REVIEW RNAi pathway integration in Caenorhabditis elegans development Sadegh Azimzadeh Jamalkandi & Ali Masoudi-Nejad Received: 30 April 2011 /Revised: 12 June 2011 /Accepted: 28 June 2011 # Springer-Verlag 2011 Abstract In this review, the pathways involving small RNAs are provided followed by a new and updated network that illustrates their interplay with diverse cellular mechanisms in Caenorhabditis elegans. The RNA silencing pathways are now recognized as key factors that connect together the many variations in biological processes, including transcrip- tional gene regulation, post-transcriptional gene silencing, translational gene silencing, apoptosis, meiosis, and antiviral defense. The utilization of small RNAs represents a specific, energy conserving, and fast mechanism of gene regulation via a core system known as RNA interference. Keywords RNAi . Integrated pathway . Silencing pathways . Small RNA . Non-coding RNA . MicroRNA Introduction At present, RNA interference (RNAi) is regarded as a strong screening tool in molecular cell biology (Meister and Tuschl 2004). This phenomenon has a short but eventful history representing its exceedingly speedy development which has indirectly helped researchers to clarify the unidentified genes via knocking down/out cellular genes to detect their function. Seemingly different levels of central dogma are under the strict control of RNAi mechanism. In addition, RNAi mechanism has diverse functions within cells, including controlling the expression of critical genes, providing an antiviral guard against invasive molecules, chromatin remodeling, and transgene silencing, which are influential but not easily understood functions (Dorer and Henikoff 1994; Ketting et al. 1999; Tabara et al. 1999; Grishok et al. 2005; Lu et al. 2005; Wassenegger 2005). Cell- and organism-scale gene regu- lations seem to be the most fascinating functions of RNAi. In the former case, the fate of the single cell and growth are brought under control but in the latter, the organism is headed toward its potential growth and developmental phases (Fig. 1). RNAi phenomenon is generally placed in the regula- tory RNome category in which small RNAs are the triggers of various silencing pathways. Interestingly, small RNA pathways, such as exogenous RNAi (exo- RNAi), endogenous RNAi (endo-RNAi) (Lee et al. 2006), microRNA (Kim 2005), 21U-RNA (Ruby et al. 2006; Batista et al. 2008), and inverted repeat-associated small RNA pathways (Dorer and Henikoff 1994; Klenov et al. 2007) are all in interplay with RNAi and as well as one another. Their functions can be generally classified into three categories: (1) defending against genome invaders, (2) hosting gene regulation at the transcriptional and post-transcriptional levels, and (3) signaling and inheriting the silencing messages systematically. First, maintaining genome stability against viral infec- tions or jumping elements could be a very critical aspect of silencing in organisms to survive and develop as an innate immunity. These invaders are capable of destabilizing chromatin, causing mutations, changing an individual’ s fitness, and survival within the population. Second is regulating the host’ s expression system through silencing in order to regulate genes at the transcriptional (DNA methylation and histone modification) and post- transcriptional levels (target mRNA degradation or transla- S. Azimzadeh Jamalkandi : A. Masoudi-Nejad (*) Laboratory of Systems Biology and Bioinformatics (LBB), Institute of Biochemistry and Biophysics and COE in Biomathematics, University of Tehran, Tehran, Iran e-mail: [email protected] URL: http://LBB.ut.ac.ir Funct Integr Genomics DOI 10.1007/s10142-011-0236-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REVIEW
RNAi pathway integration in Caenorhabditis elegansdevelopment
Sadegh Azimzadeh Jamalkandi & Ali Masoudi-Nejad
Received: 30 April 2011 /Revised: 12 June 2011 /Accepted: 28 June 2011# Springer-Verlag 2011
Abstract In this review, the pathways involving small RNAsare provided followed by a new and updated network thatillustrates their interplay with diverse cellular mechanisms inCaenorhabditis elegans. The RNA silencing pathways arenow recognized as key factors that connect together themany variations in biological processes, including transcrip-tional gene regulation, post-transcriptional gene silencing,translational gene silencing, apoptosis, meiosis, and antiviraldefense. The utilization of small RNAs represents a specific,energy conserving, and fast mechanism of gene regulationvia a core system known as RNA interference.
Keywords RNAi . Integrated pathway. Silencingpathways . Small RNA . Non-coding RNA .MicroRNA
Introduction
At present, RNA interference (RNAi) is regarded as astrong screening tool in molecular cell biology (Meister andTuschl 2004). This phenomenon has a short but eventfulhistory representing its exceedingly speedy developmentwhich has indirectly helped researchers to clarify theunidentified genes via knocking down/out cellular genesto detect their function. Seemingly different levels ofcentral dogma are under the strict control of RNAimechanism. In addition, RNAi mechanism has diversefunctions within cells, including controlling the expression
of critical genes, providing an antiviral guard againstinvasive molecules, chromatin remodeling, and transgenesilencing, which are influential but not easily understoodfunctions (Dorer and Henikoff 1994; Ketting et al. 1999;Tabara et al. 1999; Grishok et al. 2005; Lu et al. 2005;Wassenegger 2005). Cell- and organism-scale gene regu-lations seem to be the most fascinating functions of RNAi.In the former case, the fate of the single cell and growth arebrought under control but in the latter, the organism isheaded toward its potential growth and developmentalphases (Fig. 1).
RNAi phenomenon is generally placed in the regula-tory RNome category in which small RNAs are thetriggers of various silencing pathways. Interestingly,small RNA pathways, such as exogenous RNAi (exo-RNAi), endogenous RNAi (endo-RNAi) (Lee et al.2006), microRNA (Kim 2005), 21U-RNA (Ruby et al.2006; Batista et al. 2008), and inverted repeat-associatedsmall RNA pathways (Dorer and Henikoff 1994; Klenovet al. 2007) are all in interplay with RNAi and as well asone another. Their functions can be generally classifiedinto three categories: (1) defending against genomeinvaders, (2) hosting gene regulation at the transcriptionaland post-transcriptional levels, and (3) signaling andinheriting the silencing messages systematically.
First, maintaining genome stability against viral infec-tions or jumping elements could be a very critical aspect ofsilencing in organisms to survive and develop as an innateimmunity. These invaders are capable of destabilizingchromatin, causing mutations, changing an individual’sfitness, and survival within the population. Second isregulating the host’s expression system through silencingin order to regulate genes at the transcriptional (DNAmethylation and histone modification) and post-transcriptional levels (target mRNA degradation or transla-
S. Azimzadeh Jamalkandi :A. Masoudi-Nejad (*)Laboratory of Systems Biology and Bioinformatics (LBB),Institute of Biochemistry and Biophysics and COEin Biomathematics, University of Tehran,Tehran, Irane-mail: [email protected]: http://LBB.ut.ac.ir
Funct Integr GenomicsDOI 10.1007/s10142-011-0236-1

tional repression). Finally, transferring the silencing signalto near-far cells coupled with activating DNA methylationand histone modifications can pass the silencing signal ontothe offspring so as to ensure the avoidance of the potentialdangers associated with epigenetic inheritance. Additionally,impaired chromosomes during meiosis are hyper-methylated via small non-coding RNAs (ncRNAs), aspart of the meiotic silencing of unsynapsed chromosomespathway (Maine et al. 2005) which will be discussed laterin greater detail.
Small RNA-based silencing is a sequence-specific andhomology-dependent phenomenon; most of which isdouble-stranded RNA (dsRNA) dependent and is processedinto small dsRNAs by the Dicer protein. The resultingsmall dsRNAs are regarded as substrates of the silencingcomplex, which includes one of the Argonaute proteins.The silencing complex takes one strand of the small dsRNAand screens the cellular mRNAs to find the complementstrands. In the case of finding any complement mRNA,mediate its cleavage or suppresses its translation. SmallRNA-based silencing pathways in association with theRNAi pathway are discussed in the subsequent sections. Insystems biology, understanding the components of thepathways and also discovering the unrevealed componentswill help us to understand the interplay of the pathways,thus enabling the depiction of a super-pathway. In thefuture, by elucidating the exact super-pathways of organ-isms, we would be able to understand the system’s responseto its surroundings, such as the environment, drugs, andpathogens. RNAi is a distinct example of such critical
pathways that play a core role in the silencing pathways.In this review, we bring together information regardingRNAi-derived small RNA silencing phenomenon and, forthe first time, simplify the phenomenon by designingintegrated super-pathways with respect to the ribo-regulatory networks.
