. Coico, R., Kowalik, T., Quarles, J., Steven-son, B., and Taylor, R. (eds). New York: John Wiley & Sons,pp. 1B.2.1–1B.2.15.
Stoodley, P., Jorgensen, F., Williams, P., and Lappin-Scott,H.M. (1999) The role of hydrodynamics and AHL signallingmolecules as determinants of the structure of Pseudomo-nas aeruginosa biofilms. In Biofilms: The Good, the Bad,
2342 K. B. Barken et al.
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343

and the Ugly. Wimpenny, J., Gilbert, P., Walker, J.,Brading, M., and Baystun, R. (eds). Cardiff, UK: Bioline, pp.223–230.
Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D.,Warrener, P., Hickey, M.J., et al. (2000) Complete genomesequence of Pseudomonas aeruginosa PA01, an opportu-nistic pathogen. Nature 406: 959–964.
Tolker-Nielsen, T., Brinch, U.C., Ragas, P.C., Andersen, J.B.,Jacobsen, C.S., and Molin, S. (2000) Development anddynamics of Pseudomonas sp. biofilms. J Bacteriol 182:6482–6489.
Tremblay, J., Richardson, A.P., Lépine, F., and Déziel, E.(2007) Self-produced extracellular stimuli modulate thePseudomonas aeruginosa swarming motility behaviour.Environ Microbiol 9: 2622–2630.
Vallet, I., Olson, J.W., Lory, S., Lazdunski, A., and Filloux, A.(2001) The chaperone/usher pathways of Pseudomonasaeruginosa: identification of fimbrial gene clusters (cup)and their involvement in biofilm formation. Proc Natl AcadSci USA 98: 6911–6916.
Van Schaik, E.J., Giltner, C.L., Audette, G.F., Keizer, D.W.,Bautista, D.L., Slupsky, C.M., et al. (2005) DNA binding:a novel function of Pseudomonas aeruginosa type IV pili.J Bacteriol 187: 1455–1464.
Watnick, P., and Kolter, R. (2000) Biofilm, city of microbes.J Bacteriol 182: 2675–2679.
Whitchurch, C.B., Tolker-Nielsen, T., Ragas, P.C., andMattick, J.S. (2002) Extracellular DNA required for bacterialbiofilm formation. Science 295: 1487.
Whitchurch, C.B., Leech, A.J., Young, M.D., Kennedy, D.,Sargent, J.L., Bertrand, J.J., et al. (2004) Characterizationof a complex chemosensory signal transduction systemwhich controls twitching motility in Pseudomonas aerug-inosa. Mol Microbiol 52: 873–893.
Yang, L., Barken, K.B., Skindersoe, M.E., Christensen, A.B.,Givskov, M., and Tolker-Nielsen, T. (2007) Effects ofiron on DNA-release and biofilm development byPseudomonas aeruginosa. Microbiology 153: 1318–1328.
Factors involved in the formation of bacterial multicellular structures 2343
© 2008 The AuthorsJournal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 2331–2343
Related Documents











