The Role of 12-Lipoxygenase in Hypoxia-Induced Rat Pulmonary Artery Smooth Muscle Cell Proliferation IOANA R. PRESTON, NICHOLAS S. HILL, ROD R. WARBURTON, BARRY L. FANBURG Pulmonary, Critical Care and Sleep Division, Tufts-New England Medical Center, Tufts University School of Medicine, Boston, Massachusetts 02111 Running Head: 12-LO and pulmonary hypertension Contact information: Ioana R. Preston, MD Pulmonary, Critical Care and Sleep Division Tufts-New England Medical Center 750 Washington Street, Box #257 Boston, MA 02111 Tel. 617-636-7609 Fax. 617-636-5953 Email. [email protected] Articles in PresS. Am J Physiol Lung Cell Mol Physiol (September 30, 2005). doi:10.1152/ajplung.00114.2005 Copyright © 2005 by the American Physiological Society.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Role of 12-Lipoxygenase in Hypoxia-Induced Rat Pulmonary Artery Smooth
Muscle Cell Proliferation
IOANA R. PRESTON, NICHOLAS S. HILL, ROD R. WARBURTON, BARRY L.
FANBURG
Pulmonary, Critical Care and Sleep Division, Tufts-New England Medical Center,
Tufts University School of Medicine, Boston, Massachusetts 02111
Running Head: 12-LO and pulmonary hypertension
Contact information: Ioana R. Preston, MD
Pulmonary, Critical Care and Sleep Division
Tufts-New England Medical Center
750 Washington Street, Box #257
Boston, MA 02111
Tel. 617-636-7609
Fax. 617-636-5953
Email. [email protected]
Articles in PresS. Am J Physiol Lung Cell Mol Physiol (September 30, 2005). doi:10.1152/ajplung.00114.2005
Copyright © 2005 by the American Physiological Society.

12-LO and pulmonary hypertension LCMP—00114-2005.R1
2
ABSTRACT
The 12-lipoxygenase (12-LO) pathway of arachidonic acid metabolism stimulates
cell growth and metastasis of various cancer cells and the 12-LO metabolite,
12(S)-hydroxyeicosatetraenoic acid [12(S)-HETE], enhances proliferation of aortic
smooth muscle cells (SMCs). However, pulmonary vascular effects of 12-LO have
not been previous ly studied. We sought evidence for a role of 12-LO and 12-
HETE in the development of hypoxia-induced pulmonary hypertension. We found
that 12-LO gene and protein expression is elevated in lung homogenates of rats
exposed to chronic hypoxia. Immunohistochemical staining with a 12-LO antibody
revealed intense staining in endothelial cells of large pulmonary arteries, SMCs
(and possibly endothelial cells) of medium and small size pulmonary arteries, and
in alveolar walls of hypoxic lungs. 12-LO protein expression is increased in
hypoxic cultured rat pulmonary artery SMCs. 12-HETE at concentrations as low
as 10-5 µM stimulated proliferation of pulmonary artery SMCs. 12-HETE induced
ERK 1/ERK 2 phosphorylation, but had no effect on p38 kinase expression as
assessed by Western blotting. 12-HETE-stimulated SMC proliferation was
blocked by the MEK inhibitor, PD98059, but not by the p38 MAPK inhibitor,
SB202190. Hypoxia (3%) - stimulated pulmonary artery SMC proliferation was
blocked by both U0126, a MEK inhibitor, and baicalein, an inhibitor of 12-LO. We
conclude that 12-LO and its product, 12-HETE, are important intermediates in
hypoxia-induced pulmonary artery SMC proliferation and may participate in
hypoxia-induced pulmonary hypertension.
Keywords: 12-HETE, mitogen activated protein-kinases, ERK1/ERK2.

12-LO and pulmonary hypertension LCMP—00114-2005.R1
3
INTRODUCTION
Pulmonary hypertension occurs in animals and humans exposed to acute or
sustained hypoxia (26) and this is a well-established in vivo experimental model
of pulmonary hypertension. The initial event is acute hypoxic pulmonary
vasoconstriction, followed by remodeling of small and medium-sized pulmonary
arteries (46). Muscular intrapulmonary arteries become fur ther muscularized,
and distal, non-muscular arteries become newly muscularized (26). Pulmonary
artery smooth muscle cell (PASMC) proliferation is a key feature of pulmonary
vascular remodeling. The mechanism of PASMC proliferation is thought to
involve an imbalance between mediators of apoptosis and cell proliferation.
Local paracrine mediators induced by hypoxia may also participate in the
proliferative response. Among these local mediators, metabolites of arachidonic
acid derived via the lipoxygenase pathway are known to exert vascular effects.
In particular, the leukocyte-type 12-lipoxygenase (12-LO) pathway has been
shown to stimulate cell proliferation and participate in inflammatory reactions in
aortic SMCs (4, 27, 29-32, 35, 36) and in migration and metastasis of various
cancer cell lines (6, 11, 18, 23, 24, 28, 38, 39, 41, 42, 44, 45). In addition, the 12-
LO product, 12-S-hydroxyeicosatetraenoic acid (12-S-HETE), has a direct
hypertrophic effect on systemic vascular SMCs (29) and mediates hypertrophic
effects of angiotensin II (29) and chemotactic effects of platelet-derived growth
factor (27).
The 12-LO pathway has also been implicated in the proliferative response of
vascular SMCs in vivo. 12-LO gene expression is enhanced in SMCs of balloon-

12-LO and pulmonary hypertension LCMP—00114-2005.R1
4
injured rat carotid arteries and 12-LO inhibition by ribozyme-mediated cleavage of
the 12-LO mRNA significantly reduces neointimal proliferation in this model (17).
Despite the evidence that 12-LO and its products contribute to SMC
proliferation in the systemic vasculature, no studies have yet addressed the role
of 12-LO on SMC proliferation in the pulmonary circulation exposed to hypoxia.
We hypothesized that hypoxia stimulates pulmonary artery smooth muscle cell
proliferation through a mechanism that involves the 12-LO pathway. We sought
to study the effects of hypoxia on PASMCs obtained from adult rats and to
determine if the 12-LO pathway participates in this process.
MATERIALS AND METHODS
Materials. Dulbecco’s modified Eagle’s medium (DMEM) was from Sigma (St.
Louis, MO). Fetal bovine serum (FBS) was from Hyclone (Logan, UT).
Antibodies to ERK and phospho-ERK were from Sigma, and to p38 MAPK,
phospho-p38 MAPK and secondary HRP-conjugated rabbit anti-mouse
antibodies were from Cell Signaling (Beverly, MA). The specific antibody against
leukocyte type 12-LO was a gift from Dr. Jerry Nadler, University of Virginia (2).
12-S-HETE, 15-S-HETE, SB202190, PD98059 and U0126 were purchased from
Biomol Research Laboratories (Plymouth Meeting, PA). [3H]Thymidine was
purchased from Perkin-Elmer (Boston, MA) and the PCR kit was purchased from
Quiagen (Santa Clarita, CA).
Exposure of animals to chronic hypoxia. Adult male Sprague-Dawley rats
were randomized to 3 weeks of normoxia or hypobaric hypoxia (0.5 atm).