Exogenous RNAi/virus-induced RNAi; similarbut distinct
Following the discovery of long dsRNA-induced RNAi inCaenorhabditis elegans (Fire et al. 1998), further inves-tigations have led to a deeper understanding of the silencingprocess. Long dsRNAs are the major triggers of exo/endo-RNAi pathways. These long exogenous dsRNAs areprocessed by the Dicer complex (Chu and Rana 2007;Rana 2007) which creates small interfering RNAs(siRNAs), 21–23 nucleotide (nt) long, with the 2 nt over-hangs at the 3′ termini and 5′ phosphate groups naturally(Hannon and Rossi 2004). On the other hand, RDE-4(RNAi deficient, dsRNA binding protein), DCR-1 (Dicerrelated, belonging to the dsRNA specific RNase III familyof proteins), RDE-1 (RNAi deficient, PIWI/Argonaute/Zwille (PAZ)–PIWI family protein), and Dicer-relatedhelicase (DRH)-1 and DRH-2 (conserved DexH box heli-case 1 and 2, respectively) are the major components of theexo-RNAi Pathway (Grishok and Mello 2002; Tabara et al.2002) that make a complex in concert. Interestingly, co-purification of RDE-1, RDE-4, and DRH1/2 with DCR-1has already been reported (Duchaine et al. 2006). RDE-4has a dsRNA binding motif, which can bind to long dsRNApreferentially (Tabara et al. 2002), and is critical inexogenous dsRNA recognition and DCR-1 recruitment(Tabara et al. 2002; Boisvert and Simard 2009). In theDicer complex, RDE-4 interacts with RDE-1, DCR-1,DRH-1, and DRH-2 (Tabara et al. 2002). DCR-1 isresponsible for processing dsRNA into siRNAs (Meisterand Tuschl 2004). RDE-1, from the Argonaute family ofproteins (Tabara et al. 1999), is associated with primarysiRNAs, which are required for the next steps (Grishok2005; Boisvert and Simard 2009). DRH helicase proteinfamily is found in many diverse organisms ranging fromArchaea to humans as a conserved protein. The DRHfamily is most likely involved in the joining of primarysiRNAs to the Argonaute proteins and the amplification ofsecondary siRNAs or targeting/cleavage of the mRNAs(Duchaine et al. 2006; Sijen et al. 2007; Lu et al. 2009)(Fig. 2).
In the post-Dicer cleavage step, the guide strand(antisense strand) of the siRNA is loaded onto theArgonaute family protein in the small RNA-induced RNAsilencing complex (siRISC) which plays the template role
Fig. 1 Development plasticity affected by transcriptional and post-transcriptional silencing mechanism via intermediate siRNAmolecules
Funct Integr Genomics
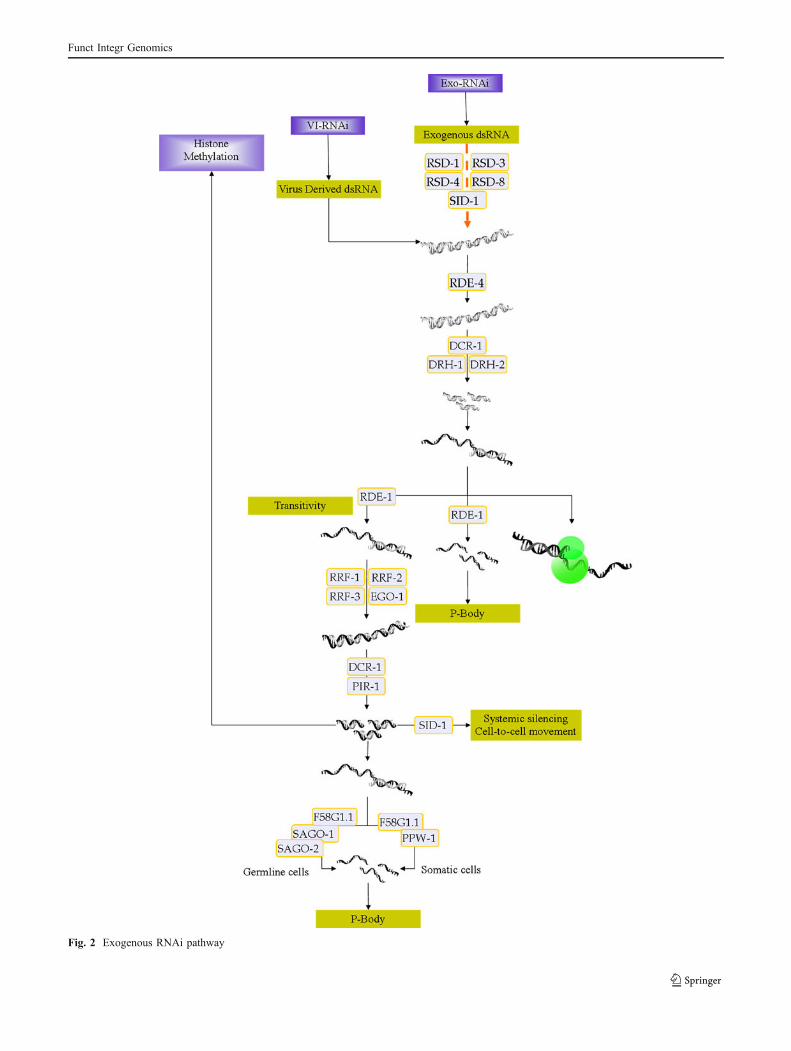
Fig. 2 Exogenous RNAi pathway
Funct Integr Genomics

in sequence-specific recognition of the target mRNA. C.elegans contains more than 26 Argonaute genes categorizedin three groups including Argonaute-like (with five mem-bers), Piwi-like (with three members), and Argonautproteins (with 18 members) (Hutvagner and Simard2008). Thermodynamic asymmetry and structural require-ments are the conditions of strand selection whereby theless thermodynamically stable positioned at its 5′ end isselected as the guide strand (Khvorova et al. 2003; Schwarzet al. 2003; Rana 2007). These features are used to produceefficient synthetic dsRNAs and siRNAs (Kim et al. 2005;Siolas et al. 2005). PIWI/Argonaute/Zwille (PAZ) and Piwiare the two domains of the Argonaute proteins. The 3′ and5′ overhangs of siRNAs are recognized by their RNAbinding domains (Ma et al. 2005; Parker et al. 2005).Argonaute proteins enfold siRNAs, pair with target mRNA,and finally slice it (Song and Joshua-Tor 2006; Boisvert andSimard 2009). The complement strand of the antisensesiRNA (sense or passenger strand) is degraded later.Subsequent to finding/binding of the mRNA target (baseparing), siRISC cleaves the target mRNA, 10–11 nt fromthe 5′ end of the guide siRNA strand. The result is specific-mRNA-OFF (specific-mRNA-silencing) and cellular deple-tion of target mRNA. The active siRISC complex will leavethe cleaved target for cellular exonucleases and continue toscreen the next targets.
Phylogenetic analysis shows three distinct groups ofArgonaute proteins as shown below (Faehnle and Joshua-Tor2007; Siomi and Siomi 2009) (Table 1):
1. AGO subfamily (AGO clade)2. PIWI subfamily (P element induced wimpy testis)3. WAGO subfamily (worm-specific Argonaute)
Accordingly, RDE-1 can interact with sense and anti-sense strands of the primary-exo-siRNAs (Yigit et al. 2006)and is the primary or first-line Argonaute of exo-RNAi path(Yigit et al. 2006; Boisvert and Simard 2009). Structuraland experimental analyses confirm the endonucleolyticcapability of RDE-1 essential for exo-siRISC activity (Toliaand Joshua-Tor 2007) with regard to the cleavage of thetarget mRNAs. RDE-1 minimizes the off-target silencingreactions, which represent the adverse and side effects ofthe silencing mechanisms (Yigit et al. 2006).
Subsequent to primary siRNA production and thespecific-mRNA-OFF process by the RDE-1 Argonauteprotein, RNA-dependent RNA polymerase (RdRP)becomes active. Remarkably, results have demonstratedthat silencing is not narrowed to the triggered cells butactually spreads throughout the animal cells and subse-quent generations (Fire et al. 1998). This example ofsignal amplification has been confirmed by the presenceof the RdRP proteins (Smardon et al. 2000). RdRPproteins are important in fungi (Martens et al. 2002),
plants (Dalmay 2000; Vaistij 2002; Azimzadeh andMasoudi-Nejad 2009), yeast (Schramke and Allshire2003), and C. elegans (Sijen et al. 2001). They generatesecondary siRNAs to enrich the siRNA pool and intensifythe silencing signal (Wilkins et al. 2005; Sijen et al. 2007).This step is also known as signal amplification, recogniz-ing aberrant RNA molecules and synthesizing a dsRNAvia a primer dependent/independent process. After theoccurrence of base pairing between primary siRNAs andtarget mRNA, RdRP creates new dsRNAs. These newdsRNAs can start and enhance the second round of thesilencing process (Naqvi et al. 2009). The second roundcan reinforce the silencing signal received from primarysiRNAs (Siomi and Siomi 2009), leading to a veryefficient silencing system. In this step, new kinds ofsiRNAs are produced via transitivity by which they areoriginated from the upstream regions of the targetedsequence (Sijen et al. 2001). The first trigger sequence isthe most abundant signal for silencing, but as distanceincreases, the frequency of transitive siRNA decreases(Simmer et al. 2002).
Presumably, RNAi deficient-3 (RDE-3; a β-nucleotidyltransferase super family polymerase) polyadenylates the 3′termini of the target mRNA ideal for RdRP recruitment andsecondary siRNA production (Chen et al. 2005). It has beenreported that secondary siRNAs carry di/triphosphates attheir 5′ end (Pak and Fire 2007; Sijen et al. 2007)recognizable by the second-line Argonauts via the assis-tance of PIR-1 phosphatase (phosphatase interacting withRNA/RNP; an RNA-phosphatase homolog) (Boisvert andSimard 2009).
