Journal of Chemical Ecology, Vol. 12, No. 3, 1986 ROLE OF SECONDARY METABOLITES IN FEEDING ASSOCIATIONS BETWEEN A PREDATORY NUDIBRANCH, TWO GRAZING NUDIBRANCHS, AND A BRYOZOAN BRAD CARTt~ and D. JOHN FAULKNER Scripps Institution of Oceanography University of California, San Diego La Jolla, California 92093 (Received April 22, 1985; accepted August 2, 1985) Abstract--The carnivorous nudibranch Roboastra tigris preys preferentially upon two nudibranchs, Tambja abdere and T. eliora, that in turn feed upon the bryozoan Sessibugula translucens. All four organisms contain tambja- mines A-D (I-IV) that were shown to be fish feeding inhibitors. When at- tacked by Roboastra, T. abdere secretes a distasteful mucus containing a total of 3 mg of the tambjamines that sometimes causes the Roboastra to break off the attack. Under similar circumstances T. eliora attempts to swim away; it presumably contains insufficient of the tambjamines to deter Roboastra. Ro- boastra follows the slime trail of nudibranchs using contact chemoreception and reverses direction when the trail is broken. The slime trail of T. abdere contains low concentrations of the tambjamines. In Y-maze experiments, T. eliora was attracted towards seawater containing S. translucens and seawater containing 10-1o M tambjamines A and B (1 : 1) but was repelled by seawater containing > 10 -8 M tambjamines A and B. At higher concentrations the mixture of tambjamines may be recognized as an alarm pheromone. Key Words--Roboastra tigris, Tambja abdere, Tambja eliora, Sessibugula translucens, tambjamines A-D, chemical defense, alarm pheromones, trail- following behavior. INTRODUCTION Gastropod mollusks of the subclass Opisthobranchia do not seem to derive any physical protection from a shell which, if present, is often reduced or covered by soft tissue. In the Nudibranchia, a shell is completely lacking. The lack of a shell is correlated with alternative defense mechanisms (Faulkner and Ghi- 795 0098-0331/86/0300-0795505.00/0 1986 Plenum Publishing Corporation

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Chemical Ecology, Vol. 12, No. 3, 1986
ROLE OF SECONDARY METABOLITES IN FEEDING ASSOCIATIONS BETWEEN A
PREDATORY NUDIBRANCH, TWO GRAZING NUDIBRANCHS, AND A BRYOZOAN
B R A D C A R T t ~ and D. J O H N F A U L K N E R
Scripps Institution of Oceanography University of California, San Diego
La Jolla, California 92093
(Received April 22, 1985; accepted August 2, 1985)
Abstract--The carnivorous nudibranch Roboastra tigris preys preferentially upon two nudibranchs, Tambja abdere and T. eliora, that in turn feed upon the bryozoan Sessibugula translucens. All four organisms contain tambja- mines A-D (I-IV) that were shown to be fish feeding inhibitors. When at- tacked by Roboastra, T. abdere secretes a distasteful mucus containing a total of 3 mg of the tambjamines that sometimes causes the Roboastra to break off the attack. Under similar circumstances T. eliora attempts to swim away; it presumably contains insufficient of the tambjamines to deter Roboastra. Ro- boastra follows the slime trail of nudibranchs using contact chemoreception and reverses direction when the trail is broken. The slime trail of T. abdere contains low concentrations of the tambjamines. In Y-maze experiments, T. eliora was attracted towards seawater containing S. translucens and seawater containing 10-1o M tambjamines A and B (1 : 1) but was repelled by seawater containing > 10 -8 M tambjamines A and B. At higher concentrations the mixture of tambjamines may be recognized as an alarm pheromone.
Key Words--Roboastra tigris, Tambja abdere, Tambja eliora, Sessibugula translucens, tambjamines A-D, chemical defense, alarm pheromones, trail- following behavior.
