INFECTION AND IMMUNITY, Mar. 2006, p. 1621–1630 Vol. 74, No. 3 0019-9567/06/$08.000 doi:10.1128/IAI.74.3.1621–1630.2006 Copyright © 2006, American Society of Mirobiology. All Rights Reserved. Role of Phagosomes and Major Histocompatibility Complex Class II (MHC-II) Compartment in MHC-II Antigen Processing of Mycobacterium tuberculosis in Human Macrophages Martha Torres, 1,5 † Lakshmi Ramachandra, 2,4 †* Roxana E. Rojas, 1 Karen Bobadilla, 5 Jeremy Thomas, 1 David H. Canaday, 1 Clifford V. Harding, 3 ‡ and W. Henry Boom 1 ‡ Division of Infectious Disease, 1 Department of Pediatrics, 2 and Department of Pathology, 3 Case Western Reserve University, Cleveland, Ohio; Rainbow Babies and Children’s Hospital, Cleveland, Ohio 4 ; and Departamento de Microbiologia, Instituto Nacional de Enfermedades Respiratorias, Mexico City, Mexico 5 Received 7 September 2005/Returned for modification 13 October 2005/Accepted 7 December 2005 Mycobacterium tuberculosis resides in phagosomes inside macrophages. In this study, we analyzed the kinetics and location of M. tuberculosis peptide-major histocompatibility complex class II (MHC-II) complexes in M. tuberculosis-infected human macrophages. M. tuberculosis peptide–MHC-II complexes were detected with poly- clonal autologous M. tuberculosis-specific CD4 T cells or F9A6 T hybridoma cells specific for M. tuberculosis antigen (Ag) 85B (96-111). Macrophages processed heat-killed M. tuberculosis more rapidly and efficiently than live M. tuberculosis. To determine where M. tuberculosis peptide-MHC-II complexes were formed intracellularly, macrophages incubated with heat-killed M. tuberculosis were homogenized, and subcellular compartments were separated on Percoll density gradients analyzed with T cells. In THP-1 cells, M. tuberculosis Ag 85B (96- 111)–DR1 complexes appeared initially in phagosomes, followed by MHC class II compartment (MIIC) and the plasma membrane fractions. In monocyte-derived macrophages, M. tuberculosis peptide–MHC-II complexes appeared only in MIIC fractions and subsequently on the plasma membrane. Although phagosomes from both cell types acquired lysosome-associated membrane protein 1 (LAMP-1) and MHC-II, THP-1 phagosomes that support formation of M. tuberculosis peptide–MHC-II complexes had increased levels of both LAMP-1 and MHC-II. Thus, M. tuberculosis phagosomes with high levels of MHC-II and LAMP-1 and MIIC both have the potential to form peptide–MHC-II complexes from M. tuberculosis antigens in human macrophages. Mycobacterium tuberculosis is an intracellular pathogen that is phagocytosed by professional phagocytes such as macro- phages and dendritic cells. A variety of receptors promote M. tuberculosis phagocytosis and these include among others the complement receptors CR1, CR3, and CR4, the mannose re- ceptor, and the scavenger receptor class A (14, 33, 42). Fol- lowing phagocytosis, M. tuberculosis resides inside plasma membrane-derived phagosomal vesicles. While the majority of bacterial phagosomes undergo a process of acidification and maturation, M. tuberculosis inhibits this process (16, 37). CD4 T cells are critical for control of M. tuberculosis in- fection in animals and humans (5, 10) and are activated when mycobacterial antigens are processed and presented by major histocompatibility complex class II (MHC-II) molecules. Pro- cessing of soluble antigens has been extensively studied. Newly synthesized MHC-II molecules bind invariant chain in the endoplasmic reticulum and target to endocytic compartments that function in antigen processing, e.g., the MHC class II compartment (MIIC) (1, 12, 22, 23, 30, 36, 39). Here the invariant chain is degraded, leaving only the class II-associated invariant chain peptide (CLIP) bound to MHC-II (8). CLIP is then replaced with an antigenic peptide generated from the degradation of the soluble antigen. The resulting peptide– MHC-II complexes are transported to the cell surface for pre- sentation to CD4 T cells. Processing of particulate antigens by the MHC-II pathway and the specific role of phagosomes in this process are still being defined. Latex bead phagosomes were shown to contain MHC-II, degrade phagosome-associated antigen, and directly mediate peptide–MHC-II complex formation (24, 26, 28, 29). More recently, Ramachandra et al. demonstrated that M. tu- berculosis phagosomes derived from M. tuberculosis-infected murine bone marrow macrophages could also mediate the formation of peptide–MHC-II complexes (27). There is also evidence suggesting that endocytic compartments play a role in the processing of certain bacteria. Following internalization of Streptococcus pyogenes, endocytic compartments mediate the formation of peptide–MHC-II complexes after receiving phago- some-derived antigen fragments (9). In the current studies, we have analyzed the intracellular compartments involved in the generation of M. tuberculosis peptide–MHC-II complexes in M. tuberculosis-pulsed human macrophages. To analyze processing of M. tuberculosis antigens in the human macrophage cell line THP-1 and in human monocyte- derived macrophages (MDMs), we have used F9A6 murine T hybridoma cells, specific for the antigen (Ag) 85B (96-111) epitope presented by HLA-DR1 or polyclonal autologous M. tuberculosis-specific CD4 T cells. Ag 85B, a mycolyltrans- ferase involved in cell wall biosynthesis (4), is a major target of human T-cell responses to M. tuberculosis and a leading vac- * Corresponding author. Mailing address: Division of Infectious Diseases, Dept. of Pediatrics, Rainbow Babies and Children’s Hospi- tal, Rm. 4007, 2101 Adelbert Rd., Cleveland, OH 44106-6008. Phone: (216) 844-5873. Fax: (216) 844-8362. E-mail: [email protected]. † These two authors (Martha Torres and Lakshmi Ramachandra) contributed equally to this work. ‡ These two authors share senior authorship. 1621

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

INFECTION AND IMMUNITY, Mar. 2006, p. 1621–1630 Vol. 74, No. 30019-9567/06/$08.00�0 doi:10.1128/IAI.74.3.1621–1630.2006Copyright © 2006, American Society of Mirobiology. All Rights Reserved.
Role of Phagosomes and Major Histocompatibility Complex Class II(MHC-II) Compartment in MHC-II Antigen Processing of
Mycobacterium tuberculosis in Human MacrophagesMartha Torres,1,5† Lakshmi Ramachandra,2,4†* Roxana E. Rojas,1 Karen Bobadilla,5 Jeremy Thomas,1
David H. Canaday,1 Clifford V. Harding,3‡ and W. Henry Boom1‡Division of Infectious Disease,1 Department of Pediatrics,2 and Department of Pathology,3 Case Western Reserve University,
Cleveland, Ohio; Rainbow Babies and Children’s Hospital, Cleveland, Ohio4; and Departamento de Microbiologia,Instituto Nacional de Enfermedades Respiratorias, Mexico City, Mexico5
Received 7 September 2005/Returned for modification 13 October 2005/Accepted 7 December 2005
Mycobacterium tuberculosis resides in phagosomes inside macrophages. In this study, we analyzed the kineticsand location of M. tuberculosis peptide-major histocompatibility complex class II (MHC-II) complexes in M.tuberculosis-infected human macrophages. M. tuberculosis peptide–MHC-II complexes were detected with poly-clonal autologous M. tuberculosis-specific CD4� T cells or F9A6 T hybridoma cells specific for M. tuberculosisantigen (Ag) 85B (96-111). Macrophages processed heat-killed M. tuberculosis more rapidly and efficiently thanlive M. tuberculosis. To determine where M. tuberculosis peptide-MHC-II complexes were formed intracellularly,macrophages incubated with heat-killed M. tuberculosis were homogenized, and subcellular compartments wereseparated on Percoll density gradients analyzed with T cells. In THP-1 cells, M. tuberculosis Ag 85B (96-111)–DR1 complexes appeared initially in phagosomes, followed by MHC class II compartment (MIIC) and theplasma membrane fractions. In monocyte-derived macrophages, M. tuberculosis peptide–MHC-II complexesappeared only in MIIC fractions and subsequently on the plasma membrane. Although phagosomes from bothcell types acquired lysosome-associated membrane protein 1 (LAMP-1) and MHC-II, THP-1 phagosomes thatsupport formation of M. tuberculosis peptide–MHC-II complexes had increased levels of both LAMP-1 andMHC-II. Thus, M. tuberculosis phagosomes with high levels of MHC-II and LAMP-1 and MIIC both have thepotential to form peptide–MHC-II complexes from M. tuberculosis antigens in human macrophages.
