Natural Haemozoin Induces Expression and Release of Human Monocyte Tissue Inhibitor of Metalloproteinase-1 Manuela Polimeni 1 , Elena Valente 1 , Daniela Ulliers 1 , Ghislain Opdenakker 2 , Philippe E. Van den Steen 2 , Giuliana Giribaldi 1. , Mauro Prato 1,3 * . 1 Dipartimento di Oncologia, Universita ` di Torino, Torino, Italy, 2 Laboratory of Immunobiology, Rega Institute, Catholic University of Leuven, Leuven, Belgium, 3 Dipartimento di Neuroscienze, Universita ` di Torino, Torino, Italy Abstract Recently matrix metalloproteinase-9 (MMP-9) and its endogenous inhibitor (tissue inhibitor of metalloproteinase-1, TIMP-1) have been implicated in complicated malaria. In vivo, mice with cerebral malaria (CM) display high levels of both MMP-9 and TIMP-1, and in human patients TIMP-1 serum levels directly correlate with disease severity. In vitro, natural haemozoin (nHZ, malarial pigment) enhances monocyte MMP-9 expression and release. The present study analyses the effects of nHZ on TIMP-1 regulation in human adherent monocytes. nHZ induced TIMP-1 mRNA expression and protein release, and promoted TNF-a, IL-1b, and MIP-1a/CCL3 production. Blocking antibodies or recombinant cytokines abrogated or mimicked nHZ effects on TIMP-1, respectively. p38 MAPK and NF-kB inhibitors blocked all nHZ effects on TIMP-1 and pro-inflammatory molecules. Still, total gelatinolytic activity was enhanced by nHZ despite TIMP-1 induction. Collectively, these data indicate that nHZ induces inflammation-mediated expression and release of human monocyte TIMP-1 through p38 MAPK- and NF- kB-dependent mechanisms. However, TIMP-1 induction is not sufficient to counterbalance nHZ-dependent MMP-9 enhancement. Future investigation on proteinase-independent functions of TIMP-1 (i.e. cell survival promotion and growth/ differentiation inhibition) is needed to clarify the role of TIMP-1 in malaria pathogenesis. Citation: Polimeni M, Valente E, Ulliers D, Opdenakker G, Van den Steen PE, et al. (2013) Natural Haemozoin Induces Expression and Release of Human Monocyte Tissue Inhibitor of Metalloproteinase-1. PLoS ONE 8(8): e71468. doi:10.1371/journal.pone.0071468 Editor: Emma H. Wilson, University of California, Riverside, United States of America Received January 29, 2013; Accepted June 29, 2013; Published August 14, 2013 Copyright: ß 2013 Polimeni et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Funding: This work was financed by intramural funds from Universita ` di Torino to G. Giribaldi, by funds from Mrs. Franca Squazza and from Societa ` Italiana di Biochimica (SIB) to M. Prato, and by Fund for Scientific Research (FWO-Vlaanderen) and Research Fund of the KULeuven (GOA/2012/017) to G. Opdenakker and P. Van den Steen. M. Prato holds a professorship granted by Universita ` di Torino and Azienda Sanitaria Locale-19 (ASL-19). P. Van den Steen holds a research professorship of the KULeuven. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Competing Interests: The authors have declared that no competing interests exist. * E-mail: [email protected] . These authors contributed equally to this work. Introduction As a consequence of the global eradication program launched by charity foundations [1], World Health Organization (WHO) officially registered in 2010 a decline in estimated malaria cases and deaths, with 655,000 deaths counted among more than 200 million clinical cases worldwide, of which 91% were due to Plasmodium falciparum [2]. Nevertheless, malaria remains an alarming emergency in developing countries, with the vast majority of cases occurring in the African Region (81%) and South-East Asia (13%) [2]. Thus it is imperative to investigate new anti-malarial drugs for primary and adjuvant therapy [3] and identify new affordable markers for early diagnosis of malaria. Human matrix metalloproteinases (MMPs) are a family of proteolytic enzymes involved in wide variety of biological functions including modulation of inflammatory response, disruption of inter-endothelial tight junctions, and degradation of sub-endothe- lial basal lamina [4–7]. As such, they are good candidate molecules and indeed there is growing evidence that MMPs play critical roles in malaria in both animal and human disease models (see [8–10] for more extensive reviews). Notably, malarial pigment (nHZ, natural haemozoin), a waste product of haemoglobin digestion by Plasmodium parasites, induces MMP-9 release from human monocytes [11–14] and endothelial cells [9,15–16], and synthetic HZ (sHZ) interacts with proMMP-9 priming its activation by MMP-3 [17]. Endogenous inhibitors of MMPs (TIMPs, tissue inhibitors of metalloproteinases) represent one mode of MMP regulation [18]; however, their involvement in malaria has been scarcely investi- gated, and their role remains debated. A few lines of evidence from animal and human models support the involvement of TIMPs in malaria. CM-sensitive mice infected with P. berghei ANKA display increased mRNA expression of TIMP-1 in the brain, and both TIMP-1 and -3 mRNA is increased in the liver and spleen, whilst mRNA levels of TIMP-2 and -4 remain unchanged [19]. Increased serum levels of TIMP-1 are also found in Rhesus macaques (Macaca mulatta) experimentally infected with P. coatneyi, a simian malaria parasite that closely mimics the biological characteristics of P. falciparum and replicates the multisystemic dysfunction of human severe malaria [20]. Human patients with severe or uncomplicated malaria have higher serum TIMP-1 levels compared to healthy controls suggesting TIMP-1 may be a valuable marker of disease severity [21]. However, the cellular source of TIMP-1 and the mechanisms underlying TIMP-1 enhancement are as of yet unidentified. Additionally, it is imperative to assess whether increased CM-associated TIMP-1 levels are sufficient to counterbalance nHZ-enhanced MMP-9. PLOS ONE | www.plosone.org 1 August 2013 | Volume 8 | Issue 8 | e71468

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Natural Haemozoin Induces Expression and Release ofHuman Monocyte Tissue Inhibitor of Metalloproteinase-1Manuela Polimeni1, Elena Valente1, Daniela Ulliers1, Ghislain Opdenakker2, Philippe E. Van den Steen2,
Giuliana Giribaldi1., Mauro Prato1,3*.
1Dipartimento di Oncologia, Universita di Torino, Torino, Italy, 2 Laboratory of Immunobiology, Rega Institute, Catholic University of Leuven, Leuven, Belgium,
3Dipartimento di Neuroscienze, Universita di Torino, Torino, Italy
Abstract
Recently matrix metalloproteinase-9 (MMP-9) and its endogenous inhibitor (tissue inhibitor of metalloproteinase-1, TIMP-1)have been implicated in complicated malaria. In vivo, mice with cerebral malaria (CM) display high levels of both MMP-9 andTIMP-1, and in human patients TIMP-1 serum levels directly correlate with disease severity. In vitro, natural haemozoin (nHZ,malarial pigment) enhances monocyte MMP-9 expression and release. The present study analyses the effects of nHZ onTIMP-1 regulation in human adherent monocytes. nHZ induced TIMP-1 mRNA expression and protein release, andpromoted TNF-a, IL-1b, and MIP-1a/CCL3 production. Blocking antibodies or recombinant cytokines abrogated or mimickednHZ effects on TIMP-1, respectively. p38 MAPK and NF-kB inhibitors blocked all nHZ effects on TIMP-1 and pro-inflammatorymolecules. Still, total gelatinolytic activity was enhanced by nHZ despite TIMP-1 induction. Collectively, these data indicatethat nHZ induces inflammation-mediated expression and release of human monocyte TIMP-1 through p38 MAPK- and NF-kB-dependent mechanisms. However, TIMP-1 induction is not sufficient to counterbalance nHZ-dependent MMP-9enhancement. Future investigation on proteinase-independent functions of TIMP-1 (i.e. cell survival promotion and growth/differentiation inhibition) is needed to clarify the role of TIMP-1 in malaria pathogenesis.
Citation: Polimeni M, Valente E, Ulliers D, Opdenakker G, Van den Steen PE, et al. (2013) Natural Haemozoin Induces Expression and Release of Human MonocyteTissue Inhibitor of Metalloproteinase-1. PLoS ONE 8(8): e71468. doi:10.1371/journal.pone.0071468
Editor: Emma H. Wilson, University of California, Riverside, United States of America
Received January 29, 2013; Accepted June 29, 2013; Published August 14, 2013
Copyright: � 2013 Polimeni et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work was financed by intramural funds from Universita di Torino to G. Giribaldi, by funds from Mrs. Franca Squazza and from Societa Italiana diBiochimica (SIB) to M. Prato, and by Fund for Scientific Research (FWO-Vlaanderen) and Research Fund of the KULeuven (GOA/2012/017) to G. Opdenakker and P.Van den Steen. M. Prato holds a professorship granted by Universita di Torino and Azienda Sanitaria Locale-19 (ASL-19). P. Van den Steen holds a researchprofessorship of the KULeuven. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
. These authors contributed equally to this work.
Introduction
As a consequence of the global eradication program launched
by charity foundations [1], World Health Organization (WHO)
officially registered in 2010 a decline in estimated malaria cases
and deaths, with 655,000 deaths counted among more than 200
million clinical cases worldwide, of which 91% were due to
Plasmodium falciparum [2]. Nevertheless, malaria remains an
alarming emergency in developing countries, with the vast
majority of cases occurring in the African Region (81%) and
South-East Asia (13%) [2]. Thus it is imperative to investigate new
anti-malarial drugs for primary and adjuvant therapy [3] and
identify new affordable markers for early diagnosis of malaria.
Human matrix metalloproteinases (MMPs) are a family of
proteolytic enzymes involved in wide variety of biological functions
including modulation of inflammatory response, disruption of
inter-endothelial tight junctions, and degradation of sub-endothe-
lial basal lamina [4–7]. As such, they are good candidate molecules
and indeed there is growing evidence that MMPs play critical roles
in malaria in both animal and human disease models (see [8–10]
for more extensive reviews). Notably, malarial pigment (nHZ,
natural haemozoin), a waste product of haemoglobin digestion by
Plasmodium parasites, induces MMP-9 release from human
monocytes [11–14] and endothelial cells [9,15–16], and synthetic
HZ (sHZ) interacts with proMMP-9 priming its activation by
MMP-3 [17].
Endogenous inhibitors of MMPs (TIMPs, tissue inhibitors of
metalloproteinases) represent one mode of MMP regulation [18];
however, their involvement in malaria has been scarcely investi-
gated, and their role remains debated. A few lines of evidence from
animal and human models support the involvement of TIMPs in
malaria. CM-sensitive mice infected with P. berghei ANKA display
increased mRNA expression of TIMP-1 in the brain, and both
TIMP-1 and -3 mRNA is increased in the liver and spleen, whilst
mRNA levels of TIMP-2 and -4 remain unchanged [19].