INTRODUCTION
G a s t r o p o d m o l l u s k s o f the subc la s s O p i s t h o b r a n c h i a do not s e e m to de r ive any
phys i ca l p r o t e c t i o n f r o m a she l l w h i c h , i f p r e sen t , is o f t en r educed o r c o v e r e d
by sof t t i ssue . In the N u d i b r a n c h i a , a she l l is c o m p l e t e l y l ack ing . T h e lack o f
a she l l is co r r e l a t ed w i th a l t e r na t i ve d e f e n s e m e c h a n i s m s ( F a u l k n e r and G h i -
795
0098-0331/86/0300-0795505.00/0 �9 1986 Plenum Publishing Corporation

796 CARTE AND FAULKNER
selin, 1983). Sea hares and dorid nudibranchs employ defense mechanisms based on dietary chemicals (Thompson et al., 1982; Schulte and Scheuer, 1982) that are effective in deterring most potential predators. However, these chemical defense mechanisms can be penetrated by specialist predators such as the ce- phalaspidean Navanax inermis (Paine, 1963) and the nudibranch Roboastra ti- gris (Farmer, 1978). The added level of complexity in the association between a carnivorous nudibranch and the nudibranchs it eats led us to investigate the role of secondary metabolites in this relationship.
Roboastra tigris is a large carnivorous nembrothid nudibranch that is known to prey upon two smaller nembrothid nudibranchs, Tambja abdere and Tambja eliora. In their attempts to avoid predation by R. tigris, the two species of Tambja employ very different escape responses: T. eliora swims away by un- dulating the body from side to side while T. abdere secretes copious amounts of a yellow mucus from numerous goblet cells found in the skin (Farmer, 1978).
Chemical studies of R. tigris, T. abdere, and T. eliora revealed that all three nudibranchs contained a similar array of pharmacologically active metab- olites that exhibited antimicrobial properties and inhibited cell division in the fertilized sea urchin egg assay (Cart6 and Faulkner, 1983). The metabolites were identified as the tambjamines A-D (I-IV) and the aldehydes V-VII (Fig- ure 1). The aldehydes V-VII were subsequently shown to be artifacts produced by hydrolysis during the extraction procedure. By comparing the antimicrobial spectra of crude extracts, it became apparent that both species of Tambja ob- tained the tambjamines (I-IV) from a single dietary source, the bryozoan Ses- sibugula translucens. Field observations confirmed these associations. The quantitative distribution of the tambjamines (I-IV) and aldehydes V-VII in the four organisms is recorded in Table 1 (from Cart6 and Faulkner, 1983). These data establish that the bryozoan S. translucens is a principal component of the diets of both species of Tambja that are in turn preyed upon by R. tigris.
These observations raised several questions. What is the role of the tamb- jamines in the close association among the bryozoan and the three nudibranchs? How do the Tambja species locate their preferred food source? Why does T. abdere use an apparent chemical defense against predation by R. tigris while T. eliora attempts to swim away? Can R. tigris detect its prey by chemorecep-
H ~0".= ~ Me ~ O M e
X H I~IHR X~'~ N f " N / " C H O
I X = H Y = H R=H V X = H Y = H II X = B r Y = H R=H VI X = B r Y = H
Ili X = H Y = H R=~;-Bu VII X= H Y = B r IV X = H Y = B r R=~;-Bu
Fie. 1 The structures of tambjamines A (I), B(II), C (III), and D (IV); and their hy- drolysis products V (from I and III), VI (from II), and VII (from IV).

NUDIBRANCH FEEDING ASSOCIATIONS 797
TABLE 1. DISTRIBUTION OF TAMBJAMINES A - D (I-IV) AND ALDEHYDES V-VII IN
Roboastra tigris, Tambja abdere, Tambja eliora, AND Sessibugula translucens EXPRESSED AS WEIGHT/ANIMAL OR % DRY WEIGHT
R. tigris T. abdere T. eliora S. translucens
Tambjamine A (I) 0.56 nag 2.22 mg 0.48 mg 0.06% Tambjamine B (II) 0.34 mg 0.94 mg 0.25 mg 0.083% Tanabjamine C (III) 0.54 mg 1.78 mg 0.22 mg 0.023% Tanabjamine D (IV) 0.36 mg 2.67 mg 0.51 mg 0.046% Aldehyde V 0.43 mg 0.61 mg 0.23 mg 0.023% Aldehyde VI 0.14 mg 0.5 mg 0.10 mg 0.071% Aldehyde VII 0.25 mg 1.39 mg 0.17 mg 0.142%
Total wt/animal 2.66 nag 10.11 mg 1.96 mg Average wt 688 mg 296 mg 91 mg Percent dry wt 0.39 % 3.42 % 2.15 % 0.45 %
tion? In this paper we report both field observations and laboratory experiments that establish the chemical relationships between the four organisms.