Mycobacterium tuberculosis is an intracellular pathogen thatis phagocytosed by professional phagocytes such as macro-phages and dendritic cells. A variety of receptors promote M.tuberculosis phagocytosis and these include among others thecomplement receptors CR1, CR3, and CR4, the mannose re-ceptor, and the scavenger receptor class A (14, 33, 42). Fol-lowing phagocytosis, M. tuberculosis resides inside plasmamembrane-derived phagosomal vesicles. While the majority ofbacterial phagosomes undergo a process of acidification andmaturation, M. tuberculosis inhibits this process (16, 37).
CD4� T cells are critical for control of M. tuberculosis in-fection in animals and humans (5, 10) and are activated whenmycobacterial antigens are processed and presented by majorhistocompatibility complex class II (MHC-II) molecules. Pro-cessing of soluble antigens has been extensively studied. Newlysynthesized MHC-II molecules bind invariant chain in theendoplasmic reticulum and target to endocytic compartmentsthat function in antigen processing, e.g., the MHC class IIcompartment (MIIC) (1, 12, 22, 23, 30, 36, 39). Here theinvariant chain is degraded, leaving only the class II-associatedinvariant chain peptide (CLIP) bound to MHC-II (8). CLIP is
then replaced with an antigenic peptide generated from thedegradation of the soluble antigen. The resulting peptide–MHC-II complexes are transported to the cell surface for pre-sentation to CD4� T cells.
Processing of particulate antigens by the MHC-II pathwayand the specific role of phagosomes in this process are stillbeing defined. Latex bead phagosomes were shown to containMHC-II, degrade phagosome-associated antigen, and directlymediate peptide–MHC-II complex formation (24, 26, 28, 29).More recently, Ramachandra et al. demonstrated that M. tu-berculosis phagosomes derived from M. tuberculosis-infectedmurine bone marrow macrophages could also mediate theformation of peptide–MHC-II complexes (27). There is alsoevidence suggesting that endocytic compartments play a role inthe processing of certain bacteria. Following internalization ofStreptococcus pyogenes, endocytic compartments mediate theformation of peptide–MHC-II complexes after receiving phago-some-derived antigen fragments (9). In the current studies, wehave analyzed the intracellular compartments involved in thegeneration of M. tuberculosis peptide–MHC-II complexes in M.tuberculosis-pulsed human macrophages.
To analyze processing of M. tuberculosis antigens in thehuman macrophage cell line THP-1 and in human monocyte-derived macrophages (MDMs), we have used F9A6 murine Thybridoma cells, specific for the antigen (Ag) 85B (96-111)epitope presented by HLA-DR1 or polyclonal autologous M.tuberculosis-specific CD4� T cells. Ag 85B, a mycolyltrans-ferase involved in cell wall biosynthesis (4), is a major target ofhuman T-cell responses to M. tuberculosis and a leading vac-
* Corresponding author. Mailing address: Division of InfectiousDiseases, Dept. of Pediatrics, Rainbow Babies and Children’s Hospi-tal, Rm. 4007, 2101 Adelbert Rd., Cleveland, OH 44106-6008. Phone:(216) 844-5873. Fax: (216) 844-8362. E-mail: [email protected].
† These two authors (Martha Torres and Lakshmi Ramachandra)contributed equally to this work.
‡ These two authors share senior authorship.
1621

cine candidate (6, 15, 19, 40). The F9A6 T hybridoma cellsallowed study of the processing of a defined antigen (Ag 85B),while polyclonal autologous M. tuberculosis-specific CD4� Tcells allowed analysis of processing of mycobacterial antigensin general. Processing of heat-killed M. tuberculosis by THP-1cells and MDMs for presentation to T cells was more rapidthan processing of live M. tuberculosis.
Subcellular fractionation coupled with T-cell assays demon-strated that phagosomes mediated the formation of M. tuber-culosis Ag 85B (96-111)–DR1 complexes in THP-1 cells, but inMDMs M. tuberculosis peptide–MHC-II complexes wereformed in MIIC but not phagosomes. Analysis of M. tuber-culosis phagosomes by flow organellometry demonstrated thatphagosomes derived from both THP-1 cells and MDMs ac-quired lysosome-associated membrane protein 1 (LAMP-1)and MHC-II, but THP-1-derived phagosomes had increasedlevels of both LAMP-1 and MHC-II. This suggests that MDMphagosomes may not be able to support formation of M. tu-berculosis peptide-MHC complexes due to its low MHC-IIlevels and altered phenotype. In conclusion, M. tuberculosispeptide-MHC-II complexes can be formed in both phago-somes and MIIC in human macrophages.
MATERIALS AND METHODS
Bacteria. M. tuberculosis H37Ra was obtained from the American Type Cul-ture Collection (ATCC 25117; Manassas, VA) and grown to log phase in Middle-brook 7H9 broth (Difco, Detroit, MI) supplemented with 1% glycerol and 10%Middlebrook albumin dextrose catalase enrichment (Difco). Bacteria were har-vested and frozen at �80°C in RPMI 1640, 10% fetal bovine serum, and 6%glycerol. Bacterial concentration was determined by counting CFU on Middle-brook 7H10 agar plates (Difco). Heat-killed M. tuberculosis was prepared byincubating M. tuberculosis at 80°C for 30 min (killing was confirmed by CFUcounting). Prior to infection of cells, all M. tuberculosis preparations were pel-leted, washed, and declumped by 10 passages through a 26-gauge needle fol-lowed by sonication for 40 seconds at 80 cycles in a sonicator (Ultrasonic Pro-cessor) and centrifugation at 800 rpm in an Eppendorf centrifuge for 2 min toremove clumps. To label live or heat-killed M. tuberculosis with fluorescein, 109
bacteria were pelleted, resuspended in 1 ml phosphate-buffered saline (pH 9.1)and mixed with 25 �l of 20 mg/ml FLUOS (Boehringer, Mannheim, Germany)in dimethyl sulfoxide for 10 min at room temperature. Labeled M. tuberculosisbacteria were washed twice and declumped prior to use. M. tuberculosis bacteriawere incubated in medium containing 10% non-heat-treated pooled humanserum with no antibiotics for 30 min at 37°C prior to incubation with cells.
Cells and media. Unless otherwise specified, all cells were cultured at 37°Cin 5% CO2. The human monocytic cell line (THP-1) was obtained from theAmerican Type Culture Collection (TIB 202) and cultured in RPMI 1640(BioWhittaker, Walkersville, MD) supplemented with 10% heat-inactivatedfetal bovine serum (HyClone, Logan, UT), 50 �M 2-mercaptoethanol, 1 mMsodium pyruvate, 2 mM L-glutamine, 10 mM HEPES buffer, nonessentialamino acids, 100 U/ml penicillin, and 100 �g/ml streptomycin (BioWhittaker,Walkersville, MD). Cultures were maintained at a cell concentration between2 � 105 and 106 cells/ml.
To induce differentiation of cells to macrophages, THP-1 cells were activatedin medium supplemented with 10 ng/ml phorbol myristate acetate (Sigma, St.Louis, MO). After 24 h, medium was removed and cells were incubated for anadditional 24 h with new medium containing 50 U/ml recombinant humangamma interferon (IFN-�) (Endogen, Woburn, MA). Activated THP-1 cellswere used in all experiments to upregulate MHC-II levels. T hybridoma cells(F9A6) specific to M. tuberculosis Ag 85B (96-11)–DR1 complexes was gen-erated in HLA-DR1 (subtype 01) transgenic mice as described previously (7,11). The T-cell hybridoma F9A6 was maintained in Dulbecco’s modifiedEagle’s medium (BioWhittaker) supplemented as described above for THP-1cells.
