RESEARCH ARTICLE Role of Sperm avb3 Integrin in Mouse Fertilization Ce ´ line Chalas Boissonnas, 1,2 Debbie Montjean, 3 Corinne Lesaffre, 1,4 Jana Auer, 1,4 Daniel Vaiman, 4 Jean-Philippe Wolf, 1,2 * and Ahmed Ziyyat 1,2 Oocyte integrins have been described as essential for fertilization. But this concept has been challenged by deletion experiments. Recently, we have shown that sperm integrin a6b1 plays a determinant role in mouse gamete interaction. In this study, we demonstrate the presence of avb3 integrin by Western blot and immunofluorescence on the sperm membrane. Oocytes and/or sperm preincubations with anti-av or anti-b3 antibodies were performed before in vitro fertilization on cumulus-intact and zona-free egg assays. We observed inhibitory effects on the fusion process mostly by means of sperm function. An anti- body directed against vitronectin inhibited gametes fusion, whereas the presence of exogenous vitronec- tin increased its efficiency. We suggest that vitronectin (on multimeric forms) can play a first nonspecific link corresponding to loosely bound spermatozoa to oocyte and that this link could be mediated by means of oocyte proteoglycans or integrins, and sperm avb3 integrin. Developmental Dynamics 239:773–783, 2010. V C 2010 Wiley-Liss, Inc. Key words: fertilization; integrin; vitronectin; sperm; mouse Accepted 24 November 2009 INTRODUCTION The initial gamete interaction model that was conceived involved one integrin, a6b1, on the oocyte (Almeida et al., 1995), and its sperm ligand, fer- tilin b, also known as ADAM2 (A Dis- integrin And Metalloprotease2; Chen and Sampson, 1999). In fact, sperm– egg interaction suggests a much more complex molecular process, involving several protein families such as ADAMs, CRISPs, SLLP1, SAMP14, SAMP32; immunoglobulins such as Izumo; integrins; GPI-APs; and tetra- spanins. Among them, two molecules have been shown to be essential for fertilization: Izumo, a sperm protein belonging to the immunoglobulin superfamily (Inoue et al., 2005), and CD9 tetraspanin on the oocyte mem- brane (Kaji et al., 2000; Le Naour et al., 2000; Miyado et al., 2000). Dele- tions of these two genes result in a lack of fusion of spermatozoa with oocytes and a dramatic reduction of the knockout (KO) mice fertility. Moreover, many integrin subunits, that may play a role in gamete inter- action, have been identified on the oocyte membrane: a2, a3, a4, a5, a6, a9, aM, av, b1, b2, b3, and b5 (Camp- bell et al., 1995; Evans et al., 1995; Zuccotti et al., 1998; Burns et al., 2002; Sengoku et al., 2004; Vjugina et al., 2009). However, the essential role of such proteins upon fertilization was called into question in several publications. Indeed, a3 and a6 integ- rin subunit null eggs from KO mice and oocyte-specific b1 integrin condi- tional KO mice were designed and investigated. These animals were fer- tile and their oocytes were normally functional in in vitro fertilization Developmental Dynamics ABBREVIATIONS AR acrosome reacted/reaction AI acrosome intact AT acid Tyrode COC cumulus oocyte complex FI fertilization index FR fertilization rate IVF in vitro fertilization PSA Pisum sativum agglutinin RT room temperature VN vitronectin ZP zona pellucida 1 Universite ´ Paris Descartes, Laboratoire Interaction Game ´tique et Fusion des Membranes, EA 2717, Institut Alfred Jost, Paris, France 2 Service d’Histologie Embryologie Biologie de la Reproduction, Ho ˆpital Cochin, Assistance Publique-Ho ˆpitaux de Paris, France 3 Institut Pasteur, Human Developmental Genetics, Paris, France 4 Inserm U567, Department of Genetics and Development, Cochin Institute, Paris, France *Correspondence to: Jean-Philippe Wolf, Universite ´ Paris Descartes, Laboratoire Interaction Game ´ tique et Fusion des Membranes, EA 2717, Institut Alfred Jost, 24 rue du Fbg Saint Jacques, 75014 Paris. E-mail: [email protected] DOI 10.1002/dvdy.22206 Published online 8 January 2010 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 239:773–783, 2010 V C 2010 Wiley-Liss, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
Role of Sperm avb3 Integrin in MouseFertilizationCeline Chalas Boissonnas,1,2 Debbie Montjean,3 Corinne Lesaffre,1,4 Jana Auer,1,4 Daniel Vaiman,4
Jean-Philippe Wolf,1,2* and Ahmed Ziyyat1,2
Oocyte integrins have been described as essential for fertilization. But this concept has been challengedby deletion experiments. Recently, we have shown that sperm integrin a6b1 plays a determinant role inmouse gamete interaction. In this study, we demonstrate the presence of avb3 integrin by Western blotand immunofluorescence on the sperm membrane. Oocytes and/or sperm preincubations with anti-av oranti-b3 antibodies were performed before in vitro fertilization on cumulus-intact and zona-free eggassays. We observed inhibitory effects on the fusion process mostly by means of sperm function. An anti-body directed against vitronectin inhibited gametes fusion, whereas the presence of exogenous vitronec-tin increased its efficiency. We suggest that vitronectin (on multimeric forms) can play a first nonspecificlink corresponding to loosely bound spermatozoa to oocyte and that this link could be mediated by meansof oocyte proteoglycans or integrins, and sperm avb3 integrin. Developmental Dynamics 239:773–783, 2010.VC 2010 Wiley-Liss, Inc.
Key words: fertilization; integrin; vitronectin; sperm; mouse
Accepted 24 November 2009
INTRODUCTION
The initial gamete interaction model
that was conceived involved one
integrin, a6b1, on the oocyte (Almeida
et al., 1995), and its sperm ligand, fer-
tilin b, also known as ADAM2 (A Dis-
integrin And Metalloprotease2; Chen
and Sampson, 1999). In fact, sperm–
egg interaction suggests a much more
complex molecular process, involving
several protein families such as
ADAMs, CRISPs, SLLP1, SAMP14,
SAMP32; immunoglobulins such as
Izumo; integrins; GPI-APs; and tetra-
spanins. Among them, two molecules
have been shown to be essential forfertilization: Izumo, a sperm proteinbelonging to the immunoglobulinsuperfamily (Inoue et al., 2005), andCD9 tetraspanin on the oocyte mem-brane (Kaji et al., 2000; Le Naouret al., 2000; Miyado et al., 2000). Dele-tions of these two genes result in alack of fusion of spermatozoa withoocytes and a dramatic reduction ofthe knockout (KO) mice fertility.Moreover, many integrin subunits,that may play a role in gamete inter-action, have been identified on the
oocyte membrane: a2, a3, a4, a5, a6,a9, aM, av, b1, b2, b3, and b5 (Camp-bell et al., 1995; Evans et al., 1995;Zuccotti et al., 1998; Burns et al.,2002; Sengoku et al., 2004; Vjuginaet al., 2009). However, the essentialrole of such proteins upon fertilizationwas called into question in severalpublications. Indeed, a3 and a6 integ-rin subunit null eggs from KO miceand oocyte-specific b1 integrin condi-tional KO mice were designed andinvestigated. These animals were fer-tile and their oocytes were normallyfunctional in in vitro fertilization
Dev
elop
men
tal D
ynam
ics
ABBREVIATIONS AR acrosome reacted/reaction AI acrosome intact AT acid Tyrode COC cumulus oocyte complex FI fertilizationindex FR fertilization rate IVF in vitro fertilization PSA Pisum sativum agglutinin RT room temperature VN vitronectin ZP zonapellucida
1Universite Paris Descartes, Laboratoire Interaction Gametique et Fusion des Membranes, EA 2717, Institut Alfred Jost, Paris, France2Service d’Histologie Embryologie Biologie de la Reproduction, Hopital Cochin, Assistance Publique-Hopitaux de Paris, France3Institut Pasteur, Human Developmental Genetics, Paris, France4Inserm U567, Department of Genetics and Development, Cochin Institute, Paris, France*Correspondence to: Jean-Philippe Wolf, Universite Paris Descartes, Laboratoire Interaction Gametique et Fusion desMembranes, EA 2717, Institut Alfred Jost, 24 rue du Fbg Saint Jacques, 75014 Paris. E-mail: [email protected]
DOI 10.1002/dvdy.22206Published online 8 January 2010 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 239:773–783, 2010
VC 2010 Wiley-Liss, Inc.