METHODS AND MATERIALS
Major collections of specimens were made in several locations in the Gulf of California: Puerto Escondido, Baja California (May 1980)--T. abdere and T. eliora; Bahia de los Angeles, Baja California (May 1980 and April 1982)-- R. tigris, T. abdere, T. eliora and S. translucens (1982 only); and Isla Partida (April 1982)--T. eliora and S. translucens. The animals were observed and collected at 2-6 m depth. Live specimens of all four organisms were collected at Bahia de los Angeles and transported to La Jolla in aerated seawater. All organisms were maintained in flowing seawater aquaria. R. tigris was main- tained on a diet of T. eliora, the two Tambja species on a diet of S. translucens, and the bryozoan was fed daily with unicellular algae (Isochrysis galbana, An- acystis marina, Rhodomonas lens, and Monochrysis lutheri, all of which were cultured in GPM media under standard conditions).
Samples of tambjamines A-D (I-IV) were isolated and purified using the procedures described previously (Cart6 and Faulkner, 1983). In some experi- ments, mixtures of compounds were employed. The mixtures were obtained by omitting the final LC separation.
Feeding Inhibition Assays. Feeding inhibition was assayed by observing the feeding response of the spotted kelpfish Gibbonsia elegans toward food pellets ("San Francisco Bay Brand" freeze-dried euphausids) treated with pure compounds at concentrations ranging from 1 to 50 #g/rag. The kelpfish, col- lected intertidally at La Jolla, were kept in aquaria and were trained to accept

798 CARTE AND FAULKNER
food from a glass tube. The test compounds were applied to the pellets (N = 10) in redistilled dichloromethane solutions, and the solvent was removed at 25~ under vacuum. Control pellets were treated with solvent only. Fish were fed the pellets in a random order and the response (eaten vs. rejected) recorded. The probability of obtaining the observed results was calculated using a chi- square distribution to determine the level at which a significant feeding avoid- ance was observed (P < 0.5 level). The following results were obtained: 3 : 2 mixture of tambjamine A (I) and tambjamine B (II); 10/~g/mg--7 of 10 pellets rejected, control--9 of 10 pellets eaten (X 2 = 7.50, P < 0.01); 5/zg/mg--3 of 10 pellets rejected, control--9 of 10 pellets eaten (X 2 = 1.25, P < 0.5). In a 3 :7 mixture of tambjamine C (III) and tambjamine D (IV); 5 tzg/mg--7 of 10 pellets rejected, control--9 of 10 pellets eaten (X 2 = 7.50, P < 0.01); 1 /~g/ mg--2 of 10 pellets rejected, control--9 of 10 pellets eaten (x 2 = 0.39, P < 0.95).
Analysis of Tambjamines A-D (I-IV) in Exudate and Slime Trail of T. abdere. A specimen of R. tigris was allowed to attack an average-sized speci- men of T. abdere in a dish containing "Instant Ocean" synthetic seawater (100 ml). After the specimen had exuded copious quantities of a yellow mucus from glands on the dorsal surface, the animals were separated and removed. The dish and its contents were extracted with dichloromethane (3 • 75 mt), the com- bined extracts dried over anhydrous sodium sulfate and the solvent removed to obtain a green oil (4.8 rag).
Two specimens of T. abdere were allowed to crawl over a bed of aquarium dolomite that had previously been washed sequentially with water, dichloro- methane, water, and synthetic seawater. The trails were marked with colored dolomite, the animals were carefully removed, and the dolomite on which the trails were laid was removed with a " lab scoop" spatula. The dolomite was washed with dichloromethane (3 x 200 ml), the combined extracts dried over anhydrous sodium sulfate, and the solvent evaporated to obtain an organic res- idue (3.1 mg).