MDMs were obtained from healthy adult donors. For certain experimentsHLA-DR1 and HLA-DR2 donors were selected. Heparinized blood was diluted1:1 with RPMI and centrifuged at 300 � g for 45 min at room temperature overa Ficoll-sodium diatrizoate solution (Ficoll-Paque: Pharmacia Fine Chemicals,
Inc.). The layer containing peripheral blood mononuclear cells (PBMCs) washarvested, and monocytes were enriched by plastic adherence for 1 h at 37°C.Adherent cells were purified by immunomagnetic bead separation according tothe manufacturer’s instructions (Miltenyi Biotec, Auburn, CA). The mean purityof positively selected CD14� monocytes was 97% as determined by flow cytom-etry. Monocytes were cultivated in RPMI 1640, 10% pooled human serum at37°C for 7 days. By this time, the cells had acquired macrophage morphology.Recruitment and participation of healthy blood donors in our research wereapproved by the University Hospitals of Cleveland and Instituto Nacional deEnfermedades Respiratorias Institutional Review Board.
Generation of antigen-specific CD4� T cells. Polyclonal autologous CD4� Tcells were obtained from purified protein derivative-positive (PPD�) donors andstimulated as described by Tan et al. (34). PBMCs (2 � 106 cells/well) werestimulated in 24-well plates with live M. tuberculosis at a multiplicity of infectionof 1:10 and 50 U/ml of interleukin-2 (IL-2). The culture was additionally sup-plemented with IL-2 on days 3, 5, and 7. After 7 to 10 days CD4� T cells werepositively selected by immunomagnetic bead separation according to the manu-facturer’s instructions (Miltenyi Biotec). The mean purity of positively selectedCD4� T cells was 96% � 2% as determined by flow cytometry. Cells wereallowed to rest for 24 h before being used in antigen processing assays.
HLA typing of donors. DNA was isolated from PBMCs with the DNeasy kit(QIAGEN) according to the manufacturer’s instructions. For HLA-DR1 andHLA-DR2 screening, Micro SSP HLA typing trays specific for HLA-DR1 orHLA-DR2 (One Lambda, Canoga Park, CA) were used.
Antigen processing and presentation assays. THP-1 cells or human MDMswere cultured in 96-well flat-bottom plates (1.5 � 105 cells/well) in completeRPMI 1640 medium containing 10% pooled human serum. Both THP-1 cells andhuman MDMs were infected with M. tuberculosis at a multiplicity of infectionranging from 1 to 40. Bacteria were pelleted onto cells by centrifugation at900 � g for 10 min at 37°C. Cells were incubated at 37°C for 10 min (providinga total pulse period of 20 min) and washed in ice-cold RPMI to remove extra-cellular bacteria. Prewarmed medium was added, and cells were incubated at37°C for up to 4 h. Cells were fixed with 1% paraformaldehyde and washed.
T hybridoma cells (F9A6; 105) or M. tuberculosis-specific CD4� T cells (105)were added to each well (200 �l total volume). After 24 h, supernatants (100 �l)were harvested. Uptake of heat-killed and live M. tuberculosis by THP-1 cells andMDMs was assessed by identification of acid-fast bacilli with Ziehl-Neelsenstaining of cells. Three hundred cells were analyzed to determine the number ofcells that had phagocytized M. tuberculosis. We also determined phagocytosis byflow cytometry by incubating MDMs and THP-1 cells with FLUOS-labeledheat-killed or live M. tuberculosis as described previously (27).
In experiments with F9A6 T hybridoma cells, IL-2 in the supernatants was mea-sured using an IL-2-dependent CTLL-2 cell line as previously described (17). Pro-liferation of CTLL-2 cells was monitored by addition of Alamar blue (Trek Diag-nostics, Westlake, OH) as an indicator dye and measured as the difference betweenabsorbances at 550 and 595 nm after 24 h. The results presented are the mean oftriplicate wells. In experiments with primary T cells, IFN-� in supernatants wasmeasured by enzyme-linked immunosorbent assay with anti-human IFN-�antibodies (Endogen, Woburn, MA), and absorbance was read on a microplatereader (Labsystems Multiskan MCC/340, Labsystems, Finland) at 450 nm.
Subcellular fractionation of macrophages for biochemical analysis and T-cellassay. THP-1 cells or MDM were replated in six-well plates at 1.5 � 106 cells/well. Four plates were used for each fractionation. Cells were pulsed with me-dium containing heat-killed M. tuberculosis (multiplicity of infection, 40) for 20min as described above, followed by various chase incubations. Cells werewashed, detached by scraping and resuspended in 1 ml of homogenization buffer(0.25 M sucrose and 10 mM HEPES, pH 7.2). Samples were homogenized in aDounce homogenizer (Wheaton) to obtain 40 to 50% lysis. Intact cells and nucleiwere removed by three consecutive spins at 200 � g for 5 min at 4°C. Thesupernatant (containing phagosomes) was collected, layered on 9 ml of 27%Percoll (Amersham Pharmacia Biotech, Uppsala, Sweden) in homogenizationbuffer and centrifuged in a T-865.1 fixed-angle rotor (Sorvall, Newton, CO) at4°C for 60 min at 32,000 � g. Gradients were manually fractionated from the topinto 30 fractions (333 �l each). Each fraction was divided into replicate aliquotsof 10, 30, or 50 �l in 96-well plates and stored frozen at �80°C.
Plasma membrane was labeled before homogenization with sulfo avidin-biotin(Pierce, Rockford, Il). Cells were incubated with 0.5 �g/ml sulfo avidin-biotin at4°C for 30 min, washed and incubated with 10 �g/ml streptavidin-fluoresceinisothiocyanate (BD Bioscience, San Diego, CA) at 4°C for 40 min. Fractions (50�l) were transferred to 96-well, clear-bottom black plates (Costar, Cambridge, MA)and analyzed on a Spectra Fluor Plus fluorimeter (Tecan, United Kingdom). �-Hex-osaminidase activity was measured by colorimetric assay. Fractions (50 �l) wereincubated with 150 �l �-hexosaminidase buffer containing 0.1 M morpho-
1622 TORRES ET AL. INFECT. IMMUN.

lineethanesulfonic acid (MES), 0.2% Triton X-100, pH 6.5, and 50 �l of 1.13mg/ml p-nitrophenyl–acetyl-�-D-glucosaminidase (Sigma-Aldrich). After 90 minat 37°C the reaction was stopped by addition of 0.5 M glycine, pH 10, and theoptical density was determined at 405 nm.
To identify phagosomes containing fluorescent bacteria, fractions (50 �l) fromcells infected with fluorescein-labeled M. tuberculosis were transferred into 96-well clear-bottom black plates (Costar) and analyzed on a Spectra Fluor Plusfluorimeter (Tecan). For T-cell analysis of Percoll gradient fractions, fractionswere frozen and thawed once. Standard medium and F9A6 T hybridoma cells orM. tuberculosis-specific CD4� T cells (105 cells/well) were added to the individualfractions to a final volume of 200 �l. In wells containing polyclonal autologous M.tuberculosis-specific CD4� T cells, 2 �g/ml of anti-human CD28 antibody fromeBioscences (San Diego, CA) was added. After 24 h supernatants were har-vested, and IL-2 or IFN-� production was assessed. Wells containing 27% Percollin homogenization buffer were used as blank controls. For identification of
MIIC, noninfected THP-1 cells were incubated with soluble Ag 85B (5 �g/ml) for1 h prior to fractionation. Fractions were frozen and thawed and then assessedfor the presence of the M. tuberculosis Ag 85B (96-111)–DR1 complexes by theT-cell assay with F9A6 T hybridoma cells.