(Miller et al., 2000; He et al., 2003).b3 integrin subunit KO mice are alsofertile (Hodivala-Dilke et al., 1999).This led to the conclusion that ‘‘noneof the integrins known to be presenton the mouse egg or to be ADAMreceptors are essential for sperm–eggbinding and fusion’’ (He et al., 2003),but their involvement in this phenom-enon was never excluded (Evans,2009). In this context, recent studyusing wild-type (WT) and b1-nulleggs and polymers of peptide from thefertilin b disintegrin domain, sug-gested that oocyte b1 integrin enhan-ces the initial adhesion of sperm tothe egg plasma membrane, subse-quent attachment and fusion beingmediated by additional egg and spermproteins present in the b1 integrincomplex (Baessler et al., 2009).Another recent study, using eitherknockdown by RNA interferencedirected against a9 integrin or anti-b1 integrin antibody revealed areduction of sperm–egg binding andfusion (Vjugina et al., 2009). In addi-tion, during total or conditional KOexperiments, the deleted femaleswere always mated with WT malesand in vitro tests were also performedusing WT sperm. But neither thepresence of these integrins on mousespermatozoa nor their involvementsin gamete interaction were ques-tioned. Yet, the expression of a6, avand b3 integrins on human spermato-zoa had been demonstrated for a longtime (Fusi et al., 1996b). Recently, wehave shown that a6b1 integrin is alsoexpressed by mouse spermatozoa andinvolved in gamete adhesion and fusion(Barraud-Lange et al., 2007b). In addi-tion, we have demonstrated that thecyclic FEE peptide, that mimics thedisintegrin binding domain of fertilinb, ligand of integrin a6b1, potentiatesthe inhibitory effect of RGD peptide,which can bind to RGD sensitive integ-rins. RGD-binding integrin subfamily(including avb1, avb3, and avb5),expressed by the oocyte have beenshown to be implicated in fertilizationbecause the use of RGD peptide duringIVF assays inhibits fertilization (Bron-son and Fusi, 1990). This suggeststhat RGD insensitive and RGD sensi-tive integrins cooperate during fertil-ization (Ziyyat et al., 2005). Bronsonet al. described a similar potentiationof the inhibitory effect of RGD pep-
tide by FEE peptide (Bronson et al.,1999). They used a linear FEE-con-taining peptide and a cyclic RGD-con-taining peptide that exhibited a syn-ergistic inhibition of both adhesionand fusion, suggesting cooperationbetween the two receptor ligands dur-ing fertilization in human. All thesedata suggests that at least two differ-ent oocyte integrins could be involvedin human gamete interaction, onerecognizing fertilin and the other rec-
ognizing RGD-containing sperm-asso-ciated proteins such as vitronectin orfibronectin, and that they may coop-erate (Bronson et al., 1999). Indeed,avb3 integrin is RGD sensitive andis known to be the receptor of vitro-nectin (VN), a component of theextracellular matrix of acrosomereacted (AR) sperm (Fusi et al.,1996a; Bronson et al., 2000). More-over, the colocalization of avb3 witha6b1 integrin on human zona
Fig. 1. Expression and dynamic distribution of avb3 integrin on mouse sperm. A,B: WesternBlot analysis of mouse sperm avb3 integrin and vitronectin expression. A: Alpha V and beta 3integrin subunits and vitronectin expression was investigated by Western blot analysis. Proteinsfrom epididymal sperm were extracted with deoxycholate/1% NP40, subjected to sodium do-decyl sulfate-polyacrylamide gel electrophoresis under reducing conditions and blotted withanti-av and anti-b3 integrin subunit and anti-VN antibodies. Specific bands (approximately 125and 135 kDa and 65 kDa) were detected on sperm extract after rabbit polyclonal antibodiesincubation against av and b3 integrin subunits and vitronectin, respectively. A: These bands cor-respond to av and b3 integrin subunits as confirmed by the use of the F9 Whole Cell Lysate aspositive control (50 mg of proteins). No specific band appeared when a nonspecific rabbit IgGwas used (Isotype lane). B: Band at 65 kDa corresponds to vitronectin. No specific bandappeared when a nonspecific rabbit IgG was used. 10 mg proteins correspond to one millionsperm. C: Dynamic distribution of alpha V beta 3 integrin on mouse sperm assessed by immu-nofluorescence. Distribution of av and b3 integrin subunits was studied by immunofluorescencemicroscopy. Spermatozoa were exposed to Pisum sativum agglutinin conjugated to fluorescein(PSA-FITC; b,e,h) after anti-av integrin subunit antibody (c,f,i) followed by anti-rat biotinylatedsecondary antibody and Alexa Fluor 594 conjugated Streptavidin. The following fluorescent pat-terns were recorded: negative noncapacitated sperm (c), 30% of capacitated acrosome intactspermatozoa (f) and 93% for capacitated acrosome reacted spermatozoa (i) were stained.Sperm nuclei were detected by DAPI (40,6-diamidine-2-phenylidole-dihydrochloride) staining(a,d,g). These images correspond to the most common staining but other distributions havebeen observed. Nonimmune control did not give any signal. Approximately similar pattern stain-ing was observed for b3 integrin except that b3 appeared on 40% of freshly recovered nonca-pacitated sperm. The same treatment was applied to sperm as for av detection but using anti-b3 antibody (a0 to i0). % indicates the frequency of showed distribution. Scale bar ¼ 1 mm.
Dev
elop
men
tal D
ynam
ics
774 BOISSONNAS ET AL.

pellucida (ZP) free oocytes supportsthis hypothesis (Ziyyat et al., 2005).
We postulate that such cooperationmight also take place on sperm,which suggests that sperm wouldexpress not only the a6b1 integrinbut also the avb3 integrin. In addi-tion, after being released during theAR, VN has been described in humanas the Velcro that binds gametes to-gether through its adhesion to avb3integrin (Fusi et al., 1996a; Bronsonet al., 2000). This suggests that avb3integrin is present on the two game-tes, what is true in humans but notyet shown in mice. Thus, we decidedto search whether avb3 is expressedby mouse sperm and involved inmouse gamete interaction or not.Western blot and immunofluores-cence analysis enabled us to demon-strate that avb3 integrin is actuallyexpressed on mouse spermatozoamembrane. Functional tests usingantibodies directed against av and b3integrin subunits demonstrated itsinvolvement in fertilization both onsperm and cumulus oocyte complex(COC) side. We also showed the roleof vitronectin in mouse sperm–egginteraction. All these findings lead usto propose a model according towhich macromolecules of VN, liber-ated during AR (Bronson et al.,2000), could bind to avb3 integrinboth on sperm and oocyte, as a Vel-cro. It would constitute a first non-specific adhesion process leading towhat is called as loosely boundsperm. Afterward, other sperm –oocyte ligands and receptors couldinteract to make a tighter and morespecific adhesion possible.
RESULTS
av and b3 Integrin Subunits
and Vitronectin Are
Expressed on Mouse Sperm
Membrane
Western blot analysis.
Expression of av and b3 integrin sub-units and vitronectin on cauda epidid-ymal sperm was evaluated by immu-noblot analysis. As shown on Figure1A, specific bands at approximately125 and 135 kDa, were detected onsperm (10 mg proteins per 106 sperm)as in the F9 Whole Cell Lysate posi-
tive control (50 mg). It corresponds tothe expected molecular weight of avand b3 integrin subunits under reduc-ing conditions. Under the same condi-tions, VN was detected using the anti-vitronectin antibody as a specificband at 65–75 kDa (Fig. 1B). Controlsusing a nonspecific rabbit and rat pol-yclonal antibodies were negative.These results demonstrated the pres-ence of av, b3 integrin subunits andVN on mouse sperm membrane.
Immunofluorescence analysis.