Increased serum levels of TIMP-1 are also found in Rhesus
macaques (Macaca mulatta) experimentally infected with P. coatneyi,
a simian malaria parasite that closely mimics the biological
characteristics of P. falciparum and replicates the multisystemic
dysfunction of human severe malaria [20]. Human patients with
severe or uncomplicated malaria have higher serum TIMP-1 levels
compared to healthy controls suggesting TIMP-1 may be a
valuable marker of disease severity [21]. However, the cellular
source of TIMP-1 and the mechanisms underlying TIMP-1
enhancement are as of yet unidentified. Additionally, it is
imperative to assess whether increased CM-associated TIMP-1
levels are sufficient to counterbalance nHZ-enhanced MMP-9.
PLOS ONE | www.plosone.org 1 August 2013 | Volume 8 | Issue 8 | e71468

Endothelial cells and monocytes are both producers of inducible
TIMP-1 protein [22–23], and their phenotype and functions can
be directly affected by malarial products such as infected red blood
cells (IRBCs) and nHZ [9]. However, it is unlikely that
endothelium is the TIMP-1 source in malaria, as neither IRBCs
nor nHZ affect TIMP-1 protein release from human microvas-
cular endothelial cells [15–16].
Here we investigate the in vitro effects of nHZ on human
monocyte TIMP-1 gene expression and protein release, the
responsible soluble mediators, the signaling pathways involved,
and the net gelatinolytic activity resulting from the altered MMP-
9/TIMP-1 balance.
Materials and Methods
MaterialsAll materials were from Sigma-Aldrich, St Louis, MO, unless
otherwise stated. Sterile plastics were from Costar, Cambridge,
UK; Percoll was from Pharmacia, Uppsala, Sweden; Diff-Quik
parasite stain was from Baxter Dade AG, Dudingen, Switzerland;
Panserin 601 monocyte medium was from PAN Biotech,
Aidenbach, Germany; ELISA kits for hTNF-a and hIL-1b assays
were from Cayman, Ann Arbor, MI; blocking anti-hTNF-a/IL-1bantibodies and rhTNF-a/IL-1b were from Merck, Darmstadt,
Germany; ELISA kit for hMIP-1a/CCL3, anti-hMIP-1a/CCL3
blocking antibodies and rhMIP-1a/CCL3 were from R&D
Systems, Minneapolis, MN; p38 MAPK inhibitor SB203580 was
from Cell Signaling Technology, Danvers, MA; ELISA kits for
hMMP-9 and hTIMP-1 were from RayBiotech, Norcross, GA;
cell culture medium RPMI, DQ-gelatin, TRIzol, M-MLV, oligo-
dT, sense and anti-sense primers, Platinum Taq DNA Polymerase
were from Invitrogen, Carlsbad, CA; DNA-free kit was from
Ambion, Austin, TX; Beacon Designer 7.0 software was from
Premier Biosoft International, Palo Alto, CA; dNTPs were from
Applied Biosystem, Foster City, CA; anti-hTIMP-1 (sc-21734)
monoclonal antibodies were from Santa Cruz Biotechnology,
Santa Cruz, CA; iCycler iQ Real Time Detection System
Software version 3.0 and electrophoresis reagents were from
Bio-Rad Laboratories, Hercules, CA; computerized densitometer
Chemidoc was from Biorad, Hercules, CA; Synergy HT
microplate reader was from Bio-Tek Instruments, Winooski, VT;
recombinant proMMP-9 and MMP-9 were produced as previ-
ously described [24].
Cultivation of Plasmodium falciparum and Isolation ofnHZPlasmodium falciparum parasites (Palo Alto strain, Mycoplasma-
free, LPS-free) were kept in culture as described [25]. After
centrifugation at 5,000g on a discontinuous Percoll-mannitol
density gradient, nHZ was collected from the 0–40% interphase.
nHZ was washed five times with 10 mM HEPES (pH 8.0)
containing 10 mM mannitol at 4uC and once with phosphate-
buffered saline (PBS). nHZ was treated with DNase to remove any
adhering nuclear material as previously described [26]. nHZ was
stored at 20% (v/v) in PBS at 220uC or immediately used for
opsonization and phagocytosis.
Preparation and Handling of MonocytesHuman monocytes were separated by Ficoll centrifugation [11]
from freshly collected buffy coats discarded from blood donations
by healthy adult donors of both sexes provided by the local blood
bank (AVIS, Associazione Volontari Italiani Sangue, Torino,
Italy). Separated lympho/monocytes were resuspended in RPMI
medium and plated on six-well plates. Each well received 86106
cells. The plates were incubated in a humidified CO2/air-
incubator at 37uC for 60 min. Thereafter, non-adherent cells
were removed by three washes with RPMI and remaining
adherent cells (,16106 monocytes/well) were again incubated
at 37uC overnight. Shortly before starting phagocytosis, wells were
washed with RPMI and Panserin 601 monocyte medium was
added (2 ml/well).
Pre-selection of NF-kB-non-activated Monocytes by FACSAnalysis and Real Time RT-PCR
Before starting experiments, a pre-selection of cell populations
was taken as a precautionary measure, as previously described
[13]. Briefly, cell cultures isolated through Ficoll separation were
analyzed by flow cytometry. Only cell populations showing at least
70% monocytes were used for following experiments. Additionally,
in order to avoid the use of NF-kB pre-activated monocytes, cells
were analyzed by Real Time RT-PCR: in each cell preparation a
cell aliquot was stimulated or not with LPS (1 mg/ml) for 4 h, and
TNF-a RNA production measured in lysates by Real Time RT-
PCR. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was
used as housekeeping gene. Only unstimulated monocyte popu-
lations (NF-kB-non-activated cells) showing at least a 3-PCR-
cycles gap of cDNA amplification between controls and LPS-
stimulated cells were used for the subsequent experiments.
Cell Culturing Experimental Conditions: Phagocytosis ofnHZ or Latex Particles and Treatment with BlockingAntibodies, Recombinant Pro-inflammatory Molecules, orCell Signalling Chemical Inhibitors
Phagocytosis assay was performed as previously described [27].
Briefly, nHZ (120 nmoles HZ haem, an amount comparable to
50 ml trophozoites on haem content basis) and 50 ml amine-
modified latex particles (2.5% solids, diameter 0.105 mm) were
added to each well of a six-well plate containing the same amounts
of human adherent monocytes (,16106 cells/well). nHZ and
latex particles were opsonized with fresh autologous serum. After
opsonization, all phagocytic meals were suspended in Panserin 601
monocyte medium. The plates were centrifuged at low speed for
5 s to start phagocytosis and incubated in a humidified CO2/air-
incubator at 37uC for 2 h - a time period maximizing phagocytosis
but not sufficient to induce haem-oxygenase-mediated degradation
of ingested haem [28]. Cells were checked by optical microscopy:
as an average, nHZ-containing monocytes were 25–35% among
the total cells, a percentage similar to in vivo levels measured in
patients with severe malaria showing high parasitaemia [29].
Additionally, the amount of nHZ phagocytosed by monocytes was
quantified by luminescence: as an average, each monocyte
ingested nHZ equivalent to ,8–10 trophozoites in term of
ingested haem, in line with previous results from our group [30].
Thereafter, non-ingested nHZ and latex particles were removed
by four washes with RPMI. The plates were then incubated in
Panserin 601 medium in a humidified CO2/air-incubator at 37uCfor the indicated times.
In selected experiments, unfed and nHZ-fed monocytes were
incubated for the indicated times with 30 ng/ml anti-hTNF-a,
anti-hIL-1b or anti-hMIP-1a/CCL3 blocking antibodies; 20 ng/
ml rhTNF-a, rhIL-1b or rhMIP-1a/CCL3; 10 mM SB203580,
15 mM quercetin, 10 mM artemisinin, or 10 mM parthenolide,
dissolved in DMSO (final solvent concentration less than 0.1%
v/v).
Complimentary co-culturing experiments with unfed and nHZ-
fed monocytes were also performed using six-well plate Transwell
systems with 0.4 mm of porosity (see Figure S2A-B). Briefly,
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 2 August 2013 | Volume 8 | Issue 8 | e71468

monocytes (0,56106 cells/transwell) were seeded in transwells
(provisionally placed in cell-free wells) and fed with nHZ for 2h.
Thereafter, transwells with nHZ-fed monocytes were moved into
new wells containing unfed monocytes (16106 cells/well) and
incubated for 2 h, a time previously shown to be sufficient to
induce early production of TNF-a, IL-1b, MIP-1a/CCL3 from
nHZ-fed cells [25,31]. As negative and positive controls, respec-
tively, unfed monocytes were also left unfed or fed with nHZ for
2 h. Then nHZ and transwells with nHZ-fed monocytes were
removed. After washing and adding fresh medium, cells were
incubated in Panserin 601 medium in a humidified CO2/air-
incubator for 24 h before collecting supernatants for subsequent
analyses.
Cytotoxicity Studies by Lactate Dehydrogenase AssayThe potential cytotoxic effects of phagocytic meals (nHZ and
latex) and treatments (anti-hTNF-a, anti-hIL-1b and anti-hMIP-
1a/CCL3 antibodies; rhTNF-a, rhIL-1b and rhMIP-1a/CCL3;
SB203580, quercetin, artemisinin, and parthenolide) were mea-
sured as the release of lactate dehydrogenase (LDH) from cells into
the extracellular medium (see Figure S1A–B).
Briefly, human adherent monocytes were left unfed or fed with
phagocytic meals for 2 h; after washings, unfed and fed cells were
incubated for 24 h in the presence or in the absence of each one of
the above mentioned treatments. Then, 1 ml of cell supernatants
was collected and centrifuged at 13000 g for 2 min. Cells were
washed with fresh medium, detached with trypsin/ethylenedi-
aminetetraacetic acid (EDTA) (0.05/0.02% v/v), washed with
PBS, resuspended in 1 ml of TRAP (82.3 mM triethanolamine,
pH 7.6), and sonicated on ice with a 10 s burst. 5 ml of cell lysates
and 50 ml of cell supernatants were diluted with TRAP and
supplemented with 0.5 mM sodium pyruvate and 0.25 mM
NADH (300 mL as a final volume) to start the reaction. The
reaction was followed measuring the absorbance at 340 nm (37uC)
with a microplate reader. Both intracellular and extracellular
enzyme activities were expressed as mmol of oxidized NADH/
min/well. Finally, cytotoxicity was calculated as the net ratio
between extracellular and total (intracellular+extracellular) LDH
activities. Cell viability was calculated as the net ratio between
intracellular and total LDH activities.
Measurement of TIMP-1 mRNA Levels by Real Time RT-PCR
After the end of phagocytosis, monocytes (16106 cells/well)
were further incubated with Panserin 601 monocyte medium in a
humidified CO2/air-incubator at 37uC for 15 h. Total cellular
RNA from 26106 cells (contained in 2 wells) was isolated from
monocytes by TRIzol, according to the manufacturer’s instruc-
tions, and eluted in 20 ml diethyl pyrocarbonate water. To remove
any contaminating DNA, RNA was treated with Ambion’s DNA-
free kit. Subsequently, 6 mg of RNA were reverse transcribed into
single-stranded cDNA using M-MLV (200 U/ml final concentra-
tion) and oligo-dT (25 mg/ml final concentration). Real Time RT-
PCR analysis was performed with the iCycler Instrument and the
iCycler iQ Real Time Detection System Software version 3.0 as
previously described [11]. For TIMP-1 (GenBank accession no.