12-LO and pulmonary hypertension LCMP—00114-2005.R1
5
Hypobaric chambers were briefly opened 3 times weekly for cleaning and to
replace food and water. Twelve hour light exposure cycles, standard rat chow
and water ad libitum were provided to all rats. Normoxic rats were kept in the
same room adjacent to the hypobaric chamber. At the end of the 3-week
exposure period, rats were anesthetized and hemodynamic measurements were
obtained as previously described (22). After euthanasia using pentobarbital
injection (120 mg/kg ip), the thorax was opened immediately and the heart and
lungs were removed. The hearts were dissected into RV free wall and left
ventricle plus septum (LV+S) and the ratio of RV/(LV+S) was used as an index of
RV hypertrophy. The left lung was frozen in liquid nitrogen and stored at -80°C
for further measurements. Double fixation of the right lung was achieved in the
distended state by infusion of 4% aqueous buffered formalin into the trachea at
25 cm H2O pressure and into the pulmonary trunk at 5 cm H2O pressure. The
right lung was then processed for paraffin embedding. 5 µm sections were
immunostained with the leukocyte-type 12-LO antibody and counterstained with
hematoxylin-eosin. Vessels with a layer of smooth muscle cells comprising ≥ 3/4
of the vessel perimeter were categorized as fully muscularized. Vessels with a
smooth muscle layer comprising <3/4 of the vessel perimeter were categorized
as partially muscularized and vessels that had no identifiable smooth muscle
cells were categorized as nonmuscularized vessels.
Immunohistochemical detection of leukocyte-type 12-LO in rat lungs. The
immunohistochemical methods were carried out using the technique described
previously (31). Briefly, 5 µm paraffin-embedded tissue sections were mounted

12-LO and pulmonary hypertension LCMP—00114-2005.R1
6
on Probe-on slides (Biotek Solutions) and dried for 1h in 56°C oven and
overnight at 45-50°C, deparaffinized in xylene and rehydrated in graduated
alcohol to distilled water. The slides were loaded into a Techmate Slide holder
and placed into 0.1 mol/l citrate buffer solution for HIER. The slides were
steamed in 0.1M citrate buffer for 20 min and then allowed to cool for 5 min.
After treatment with the first and second antibody, slides were stained using a
modified ABC technique using 3,3’-diaminobenzidine tetrahydrochloride (DAB)
as the chromogen and conterstained using Mayer’s hematoxylin. Staining was
performed using a Bioteck Techmate 1000 Immunostainer (Teckmate, Santa
Barbara, CA) with Biotek solutions and an ABC detection system (Teckmate).
We used the 12-LO antibody at 1:500 concentration. Parallel controls were run
without primary antibody.
Isolation and culture of pulmonary artery smooth muscle cells. For isolation
of PASMCs, freshly excised lobar pulmonary arteries obtained from adult male
Sprague Dawley rats were stripped of adventitia. Vascular segments were then
cut open and endothelium was removed by gentle scraping of the luminal surface
of the vessel. Rat PASMCs were cultured from explants as previously described
(1, 9). SMC phenotype was assessed by the hill and valley morphology (1).
Cells were grown in DMEM supplemented with 10%FBS, penicillin (100 units/ml),
streptomycin (100 units/ml), and fungizone (1.25 µg/ml) and were passaged
every 1-2 weeks at a 1:3 ratio using trypsin. Passage 3-8 cells at 80%
confluence were used in all reported experiments. Before exposure to hypoxia or
treatment with different agents, cells were starved in 0.1% FBS DMEM with

12-LO and pulmonary hypertension LCMP—00114-2005.R1
7
antibiotics for 72 hours. Cells were exposed to hypoxia or treated with different
agents in 0.1% FBS DMEM with antibiotics, unless otherwise specified. Medium
was changed every 2 -3 days.
Exposure of PASMCs to hypoxia. PASMCs were exposed to 3% hypoxia in a
humidified modular incubator chamber (Billups-Rothenberg, Del Mar, CA) that
was maintained at 37°C. The incubator chamber was sealed and purged with
3%O2, 5%CO2, balance N2 for 15 minutes. Normoxic control PASMCs were
exposed to 95% ambient air, 5%CO2 for the entire incubation period. A portable
gas analyzer (Hudson Ventronics Division) was used to ensure that the O2
concentration inside the chamber was 3%. Hypoxia had no effect on the pH of
the medium, which was maintained at 7.36 ± 0.3.
[3H]Thymidine incorporation. PASMCs were plated in complete growth media
containing 10% FBS in 96-well plates at a density of 4 x 103 cells/cm2. After
plates were 80% confluent, cells were starved in 0.1% FBS for 72 hours. Cells
were then either exposed to hypoxia or normoxia, or treated with different agents
30 minutes before the beginning of the hypoxia exposure. Twenty four hours
before the end of the experiment, [3H]thymidine (0.1 mCi/ml) was added.
Incorporation of labeled thymidine was stopped by aspirating the medium and
trypsinizing the cells for 15 minutes at 37ºC, after which cells were transferred
onto a filter membrane using a cell harvester. Radioactivity was counted in a
microplate liquid scintillation counter (Perkin-Elmer) after 20 µl of scintillation
liquid was added to each well.
Cell proliferation. PASMCs were plated at 2 x 104 cells/well in 6-well plates in

