27 Role of Minerals in Halophyte Feeding to Ruminants SALAH A. ATTIA-ISMAIL Desert Research Center, Matareya, 11753 Cairo, Egypt 1 INTRODUCTION Halophyte plants are those that grow naturally in saline environment. Halophytes grow in many arid and semi-arid regions around the world and are distributed from coastal areas to mountains and deserts. Saline conditions may include either salt- affected soil or saline water irrigation. Economically speaking, the most productive halophytic plant species yield from 10 to 20 ton/ha of biomass when irrigated with seawater [Glenn et al., 1999]. Also, the productivity of cultivated halophytes, as compared with traditional crops, is high [Jaradat, 2003]. The average area of salt-affected soils might be calculated on the basis of FAO averages to be 450 million hectares. That means we are talking in about 4–5 billion tons of biomass. Halophytic plants have many uses: They can be used as animal feeds, vegetables, and drugs, as well as for sand dune fixation, wind shelter, soil cover, cultivation of swampy saline lands, laundry detergents, paper production, and so on [El Shaer and Attia-Ismail, 2002]. Feed shortage especially in coastal areas has led to the exploration and exploitation of marginal resources such as halophytic plants. Halophytes can, then, play an important role in the welfare of people in such regions. As an animal feed component, halophytes are promising because they have the potentiality of being good feed resource. The potentialities of halophytes as animal feed components were recognized as early as the 1880s [Hutchings, 1965]. Yet, this potentiality does not go far because several constraints are limiting and have to be worked out. One of these constraints is the high contents of salts and conse- quently the high mineral composition. Trace Elements as Contaminants and Nutrients: Consequences in Ecosystems and Human Health, Edited by M. N. V. Prasad Copyright # 2008 John Wiley & Sons, Inc. 701

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

27 Role of Minerals in HalophyteFeeding to Ruminants
SALAH A. ATTIA-ISMAIL
Desert Research Center, Matareya, 11753 Cairo, Egypt
1 INTRODUCTION
Halophyte plants are those that grow naturally in saline environment. Halophytesgrow in many arid and semi-arid regions around the world and are distributed fromcoastal areas to mountains and deserts. Saline conditions may include either salt-affected soil or saline water irrigation.
Economically speaking, the most productive halophytic plant species yield from10 to 20 ton/ha of biomass when irrigated with seawater [Glenn et al., 1999].Also, the productivity of cultivated halophytes, as compared with traditional crops,is high [Jaradat, 2003]. The average area of salt-affected soils might be calculatedon the basis of FAO averages to be 450 million hectares. That means we aretalking in about 4–5 billion tons of biomass. Halophytic plants have many uses:They can be used as animal feeds, vegetables, and drugs, as well as for sanddune fixation, wind shelter, soil cover, cultivation of swampy saline lands, laundrydetergents, paper production, and so on [El Shaer and Attia-Ismail, 2002].
Feed shortage especially in coastal areas has led to the exploration and exploitationof marginal resources such as halophytic plants. Halophytes can, then, play animportant role in the welfare of people in such regions.
As an animal feed component, halophytes are promising because they have thepotentiality of being good feed resource. The potentialities of halophytes as animalfeed components were recognized as early as the 1880s [Hutchings, 1965]. Yet,this potentiality does not go far because several constraints are limiting and haveto be worked out. One of these constraints is the high contents of salts and conse-quently the high mineral composition.
Trace Elements as Contaminants and Nutrients: Consequences in Ecosystems andHuman Health, Edited by M. N. V. PrasadCopyright # 2008 John Wiley & Sons, Inc.
701

2 ASH AND MINERAL CONTENTS OF HALOPHYTES
The high content of ash is a typical characteristic of halophytes. Mineralcontents differ in halophytes from those of regular forages. This has led to scientificconcerns of with regard to what extent this may affect their feeding value toanimals. Also, the mineral compositions of the high ash contents of halophyteshave been controversial: Do they support animals’ requirements of certain minerals,do they might exceed the requirements to the extent that they may represent a poison-ing threat to animals? Mineral contents of halophytes vary considerably. Thevariations in mineral contents of halophytes depend on several factors. Amongthe many factors that cause the variations are the plant species, stage of growth,and season.
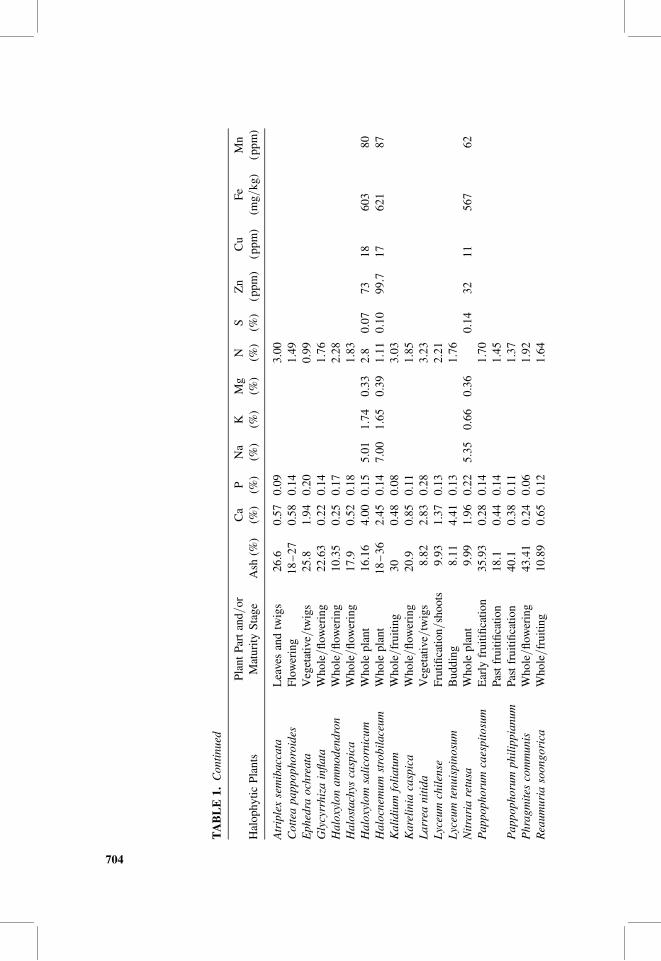
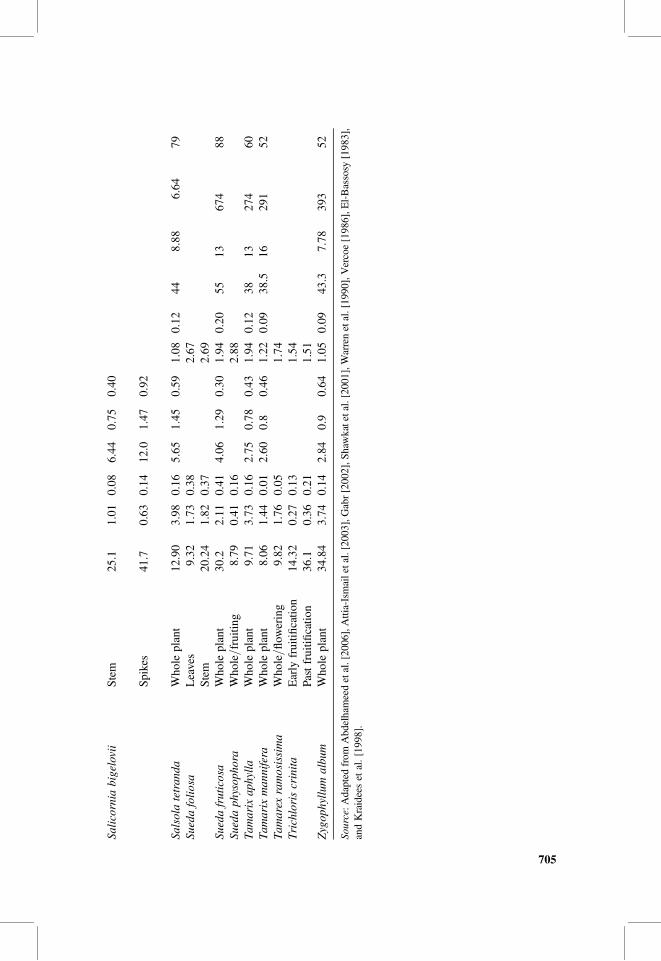
Table 1 shows the ash and mineral contents of different parts of some halophytesaround the world. Halophytes may represent a good source of minerals to ruminantanimals. High ash contents of halophytes, unless associated with nondigestible com-ponents such as silica, may compensate for mineral deficiency usually seen inanimals in areas depending mainly on rangeland grazing as well as desert andcoastal areas. Table 1 shows that ash contents of halophytes vary for whole plants,maturity stage, and plant part. It runs up to more than 40% of plant materials.Plant parts differ also in their ash contents. Leaves and twigs of Atriplex repanda,for instance, have 23.1%, whereas the fruits contain 32.65% (Table 1); this represents41% increment in ash content. Salicornia bigelovii spikes have 66% more ash thanthe stem. The stem of Sueda foliosa has 117% more ash than leaves.
Stage of plant maturity affects ash contents as well. Pappophorum caespitosumis an example. Its ash contents decreased after fructification by 98.5%. Trichloriscrinita is a contrast case where ash contents increased after fruitification by 152%.The variability in ash contents might be due to the physiological distribution andpools of minerals in different plant parts. Physiological and biochemical processesthat take place in plants vary according to the plant species, and this mayexplain the contrast in the percentage of ash content increment or decrement due tothe process of fruitification between both of Pappophorum caespitosum andTrichloris crinita.
3 FACTORS AFFECTING MINERAL CONTENTSOF HALOPHYTES
Halophyte plants either grow naturally in saline environment (salt-affected soils orunder the circumstances of the irrigation with saline water) or need be cultivated.Either way has its impacts on ash contents and mineral composition of the plants.Minerals in halophytes differ widely according to the particularities of the environ-ment they grow within. Type of soil (saline or sodic), level of salinity (in eithersoil or irrigation water), mineral profile of the soil, plant species, season, stage ofgrowth, part of the plant, and so on, are among factors affecting ash and mineralcontents of halophytes. Mineral contents are, therefore, affected accordingly.
702 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS

TA
BL
E1.
Ash
and
Min
eral
Con
tent
sof
Som
eH
alop
hyti
cP
lant
sfr
omD
iffe
rent
Par
tsof
the
Wor
ld
Hal
ophy
ticP
lant
sP
lant
Part
and/
orM
atur
ityS
tage
Ash
(%)
Ca
(%)
P (%)
Na
(%)
K (%)
Mg
(%)
N (%)
S (%)
Zn
(ppm
)C
u(p
pm)
Fe(m
g/kg
)M
n(p
pm)
Aca
cia
aneu
raL
eave
s5.
151.
390.
070.
010.
260.
1776
863
Aca
cia
bidw
illi
Lea
ves
2.43
0.09
0.06
0.24
0.21
3749
Aca
cia
conc
urre
nsL
eave
s0.
970.
050.
250.
260.
1847
917
Aca
cia
dean
eiL
eave
s0.
700.
090.
090.
270.
1537
64A
caci
afim
bria
taL
eave
s0.
300.
060.
130.
250.
1233
83A
caci
agl
auco
carp
aL
eave
s1.
010.
050.
080.
230.
1445
53A
caci
ale
ptoc
arpa
Lea
ves
0.97
0.06
0.25
0.37
0.18
7562
2A
caci
am
elan
oxyl
onL
eave
s0.
810.
090.
160.
280.
1664
457
Aca
cia
salic
ina
Lea
ves
3.52
0.11
0.02
0.29
1.13
5530
Aca
cia
salig
naW
hole
8.83
3.75
1.15
1.05
6.14
2.21
140.
5A
caci
ast
enop
hylla
Lea
ves
2.08
0.10
0.16
0.35
0.58
123
42A
naba
sis
brev
ifolia
Who
le/fl
ower
ing
17.5
3.19
0.08
1.52
Ari
stid
am
endo
cina
Ear
lyfr
uitifi
catio
n22
.04
0.33
0.12
1.10
Atr
iple
xar
gent
ina
Lea
ves
and
twig
s6.
950.
670.
082.
71A
trip
lex
atac
amen
sis
Lea
ves
and
twig
s27
0.68
0.11
2.55
Atr
iple
xco
quim
bens
isL
eave
san
dtw
igs
30.5
60.
870.
102.
69A
trip
lex
halim
usW
hole
plan
t29
.20
1.69
0.32
3.91
0.57
0.32
2.11
0.17
6410
503
51A
trip
lex
lam
paT
wig
s30
0.15
0.01
1.80
Ari
plex
num
mul
aria
Who
lepl
ant
18.9
12.
081.
174.
992.
9915
.63
2.03
133.
560
.52
Atr
iple
xre
pand
aL
eave
san
dtw
igs
23.1
1.9
0.12
3.31
Fru
its32
.65
0.48
0.14
1.65
(Con
tinue
d)
703
TA
BL
E1.
Con
tinue
d
Hal
ophy
ticP
lant
sP
lant
Part
and/
orM
atur
ityS
tage
Ash
(%)
Ca
(%)
P (%)
Na
(%)
K (%)
Mg
(%)
N (%)
S (%)
Zn
(ppm
)C
u(p
pm)
Fe(m
g/kg
)M
n(p
pm)
Atr
iple
xse
mib
acca
taL
eave
san
dtw
igs
26.6
0.57
0.09
3.00
Cot
tea
papp
opho
roid
esF
low
erin
g18
–27
0.58
0.14
1.49
Eph
edra
ochr
eata
Veg
etat
ive/
twig
s25
.81.
940.
200.
99G
lycy
rrhi
zain
flata
Who
le/fl
ower
ing
22.6
30.
220.
141.
76H
alox
ylon
amm
oden
dron
Who
le/fl
ower
ing
10.3
50.
250.
172.
28H
alos
tach
ysca
spic
aW
hole
/flow
erin
g17
.90.
520.
181.
83H
alox
ylom
salic
orni
cum
Who
lepl
ant
16.1
64.
000.
155.
011.
740.
332.
80.
0773
1860
380
Hal
ocne
mum
stro
bila
ceum
Who
lepl
ant
18–
362.
450.
147.
001.
650.
391.
110.
1099
.717
621
87K
alid
ium
folia
tum
Who
le/f
ruiti
ng30
0.48
0.08
3.03
Kar
elin
iaca
spic
aW
hole
/flow
erin
g20
.90.
850.
111.
85L
arre
ani
tida
Veg
etat
ive/
twig
s8.
822.
830.
283.
23L
yceu
mch
ilens
eF
rutifi
catio
n/sh
oots
9.93
1.37
0.13
2.21
Lyc
eum
tenu
ispi
nosu
mB
uddi
ng8.
114.
410.
131.
76N
itrar
iare
tusa
Who
lepl
ant
9.99
1.96
0.22
5.35
0.66
0.36
0.14
3211
567
62P
appo
phor
umca
espi
tosu
mE
arly
frui
tifica
tion
35.9
30.
280.
141.
70Pa
stfr
uitifi
catio
n18
.10.
440.
141.
45P
appo
phor
umph
ilipp
ianu
mPa
stfr
uitifi
catio
n40
.10.
380.
111.
37P
hrag
mite
sco
mm
unis
Who
le/fl
ower
ing
43.4
10.
240.
061.
92R
eaum
uria
soon
gori
caW
hole
/fru
iting
10.8
90.
650.
121.
64
704
Salic
orni
abi
gelo
vii
Ste
m25
.11.
010.
086.
440.
750.
40
Spi
kes
41.7
0.63
0.14
12.0
1.47
0.92
Sals
ola
tetr
anda
Who
lepl
ant
12.9
03.
980.
165.
651.
450.
591.
080.
1244
8.88
6.64
79Su
eda
folio
saL
eave
s9.
321.
730.
382.
67S
tem
20.2
41.
820.
372.
69Su
eda
frut
icos
aW
hole
plan
t30
.22.
110.
414.
061.
290.
301.
940.
2055
1367
488
Sued
aph
ysop
hora
Who
le/f
ruiti
ng8.
790.
410.
162.
88Ta
mar
ixap
hylla
Who
lepl
ant
9.71
3.73
0.16
2.75
0.78
0.43
1.94
0.12
3813
274
60Ta
mar
ixm
anni
fera
Who
lepl
ant
8.06
1.44
0.01
2.60
0.8
0.46
1.22
0.09
38.5
1629
152
Tam
arex
ram
osis
sim
aW
hole
/flow
erin
g9.
821.
760.
051.
74Tr
ichl
oris
crin
itaE
arly
frui
tifica
tion
14.3
20.
270.
131.
54Pa
stfr
uitifi
catio
n36
.10.
360.
211.
51Z
ygop
hyllu
mal
bum
Who
lepl
ant
34.8
43.
740.
142.
840.
90.
641.
050.
0943
.37.
7839
352
Sour
ce:A
dapt
edfr
omA
bdel
ham
eed
etal
.[20
06],
Atti
a-Is
mai
leta
l.[2
003]
,Gab
r[2
002]
,Sha
wka
teta
l.[2
001]
,War
ren
etal
.[19
90],
Ver
coe
[198
6],E
l-B
asso
sy[1
983]
,an
dK
raid
ees
etal
.[1
998]
.
705