Immunofluorescence was used tolocalize av and b3 integrin subunitson the head of mouse sperm. In paral-lel, acrosomal status was assessedusing Pisum sativum agglutinin(PSA) staining protocol. The detectionof the av integrin subunit variedaccording to the sperm state of matu-rity. Indeed, freshly recovered nonca-pacitated sperm did not show any dis-tinct fluorescence (Fig. 1C.c). Thestaining detection appeared duringcapacitation, whereas only 30% ofcapacitated acrosome intact (AI) sper-matozoa were stained (Fig. 1C.f), 93%
of capacitated AR spermatozoa group(Fig. 1C.i) showed a specific labeling.Three different staining patternscould be observed: av integrin subunitcould be seen spreading over the acro-some, over the whole head or could belocalized on the equatorial region.Whereas some spermatozoa showed apunctual staining others had a moreuniform covering over the entirehead. The most frequent pattern wasthe overall covering of the acrosomalregion (75% of capacitated acrosome-reacted spermatozoa). Similar distri-bution, staining, and relative propor-tions were found for b3 integrin subu-nit, except that it appeared lessdependent on the sperm maturity sta-tus. Forty percent of freshly recoverednoncapacitated sperm were stained(Fig. 1C.c0). Nonimmune controls(Fig. 1C) and secondary antibodiesgave no signal (data not shown).
avb3 Integrin Is Functional
During Mouse Fertilization
Because the presence of av and b3integrin subunits as well as that ofVN was demonstrated on the mouse
Fig. 2. Effects of alpha V integrin subunit antibody (50 mg/ml) on mouse cumulus-intact andzona free in vitro fertilization (IVF) assays. A: Fertilization rate or mean (6 SEM) percentage offertilized eggs following cumulus-intact IVF assay at 106 spermatozoa per ml for 3 hr. B: Fertil-ization index or mean (6 SEM) of sperm number fused by egg on zona-free IVF assay at 105
spermatozoa per ml for 3 hr. No antibodies (CTRL), in vitro fertilization in the presence of anti-body but without preincubations (IVF), groups with cumulus oocyte complex (COC), oocyte (O),sperm (S), sperm and COC (S/COC) or sperm and oocyte (S/O) preincubations. ( ), number ofoocytes. The studies were repeated at least three times.
Dev
elop
men
tal D
ynam
ics
INTEGRINS AND MOUSE FERTILIZATION 775

sperm, we next investigated theirinvolvement in fertilization processby studying the effects of their inhibi-tion using their respective antibodiesor the addition of VN on the fertiliza-tion rate. Fertilization rate (FR; thepercentage of eggs fused with at leastone sperm) and/or the fertilizationindex (FI; the mean number of fusedsperm per egg) were assessed in cu-mulus-intact and zona-free eggs in-semination, respectively. To discrimi-nate the origin of the antibody effect,whether it was acting on integrin sub-units from oocyte, sperm, or both,gametes were separately preincu-bated with the antibodies. Weobserved the involvement of cumulusoocyte complex (COC) av integrinsubunit in the fertilization process.Indeed, preincubation of COC withthe anti-av antibody at 50 mg/ml sig-nificantly inhibited the FR, as com-pared to the control group (32.9 65.3% vs. 79.36 3.9%, respectively; P <0.0001, Fig. 2A). After sperm preincu-bation, the FR was also significantlydecreased when compared with thecontrol group (23.7 6 3.9%; P <0.0001). The inhibitory effect of theantibody was not further increasedwhen both gametes were preincu-bated, suggesting that there is neitherpotentialization nor addition of inhibi-tory effects on both gametes (Fig. 2A).
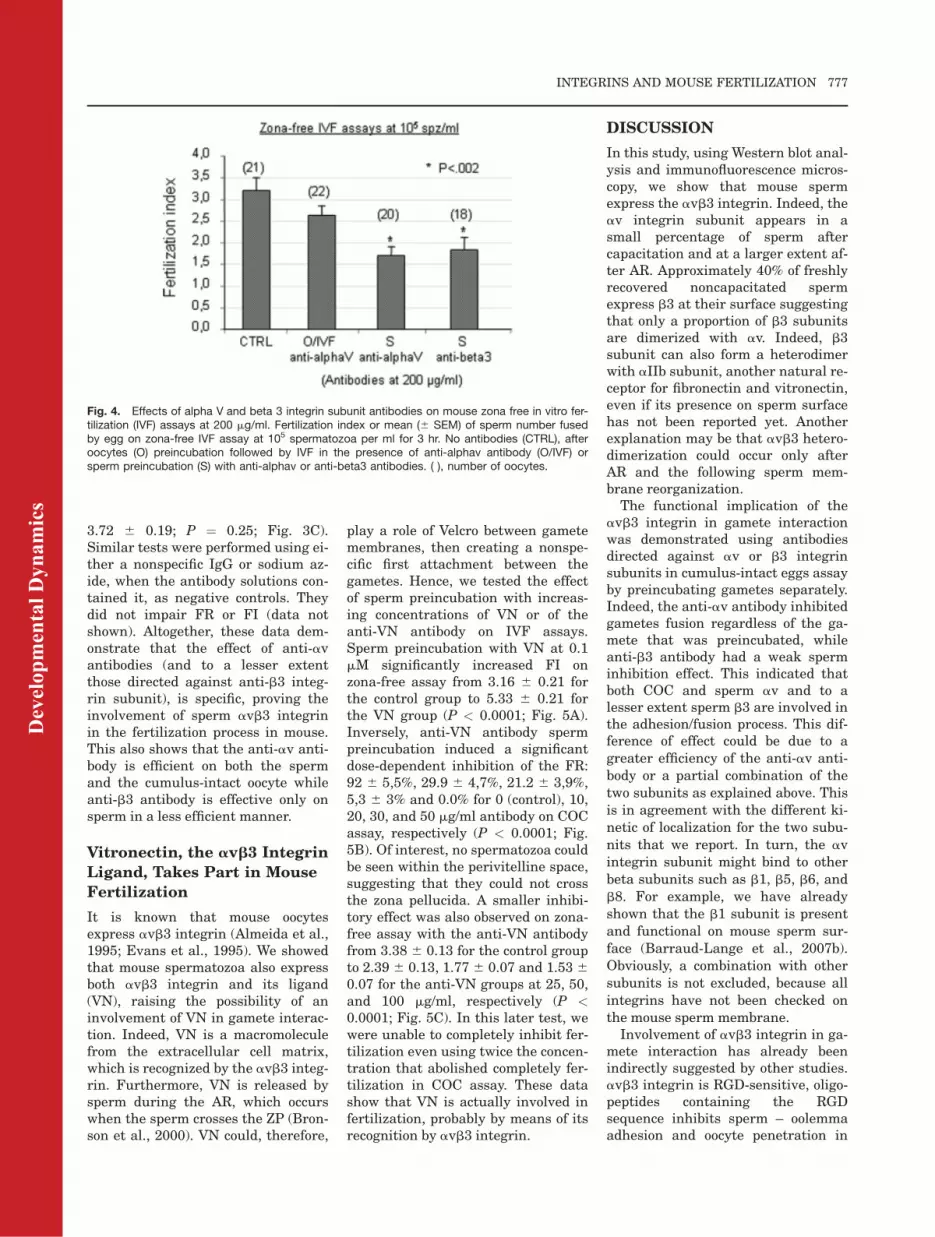
We confirmed on zona-free eggs in-semination assays the effect of theanti-av integrin subunit antibodies(50 mg/ml) on sperm when preincu-bated alone or with oocytes and whenthe antibody was present in theinseminating medium during the IVF(FI: 2.22 6 0.19, 2.40 6 0.17 and 2.776 0.16, respectively, as compared tothe control group at 4.01 6 0.17; P <0.0001; Fig. 2B). After sperm preincu-bation at 200 mg/ml, the inhibitoryeffect remained partial (Fig. 4). But,anti-av mAb (at 50 mg/ml and 200 mg/ml) had no effect on zona-free oocytes(Figs. 2B, 4).
The same experiments were carriedout using the anti-b3 antibody (10 mg/ml), which triggers a modest but sig-nificant decrease of the FR betweenthe control group and the group inwhich IVF was performed in the pres-ence of the antibody (54.5 6 5.3%,32.4 6 5.7%; P ¼ 0.039; Fig. 3A). Nodifferences were observed when theother groups were compared with the
controls (Fig. 3A). The modest inhibi-tory effect of the antibody (10 mg/ml)was also observed on FI in zona-freeassays, only when sperm was preincu-bated alone or with oocytes, and inIVF group (FI: 2.68 6 0.17; P <0.0001; 3.01 6 0.15; P ¼ 0.01; and2.85 6 0.15; P ¼ 0.002 compared withthe control group, following analysis
of variance (ANOVA) and post hoctests at 3.63 6 0.13; Fig. 3B). Incuba-tion of oocytes alone did not induceany significant inhibition. Usingthe antibody at 50 mg/ml, theseresults remained unchanged exceptfor IVF group in which no differencewas observed when compared withthe control group (FI: 3.04 6 0.21 and
Fig. 3. Effects of beta 3 integrin subunit antibody on mouse cumulus-intact (10 mg/ml) andzona free in vitro fertilization (IVF) assays (10 and 50 mg/ml). A: Fertilization rate or mean (6SEM) percentage of fertilized eggs following cumulus-intact IVF assay at 106 spermatozoa perml for 3 hr. B: Fertilization index or mean (6 SEM) of sperm number fused by egg on zona-freeIVF assay at 105 spermatozoa per ml for 3 hr. No antibodies (CTRL), in vitro fertilization in thepresence of antibody but without preincubations (IVF), groups with cumulus oocyte complex(COC), oocyte (O), sperm (S), sperm and COC (S/COC) or sperm and oocyte (S/O) preincuba-tions (antibody concentration:10 mg/ml). The studies were repeated at least three times. C: Thesame experiment that this described in B but at 50 mg/ml of antibody. ( ), number of oocytes.The study was repeated two times.