12-LO and pulmonary hypertension LCMP—00114-2005.R1
8
DMEM and 10%FBS with antibiotics, growth-arrested for 72 hours, then exposed
to normoxia or hypoxia for various times. Cells were removed from the wells by
trypsin digestion. Cells were counted using a Fischer hemocytometer
(Pittsburgh, Pa). In each of the three experiments, the results from 3 wells were
averaged to obtain a single cell count (± SD) for each time point.
Western blot analysis. Protein determinations from both lung homogenates
and cell lysates were made using the Bradford method (3). To prepare lysates,
cells were washed in phosphate-buffered saline and solubilized with 50 mM
Hepes solution (pH 7.4) containing 1% (v/v) Triton X-100, 4 mM EDTA, 1 mM
sodium fluoride, 0.1 mM sodium orthovanidate, 1 mM tetrasodium
pyrophosphate, 2 mM pheylmethylsulsonyl fluoride, 10 µg/ml leupeptin and 10
µg/ml aprotinin. Cell lysates (20 µg of protein) were electrophoresed through
12% SDS-polyacrylamide gel and electroblotted onto a polyvinylidine difluoride
membrane. The membrane was blocked and incubated with primary antibodies
for phospho-specific ERK1/2, ERK, phospho-specific p38 MAPK, p38 MAPK or
12-LO. The polyclonal antibody against amino acids 646 to 662 peptide
sequence of porcine leukocyte 12-LO protein was raised in rabbits and has been
shown to have excellent cross-reactivity with rat 12-LO (49). The levels of
proteins and phosphoproteins were detected with horseradish peroxide-linked
secondary antibodies and the ECL System (Amersham Biosciences).
RT-PCR for 12-LO. Lungs were homogenized and total RNA was extracted
from the lung tissues by using the RNeasy Total RNA Isolation Kit (Quiagen,
Valencia, CA). The PCR method is specific for the leukocyte-type 12-LO and

12-LO and pulmonary hypertension LCMP—00114-2005.R1
9
does not cross-react with platelet-type 12-LO (17), which is the product of a
separate, linked gene. The primers for the rat 12-LO gene were: sense, 5’-TGG
GGC AAC TGG AAG G-3’, located at the 324 to 339 position of rat 12-LO, and
antisense, 5’-AGA GCG CTT CAG CAC CAT–3’ (2), located at the 718 to 735
position. PCR reaction was carried out under the following conditions: 38 cycles
of denaturation at 94ºC for 45 seconds, annealing at 68ºC for 60 seconds, and
extension at 72ºC for 60 seconds. A final extension was performed at 72ºC for
10 minutes. PCR products were separated by 3% polyacrylamide gel
electrophoresis in Tri-borate-EDTA buffer. For internal control, we performed RT-
PCR, then PCR for tubulin.
Statistical Analysis. Data are expressed as mean (± SD). Statistical analysis
was performed using the unpaired Student’s t-test, or analysis of variance, as
indicated. Differences were considered to be significant when p < 0.05.
RESULTS
12-LO mRNA and protein expression is increased in hypoxic rat lungs. Adult
rats exposed to 3 weeks of hypobaric hypoxia (0.5 atm) developed pulmonary
hypertension, as demonstrated by an increase in systolic pulmonary artery
pressures (27.5 ± 0.8 mmHg in normoxic rats and 42 ± 2 mmHg in hypoxic rats,
p < 0.05) and RV hypertrophy (RV/LV+S was 0.28 ± 0.005 in normoxic rats and
0.46 ± 0.02 in hypoxic rats, p < 0.05, N=5 animals per group). Lung
homogenates from these animals showed increases in both 12-LO mRNA (Fig 1
A) and protein (Fig 1 B, C) as compared to homogenates from normoxic controls.

12-LO and pulmonary hypertension LCMP—00114-2005.R1
10
Histologic examination (hematoxylin-eosin staining) revealed thickening of the
small pulmonary arterioles and no evidence of inflammation (Fig 2).
Evidence of increased 12-LO expression in hypoxic rat lung. In normoxic
rats, medium size and small pulmonary arteries were nonmuscularized (Fig 2 C).
After 3 weeks of exposure to hypoxia, many of these arteries became either
partially, or fully muscularized (Fig 2 F). Immunohistochemistry of rat lungs
showed intense 12-LO staining of bronchial epithelial cells and alveolar
macrophages that remained unchanged with hypoxia (Fig 2 A, C, D, F). In
normoxic lungs, there was no 12-LO staining of the pulmonary vasculature (Fig 2
B, C). In contrast, there was intense 12-LO immunostaining of endothelial cells
of large pulmonary arteries (Fig 2 D, E), SMCs (and possibly endothelial cells) of
small pulmonary arteries, and alveolar pneumocytes of hypoxic lungs (Fig 2 F).
Hypoxia stimulates rat PASMC proliferation. To provide an in vitro correlate
for the hypoxia-induced medial hypertrophy of pulmonary arteries seen in vivo,
we determined whether cultured SMCs proliferate during exposure to hypoxia.
PASMCs were growth-arrested in 0.1%FBS prior to being exposed to hypoxia or
normoxia with or without 10% serum. Hypoxia stimulated DNA synthesis, as
assessed by incorporation of [3H]thymidine into cells in both 0.1% and 10 %
serum (Fig 3 A, B). The proliferative effect of hypoxia was validated by
demonstrating an increase in cell number in hypoxic compared to control SMCs
(see Fig 3 C for representative experiment).
12-LO is up-regulated in hypoxic rat PASMCs in culture. Because PASMCs
are believed to participate in the pulmonary vascular response to hypoxia, we

12-LO and pulmonary hypertension LCMP—00114-2005.R1
11
next evaluated the influence of hypoxia on 12-LO protein expression in vitro.
Hypoxia up-regulated 12-LO protein expression in PASMCs at 2 hours and this
was maintained at 48 hours (Fig 4 A,B).
12-HETE stimulates PASMC proliferation through activation of ERK. Having
established that hypoxia enhances 12-LO expression and stimulates SMC
proliferation, we next sought to evaluate the effects of the 12-LO product, 12-
HETE, on SMC proliferation. Figure 5 A shows that 12-HETE directly stimulates
PASMC proliferation at concentrations as low as 10-5 µM. In contrast, 15-HETE,
a product resulting from arachidonic acid metabolism by 15-LO, did not have a
proliferative effect. In addition, 12-HETE-induced PASMC proliferation was
completely blocked by PD98059, a specific MEK inhibitor (an upstream factor of
ERK) (Fig 5 B). These results suggest that ERK and 12-LO are linked in the
proliferative pathway and that 12-LO up-regulation and 12-HETE formation
precede activation of ERK.
12-HETE stimulates ERK activation. In order to establish if ERK is
downstream of 12-HETE, we measured ERK activation in PASMCs stimulated
with 12-HETE. Fig 6 A and B demonstrate that 12-HETE directly activates the
ERK pathway by inducing phosphorylation of ERK1/ERK2. This is a biphasic
action with peaks at 10 and 90 minutes. Another signaling pathway that has
been shown to be activated during cell proliferation is p38 MAPK (10, 13). We
found that 12-HETE did not induce p38 MAPK phosphorylation (Fig 6 C, D).