Preparation of phagosomes for flow organellometry. THP-1 cells or MDMswere plated in six-well plates at 1.5 � 106 cells/well. Four plates were used foreach condition. Cells were pulsed with fluorescein-labeled heat-killed M. tuber-culosis (multiplicity of infection, 40) for 20 min as described above and chased foran additional 30 min. Cells were washed, detached by scraping, and resuspendedin homogenization buffer. Cells were homogenized, and intact cells and nucleiremoved by centrifugation. Phagosomes were pelleted from the supernatant at500 � g at 4°C for 15 min and resuspended in 1 ml homogenization buffer.Resuspended phagosomes were isolated on 27% Percoll density gradients. Frac-tions previously identified to contain phagosomes (see above) were collected andcombined with equal volumes of 2% paraformaldehyde for 10 min. The suspen-
FIG. 1. Processing and presentation of M. tuberculosis by human macrophages: assessment of specificity of F9A6 T-cell hybridoma and kineticsof processing of heat-killed and live M. tuberculosis. Macrophages were pulsed with heat-killed or live M. tuberculosis for 20 min at a multiplicityof infection of 40, chased for various periods at 37°C, and fixed. F9A6 T hybridoma cells or polyclonal autologous M. tuberculosis-specific CD4�
T cells were used to detect M. tuberculosis Ag 85B (96-111)–DR1 or M. tuberculosis peptide–MHC-II complexes, respectively. A. Processing byIFN-�-activated THP-1 cells, HLA-DR1� or HLA-DR2� MDMs with and without heat-killed M. tuberculosis to F9A6 T hybridoma cells forassessment of specificity of the F9A6 T hybridoma cells. B. Kinetics of processing and presentation of heat-killed and live M. tuberculosis byIFN-�-activated THP-1 cells to F9A6 cells. C. Kinetics of processing and presentation of heat-killed and live M. tuberculosis by HLA-DR1� MDMsto F9A6 cells. D. Kinetics of processing and presentation of heat-killed and live M. tuberculosis by MDMs to polyclonal autologous M.tuberculosis-specific CD4� T cells. For experiments using F9A6 T hybridoma cells, supernatants were assessed for IL-2 content using a CTLL-2proliferation assay that was monitored with Alamar blue, an indicator dye. For experiments using polyclonal autologous M. tuberculosis-specificCD4� T cells, supernatants were assessed for IFN-� content by enzyme-linked immunosorbent assay (ELISA). Data points are means of triplicatesamples � standard deviation. Data are representative of four independent experiments.
VOL. 74, 2006 PROCESSING OF M. TUBERCULOSIS BY HUMAN MACROPHAGES 1623

sion was mixed with an equal volume of 0.4 M lysine in phosphate-bufferedsaline, and phagosomes were washed twice by pelleting and then resuspended in0.5 ml phosphate-buffered saline (29). Phagosomes were stained with antibodiesand isotype controls in 96-well plates using a buffer containing saponin to allowaccess to luminal epitopes and analyzed using a Coulter EPICS Elite ESPinstrument (Beckman Coulter, San Diego, CA).
Antibodies and reagents. The following antibodies were used for stainingphagosomes: Cy5-conjugated mouse anti-human LAMP-1 monoclonal antibody,Cy5-conjugated mouse anti-HLA-DR monoclonal antibody, Cy5-conjugatedmouse immunoglobulin G2b isotype control, Cy5-conjugated mouse immuno-globulin G1 isotype control (BD Biosciences). Recombinant Ag 85B of M.tuberculosis was obtained from David Lakey, synthetic peptide 96-111 was ob-tained from Mimotopes Ltd. (Melbourne, Australia), and anti-human CD28antibody was obtained from eBiosciences (San Diego, CA).
RESULTS
Characterization of M. tuberculosis processing and presen-tation. Processing of M. tuberculosis was analyzed in THP-1cells and MDMs using an M. tuberculosis-specific T-cell hybrid-oma, F9A6. The F9A6 T-cell hybridoma was specific for M.tuberculosis Ag 85B (96-111) presented by HLA-DR1 and re-
sponded to THP-1 cells (which express HLA-DR1) andMDMs from HLA-DR1-positive donors following incubationwith heat-killed M. tuberculosis (Fig. 1A). No response wasobserved in the absence of M. tuberculosis or when HLA-DR1� MDMs were incubated with heat-killed M. tuberculosis,confirming the antigen specificity and HLA-DR1 restriction ofthe F9A6 T-cell hybridoma.
To determine the kinetics of processing of M. tuberculosis an-tigens, F9A6 T hybridoma cells and M. tuberculosis-specific CD4�
T cells were used to detect peptide–MHC-II complexes. Whilethe F9A6 T-cell hybridoma allowed analysis of processing of aspecific M. tuberculosis antigen, polyclonal autologous M. tuber-culosis-specific CD4� T cells provided an assessment of a broadspectrum of M. tuberculosis antigens. THP-1 cells and MDMswere pulsed for 20 min with heat-killed M. tuberculosis or live M.tuberculosis at a multiplicity of infection of 40 and chased forvarious lengths of time for up to 4 h (Fig. 1B to D).
Processing of heat-killed M. tuberculosis was initiated rapidlyin both THP-1 cells and MDMs, and M. tuberculosis Ag 85B
FIG. 2. Characterization of subcellular fractions of macrophages isolated on 27% Percoll density gradients. THP-1 cells were incubated withand without fluorescein-labeled or unlabeled heat-killed M. tuberculosis for 20 min or with soluble M. tuberculosis Ag 85B for 30 min, washed,homogenized, and fractionated on 27% Percoll gradients. A. Distribution of fluorescein-labeled heat-killed M. tuberculosis phagosomes. Data arerepresentative of six independent experiments. B. Distribution of plasma membrane when THP-1 (incubated with heat-killed M. tuberculosis)plasma membranes were labeled with sulfo-NHS-LC-biotin, washed and incubated with streptavidin-fluorescein at 4°C prior to fractionation. Dataare representative of 3 independent experiments. C. �-Hexosaminidase activity (a marker of lysosomal enzyme distribution). Data shown are froma representative experiment. This control was included in every fractionation experiment. D. Distribution of M. tuberculosis Ag 85B (96-111)–DR1complexes in cells incubated with soluble M. tuberculosis Ag 85B as assessed by the F9A6 T hybridoma assay. Data are representative of threeindependent experiments.
1624 TORRES ET AL. INFECT. IMMUN.

(96-111)–DR1 complexes were expressed at high levels on thecell surface by 60 min (Fig. 1B and C). M. tuberculosis-specificCD4� T-cell responses to MDMs incubated with heat-killedM. tuberculosis was also observed by 60 min (Fig. 1D). Incontrast, processing of live M. tuberculosis by both THP-1 cellsand MDMs was poor, with weak T-cell responses even at achase incubation of 240 min (Fig. 1B and C). However, after18 h of chase, live M. tuberculosis processing by both THP-1cells and MDMs was similar to heat-killed M. tuberculosis (dataonly shown for THP-1 cells in Fig. 1B). It should be noted thatTHP-1 cells and MDMs were equally efficient in phagocytosisof live and heat-killed M. tuberculosis as assessed by identifi-cation of acid-fast bacilli with Ziehl-Neelsen staining of cellsand analysis of cells incubated with FLUOS-labeled heat-killedand live M. tuberculosis by flow cytometry. In conclusion, hu-man macrophages process and present heat-killed M. tuber-culosis more rapidly and efficiently than live M. tuberculosis.
Subcellular fractionation of THP-1 cells and MDMs incubatedwith M. tuberculosis. Studies of phagosomal antigen processingrequire separation of phagosomes and other subcellular organ-elles for subsequent analysis. Following various treatments,THP-1 cells and MDMs were fractionated on Percoll densitygradients. The distribution of organelles was carefully deter-mined as previously described with a few modifications (27).To identify phagosomes, cells were incubated with FLUOS-labeled M. tuberculosis. To identify the plasma membrane, cellswere incubated with sulfo-NHS-LC-biotin, followed by incu-bation with streptavidin-fluorescein at 4°C. To identify MIICvesicles, cells were incubated with soluble Ag 85B to labelMIIC vesicles with M. tuberculosis Ag 85B (96-111)–DR1 com-plexes. Cells were homogenized, and phagosomes and otherorganelles were separated on 27% Percoll density gradients.