Dev
elop
men
tal D
ynam
ics
776 BOISSONNAS ET AL.
3.72 6 0.19; P ¼ 0.25; Fig. 3C).Similar tests were performed using ei-ther a nonspecific IgG or sodium az-ide, when the antibody solutions con-tained it, as negative controls. Theydid not impair FR or FI (data notshown). Altogether, these data dem-onstrate that the effect of anti-avantibodies (and to a lesser extentthose directed against anti-b3 integ-rin subunit), is specific, proving theinvolvement of sperm avb3 integrinin the fertilization process in mouse.This also shows that the anti-av anti-body is efficient on both the spermand the cumulus-intact oocyte whileanti-b3 antibody is effective only onsperm in a less efficient manner.
Vitronectin, the avb3 Integrin
Ligand, Takes Part in Mouse
Fertilization
It is known that mouse oocytesexpress avb3 integrin (Almeida et al.,1995; Evans et al., 1995). We showedthat mouse spermatozoa also expressboth avb3 integrin and its ligand(VN), raising the possibility of aninvolvement of VN in gamete interac-tion. Indeed, VN is a macromoleculefrom the extracellular cell matrix,which is recognized by the avb3 integ-rin. Furthermore, VN is released bysperm during the AR, which occurswhen the sperm crosses the ZP (Bron-son et al., 2000). VN could, therefore,
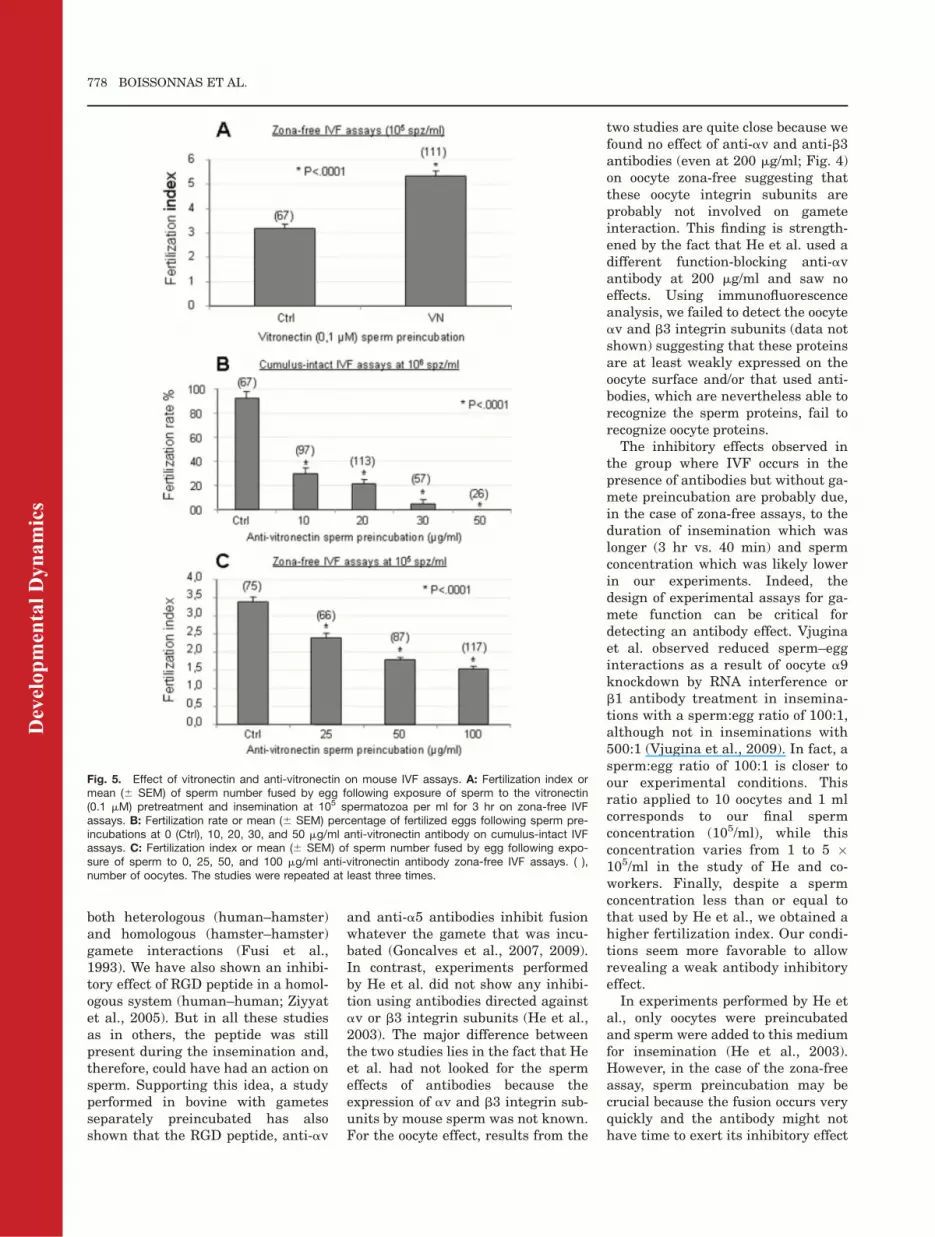
play a role of Velcro between gametemembranes, then creating a nonspe-cific first attachment between thegametes. Hence, we tested the effectof sperm preincubation with increas-ing concentrations of VN or of theanti-VN antibody on IVF assays.Sperm preincubation with VN at 0.1mM significantly increased FI onzona-free assay from 3.16 6 0.21 forthe control group to 5.33 6 0.21 forthe VN group (P < 0.0001; Fig. 5A).Inversely, anti-VN antibody spermpreincubation induced a significantdose-dependent inhibition of the FR:92 6 5,5%, 29.9 6 4,7%, 21.2 6 3,9%,5,3 6 3% and 0.0% for 0 (control), 10,20, 30, and 50 mg/ml antibody on COCassay, respectively (P < 0.0001; Fig.5B). Of interest, no spermatozoa couldbe seen within the perivitelline space,suggesting that they could not crossthe zona pellucida. A smaller inhibi-tory effect was also observed on zona-free assay with the anti-VN antibodyfrom 3.38 6 0.13 for the control groupto 2.39 6 0.13, 1.77 6 0.07 and 1.53 60.07 for the anti-VN groups at 25, 50,and 100 mg/ml, respectively (P <0.0001; Fig. 5C). In this later test, wewere unable to completely inhibit fer-tilization even using twice the concen-tration that abolished completely fer-tilization in COC assay. These datashow that VN is actually involved infertilization, probably by means of itsrecognition by avb3 integrin.