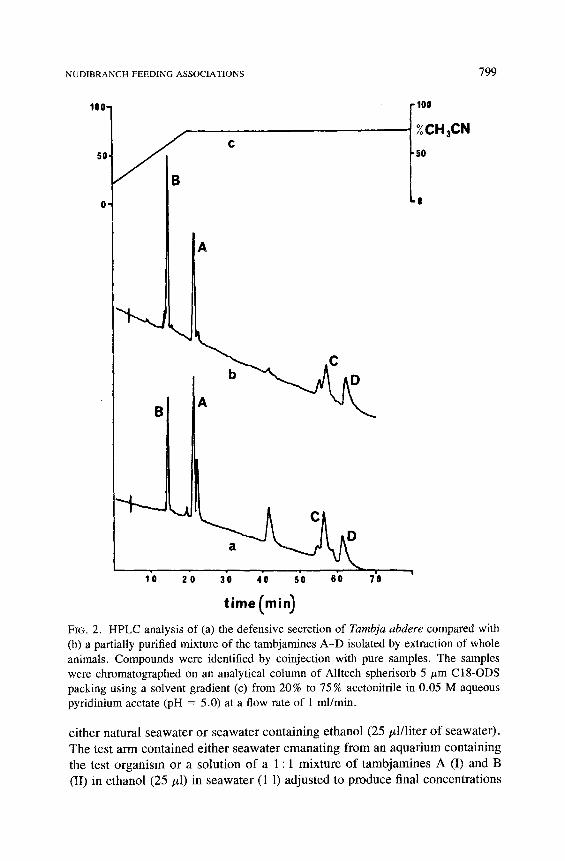
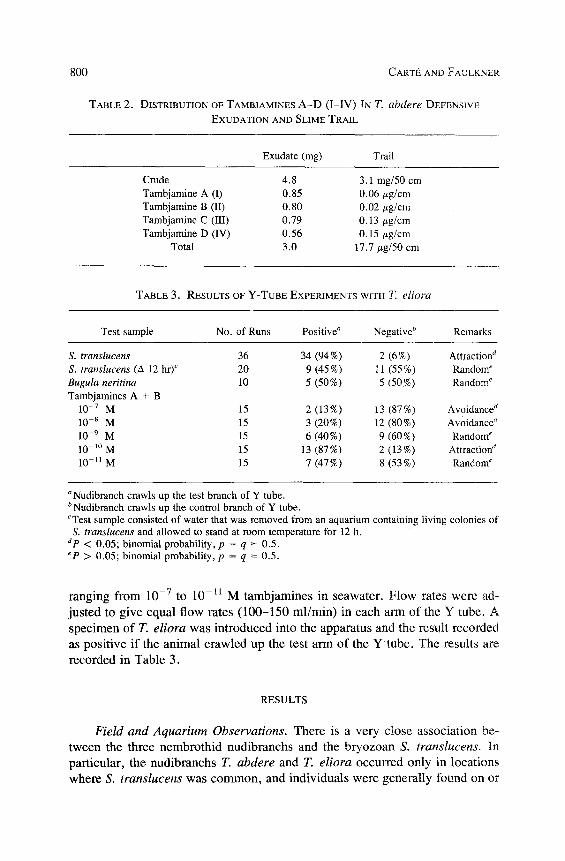
The concentrations of the tambjamines A-D (I-IV) were determined by analytical LC. Separation of the tambjamines (I-IV) was accomplished by LC on an Alltech spherisorb 5-/~m C18-ODS analytical column using a linear sol- vent gradient from 20% to 75 % acetonitrile in 0.05 M pyridinium acetate buffer (pH 5.0) (see Figure 2). The tambjamines were detected by UV absorption at 400 nm. Standard response curves of concentrations vs. peak area (height • width at half height) for each pure compound were used to calculate concentra- tions of the tambjamines in the exudate and the slime trail (see Table 2).
Y-Tube Experiments. A Y-tube apparatus was constructed from clear Plex- iglas tubing (3.75 cm ID) and " a g e d " in seawater. Prior to each experiment, the Y tube was thoroughly washed in natural seawater using a bottle brush to remove any attached materials. A coin flip was used to determine which arm of the Y tube would be the control arm. The control arm contained a stream of
NUDIBRANCH FEEDING ASSOCIATIONS 799
100-
50'
0"
J
B
A
.lOO
~CH3CN
'50
.0
1"0 2 0 3 0 4"0 5"0 6"0 7'0
timeCmin ) FIG. 2. HPLC analysis of (a) the defensive secretion of Tambja abdere compared with (b) a partially purified mixture of the tambjamines A-D isolated by extraction of whole animals. Compounds were identified by coinjection with pure samples. The samples were chromatographed on an analytical column of Alltech spherisorb 5 #m CI8-ODS packing using a solvent gradient (c) from 20% to 75% acetonitrile in 0.05 M aqueous pyridinium acetate (pH = 5.0) at a flow rate of 1 ml/min.
either natural seawater or seawater containing ethanol (25 #l / l i ter o f seawater). The test arm contained either seawater emanating from an aquarium containing the test organism or a solution of a 1 : 1 mixture of tambjamines A (I) and B (II) in ethanol (25 t~l) in seawater (1 1) adjusted to produce final concentrations
800
TABLE 2.