BC000866) primers (forward: 59-AGA CGG CCT TCT GCA
ATT CC-39, reverse: 59-GCT GGT ATA AGG TGG TCT GGT
T-39), oligonucleotide sequences were identified using Beacon
Designer Software package and designed to be intron-spanning
allowing the differentiation between cDNA and genomic DNA-
derived PCR products. GAPDH (GenBank accession no.
BC020308) was used as housekeeping gene; primer sequences
were from the Bio-Rad library: forward: 59-GAA GGT GAA
GGT CGG AGT-39 and reverse: 59-CAT GGG TGG AAT CAT
ATT GGA A-39. For each 25 ml PCR reaction mix: 1 ml cDNA
(corresponding to 105 cells); 1.0 ml sense primer (10 mM); 1.0 ml
anti-sense primer (10 mM); 0.5 ml dNTP (10 mM); 1.5 ml MgCl2(50 mM); 1.25 U Platinum Taq DNA Polymerase; 2.5 ml Buffer
(10x); 1.7 ml SYBR Green (stock 1:10,000); and 14.55 ml PCR-
grade water were mixed together. DNA polymerase was pre-
activated for 2 min at 94uC. TIMP-1 amplification was performed
by 50 cycles with denaturation at 54uC for 30 s, annealing at 57uCfor 40 s and extension at 72uC for 30 s; GAPDH amplification
was performed by 40 cycles with denaturation at 94uC for 30 s,
annealing at 65uC for 30 s and extension at 72uC for 30 s for
GAPDH. Relative quantification for TIMP-1, expressed as fold
variation over untreated control cells, was calculated with the
efficiency-corrected quantification model [32] after determining
the difference between CT of the given gene A (TIMP-1) and that
of the calibrator gene B (GAPDH). CT values are means of
triplicate measurements. To validate the use of the method we
tested serial dilutions of cDNA from monocytes, stimulated for
15 h by 20 ng/ml rhTNF-a. The specificity of PCR was
confirmed by melt curve analysis. The melting temperatures for
each amplification product were 87.4uC for TIMP-1 and 86.5uCfor GAPDH.
Assay of TIMP-1 Protein Levels by Western BlottingAfter the end of phagocytosis, monocytes (16106 cells/well)
were further incubated with Panserin 601 monocyte medium in a
humidified CO2/air-incubator at 37uC for 24 h. Thereafter, 15 ml
of cell supernatants were separated on a 12% polyacrylamide
denaturing gel, transferred to a polyvinylidene difluoride mem-
brane, and probed with monoclonal anti-TIMP-1 antibodies.
After staining with secondary anti-mouse HRP-conjugated anti-
bodies, bands were visualized by ECL staining. Densitometric
analysis of the bands considered to reflect total protein levels was
performed using a computerized densitometer with protein levels
presented in relative units compared to background.
Measurement of TNF-a, IL-1b, MIP-1a/CCL3, MMP-9 andTIMP-1 Production by ELISA
After the end of phagocytosis, monocytes (16106 cells/well)
were further incubated with Panserin 601 monocyte medium in a
humidified CO2/air-incubator at 37uC up to 24 h. Cell superna-
tants were collected 6 h, 15 h and 24 h after phagocytosis. The
levels of soluble TNF-a, IL-1b, MIP-1a/CCL3, MMP-9 and
TIMP-1 were assayed in 100 ml of monocyte supernatants by
specific ELISA. Standard calibration curves were generated with
rhTNF, rhIL-1b, rhMIP-1a/CCL3, rhMMP-9 and rhTIMP-1
according to the manufacturer’s instructions.
Assay of Total Gelatinolytic Activity by a FluorogenicGelatin Conversion Assay
After phagocytosis, monocytes (16106 cells/well) were further
incubated with Panserin 601 monocyte medium in a humidified
CO2/air-incubator at 37uC for 24 h. Thereafter, the total
gelatinolytic activity [the net overall activity of gelatinases
(MMP-2 and MMP-9) and their inhibitors (TIMP-2 and TIMP-
1, respectively)], was measured in cultured supernatants using DQ-
gelatin as recently described [33]. Briefly, DQ-gelatin was
dissolved at a stock concentration of 1 mg/ml in pure water.
Serial dilutions of the samples were prepared in a black microtiter
plate, and DQ-gelatin was added at a final dilution of 2.5 mg/ml in
50 mM Tris/HCl pH 7.6, 150 mM NaCl, 5 mM CaCl2, 0.01%
Tween-20. Immediately thereafter, the plate was placed in the
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 3 August 2013 | Volume 8 | Issue 8 | e71468

fluorescence reader and fluorescence measured every 10 min for
2 h at 37uC (ex. 485 nm/em. 530 nm). As a standard, serial
dilutions of recombinant MMP-9 activated by MMP-3 were
included in each plate. The results were expressed as gelatinolytic
activity units per ml, with 1 unit corresponding to 37 ng/ml
activated rhMMP-9.
Statistical AnalysisFor each set of experiments, data are shown as means+SEM
(Real Time RT-PCR, densitometry, ELISA and fluorogenic
gelatin conversion assay) or as a representative image (Western
blotting) of three independent experiments with similar results. All
data were analyzed by a one-way Analysis of Variance (ANOVA)
followed by Tukey’s post-hoc test (software: SPSS 16.0 for
Windows, SPSS Inc., Chicago, IL).
Results
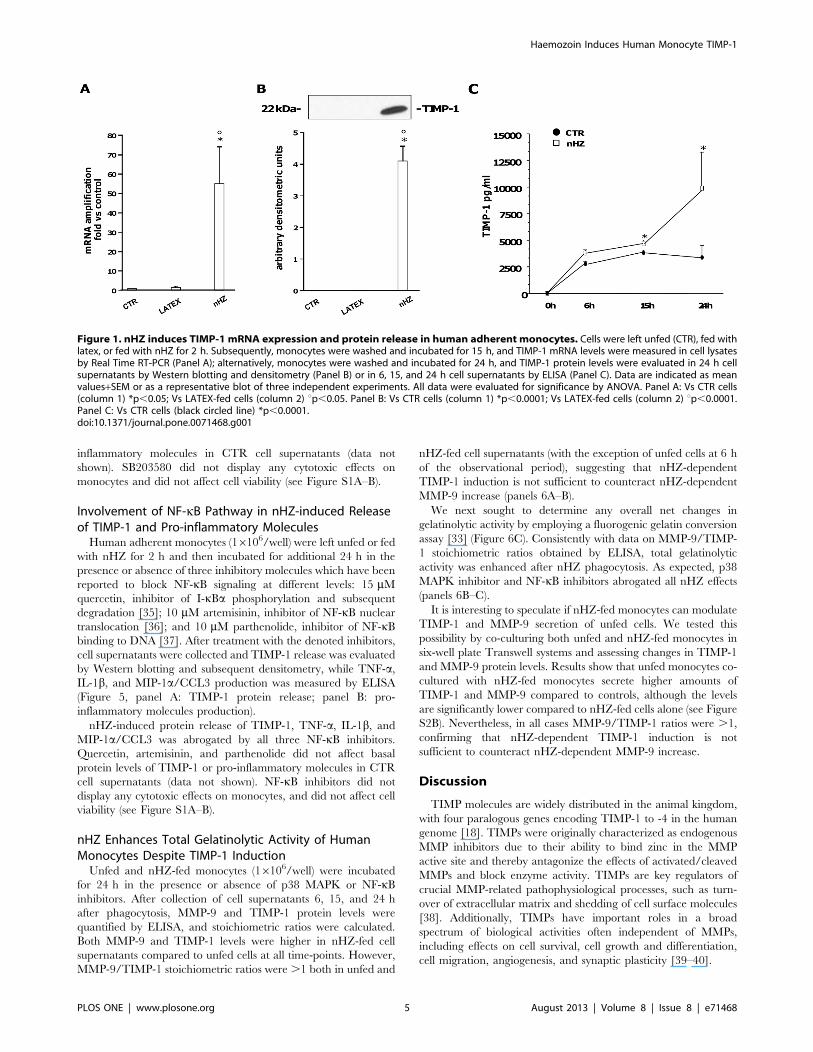
nHZ Induces TIMP-1 mRNA Expression and ProteinRelease in Human Adherent Monocytes
Human adherent monocytes (16106/well) were left unfed
(control cells, CTR), fed with latex as a control meal (LATEX-
FED), or fed with nHZ (nHZ-FED) for 2 h. At the end of
phagocytosis cells were washed and incubated for 15 h. Thereaf-
ter, mRNA was extracted and TIMP-1 expression was measured
by Real Time RT-PCR (Figure 1A). nHZ enhanced .50-fold
TIMP-1 mRNA expression compared to CTR, while latex
particles did not significantly alter basal TIMP-1 mRNA levels.
Alternatively, to evaluate TIMP-1 protein secretion, monocytes
were washed and incubated for 24 h after phagocytosis, with cell
supernatants collected at selected time-points (0, 6, 15, and 24 h)
during the incubation. TIMP-1 protein levels were evaluated by
Western blotting and densitometry (24 h end-point studies,
Figure 1B) and by ELISA (0–24 h time course studies,
Figure 1C). TIMP-1 protein release was induced de novo in nHZ-
FED cells, whereas Western blotting analysis did not detect
apparent levels of TIMP-1 in 24 h supernatants of CTR and
LATEX-FED cells. To verify these results, we used a more
sensitive ELISA assay to examine levels of TIMP-1 at earlier time-
points. Over time, CTR cells released low levels of TIMP-1
protein (up to ,3,5 ng/ml at the end of the 24 h observational
period), which were significantly increased upon nHZ treatment in
the nHZ-FED cells at 15 h and 24 h post phagocytosis (up to
,10,5 ng/ml at the end of the 24 h observational period).
Notably, nHZ and latex did not display any cytotoxic effects on
monocytes, and did not affect cell viability (see Figure S1A–B).
We next questioned whether nHZ-FED monocytes could
modulate TIMP-1 secretion from unfed cells. To address this
possibility, we co-cultured both unfed and nHZ-fed monocytes in
six-well plate Transwell systems with 0.4 mm of porosity, plating
unfed monocytes (16106 cells/well) at the bottom of the wells and
seeding nHZ-fed human adherent monocytes (0,56106 cells/well)
onto the inserts. Co-cultures were incubated for 2 h before
removal of the inserts. After washing, unfed monocytes were
further incubated for 24 h. Then, cell supernatants were collected
and analysed for TIMP-1 secretion. Results (see Figure S2A) show
that unfed monocytes co-cultured with nHZ-fed monocytes secrete
higher amounts of TIMP-1 than control cells, albeit significantly
lower in comparison to the amounts released by nHZ-fed cells
cultured alone.