12-LO and pulmonary hypertension LCMP—00114-2005.R1
12
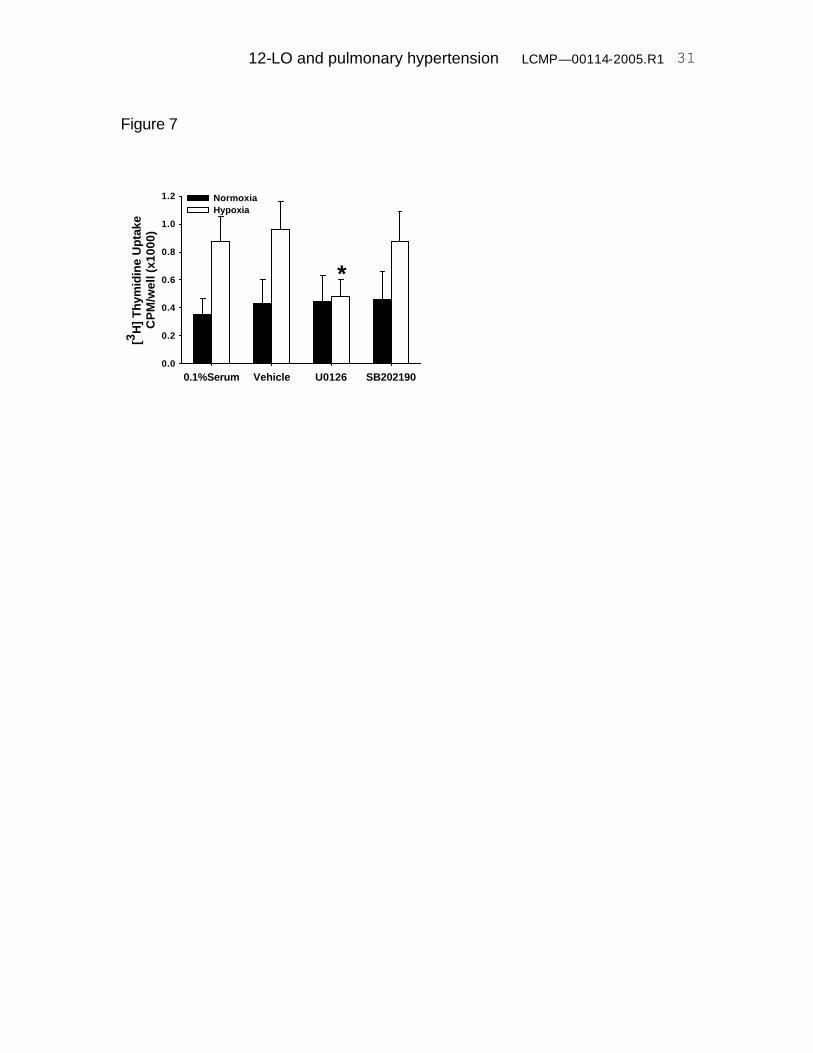
Hypoxia –induced PASMC proliferation is blocked by the MEK-1 inhibitor,
PD98059. In order to determine the cellular pathways that may be activated
during cell exposure to hypoxia, we evaluated the MAPK signaling pathway, in
particular ERK1/2 MAPK and p38 MAPK activation. PASMCs were exposed to
72 hours of hypoxia (3%O2) in the presence or absence of a specific MEK
inhibitor, U0126, as well as in the presence or absence of a specific p38 MAPK
inhibitor, SB202190. Fig 7 demonstrates inhibition of hypoxia-stimulated cell
proliferation by the MEK inhibitor and lack of effect by the p38 MAPK inhibitor.
Hypoxia-induced PASMC proliferation is blocked by 12-LO inhibition. A
partial blockade of hypoxia-stimulated SMC proliferation was obtained with the
12-LO inhibitor, baicalein (Fig 8). Studies using trypan blue exclusion
demonstrated that concentrations of baicalein from 0.5-10 µM had no effect on
cell viability (data not shown).
DISCUSSION
It is now generally accepted that hypoxia induces pulmonary hypertension
that is associated with pulmonary vascular remodeling and, in particular, smooth
muscle cell hyperplasia and hypertrophy. A large number of intermediate cell
signaling pathways by which this occurs have been proposed (1, 7-9, 14, 15, 19,
20, 22). The present study provides evidence that the 12-LO pathway via its
metabolite, 12-HETE, contributes to hypoxia-induced pulmonary arterial smooth
muscle cell remodeling. We found up-regulation of 12-LO mRNA and protein
expression in lung homogenates of rats exposed to chronic hypoxia and that 12-

12-LO and pulmonary hypertension LCMP—00114-2005.R1
13
HETE, but not 15-HETE, stimulates smooth muscle cell proliferation in vitro. We
also found increased 12-LO immunostaining of pulmonary arteries and
pneumocytes of hypoxic, but not normoxic rat lungs. Furthermore, we
substantiated the contribution of the 12-LO pathway to hypoxia-induced
pulmonary vascular SMC proliferation by demonstrating that inhibition of 12-LO
blunts the proliferative response.
The LOs comprise a family of non-heme iron-containing dioxygenases that
catalyze the stereospecific oxygenation of the 5-, 12- or 15-carbon atoms of
arachidonic acid. There are four functionally distinct 12-LOs that vary in cell and
tissue distribution and catalytic activity and each is a product of linked, but
separate genes. These consist of “platelet-type” 12-LO (P-12LO), “leukocyte-
type” 12-LO (L-12LO), epidermal-type 12-LO (e-12LO) (25) and 12(R)-LO. The
12-LO isoforms and their metabolite 12-HETE have been described in a variety
of rat, mouse and human tissues (37). Although macrophages have the capacity
to synthesize the leukocyte-type 12-LO in rat lungs (5), we do not believe that
macrophages are the source of the increased 12-LO in our in vivo rat model
because histological studies did not reveal an inflammatory pattern (unpublished
data), although this does not exclude the possibility.
12-LO products such as 12-HETE have been implicated in tumor cell
proliferation, particularly that of prostatic, pancreatic and breast cancers (11, 12,
23, 28). At least some of these effects may be due to inhibition of apoptosis (41).
No cellular receptor for 12-HETE has yet been identified, but some potential
pathways by which it may stimulate cellular activities have been explored. For