CARTI~ AND FAULKNER
DISTRIBUTION OF TAMBJAMINES A - D (I-IV) IN T. abdere DEFENSIVE EXUDATION AND SLIME TRAIL
Exudate (ms) Trail
Crude 4,8 3.1 mg/50 cm Tambjamine A (I) 0.85 0.06 tzg/cm Tambjamine B (II) 0.80 0.02 tzg/cm Tambjamine C (III) 0.79 0.13/zg/cm Tambjamine D (IV) 0.56 0.15 #g/cm
Total 3.0 17.7 #g/50 cm
TABLE 3. RESULTS OF Y-TUBE EXPERIMENTS WITH T. eliora
Test sample No. of Runs Positive a Negative b Remarks
S. translucens 36 34 (94%) 2 (6%) Attraction d S. translucens (A 12 hr) c 20 9 (45%) 11 (55%) Random ~ Bugula neritina 10 5 (50%) 5 (50%) Random e Tambjamines A + B
10 -7 M 15 2 (13%) 13 (87%) Avoidance d 10 -8 M 15 3 (20%) 12 (80%) Avoidance d 10 9 M 15 6 (40%) 9 (60%) Random e 10 -1~ M 15 13 (87%) 2 (13%) Attraction d 10 -I1 M 15 7 (47%) 8 (53%) Random ~
aNudibranch crawls up the test branch of Y tube. bNudibranch crawls up the control branch of Y tube. CTest sample consisted of water that was removed from an aquarium containing living colonies of S. translucens and allowed to stand at room temperature for 12 h.
dp < 0.05; binomial probability, p = q = 0.5. ~P > 0.05; binomial probability, p = q = 0.5.
ranging f rom 10 -7 to 10 -11 M t a m b j a m i n e s in seawater . F low rates were ad-
jus t ed to g ive equal f low rates ( 100 -150 ml /min ) in each arm o f the Y tube. A
s p e c i m e n o f T. eIiora was in t roduced into the appara tus and the result r ecorded
as pos i t ive i f the an imal c r awled up the tes t a rm o f the Y tube. The results are
r ecorded in Table 3.
RESULTS
Field and Aquarium Observations. There is a very c lose assoc ia t ion be-
t w e e n the th ree n e m b r o t h i d nud ib ranchs and the b r y o z o a n S. translucens. In
part icular , the nud ib ranchs T. abdere and T. eliora occur red only in locat ions
w h e r e S. translucens was c o m m o n , and indiv iduals w e r e genera l ly found on or

NUDIBRANCH FEEDING ASSOCIATIONS 801
near the bryozoan. Although Roboastra tigris was known to feed on a variety of smaller nudibranchs (J.R. Lance, personal communication), it prefers to prey on T. abdere and T. eliora and was most commonly found in areas where the Tambja nudibranchs, and therefore the bryozoan S. translucens, also occurred. However, a few specimens of R. tigris were encountered in habitats seemingly devoid of either Tambja species.
Field observations of the feeding habits of R. tigris suggested that the nu- dibranch could easily detect and follow fresh slime trails of T. abdere, T. eliora, and several other nudibranchs. The trail-following behavior is similar to that described by Paine (1963) for the Cephalaspidean opisthobranch Navanax iner- mis. To follow a slime trail, R. tigris "fee ls" for the trail with external, anterior located cephalic tentacles while sweeping its head from side to side. If the trail is broken, R. tigris will usually tum around and follow the trail in the reverse direction. It appears that R. tigris relies on direct contact between the cephalic tentacles and the slime trail and is not capable of detecting the resumption of the trail by distant chemoreception.