Enhancer of Glp-one (glp-1) (EGO-1), RNA-dependentRNA polymerase family (RRF)-1, RRF-2, and RRF-3 arethe four RdRPs of C. elegans, where EGO-1 is responsiblefor the adult germline exo-RNAi (Smardon et al. 2000;Maniar and Fire 2011). RRF-2 and RRF-3 are required forsoma and germline RNAi and transitivity (Sijen et al.2001). Perhaps there is competition between EGO-1 andRRF-1 (Simmer et al. 2002; Maniar and Fire 2011). RRF-1is essential for secondary siRNA production by somaticexo-RNAi (Sijen et al. 2001). RRF-3 interacting with thecomplex and the rrf-1 mutants results in a phenotypesimilar to the enhancer of the RNAi (ERI) mutant strain(eri-1) crucial to endo-RNAi (Duchaine et al. 2006; Lee etal. 2006; Boisvert and Simard 2009). In the primer-independent RdRP phenomenon where dsRNA-Dicercleavage is not required, 21 nt and single-strandedtriphosphorylated small RNAs are typically produced (Aokiet al. 2007; Pak and Fire 2007; Sijen et al. 2007; Siomi andSiomi 2009). The components and the mechanism of theprocess, in which the RdRP protein is directly involved inthe pathway, are still unknown. Seemingly, RdRP-generatedsecondary siRNAs are self-terminated, Dicer-non-processed,
Funct Integr Genomics

Table 1 Components of the small RNA silencing pathways including their functions
Components Description Pathway Ref
1 ALG-1 Argonaute-like gene-1 Chromatin remodeling, MIR Ding et al. (2005)
2 ALG-2 Argonaute-like gene-2 MIR Jannot et al. (2008)
3 CSR-1 Chromosome segregation and RNAi Endo-RNAi Rocheleau et al. (2008)
4 DCR-1 Dicer related-1 MSUD, MIR, Endo/Exo-RNAi,VI-RNAi, chromatin remodeling
Knight and Bass (2001);Tabara et al. (2002);Duchaine et al. (2006)
5 DRH-1 Dicer-related helicase-1 Endo/exo-RNAi, VI-RNAi Tabara et al. (2002)
6 DRH-2 Dicer-related helicase-2 Endo/exo-RNAi, VI-RNAi Tabara et al. (2002)
7 DRH-3 Dicer-related helicase-3 Endo-RNAi, VI-RNAi Nakamura et al. (2007)
8 DROSHA Nuclear RNase III MIR pathway Han et al. (2009)
9 EGO-1 Enhancer of glp-1 Exo-RNAi, MSUD Vought et al. (2005)
10 ERGO-1 Endogenous RNAi-deficient Argonaute-1 Endo/exo-RNAi, VI-RNAi Lu et al. (2009)
11 ERI-1 Enhancer of RNAi-1 Endo-RNAi Grishok (2005)
12 ERI-3 Enhancer of RNAi-3 Endo-RNAi Duchaine et al. (2006)
13 ERI-5 Enhancer of RNAi-5 Endo-RNAi Parker and Barford (2006)
14 ERI-6 Enhancer of RNAi-6 Negative regulator of exo-RNAi Fischer et al. (2008)
15 ERI-7 Enhancer of RNAi-7 Negative regulator of exo-RNAi Fischer et al. (2008)
16 EXP-5 Ran-GTPase cofactor dependant exporter MIR Naqvi et al. (2009)
17 F58G1.1 AGO mutant allele Germline and somatic exo-RNAi Hutvagner and Simard (2008)
18 HPL-1 Heterochromatin Protein Like-1 Chromatin remodeling Ouellet and Roy (2007)
19 MUT-7 Mutator-7 Chromatin remodeling Ketting et al. (1999);Tops et al. (2005)
20 MUT-8 Mutator-8 Chromatin remodeling Tops et al. (2005)
21 MUT-14 Mutator-14 Exo-RNAi, transposon silencingin germline
Tijsterman et al. (2002a, b)
22 PASHA dsRNA binding protein MIR Kadener et al. (2009)
23 PIR-1 RNA-phosphatase homolog-1, T23G7.5 Endo/exo-RNAi, VI-RNAi Duchaine et al. (2006)
24 PPW-1 PAZ/PIWI-related protein-1 Exo-RNAi, VI-RNAi Tijsterman et al. (2002a, b)
25 PPW-2 PAZ/PIWI-related protein-2 Chromatin remodeling Vastenhouw et al. (2003)
26 PRG-1 PIWI-related gene-1 21U-RNA Batista et al. (2008)
27 RDE-1 RNAi deficient-1 Exo-RNAi, VI-RNAi,chromatin remodeling
Steiner et al. (2009)
28 RDE-2 RNAi deficient-2 Chromatin remodeling Tops et al. (2005)
29 RDE-3 RNAi deficient-3, β-nucleotidyltransferase Exo-RNAi, VI-RNAi Tabara et al. (1999)
30 RDE-4 RNAi deficient-4 Exo-RNAi, VI-RNAi,chromatin remodeling
Tabara et al. (2002)
31 RHA-1 RNA helicase A MSUD Robert et al. (2005)
32 RRF-1 RNA-dependent RNA polymerase family-1 Exo-RNAi, VI-RNAi,chromatin remodeling
Fischer et al. (2008)
33 RRF-2 RNA-dependent RNA polymerase family-2 Soma and germline RNAiand transitivity
Lee et al. (2006)
34 RRF-3 RNA-dependent RNA polymerase family-3 Soma and germline exo-RNAiand transitivity, endo-RNAi
Simmer et al. (2002)
35 RSD-1 RNA spreading deficient-1 RNA uptake Grishok (2005)
36 RSD-3 RNA spreading deficient-3 RNA uptake Grishok (2005)
37 RSD-4 RNA spreading deficient-4 RNA uptake Grishok (2005)
38 RSD-6 RNA spreading deficient-6 RNA uptake Grishok (2005)
39 RSD-8 RNA spreading deficient-8 RNA uptake Grishok (2005)
40 SAGO-1 Synthetic secondary siRNA defective AGO-1 Somatic exo-RNAi, VI-RNAi Farazi et al. (2008)
41 SAGO-2 Synthetic secondary siRNA defective AGO-2 Somatic exo-RNAi, VI-RNAi Farazi et al. (2008)
42 SID-1 Systemic RNAi deficient-1 RNA transporter Feinberg and Hunter (2003)
Funct Integr Genomics

and limited to transitivity (Aoki et al. 2007; Pak and Fire2007; Sijen et al. 2007; Siomi and Siomi 2009).
Consequently, target mRNA is used as a templatesequence in the formation of dsRNA by primary siRNAprimers (Lipardi et al. 2001). Secondary siRNAs carryingbi\triphosphates will be loaded onto the SAGO proteins(secondary siRNA defective Argonaute protein) during thenext phase of the process (Yigit et al. 2006). Second-lineArgonaute proteins known as SAGO bind to secondarysiRNAs triggering the downstream silencing events (Siomiand Siomi 2009). Among second-line Argonaut proteins,the PAZ/PIWI-related protein (PPW)-1 in the germlineRNAi, secondary siRNA defective Argonaute protein(SAGO)-1 and SAGO-2 in the somatic RNAi, and theArgonaute mutant allele (F58G1.1) in both germline andsomatic exo-RNAi (Yigit et al. 2006) bind to secondarysiRNAs produced via the RdRP phenomenon. SAGOproteins lack catalytic domains, which suggest that certainother components are required for signal cleavage andamplification (Yigit et al. 2006).
On the other hand, cell-to-cell and also cell-to-the-subsequent generations’ movement of silencing signalshave been reported in nematodes (Wilkins et al. 2005; Joseand Hunter 2007). Cell-to-cell signal movement in C.elegans depends on the systemic RNAi-deficient trans-membrane passive channel protein (systemic RNA interfer-ence defective-1, SID-1) (Winston et al. 2002; Feinberg andHunter 2003; Obbard et al. 2009). Silencing signals havebeen detected even after about 80 generations (Vastenhouwet al. 2006). This phenomenon is shared only by plants, butnot Drosophila and mammals (Boisvert and Simard 2009).SID-1 permits passive uptake of dsRNA (Winston et al.2002; Feinberg and Hunter 2003) and has been shown to beessential for systemic silencing of transgenic/exo-RNAi(Winston et al. 2002). In fact, five other RNA spreading-deficient (RSD) proteins including RSD-1, RSD-3, RSD-4,RSD-6, and RSD-8 have a crucial role with regard tosystemic silencing in C. elegans (Tijsterman et al. 2004).RSD-4 and RSD-8 are needed for total cellular uptake ofdsRNA, but RSD-1, RSD-3, and RSD-6 are needed just forthe initial step of dsRNA uptake from the gut into somaticcells, and not for the next steps of RNA spreading(Tijsterman et al. 2004). Systemic RNAi and transitivity,in association with the silencing pathways, function tomaintain the silencing signal (Simmer et al. 2002).