4 SALT-AFFECTED SOILS
Saline soils are the soils that have high percentages of salts. Saline soils have an elec-trical conductivity at the saturation of soil extract of more than 4 dS/m [FAO, 1988].Salt-affected soils are distributed all over the world. Salt-affected soils are classifiedinto two main classes according to the type of salts present in soils: saline soils(solonchak) and sodic soils (solonetz) [Szabolcs, 1974]. FAO [2007] estimated thatthe solonchak soils area varies from 260 to 340 million hectares and are mostlyfound in Saharan Africa, East Africa, Central Asia, Australia, and South America.The solonetz soils are associated with solonchak soils and are estimated to bearound 135 million hectares (http://www.fao.org/ag/agl/agll/wrb/wrbmaps/htm/).Saline soils were classified into three main groups according to the dominantcations: the sodium-dominated soils, the magnesium-dominated soils, and thecalcium-dominated soils (http://www.fao.org/DOCREP/003/Y1899E/y1899e00.htm#toc). In each class of these soils the ratios of cations differ.
The most common salts present in saline soils are sodium, potassium, calcium andmagnesium in the form of chlorides, sulfates or bicarbonates. The sodium and chlo-ride are present in the highest percentage particularly in highly saline soils. The mostcommon sodic salts present in sodic soils are the carbonates [FAO, 1988] particularlysodium.
5 IRRIGATION WITH SALINE WATER
Saline water contains large amounts of dissolved salts. For water to be saline, itshould include more than 1000 ppm dissolved salts; this value can be as high as35,000 ppm for typical seawater. Saline water usually goes along with saline soilsor in some cases independently of saline soils. Saline water varies in its salinity con-tents according to the source (seawater, underground water, or drainage water). Salinedrainage waters may contain toxic levels of trace elements as well as the macro-minerals [Retana et al., 1993]. The minerals in saline water differ according to itssource. Brackish water may contain toxic levels of certain minerals according to theeffluent. Major elements of seawater are Ca, 419 ppm; Mg, 1304 ppm; Na, 10,710;K, 390; Cl, 19,350 (http://www.palomar.edu/oceanography/salty_ocean.htm).
6 SALINITY LEVEL
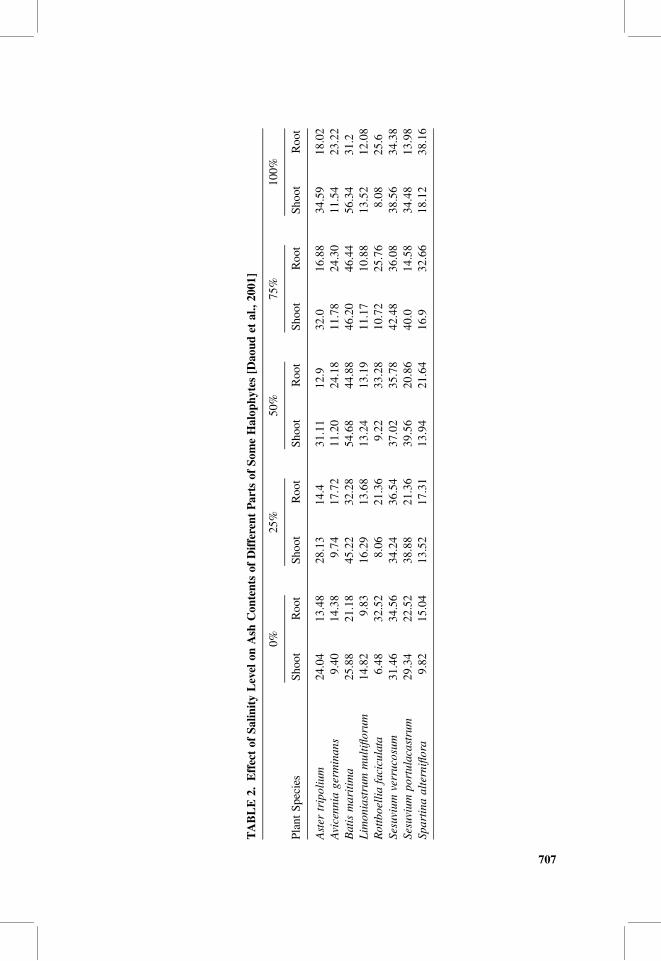
Salinity level either in irrigation water or contained in soils is a major determinantfactor in plant growth and, therefore, in ash contents and mineral profile in plants.Brown et al. [2006] found that salinity level affects nutrient uptake in Spartinaalterniflora. Daoud et al. [2001] mentioned that includer halophytes may accumulatesalt and, therefore, increase mineral concentrations at increasing salinity level, whileexcluder halophytes may also accumulate salt but at lower rates. Table 2 shows thateffect clearly. However, it appears that different parts of the plant respond differently.
706 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS
TA
BL
E2.
Eff
ect
ofSa
linit
yL
evel
onA
shC
onte
nts
ofD
iffe
rent
Par
tsof
Som
eH
alop
hyte
s[D
aoud
etal
.,20
01]
0%25
%50
%75
%10
0%
Pla
ntS
peci
esS
hoot
Roo
tS
hoot
Roo
tS
hoot
Roo
tS
hoot
Roo
tS
hoot
Roo
t
Ast
ertr
ipol
ium
24.0
413
.48
28.1
314
.431
.11
12.9
32.0
16.8
834
.59
18.0
2A
vice
nnia
germ
inan
s9.
4014
.38
9.74
17.7
211
.20
24.1
811
.78
24.3
011
.54
23.2
2B
atis
mar
itim
a25
.88
21.1
845
.22
32.2
854
.68
44.8
846
.20
46.4
456
.34
31.2
Lim
onia
stru
mm
ultifl
orum
14.8
29.
8316
.29
13.6
813
.24
13.1
911
.17
10.8
813
.52
12.0
8R
ottb
oelli
afa
cicu
lata
6.48
32.5
28.
0621
.36
9.22
33.2
810
.72
25.7
68.
0825
.6Se
suvi
umve
rruc
osum
31.4
634
.56
34.2
436
.54
37.0
235
.78
42.4
836
.08
38.5
634
.38
Sesu
vium
port
ulac
astr
um29
.34
22.5
238
.88
21.3
639
.56
20.8
640
.014
.58
34.4
813
.98
Spar
tina
alte
rnifl
ora
9.82
15.0
413
.52
17.3
113
.94
21.6
416
.932
.66
18.1
238
.16
707