12-LO and pulmonary hypertension LCMP—00114-2005.R1
14
example, the mechanisms implicated in tumor invasion and metastasis that are
due, at least in part, to generation of 12-HETE, are believed to involve activation
of MAPK signaling pathways, in particular the ERK pathway (11, 39, 40, 44). In
addition, ERK activation has been demonstrated in the normal growth of
mesangial cells in association with the stimulation of the 12-LO pathway (34) and
in systemic vascular SMCs stimulated by arachidonic acid and its metabolites
(33).
Although we believe that ours is the first study exploring the role of 12-LO
and 12-HETE in pulmonary vascular smooth muscle cell proliferation, previous
studies have explored the influence of 12-LO and its product, 12-HETE, on
systemic vascular cells. 12-HETE is a known mitogenic factor for microvascular
endothelial cells (43) and systemic smooth muscle cells (29). In addition, 12-LO
participates in angiotensin II – induced vascular smooth muscle cell hypertrophy
(29, 30) and 15-LO has been shown to activate MAPK in systemic vascular
smooth muscle cells (33). The previously reported proliferative effects of 12-
HETE on systemic SMCs are consistent with our results demonstrating that 12-
HETE has a direct stimulatory effect on PASMC growth. This effect, similar to
the hypoxic stimulus, seems to involve ERK MAPK activation, as evidenced by
the increased phosphorylation of ERK that we observed during 12-HETE
treatment. What seems to be unique for the PASMCs is the biphasic activation of
ERK, which has not been reported in other cell systems. This possibly leads to a
positive feedback mechanism involving activation of the ERK pathway at multiple
steps. Furthermore, the proliferative effect of both hypoxia and 12-HETE on

12-LO and pulmonary hypertension LCMP—00114-2005.R1
15
PASMCs was completely blocked by the ERK inhibitor, PD98059, suggesting
that the ERK pathway is critical in 12-HETE-induced PASMC proliferation.
Although 15-HETE and 12-HETE have similar effects in tumor cells,
stimulating tumor invasion and metastasis, we found that, unlike 12-HETE, 15-
HETE exerts no proliferative effects in PASMCs. This indicates that some actions
of the various lipoxygenases are cell-specific.
Prior studies have observed activation of lipoxygenases in the lungs of
animals exposed to hypoxia, but these focused on pulmonary vasoconstriction
and examined gene regulation only to a limited extent. For example, Zhu et al.
(52) found activation of 15-lipoxygenase in the lungs of neonatal rabbits exposed
to hypoxia and enhanced constriction of pulmonary vascular rings from these
animals to 15-HETE. These data suggest that different lipoxygenases may play
different roles in various species.
The effects of hypoxia on proliferation of cultured PASMCs has been less
extensively studied than those on pulmonary vessels in intact experimental
models, and results have been less consistent. Whereas fibroblasts from
pulmonary arteries consistently proliferate in response to hypoxia (48), smooth
muscle cells behave differently. For example, bovine PAs have 2 distinct
populations of cells, only one of which proliferates in hypoxia (16). On the other
hand, hypoxia stimulates proliferation of cultured human PASMCs, even in the
absence of serum (50). In addition, when combined with serum, moderate
hypoxia (5%O2) synergistically stimulates human PASMC growth (9). Consistent
with human PASMCs, the rat PASMCs isolated from lobar PAs in our study

12-LO and pulmonary hypertension LCMP—00114-2005.R1
16
proliferated in response to hypoxic exposure (3%O2) and manifested synergy in
response to the combination of hypoxia and serum.
Prior studies on pulmonary arteries from chronically hypoxic rats have shown
increases in JNK, ERK and p38 MAPK activities, indicating involvement of
MAPKs in hypoxia-induced remodeling (19). Hypoxia also increases
phosphorylated-MAPK immunostaining in both large and small intrapulmonary
arteries (19). Our finding that a MEK inhibitor blocks hypoxia-stimulated PASMC
growth supports the idea that MAPKs are involved in the response. On the other
hand, p38 MAPK does not appear to be involved, as suggested by the lack of a
change in expression of phosphorylated p38 and lack of effect of a specific p38
MAPK inhibitor on PASMC proliferation. In contrast, p38 MAPK is activated in
hypoxic cultured PA endothelial cells (21), suggesting that signaling pathways
responding to hypoxic exposure are cell-specific.
Limitations of our study include the fact that our focus was on the role of 12-
LO in the pulmonary vasculature during hypoxia and we did not examine the
roles of other lipoxygenases, such as 5-LO, or 15-LO. The 5-LO pathway
appears to participate in the development of hypoxia-induced pulmonary
hypertension, because inhibition of 5-lipoxygenase-activating protein (FLAP)
reduces pulmonary vascular reactivity and pulmonary hypertension in hypoxic
rats (47), with the most intense FLAP immunoreactivity in pulmonary arterial
endothelial cells. Also, chronic hypoxia activates 15-LO in neonatal rabbit lungs,
with increased 15-LO being found in both pulmonary arterial endothelial cells and
SMCs (52). Thus, it is possible that an array of lipoxygenases is increased by

12-LO and pulmonary hypertension LCMP—00114-2005.R1
17
hypoxia and each of them has different cellular effects on different cell types. It is
also possible that the type of lipoxygenase that participates in hypoxia is species-
specific, since rabbit 15-LO has 70% homology with rat 12-LO (51).
We conclude that the 12-LO pathway and its metabolic product, 12-HETE,
participate in hypoxia-induced rat PASMC proliferation in vitro, most likely via the
ERK1/ERK2 MAPK pathway and apparently without the involvement of p38
MAPK. We speculate that the 12-LO pathway also contributes to the
development of hypoxia-induced pulmonary hypertension in vivo, but further
studies will be necessary to substantiate this.
Grants: This study was supported by NIH/NHLBI HL32723-15 (BLF) and a
Charlton Award from Tufts University School of Medicine (IRP).

12-LO and pulmonary hypertension LCMP—00114-2005.R1
18
REFERENCES
1. Arjona AA, Hsu CA, Wrenn DS, and Hill NS. Effects of natriuretic peptides on vascular smooth-muscle cells derived from different vascular beds. Gen Pharmacol 28: 387-392, 1997. 2. Bleich D, Chen S, Gu JL, Thomas L, Scott S, Gonzales N, Natarajan R, and Nadler JL. Interleukin-1 beta regulates the expression of a leukocyte type of 12-lipoxygenase in rat islets and RIN m5F cells. Endocrinology 136: 5736-5744, 1995. 3. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254, 1976. 4. Brinkman HJ, van Buul-Wortelboer MF, and van Mourik JA. Involvement of cyclooxygenase- and lipoxygenase-mediated conversion of arachidonic acid in controlling human vascular smooth muscle cell proliferation. Thromb Haemost 63: 291-297, 1990. 5. Chen XS, Kurre U, Jenkins NA, Copeland NG, and Funk CD. cDNA cloning, expression, mutagenesis of C-terminal isoleucine, genomic structure, and chromosomal localizations of murine 12-lipoxygenases. J Biol Chem 269: 13979-13987, 1994. 6. Chen Y, Hagmann W, and Honn KV. Regulation of 12(S)-HETE production in tumor cells. In: Eicosanoids and other bioactive lipids in cancer, inflammation, and radiation injury, edited by Konn KV. New York: Plenum press, 1997, p. 159-166. 7. Christou H, Morita T, Hsieh CM, Koike H, Arkonac B, Perrella MA, and Kourembanas S. Prevention of hypoxia-induced pulmonary hypertension by enhancement of endogenous heme oxygenase-1 in the rat. Circ Res 86: 1224-1229, 2000. 8. Cogo A, Napolitano G, Michoud MC, Barbon DR, Ward M, and Martin JG. Effects of hypoxia on rat airway smooth muscle cell proliferation. J Appl Physiol 94: 1403-1409, 2003. 9. Cooper AL and Beasley D. Hypoxia stimulates proliferation and interleukin-1alpha production in human vascular smooth muscle cells. Am J Physiol 277: H1326-1337, 1999. 10. Denhardt DT. Signal-transducing protein phosphorylation cascades mediated by Ras/Rho proteins in the mammalian cell: the potential for multiplex signalling. Biochem J 318 (Pt 3): 729-747, 1996. 11. Ding XZ, Tong WG, and Adrian TE. 12-lipoxygenase metabolite 12(S)-HETE stimulates human pancreatic cancer cell proliferation via protein tyrosine phosphorylation and ERK activation. Int J Cancer 94: 630-636, 2001. 12. Ding XZ, Tong WG, and Adrian TE. Cyclooxygenases and lipoxygenases as potential targets for treatment of pancreatic cancer. Pancreatology 1: 291-299, 2001. 13. Duan C. The chemotactic and mitogenic responses of vascular smooth