The ERI, ERI-1, and ERI-6/7 (Fischer et al. 2008),together with the endogenous RNAi-deficient ArgonautePIWI subfamily (ERGO)-1 act as negative regulators ofexo-RNAi (Grishok 2005; Ruby et al. 2006; Lu et al. 2009;Siomi and Siomi 2009). ERI-1 as a siRNase degradessiRNA via its 3′-overhang to prevent its further entry intoRISC, thus limiting the silencing duration, specificity, andfunction (Simmer et al. 2002).
Virus-induced RNAi
The virus-induced RNAi (VI-RNAi) path in worms is acrucial protection system affecting genomic invaders.Innate immune response against viruses has evolvedthrough the VI-RNAi pathway (Ding and Voinnet 2007).Viral dsRNAs are the key initiators (Wilkins et al. 2005;Boisvert and Simard 2009) of the pathway which starts inthe cytoplasm once exo-dsRNAs enter the cells. VI-RNAipathway is identified in plants (Wang and Metzlaff 2005),Drosophila (Galiana-Arnoux et al. 2006; Rij et al. 2006;Wang et al. 2006; Zambon et al. 2006), worms (Lu et al.2005; Schot et al. 2005; Wilkins et al. 2005), and fungi(Segers et al. 2007; Hammond et al. 2008), but not inSaccharomyces (Obbard et al. 2009). It is seemingly apathway common to many eukaryotes.
The VI-RNAi pathway in C. elegans overlaps withexo-RNAi (Lu et al. 2005; Schot et al. 2005; Wilkins et al.2005) in which ERGO-1 and DRH-2 are both its negativeregulators (Lu et al. 2009). Viral RNA biogenesis isindependent of DRH-1 and RDE-1 proteins but dependenton RDE-4 and DCR-1 (Lu et al. 2009). Therefore, bothexo-RNAi and VI-RNAi pathway share certain compo-nents that will be mentioned (Grishok 2005). DRH-1 actsdownstream of DCR-1 and RDE-4 in this pathway and isnot critical for viral RNA sensing (Lu et al. 2009). DRH-2is a negative regulator of VI-RNAi, while DRH-3 isnecessary for germline antiviral RNAi (Tabara et al. 2002;Nakamura et al. 2007). Moreover, the potential antiviralrole of the RDE-1 protein via viral replication has beenreported (Lu et al. 2005) during the first step of the exo-RNAi pathway (Grishok et al. 2000). DRH-2 is producedas a negative regulator of the antiviral RNAi path and isnot interconnected with other small non-coding RNApathways (Lu et al. 2009). DRH-1 is involved in virussensing after RDE-4 derived viRNAs (Ding and Voinnet2007) together with RDE-1 (Lu et al. 2009). This proteinis dispensable for exo-RNAi but necessary for VI-RNAi(Lu et al. 2009) (Fig. 2). Surprisingly, some viruses havevarious inhibitors known as viral suppressors of RNAi bywhich they can suppress the host silencing defense system(Li and Ding 2006; Ding and Voinnet 2007; Li and Ding2001) in plants (Li et al. 1999) and animals (Li et al.2002).
Endogenous RNAi
In addition to exo-RNAi, there is a pathway called endo-RNAi that targets endogenous dsRNAs derived fromoverlapping genes, long hairpins, and transposableelement-derived dsRNAs (Chung et al. 2008; Czech et al.2008; Okamura et al. 2008) (Fig. 3c). Generally, endo-
Funct Integr Genomics

siRNAs have similar features to exo-siRNAs (Obbard et al.2009), but in the former pathway, both primary andsecondary siRNAs are produced.
PIR-1 and DRH-3, highly conserved proteins, are inconcert with the essential enzyme DCR-1 (Ambros et al.2003a, b) and are crucial to the endo-RNAi pathwayshowing vital developmental roles. The DRH-3 proteinfunctions in maintaining germline chromosomal integrityand compaction (Duchaine et al. 2006; Nakamura et al.2007), promoting chromosome segregation, germlineproliferation, fertility (sperm function and gametogenesis)(Duchaine et al. 2006), and protecting against X-rayirradiation (Nakamura et al. 2007). It also functions incollaboration with DCR-1 and ERI proteins (Duchaine etal. 2006). Interestingly, ERI proteins operate together withRRF-3 (RdRP), DRH-3, and DCR-1 in the endo-RNAipathway (Duchaine et al. 2006). ERI-3 (siRNase; exonu-clease mentioned in exo-RNAi in more details) and ERI-5enhance the ERI-1/DCR-1 interaction in the ERI/DCR-1complex (Duchaine et al. 2006).
DRH-3 and also DRH-1 and DRH-2 have high homol-ogy degree and are essential for endo-RNAi (Duchaine etal. 2006). Co-purification of DRH-3 with DCR-1 haspreviously been reported (Duchaine et al. 2006), and ithas been found that DCR-1/RRF-3/ERI-1/ERI-3/ERI-5/DRH-3 proteins are involved in the ERI/DCR-1 complex(Bleys et al. 2006; Duchaine et al. 2006).
In secondary siRNA production by C. elegans, RdRPproteins use target mRNA to produce siRNAs in a primer-independent process (Siomi and Siomi 2009). RRF-3 is theRdRP protein in the Endo-RNAi pathway. The producedsiRNAs are then loaded onto the second-line Argonauteproteins. All secondary siRNAs have negative polarity andare included in reinforcement of the silencing signal (Aokiet al. 2007; Pak and Fire 2007; Sijen et al. 2007; Siomi andSiomi 2009).
ERGO-1 and chromosome segregation and RNAi-1 (CSR-1) are the other components of the endo-RNAi pathway(Kennedy et al. 2004; Duchaine et al. 2006; Hutvagner andSimard 2008) that play a similar role to that of RDE-1 in theexo-RNAi pathway as the first-line Argonaute proteins(Boisvert and Simard 2009). The Argonaute protein, CSR-1,functions in both the RNAi and chromosome segregationpathways (Yigit et al. 2006). ERGO-1 is another proteinessential to this pathway (Nakamura et al. 2007).
26- and 22G-RNA generation pathway
In large-scale sequencing of 21U-RNAs, a new class ofendogenous siRNAs named 26- and 22G-RNAs wasdiscovered (Ruby et al. 2006). In this pathway, 26G-RNAs are first produced as primary siRNAs which derive
the 22G-RNA generation as secondary siRNAs (Gu et al.2009). At the first step, RDE-4 (dsRNA binding), DCR-1,and RRF-3 (RdRP protein) are required to 26G-RNAproduction. Then, 26G-RNAs act as cofactors for ERGO-1 (Argonaute) and trigger downstream pathway of 22G-RNAs. Seemingly, RDE-4, DCR-1, RRF-3, and ERGO-1are essential in both 26- and 22G-RNA generation at thefirst step. But in the next, two routes can be modeled (Gu etal. 2009): WAGO-1/22G-RNA or CSR-1/22G-RNA path-ways (Vasale et al. 2010). Interestingly, RRF-1 and WAGO-1 are required for 22G-RNA production and not 26G-RNAproposing that they may role just in secondary siRNAgeneration (Fig. 6). This path acts against transposableelements and aberrant endogenous and exogenous tran-scripts. The second path including CSR-1/22G-RNA iscritical in chromosome segregation (Gu et al. 2009; Conineet al. 2010; Vasale et al. 2010). It has been reported thatduring embryogenesis of the nematode, small amount ofERGO-1/26G-RNAs can induce 22G-RNA biogenesis inorder to persist it into the further developmental stages.Then 22G-RNAs can intensify the silencing signal intransitivity and self-amplifying mode playing their criticalrole in development of the nematode (Sijen et al. 2001;Vasale et al. 2010). However, the detailed process of thepathway is under discovery yet. We know that these newsmall RNAs are distinct from others in their 5-phosphory-lated guanine which is the basis of their nomenclature.
MicroRNA
MiRNA research has been extensively studied in the lastfew decades, with initial investigations focusing on lin-4and let-7 in C. elegans. From an evolutionary point of view,miRNAs are conserved ribo-regulators that function in mosteukaryotes through the downregulation of specific targets atthe post-transcriptional level, either by target degradation ortranslational suppression (Ketting et al. 2001; Ambros et al.2003a, b). Currently, the core of these small RNA networksare the RNAi phenomena as represented by the firstidentified small ncRNAs (Lee et al. 1993).
Biogenesis of microRNAs (miRNAs) occurs via thecanonical pathway or the mirtron pathway (Naqvi et al.2009) (Fig. 3b). In canonical biogenesis, miRNAs aretranscribed from miRNA genes (MIR genes) by RNApolymerase II (Ambros et al. 2003a, b), which make upabout 1% of the total predicted genes in C. elegans (Grad etal. 2003; Lim et al. 2003; Bartel 2004; Naqvi et al. 2009)and are found on all chromosomes in independent tran-scription units as well as the intronic regions in worms(Bartel 2004; Chu and Rana 2007).