Phagosomes consistently appeared in fractions 27 to 30 inthe high-density region of 27% Percoll gradients (Fig. 2A),whereas plasma membrane-associated fluorescence was foundin fractions 9 to 12 in the low-density region (Fig. 2B). �-Hexosaminidase, a lysosomal enzyme, was primarily found inhigh-density phagolysosomal/lysosomal fractions (Fig. 2C,fractions 25 to 27). Similar levels of �-hexosaminidase werefound in fractions derived from cells incubated with eitherheat-killed or live M. tuberculosis. FA96 T hybridoma cellsdetected M. tuberculosis Ag 85B (96-11)–DR1 complexes inMIIC endocytic vesicles in fractions 18 to 27 and sometimes onthe plasma membrane in fractions 10 and 11 (Fig. 2D). Thesame results were obtained with fractionation of MDMs (datanot shown).
Analysis of subcellular fractions for M. tuberculosis peptide–MHC-II complexes using a T-cell assay. To determine the site(s)where M. tuberculosis peptide–MHC-II complexes were formedduring M. tuberculosis processing, we used a technique previously
FIG. 3. M. tuberculosis Ag 85B (96-111)–DR1 complexes appear inphagosomes and MIIC fractions in THP-1 cells incubated with heat-killed M. tuberculosis. THP-1 cells were pulsed with M. tuberculosis(multiplicity of infection, 40) for 20 min, washed, and chased for 0 min
(A), 30 min (B), and 2 h (C) before homogenization. Subcellularorganelles were separated on 27% Percoll density gradients. M. tuber-culosis Ag 85B (96-111)–DR1 complexes were detected using F9A6 Thybridoma cells. Culture supernatants were harvested after 24 h andIL-2 production was measured by using a CTLL-2 proliferation assay.Diagrams at the top summarize the positions of different compart-ments in the Percoll gradients (Fig. 2). Data are representative of fourindependent experiments.
VOL. 74, 2006 PROCESSING OF M. TUBERCULOSIS BY HUMAN MACROPHAGES 1625

developed to detect peptide–MHC-II complexes in latex beadand M. tuberculosis phagosomes and other organelles in murinecells (12, 26, 27). This approach uses subcellular fractionation toisolate organelle membranes, which are disrupted by freezing andthawing to expose the lumenal antigen-presenting domain ofMHC-II molecules and then probed for peptide–MHC-II com-plexes using T cells.
High levels of peptide–MHC-II complexes appeared on thecell surface following a 60-min chase with heat-killed M. tuber-culosis (Fig. 1). Therefore, in order to determine where pep-tide–MHC-II complexes are formed intracellularly, chase pe-riods of less then 60 min are needed. THP-1 cells and MDMswere pulsed for 20 min with heat-killed M. tuberculosis, chasedfor either 0, 30 or 120 min and fractionated on 27% Percollgradients. THP-1-derived fractions were incubated with F9A6T hybridoma cells, and detection of M. tuberculosis Ag 85B(96-111)–DR1 complexes was monitored by IL-2 secretion(Fig. 3). MDM-derived fractions were incubated with M. tu-berculosis-specific CD4� T cells, and detection of M. tubercu-losis peptide–MHC-II complexes was monitored by IFN-� se-cretion (Fig. 4). We were unable to analyze subcellularfractions of macrophages infected with live M. tuberculosis forup to 6 h due to insufficient signal for T-cell assays. Fraction-ation of cells infected with live M. tuberculosis overnight wasnot technically possible.
When THP-1 cells were pulsed with heat-killed M. tubercu-losis for 20 min with no chase incubation, M. tuberculosis Ag85B (96-111)–DR1 complexes were detected primarily inphagosomes (fractions 26 to 28) with little expression on theplasma membrane (Fig. 3A). After a 20-min pulse and 30-minchase, M. tuberculosis Ag 85B (96-111)–DR1 complexes weredetected in both phagosomes (fractions 26–29) and MIIC frac-tions (fractions 18 to 27) as well as on the plasma membrane(fractions 9 to 14, Fig. 3B). When the chase period was in-creased to 120 min, M. tuberculosis Ag 85B (96 to 111)–DR1complexes were detected primarily in plasma membrane frac-tions with some complexes still present in MIIC compartments(Fig. 3C). When 0.5 �M M. tuberculosis Ag 85B (96-111) pep-tide was added to the fractions during the T-cell assay, F9A6cells responded to all fractions containing MHC-II, providing apositive control and confirming the ability of F9A6 T cells torespond to all fractions where M. tuberculosis Ag 85B (96-111)–DR1 complexes were present (data not shown).
Thus, in THP-1 cells, M. tuberculosis Ag 85B (96-111)–DR1complexes were initially formed in M. tuberculosis phagosomes(Fig. 3A). The presence of complexes in the MIIC compart-ment at the 30-min chase point (Fig. 3B) suggests either thatthe MIIC compartment receives phagosome-derived antigenfragments for generation of M. tuberculosis Ag 85B (96-11)–DR1 complexes within the MIIC compartment or that com-plexes formed in the phagosomes traffic through the MIICcompartment en route to the plasma membrane.
When MDMs were pulsed with heat-killed M. tuberculosis
FIG. 4. M. tuberculosis peptide–MHC-II complexes appear inMIIC fractions in MDMs incubated with heat-killed M. tuberculosis.MDMs were pulsed with M. tuberculosis (multiplicity of infection, 40)for 20 min, washed, and chased for 0 min (A), 30 min (B), and 2 h(C) before homogenization. Subcellular organelles were separated on27% Percoll density gradients. M. tuberculosis peptide–MHC-II com-plexes were detected using polyclonal autologous M. tuberculosis-
specific CD4� T cells. Culture supernatants were harvested after 24 h,and IFN-� production was measured by ELISA. Diagrams at the topsummarize the position of different compartments in the Percoll gra-dients. Data are representative of three independent experiments.
1626 TORRES ET AL. INFECT. IMMUN.
for 20 min without chase, M. tuberculosis peptide–MHC-IIcomplexes appeared in the MIIC compartment (fractions 21 to27) with little expression on plasma membrane (Fig. 4A). Asthe chase period was increased to 30 and 120 min, M. tuber-culosis peptide–MHC-II complexes in MIIC declined and com-plexes increased in the plasma membrane fractions (Fig. 4Band C). Polyclonal M. tuberculosis-specific T cells did not re-spond to fractions from MHC-II mismatched donors. We con-clude that in MDMs, M. tuberculosis peptide–MHC-II com-plexes are formed in the MIIC compartment that receivesphagosome-derived antigen fragments.