12-LO and pulmonary hypertension LCMP—00114-2005.R1
19
muscle cells to insulin-like growth factor-I require the activation of ERK1/2. Mol Cell Endocrinol 206: 75-83, 2003. 14. Eddahibi S, Fabre V, Boni C, Martres MP, Raffestin B, Hamon M, and Adnot S. Induction of serotonin transporter by hypoxia in pulmonary vascular smooth muscle cells. Relationship with the mitogenic action of serotonin. Circ Res 84: 329-336, 1999. 15. Fanburg BL and Lee SL. A role for the serotonin transporter in hypoxia-induced pulmonary hypertension. J Clin Invest 105: 1521-1523, 2000. 16. Frid MG, Aldashev AA, Dempsey EC, and Stenmark KR. Smooth muscle cells isolated from discrete compartments of the mature vascular media exhibit unique phenotypes and distinct growth capabilities. Circ Res 81: 940-952, 1997. 17. Gu JL, Pei H, Thomas L, Nadler JL, Rossi JJ, Lanting L, and Natarajan R. Ribozyme-mediated inhibition of rat leukocyte-type 12-lipoxygenase prevents intimal hyperplasia in balloon-injured rat carotid arteries. Circulation 103: 1446-1452, 2001. 18. Herbertsson H and Hammarstrom S. Cytosolic 12(S)-hydroxy-5,8,10,14-eicosatetraenoic acid binding sites in carcinoma cells. Adv Exp Med Biol 400A: 287-293, 1997. 19. Jin N, Hatton N, Swartz DR, Xia X, Harrington MA, Larsen SH, and Rhoades RA. Hypoxia activates jun-N-terminal kinase, extracellular signal-regulated protein kinase, and p38 kinase in pulmonary arteries. Am J Respir Cell Mol Biol 23: 593-601, 2000. 20. Karamsetty MR, Pietras L, Klinger JR, Lanzillo JJ, Leiter JC, Ou LC, and Hill NS. The role of endothelin-1 in strain-related susceptibility to develop hypoxic pulmonary hypertension in rats. Respir Physiol 128: 219-227, 2001. 21. Kayyali US, Pennella CM, Trujillo C, Villa O, Gaestel M, and Hassoun PM. Cytoskeletal changes in hypoxic pulmonary endothelial cells are dependent on MAPK-activated protein kinase MK2. J Biol Chem 277: 42596-42602, 2002. 22. Klinger JR, Warburton RR, Pietras L, and Hill NS. Brain natriuretic peptide inhibits hypoxic pulmonary hypertension in rats. J Appl Physiol 84: 1646-1652, 1998. 23. Liu B, Maher RJ, Hannun YA, Porter AT, and Honn KV. 12(S)-HETE enhancement of prostate tumor cell invasion: selective role of PKC alpha. J Natl Cancer Inst 86: 1145-1151, 1994. 24. Liu B, Timar J, Howlett J, Diglio CA, and Honn KV. Lipoxygenase metabolites of arachidonic and linoleic acids modulate the adhesion of tumor cells to endothelium via regulation of protein kinase C. Cell Regul 2: 1045-1055, 1991. 25. McDonnell M, Davis W, Jr., Li H, and Funk CD. Characterization of the murine epidermal 12/15-lipoxygenase. Prostaglandins Other Lipid Mediat 63: 93-107, 2001. 26. Meyrick B and Reid L. The effect of chronic hypoxia on pulmonary arteries in young rats. Exp Lung Res 2: 257-271, 1981. 27. Natarajan R, Bai W, Rangarajan V, Gonzales N, Gu JL, Lanting L, and Nadler JL. Platelet-derived growth factor BB mediated regulation of 12-