DISCUSSION
In this study, using Western blot anal-ysis and immunofluorescence micros-copy, we show that mouse spermexpress the avb3 integrin. Indeed, theav integrin subunit appears in asmall percentage of sperm aftercapacitation and at a larger extent af-ter AR. Approximately 40% of freshlyrecovered noncapacitated spermexpress b3 at their surface suggestingthat only a proportion of b3 subunitsare dimerized with av. Indeed, b3subunit can also form a heterodimerwith aIIb subunit, another natural re-ceptor for fibronectin and vitronectin,even if its presence on sperm surfacehas not been reported yet. Anotherexplanation may be that avb3 hetero-dimerization could occur only afterAR and the following sperm mem-brane reorganization.The functional implication of the
avb3 integrin in gamete interactionwas demonstrated using antibodiesdirected against av or b3 integrinsubunits in cumulus-intact eggs assayby preincubating gametes separately.Indeed, the anti-av antibody inhibitedgametes fusion regardless of the ga-mete that was preincubated, whileanti-b3 antibody had a weak sperminhibition effect. This indicated thatboth COC and sperm av and to alesser extent sperm b3 are involved inthe adhesion/fusion process. This dif-ference of effect could be due to agreater efficiency of the anti-av anti-body or a partial combination of thetwo subunits as explained above. Thisis in agreement with the different ki-netic of localization for the two subu-nits that we report. In turn, the avintegrin subunit might bind to otherbeta subunits such as b1, b5, b6, andb8. For example, we have alreadyshown that the b1 subunit is presentand functional on mouse sperm sur-face (Barraud-Lange et al., 2007b).Obviously, a combination with othersubunits is not excluded, because allintegrins have not been checked onthe mouse sperm membrane.Involvement of avb3 integrin in ga-
mete interaction has already beenindirectly suggested by other studies.avb3 integrin is RGD-sensitive, oligo-peptides containing the RGDsequence inhibits sperm – oolemmaadhesion and oocyte penetration in
Fig. 4. Effects of alpha V and beta 3 integrin subunit antibodies on mouse zona free in vitro fer-tilization (IVF) assays at 200 mg/ml. Fertilization index or mean (6 SEM) of sperm number fusedby egg on zona-free IVF assay at 105 spermatozoa per ml for 3 hr. No antibodies (CTRL), afteroocytes (O) preincubation followed by IVF in the presence of anti-alphav antibody (O/IVF) orsperm preincubation (S) with anti-alphav or anti-beta3 antibodies. ( ), number of oocytes.
Dev
elop
men
tal D
ynam
ics
INTEGRINS AND MOUSE FERTILIZATION 777
both heterologous (human–hamster)and homologous (hamster–hamster)gamete interactions (Fusi et al.,1993). We have also shown an inhibi-tory effect of RGD peptide in a homol-ogous system (human–human; Ziyyatet al., 2005). But in all these studiesas in others, the peptide was stillpresent during the insemination and,therefore, could have had an action onsperm. Supporting this idea, a studyperformed in bovine with gametesseparately preincubated has alsoshown that the RGD peptide, anti-av
and anti-a5 antibodies inhibit fusionwhatever the gamete that was incu-bated (Goncalves et al., 2007, 2009).In contrast, experiments performedby He et al. did not show any inhibi-tion using antibodies directed againstav or b3 integrin subunits (He et al.,2003). The major difference betweenthe two studies lies in the fact that Heet al. had not looked for the spermeffects of antibodies because theexpression of av and b3 integrin sub-units by mouse sperm was not known.For the oocyte effect, results from the
two studies are quite close because wefound no effect of anti-av and anti-b3antibodies (even at 200 mg/ml; Fig. 4)on oocyte zona-free suggesting thatthese oocyte integrin subunits areprobably not involved on gameteinteraction. This finding is strength-ened by the fact that He et al. used adifferent function-blocking anti-avantibody at 200 mg/ml and saw noeffects. Using immunofluorescenceanalysis, we failed to detect the oocyteav and b3 integrin subunits (data notshown) suggesting that these proteinsare at least weakly expressed on theoocyte surface and/or that used anti-bodies, which are nevertheless able torecognize the sperm proteins, fail torecognize oocyte proteins.The inhibitory effects observed in
the group where IVF occurs in thepresence of antibodies but without ga-mete preincubation are probably due,in the case of zona-free assays, to theduration of insemination which waslonger (3 hr vs. 40 min) and spermconcentration which was likely lowerin our experiments. Indeed, thedesign of experimental assays for ga-mete function can be critical fordetecting an antibody effect. Vjuginaet al. observed reduced sperm–egginteractions as a result of oocyte a9knockdown by RNA interference orb1 antibody treatment in insemina-tions with a sperm:egg ratio of 100:1,although not in inseminations with500:1 (Vjugina et al., 2009). In fact, asperm:egg ratio of 100:1 is closer toour experimental conditions. Thisratio applied to 10 oocytes and 1 mlcorresponds to our final spermconcentration (105/ml), while thisconcentration varies from 1 to 5 �105/ml in the study of He and co-workers. Finally, despite a spermconcentration less than or equal tothat used by He et al., we obtained ahigher fertilization index. Our condi-tions seem more favorable to allowrevealing a weak antibody inhibitoryeffect.In experiments performed by He et
al., only oocytes were preincubatedand sperm were added to this mediumfor insemination (He et al., 2003).However, in the case of the zona-freeassay, sperm preincubation may becrucial because the fusion occurs veryquickly and the antibody might nothave time to exert its inhibitory effect
Fig. 5. Effect of vitronectin and anti-vitronectin on mouse IVF assays. A: Fertilization index ormean (6 SEM) of sperm number fused by egg following exposure of sperm to the vitronectin(0.1 mM) pretreatment and insemination at 105 spermatozoa per ml for 3 hr on zona-free IVFassays. B: Fertilization rate or mean (6 SEM) percentage of fertilized eggs following sperm pre-incubations at 0 (Ctrl), 10, 20, 30, and 50 mg/ml anti-vitronectin antibody on cumulus-intact IVFassays. C: Fertilization index or mean (6 SEM) of sperm number fused by egg following expo-sure of sperm to 0, 25, 50, and 100 mg/ml anti-vitronectin antibody zona-free IVF assays. ( ),number of oocytes. The studies were repeated at least three times.
Dev
elop
men
tal D
ynam
ics
778 BOISSONNAS ET AL.

which is not immediate and requiresa delay to be effective.
In the case of cumulus-intactassays, the effect observed in thegroup in which IVF is performed inthe presence of antibodies is probablydue, in addition to the possible effecton the cumulus cells that express atleast av integrin subunit on their sur-face (Tamba et al., 2008), to the delayrequired for sperm to cross the cumu-lus and the ZP before gametes fusion.It represents a necessary sperm pre-incubation step. In this regard, thiseffect is closer to that observed aftersperm preincubation.
When only COC was preincubated,sperm effect could be due to residualantibodies remaining trapped by theviscous extracellular matrix. It is cer-tainly difficult to completely get rid ofantibodies with which it wasincubated.
Sperm and cumulus cells inhibitoryeffect of anti-av and anti-b3 integrinsubunits antibodies remains partial,confirming that they participate butare not the only molecules involved ingamete interaction.
Deletions of av integrin subunitgene led to the death of the majorityof the fetuses at day 9.5, and thosewhich survived until birth died frombrain or intestine hemorrhage (Baderet al., 1998). These data highlight thenecessity to explore the role of av sub-unit in mouse fertilization. The factthat b3 integrin subunit null mice areviable and fertile (Hodivala-Dilke etal., 1999) does not exclude a possibleredundancy for this integrin subunit.Conditional deletions of these integ-rin subunits have never been investi-gated, but even when such experi-ments were performed for otherintegrin subunits, focus was only onthe oocyte, never on the spermatozoonor on the cumulus cells (Miller et al.,2000; He et al., 2003).