Role of TNF-a, IL-1b, and MIP-1a/CCL3 in nHZ-dependentTIMP-1 Induction
Protein levels of TNF-a, IL-1b, and MIP-1a/CCL3 released
from CTR, LATEX-FED and nHZ-FED monocytes (16106/well)
were measured over 24 h post phagocytosis by ELISA (Figure 2).
During the observational period, CTR and LATEX-FED cells
released low amounts of all three pro-inflammatory molecules in a
time-dependent manner, reaching less than 5 ng/ml TNF-a and
less than 3 ng/ml IL-1b or MIP-1a/CCL3 at 24 h as evaluated
for both conditions. No significant differences between CTR and
LATEX-FED cells were found. After phagocytosis of nHZ, the
production of pro-inflammatory molecules was significantly higher
than in CTR or LATEX-FED cells, reaching ,15 ng/ml TNF-aand ,12 ng/ml IL-1b and MIP-1a/CCL3 at 15 h, as well as
,22 ng/ml TNF-a or IL-1b and ,15 ng/ml MIP-1a/CCL3 at
24 h.
As a next step, a double blocking and mimicking approach was
used to investigate the role of IL-1b, TNF-a, and MIP-1a/CCL3
(alone or combined) in the nHZ-dependent induction of TIMP-
1 mRNA expression and protein release (Figure 3).
After the end of phagocytosis CTR and nHZ-FED cells
(16106/well) were left untreated or treated with single or
combined doses (30 ng/ml) of anti-hTNF-a, anti-hIL-1b and
anti-hMIP-1a/CCL3 blocking antibodies for 15–24 h. We ana-
lyzed TIMP-1 mRNA expression and protein release by Real
Time RT-PCR in cell lysates (15 h) and Western blotting in cell
supernatants (24 h), respectively. The effects of nHZ on either
mRNA expression or protein release were reduced by single
blocking antibodies, and were fully abrogated by the combination
of all three antibodies (Panels 3A and 3C). Neither basal TIMP-
1 mRNA nor TIMP-1 protein levels in CTR cells were affected by
any treatments with blocking antibodies and by non-immune Ig
used as a negative control (data not shown).
Alternatively, after the end of phagocytosis CTR and nHZ-FED
cells were left untreated or treated with single or combined doses
(20 ng/ml) of rhTNF-a, rhIL-1b and rhMIP-1a/CCL3. Again,
TIMP-1 mRNA expression and protein release were analyzed as
previously indicated. The effects of nHZ on either mRNA
expression or protein release were only partially mimicked by
adding the single recombinant molecules to CTR cells, whereas
they were fully recapitulated by the simultaneous addition of all
three recombinant molecules (Panels 3B and 3D). None of
treatments with recombinant molecules further induced TIMP-1
mRNA or protein levels in nHZ-FED cells, possibly because a
plateau was already reached after nHZ phagocytosis (data not
shown).
Of note, blocking antibodies and recombinant molecules did not
display any cytotoxic effects on monocytes and did not affect cell
viability (see Figure S1A-B).
Involvement of p38 MAPK Pathway in nHZ-inducedRelease of TIMP-1 and Pro-inflammatory Molecules
Human adherent monocytes (16106/well) were left unfed or fed
with nHZ for 2 h and then incubated for an additional 24 h in the
presence or absence of p38 MAPK inhibitor SB203580 (10 mM)
[34]. Supernatants were collected and TIMP-1 release was
evaluated by Western blotting and subsequent densitometry,
whereas TNF-a, IL-1b, and MIP-1a/CCL3 production was
measured by ELISA (Figure 4, panel A: TIMP-1 protein release;
panel B: production of pro-inflammatory molecules).
Addition of SB203580 abrogated nHZ-induced protein release
of TIMP-1, TNF-a, IL-1b, and MIP-1a/CCL3. SB203580
treatment did not affect basal protein levels of TIMP-1 or pro-
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 4 August 2013 | Volume 8 | Issue 8 | e71468
inflammatory molecules in CTR cell supernatants (data not
shown). SB203580 did not display any cytotoxic effects on
monocytes and did not affect cell viability (see Figure S1A–B).
Involvement of NF-kB Pathway in nHZ-induced Releaseof TIMP-1 and Pro-inflammatory Molecules
Human adherent monocytes (16106/well) were left unfed or fed
with nHZ for 2 h and then incubated for additional 24 h in the
presence or absence of three inhibitory molecules which have been
reported to block NF-kB signaling at different levels: 15 mM
quercetin, inhibitor of I-kBa phosphorylation and subsequent
degradation [35]; 10 mM artemisinin, inhibitor of NF-kB nuclear
translocation [36]; and 10 mM parthenolide, inhibitor of NF-kB
binding to DNA [37]. After treatment with the denoted inhibitors,
cell supernatants were collected and TIMP-1 release was evaluated
by Western blotting and subsequent densitometry, while TNF-a,
IL-1b, and MIP-1a/CCL3 production was measured by ELISA
(Figure 5, panel A: TIMP-1 protein release; panel B: pro-
inflammatory molecules production).
nHZ-induced protein release of TIMP-1, TNF-a, IL-1b, and
MIP-1a/CCL3 was abrogated by all three NF-kB inhibitors.
Quercetin, artemisinin, and parthenolide did not affect basal
protein levels of TIMP-1 or pro-inflammatory molecules in CTR
cell supernatants (data not shown). NF-kB inhibitors did not
display any cytotoxic effects on monocytes, and did not affect cell
viability (see Figure S1A–B).
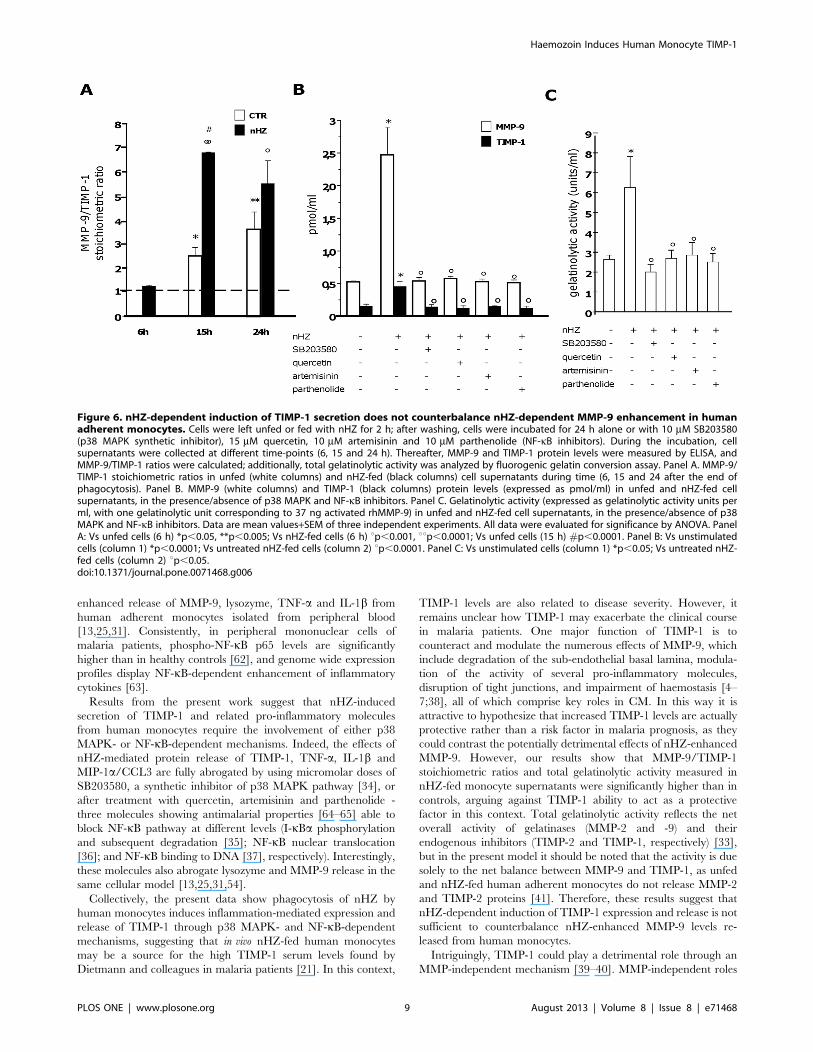
nHZ Enhances Total Gelatinolytic Activity of HumanMonocytes Despite TIMP-1 Induction
Unfed and nHZ-fed monocytes (16106/well) were incubated
for 24 h in the presence or absence of p38 MAPK or NF-kB
inhibitors. After collection of cell supernatants 6, 15, and 24 h
after phagocytosis, MMP-9 and TIMP-1 protein levels were
quantified by ELISA, and stoichiometric ratios were calculated.
Both MMP-9 and TIMP-1 levels were higher in nHZ-fed cell
supernatants compared to unfed cells at all time-points. However,
MMP-9/TIMP-1 stoichiometric ratios were .1 both in unfed and
nHZ-fed cell supernatants (with the exception of unfed cells at 6 h
of the observational period), suggesting that nHZ-dependent
TIMP-1 induction is not sufficient to counteract nHZ-dependent
MMP-9 increase (panels 6A–B).
We next sought to determine any overall net changes in
gelatinolytic activity by employing a fluorogenic gelatin conversion
assay [33] (Figure 6C). Consistently with data on MMP-9/TIMP-
1 stoichiometric ratios obtained by ELISA, total gelatinolytic
activity was enhanced after nHZ phagocytosis. As expected, p38
MAPK inhibitor and NF-kB inhibitors abrogated all nHZ effects
(panels 6B–C).
It is interesting to speculate if nHZ-fed monocytes can modulate
TIMP-1 and MMP-9 secretion of unfed cells. We tested this
possibility by co-culturing both unfed and nHZ-fed monocytes in
six-well plate Transwell systems and assessing changes in TIMP-1
and MMP-9 protein levels. Results show that unfed monocytes co-
cultured with nHZ-fed monocytes secrete higher amounts of
TIMP-1 and MMP-9 compared to controls, although the levels
are significantly lower compared to nHZ-fed cells alone (see Figure
S2B). Nevertheless, in all cases MMP-9/TIMP-1 ratios were .1,
confirming that nHZ-dependent TIMP-1 induction is not
sufficient to counteract nHZ-dependent MMP-9 increase.
Discussion
TIMP molecules are widely distributed in the animal kingdom,
with four paralogous genes encoding TIMP-1 to -4 in the human
genome [18]. TIMPs were originally characterized as endogenous
MMP inhibitors due to their ability to bind zinc in the MMP
active site and thereby antagonize the effects of activated/cleaved
MMPs and block enzyme activity. TIMPs are key regulators of
crucial MMP-related pathophysiological processes, such as turn-
over of extracellular matrix and shedding of cell surface molecules
[38]. Additionally, TIMPs have important roles in a broad
spectrum of biological activities often independent of MMPs,
including effects on cell survival, cell growth and differentiation,
cell migration, angiogenesis, and synaptic plasticity [39–40].