The initial transcript called the primary miRNA (pri-miRNA) contains a hairpin structure. The mature miRNA
Funct Integr Genomics

Fig. 3 a MSUD pathway, b MIR pathway, c endo-RNAi pathway, d 21U-RNA pathway
Funct Integr Genomics

sequence is buried (masked) in the pri-miRNA sequenceand is revealed through several processing levels. A 5′cap and 3′ poly-A tail are added to the pri-miRNAtranscript (Cai et al. 2004; Kim 2005). In C. elegans,miRNA maturation starts in the nucleus and terminates inthe cytoplasm. The Drosha (RNase III; ortholog ofDrosophila Drosha gene) along with the Pasha (dsRNAbinding protein) proteins are the initiators of this pathway(Lee et al. 2003). They recognize the pri-miRNA bycleaving it at the base of its stem structure and generate asmaller stem loop-miRNA molecule called the precursormiRNA (pre-miRNA; ~70 nt long) containing a 5′phosphate end and a 2-nt 3′ overhang feature (Chu andRana 2007). Pasha leads in directing Drosha to the cuttingsite of the 11th nucleotide at the base of the stem (Han etal. 2006). The RAS-related nuclear small GTPase proteincofactor dependent exporter protein (EXP-5) then trans-ports these processed pre-miRNAs out into the cytoplasm(Bohnsack et al. 2004).
In a second pathway, it has been reported that miRNAbiogenesis can originate from intron sequences, known asmirtrons (also known as miRtrons) (Lin et al. 2006;Okamura et al. 2007; Ruby et al. 2007; Naqvi et al.2009). These miRtrons can originate either from the non-coding genes (more than 80%) or the protein codingregions (Rodriguez et al. 2004; Kim and Kim 2007) andare able to produce hairpin structures with overhanging 3′2-nt recognizable by EXP-5 and Dicer endonuclease.Experimentally, it has not yet been proven if miRtronbiogenesis is independent of the Drosha and pasha proteins(Rodriguez et al. 2004; Wang et al. 2007; Naqvi et al.2009). EXP-5 transports these miRtron molecules into thecytoplasm, where subsequent downstream processing is thesame for both canonically and miRtron-derived miRNAs.Therefore, the pre-miRNA pool in the cytoplasm isregarded as homogenous. DCR-1 can act on these pre-miRNAs and generate mature miRNAs (~21–22 nt long)with 3′ 2-nt overhangs (Lee et al. 1993). After Dicercleavage, a small imperfect RNA duplex (miRNA–miRNA*), approximately 19–22 nt long is generated.Subsequently, the strand with less stability (thermodynamicstability) at its 5′ termini is loaded onto the Argonauteprotein (Khvorova et al. 2003) in the miRISC complex(microRNA-induced RNA silencing complex), thus acti-vating the complex to screen cytoplasmic mRNAs to findthe complementary target (a perfect or imperfect comple-ment) for the mature miRNA strand (Holmes and Cohen2007). Within the mature miRNA, there is a region of 2–8 nt at the 5′ end named the “seed” region, which is a keydeterminant of sequence specificity (Doench and Sharp2004). Target recognition studies show that in some casesone target mRNA has more than one miRNA recognitionsite enhancing the flexibility of the silencing phenomenon
as a stringent regulating mechanism (Lai 2002; Bartel 2004;Naqvi et al. 2009). Also, miRNA binding via RISC canenhance deadenylation/decapping of the target transcripts inC. elegans (Wakiyama et al. 2007).
Moreover, in various organisms, the negative regulation ofkey enzymes of the RNAi pathway, such as the Dicer andArgonaute (Xie et al. 2003), upregulation of translation(Vasudevan and Steitz 2007; Vasudevan et al. 2007), reverseimport of miRNAs into the nucleus (Hwang et al. 2007),introduces more complexity to this scenario and opens up newhorizons and possibilities for regulation (Naqvi et al. 2009).
Similar to the respective roles of RDE-1 and ERGO-1 inthe exo- and endo-RNAi pathways, Argonaute-like gene(ALG)-1 and ALG-2 are also Argonaute proteins involvedin the MIR pathway of C. elegans (Hutvágner et al. 2004;Hutvagner and Simard 2008).
The endogenous miRNA and exo-miRNA pathways arevarious sources of miRNAs in which are involved in thetranslational repression or degradation of target mRNA. Somereports indicate that miRNAs cause mRNA targets to relocateto P-bodies for further decay or storage (Boag et al. 2008;Noble et al. 2008). Such processing bodies contain concen-trated amounts of non-translated mRNAs and associatedproteins required for mRNA sequestration, translationalrepression, decapping, and degradation (Fillman and Lykke-Andersen 2005; Boag et al. 2008). The existence of suchbodies within the cells shows specificity and an acceleratedmode of action. P-bodies are probably a storage site forsequestered mRNAs, which are released under conditions ofstress (Bhattacharyya et al. 2006; Chu and Rana 2007).
Reports show that many of the conserved miRNAsperform crucial functions in the organism’s development(Shabalina and Koonin 2008). Prediction-based algorithmshave been used to predict their genes, mature miRNAs, andpossible targets within the cells; however, many of thesepredictions are yet to be confirmed (Lewis et al. 2005;Rajewsky 2006; Gaidatzis et al. 2007). The existence ofapproximately 150 miRNAs has already been established inC. elegans, all of which are crucial to neural development,synaptic function, neuromuscular junctions’ efficient sig-naling, cell lineage division, cuticle formation, and tumorgrowth in a strictly timed and coordinated manner. Lin-28,lin-41, lin-7, lsy-6, miR-84, miR-48, and miR-241 areexamples of such small regulators (Chatterjee et al. 2011).Besides, there are many families of C. elegans miRNAsthat are not essential for development or viability (Alvarez-Saavedra and Horvitz 2010).
21U-RNAs
In the procession of small RNA discoveries, followinginvestigations in germline cells, 21U-RNAs are the latest
Funct Integr Genomics

identified small RNAs of C. elegans (Fig. 3d) (Ruby et al.2006). As their names suggest, they carry a uridine mono-phosphate at their 5′ end (21U). They are exactly 21 nt longtailed with a 5′ uridine, sensitive to alkaline hydrolysis andphosphatase treatment, and are also capable of being asubstrate for the ligase enzyme. Furthermore, they aremodified with respect to their 2′ or 3′ oxygen (Ruby et al.2006). Presumably, these small RNAs are involved inchromatin remodeling, stabilizing, and transposon silencing.They are autonomously transcribed from the genome and nodsRNA is detected during the biogenesis of these smallRNAs, but it appears that the 5′ uridine acts as an indicatorfor their recognition by special components (Naqvi et al.2009). Most of them are mapped to intergenic and intronicregions (Naqvi et al. 2009). Additionally, two highlyconserved upstream sequences among 21U-RNAs of thesame species have been identified which are separated by alinker sequence (~34, 4, and 19–27 bp, respectively) and arerequired for transcription (Ruby et al. 2006; Houwing et al.2007) or act as promoter elements (Farazi et al. 2008). Nohomology has been detected between the 21U-RNAs andmRNAs, leading to a lack of mechanism in their probablerole in genome stability (Naqvi et al. 2009). Also, mature21U-RNAs are not conserved but their loci share aconserved motif in the regulatory sequence of their upstreamcis-acting elements, including the promoter region (Ruby etal. 2006). They are encoded by approximately 13,000 lociclustered on two parts of chromosome IV. Only one of the21U RNAs has been identified as being transposon-derived,suggesting that transposon silencing is not its main probablefunction (Batista et al. 2008). Detailed information regardingtheir biogenesis and function is not well-defined yet.
Reports indicate that these 21U-RNAs are the Piwi-interacting RNAs (piRNAs) which have been identified inmost animals and function in germline maintenance andfertility (Klattenhoff and Theurkauf 2008; Batista et al.2008). Characterized 21U-RNAs share several featuressimilar to the piRNAs of flies and mammals (Ruby et al.2006; Horwich et al. 2007; Ohara et al. 2007; Saito et al.2007). These small RNAs are exclusively detected in thegermline where the PIWI-related gene-1 protein is recog-nized as one of its components (Das et al. 2008; Batista etal. 2008). This pathway is independent of DCR-1 (Siomiand Siomi 2009). 21U-RNAs show diverse sequencecomposition with no distinct target sites (Aravin et al.2006; Girard et al. 2006).
Meiotic silencing of unsynapsed chromosomes
Meiotic silencing of unsynapsed chromosomes (MSUD) inC. elegans is another small RNA related pathway. Unpairedregions between chromosomes create loops which are
prone to methylation on H3K9, the result of which issilencing (Bean et al. 2004; Zaratiegui et al. 2007) (Fig. 3a).Interestingly, unsynapsed regions are transcribed intosingle-stranded RNA (ssRNA) which is then converted todsRNA by the activity of an RdRP protein known as EGO-1. Presumably, EGO-1 binds to ssRNA and recruits histonemodifiers, such as histone methyltransferase and RISC(Maine et al. 2005; Maniar and Fire 2011). This pathway isalso dependent on the DCR-1 and Drosha-1 proteinsprocessing details of which have not yet been unraveled(Maine et al. 2005; Zaratiegui et al. 2007). Recently, aninterlacement between the chromatin remodeling machin-ery, MSUD, and RNAi pathways has been reported (Robertet al. 2005; Costa et al. 2006). MSUD of mice can modifythe XY body besides the unsynapsed autosomes in themale/female germlines (Turner et al. 2005; Zaratiegui et al.2007).