Our data reveal that the avb3integrin plays a role in gamete inter-action, and most probably by meansof VN. Hence, we evaluated the role ofthis latter using exogenous VN or ananti-VN antibody. Anti-VN antibodyinhibited the gamete fusion on cumu-lus-intact (completely at 50 mg/ml)and on ZP-free IVF assays in a dose-dependent manner. The fact thatanti-VN antibody did not completelyabolish fertilization in zona-free
assays suggests that a substantialpart of its effect possibly takes placethrough an action on the cumuluscells or on ZP sperm interaction. Inaddition, this result could beexplained by membrane changes dueto ZP removal. Actually, VN acts bymeans of its receptors, oocyte integ-rins, which organization is modifiedby ZP removal (Ziyyat et al., 2006;Barraud-Lange et al., 2007b). Addi-tion of 0.1 mM VN to the medium dur-ing zona-free oocytes inseminationpromoted human gametes fusion, asalready observed by Fusi et al. (Fusiet al., 1996a). They showed that thisfusion promotion was probably due tothe VN promotion of calcium depend-ent sperm – oolemma binding. Theyhave also observed that repeatedwashings of the capacitated sperma-tozoa led to a reduced binding, whichcould be restored by the addition ofVN to the medium. In our study, pro-longed sperm incubation (3 hr) withthe same VN concentration (0.1 mM)caused their aggregation (data notshown). This is probably due to thedynamic emergence of VN receptors(integrin avb3 and others) duringcapacitation and AR. The fact thatavb3 integrin appears mostly at themoment of AR and that it is preciselyat this time that VN is released(Bronson et al., 2000), leads us to pro-pose the following scenario: VN isreleased during acrosome reactionwhen the sperm enters the perivitel-line space. Gamete integrins capturesome of the released VN, which afterdimeric or multimeric formation couldthen play a role as Velcro betweenoocyte and sperm avb3 integrins.Such nonspecific adhesion would ena-ble oocyte and spermatozoon to sticktogether, the time necessary for a6b1integrin, fertilin b, cyritestin andprobably other molecules to interact.As VN possesses only a single RGDsequence, we hypothesize that a VNmolecule binds to another, through anon-RGD motif to form at least adimer. The RGD sequence located atthe carboxy-terminal boundary of thesomatomedin B domain of VN (Suzukiet al., 1984) appears to mediate bind-ing of VN to distinct members ofintegrins, in particular to the VN re-ceptor, and is responsible for the cellattachment activity of VN and otheradhesive proteins (Ruoslahti and
Pierschbacher, 1986). Multimericforms of VN have been shown to exist(Sane et al., 1988, 1990; Hess et al.,1995). In this scenario, within themultimeric molecule, the RGD site ofeach VN molecule remaining free canmake a bridge between the two game-tes through the recognition of theseRGD sites by integrins. This scenariowould also explain the phenomenon ofcalcium-dependent sperm agglutina-tion in the presence of a large concen-tration of VN previously described(Fusi et al., 1996a; Fig. 6A). A secondnonexclusive scenario could be thatsperm capture some released VN bymeans of RGD site. VN is a conforma-tionally labile molecule that possessesa cryptic heparin binding site whichis not expressed in the soluble form,but which makes its appearancewhen VN is immobilized. Human andmouse eggs have been shown to syn-thesize proteoglycans (Tesarik andKopecny, 1986; Zhuo and Kimata,2001; Russell and Salustri, 2006;Romanato et al., 2008) which couldprovide a secondary binding site forVN (Fig. 6B). Because the used anti-bodies did not allow either to stain orinhibit oocyte integrins (at least onthe zona-free oocytes), we favor thissecond scenario.We propose that these models
account for the phenomena that occurin vitro. Two categories of spermato-zoa are observed on the oolemma: the‘‘loosely bound’’ that could correspondto spermatozoa linked to the oocytemembrane only through VN, and the‘‘tightly bound’’ that could be doublylinked to the oolemma by both a6b1and avb3 integrins and probablyother molecules.Homozygous null mice completely
deficient in VN demonstrate normalsurvival, development, and fertilitysuggesting that its role in these proc-esses may overlap with other adhe-sive matrix components (Zheng et al.,1995). Fibronectin, which is alsoexpressed by sperm, could providethis redundancy. Because of the em-bryonic lethality of fibronectin nullmutant mouse, the essential nature ofits role in fertilization could not beassessed (George et al., 1993). How-ever, its participation in fertilizationhas been widely documented in sev-eral species (Fusi et al., 1992, 1996a;Thys et al., 2009).
Dev
elop
men
tal D
ynam
ics
INTEGRINS AND MOUSE FERTILIZATION 779

We and others demonstrated par-tial gamete membrane symmetrywith a6b1 integrin, avb3 integrinand fertilinb (personal observations)expressed by both gametes. This par-tial gamete membrane symmetry, of-ten unknown or overlooked or notenough taken into account, could besignificantly enhanced by the mem-brane fragment transfer that occursin perivitelline space by trogocytosisand/or exosome-like vesicles (Bar-raud-Lange et al., 2007a; Miyado
et al., 2008). According to this lastpublication, exosomes derived fromWT CD9 oocytes can associate withsperm and make them able to fusewith either WT or CD9 null oocytes.But this assertion concerning func-tional role of these transferred exo-somes has been questioned by otherwork (Gupta et al., 2009). However,CD9, which is not expressed bysperm (Chen et al., 1999), is actuallytransferred from the oocyte to thesperm present in the perivitelline
space. We have shown that thistransfer affects entire CD9-contain-ing membrane fragments (Barraud-Lange et al., 2007a). Because integ-rins are associated with CD9 tetra-spanin that controls their reorgan-ization during fertilization (Ziyyatet al., 2006), it is likely that integ-rins are also transferred with CD9tetraspanin. After membrane frag-ments transfer and adhesion step, itwill then form the molecular com-plexes common to both membranes
Fig. 6. Hypothetical models of participation of vitronectin (VN), integrins and related molecules on mouse sperm–egg adhesion and fusion. Impor-tant sperm molecules for interaction with oocyte oolemma are not detected on the surface of freshly recovered sperm. When sperm are capaci-tated, a part of some proteins such as integrins appear at the surface (1). It is the acrosome reaction that reveals all the molecules involved in theprocess of membrane–membrane interaction and in particular integrins and IZUMO (2). At this moment, a part of sperm released VN is capturedby sperm avb3 integrin. As VN possesses only a single RGD sequence, two nonexclusive scenarios are possible: A: This VN would play a role asVelcro between oocyte and sperm avb3 integrins (3). In this case, VN acts as multimeric form (at least dimer) in which VN molecules bind to themby a site other than RGD to kept free the RGD site to link oocyte avb3 integrin. B: Alternatively and more likely, mouse eggs which have beenshown to synthesize proteoglycans could provide a secondary binding site for VN. Indeed, VN is a conformationally labile molecule that possessesa cryptic heparin binding site which is not expressed in the soluble form, but which makes its appearance when VN adheres to solid phase surfa-ces. When spermatozoon is in the perivitelline space, due to repeated contact with oocyte membrane or Exosome-like vesicles present in the peri-vitelline space, it captures oocyte membrane fragments containing the tetraspanin CD9 and its partners. We postulate that this CD9 allows thesperm to organize molecular complexes that would deal with those existing on the oocyte side.
Dev
elop
men
tal D
ynam
ics
780 BOISSONNAS ET AL.

in which the absence of a moleculeon one side could be offset by itspresence on the other side. This isprobably the reason why the absenceof one of these ‘‘not essential but con-tributing’’ molecules does not preventgamete interaction (Miller et al.,2000; He et al., 2003). In contrast,molecules whose absence leads to adrastic effect play a much more im-portant role, as it is the case of CD9to which the organizer role of molec-ular complex has been widelyattributed.
EXPERIMENTAL
PROCEDURES
Antibodies and Peptides
For Western blot analysis, rabbit pol-yclonal antibodies against mouse avintegrin subunit (Fitzgerald, USA),b3 integrin subunit (Biosource, USA),and vitronectin (Santa Cruz Biotech-nology, USA) were used, the negativecontrol being rabbit IgGHþL and thesecondary antibody a biotinylatedanti-rabbit (Abcys S.A., France). Thebands were detected with streptavi-din conjugated to HRP (horseradishperoxidase [HRP]) enzyme (Immuno-tech, France).
To perform functional blocking testsand immunofluorescence experi-ments, a rat monoclonal antibodyagainst the mouse av integrin subunit(RMV-7), its isotype, a rat IgG1k (R3-34), a hamster monoclonal antibodyagainst the mouse b3 integrin subu-nit (2C9-G2) and its isotype, and anhamster IgG1 were purchased fromBD Pharmingen (USA). Vitronectinand anti-vitronectin antibody (H-270)were purchased from Sigma andSanta Cruz, respectively. For immu-nolabeling purpose, an anti-rat biotin-ylated secondary antibody (BD Phar-mingen) and an Alexa Fluor 594conjugated streptavidin (Invitrogen,France) were used to detect the sig-nal. Detection of the AR was per-formed using Pisum sativum aggluti-nin conjugated to fluorescein (PSA-FITC, Invitrogen).
Western Blot Analysis
Capacitated sperm were washed twicein phosphate buffered saline (PBS),the pellet snap-frozen in liquidN2 and stored at �80�C for further
use. Sperm aliquots were lysed in50 mM Tris (pH 8.0), 150 mM NaCl,1 mM ethylenediaminetetraaceticacid (EDTA), 0.25% sodium deoxycho-late, and 1% NP40, supplementedwith Protease Inhibitor Cocktail(Sigma) for 1 hr on ice, gently soni-cated, and then boiled for 5 min withan equal volume of 2� NuPAGE LDSsample buffer (Invitrogen) supple-mented with 5% b-mercaptoethanol.F9 Whole Cell Lysate (Abcam,France) was used at 50 mg proteins asa positive control for the av and b3integrin subunits detection. Sampleproteins were separated by a precastgel electrophoresis (SoftGel, USA)and electro-transferred to Immobilon-P membranes (Millipore, France).Membranes were blocked for 1 hrwith 2% casein before incubation withappropriate primary antibodies (rab-bit anti-av integrin: 1:5,000; rabbitanti-b3 integrin: 0.5 mg/ml, and anonspecific rabbit IgG: 0.5 mg/ml)for 90 min at 37�C, followed bybiotinylated appropriate secondaryantibodies (0.5 mg/ml) and HRP-Streptavidin (0.02 mg/ml), for 1 hrand 45 min at 37�C, respectively. Vi-tronectin expression was revealed byanti-vitronectin antibody at 1:200dilution and by secondary HRP conju-gated (0.5 mg/ml) antibody. HRP activ-ity was revealed by ECL detection kit(GE Healthcare, France) and autora-diography (BioMax Light Film,Kodac). Film exposure was less than3 min.