Figure 1. nHZ induces TIMP-1 mRNA expression and protein release in human adherent monocytes. Cells were left unfed (CTR), fed withlatex, or fed with nHZ for 2 h. Subsequently, monocytes were washed and incubated for 15 h, and TIMP-1 mRNA levels were measured in cell lysatesby Real Time RT-PCR (Panel A); alternatively, monocytes were washed and incubated for 24 h, and TIMP-1 protein levels were evaluated in 24 h cellsupernatants by Western blotting and densitometry (Panel B) or in 6, 15, and 24 h cell supernatants by ELISA (Panel C). Data are indicated as meanvalues+SEM or as a representative blot of three independent experiments. All data were evaluated for significance by ANOVA. Panel A: Vs CTR cells(column 1) *p,0.05; Vs LATEX-fed cells (column 2) up,0.05. Panel B: Vs CTR cells (column 1) *p,0.0001; Vs LATEX-fed cells (column 2) up,0.0001.Panel C: Vs CTR cells (black circled line) *p,0.0001.doi:10.1371/journal.pone.0071468.g001
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 5 August 2013 | Volume 8 | Issue 8 | e71468

It has only recently come to light that TIMPs are involved in
malaria disease state. Currently, our understanding of TIMPs in
malaria manifests from only a few studies showing dysregulated
TIMP levels in different in vivo and in vitro malaria models. In vivo,
brain and spleen TIMP-1 mRNA levels are increased in CM-
sensitive mice infected with P. berghei ANKA [19]. Further,
increased TIMP-1 serum levels are associated with severe malaria
in Rhesus macaques infected with P. coatneyi [20]. Moreover, in a
case-control study on Gabonese children, TIMP-1 serum levels
were significantly higher in malaria patients than in healthy
controls and correlated with disease severity, suggesting a potential
role for TIMP-1 as a diagnostic marker [21]. In vitro, P. falciparum-
IRBCs and nHZ promoted TIMP-2 but not TIMP-1 protein
release from human microvascular endothelial cells [15–16]. On
the other hand, in human monocytes, nHZ did not affect TIMP-2
production [41], whereas no data on TIMP-1 regulation by nHZ
were available so far.
The present work explores the effects of phagocytosed nHZ on
monocyte TIMP-1 expression and release, and identifies cellular
mechanisms and soluble mediators involved. Untreated cells, used
as negative controls, express low TIMP-1 mRNA levels and
secrete negligible TIMP-1 protein, consistent with previous reports
[23,42]. Interestingly, 15 h after phagocytosis of nHZ, TIMP-
1 mRNA expression is significantly enhanced, and higher TIMP-1
protein levels are detected in cell supernatants up to 24 h post
nHZ phagocytosis. The effect is specific for nHZ and not due to
phagocytosis per se since latex particles do not induce TIMP-1
expression and secretion.
These data give insight into the molecular mechanisms that
enable nHZ to alter the phenotype of human monocytes. Indeed,
nHZ was previously shown to induce gene expression of a large
number of pro-inflammatory molecules including cytokines (IL-1b,
TNF-a, IL-1RA) and chemokines (MIP-1a/CCL3, MIP-1b/
CCL4, MCP-1/CCL2, IL-8/CXCL8, ENA-78/CXCL5,
GROa/CXCL1, GROb/CXCL2, GROc/CXCL3) [43]. Also,
nHZ was reported to promote expression and release of MMP-9
[11–13] and lysozyme [24,31]. Notably, it has been proposed that
IL-1b, TNF-a and MIP-1a/CCL3 function as soluble mediators
for nHZ-dependent upregulation of either MMP-9 [11–12,14] or
lysozyme [24,31]. Here we show that these three pro-inflamma-
tory molecules are also involved in nHZ-induced expression and
release of TIMP-1. Either 15 or 24 h after phagocytosis of nHZ,
the levels of TNF-a, IL-1b, and MIP-1a/CCL3 in cell superna-
tants are significantly higher compared to controls. Treating with
blocking antibodies directed against these three molecules
abrogates the effects of nHZ on monocyte TIMP-1 mRNA
expression and protein release. Consistently, unfed cells treated
with recombinant TNF-a, IL-1b, and MIP-1a/CCL3 display
TIMP-1 mRNA levels in lysates and TIMP-1 protein levels in
supernatants similar to those of nHZ-fed cells. These results are in
line with previous data showing TIMP-1 induction by IL-1b [44].
Apparently, all three molecules are required in combination to
obtain nHZ-dependent TIMP-1 induction, since only partial
abrogation/emulation of nHZ effects is obtained with single doses
of blocking antibodies/recombinant molecules, whereas the effect
is fully reached after using combined doses. Nevertheless, nHZ
induces a plethora of pro-inflammatory molecules [43], some of
which correlate with TIMP-1 gene expression (i.e. MCP-1/CCL2,
MIP-1b/CCL4, and RANTES/CCL5) [45–46]. Therefore, the
involvement of additional cytokines and chemokines as redundant
soluble mediators for nHZ-promoted TIMP-1 expression and
release cannot be excluded.
We next wanted to investigate the molecular mechanisms
underlying nHZ-induced upregulation of TIMP-1 and related pro-
inflammatory molecules. It has been previously reported that IL-
1b induces both TIMP-1 and MMP-9 in monocytes and
monocytic leukemia cells [44,47]. In a separate study using
human monocytes, gene expression of TIMP-1, MMP-9 and
TNF-a, required p38 MAPK and NF-kB pathway activation [48–
49]. Interestingly, several in vitro and in vivo studies have raised the
possibility that either MAPK or NF-kB routes are involved in
malaria.
To date we understand very little about nHZ-dependent
activation of MAPKs. However, recent emerging evidence in a
murine malaria model shows that nHZ induces the activation of
p38 [49] and ERK1/2 [50–53] MAPKs, but does not induce the
activation of other signaling pathways such as the JNK-2/STAT
pathway [47–49]. Evidence from a human malaria model shows
Figure 2. nHZ enhances production of TNF-a, IL-1b, and MIP-1a/CCL3 by human adherent monocytes. Cells were left unfed(CTR), fed with latex particles (LATEX), or fed with nHZ (nHZ) for 2 h;therefore, production of TNF-a (upper panel), IL-1b (central panel), andMIP-1a/CCL3 (lower panel) was monitored in cell supernatants 0, 15 and24 h after the end of phagocytosis. Data are means+SEM of threeindependent experiments. Production of pro-inflammatory molecules isindicated as ng/ml. All data were evaluated for significance by ANOVA.Vs control cells (CTR) *p,0.0001; Vs LATEX-fed cells (LATEX) up,0.0001.doi:10.1371/journal.pone.0071468.g002
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 6 August 2013 | Volume 8 | Issue 8 | e71468

nHZ activates p38 MAPK signaling, similar to the murine model,
whilst nHZ does not appear to activate the ERK1/2 or JNK-1/2
signaling pathways in the human model [25,31,54]. Interestingly,
a study using syncytiotrophoblast cells shows nHZ-dependent
phosphorylation of ERK1/2, along with IRBC-dependent phos-
phorylation of JNK-1, with both events being causally related to
production of pro-inflammatory molecules (TNF-a, MIP-1a/
CCL3, IL-8/CXCL8) [55–56]. Moreover, our group recently
showed intravenous injection of nHZ in malaria-free mice induces
an inflammatory frame similar to that observed in experimental
malaria-associated acute respiratory distress syndrome, pointing to
nHZ as a prominent inflammatory virulent factor in lung
pathology [26]. On the other hand, Plasmodium falciparum
glycosylphosphatidylinositol (PfGPI) promotes phosphorylation of
all major MAPK routes, including p38, ERK1/2, and JNK-2/
STAT-1 [57–58]. Inhibition of PfGPI-dependent activation of
MAPKs decreases inflammatory responses and enhances phago-
cytic clearance of IRBCs in mice infected by Plasmodium berghei or
chabaudi chabaudi [59]. Future research defining the differential
roles of nHZ, IRBCs, and GPI in regulating MAPKs and
inflammation in malaria, both in vitro and in vivo, will be certainly
welcomed.
Along with nHZ-dependent regulation of MAPKs, emerging
evidence suggests malarial pigment plays a role in activating the
NF-kB pathway. In murine macrophages fed with nHZ or sHZ,
NF-kB activation is required to upregulate expression of inducible
nitric oxide synthase [51] and production of several chemokines,
including MIP-1a/CCL3, MIP-1b/CCL4, MIP-2/CXCL2, and
MCP-1/CCL2 [52]. Gene expression profiling of sHZ-laden
RAW 264.7 macrophage cells display altered NF-kB signal
transduction, enhanced inflammatory response and a severe
MMP-9/TIMP-1 imbalance in favor of ECM proteolysis [60].