At higher salinity levels that reached seawater salinity, sodium concentrationsincreased in shoots while potassium concentrations decreased in roots (Table 2).Noticeably, the salinity level did not affect the concentrations of magnesium orcalcium in their experiments. The distribution of ash and, therefore, mineral contentsin different plant pools might be a mechanism by which the plants adapt to a salineenvironment [Gorham, 1996].
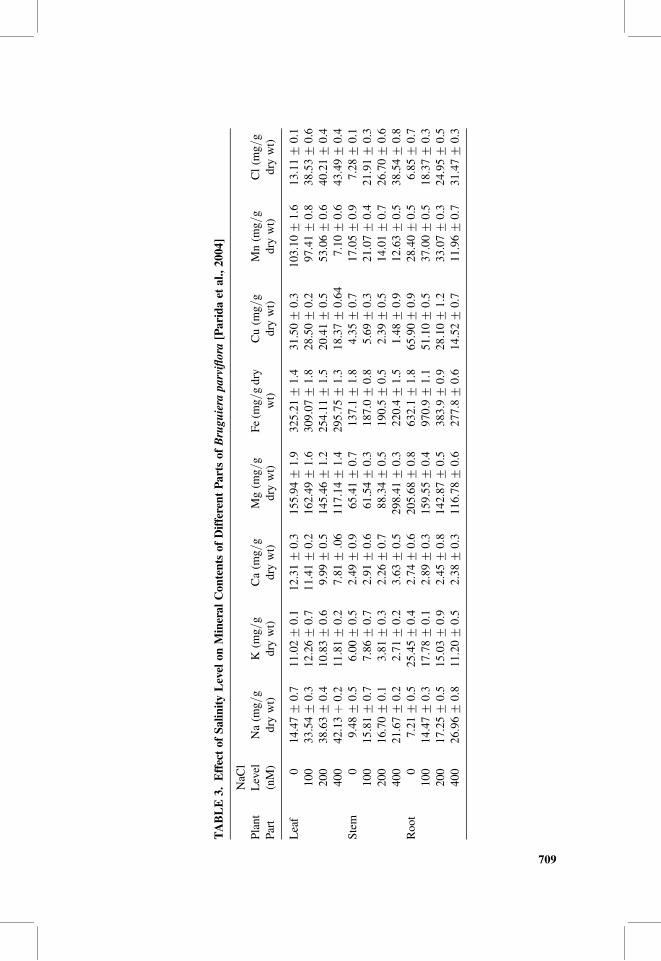
Parida et al. [2004] used Bruguiera parviflora to test the effect of salinity leveltolerance, by adding increasing levels of sodium chloride, on some mineral contentsof different parts of this mangrove (Table 3). The increase in sodium chloride concen-trations led to increases in the concentrations of both of Na and Cl ions in all testedparts of the plant. Khan [2001] reported that the increase in sodium chloride uptakecounteracts the uptake of potassium, calcium, and manganese in some plants.
Certain mineral ratios are affected by salinity level as a consequence of increasingsalinity level like the K/Na, Ca/Na, and many other ratios.
7 PLANT SPECIES
Halophyte plant species differ in their ability to accumulate minerals [Reboreda andCacador, 2007]. The ability of halophytes to accumulate minerals depends on themechanism by which halophytes distribute minerals in different plant compartments.Minerals in halophytes are mainly accumulated in the roots with small quantitiestranslocated to the stems and leaves [Windham et al., 2003]. Halophytes havehigher potentiality to phytoextract minerals from the soil than do glycophytes[Jordan et al., 2002].
Legumes, generally, have different mineral profile (being high in calcium,potassium, magnesium) than grasses which tend to be higher in manganese andmolybdenum when grown under the same conditions.
8 MINERAL ROLE IN RUMINANT NUTRITION
Minerals, in general, either major or minor, play a crucial role in the lives of animalsand affect their production to a greater extent. Minerals have four vital functions inthe bodies of animals. They have a structural function in bones and other structuraltissues of the body, physiological function in body fluids as electrolytes, regulatoryfunction as they regulate several metabolic processes in the body, and catalytic func-tion when they enhance the enzyme activities [Underwood and Suttle, 1999].
9 RECOMMENDED MINERAL ALLOWANCES
Minerals required by animals for proper functioning of the body and for the optimalproduction are either macro- or micro-minerals. Macro-minerals are known to be Ca,
708 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS
TA
BL
E3.
Eff
ect
ofSa
linit
yL
evel
onM
iner
alC
onte
nts
ofD
iffe
rent
Par
tsof
Bru
guie
rapa
rvifl
ora
[Par
ida
etal
.,20
04]
Pla
ntPa
rt
NaC
lL
evel
(nM
)N
a(m
g/g
dry
wt)
K(m
g/g
dry
wt)
Ca
(mg/
gdr
yw
t)M
g(m
g/g
dry
wt)
Fe(m
g/g
dry
wt)
Cu
(mg/
gdr
yw
t)M
n(m
g/g
dry
wt)
Cl
(mg/
gdr
yw
t)
Lea
f0
14.4
7+
0.7
11.0
2+
0.1
12.3
1+
0.3
155.
94+
1.9
325.
21+
1.4
31.5
0+
0.3
103.
10+
1.6
13.1
1+
0.1
100
33.5
4+
0.3
12.2
6+
0.7
11.4
1+
0.2
162.
49+
1.6
309.
07+
1.8
28.5
0+
0.2
97.4
1+
0.8
38.5
3+
0.6
200
38.6
3+
0.4
10.8
3+
0.6
9.99
+0.
514
5.46
+1.
225
4.11
+1.
520
.41+
0.5
53.0
6+
0.6
40.2
1+
0.4
400
42.1
3þ
0.2
11.8
1+
0.2
7.81
+.0
611
7.14
+1.
429
5.75
+1.
318
.37+
0.64
7.10
+0.
643
.49+
0.4
Ste
m0
9.48
+0.
56.
00+
0.5
2.49
+0.
965
.41+
0.7
137.
1+
1.8
4.35
+0.
717
.05+
0.9
7.28
+0.
110
015
.81+
0.7
7.86
+0.
72.
91+
0.6
61.5
4+
0.3
187.
0+
0.8
5.69
+0.
321
.07+
0.4
21.9
1+
0.3
200
16.7
0+
0.1
3.81
+0.
32.
26+
0.7
88.3
4+
0.5
190.
5+
0.5
2.39
+0.
514
.01+
0.7
26.7
0+
0.6
400
21.6
7+
0.2
2.71
+0.
23.
63+
0.5
298.
41+
0.3
220.
4+
1.5
1.48
+0.
912
.63+
0.5
38.5
4+
0.8
Roo
t0
7.21
+0.
525
.45+
0.4
2.74
+0.
620
5.68
+0.
863
2.1+
1.8
65.9
0+
0.9
28.4
0+
0.5
6.85
+0.
710
014
.47+
0.3
17.7
8+
0.1
2.89
+0.
315
9.55
+0.
497
0.9+
1.1
51.1
0+
0.5
37.0
0+
0.5
18.3
7+
0.3
200
17.2
5+
0.5
15.0
3+
0.9
2.45
+0.
814
2.87
+0.
538
3.9+
0.9
28.1
0+
1.2
33.0
7+
0.3
24.9
5+
0.5
400
26.9
6+
0.8
11.2
0+
0.5
2.38
+0.
311
6.78
+0.
627
7.8+
0.6
14.5
2+
0.7
11.9
6+
0.7
31.4
7+
0.3
709