Sperm Immunolabeling and
Fluorescence Microscopy
Freshly recovered, capacitated sper-matozoa or calcium ionophoreinduced AR spermatozoa (10 mM Cal-cium ionophore A23187, for 30 min at37�C, Sigma) were washed in PBScontaining 1% bovine serum albumin(BSA), centrifuged at 300 g for 10min. After washing, the unfixed sper-matozoa were incubated in PBS con-taining 10 mg/ml of rat anti-av orhamster anti-b3 integrin antibodiesfor 70 min at 37�C and then with 10mg/ml of the anti-rat or anti-hamsterbiotinylated antibody (for av and b3,respectively) for 45 min at room tem-perature (RT). To detect the staining,Alexa 594 coupled streptavidin (10mg/ml) was applied during 30 min at
RT. Negative controls were performedby omitting the first antibody. Thecells were then submitted to the PSA-staining protocol for the sequentialdetection of integrin distribution andacrosomal status. The spermatozoawere stained with fluorescein isothio-cyanate (FITC) -conjugated lectinPSA (25 mg/ml in PBS) for 10 min. Af-ter repeated washing with double dis-tilled water, a drop of sperm suspen-sion was smeared on slide, air-dried,mounted with Vectashield/DAPI(40,6-diamidine-2-phenylidole-dihydro-chloride) and covered with a coverslipfor analysis. Detection was performedusing a Zeiss Axiophot epifluorescencemicroscope and images were digitallyacquired with a camera (Coolpix 4500,Nikon).
Gamete Preparation and
In Vitro Fertilization
Oocytes.
B6CBA F1 female mice (5–8 weeksold), purchased from Charles RiverLaboratories (France), were supero-vulated with 5 IU PMSG and 5 IUhCG (Intervet, France) 48 hr apart.At 12 to 14 hr after hCG injection, theanimals were killed by cervical dislo-cation. Cumulus oophorus were col-lected by tearing the ampulla wall ofthe oviduct, placed in Whittingham’smedium (Whittingham, 1971) supple-mented with 3% BSA (Sigma), andmaintained at 37�C under 5% CO2 inair under mineral oil (Sigma). Whenexperiments were run with zona-freeoocytes, cumulus cells were removedby a brief exposure to hyaluronidase(0.01%, Sigma). The ZP was then dis-solved with acidic Tyrode’s (AT) solu-tion (pH 2.5) (Sigma) under visualmonitoring. The zona-free eggs wererapidly washed five times and kept at37�C under 5% CO2 in air for 2 to 3 hrto recover their fertilization ability.
Sperm preparation.
Mouse spermatozoa were obtainedfrom the caudae epididymis of B6CBAF1 male mice (8–13 weeks old) andcapacitated at 37�C under 5% CO2 for90 min in a 500-ml drop of Whitting-ham’s medium supplemented with 30mg/ml BSA, under mineral oil.
Dev
elop
men
tal D
ynam
ics
INTEGRINS AND MOUSE FERTILIZATION 781

In vitro fertilization.
Cumulus-intact and zona-free eggswere inseminated with capacitatedspermatozoa for 3 hr in a 50-ml dropof medium at a final concentration of106/ml or 105/ml, respectively. Then,they were washed and directlymounted in Vectashield/DAPI for ob-servation under UV light (Zeiss Axio-skop 20 microscope). The oocytes wereconsidered fertilized when theyshowed at least one fluorescent decon-densed sperm head within theircytoplasm.
To test the effect of antibodies onthe fertilization rates (FR) or fertiliza-tion index (FI), before in vitro fertil-ization (IVF), either one or bothgametes were separately preincu-bated for 30 min in medium supple-mented with anti-av or anti-b3 anti-bodies at 50 or 10 and 50 mg/ml,respectively. Oocytes were thenwashed, and sperm were diluted1:100 into the final droplet containingthe oocytes. In parallel, inseminationwas performed in medium containingantibodies without any gametepreincubations.
Effects of VN or anti-VN antibodywere assessed by sperm preincuba-tion in a medium containing eitherVN at 0.1 mM or the anti-VN antibodyat 10, 20, 30, or 50 mg/ml for 30 min.
Statistical Analysis
Statistical analysis was performedusing standard ANOVA. Differenceswere considered significant at P< 0.05.
ACKNOWLEDGMENTSWe thank K. Senni for constructivediscussion about the role of thevitronectin.
REFERENCES
Almeida EA, Huovila AP, Sutherland AE,Stephens LE, Calarco PG, Shaw LM,Mercurio AM, Sonnenberg A, PrimakoffP, Myles DG, et al. 1995. Mouse eggintegrin alpha 6 beta 1 functions as asperm receptor. Cell 81:1095–1104.
Bader BL, Rayburn H, Crowley D, HynesRO. 1998. Extensive vasculogenesis,angiogenesis, and organogenesis pre-cede lethality in mice lacking all alphav integrins. Cell 95:507–519.
Baessler KA, Lee Y, Sampson NS. 2009.Beta1 integrin is an adhesion proteinfor sperm binding to eggs. ACS ChemBiol 4:357–366.
Barraud-Lange V, Naud-Barriant N, Bom-sel M, Wolf JP, Ziyyat A. 2007a. Trans-fer of oocyte membrane fragments tofertilizing spermatozoa. FASEB J 21:3446–3449.
Barraud-Lange V, Naud-Barriant N, Saf-far L, Gattegno L, Ducot B, Drillet AS,Bomsel M, Wolf JP, Ziyyat A. 2007b.Alpha6beta1 integrin expressed bysperm is determinant in mouse fertil-ization. BMC Dev Biol 7:102.
Bronson RA, Fusi F. 1990. Sperm-oolem-mal interaction: role of the Arg-Gly-Asp(RGD) adhesion peptide. Fertil Steril54:527–529.
Bronson RA, Fusi FM, Calzi F, Doldi N,Ferrari A. 1999. Evidence that a func-tional fertilin-like ADAM plays a role inhuman sperm-oolemmal interactions.Mol Hum Reprod 5:433–440.
Bronson R, Peresleni T, Golightly M, Pre-issner K. 2000. Vitronectin is seques-tered within human spermatozoa andliberated following the acrosome reac-tion. Mol Hum Reprod 6:977–982.
Burns KH, Owens GE, Fernandez JM,Nilson JH, Matzuk MM. 2002. Charac-terization of integrin expression in themouse ovary. Biol Reprod 67:743–751.
Campbell S, Swann HR, Seif MW, KimberSJ, Aplin JD. 1995. Cell adhesion mole-cules on the oocyte and preimplantationhuman embryo. Hum Reprod 10:1571–1578.
Chen H, Sampson NS. 1999. Mediation ofsperm-egg fusion: evidence that mouseegg alpha6beta1 integrin is the receptorfor sperm fertilinbeta. Chem Biol 6:1–10.
Chen MS, Tung KS, Coonrod SA, Takaha-shi Y, Bigler D, Chang A, Yamashita Y,Kincade PW, Herr JC, White JM. 1999.Role of the integrin-associated proteinCD9 in binding between sperm ADAM 2and the egg integrin alpha6beta1: impli-cations for murine fertilization. ProcNatl Acad Sci U S A 96:11830– 11835.
Evans JP. 2009. Egg integrins: back inthe game of mammalian fertilization.ACS Chem Biol 4:321–323.
Evans JP, Schultz RM, Kopf GS. 1995.Identification and localization of integ-rin subunits in oocytes and eggs of themouse. Mol Reprod Dev 40:211–220.
Fusi FM, Vignali M, Busacca M, BronsonRA. 1992. Evidence for the presence ofan integrin cell adhesion receptor onthe oolemma of unfertilized humanoocytes. Mol Reprod Dev 31:215–222.
Fusi FM, Bernocchi N, Ferrari A, Bron-son RA. 1996a. Is vitronectin the velcrothat binds the gametes together? MolHum Reprod 2:859–866.