NF-kB activation is also mandatory for nHZ-induced MMP-9
expression in human THP-1 monocyte cell line [61] and for nHZ-
Figure 3. Role of TNF-a, IL-1b, and MIP-1a/CCL3 in nHZ-induced mRNA expression and protein release of human monocytic TIMP-1.Human adherent monocytes were left unfed (negative controls) or fed with nHZ (positive controls). Thereafter, cells were incubated for 15 h (mRNAstudies, Panels A-B) or 24 h (protein studies, Panels C-D) in the presence/absence of the following treatments: single or combined doses (30 ng/ml) ofanti-hTNF-a, anti-hIL-1b and anti-hMIP-1a/CCL3 blocking antibodies (Panels A and C, blocking approach); and single or combined doses (20 ng/ml) ofrhTNF-a, rhIL-1b and rhMIP-1a/CCL3 (Panels B and D, mimicking approach). After lysis, TIMP-1 mRNA expression was measured by Real Time RT-PCR;alternatively, TIMP-1 protein levels in cell supernatants were evaluated by Western blotting and densitometry. Data are indicated as meanvalues+SEM or as a representative blot of three independent experiments. All data were evaluated for significance by ANOVA. Panel A: Vsunstimulated cells (column 1) *p,0.01; Vs untreated nHZ-fed cells (column 2) up,0.05. Panel B: Vs unstimulated cells (column 1) *p,0.05, **p,0.01.Panel C: Vs unstimulated cells (column 1) *p,0.05, **p,0.0001; Vs untreated nHZ-fed cells (column 2) up,0.01, uup,0.0001; Vs anti-TNFa-treatednHZ-fed cells (column 3) 1p,0.05. Panel D: Vs unstimulated cells (column 1) *p,0.05, **p,0.01, ***p,0.001.doi:10.1371/journal.pone.0071468.g003
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 7 August 2013 | Volume 8 | Issue 8 | e71468
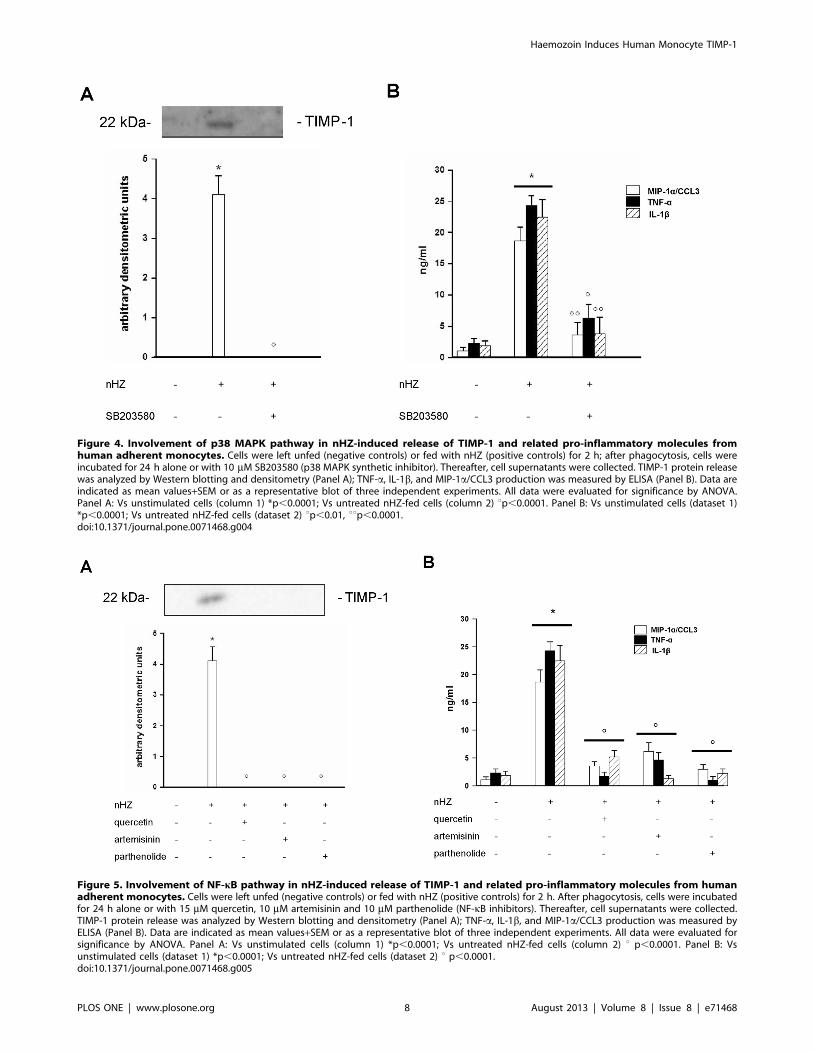
Figure 4. Involvement of p38 MAPK pathway in nHZ-induced release of TIMP-1 and related pro-inflammatory molecules fromhuman adherent monocytes. Cells were left unfed (negative controls) or fed with nHZ (positive controls) for 2 h; after phagocytosis, cells wereincubated for 24 h alone or with 10 mM SB203580 (p38 MAPK synthetic inhibitor). Thereafter, cell supernatants were collected. TIMP-1 protein releasewas analyzed by Western blotting and densitometry (Panel A); TNF-a, IL-1b, and MIP-1a/CCL3 production was measured by ELISA (Panel B). Data areindicated as mean values+SEM or as a representative blot of three independent experiments. All data were evaluated for significance by ANOVA.Panel A: Vs unstimulated cells (column 1) *p,0.0001; Vs untreated nHZ-fed cells (column 2) up,0.0001. Panel B: Vs unstimulated cells (dataset 1)*p,0.0001; Vs untreated nHZ-fed cells (dataset 2) up,0.01, uup,0.0001.doi:10.1371/journal.pone.0071468.g004
Figure 5. Involvement of NF-kB pathway in nHZ-induced release of TIMP-1 and related pro-inflammatory molecules from humanadherent monocytes. Cells were left unfed (negative controls) or fed with nHZ (positive controls) for 2 h. After phagocytosis, cells were incubatedfor 24 h alone or with 15 mM quercetin, 10 mM artemisinin and 10 mM parthenolide (NF-kB inhibitors). Thereafter, cell supernatants were collected.TIMP-1 protein release was analyzed by Western blotting and densitometry (Panel A); TNF-a, IL-1b, and MIP-1a/CCL3 production was measured byELISA (Panel B). Data are indicated as mean values+SEM or as a representative blot of three independent experiments. All data were evaluated forsignificance by ANOVA. Panel A: Vs unstimulated cells (column 1) *p,0.0001; Vs untreated nHZ-fed cells (column 2) u p,0.0001. Panel B: Vsunstimulated cells (dataset 1) *p,0.0001; Vs untreated nHZ-fed cells (dataset 2) u p,0.0001.doi:10.1371/journal.pone.0071468.g005
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 8 August 2013 | Volume 8 | Issue 8 | e71468
enhanced release of MMP-9, lysozyme, TNF-a and IL-1b from
human adherent monocytes isolated from peripheral blood
[13,25,31]. Consistently, in peripheral mononuclear cells of
malaria patients, phospho-NF-kB p65 levels are significantly
higher than in healthy controls [62], and genome wide expression
profiles display NF-kB-dependent enhancement of inflammatory
cytokines [63].
Results from the present work suggest that nHZ-induced
secretion of TIMP-1 and related pro-inflammatory molecules
from human monocytes require the involvement of either p38
MAPK- or NF-kB-dependent mechanisms. Indeed, the effects of
nHZ-mediated protein release of TIMP-1, TNF-a, IL-1b and
MIP-1a/CCL3 are fully abrogated by using micromolar doses of
SB203580, a synthetic inhibitor of p38 MAPK pathway [34], or
after treatment with quercetin, artemisinin and parthenolide -
three molecules showing antimalarial properties [64–65] able to
block NF-kB pathway at different levels (I-kBa phosphorylation
and subsequent degradation [35]; NF-kB nuclear translocation
[36]; and NF-kB binding to DNA [37], respectively). Interestingly,
these molecules also abrogate lysozyme and MMP-9 release in the
same cellular model [13,25,31,54].
Collectively, the present data show phagocytosis of nHZ by
human monocytes induces inflammation-mediated expression and
release of TIMP-1 through p38 MAPK- and NF-kB-dependent
mechanisms, suggesting that in vivo nHZ-fed human monocytes
may be a source for the high TIMP-1 serum levels found by
Dietmann and colleagues in malaria patients [21]. In this context,
TIMP-1 levels are also related to disease severity. However, it
remains unclear how TIMP-1 may exacerbate the clinical course
in malaria patients. One major function of TIMP-1 is to
counteract and modulate the numerous effects of MMP-9, which
include degradation of the sub-endothelial basal lamina, modula-
tion of the activity of several pro-inflammatory molecules,
disruption of tight junctions, and impairment of haemostasis [4–
7;38], all of which comprise key roles in CM. In this way it is
attractive to hypothesize that increased TIMP-1 levels are actually
protective rather than a risk factor in malaria prognosis, as they
could contrast the potentially detrimental effects of nHZ-enhanced
MMP-9. However, our results show that MMP-9/TIMP-1
stoichiometric ratios and total gelatinolytic activity measured in
nHZ-fed monocyte supernatants were significantly higher than in
controls, arguing against TIMP-1 ability to act as a protective
factor in this context. Total gelatinolytic activity reflects the net
overall activity of gelatinases (MMP-2 and -9) and their
endogenous inhibitors (TIMP-2 and TIMP-1, respectively) [33],
but in the present model it should be noted that the activity is due
solely to the net balance between MMP-9 and TIMP-1, as unfed
and nHZ-fed human adherent monocytes do not release MMP-2
and TIMP-2 proteins [41]. Therefore, these results suggest that
nHZ-dependent induction of TIMP-1 expression and release is not
sufficient to counterbalance nHZ-enhanced MMP-9 levels re-
leased from human monocytes.
Intriguingly, TIMP-1 could play a detrimental role through an
MMP-independent mechanism [39–40]. MMP-independent roles
Figure 6. nHZ-dependent induction of TIMP-1 secretion does not counterbalance nHZ-dependent MMP-9 enhancement in humanadherent monocytes. Cells were left unfed or fed with nHZ for 2 h; after washing, cells were incubated for 24 h alone or with 10 mM SB203580(p38 MAPK synthetic inhibitor), 15 mM quercetin, 10 mM artemisinin and 10 mM parthenolide (NF-kB inhibitors). During the incubation, cellsupernatants were collected at different time-points (6, 15 and 24 h). Thereafter, MMP-9 and TIMP-1 protein levels were measured by ELISA, andMMP-9/TIMP-1 ratios were calculated; additionally, total gelatinolytic activity was analyzed by fluorogenic gelatin conversion assay. Panel A. MMP-9/TIMP-1 stoichiometric ratios in unfed (white columns) and nHZ-fed (black columns) cell supernatants during time (6, 15 and 24 after the end ofphagocytosis). Panel B. MMP-9 (white columns) and TIMP-1 (black columns) protein levels (expressed as pmol/ml) in unfed and nHZ-fed cellsupernatants, in the presence/absence of p38 MAPK and NF-kB inhibitors. Panel C. Gelatinolytic activity (expressed as gelatinolytic activity units perml, with one gelatinolytic unit corresponding to 37 ng activated rhMMP-9) in unfed and nHZ-fed cell supernatants, in the presence/absence of p38MAPK and NF-kB inhibitors. Data are mean values+SEM of three independent experiments. All data were evaluated for significance by ANOVA. PanelA: Vs unfed cells (6 h) *p,0.05, **p,0.005; Vs nHZ-fed cells (6 h) up,0.001, uup,0.0001; Vs unfed cells (15 h) #p,0.0001. Panel B: Vs unstimulatedcells (column 1) *p,0.0001; Vs untreated nHZ-fed cells (column 2) up,0.0001. Panel C: Vs unstimulated cells (column 1) *p,0.05; Vs untreated nHZ-fed cells (column 2) up,0.05.doi:10.1371/journal.pone.0071468.g006
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 9 August 2013 | Volume 8 | Issue 8 | e71468

for TIMP-1 in biological processes include anti-apoptotic effects of
TIMP-1 in several human cells, such as Burkitt’s lymphoma [66],
breast epithelial cells [67], and cardiomyocytes [68]. Recently
CD63, a member of the tetraspanin family, was identified as a cell-
binding partner for TIMP-1 in human mammary epithelial cells,
and CD63 down-regulation with shRNA resulted in reduced
TIMP-1 binding and restored cell apoptosis [69]. Interestingly,
nHZ-fed monocytes do not undergo apoptosis, despite increased
inflammation and functional impairment [30,43]. CD63 is
constitutively expressed by human monocytes [70]. Thus, it is
intriguing to speculate that nHZ-enhanced TIMP-1 levels might
prevent apoptosis of functionally impaired nHZ-fed cells through
CD63-dependent mechanisms. Additionally, there are several
reports describing MMP-independent abilities of TIMP-1 to
inhibit cell growth and differentiation [39–40,71]. These TIMP-
1 properties may be crucial for nHZ-fed monocytes, since these
cells have been shown not to mature to dendritic cells [72] and not
to coordinate erythropoiesis [73].