Chromatin remodeling
Repeat-associated gene silencing, centromere and bi-directional transcriptional silencing, RNA-directed DNAmethylation, co-suppression, transgene array silencing,epigenetic modifications, chromatin-dependent gene silenc-ing, and position effect variegation, being the focus ofdiscussion in this section, are as a whole responsible formaintaining and protecting genome stability and assemblyand are also involved in the acute regulation of geneexpression (Lippman and Martienssen 2004).
First, it would be worthwhile to review the maincharacteristics and functions of the heterochromatin.Some heterochromatic regions are constantly silent, suchas the centromeres and telomeres, but some other regionshave some degrees of plasticity. In the constantly silentregions, there are special characters like repetitivesequences, which are also the origin of small RNAs thatmaintain the silent mode of the heterochromatin for thecell offspring cells epigenetically. The flexible process ofchromatin remodeling is also dependent on special smallRNAs. Besides, interplay between heterochromatinizationand silencing regulation has been investigated (Allo et al.2009; Luco et al. 2010; Tilgner and Guigó 2010). Certainassociations (arising from evolution) exist between het-erochromatin, DNA, and transcription, which are collec-tively known as the self-guard phenomenon that involvesDNAwithin the cell. DNA is packed in the center and atboth ends; it also assembles against the offensive orunnecessary fragments to decrease the usage of energy andreduce crossover. Repeat sequences in the genome areindications of offensive elements, which must therefore besilenced. So transposable elements, repetitive sequences,and repeat-associated transgenes should be silenced at the
Funct Integr Genomics

DNA or chromatin level. DNA and histone methylationare the major processes used for this purpose.
It has already been established that RNAi is involved inrepeat-associated gene silencing and heterochromatinremodeling. Regions of repetitive DNA generate repeat-associated siRNAs, critical for transcriptional silencing.Centromere is a heterochromatic DNA rich in repetitivesequences and is important for epigenetic modification,meiosis, and mitosis. Maintained epigenetic characteristicsof the centromere are very important with regard to the fateof the cell, and RNAi plays a crucial role in this process,endogenously.
DNA, histone, and non-histone proteins are boundtogether in the condensed structure of heterochromatinand are thus inaccessible for transcription (Jenuwein andAllis 2001) and are thus inaccessible for transcriptionthrough heterochromatinization (Richards and Elgin2002). In fact, heterochromatinization is involved inaltering chromatin structure, reducing rate of recombinationand making it transcriptionally less active (Richards andElgin 2002; Grewal and Moazed 2003). Obviously, H3Lys9(H3k9) histone modification is a shared feature of tran-scriptionally silent heterochromatin regions (Jenuwein2001; Peters et al. 2003; Buhler and Moazed 2007). Allthe epigenetic regulation of the genome switching fromeuchromatin to heterochromatin and vice versa has manykey regulatory and transcriptionally alterations in organ-ism’s development and differentiation as a response ofgenome epigenetic regulation. One such triggers are thesmall ncRNAs, which work at the two distinct levels ofDNA and histone.
Studies show that transcriptionally silent elementstranscribe to a single-stranded RNA as a template forproducing dsRNA, or siRNAs alternatively. Then, nascentchromatin-bound RNAs should be synthesized and serve astemplates for siRNA-dependent recruitment of chromatinmodifying factors (Buhler and Moazed 2007). Now, it isclear that small ncRNAs are generated from heterochro-matic DNA repeats (Volpe et al. 2002; Fukagawa et al.2004; Kanellopoulou et al. 2005; Buhler and Moazed 2007)and obviously play important roles in heterochromaticassembly. They are transcribed from heterochromatinregions which are rich in transposable elements, rich inrepetitive sequences, centromeres, and telomeres that havealso distinct histone modifications.
The transgene silencing pathway (Fig. 4) is under thecontrol of DCR-1, RDE-1, RDE-4, RRF-1, ALG-1, andheterochromatin protein like-1 (HPL-1; one of the HP-1proteins in C. elegans) are also included (Grishok et al.2005). Soma transgene silencing is dependent on RDE-1 incontrast to germline RNAi-induced transgene silencing(RNAi-TGS) (Grishok et al. 2005). HPL-1, a chromatinsilencing component, inhibits the pre-edited germline trans-
gene mRNA containing intron fragments (Couteau et al.2002; Grishok et al. 2005), which then decreases theacetylation of the transgene-associated histones (Grishoket al. 2005). Transgene silencing in the germline is a stablephenomenon which is inherited epigenetically through thechromatin (Kelly et al. 1997; Strome et al. 2001; Grishoket al. 2005). Germline transgene silencing is interlacedwith H3-K9 methylation (Kelly et al. 2002). Clearly,transgene silencing functions differently in soma andgermline cells of C. elegans. Also, dsRNA productionhas been reported in soma cells, but subsequentlyadenosine deaminase acting on dsRNA edits and modifiestransgene-derived dsRNA, thus suppressing the RNAipathway (Knight and Bass 2002; Grishok et al. 2005). Insoma transgene silencing, the activity of the RdRP protein,RDE-1, seems to be essential (Grishok et al. 2005).Presumably, the SOP-2 protein (suppressor of PAL-1(posterior ALae in males-1)) which can bind to single-and double-stranded RNAs is involved in this pathway(Grishok et al. 2005). ALG-1, besides its crucial role inmiRNA generation, plays another critical role in thetransgene silencing pathway, separating it from the PTGSpathways (Grishok et al. 2001, 2005).
Fig. 4 The transgene silencing pathway
Funct Integr Genomics

In addition to transgene silencing via the TGS-RNAipathway, transposon silencing is another related path-way (Grishok et al. 2005). Accordingly, transposonsilencing (Fig. 5) could occur through transcriptionaland also post-transcriptional silencing pathways (Sijenand Plasterk 2003; Vastenhouw et al. 2003; Grishok et al.2005) because the repeatedly reported transposon silenc-ing and RNAi pathways share common factors (Kettinget al. 1999; Tabara et al. 1999; Vastenhouw and Plasterk2004; Grishok et al. 2005). Twelve percent of the C.elegans genome is composed of transposable sequences(Consortium CeS 1998; Vastenhouw and Plasterk 2004).Some transposons, such as the Tc1/mariner, are tissuespecific and are active in soma but silenced in thegermline (Emmons et al. 1984; Vastenhouw and Plasterk2004). The potential presence of DCR-1 in siRNAbiogenesis is confirmed by the transposable dsRNA(Ambros et al. 2003a, b; Sijen and Plasterk 2003;Vastenhouw and Plasterk 2004), whereby cells can trapand silence the transposons including their dsRNAs(Vastenhouw and Plasterk 2004).
Mutator-7 (MUT-7; a potent 3′ to 5′ endo-ribonuclease)is the first protein to be identified in silencing of
transposable elements and is reported in both thenucleus and cytosol, with the cytosolic complex beinglarger in size. This increase in complex size is probablydue to the involvement of RDE-2/MUT-8 proteins indownstream production of siRNA but not in theupstream process, indicating their probable functionalityin siRNA amplification (Tops et al. 2005). Therefore,the RDE-2/MUT-8 product is expected to interact withMUT-7 in the cytosolic environment. Additionally, theprobable involvement of the PPW-2 protein is recom-mended in binding to the siRNAs and guiding themtowards silencing (Vastenhouw et al. 2003; Vastenhouwand Plasterk 2004). Finally, transposing through thegenome would be specifically effective for cell’s fateand generally for the organism’s fitness deleterious duringthe evolution.
Integrated pathways
In this section, we unify the individual RNA pathwaysso as to show an integrated scheme (Fig. 6). Thesuggested scheme presents the interplay of the transcrip-tional and post-transcriptional gene silencing levels viavarious small ncRNA silencers. The reconstruction of theintegrated silencing pathways (super-pathways) of thesesmall RNAs provides a general perspective of cellularsignal transduction and a first step toward the integrationof metabolic pathways. We now know that there is a clearconnection between these small RNAs and the individu-al’s growth and development. But the intermediatecomponents among these pathways and the environmentremain unidentified and are yet to be elucidated. Thefunctional importance of these pathways in growth anddevelopment, cancer, differentiation, and apoptosis hasmotivated scientists to develop new methods in sequenc-ing and analysis, in order to understand these smallmolecules in greater detail. As recently reported, DCR-1can be proteolysed by the C. elegans cell death caspase-3protein to destroy its dsRNase activity and activating itsintrinsic DNase activity which can cleave DNA fragmentduring apoptosis (Okamura and Lai 2010). This cleavagemay also attenuate small RNA production pathways(Fig. 6).