12-LO and pulmonary hypertension LCMP—00114-2005.R1
20
lipoxygenase in porcine aortic smooth muscle cells. J Cell Physiol 169: 391-400, 1996. 28. Natarajan R, Esworthy R, Bai W, Gu JL, Wilczynski S, and Nadler J. Increased 12-lipoxygenase expression in breast cancer tissues and cells. Regulation by epidermal growth factor. J Clin Endocrinol Metab 82: 1790-1798, 1997. 29. Natarajan R, Gonzales N, Lanting L, and Nadler J. Role of the lipoxygenase pathway in angiotensin II-induced vascular smooth muscle cell hypertrophy. Hypertension 23: I142-147, 1994. 30. Natarajan R, Gu JL, Rossi J, Gonzales N, Lanting L, Xu L, and Nadler J. Elevated glucose and angiotensin II increase 12-lipoxygenase activity and expression in porcine aortic smooth muscle cells. Proc Natl Acad Sci U S A 90: 4947-4951, 1993. 31. Natarajan R, Pei H, Gu JL, Sarma JM, and Nadler J. Evidence for 12-lipoxygenase induction in the vessel wall following balloon injury. Cardiovasc Res 41: 489-499, 1999. 32. Natarajan R, Rosdahl J, Gonzales N, and Bai W. Regulation of 12-lipoxygenase by cytokines in vascular smooth muscle cells. Hypertension 30: 873-879, 1997. 33. Rao GN, Baas AS, Glasgow WC, Eling TE, Runge MS, and Alexander RW. Activation of mitogen-activated protein kinases by arachidonic acid and its metabolites in vascular smooth muscle cells. J Biol Chem 269: 32586-32591, 1994. 34. Reddy MA, Adler SG, Kim YS, Lanting L, Rossi J, Kang SW, Nadler JL, Shahed A, and Natarajan R. Interaction of MAPK and 12-lipoxygenase pathways in growth and matrix protein expression in mesangial cells. Am J Physiol Renal Physiol 283: F985-994, 2002. 35. Reddy MA, Kim YS, Lanting L, and Natarajan R. Reduced growth factor responses in vascular smooth muscle cells derived from 12/15-lipoxygenase-deficient mice. Hypertension 41: 1294-1300, 2003. 36. Reddy MA, Thimmalapura PR, Lanting L, Nadler JL, Fatima S, and Natarajan R. The oxidized lipid and lipoxygenase product 12(S)-hydroxyeicosatetraenoic acid induces hypertrophy and fibronectin transcription in vascular smooth muscle cells via p38 MAPK and cAMP response element-binding protein activation. Mediation of angiotensin II effects. J Biol Chem 277: 9920-9928, 2002. 37. Reddy RG, Yoshimoto T, Yamamoto S, Funk CD, and Marnett LJ. Expression of porcine leukocyte 12-lipoxygenase in a baculovirus/insect cell system and its characterization. Arch Biochem Biophys 312: 219-226, 1994. 38. Szekeres CK, Tang K, Trikha M, and Honn KV. Eicosanoid activation of extracellular signal-regulated kinase1/2 in human epidermoid carcinoma cells. J Biol Chem 275: 38831-38841, 2000. 39. Szekeres CK, Trikha M, and Honn KV. 12(S)-HETE, pleiotropic functions, multiple signaling pathways. Adv Exp Med Biol 507: 509-515, 2002. 40. Szekeres CK, Trikha M, Nie D, and Honn KV. Eicosanoid 12(S)-HETE activates phosphatidylinositol 3-kinase. Biochem Biophys Res Commun 275:

12-LO and pulmonary hypertension LCMP—00114-2005.R1
21
690-695, 2000. 41. Tang DG, Chen YQ, and Honn KV. Arachidonate lipoxygenases as essential regulators of cell survival and apoptosis. Proc Natl Acad Sci U S A 93: 5241-5246, 1996. 42. Tang DG, Grossi IM, Tang KQ, Diglio CA, and Honn KV. Inhibition of TPA and 12(S)-HETE-stimulated tumor cell adhesion by prostacyclin and its stable analogs: rationale for their antimetastatic effects. Int J Cancer 60: 418-425, 1995. 43. Tang DG, Renaud C, Stojakovic S, Diglio CA, Porter A, and Honn KV. 12(S)-HETE is a mitogenic factor for microvascular endothelial cells: its potential role in angiogenesis. Biochem Biophys Res Commun 211: 462-468, 1995. 44. Tang DG, Tarrien M, Dobrzynski P, and Honn KV. Melanoma cell spreading on fibronectin induced by 12(S)-HETE involves both protein kinase C- and protein tyrosine kinase-dependent focal adhesion formation and tyrosine phosphorylation of focal adhesion kinase (pp125FAK). J Cell Physiol 165: 291-306, 1995. 45. Timar J, Bazaz R, Kimler V, Haddad M, Tang DG, Robertson D, Tovari J, Taylor JD, and Honn KV. Immunomorphological characterization and effects of 12-(S)-HETE on a dynamic intracellular pool of the alpha IIb beta 3-integrin in melanoma cells. J Cell Sci 108 (Pt 6): 2175-2186, 1995. 46. Voelkel N and Reeves J. Primary pulmonary hypertension. In: Pulmonary vascular diseases, edited by Moser K. New York: Marcel Dekker, 1997, p. 573-649. 47. Voelkel NF, Tuder RM, Wade K, Hoper M, Lepley RA, Goulet JL, Koller BH, and Fitzpatrick F. Inhibition of 5-lipoxygenase-activating protein (FLAP) reduces pulmonary vascular reactivity and pulmonary hypertension in hypoxic rats. J Clin Invest 97: 2491-2498, 1996. 48. Welsh DJ, Scott P, Plevin R, Wadsworth R, and Peacock AJ. Hypoxia enhances cellular proliferation and inositol 1,4, 5-triphosphate generation in fibroblasts from bovine pulmonary artery but not from mesenteric artery. Am J Respir Crit Care Med 158: 1757-1762, 1998. 49. Wen Y, Gu J, Liu Y, Wang PH, Sun Y, and Nadler JL. Overexpression of 12-lipoxygenase causes cardiac fibroblast cell growth. Circ Res 88: 70-76, 2001. 50. Yang X, Sheares KK, Davie N, Upton PD, Taylor GW, Horsley J, Wharton J, and Morrell NW. Hypoxic induction of cox-2 regulates proliferation of human pulmonary artery smooth muscle cells. Am J Respir Cell Mol Biol 27: 688-696, 2002. 51. Yoshimoto T, Suzuki H, Yamamoto S, Takai T, Yokoyama C, and Tanabe T. Cloning and sequence analysis of the cDNA for arachidonate 12-lipoxygenase of porcine leukocytes. Proc Natl Acad Sci U S A 87: 2142-2146, 1990. 52. Zhu D, Medhora M, Campbell WB, Spitzbarth N, Baker JE, and Jacobs ER. Chronic hypoxia activates lung 15-lipoxygenase, which catalyzes production of 15-HETE and enhances constriction in neonatal rabbit pulmonary arteries. Circ Res 92: 992-1000, 2003.

12-LO and pulmonary hypertension LCMP—00114-2005.R1
22
Figure legends:
Fig 1. Chronic hypoxia up-regulates 12-LO in rat lung homogenates. A) RT-
PCR for 12-LO mRNA shows up-regulation in hypoxic rat lungs. For internal
control, we performed RT-PCR for tubulin, together with an internal standard for
tubulin (N=normoxia, H=hypoxia). B) Western blot analysis demonstrates that 12-
LO protein is up-regulated in hypoxic rat lung homogenates compared with
normoxic controls. C) Densitometric analysis of Western blot. N = 5 animals per
group, *p<0.05.
Fig 2. Increased 12-LO immunostaining in hypoxic rat lung. Immunostaining
with an antibody to leukocyte-type 12-LO and H and E counterstaining of normoxic
lung (A, B, C) compared with hypoxic lung sections (D, E, F) (100X magnification).
There is intense immunostaining of bronchial epithelial cells at baseline (Fig 2 A,
arrow), with no change by hypoxia (Fig 2 B, arrow). Hypoxia was associated with
increased immunostaining of endothelial cells of large pulmonary arteries (Fig 2 B,
arrow head and C, arrow), smooth muscle cells and possibly endothelial cells of
medium size and small pulmonary arteries (Fig 2 F, arrows) and pneumocytes of
alveolar walls. Arrow heads from fig 2 C and F depict intense staining of alveolar
macrophages, unchanged by hypoxia.
Fig 3. Hypoxia stimulates PASMC proliferation. Hypoxia stimulated cell