In conclusion, the present work expands the available evidence
on the expression of TIMPs in malaria, providing new information
on the mechanisms underlying nHZ-dependent TIMP-1 increase.
Future investigation is needed to ascertain whether nHZ-enhanced
TIMP-1 may contribute to worsen the clinical course in malaria
patients as a consequence of its MMP-independent anti-apoptotic
or growth/differentiation-inhibitory properties. Further, previous
evidence correlating the number of circulating nHZ-laden
monocytes in patients to parasitaemia degree and malaria severity
[29] seems to support such a hypothesis. More extensive research
on the functional role of TIMP-1 in malaria, along with a better
understanding of MMP-independent TIMP functions is necessary
in order to find new tools for differential diagnosis and therapy of
severe malaria.
Supporting Information
Figure S1 Phagocytic meals and treatments do notdisplay cytotoxicity and do not affect viability of humanadherent monocytes. Cells were left unfed or fed with nHZ
and latex for 2 h; after washing, nHZ-fed cells were incubated for
24 h alone, whereas unfed cells were incubated for 24 h with
30 ng/ml of anti-hTNF-a, anti-hIL-1b, anti-hMIP-1a/CCL3
blocking antibodies; 20 ng/ml of rhTNF-a, rhIL-1b, rhMIP-1a/
CCL3; 10 mM SB203580; 15 mM quercetin; 10 mM artemisinin;
and 10 mM parthenolide. Thereafter, cell supernatants and lysates
were collected and LDH activity was measured by a spectrometric
assay. Panel A. Cytotoxicity of phagocytic meals and treatments,
expressed as percentage of (extracellular LDH activity)/(total
LDH activity) ratio versus controls (unfed/untreated monocytes).
Panel B. Viability of cells after exposure to phagocytic meals and
treatments, expressed as percentage of (intracellular LDH
activity)/(total LDH activity) ratio versus controls (unfed untreated
monocytes). Data are mean values+SEM of three independent
experiments. All data were evaluated for significance by ANOVA:
no significant differences were found.
(TIF)
Figure S2 Co-culturing with nHZ-fed monocytes induc-es unfed cells to release TIMP-1 and MMP-9. Unfed
monocytes (16106 cells/well) were plated at the bottom of the
wells, with nHZ-fed human adherent monocytes (0,56106 cells/
well) seeded onto the inserts. Co-cultures were incubated for 2 h
before removal of the inserts. After washings, unfed monocytes
were further incubated for 24 h. Non-co-cultured unfed and nHZ-
cells were also used as negative and positive controls, respectively.
Cell supernatants were collected and analysed for TIMP-1 and
MMP-9 secretion by ELISA. Panel A. Secretion of TIMP-1 (white
columns), expressed as pg/ml. Panel B. Secretion of MMP-9
(white columns) and TIMP-1 (black columns), expressed as pmol/
ml. Data are mean values+SEM of three independent experi-
ments. All data were evaluated for significance by ANOVA. Panel
A: Vs non-co-cultured unfed cells (column 1) *p,0.0001; Vs non-
co-cultured nHZ-fed cells (column 3) up,0.0001. Panel B: Vs non-
co-cultured unfed cells (column 1) *p,0.0001.
(TIF)
Acknowledgments
Thanks are due to Nathalie Geurts and Erik Martens for help with the
fluorogenic gelatin conversion assay; to Valentina Gallo for help with
FACS and Real Time RT-PCR analyses; to Giulia Rossana Gulino for
help with LDH experiments; to Nicoletta Basilico and colleagues for
providing supplementary nHZ and six-well plate Transwell systems for co-
culturing experiments; to Mary Lynn Dear and Andrew Giles for
comments on the manuscript; and to Associazione Volontari Italiani
Sangue (AVIS Torino) for providing fresh blood.
Author Contributions
Conceived and designed the experiments: GG M. Prato. Performed the
experiments: M. Polimeni EV DU GG M. Prato. Analyzed the data: M.
Polimeni GO PEVDS GG M. Prato. Contributed reagents/materials/
analysis tools: EV GO PEVDS GG M. Prato. Wrote the paper: M.
Polimeni GG M. Prato.
References
1. Khadjavi A, Giribaldi G, Prato M (2010) From control to eradication of malaria:
the end of being stuck in second gear? Asian Pac J Trop Med 3: 412–420.
2. WHO (2011) World Malaria Report.
3. Higgins SJ, Kain KC, Liles WC (2011) Immunopathogenesis of falciparum
malaria: implications for adjunctive therapy in the management of severe and
cerebral malaria. Expert Rev Anti Infect Ther 9: 803–819.
4. Lucchi NW, Jain V, Wilson NO, Singh N, Udhayakumar V, et al (2011)
Potential serological biomarkers of cerebral malaria. Dis Markers 31: 327–335.
5. Nagase H, Woessner JF (1999) Matrix metalloproteinases. J Biol Chem 274:
21491–21494.
6. Cauwe B, Van den Steen PE, Opdenakker G (2007) The biochemical,
biological, and pathological kaleidoscope of cell surface substrates processed by
matrix metalloproteinases. Crit Rev Biochem Mol Biol 42: 113–185.
7. Seo JH, Guo S, Lok J, Navaratna D, Whalen MJ, et al (2012) Neurovascular
matrix metalloproteinases and the blood-brain barrier. Curr Pharm Des 18:
3645–3648.
8. Szklarczyk A, Stins M, Milward EA, Ryu H, Fitzsimmons C, et al (2007) Glial
activation and matrix metalloproteinase release in cerebral malaria. J Neurovirol
13: 2–10.
9. Prato M, Giribaldi G (2011b) Matrix Metalloproteinase-9 and Haemozoin:
Wedding Rings for Human Host and Plasmodium falciparum Parasite inComplicated Malaria. J Trop Med 2011: 628435.
10. Geurts N, Opdenakker G, Van den Steen PE (2012) Matrix metalloproteinases
as therapeutic targets in protozoan parasitic infections. Pharmacol Ther 133:
257–279.
11. Prato M, Giribaldi G, Polimeni M, Gallo V, Arese P (2005) Phagocytosis ofhemozoin enhances matrix metalloproteinase-9 activity and TNF-alpha
production in human monocytes: role of matrix metalloproteinases in thepathogenesis of falciparum malaria. J Immunol 175: 6436–6442.
12. Prato M, Gallo V, Giribaldi G, Arese P (2008) Phagocytosis of haemozoin(malarial pigment) enhances metalloproteinase-9 activity in human adherent
monocytes: role of IL-1beta and 15-HETE. Malar J 7: 157.
13. Prato M, Gallo V, Giribaldi G, Aldieri E, Arese P (2010a) Role of the NF-kBtranscription pathway in the haemozoin- and 15-HETE-mediated activation of
matrix metalloproteinase-9 in human adherent monocytes. Cell Microbiol 12:
1780–1791.
14. Giribaldi G, Valente E, Khadjavi A, Polimeni M, Prato M (2011) Macrophageinflammatory protein-1alpha mediates matrix metalloproteinase-9 enhancement
in human adherent monocytes fed with malarial pigment. Asian Pac J Trop Med
4: 925–930.
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 10 August 2013 | Volume 8 | Issue 8 | e71468

15. Prato M, D’Alessandro S, Van den Steen PE, Opdenakker G, Arese P, et al
(2011a) Natural haemozoin modulates matrix metalloproteinases and inducesmorphological changes in human microvascular endothelium. Cell Microbiol
13: 1275–1285.
16. D’Alessandro S, Basilico N, Prato M (2013) Effects of Plasmodium falciparum-infected erythrocytes on matrix metalloproteinase-9 regulation in human
microvascular endothelial cells. Asian Pac J Trop Med 6: 195–199.
17. Geurts N, Martens E, Van Aelst I, Proost P, Opdenakker G, et al (2008) Beta-
hematin interaction with the hemopexin domain of gelatinase B/MMP-9provokes autocatalytic processing of the propeptide, thereby priming activation
by MMP-3. Biochemistry 47: 2689–2699.
18. Brew K, Nagase H (2010) The tissue inhibitors of metalloproteinases (TIMPs):an ancient family with structural and functional diversity. Biochim Biophys Acta
1803: 55–71.
19. Van den Steen PE, Van Aelst I, Starckx S, Maskos K, Opdenakker G, et al
(2006b) Matrix metalloproteinases, tissue inhibitors of MMPs and TACE inexperimental cerebral malaria. Lab Invest 86: 873–888.
20. Moreno A, Cabrera-Mora M, Garcia A, Orkin J, Strobert E, et al (2013)
Plasmodium coatneyi in Rhesus Macaques Replicates the MultisystemicDysfunction of Severe Malaria in Humans. Infect Immun 81: 1889–1904.
21. Dietmann A, Helbok R, Lackner P, Issifou S, Lell B, et al (2008) Matrix
metalloproteinases and their tissue inhibitors (TIMPs) in Plasmodium falciparummalaria: serum levels of TIMP-1 are associated with disease severity. J Infect Dis
197: 1614–1620.
22. Hanemaaijer R, Koolwijk P, le Clercq L, de Vree WJ, van Hinsbergh VW
(1993) Regulation of matrix metalloproteinase expression in human vein andmicrovascular endothelial cells. Effects of tumour necrosis factor alpha,
interleukin 1 and phorbol ester. Biochem J 296 (Pt 3): 803–809.
23. Zhang Y, McCluskey K, Fujii K, Wahl LM (1998) Differential regulation ofmonocyte matrix metalloproteinase and TIMP-1 production by TNF-alpha,
granulocyte-macrophage CSF, and IL-1 beta through prostaglandin-dependent
and -independent mechanisms. J Immunol 161: 3071–3076.
24. Van den Steen PE, Van Aelst I, Hvidberg V, Piccard H, Fiten P, et al (2006a)The hemopexin and O-glycosylated domains tune gelatinase B/MMP-9
bioavailability via inhibition and binding to cargo receptors. J Biol Chem 281:18626–18637.
25. Polimeni M, Valente E, Aldieri E, Khadjavi A, Giribaldi G, et al (2012)
Haemozoin induces early cytokine-mediated lysozyme release from human
monocytes through p38 MAPK- and NF-kappaB-dependent mechanisms. PLoSOne 7: e39497.
26. Deroost K, Tyberghein A, Lays N, Noppen S, Schwarzer E, et al (2013)
Hemozoin induces lung inflammation and correlates with malaria-associatedacute respiratory distress syndrome. Am J Resp Cell Mol Biol 48: 589–600.
27. Schwarzer E, Turrini F, Ulliers D, Giribaldi G, Ginsburg H, et al (1992)
Impairment of macrophage functions after ingestion of Plasmodium falciparum-
infected erythrocytes or isolated malarial pigment. J Exp Med 176: 1033–1041.
28. Schwarzer E, Kuhn H, Valente E, Arese P (2003) Malaria-parasitizederythrocytes and hemozoin nonenzymatically generate large amounts of
hydroxy fatty acids that inhibit monocyte functions. Blood 101: 722–728.