P, Na, Cl, Mg, K, and S, while the micro-minerals are Fe, I, Cu, Zn, Mo, Mn,Co, and Se.
Recent studies have revealed some of the requirements of goats and sheep in dryareas. NRC [1981] recommended mineral requirements for goats and for sheep[NRC, 1985]. Table 4 shows some recommended mineral requirements for goatsand sheep. Clear variations are present when matching mineral supply of halophytesto the requirements of these animal species. Depending on halophytes as the onlymineral source for the animals may result in either mineral deficiency of someelements, toxicity of some others, or malabsorption of some other elementsbecause of the presence of different proportions of them (e.g., Ca/P, K/Na,Ca/Na ratios). However, mineral requirements differ according to several conditionssuch as the season (summer versus winter, especially for the desert animal) and thephysiological state (pregnant, lactating, growing, etc.).
10 MINERALS DEFICIENCY IN HALOPHYTE INCLUDED DIETS
Ash contents of halophytes may limit animal feed intake and digestibility coefficientsof ruminant diets. Therefore, some conservation is placed on the use of these plantspecies in livestock ratios. Adequate trace mineral intake and absorption is requiredfor a variety of metabolic functions including immunity, reproduction, and so on.
Animals, especially those inhabiting desert areas, suffer from high environmentalheat stress. A typical characteristic of animals in desert areas is to dissipate heatthrough several adaptive mechanisms. One of them is to sweat. Sweating in such
TABLE 4. Recommended Mineral Requirements of Sheep and Goats
NRC
Mineral Lactating Goats, 1981 Sheep, 1985
Na (% of DM) 0.09–0.18 0.09–0.18Cl (% of DM) — —Ca (% of DM) 0.20–0.80 0.20–0.82P (% of DM) 0.20–0.40 0.16–0.38Mg (% of DM) 0.12–0.18 0.12–0.18K (% of DM) 0.50–0.80 0.50–0.80S (% of DM) 0.14–0.26 0.14–0.26I (ppm) 0.10–0.80a 0.10–0.80Fe (ppm) 30–50a 30–50Cu (ppm) 10–20a 7–11Mo (ppm) 0.50–1.0a 0.5Co (ppm) 0.10–0.20a 0.1–0.2Mn (ppm) 20–40a 20–40Zn (ppm) 20–33a 20–33Se (ppm) 0.10–0.20a 0.1–0.2
aRick Machen, Texas Agriculture Extension Service.
710 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS

animals [Farid, 1989] is accompanied by the excretion of salts from the body. Thisauthor cited (from MacFarlane et al. [1963]) that camels’ sweat is rich in bicarbonates(pH 8.2–8.5) and is particularly high in potassium, four times as much as sodium.When animals are exposed to high environmental temperatures, the sweating rateincreases as a result (about 0.5 kg/p day in sheep and goats). Thus, higher concen-trations of urea, sodium, potassium, and chloride might be found in the sweat ofanimals [Farid, 1989]. Sweat was found to contain different amounts of Mg, Na,K, Ca, and Cl in humans [Verde et al., 1982].
Animals in these areas suffer from mineral deficiency or are exposed to high con-centrations of various minerals in either feedstuffs or drinking water (if brackish wateris used) or both. For example, Walker [1980] found that the concentration of calciumand potassium is usually higher than that of other minerals, with the average being1–1.5% in Southern African browsers. The deficiency results from either excessivesweating or through the feeding on halophyte plants that are either deficient incertain minerals or that may have imbalanced mineral ratios. The opposite appliesfor mineral toxicity. Figure 1 shows the symptoms of mineral deficiency inanimals fed on Atriplex halimus. The clear calcium deficiency is manifested inthese pictures [Khattab, 2007]. Figure 2 also shows lambs in a state of convulsion,exhibiting retracted head on chest with extension of limbs, while Fig. 3 showslambs in a state of muscular deformities abduction at the foreleg. Figure 4 alsoshows lambs in a state of paralysis of the hind legs; these lambs exhibit incoordina-tion of movement, tremors, convulsion, reluctance to move, aimless movement, andwandering.
Goats browsing naturally growing halophytes in Mexico [Haenlein and Ramirez,2007] were deficient in minerals intake. They found a significant low supply of Mg,Cu, Mn, and Zn from the range plants browsed by goats compared to the requirementsof these minerals. On the other hand, sheep selected different range plants and haddeficiency in Ca, Mg, K, Cu, and Mn. Similarly, phosphorus deficiencies inacacias were reported by Vercoe [1986] and Craig et al. [1991] which led to an imbal-ance in the Ca/P ratio in foliage. Panicum turgidum was fed to camels in the south-eastern region of Egypt [Badawy, 2005]. Mineral composition of this halophyte is
Figure 1. Manifestation of calcium deficiency in lambs fed Atriplex halimus. Courtesy ofDr. Khattab, Desert Research Center, Cairo, Egypt.
10 MINERALS DEFICIENCY IN HALOPHYTE INCLUDED DIETS 711

presented in Table 5. The major problem encountered was the extreme low Ca/P ratiowhich reached 1:19 in wet season and 1:32 in dry season. This ratio is far below thatgenerally recommended (2:1) for domestic animals. They found a similar trend in Mgconcentrations. It tended to be higher in range during dry season than in wet season.
Figure 2. Lambs in a state of convulsion, retracted head on chest with extension of limbs.Courtesy of Dr. Khattab, Desert Research Center, Cairo, Egypt.
Figure 3. Lambs in a state of a muscular detormities adduction at the foreleg and abduction atthe foreleg. Courtesy of Dr. Khattab, Desert Research Center, Cairo, Egypt.
Figure 4. Lambs in a state of paralysis of the hind legs. These lambs exhibit incoordination ofmovement, tremors, convulsion, reluctance to move, aimless movement, and wandering.Courtesy of Dr. Khattab, Desert Research Center, Cairo, Egypt.
712 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS
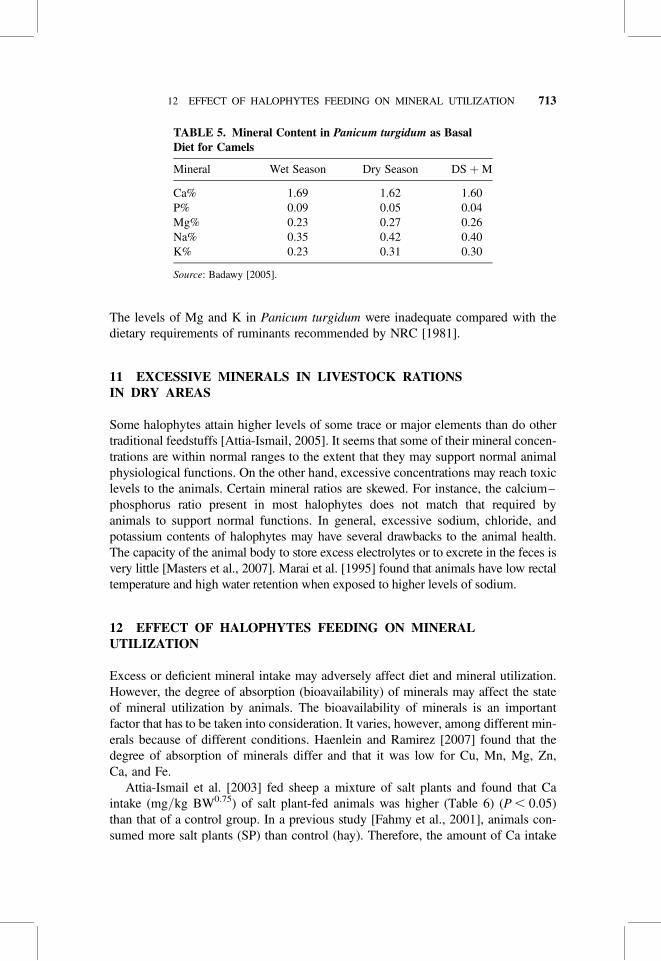
The levels of Mg and K in Panicum turgidum were inadequate compared with thedietary requirements of ruminants recommended by NRC [1981].
11 EXCESSIVE MINERALS IN LIVESTOCK RATIONSIN DRY AREAS
Some halophytes attain higher levels of some trace or major elements than do othertraditional feedstuffs [Attia-Ismail, 2005]. It seems that some of their mineral concen-trations are within normal ranges to the extent that they may support normal animalphysiological functions. On the other hand, excessive concentrations may reach toxiclevels to the animals. Certain mineral ratios are skewed. For instance, the calcium–phosphorus ratio present in most halophytes does not match that required byanimals to support normal functions. In general, excessive sodium, chloride, andpotassium contents of halophytes may have several drawbacks to the animal health.The capacity of the animal body to store excess electrolytes or to excrete in the feces isvery little [Masters et al., 2007]. Marai et al. [1995] found that animals have low rectaltemperature and high water retention when exposed to higher levels of sodium.
12 EFFECT OF HALOPHYTES FEEDING ON MINERALUTILIZATION
Excess or deficient mineral intake may adversely affect diet and mineral utilization.However, the degree of absorption (bioavailability) of minerals may affect the stateof mineral utilization by animals. The bioavailability of minerals is an importantfactor that has to be taken into consideration. It varies, however, among different min-erals because of different conditions. Haenlein and Ramirez [2007] found that thedegree of absorption of minerals differ and that it was low for Cu, Mn, Mg, Zn,Ca, and Fe.
Attia-Ismail et al. [2003] fed sheep a mixture of salt plants and found that Caintake (mg/kg BW0.75) of salt plant-fed animals was higher (Table 6) (P , 0.05)than that of a control group. In a previous study [Fahmy et al., 2001], animals con-sumed more salt plants (SP) than control (hay). Therefore, the amount of Ca intake
TABLE 5. Mineral Content in Panicum turgidum as BasalDiet for Camels
Mineral Wet Season Dry Season DS þM
Ca% 1.69 1.62 1.60P% 0.09 0.05 0.04Mg% 0.23 0.27 0.26Na% 0.35 0.42 0.40K% 0.23 0.31 0.30
Source: Badawy [2005].
12 EFFECT OF HALOPHYTES FEEDING ON MINERAL UTILIZATION 713