Fusi FM, Tamburini C, Mangili F, Monte-sano M, Ferrari A, Bronson RA. 1996b.The expression of alpha v, alpha 5, beta1, and beta 3 integrin chains on ejacu-lated human spermatozoa varies withtheir functional state. Mol Hum Reprod2:169–175.
Fusi FM, Vignali M, Gailit J, BronsonRA. 1993. Mammalian oocytes exhibitspecific recognition of the RGD (Arg-Gly-Asp) tripeptide and express oolem-
mal integrins. Mol Reprod Dev 36:212–219.
George EL, Georges-Labouesse EN, Patel-King RS, Rayburn H, Hynes RO. 1993.Defects in mesoderm, neural tube andvascular development in mouseembryos lacking fibronectin. Develop-ment 119:1079–1091.
Goncalves RF, Wolinetz CD, Killian GJ.2007. Influence of arginine-glycine-as-partic acid (RGD), integrins (alpha(V)and alpha(5)) and osteopontin on bovinesperm-egg binding, and fertilization invitro. Theriogenology 67:468–474.
Goncalves RF, Bertolla RP, Mortara RA,Barnabe VH. 2009. alpha(6), beta(1),and beta(3) integrins expressed bysperm may be involved in cattle fertil-ization. Reprod Fertil Dev 21:201.
Gupta S, Primakoff P, Myles DG. 2009.Can the presence of wild-type oocytesduring insemination rescue the fusiondefect of CD9 null oocytes? Mol ReprodDev 76:602.
He ZY, Brakebusch C, Fassler R, KreidbergJA, Primakoff P, Myles DG. 2003. Noneof the integrins known to be present onthe mouse egg or to be ADAM receptorsare essential for sperm-egg binding andfusion. Dev Biol 254:226–237.
Hess S, Kanse SM, Kost C, Preissner KT.1995. The versatility of adhesion recep-tor ligands in haemostasis: morpho-regulatory functions of vitronectin.Thromb Haemost 74:258–265.
Hodivala-Dilke KM, McHugh KP, TsakirisDA, Rayburn H, Crowley D, Ullman-Cullere M, Ross FP, Coller BS, Teitel-baum S, Hynes RO. 1999. Beta3-integ-rin-deficient mice are a model forGlanzmann thrombasthenia showingplacental defects and reduced survival.J Clin Invest 103:229–238.
Inoue N, Ikawa M, Isotani A, Okabe M.2005. The immunoglobulin superfamilyprotein Izumo is required for sperm tofuse with eggs. Nature 434:234–238.
Kaji K, Oda S, Shikano T, Ohnuki T,Uematsu Y, Sakagami J, Tada N, Miya-zaki S, Kudo A. 2000. The gametefusion process is defective in eggs ofCd9-deficient mice. Nat Genet 24:279–282.
Le Naour F, Rubinstein E, Jasmin C, Pre-nant M, Boucheix C. 2000. Severelyreduced female fertility in CD9-defi-cient mice. Science 287:319–321.
Miller BJ, Georges-Labouesse E, Primak-off P, Myles DG. 2000. Normal fertiliza-tion occurs with eggs lacking theintegrin alpha6beta1 and is CD9-de-pendent. J Cell Biol 149:1289–1296.
Miyado K, Yamada G, Yamada S, HasuwaH, Nakamura Y, Ryu F, Suzuki K,Kosai K, Inoue K, Ogura A, Okabe M,Mekada E. 2000. Requirement of CD9on the egg plasma membrane for fertil-ization. Science 287:321–324.
Miyado K, Yoshida K, Yamagata K, Saka-kibara K, Okabe M, Wang X, MiyamotoK, Akutsu H, Kondo T, Takahashi Y,Ban T, Ito C, Toshimori K, NakamuraA, Ito M, Miyado M, Mekada E, Ume-zawa A. 2008. The fusing ability of
Dev
elop
men
tal D
ynam
ics
782 BOISSONNAS ET AL.

sperm is bestowed by CD9-containingvesicles released from eggs in mice.Proc Natl Acad Sci U S A 105:12921–12926.
Romanato M, Julianelli V, Zappi M, CalvoL, Calvo JC. 2008. The presence of hep-aran sulfate in the mammalian oocyteprovides a clue to human sperm nucleardecondensation in vivo. Hum Reprod23:1145–1150.
Ruoslahti E, Pierschbacher MD. 1986.Arg-Gly-Asp: a versatile cell recognitionsignal. Cell 44:517–518.
Russell DL, Salustri A. 2006. Extra-cellular matrix of the cumulus-oocytecomplex. Semin Reprod Med 24:217–227.
Sane DC, Moser TL, Pippen AM, ParkerCJ, Achyuthan KE, Greenberg CS.1988. Vitronectin is a substrate fortransglutaminases. Biochem BiophysRes Commun 157:115–120.
Sane DC, Moser TL, Parker CJ, SeiffertD, Loskutoff DJ, Greenberg CS. 1990.Highly sulfated glycosaminoglycansaugment the cross-linking of vitronectinby guinea pig liver transglutaminase.Functional studies of the cross-linkedvitronectin multimers. J Biol Chem265:3543–3548.
Sengoku K, Takuma N, Miyamoto T,Horikawa M, Ishikawa M. 2004. Integ-rins are not involved in the process of
human sperm-oolemmal fusion. HumReprod 19:639–644.
Suzuki S, Pierschbacher MD, HaymanEG, Nguyen K, Ohgren Y, Ruoslahti E.1984. Domain structure of vitronectin.Alignment of active sites. J Biol Chem259:15307–15314.
Tamba S, Yodoi R, Segi-Nishida E, Ichi-kawa A, Narumiya S, Sugimoto Y.2008. Timely interaction between pros-taglandin and chemokine signaling is aprerequisite for successful fertilization.Proc Natl Acad Sci U S A 105:14539–14544.
Tesarik J, Kopecny V. 1986. Late preovu-latory synthesis of proteoglycans by thehuman oocyte and cumulus cells andtheir secretion into the oocyte-cumulus-complex extracellular matrices. Histo-chemistry 85:523–528.
Thys M, Nauwynck H, Maes D, HoogewijsM, Vercauteren D, Favoreel H, Van SoomA. 2009. Expression and putative func-tion of fibronectin and its receptor (integ-rin alpha5 beta1) in male and femalegametes during bovine fertilization invitro. Reproduction 138:471–482.
Vjugina U, Zhu X, Oh E, Bracero NJ,Evans JP. 2009. Reduction of mouse eggsurface integrin alpha9 subunit(ITGA9) reduces the egg’s ability tosupport sperm-egg binding and fusion.Biol Reprod 80:833–841.
Whittingham DG. 1971. Culture of mouseova. J Reprod Fertil Suppl 14:7–21.
Zheng X, Saunders TL, Camper SA,Samuelson LC, Ginsburg D. 1995. Vitro-nectin is not essential for normal mam-malian development and fertility. ProcNatl Acad Sci U S A 92:12426–12430.
Zhuo L, Kimata K. 2001. Cumulus oopho-rus extracellular matrix: its construc-tion and regulation. Cell Struct Funct26:189–196.
Ziyyat A, Naud-Barriant N, Barraud-Lange V, Chevalier F, Kulski O, Lem-kecher T, Bomsel M, Wolf JP. 2005.Cyclic FEE peptide increases humangamete fusion and potentiates its RGD-induced inhibition. Hum Reprod 20:3452–3458.
Ziyyat A, Rubinstein E, Monier-Gavelle F,Barraud V, Kulski O, Prenant M, Bou-cheix C, Bomsel M, Wolf JP. 2006. CD9controls the formation of clusters thatcontain tetraspanins and the integrin{alpha}6{beta}1, which are involved inhuman and mouse gamete fusion. JCell Sci 119:416–424.
Zuccotti M, Giorgi Rossi P, Fiorillo E,Garagna S, Forabosco A, Redi CA.1998. Timing of gene expression andoolemma localization of mouse alpha6and beta1 integrin subunits duringoogenesis. Dev Biol 200:27–34.
Dev
elop
men
tal D
ynam
ics
INTEGRINS AND MOUSE FERTILIZATION 783
Related Documents








![Sperm DNA Fragmentation is Significantly Increased in ... · Sperm DNA fragmentation assessment The sperm DNA damage was evaluated by Sperm Chromatin Dispersion (SCD) test [23] using](https://static.cupdf.com/doc/110x72/5f3a6b0098469b5f937b3512/sperm-dna-fragmentation-is-significantly-increased-in-sperm-dna-fragmentation.jpg)