29. Were T, Davenport GC, Yamo EO, Hittner JB, Awandare GA, et al (2009)Naturally acquired hemozoin by monocytes promotes suppression of RANTES
in children with malarial anemia through an IL-10-dependent mechanism.
Microbes Infect 11: 811–819.
30. Prato M, Gallo V, Valente E, Khadjavi A, Mandili G, et al (2010b) Malarialpigment enhances Heat Shock Protein-27 in THP-1 cells: new perspectives for
in vitro studies on monocyte apoptosis prevention. Asian Pac J Trop Med 3:934–938.
31. Polimeni M, Valente E, Aldieri E, Khadjavi A, Giribaldi G, et al (2013) Role of
15-HETE in haemozoin-induced lysozyme release from human adherent
monocytes. Biofactors 39: 304–314.
32. Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29: e45.
33. Vandooren J, Geurts N, Martens E, Van den Steen PE, Jonghe SD, et al (2011)
Gelatin degradation assay reveals MMP-9 inhibitors and function of O-glycosylated domain. World J Biol Chem 2: 14–24.
34. Lee JC, Kumar S, Griswold DE, Underwood DC, Votta BJ, et al (2000)
Inhibition of p38 MAP kinase as a therapeutic strategy. Immunopharmacol 47:
185–201.
35. Nair MP, Mahajan S, Reynolds JL, Aalinkeel R, Nair H, et al (2006) Theflavonoid quercetin inhibits proinflammatory cytokine (tumor necrosis factor
alpha) gene expression in normal peripheral blood mononuclear cells viamodulation of the NF-kappa beta system. Clin Vaccine Immunol 13: 319–328.
36. Aldieri E, Atragene D, Bergandi L, Riganti C, Costamagna C, et al (2003)
Artemisinin inhibits inducible nitric oxide synthase and nuclear factor NF-kBactivation. FEBS Lett 552: 141–144.
37. Garcıa-Pineres AJ, Castro V, Mora G, Schmidt TJ, Strunck E, et al (2001)Cysteine 38 in p65/NF-kappaB plays a crucial role in DNA binding inhibition
by sesquiterpene lactones. J Biol Chem 276: 39713–39720.
38. Nagase H, Visse R, Murphy G (2006) Structure and function of matrixmetalloproteinases and TIMPs. Cardiovasc Res 69: 562–573.
39. Chirco R, Liu XW, Jung KK, Kim HR (2006) Novel functions of TIMPs in cell
signaling. Cancer Metastasis Rev 25: 99–113.
40. Stetler-Stevenson WG (2008) Tissue inhibitors of metalloproteinases in cell
signaling: metalloproteinase-independent biological activities. Sci Signal 1: re6.
41. Prato M (2011) Malarial pigment does not induce MMP-2 and TIMP-2 protein
release by human monocytes. Asian Pac J Trop Med 4: 756.
42. Bar-Or A, Nuttall RK, Duddy M, Alter A, Kim HJ, et al (2003) Analyses of all
matrix metalloproteinase members in leukocytes emphasize monocytes as major
inflammatory mediators in multiple sclerosis. Brain 126: 2738–2749.
43. Giribaldi G, Prato M, Ulliers D, Gallo V, Schwarzer E, et al (2010) Involvement
of inflammatory chemokines in survival of human monocytes fed with malarial
pigment. Infect Immun 78: 4912–4921.
44. Opdenakker G, Masure S, Proost P, Billiau A, van Damme J (1991b) Natural
human monocyte gelatinase and its inhibitor. FEBS Lett 284: 73–78.
45. Yamamoto T, Eckes B, Mauch C, Hartmann K, Krieg T (2000) Monocyte
chemoattractant protein-1 enhances gene expression and synthesis of matrix
metalloproteinase-1 in human fibroblasts by an autocrine IL-1 alpha loop.
J Immunol 164: 6174–6179.
46. Johnatty RN, Taub DD, Reeder SP, Turcovski-Corrales SM, Cottam DW, et al
(1997) Cytokine and chemokine regulation of proMMP-9 and TIMP-1
production by human peripheral blood lymphocytes. J Immunol 158: 2327–
2333.
47. Opdenakker G, Masure S, Grillet B, Van Damme J (1991a) Cytokine-mediated
regulation of human leukocyte gelatinases and role in arthritis. Lymphokine
Cytokine Res 10: 317–324.
48. Nguyen J, Gogusev J, Knapnougel P, Bauvois B (2006) Protein tyrosine kinase
and p38 MAP kinase pathways are involved in stimulation of matrix
metalloproteinase-9 by TNF-alpha in human monocytes. Immunol Lett 106:
34–41.
49. Jovanovic DV, Di Battista JA, Martel-Pelletier J, Reboul P, He Y, et al (2001)
Modulation of TIMP-1 synthesis by antiinflammatory cytokines and prosta-
glandin E2 in interleukin 17 stimulated human monocytes/macrophages.
J Rheumatol 28: 712–718.
50. Cambos M, Bazinet S, Abed E, Sanchez-Dardon J, Bernard C, et al (2010) The
IL-12p70/IL-10 interplay is differentially regulated by free heme and hemozoin
in murine bone-marrow-derived macrophages. Int J Parasitol 40: 1003–1012.
51. Jaramillo M, Gowda DC, Radzioch D, Olivier M (2003) Hemozoin increases
IFN-gamma-inducible macrophage nitric oxide generation through extracellular
signal-regulated kinase- and NF-kappa B-dependent pathways. J Immunol 171:
4243–4253.
52. Jaramillo M, Godbout M, Olivier M (2005) Hemozoin induces macrophage
chemokine expression through oxidative stress-dependent and -independent
mechanisms. J Immunol 2005. 174: 475–484.
53. Griffith JW, Sun T, McIntosh MT, Bucala R (2009) Pure Hemozoin is
inflammatory in vivo and activates the NALP3 inflammasome via release of uric
acid. J Immunol 183: 5208–5220.
54. Khadjavi A, Valente E, Giribaldi G, Prato M (2013) Involvement of p38 MAPK
in natural haemozoin- and 15-HETE-dependent MMP-9 enhancement in
human adherent monocytes. Cell Biochem Funct. Epub 2013 Mar 7. DOI:
10.1002/cbf.2963.
55. Lucchi NW, Peterson DS, Moore JM (2008) Immunologic activation of human
syncytiotrophoblast by Plasmodium falciparum. Malar J 7: 42.
56. Lucchi NW, Sarr D, Owino SO, Mwalimu SM, Peterson DS, et al (2011)
Natural hemozoin stimulates syncytiotrophoblast to secrete chemokines and
recruit peripheral blood mononuclear cells. Placenta 32: 579–585.
57. Zhu J, Wu X, Goel S, Gowda NM, Kumar S, et al (2009) MAPK-activated
protein kinase 2 differentially regulates plasmodium falciparum glycosylpho-
sphatidylinositol-induced production of tumor necrosis factor-{alpha} and
interleukin-12 in macrophages. J Biol Chem 284: 15750–15761.
58. Lu Z, Serghides L, Patel SN, Degousee N, Rubin BB, et al (2006) Disruption of
JNK2 decreases the cytokine response to Plasmodium falciparum glycosylpho-
sphatidylinositol in vitro and confers protection in a cerebral malaria model.
J Immunol 177: 6344–6352.
59. Serghides L, Patel SN, Ayi K, Lu Z, Gowda DC, et al (2009) Rosiglitazone
modulates the innate immune response to Plasmodium falciparum infection and
improves outcome in experimental cerebral malaria. J Infect Dis 199: 1536–
1545.
60. Schrimpe AC, Wright DW (2009) Comparative analysis of gene expression
changes mediated by individual constituents of hemozoin. Chem Res Toxicol
22: 433–445.
61. Dell’agli M, Galli GV, Bulgari M, Basilico N, Romeo S, et al (2010)
Ellagitannins of the fruit rind of pomegranate (Punica granatum) antagonize
in vitro the host inflammatory response mechanisms involved in the onset of
malaria. Malar J 9: 208. DOI: 10.1186/1475-2875-9-208.
62. Punsawad C, Krudsood S, Maneerat Y, Chaisri U, Tangpukdee N, et al (2012)
Activation of nuclear factor kappa B in peripheral blood mononuclear cells from
malaria patients. Malar J 11: 191.
63. Ockenhouse CF, Hu WC, Kester KE, Cummings JF, Stewart A, et al (2006)
Common and divergent immune response signaling pathways discovered in
peripheral blood mononuclear cell gene expression patterns in presymptomatic
and clinically apparent malaria. Infect Immun 74: 5561–5573.
64. Burrows JN, Chibale K, Wells TN (2011) The state of the art in anti-malarial
drug discovery and development. Curr Top Med Chem 11: 1226–1254.
65. Khalid SA, Farouk A, Geary TG, Jensen JB (1986) Potential antimalarial
candidates from African plants: and in vitro approach using Plasmodium
falciparum. J Ethnopharmacol 15: 201–209.
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 11 August 2013 | Volume 8 | Issue 8 | e71468

66. Guedez L, Stetler-Stevenson WG, Wolff L, Wang J, Fukushima P, et al (1998) In
vitro suppression of programmed cell death of B cells by tissue inhibitor of
metalloproteinases-1. J Clin Invest 102: 2002–2010.
67. Li G, Fridman R, Kim HR (1999) Tissue inhibitor of metalloproteinase-1
inhibits apoptosis of human breast epithelial cells. Cancer Res 59: 6267–6275.
68. Singla DK, McDonald DE (2007) Factors released from embryonic stem cells
inhibit apoptosis of H9c2 cells. Am J Physiol Heart Circ Physiol 293: H1590–
1595.
69. Jung KK, Liu XW, Chirco R, Fridman R, Kim HR (2006) Identification of
CD63 as a tissue inhibitor of metalloproteinase-1 interacting cell surface protein.
EMBO J 25: 3934–3942.
70. Pols MS, Klumperman J (2009) Trafficking and function of the tetraspanin
CD63. Exp Cell Res 315: 1584–1592.71. Egea V, Zahler S, Rieth N, Neth P, Popp T, et al (2012) Tissue inhibitor of
metalloproteinase-1 (TIMP-1) regulates mesenchymal stem cells through let-7f
microRNA and Wnt/b-catenin signaling. Proc Natl Acad Sci USA 109: E309–316.
72. Urban BC, Todryk S (2006) Malaria pigment paralyzes dendritic cells. J Biol 5:4.
73. Giribaldi G, Ulliers D, Schwarzer E, Roberts I, Piacibello W, et al (2004)
Hemozoin- and 4-hydroxynonenal-mediated inhibition of erythropoiesis.Possible role in malarial dyserythropoiesis and anemia. Haematologica 89:
492–493.
Haemozoin Induces Human Monocyte TIMP-1
PLOS ONE | www.plosone.org 12 August 2013 | Volume 8 | Issue 8 | e71468
Related Documents