12-LO and pulmonary hypertension LCMP—00114-2005.R1
23
proliferation as demonstrated by increased thymidine uptake in 0.1% and 10%
serum (A and B) and cell count (C) in rat PASMCs exposed to hypoxia, compared
with their normoxic controls. N = 6, *p<0.05.
Fig 4. Hypoxia up-regulates 12-LO protein in rat PASMCs in culture. A) 12-LO
immunoblot was compared with Western blot for actin. B) Densitometric ratio
demonstrated that 12-LO is up-regulated by hypoxia starting at 2 hours. N = 5,
*p<0.05. Normoxic PASMCs did not show an increase in 12-LO expression with
time in culture (results not shown).
Fig 5. 12-HETE stimulates cell proliferation of cultured rat PASMCs. A)
PASMCs were treated for 24 hours with 12-HETE at 10-7-1µM. In contrast, 15-
HETE has no effect on PASMCs. B) Pretreatment with PD98059 at 10µM 30 min
before the 12-HETE treatment (0.001-1µM), completely blocked cell proliferation.
N = 6, *p<0.05.
Fig 6. 12-HETE stimulates phosphorylation of ERK. PASMCs were treated with
0.1 µM 12-HETE for 5 to 90 minutes. Western blot and densitometric ratio of
phophorylated ERK and total ERK (A and B) showed that ERK phosphorylation
has a biphasic pattern. p38 MAPK phosphorylation is not affected by 12-HETE (C
and D). N = 5, *p<0.05 compared with time 0.
Fig 7. Hypoxia-induced PASMC proliferation is dependent on ERK. Hypoxia-

12-LO and pulmonary hypertension LCMP—00114-2005.R1
24
induced stimulation of DNA synthesis was blocked by the MEK inhibitor (U0126,
5µM), while the p38 MAPK inhibitor (SB202190 10µM) had no effect. N = 6,
*p=0.02 compared with vehicle-treated hypoxic PASMCs.
Fig 8. 12-LO participates in hypoxia-induced PASMC proliferation. Hypoxia-
induced PASMC proliferation was blocked by the specific 12-LO inhibitor baicalein.
N = 6, *p<0.05 between normoxia and hypoxia, †p<0.05 between baicalein-treated
and vehicle-treated hypoxic experiments.

12-LO and pulmonary hypertension LCMP—00114-2005.R1
25
Figure 1 A, B, C
- 12-LO immunoblot - IgG
- 12-LO PCR
- Tubulin - Internal Standard
N N N H H A
B
Normoxia Hypoxia
Den
sito
met
ric
Rat
io(1
2-L
O/Ig
G)
0
1
2
3
(3 weeks)
*C

12-LO and pulmonary hypertension LCMP—00114-2005.R1
26
Figure 2 A, B, C, D, E, F

12-LO and pulmonary hypertension LCMP—00114-2005.R1
27
Figure 3 A, B, C
A
B
C
Days
0 1 2 3 4 5 6 7 8 9 10
Cel
l Nu
mb
erx1
000/
cm2
0
10
20
30
40Normoxia, 0.1%SerumHypoxia 3%O2, 0.1%Serum
*
* * * *
[3H
]Th
ymid
ine
Up
take
CP
M/w
ell (
x100
0)
0
4
8
12
16Normoxia, 0.1%SerumHypoxia (3%O2), 0.1%Serum
Day 1 Day 3
*
*
[3H
]Th
ymid
ine
Up
take
CP
M/w
ell (
x100
0)
0
20
40
60
80Normoxia, 10%SerumHypoxia 3%O2, 10%Serum
Day 1 Day 3
*
*
**

12-LO and pulmonary hypertension LCMP—00114-2005.R1
28
Figure 4 A, B
0
Den
sito
met
ric
Rat
io(1
2-L
O/A
ctin
)
0
2
4
6 Rat PASMCs
0 1h 2h 24h 48h
* **
- 12-LO immunoblot - Actin
A
B
0 1 1 2 2 24 24 48 hours

12-LO and pulmonary hypertension LCMP—00114-2005.R1
29
Figure 5 A, B
[3H
]Thy
mid
ine
Upt
ake
CP
M/w
ell (
x100
0)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
10 -7 10 -6 10 -5 10 -4 10 -3 10 -2 0.1 1 10-3 10 -2 0.1
12-HETE (µM) 15-HETE (µM)
Cont
rol
* ** * *
*
A
B
12 HETE (µM)
[3H
]Th
ymid
ine
Up
take
CP
M/w
ell (
x100
0)
0
2
4
6
ControlPD98059
0 0.001 0.01 0.1 1
* * **

12-LO and pulmonary hypertension LCMP—00114-2005.R1
30
Figure 6 A, B, C, D
- pERK1/2 immunoblot - ERK1/2 immunoblot
- p p38 MAPK immunoblot - p38 MAPK immunoblot
A
B
C
0 5 10 15 30 60 90 min
Minutes
0 5 10 15 30 60 90
Rat
io(p
P38
MA
PK
/P38
MA
PK
)
0.0
0.4
0.8
1.2D
Minutes
0 5 10 15 30 60 90
Rat
io(p
ER
K/E
RK
)
0.0
0.4
0.8
1.2
1.6
2.0
* *
* *
*
0 5 1 15 30 60 90 min
12-LO and pulmonary hypertension LCMP—00114-2005.R1
31
Figure 7
0.1%Serum Vehicle U0126 SB202190
[3 H]
Thym
idin
e U
ptak
eC
PM
/wel
l (x1
000)
0.0
0.2
0.4
0.6
0.8
1.0
1.2 NormoxiaHypoxia
*

12-LO and pulmonary hypertension LCMP—00114-2005.R1
32
Figure 8
Baicalein (µM)
0 0.5 1 2 3
[3 H]T
hym
idin
e U
pta
keC
PM
/wel
l (x
1000
)
0
1
2
3
4
5
6NormoxiaHypoxia
**
** *
†
Related Documents







![BMC Cancer BioMed Central · 2017. 8. 27. · intrinsic radioresistance; tumor-cell proliferation; and hypoxia [31]. Forty-two percent of prostatic tumors have abnormal PTEN/Akt expression](https://static.cupdf.com/doc/110x72/60d1bf9958aa13749c035126/bmc-cancer-biomed-central-2017-8-27-intrinsic-radioresistance-tumor-cell-proliferation.jpg)