of the SP group and, hence, the balance were higher (P , 0.05) than control. Yet,serum Ca concentrations (mg%) of the SP group was almost similar to that of thecontrol group. Sodium (Na) intake of salt plant-fed groups was, logically, higherthan control group (Table 6).
Fahmy [1998] found Na intake to vary from 805 to 1507 mg/kg BW0.75 forsilages made of a mixture of salt plants; a range close to that was obtained in thework carried out by Attia-Ismail et al. [2003]. The Na balance in the SP-fedanimals was, however, lower than that of the control group. The excreted Na inthis experiment might be higher for the SP-fed animals than for the control group.Serum Na did not differ (Table 6). Potassium utilization followed the same trendas Na did except that serum K of control was higher than the SP-fed animals.
13 EFFECT OF MINERALS ON RUMEN FUNCTION
The role of mineral elements in rumen microbial activities has not generally receivedenough attention in work dealing with mineral metabolism in ruminants. Minerals,VFA, lactate, and glucose are the primary solutes in ruminal fluid. Short-chainfatty acids contribute to the osmotic pressure in the rumen. Major minerals contribute
TABLE 6. Mineral Utilization of Sheep Fed a Mixture of HalophyticPlants [Attia-Ismail et al., 2003]
Halophyte-Fed Sheep Control Group
Calcium Utilization
Ca intake (mg/BW0.75) 914a 484b
Ca balance (mg/BW0.75) 914a 341b
Serum Ca (mg%) 8.2a 10.9a
Sodium Utilization
Na intake (mg/BW0.75) 1546a 1068b
Na balance (mg/BW0.75) 252a 316a
Serum Na (mg%) 421a 431a
Potassium Utilization
K intake (mg/BW0.75) 1138 1049K balance (mg/BW0.75) 8b 61a
Serum K (mg%) 25.5a 61b
Zinc Utilization
Zn intake (mg/BW0.75) 45.2a 31.2b
Zn balance (mg/BW0.75) 40.4a 28.6b
Serum Zn (mg%) 1.22a 0.85b
a,bValues bearing different superscripts in the same row differ significantly( p , 0.05).
714 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS
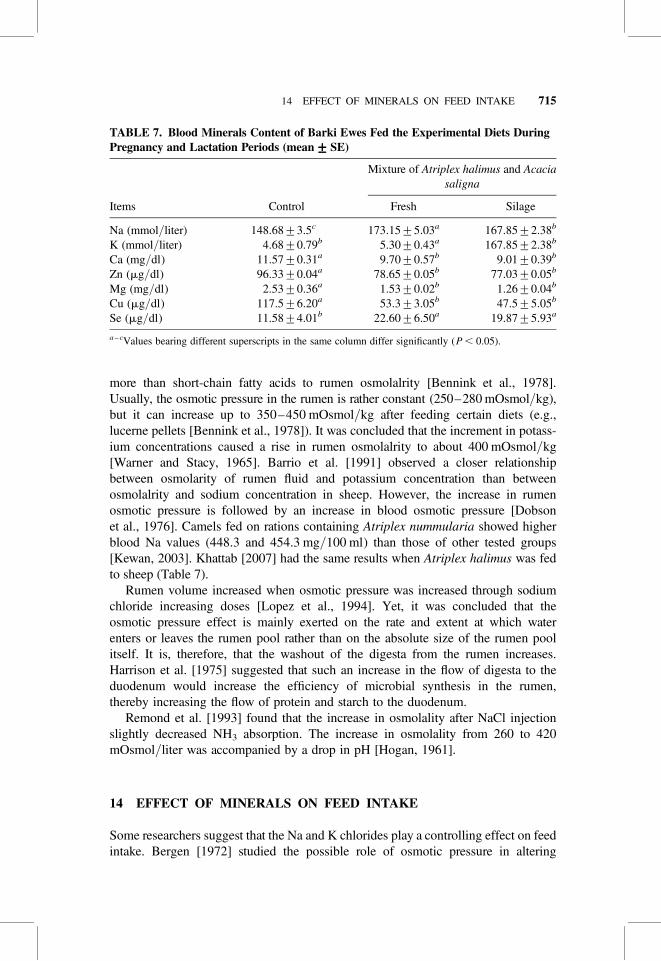
more than short-chain fatty acids to rumen osmolalrity [Bennink et al., 1978].Usually, the osmotic pressure in the rumen is rather constant (250–280 mOsmol/kg),but it can increase up to 350–450 mOsmol/kg after feeding certain diets (e.g.,lucerne pellets [Bennink et al., 1978]). It was concluded that the increment in potass-ium concentrations caused a rise in rumen osmolalrity to about 400 mOsmol/kg[Warner and Stacy, 1965]. Barrio et al. [1991] observed a closer relationshipbetween osmolarity of rumen fluid and potassium concentration than betweenosmolalrity and sodium concentration in sheep. However, the increase in rumenosmotic pressure is followed by an increase in blood osmotic pressure [Dobsonet al., 1976]. Camels fed on rations containing Atriplex nummularia showed higherblood Na values (448.3 and 454.3 mg/100 ml) than those of other tested groups[Kewan, 2003]. Khattab [2007] had the same results when Atriplex halimus was fedto sheep (Table 7).
Rumen volume increased when osmotic pressure was increased through sodiumchloride increasing doses [Lopez et al., 1994]. Yet, it was concluded that theosmotic pressure effect is mainly exerted on the rate and extent at which waterenters or leaves the rumen pool rather than on the absolute size of the rumen poolitself. It is, therefore, that the washout of the digesta from the rumen increases.Harrison et al. [1975] suggested that such an increase in the flow of digesta to theduodenum would increase the efficiency of microbial synthesis in the rumen,thereby increasing the flow of protein and starch to the duodenum.
Remond et al. [1993] found that the increase in osmolality after NaCl injectionslightly decreased NH3 absorption. The increase in osmolality from 260 to 420mOsmol/liter was accompanied by a drop in pH [Hogan, 1961].
14 EFFECT OF MINERALS ON FEED INTAKE
Some researchers suggest that the Na and K chlorides play a controlling effect on feedintake. Bergen [1972] studied the possible role of osmotic pressure in altering
TABLE 7. Blood Minerals Content of Barki Ewes Fed the Experimental Diets DuringPregnancy and Lactation Periods (mean+++++SE)
Mixture of Atriplex halimus and Acaciasaligna
Items Control Fresh Silage
Na (mmol/liter) 148.68+3.5c 173.15+5.03a 167.85+2.38b
K (mmol/liter) 4.68+0.79b 5.30+0.43a 167.85+2.38b
Ca (mg/dl) 11.57+0.31a 9.70+0.57b 9.01+0.39b
Zn (mg/dl) 96.33+0.04a 78.65+0.05b 77.03+0.05b
Mg (mg/dl) 2.53+0.36a 1.53+0.02b 1.26+0.04b
Cu (mg/dl) 117.5+6.20a 53.3+3.05b 47.5+5.05b
Se (mg/dl) 11.58+4.01b 22.60+6.50a 19.87+5.93a
a– cValues bearing different superscripts in the same column differ significantly (P , 0.05).
14 EFFECT OF MINERALS ON FEED INTAKE 715