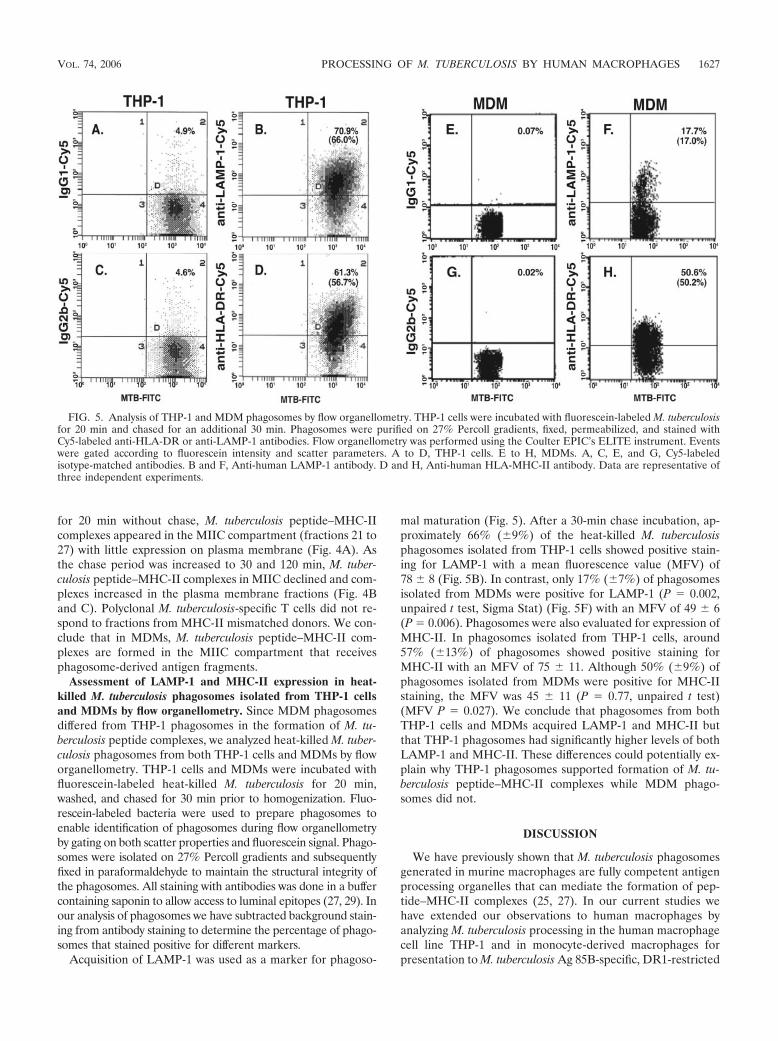
Assessment of LAMP-1 and MHC-II expression in heat-killed M. tuberculosis phagosomes isolated from THP-1 cellsand MDMs by flow organellometry. Since MDM phagosomesdiffered from THP-1 phagosomes in the formation of M. tu-berculosis peptide complexes, we analyzed heat-killed M. tuber-culosis phagosomes from both THP-1 cells and MDMs by floworganellometry. THP-1 cells and MDMs were incubated withfluorescein-labeled heat-killed M. tuberculosis for 20 min,washed, and chased for 30 min prior to homogenization. Fluo-rescein-labeled bacteria were used to prepare phagosomes toenable identification of phagosomes during flow organellometryby gating on both scatter properties and fluorescein signal. Phago-somes were isolated on 27% Percoll gradients and subsequentlyfixed in paraformaldehyde to maintain the structural integrity ofthe phagosomes. All staining with antibodies was done in a buffercontaining saponin to allow access to luminal epitopes (27, 29). Inour analysis of phagosomes we have subtracted background stain-ing from antibody staining to determine the percentage of phago-somes that stained positive for different markers.
Acquisition of LAMP-1 was used as a marker for phagoso-
mal maturation (Fig. 5). After a 30-min chase incubation, ap-proximately 66% (�9%) of the heat-killed M. tuberculosisphagosomes isolated from THP-1 cells showed positive stain-ing for LAMP-1 with a mean fluorescence value (MFV) of78 � 8 (Fig. 5B). In contrast, only 17% (�7%) of phagosomesisolated from MDMs were positive for LAMP-1 (P � 0.002,unpaired t test, Sigma Stat) (Fig. 5F) with an MFV of 49 � 6(P � 0.006). Phagosomes were also evaluated for expression ofMHC-II. In phagosomes isolated from THP-1 cells, around57% (�13%) of phagosomes showed positive staining forMHC-II with an MFV of 75 � 11. Although 50% (�9%) ofphagosomes isolated from MDMs were positive for MHC-IIstaining, the MFV was 45 � 11 (P � 0.77, unpaired t test)(MFV P � 0.027). We conclude that phagosomes from bothTHP-1 cells and MDMs acquired LAMP-1 and MHC-II butthat THP-1 phagosomes had significantly higher levels of bothLAMP-1 and MHC-II. These differences could potentially ex-plain why THP-1 phagosomes supported formation of M. tu-berculosis peptide–MHC-II complexes while MDM phago-somes did not.
DISCUSSION
We have previously shown that M. tuberculosis phagosomesgenerated in murine macrophages are fully competent antigenprocessing organelles that can mediate the formation of pep-tide–MHC-II complexes (25, 27). In our current studies wehave extended our observations to human macrophages byanalyzing M. tuberculosis processing in the human macrophagecell line THP-1 and in monocyte-derived macrophages forpresentation to M. tuberculosis Ag 85B-specific, DR1-restricted
FIG. 5. Analysis of THP-1 and MDM phagosomes by flow organellometry. THP-1 cells were incubated with fluorescein-labeled M. tuberculosisfor 20 min and chased for an additional 30 min. Phagosomes were purified on 27% Percoll gradients, fixed, permeabilized, and stained withCy5-labeled anti-HLA-DR or anti-LAMP-1 antibodies. Flow organellometry was performed using the Coulter EPIC’s ELITE instrument. Eventswere gated according to fluorescein intensity and scatter parameters. A to D, THP-1 cells. E to H, MDMs. A, C, E, and G, Cy5-labeledisotype-matched antibodies. B and F, Anti-human LAMP-1 antibody. D and H, Anti-human HLA-MHC-II antibody. Data are representative ofthree independent experiments.
VOL. 74, 2006 PROCESSING OF M. TUBERCULOSIS BY HUMAN MACROPHAGES 1627

T hybridoma cells (F9A6) or polyclonal autologous M. tuber-culosis-specific CD4� T cells.
Processing of heat-killed M. tuberculosis for presentation toT cells (Fig. 1B to D) was initiated rapidly in both THP-1 cellsand MDMs, and complexes could be observed on the cellsurface by 60 min. In contrast, processing of live M. tuberculosiswas poor and few complexes appeared on the cell surface evenafter prolonged chase incubations. However, after 18 h ofchase, live M. tuberculosis processing was similar to that ofheat-killed M. tuberculosis (Fig. 1B), indicating that M. tuber-culosis viability influences the process involved in early eventsof antigen processing.
To determine if live and heat-killed M. tuberculosis differ-entially regulated MHC-II levels on THP-1 cells and MDMs,we assessed MHC-II surface levels on MDMs and THP-1 cellsinfected with heat-killed or live M. tuberculosis at an early timepoint (2 h) and at a late time point (18 h). While no differenceswere seen at the early time point, overnight incubation of cellswith live M. tuberculosis resulted in decreased surface MHC-II(data not shown). Therefore, levels of surface MHC-II do notcorrelate with the differences in processing of heat-killed andlive M. tuberculosis at early or late time points.
Pancholi et al. (21) have previously reported good antigenprocessing and presentation in macrophages infected for 2days with live BCG and poor antigen presentation in macro-phages infected for 4 days (chronic infection). While our re-sults (18 h time point) are similar to what they found on day 2,our studies have focused on early events in antigen processingand presentation. Since live M. tuberculosis can inhibit phago-some maturation, our observations suggest that live M. tuber-culosis may be able to inhibit both phagosome maturation andMHC-II antigen processing. However, phagosomal processingof M. tuberculosis Ag 85B in murine macrophages is modulatedindependently of mycobacterial viability and phagosome mat-uration (25).
To analyze the intracellular sites of M. tuberculosis peptide–MHC-II complex formation, such as MIIC and M. tuberculosisphagosomes, subcellular fractionation was coupled with M.tuberculosis-specific T-cell assays or flow organellometry.Plasma membrane and phagosome fractions were well sepa-rated on 27% Percoll gradients (Fig. 2), and no significantplasma membrane contamination of the phagosomal fractionswas observed (no fluorescein isothiocyanate signal in phago-somal fractions, Fig. 2B). Since approximately 60% of totalMHC-II is found on the plasma membrane in murine macro-phages (13), contamination of phagosomes with plasma mem-brane would have interfered with evaluation of phagosomalMHC-II levels. The distribution of phagosomes and MIIC didoverlap on 27% Percoll gradients (Fig. 2). Since MIIC containsintracellular MHC-II molecules, in some experiments wherephagosomes were isolated for evaluation of MHC-II levels byflow organellometry, differential centrifugation to separatephagosomes from smaller membrane structures, e.g., MIIC,was carried out prior to fractionation on 27% Percoll gradients(27). This approach provided us with pure preparations of M.tuberculosis phagosomes for our flow organellometry analyses.