The study of C. elegans has provided the basis foridentifying 40 components of small RNA pathways(Table 1), but many black boxes remain. The transcriptionaland post-transcriptional silencing pathways in epigeneticand systemic signaling are triggered by small RNApathway components and uncovering the upstream andtrans-acting elements behind the different pathways willcontinue to provide a basis for interesting and usefulresearch.Fig. 5 The transposon silencing pathway via the RNAi mechanism
Funct Integr Genomics
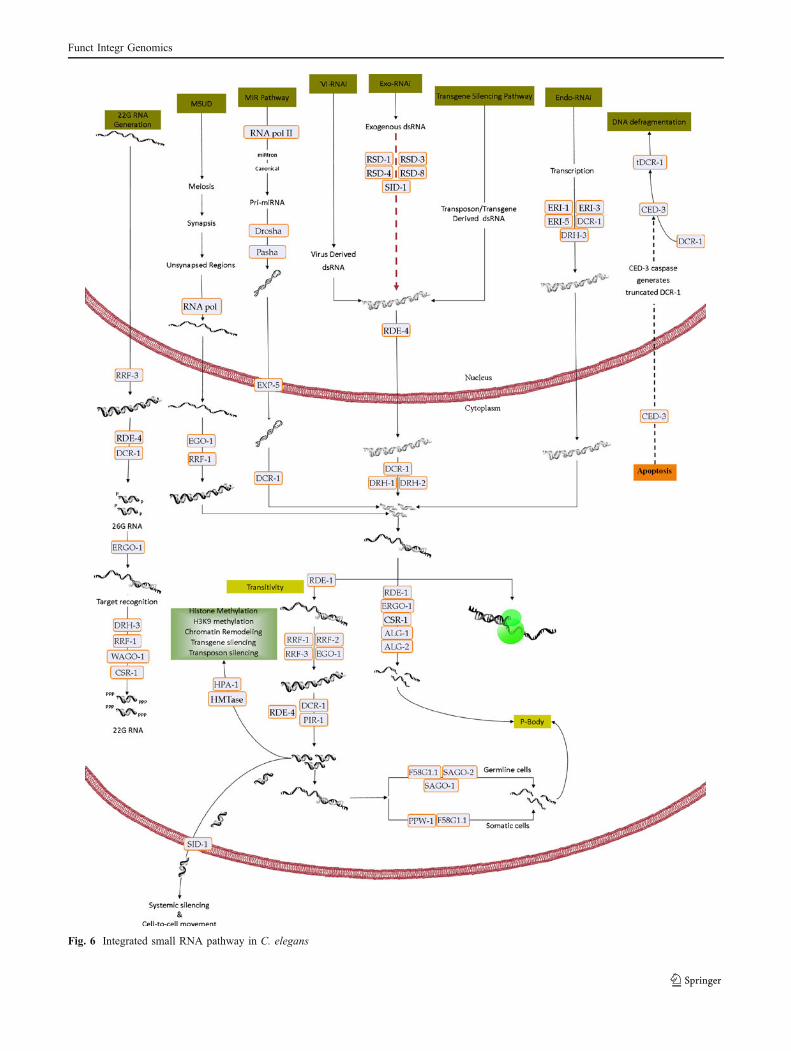
Fig. 6 Integrated small RNA pathway in C. elegans
Funct Integr Genomics

Acknowledgments The authors would like to thank RogayehShakoory for her assistance and guidance throughout the course ofthis work. We also express our appreciation to Dr. Parvin Shariati fromIran National Institute for Genetic Engineering and Biotechnology forcritical reading of the manuscript. Part of this work has been supportedby Iran National Science Foundation (www.insf.org).
References
Allo M, Buggiano V, Fededa JP, Petrillo E, Schor I, Mdl M, AgirreE, Plass M, Eyras E, Elela SA, Klinck R, Chabot B, KornblihttAR (2009) Control of alternative splicing through siRNA-mediated transcriptional gene silencing. Nat Structural Mol Biol1620:1–9
Alvarez-Saavedra E, Horvitz HR (2010) Many families of C. elegansmicroRNAs are not essential for development or viability. CurrBiol 20:367–373
Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D (2003a)MicroRNAs and other tiny endogenous RNAs in C. elegans.Curr Biol 13:807–818
Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X,Dreyfuss G, Eddy SR, Griffiths-Jones S, Marshall M, Matzke M,Ruvkun G, Tuschl T (2003b) A uniform system for microRNAannotation. RNA 9:277–279
Aoki K, Moriguchi H, Yoshioka T, Okawa K, Tabara H (2007) In vitroanalyses of the production and activity of secondary smallinterfering RNAs in C. elegans. EMBO J 26:5007–5019
Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M (2006) A novelclass of small RNAs bind to MILI protein in mouse testes. Nature442:203–207
Azimzadeh J, Masoudi-Nejad A (2009) Reconstruction of Arabidopsisthaliana fully integrated small RNA pathway. Functional IntegrGenomics 9(4):419–432
Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, andfunction. Cell 116(2):281–297
Batista PJ, Ruby G, Claycomb JM, Chiang R, Fahlgren N, KasschauKD, Chaves DA, Gu W, Vasale JJ, Duan S, Conte D Jr, Luo S,Scroth GP, Carrington JC, Bartel DP, Mello CC (2008) PRG-1and 21U-RNAs interact to form the piRNA complex required forfertility in C. elegans. Mol Cell 31(1):67–78
Bean CJ, Schaner CE, Kelly WG (2004) Meiotic pairing andimprinted X chromatin assembly in Caenorhabditis elegans.Nat Genet 36:100–105
Bhattacharyya SN, Habermacher R, Martine U, Closs EI, FilipowiczW (2006) Relief of microRNA-mediated translational repressionin human cells subjected to stress. Cell 125:1111–1124
Bleys A, Van Houdt H, Depicker A (2006) Transitive and systemicRNA silencing: both involving an RNA amplification mecha-nism? In: Nellen W, Hammann C (eds) Small RNAs: analysis andregulatory functions. Nucleic Acids and Molecular Biology. Vol17. Springer, Berlin, pp 119–139
Boag PR, Atalay A, Robida S, Reinke V, Blackwell TK (2008)Protection of specific maternal messenger RNAs by the P bodyprotein CGH-1 (Dhh1/RCK) during Caenorhabditis elegansoogenesis. J Cell Biol 182(3):543–557
Bohnsack MT, Czaplinski K, Gorlich D (2004) Exportin 5 is aRanGTP-dependent dsRNA-binding protein that mediates nuclearexport of pre-miRNAs. RNA 10(2):185–191
Boisvert M-EL, Simard MJ (2009) RNAi pathway in C. elegans: theArgonautes and collaborators. Curr Top Microbiol Immunol320:21–36
Buhler M, Moazed D (2007) Transcription and RNAi in heterochro-matic gene silencing. Nat Structural Mol Biol 14:1041–1048
Cai X, Hagedorn C, Cullen B (2004) Human microRNAs areprocessed from capped, polyadenylated transcripts that can alsofunction as mRNAs. RNA 10(12):1957–1966
CeS C (1998) Genome sequence of the nematode C. elegans: aplatform for investigating biology. Science 282:2012–2018
Chatterjee S, Fasler M, Büssing I, Großhans H (2011) Target-mediatedprotection of endogenous microRNAs in C. elegans. Dev Cell20:388–396
Chen C-CG, Simard MJ, Tabara H, Brownell DR, McCollough JA,Mello CC (2005) A member of the polymerase nucleotidyltrans-ferase superfamily is required for RNA interference in C.elegans. Curr Biol 15:378–383
Chu C-Y, Rana TM (2007) Small RNAs: regulators and guardians ofthe genome. Cell Physiol 213:412–419
Chung W-J, Okamura K, Martin R, Lai EC (2008) Endogenous RNAinterference provides a somatic defense against Drosophilatransposons. Curr Biol 18:795–802
Conine CC, Batista PJ, Gu W, Claycomb JM, Chaves DA,Shirayama M, Mello CC (2010) Argonautes ALG-3 andALG-4 are required for spermatogenesis-specific 26G-RNAsand thermotolerant sperm in Caenorhabditis elegans. PNAS 107(8):3588–3593
Costa Y, Speed RM, Gautier P, Semple CA, Maratou K, Turner JM,Cooke HJ (2006) Mouse MAELSTROM: the link betweenmeiotic silencing of unsynapsed chromatin and microRNApathway? Hum Mol Genet 15:2324–2334
Couteau F, Guerry F, Muller F, Palladino F (2002) A heterochromatinprotein 1 homologue in Caenorhabditis elegans acts in germlineand vulval development. EMBO Rep 3(31):235–241
Czech B, Malone CD, Zhou R, Stark A (2008) An endogenous smallinterfering RNA pathway in Drosophila. Nature 453:729–731
Dalmay T (2000) An RNA-dependent RNA polymerase gene inArabidopsis is required for posttranscriptional gene silencingmediated by a transgene but not by a virus. Cell 101:543–553
Das PP, Bagijn MP, Goldstein LD, Woolford JR (2008) Piwi andpiRNAs act upstream of an endogenous siRNA pathway tosuppress Tc3 transposon mobility in the Caenorhabditis elegansgermline. Mol Cell 31:79–90
Ding SW, Voinnet O (2007) Antiviral immunity directed by smallRNAs. Cell 7:39
Ding L, Spencer A, Morita K, Han M (2005) The developmentaltiming regulator AIN-1 interacts with miRISCs and may targetthe Argonaute protein ALG-1 to cytoplasmic P bodies in C.elegans. Mol Cell 19(4):437–447
Doench JG, Sharp PA (2004) Specificity of microRNA target selectionin translational repression. Genes Dev 18:504–511
Dorer DR, Henikoff S (1994) Expansions of transgene repeats causeheterochromatin formation and gene silencing in Drosophila. Cell77:993–1002
Duchaine TF, Wohlschlegel JA, Kennedy S, Bei Y, Conte D, Pang K,Brownell DR, Harding S, Mitani S, Ruvkun G, Yates JR, MelloCC (2006) Functional proteomics reveals the biochemical nicheof C. elegans DCR-1 in multiple small-RNA-mediated pathways.Cell 124(2):343–354