voluntary feed intake when rumen osmolalrity was artificially elevated to morethan 400 mOsm/kg with NaCl or NaAc just before feeding. He suggested that theshort duration of elevated osmotic pressure when feeding roughages is not animportant factor in controlling feed intake. Kato et al. [1979] reached the same con-clusion when they infused sheep with potassium chloride or sodium chloride duringmeals. On the other hand, when Ternouth and Beattie [1971] infused NaCl, KCl, orsodium salts of organic acids into fistulated sheep, they found that the reduction infood intake is related to osmolalrity of the added fluid and that the reduction was moresignificant when potassium chloride was infused than with sodium chloride. Theyexplained that on the basis of the probable relation of the slower absorption of potassiumcompared with sodium. An elevation in osmotic pressure in the rumen is sensed bythe wall of the reticulo-rumen to inhibit feed intake [Carter and Grovum, 1990].Forbes [1995] concluded that the relative importance of osmolalrity as a satiety factorin ruminants is still uncertain. However, the results of the effect of increased saltintake are inconsistent.
Fahmy et al. [2001] fed a mixture of sun-dried salt plants [22.5% as Zygophyllumalbum, 22.5% as Halocnemum strobilaceum, 45% as Tamarix mannifera, and 10%molasses (w/w)] to growing sheep. The mixture represented 40% of the totalration. The salt-plant-fed group had a higher feed intake than that of the Berseemhay-fed group. Total dry matter intake was comparable and did not differ significantlyamong animal groups. Khattab [2007] fed a mixture of fresh and silage of bothAtriplex halimus and Acacia saligna to sheep lambs. Feed intakes were 1.57 forcontrol, 1.45 for fresh atriplex and acacia, and 1.62 kg/hr/day for ensiled atriplexand acacia. Sodium intakes were 10.81, 24.13, and 21.54 while potassium intakeswere 22.62, 15.74, and 11.69 g/hr/day, respectively.
15 EFFECT OF MINERALS ON WATER INTAKEAND NUTRIENT UTILIZATION
The salt content in halophytes can influence the animal’s water requirement becauseadditional water is required to excrete their high salt content. The high salt levels ofthe foliage increase the water requirements of grazing animals [Wilson, 1974].Gihad [1993] reported that drinking water increased by 61.4% when sheep werefed Atriplex halimus instead of clover hay. Puri and Garg [2001] observed that sup-plementation of sodium chloride increased voluntary water intake, rumen volume,and outflow rate from the rumen. Rapid influx of water to neutralize osmoticpressure swells the ruminal papillae and can pull patches of the ruminal epitheliuminto the rumen by stripping the internal surface layers of the rumen wall fromthe underlying layers, as illustrated vividly in histological studies by Eadie andMann [1970]. Swingle et al. [1996] observed that lambs fed halophyte diets con-sumed up to 110% more water per day and 50% more water per kilogram of drymatter intake.
716 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS

16 EFFECT OF MINERALS ON MICROBIAL COMMUNITYIN THE RUMEN
It is proposed that the increased salt intake and, hence, the osmotic pressure of therumen may affect the microorganism population and metabolism. Increasingosmotic pressure in the rumen is believed to be unfavorable to the growth of protozoa.Also, the increased rates of outflow are believed to contribute to decreased protozoalpopulation [Warner and Stacy, 1977]. The artificial rise of rumen osmotic pressure[Bergen, 1972] up to 400 mOsmol/kg caused cellulose digestion to be inhibited.Shawket et al. [2001] reported that increased salt intake increased dilution rate ofthe rumen fluid phase and negatively affected the protozoal population. Kewan[2003] reported that the total protozoal count (�103) per milliliter was significantlyhigher (P � 0.05) for camels fed on rations that contained Atriplex nummularia thanfor those fed on Acacia saligna and treated rice straw rations. Total rumen protozoalcount in camels fed on berseem hay ration were significantly (P � 0.05) higher thanthose fed on the other experimental rations.
In conclusion, halophytic plants may be used as a feed component in ruminant rationsin spite of the precautions placed on their use in animal feeds. However, if these plants aretreated the right wayaccording to the limitations of each plant species, they would providea plausible feed resource for ruminants in areas like arid, semi-arid, and coastal areas.This way the feed shortage in these areas will be relieved.
REFERENCES
Abdelhameed Afaf AE, Shawkat SM, Hafez IM. 2006. Physiological studies on the effect offeeding salt plants in ewes under semi arid conditions. 4th Sci. Conf. Physiol. Applic. forAnim. Wealth Dev., Cairo, Egypt, July 29–30, pp. 113–132.
Attia-Ismail SA. 2005. Factors limiting and methods of improving nutritive and feeding valuesof halophytes in arid, semi-arid and coastal areas. Conference on Biosaline Agriculture &High Salinity Tolerance, January 9–14, 2005, Mugla, Turkey.
Attia-Ismail SA, Fayed AM, Fahmy AA. 2003. Some mineral and nitrogen utilization of sheepfed salt plant and monensin. Egyptian J Nutr Feeds 6:151–161.
Badawy HS. 2005. Nutritional studies on camels fed on natural ranges in Shalatin–HalaibTriangle region. Ph.D. Cairo University.
Barrio JP, Bapat ST, Forbes JM. 1991. The effect of drinking water on food intake responses tomanipulations of rumen osmolality in sheep. Proc Nutr Soc 50:98A.
Bennink MR, Tyler TR, Ward GM, Johnson DE. 1978. Ionic milieu of bovine and ovine rumenas affected by diet. J Dairy Sci 61:315–323.
Bergen WG. 1972. Rumen osmolality as a factor in feed intake control of sheep. J Anim Sci 34:1054–1060.
Brown CE, Pezeshkia SR, DeLaune RD. 2006. The effects of salinity and soil drying on nutri-ent uptake and growth of Spartina alterniflora in a simulated tidal system. Environ Exp Bot58:140–148.
REFERENCES 717