Once the prey has been located, R. tigris rapidly everts a blue buccal hood that engulfs the prey. In attempting to avoid predation by R. tigris, the two Tambja species use different escape strategies. Tambja eliora attempts to escape by writhing vigorously, while T. abdere excretes an apparently distasteful yel- low mucus (Farmer, 1978). In aquarium situations, T. abdere was more suc- cessful in repelling attacks of R. tigris, but in field situations, where currents and wave motion were present, the swimming motion of T. eliora seemed equally effective because the slime trail is thereby broken.
Chemical Studies. We had previously found that specimens of T. abdere contained an average of approximately 10 mg of a mixture of tambjamines while specimens of T. eliora contained less than 2 mg of the same mixture of tamb- jamines (Cart6 and Faulkner, 1983). In order to demonstrate their deterrent value, mixtures of the tambjamines were assayed for feeding inhibition against the spotted kelpfish Gibbonsia elegans and were found to be effective at con- centrations of 5-10/zg/mg pellet for a 3 : 2 mixture of tambjamines A and B and at 1-5 #g/mg pellet for a 3 : 7 mixture of tambjamines C and D. The al- dehydes V-VII, which are formed by hydrolysis of the tambjamines, are not effective feeding inhibitors at concentrations less than 20 #g/rag. These data indicate that the tambjamines are relatively effective fish feeding inhibitors (cf., Thompson et al., 1982) and might be considered to provide protection against predators other than Roboastra. Since the differences in the escape responses of T. eliora and T. abdere might be an outcome of the different quantities of the tambjamines in the two animals, it was important to determine the quantity of tambjamines in the defensive secretion of T. abdere. L C analysis of the exudate from an average-sized specimen of T. abdere revealed that the defen- sive secretion contained a total of 3 mg of a mixture of tambjamines A-D (I- IV) (Table 2).

802 CARTI~ AND FAULKNER
Since R. tigris had been observed to hunt Tambja species in preference to other nudibranchs, we suspected that the slime trails of Tambja might contain distinctive chemicals. A 50-cm length of the dolomite on which the trail of T. abdere had been secreted was extracted with dichloromethane to obtain a total of 18/zg of the tambjamines A-D (I-IV) (Table 2).
Although our field observations suggested that R. tigris could not detect the tambjamines by distant chemoreception, there was a good reason to believe that the Tambja species could find their food source, Sessibugula translucens, by using chemical detection. To test this hypothesis, a number of Y-maze ex- periments, modeled on those of Davenport and Hickock (1951), were per- formed using T. eliora as the test animal. T. eliora were clearly attracted by water that had been passed over S. translucens, but they were unaffected by water passing over Bugula neritina, a species of bryozoan that is common in areas populated by Tambja and that appears to be eaten only when S. translu- cens is unavailable. Water removed from an aquarium containing S. translucens and allowed to stand for 12 hr no longer elicited a positive response, indicating that the chemical responsible for attraction was not stable in seawater. These results suggested that the tambjamines (I-IV), which are easily hydrolyzed to the aldehydes V-VII, might be responsible for the observed chemotactic re- sponse of T. eliora. A 1 : 1 mixture of tambjamines A (I) and B (II) was tested at several concentrations from 10 -7 M to 10-11 M. At very low concentrations, the tambjamines elicited no response, but the nudibranchs were attracted to a 10-10 M solution of tambjamines A and B in seawater. At concentrations greater than 10 -9 M, T. eliora was repelled by the test solution (see Table 3).
DISCUSSION
Our previous chemical studies had established the presence of the tamb- jamines in the bryozoan Sessibugula translucens and the three nembrothid nu- dibranchs Tambja abdere, Tambja eliora, and Roboastra tigris (Cart6 and Faulkner, 1983). These results, together with our random field observations and those of Farmer (1978) and Lance (personal communication), clearly estab- lished that T. abdere and T. eliora favored the bryozoan S. translucens as a food source and were, in turn, eaten by R. tigris. The Y-maze experiments demonstrate that the Tambja species can locate their preferred food source by detection of low concentrations of the tambjamines that are produced by the bryozoan S. translucens. While it is not firmly established that R. tigris can distinguish the slime trails of Tambja species from those of other species by detecting tambjamines, their presence in low concentrations in the slime trail of T. abdere certainly suggests that this is the case. It would be difficult to test this hypothesis because we would require a synthetic slime trail sufficiently similar to the natural material that it would be followed by Roboastra.