To determine where M. tuberculosis peptide–MHC-II com-plexes were formed intracellularly, macrophages were incu-bated with heat-killed M. tuberculosis for various periods andfractionated on Percoll density gradients. Fractionation of
THP-1 cells and MDMs infected with live M. tuberculosis wasunsuccessful due to insufficient signal for T-cell assays. InTHP-1 fractions, M. tuberculosis Ag 85B (96-11)–DR1 com-plexes appeared in phagosomes soon after pulse incubationwith heat-killed M. tuberculosis (20 min) (Fig. 3A). Subse-quently, complexes were additionally found in MIIC andplasma membrane fractions (Fig. 3B and C). These observa-tions differ from those seen in murine macrophages pulsedwith heat-killed M. tuberculosis, where M. tuberculosis Ag 85Bpeptide–MHC-II complexes were detected in phagosomes andplasma membrane fractions but not in MIIC fractions. Thepresence of M. tuberculosis Ag 85B (96-111)–DR1 complexesin the MIIC fractions suggests that peptide–MHC-II com-plexes were either additionally generated in these compart-ments following export of M. tuberculosis protein fragmentsfrom the phagosome to the MIIC compartment (either directlyor indirectly) or that peptide–MHC-II complexes formed inphagosomes trafficked via the MIIC en route to the plasmamembrane.
In MDM fractions, no M. tuberculosis peptide–MHC-IIcomplexes were ever observed in phagosomes (Fig. 4). Weconsistently saw complexes in the MIIC and plasma mem-brane fractions. Therefore, in MDMs, M. tuberculosis pep-tide–MHC-II complexes are formed in MIIC following di-rect or indirect export of protein fragments fromphagosomes to MIIC compartments. Several observationsdo support a role for endocytic compartments (e.g., MIIC)in the formation of complexes derived from phagocytic an-tigens. For example, export of Ag 85 in addition to otherproteins and lipids, from the M. tuberculosis phagosome toother intracellular compartments, has been reported by sev-eral investigators (3, 20, 31, 35, 41). In addition, in murinemacrophages infected with Streptococcus pyogenes, peptide–MHC-II complexes were consistently observed in endocyticcompartments and not in phagosomes (9).
With polyclonal autologous CD4 T cells we are analyzing abroad spectrum of M. tuberculosis peptide–MHC-II complexesin MDMs. This approach is useful to generalize our observa-tions of M. tuberculosis processing. Since antigens may differ insubcellular processing compartmentalization, this may explainthe differences seen between THP-1 cells and MDMs. Alter-natively, MDM phagosomes may be incapable of supportingthe formation of M. tuberculosis peptide–MHC-II complexesand differ from THP-1 phagosomes in their MHC-II levels andphenotype.
In order to analyze MDM and THP-1 phagosomes we car-ried out flow organellometry of phagosomes containing fluo-rescein-labeled heat-killed M. tuberculosis. Most of the THP-1phagosomes were LAMP-1 positive compared to MDMphagosomes, and the levels of LAMP-1 in THP-1 phagosomeswere also higher than that seen in MDM phagosomes. SinceTHP-1 cells were activated with IFN-� to increase MHC-IIlevels (to facilitate T-cell assays) and IFN-� is known to pro-mote phagosome maturation (2, 18, 32, 38), we do not knowwhether the difference in LAMP-1 levels is due to IFN-� ac-tivation of THP-1 cells or due to more intrinsic differencesbetween the two cell types.
Within the time frame of active antigen processing, similarpercentages of heat-killed M. tuberculosis phagosomes from bothTHP-1 cells and MDMs acquired MHC-II molecules that could
1628 TORRES ET AL. INFECT. IMMUN.

potentially bind peptides derived from M. tuberculosis antigens.However, the levels of MHC-II in MDM phagosomes were lowerthan those of THP-1 phagosomes despite the fact that IFN-�-activated THP-1 cells had surface MHC-II levels that were only80% � 8% of that seen in MDMs. Since MDM phagosomesappear to be different from THP-1 phagosomes and acquirefewer MHC-II molecules (Fig. 5), it is possible that MDM phago-somes are incapable of supporting the formation of M. tuber-culosis peptide–MHC-II complexes.
There are other alternative explanations for the differencesin processing between IFN-�-activated THP-1 cells and MDMs.IFN-� activation could have induced the expression of differ-ent receptors on THP-1 cells and altered the mode of entry ofM. tuberculosis and the subsequent processing events. IFN-�could have also altered trafficking events in THP-1 cells, re-sulting in the differences that we have observed.
In conclusion, we have observed that human macrophagesprocess and present heat-killed M. tuberculosis far more rapidlythan live M. tuberculosis. While M. tuberculosis phagosomescan mediate the formation of peptide–MHC-II complexes inTHP-1 cells, it is the MIIC compartment that receives phago-some-derived antigen fragments for generation of M. tubercu-losis peptide–MHC-II complexes in MDMs. Future studies willhelp determine whether the maturation status of the phago-some dictates its ability to generate peptide–MHC-II com-plexes in human macrophages.
ACKNOWLEDGMENTS
This work was supported by a Public Health Service grant from theFogarty International Center, National Institutes of Health, AIDSInternational Training and Research Program (TW00011) to M.T.,ALA grant RG-045-N to L.R., National Institutes of Health grantsAI-35726 and AI-34343 to C.V.H. and AI-27243 to W.H.B., and con-tract AI-95383 to the Tuberculosis Research Unit to W.H.B.
REFERENCES
1. Amigorena, S., J. R. Drake, P. Webster, and I. Mellman. 1994. Transientaccumulation of new class II MHC molecules in a novel endocytic compart-ment in B lymphocytes. Nature 369:113–120.
2. Appelberg, R., and I. M. Orme. 1993. Effector mechanisms involved incytokine-mediated bacteriostasis of Mycobacterium avium infections in mu-rine macrophages. Immunology 80:352–359.
3. Beatty, W. L., and D. G. Russell. 2000. Identification of mycobacterial sur-face proteins released into subcellular compartments of infected macro-phages. Infect. Immun. 68:6997–7002.
4. Belisle, J. T., V. D. Vissa, T. Sievert, K. Takayama, P. J. Brennan, and G. S.Besra. 1997. Role of the major antigen of Mycobacterium tuberculosis in cellwall biogenesis. Science 276:1420–1422.
5. Boom, W. H., D. H. Canaday, S. A. Fulton, A. J. Gehring, R. E. Rojas, andM. Torres. 2003. Human immunity to M. tuberculosis: T-cell subsets andantigen processing. Tuberculosis 83:98–106.
6. Britton, W. J., and U. Palendira. 2003. Improving vaccines against tubercu-losis. Immunol. Cell Biol. 81:33–45.
7. Canaday, D. H., A. Gehring, E. G. Leonard, B. Eilertson, J. R. Schreiber,C. V. Harding, and W. H. Boom. 2003. T-cell hybridomas from HLA-trans-genic mice as tools for analysis of human antigen processing. J. Immunol.Methods 281:129–142.
8. Cresswell, P. 1996. Invariant chain structure and MHC class II function. Cell84:505–507.
9. Delvig, A., L. Ramachandra, C. V. Harding, and J. H. Robinson. 2003.Localization of peptide/MHC class II complexes in macrophages followingantigen processing of viable Streptococcus pyogenes. Eur. J. Immunol. 32:2353–2360.
10. Flynn, J. L., and J. Chan. 2001. Immunology of tuberculosis. Annu. Rev.Immunol. 19:93–129.
11. Gehring, A. J., R. E. Rojas, D. H. Canaday, D. L. Lakey, C. V. Harding, andW. H. Boom. 2003. The Mycobacterium tuberculosis 19-kilodalton lipopro-tein inhibits gamma interferon-regulated HLA-DR and Fc gamma R1 onhuman macrophages through Toll-like receptor 2. Infect. Immun. 71:4487–4497.