Carter RR, Grovum WL. 1990. A review of the physiological significance of hypertonic bodyfluids on feed intake and ruminal function: Salivation, motility and microbes. J Anim Sci68:2811–2832.
Craig GF, Bell DT, Atkins CA. 1991. Nutritional characteristics of selected species of Acaciagrowing in naturally saline areas of Western Australia. Aust J Exp Agric 31:341–345.
Daoud S, Harrouni MC, Bengueddour R. 2001. Biomass production and ion compositionof some halophytes irrigated with different seawater dilutions. First InternationalConference on Saltwater Intrusion and Coastal Aquifers—Monitoring, Modeling, andManagement. Essaouira, Morocco, April 23–25, 2001.
Dobson A, Sellers AF, Gatewood VH. 1976. Absorption and exchange of water across rumenepithelium. Am J Physio 231:1588–1594.
Eadie JM, Hyldgaard-Jensen SO, Mann RS. 1970. Observations on the microbiology and bio-chemistry of the rumen in cattle. Br J Nutr 24:157.
El-Bassosy AA. 1983. A study of the nutritive value of some range plants from El-Saloomand Mersa Mattroh. Ph.D. Thesis, Faculty of Agriculture, Ain Shams University, Egypt.
El Shaer MH, Attia-Ismail SA. 2002. Halophytes as animal feeds: Potentiality, constraints, andprospects. International Symposium on Optimum Utilization in Salt Affected Ecosystems inArid and Semi-arid Regions, Cairo, Egypt, April 8–11, 2002.
Fahmy AA. 1998. Nutritional studies on halophytes and agricultural wastes as feed sup-plements for small ruminants in Sinai. Ph.D. Thesis, Cairo University, Egypt.
Fahmy AA, Attia-Ismail SA, Fayed AM. 2001. Effect of Monensin on salt plant utilization andsheep performance. Egyptian J Nutr Feeds 4:581–590.
FAO. 1988. Salt-affected soils and their management. By Abrol IP, Yadav JSP, and MassoudFI. FAO Soils Bull. 39.
FAO. 2007. http://www.fao.org/ag/agl/agll/wrb/wrbmaps/htm/
Farid MFA. 1989. Water and minerals problems of the dromedary camel (an overview).Options Mediterraneenn—Ser Semin 2:111–124.
Forbes JM. 1995. Voluntary food intake and diet selection in farm animals. CAB International,Wallingford, Oxon OX 108DE, UK.
Gabr MG. 2002. First experience of Matrouh Resource Management Project in salt bush utilizationfor animal feeding. International Symposium on Optimum Utilization in Salt AffectedEcosystems in Arid and Semi Arid Regions, Cairo, Egypt, April 8–11, 2002, pp. 419–425.
Gihad EA. 1993. Utilization of high salinity tolerant plants and saline water by desert animals.In: Lieth H, Al-Masoom A, editors. Towards the rational use of high salinity tolerant plant,Klumer Academic Publishers. Dordrecht. T. Vs Vol. 1:445–447.
Glenn EP, Brown JJ, Blumwald E. 1999. Salt tolerance and crop potential of halophytes. CritRev Plant Sci 18:227–255.
Gorham J. 1996. Mechanisms of salt tolerance of halophytes. In: Choukr-Allah R, MalcolmCV, Hamdy A, editors. Halophytes and Biosaline Agriculture. New York: MarcelDekker, pp. 31–53.
Haenlein GFW, Ramirez RG. 2007. Potential mineral deficiencies on arid rangelands for smallruminants with special reference to Mexico. Small Ruminant Res 68:35–41.
Harrison DJ, Beever DE, Thomson DJ, Osbourn DF. 1975. Manipulation of rumen ermentationin sheep by increasing the rate of flow of water from the rumen. J Agric Sci, Cambridge85:93–101.
718 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS

Hogan JP. 1961. The absorption of ammonia through the rumen of the sheep. Aust J Biol Sci14:448.
Hutchings SS. 1965. Grazing management of salt-desert shrub ranges in the Western UnitedStates. Proc 9th Int Grass Cong 2:1619–1625.
Jaradat AA. 2003. Halophytes for sustainable biosaline. In: Al Sharhan AA, et al., editors.Farming Systems in the Middle East. Rotterdam, The Netherlands: A.A. Baikema/Swets& Zeitlinger. Desertification for the third millennium, pp. 187–203.
Jordan FL, Robin-Abbott MM, Raina M, Glenn P. 2002. A comparison of chelator-facilitatedmetal uptake by a halophyte and a glycophyte. Environ Toxicol Chem 21:2698–2704.
Kato S, Sasaki Y, Tsuda T. 1979. Food intake and rumen osmolality in the sheep. AnnRecherches Veterinaire 10:229–230.
Kewan KZ. 2003. Studies on camel nutrition. Ph.D. Thesis, Faculty of Agriculture, AlexandriaUniversity, Egypt.
Khan MA. 2001. Experimental assessment of salinity tolerance of Ceriops tagal seedlings andsaplings from the Indus delta. Pakistan Aquat Bot 70:259–268.
Khattab IMA. 2007. Studies on halophytic forages as sheep fodder under arid and semi aridconditions in Egypt. Ph.D. Thesis. Alexandria University.
Kraidees MS, Abouheif MA, Al-Saiady MY, Tag-Eldin A, Metwally H. 1998. The effect ofdietary inclusion of halophyte Salicornia bigelovii Torr. on growth performance andcarcass characteristics of lambs. Anim Feed Sci Technol 76:149–159.
Lopez S, Hovell B, Macleod NA. 1994. Osmotic pressure, water kinetics and volatile fatty acidabsorption in the rumen of sheep sustained by intragastric infusion. British J Nutr71:153–168.
MacFarlane WV, Morris RJH, Howard B. 1963. Turnover and distribution of water in desertcamels, sheep, cattle and kangaroo. Nature London 197:270–271.
Marai IFM, Habeeb AA, Kamal TH. 1995. Response of livestock to excess sodium intake. In:Phillips CJC, Chiy PC, editors. Sodium in Agriculture. Canterbury: ChalcombePublications, pp. 173–180.
Masters DG, Benes SE, Norman HC. 2007. Biosaline agriculture for forage and livestock pro-duction. Agric Ecosystems Environ 119:234–248.
NRC. 1981. Nutrient Requirements of Goats. Washington, DC: National Research Council,National Academic Press.
NRC. 1985. Nutrient Requirements of Sheep. Washington, DC: National Research Council,National Academic Press.
Parida AK, Das AB, Maritta B. 2004. Effect of salt on growth, ion accumulation, photosyn-thesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees—Struct Funct18:167–174.
Puri JP, Garg SK. 2001. Effect of osmotic agent supplementation in the diet of buffalo on somerumen functions and blood electrolytes. Indian J Anim Sci 71(10):927–931.
Reboreda R, Cacador I. 2007. Halophyte vegetation influences in salt marsh retention capacityfor heavy metals. Environ Pollut 146:147–154.
Remond D, Chaise JP, Delval E, Poncet C. 1993. Net transfer of urea and ammonia across theruminal wall of sheep. J Amin Sci 71:2785–2792.
Retana J, Parker DR, Amrhein C, Page AL. 1993. Growth and trace element concentrations offive plant species grown in a highly saline soil. J Environ Sci 22:805–811.
REFERENCES 719

Shawkat SM, Khatab IM, Borhami BE, El-Shazly KA. 2001. Performance of growing goatsfed halophytic pasture with different energy sources. Egyptian J Nutr Feeds 4:215–264.
Swingle RS, Glenn EP, Squires V. 1996. Growth performance of lambs fed mixed diets con-taining halophyte ingredients. Anim Feed Sci Technol 63:137–148.
Szabolcs I. 1974. Salt Affected Soils in Europe. The Hague: Martinus Nijhoff.
Ternouth JH, Beattie A. 1971. Studies of the food intake of sheep at a single meal. Br J Nutr25:153.
Underwood EJ, Suttle NF. 1999. The Mineral Nutrition of Livestock. New York: CABIPublishing.
Vercoe TK. 1986. Fodder potential of selected Australian tree species. In: Australian Acacias inDeveloping Countries, Proc. Of an international workshop held at the Forestry TrainingCenter Gympie, Qld, Australia, 4–7 August, 1986 (J.W. Turnbull, Ed.), pp. 95–100.ACIAR Proceedings no. 16, Australian Center for International Research, Canberra.
Verde T, Shephard RJ, Corey P, Moore R. 1982. Sweat composition in exercise and in heat.J Appl Physiol 53:1540–1545.
Walker BH. 1980. A review of browse and role in livestock production in Southern Africa. In:Le Houerou HN, editor. Browse in Africa, the current state of knowledge. ILCA AddisAbaba, Ehtiopia.
Warner ACI, Stacy BD. 1965. Solutes in the rumen of the sheep. Quart J Exp Physiol50:169–184.
Warner ACI, Stacy BD. 1977. Influence of ruminal and plasma osmotic pressure on salivarysecretion in sheep. Q J Expe Physiol 62:133–142.
Warren BE, Bunny CI, Bryant ER. 1990. A preliminary examination of the nutritive value offour saltbush (Atriplex) species. Proc. Austr.
Warren BE, Casson T, Ryall DH. 1994. Production from grazing sheep on revegetated saltlandin Western Australia. In: Squires VR, Ayoub.
Wilson AD. 1974. Water requirements and water turnover of sheep grazing semi-arid pasturecommunities in New South Wales. Aust J Agric Res 25:339.
Windham L, Weis JS, Weis P. 2003. Uptake and distribution of metals in two dominant saltmarsh macrophytes, Spartina alterniflora (cordgrass) and Phragmites australis (commonreed). Estuar Coast Shelf Sci 56:63–72.
720 ROLE OF MINERALS IN HALOPHYTE FEEDING TO RUMINANTS
Related Documents