NUDIBRANCH FEEDING ASSOCIATIONS 803
The exudate of T. abdere contains a very large quantity of the mixture of tambjamines, representing approximately 30% of the metabolites stored by a typical animal. The exudate is clearly distasteful enough to deter most potential predators and can sometimes repel the specialist predator R. tigris (Farmer, 1978). However, the use of such a great proportion of the stored tambjamines to repel a single attack must render the animal susceptible to repeated attacks if it cannot rapidly replace the active ingredients of its defensive secretion. Since the entire tambjamine content of an average specimen of T. eliora is less than that used by T. abdere during a single, sometimes unsuccessful encounter with R. tigris, it is not surprising that T. eliora has adopted an escape strategy that is not dependent on a chemical feeding inhibitor.
The Y-maze experiments demonstrated that T. eliora avoided a 10 -8 M solution of a 1 : 1 mixture of tambjamines A and B in seawater but were attracted by a 10-~o M solution of the same compounds. The ability to detect a two order of magnitude difference in the concentration of tambjamines in seawater may assist T. eliora in avoiding predation. While it is unlikely that R. tigris exudes a sufficient quantity of the tambjamines to be detected at a distance, an encoun- ter between Roboastra and a Tambja species might result in the localized release of tambjamines, thus warning other potential prey of the presence of a predator, similar to a situation found among some fishes (von Frisch, 1941) and for the marine mud snail Nassarius obsoletus (Atema and Stenzler, 1977). Concentra- tion-dependent properties have also been observed for ant trail pheromones (Tumlinson et al., 1971).
The ease with which the tambjamines (I-IV) are hydrolyzed to obtain the less active aldehydes V-VII provides a natural mechanism to remove old trails and exudations that might otherwise provide false information. This is consid- ered an important factor in maintaining the efficacy of a chemical messenger.
Acknowledgments--We thank James R. Lance for providing information on the distribution and behavior of the nudibranchs. The feeding inhibition assay was devised by Joseph R. Pawlik who has also critically reviewed this manuscript. The research was supported by a grant from the National Science Foundation (CHE81-21471).
REFERENCES
ATEMA, J., and STENZLER, D. 1977. Alarm substance of the marine and mud snail, Nassarius obsoletus: Biological characterization and possible evolution. J. Chem. Ecol. 3:173-187.
CARTI~, B., and FAULKNER, D.J. 1983. Defensive metabolites from three nembrothid nudibranchs. J. Org. Chem. 48:2314-2318.
DAVENPORT, D. 1950. Studies in the physiology of commensalism. 1. The polynoid genus Arc- tono~. Biol. Bull. 98:81-93.
DAVENPORT, D., and HICKOK, J.R. 1951. Studies in the physiology of commensalism. 2. The polynoid genera Arctono~ and Halosydna. Biol. Bull. 100:71-83.

804 CARTr~ AND FAULKNER
FARMER, W.M. 1978. Tambja and Roboastra (Mollusca: Opisthobranchia) from the Gulf of Cal- ifornia and Galapagos Islands. Veliger 20:375-385.
FAULKNER, D.J., and GHISEL1N, M.T. 1983. Chemical defense and evolutionary ecology of dorid nudibranchs and some other opisthobranch gastropods. Mar. Ecol. Prog. Ser. t3:295-301.
PAINE, R.T. 1963. Food recognition and predation on opisthobranchs by Navanax inerrnis (Gas- tropoda: Opisthobranchia). Veliger 6:1-9.
SCHtJLTE, G.R., and SCHEUER, P.J. 1982. Defense allomones of some marine mollusks. Tetrahe- dron 38:1857-1863.
THOMPSON, J.E., WALKER, R.P., WRATTEN, S.J., and FAULKNER, D.J. 1982. A chemical defense mechanism for the nudibranch Cadlina luteomarginata. Tetrahedron 38:1865-1873.
TUMLINSON, J.H., SILVERSTEIN, R.M., MOSER, J.C., BROWNLEE, R.G., and RUTH, J.M. 1971. Identification of the trail pheromone of a leaf-cutting ant, Atta texana. Nature 234:348-349.
Related Documents