12. Harding, C. V., and H. J. Geuze. 1993. Immunogenic peptides bind to classII MHC molecules in an early lysosomal compartment. J. Immunol. 151:3988–3998.
13. Harding, C. V., and E. R. Unanue. 1989. Antigen processing and intracellularIa. Possible roles of endocytosis and protein synthesis in Ia function. J. Im-munol. 142:12–19.
14. Hirsch, C. S., J. J. Ellner, D. G. Russell, and E. A. Rich. 1994. Complementreceptor-mediated uptake and tumor necrosis factor-alpha-mediated growthinhibition of Mycobacterium tuberculosis by human alveolar macrophages.J. Immunol. 152:743–753.
15. Horwitz, M. A. 1997. A new TB vaccine. Immunologist 5:15–20.16. Jutras, I. D. M. 2005. Phagocytosis: at the crossroads of innate and adaptive
immunity. Annu. Rev. Cell Dev. Biol. 21:511–527.17. Kwack, K., and R. G. Lynch. 2000. A new non-radioactive method for IL-2
bioassay. Mol. Cells 10:575–578.18. MacMicking, J. D., G. A. Taylor, and J. D. McKinney. 2003. Immune control
of tuberculosis by IFN-gamma-inducible LRG-47. Science 302:654–659.19. Miki, K., T. Nagata, T. Tanaka, Y.-H. Kim, M. Uchijima, N. Ohara, S.
Nakamura, M. Okada, and Y. Koide. 2004. Induction of protective cellularimmunity against Mycobacterium tuberculosis by recombinant attenuatedself-destructing Listeria monocytogenes strains harboring eukaryotic expres-sion plasmids for antigen 85 complex and MPB/MPT51. Infect. Immun.72:2014–2021.
20. Neyrolles, O., K. Gould, M. P. Gares, S. Brett, R. Janssen, P. O’Gaora, J. L.Herrmann, M. C. Prevost, E. Perret, J. E. Thole, and D. Young. 2001.Lipoprotein access to MHC class I presentation during infection of murinemacrophages with live mycobacteria. J. Immunol. 166:447–457.
21. Pancholi, P., M. A. Bhardwaj, and R. Steinman. 1993. Sequestation fromimmune CD4� T cells of mycobacteria growing in human macrophages.Science 260:984–986.
22. Peters, P. J., J. J. Neefjes, V. Oorschot, H. L. Ploegh, and H. J. Geuze. 1991.Segregation of MHC class II molecules from MHC class I molecules in theGolgi complex for transport to lysosomal compartments. Nature. 349:669–676
23. Qiu, Y., X. Xu, A. Wandinger-Ness, D. P. Dalke, and S. K. Pierce. 1994.Separation of subcellular compartments containing distinct functional formsof MHC class II. J. Cell Biol. 125:595–605.
24. Ramachandra, L., R. S. Chu, D. Askew, E. H. Noss, D. H. Canaday, N. S.Potter, A. Johnsen, A. M. Krieg, J. G. Nedrud, W. H. Boom, and C. V.Harding. 1999. Phagocytic antigen processing and effects of microbial prod-ucts on antigen processing and T-cell responses. Immunol. Rev. 168:217–239.
25. Ramachandra, L., J. Smialek, S. S. Shank, M. Convery, W. H. Boom, andC. V. Harding. 2005. Phagosomal processing of Mycobacterium tuberculosisantigen 85B is modulated independently of mycobacterial viability andphagosome maturation. Infect. Immun. 73:1097–1105.
26. Ramachandra, L., E. Noss, W. H. Boom, and C. V. Harding. 1999. Phagocyticprocessing of antigens for presentation by class II major histocompatibilitycomplex molecules.. Cell. Microbiol. 1:205–214.
27. Ramachandra, L., E. Noss, W. H. Boom, and C. V. Harding. 2001. Processingof Mycobacterium tuberculosis antigen 85B involves intraphagosomal for-mation of peptide-major histocompatibility complex II complexes and isinhibited by live bacilli that decrease phagosome maturation. J. Exp. Med.194:1421–1432.
28. Ramachandra, L., R. Song, and C. V. Harding. 1999. Phagosomes are fullycompetent antigen processing organelles that mediate the formation of pep-tide–class II MHC complexes. J. Immunol. 162:3263–3272.
29. Ramachandra, L., R. M. Sramkoski, D. H. Canaday, W. H. Boom, and C. V.Harding. 1998. Flow analysis of MHC molecules and other membrane pro-teins in isolated phagosomes. J. Immunol. Methods 213:53–71.
30. Rudensky, A. Y., M. Maric, S. Eastmen, L. Shoemaker, P. C. DeRoos, andJ. S. Blum. 1994. Intracellular assembly and transport of endogenous pep-tide-MHC class II complexes. Immunity 1:585–594.
31. Schaible, U. E., K. Hagens, K. Fischer, H. L. Collins, and S. H. Kaufmann.2000. Intersection of group I CD1 molecules and mycobacteria in differentintracellular compartments of dendritic cells. J. Immunol. 164:4843–4852.
32. Schaible, U. E., S. Sturgill-Koszycki, P. H. Schlesinger, and D. G. Russell.1998. Cytokine activation leads to acidification and increases maturation ofMycobacterium avium-containing phagosomes in murine macrophages.J. Immunol. 160:1290–1296.
33. Schlesinger, L. S. 1993. Macrophage phagocytosis of virulent but not atten-uated strains of Mycobacterium tuberculosis is mediated by mannose recep-tors in addition to complement receptors. J. Immunol. 150:2920–2930.
34. Tan, J. S., D. H. Canaday, W. H. Boom, K. N. Balaji, S. K. Schwander, andE. A. Rich. 1997. Human alveolar T lymphocyte responses to Mycobacteriumtuberculosis antigens: role for CD4� and CD8� cytotoxic T cells and rela-tive resistance of alveolar macrophages to lysis. J. Immunol. 159:290–297.
35. Teitelbaum, R., M. Cammer, M. L. Maitland, N. E. Freitag, J. Condeelis,and B. R. Bloom. 1999. Mycobacterial infection of macrophages results inmembrane-permeable phagosomes. Proc. Natl. Acad. Sci. USA 96:15190–15195.
36. Tulp, A., D. Verwoerd, B. Dobberstein, H. L. Ploegh, and J. Pieters. 1994.
VOL. 74, 2006 PROCESSING OF M. TUBERCULOSIS BY HUMAN MACROPHAGES 1629

Isolation and characterization of the intracellular MHC class II compart-ment. Nature 369:120–126.
37. Vergne, I., J. Chua, S. B. Singh, and V. Deretic. 2004. Cell biology of myco-bacterium tuberculosis phagosome. Annu. Rev. Cell Dev. Biol. 20:367–394.
38. Via, L. E., R. A. Fratti, M. McFalone, E. Pagan-Ramos, D. Deretic, and V.Deretic. 1998. Effects of cytokines on mycobacterial phagosome maturation.J. Cell Sci. 111:897–905.
39. West, M. A., J. M. Lucocq, and C. Watts. 1994. Antigen processing and classII MHC peptide-loading compartments in human B-lymphoblastoid cells.Nature 369:147–151.
40. Wiker, H. G., and M. Harboe. 1992. The antigen 85 complex: a major secretionproduct of Mycobacterium tuberculosis. Microbiol. Rev. 56:648–661.
41. Xu, S., A. Cooper, S. Sturgill-Koszycki, T. van Heyningen, D. Chatterjee, I.Orme, P. Allen, and D. G. Russell. 1994. Intracellular trafficking in Myco-bacterium tuberculosis and Mycobacterium avium-infected macrophages.J. Immunol. 153:2568–2578.
42. Zimmerli, S., S. Edwards, and J. D. Ernst. 1996. Selective receptor blockadeduring phagocytosis does not alter the survival and growth of Mycobacteriumtuberculosis in human macrophages. Am. J. Respir. Cell Mol. Biol. 15:760–770.
Editor: W. A. Petri, Jr.
1630 TORRES ET AL. INFECT. IMMUN.
Related Documents