Emmons S, Emmons W, Yesner L (1984) High-frequency excision oftransposable element Tc1 in the nematode Caenorhabditiselegans is limited to somatic cells. Cell 36:599–605
Faehnle CR, Joshua-Tor L (2007) Argonautes confront new smallRNAs. Curr Opin Chem Biol 11:569–577
Farazi TA, Juranek SA, Tuschl T (2008) The growing catalog of smallRNAs and their association with distinct Argonaute/Piwi familymembers. Development 135:1201–1214
Feinberg EH, Hunter CP (2003) Transport of dsRNA into cells by thetransmembrane protein SID-1. Science 301:1545–1547
Fillman C, Lykke-Andersen J (2005) RNA decapping inside andoutside of processing bodies. Curr Opin Cell Biol 17:326–331
Funct Integr Genomics

Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC(1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391(6669):806–811
Fischer SE, Butler MD, Pan Q, Ruvkun G (2008) Trans-splicing in C.elegans generates the negative RNAi regulator ERI-6/7. Nature455(7212):491–496
Fukagawa T, Nogami M, Yoshikawa M, Ikeno M (2004) Dicer isessential for formation of the heterochromatin structure invertebrate cells. Nat Cell Biol 6:784–791
Gaidatzis D, Ev N, Hausser J, Zavolan M (2007) Inference of miRNAtargets using evolutionary conservation and pathway analysis.BMC Bioinforma 8:69
Galiana-Arnoux D, Dostert C, Schneemann A, Hoffmann JA, Imler JL(2006) Essential function in vivo for Dicer-2 in host defenseagainst RNA viruses in Drosophila. Nat Immunol 7:590–597
Girard A, Sachidanandam R, Hannon GJ, Carmell MA (2006) Agermline-specific class of small RNAs binds mammalian Piwiproteins. Nature 442:199–202
Grad Y, Aach J, Hayes GD, Reinhart BJ (2003) Computational andexperimental identification of C. elegans microRNAs. Mol Cell11(5):1253–1263
Grewal SI, Moazed D (2003) Heterochromatin and epigenetic controlof gene expression. Science 301:798–802
Grishok A (2005) RNAi mechanisms in Caenorhabditis elegans.FEBS Lett 579(26):5932–5939
Grishok A, Mello CC (2002) RNAi (nematodes: Caenorhabditiselegans). Adv Genet 46:339–360
Grishok A, Tabara H, Mello CC (2000) Genetic requirements forinheritance of RNAi in C. elegans. Science 287(5462):2494–2497
Grishok A, Pasquinelli AE, Conte D, Li N, Parrish S, Ha I, Baillie DL, FireA, Ruvkun G, Mello CC (2001) Genes and mechanisms related toRNA interference regulate expression of the small temporal RNAsthat control C. elegans developmental timing. Cell 106(1):23–34
Grishok A, Sinskey JL, Sharp PA (2005) Transcriptional silencing of atransgene by RNAi in the soma of C. elegans. Genes Dev 19(6):683–696
Gu W, Shirayama M, Darryl Conte J, Vasale J, Batista PJ, ClaycombJM, Moresco JJ, Youngman EM, Keys J, Stoltz MJ, Chen C-CG,Chaves DA, Duan S, Kasschau KD, Fahlgren N, Yates JR III,Mitani S, Carrington JC, Mello CC (2009) Distinct Argonaute-mediated 22G-RNA pathways direct genome surveillance in theC. elegans germline. Mol Cell 36:231–244
Hammond TM, Andrewski MD, Roossinck MJ, Keller NP (2008)Aspergillus mycoviruses are targets and suppressors of RNAsilencing. Eukaryot Cell 7:350–357
Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, Sohn SY, Cho Y,Zhang BT, Kim VN (2006) Molecular basis for the recognition ofprimary microRNAs by the Drosha–DGCR8 complex. Cell 125(5):887–901
Han J, Pedersen JS, Kwon SC, Belair CD, Kim YK, Yeom KH, YangWY, Haussler D, Blelloch R, Kim VN (2009) Posttranscriptionalcrossregulation between Drosha and DGCR8. Cell 136(1):75–84
Hannon GJ, Rossi JJ (2004) Unlocking the potential of the humangenome with RNA interference. Nature 431:371–378
Holmes RJ, Cohen PE (2007) Small RNAs and RNAi pathways inmeiotic prophase I. Chromosome Res 15:653–665
Horwich MD, Li C, Matranga C, Vagin V, Farley G, Wang P, ZamorePD (2007) The Drosophila RNA methyltransferase, DmHen1,modifies germline piRNAs and single-stranded siRNAs in RISC.Curr Biol 17:1265–1272
Houwing S, Kamminga LM, Berezikov E, Cronembold D, Girard A(2007) A role for Piwi and piRNAs in germ cell maintenance andtransposon silencing in zebrafish. Cell 129:69–82
Hutvagner G, Simard MJ (2008) Argonaute proteins: key players inRNA silencing. Nat Reviews Mol Cell Biol 9:22–32
Hutvágner G, Simard M, Mello C, Zamore P (2004) Sequence-specific inhibition of small RNA function. PLoS Biol 2:E98
Hwang HW, Wentzel EA, Mendell JT (2007) A hexanucleotide elementdirects microRNA nuclear import. Science 315(5808):97–100
Jannot G, Boisvert M-EL, Banville IH, Simard MJ (2008) Twomolecular features contribute to the Argonaute specificity for themicroRNA and RNAi pathways in C. elegans. RNA 14:829–835
Jenuwein T (2001) Re-SET-ting heterochromatin by histone methyl-transferases. Trends Cell Biol Chem 11:266–273
Jenuwein T, Allis CD (2001) Translating the histone code. Science293:1074–1080
Jose AM, Hunter CP (2007) Transport of sequence-specific RNAinterference information between cells. Annu Rev Genet 41:305–330
Kadener S, Rodriguez J, Abruzzi KC, Khodor YL, Sugino K, MarrMT 2nd, Nelson S, Rosbash M (2009) Genome-wide identifica-tion of targets of the Drosha-pasha/DGCR8 complex. RNA 15(4):537–545
Kanellopoulou C, Muljo SA, Kung AL, Ganesan S (2005) Dicer-deficient mouse embryonic stem cells are defective in differen-tiation and centromeric silencing. Genes Dev 19:489–501
Kelly WG, Xu S, Montgomery MK, Fire A (1997) Distinct require-ments for somatic and germline expression of a generallyexpressed Caenorhabditis elegans gene. Genetics 146:227–238
Kelly WG, Schaner CE, Dernburg AF, Lee MH, Kim SK, VilleneuveAM, Reinke V (2002) X-chromosome silencing in the germlineof C. elegans. Development 129(2):479–492
Kennedy S, Wang D, Ruvkun G (2004) A conserved siRNA-degrading RNase negatively regulates RNA interference in C.elegans. Nature 427(6975):645–649
Ketting RF, Haverkamp TH, van Luenen HG, Plasterk RH (1999)Mut-7 of C. elegans, required for transposon silencing and RNAinterference, is a homolog of Werner syndrome helicase andRNaseD. Cell 99(2):133–141
Ketting RF, Fischer SE, Bernstein E, Sijen T, Hannon GJ, Plasterk RH(2001) Dicer functions in RNA interference and in synthesis ofsmall RNA involved in developmental timing in C. elegans.Genes Dev 15(20):2654–2659
Khvorova A, Reynolds A, Jayasena SD (2003) Functional siRNAsand miRNAs exhibit strand bias. Cell 115:209–216
Kim VN (2005) MicroRNA biogenesis: coordinated cropping anddicing. Nat Mol Cell Biol 6:376–385
Kim Y-K, Kim VN (2007) Processing of intronic microRNAs. EMBOJ 26:775–783
Kim DH, Behlke MA, Rose SD, Chang MS, Choi S, Rossi JJ (2005)Synthetic dsRNA Dicer substrates enhance RNAi potency andefficacy. Nat Biotechnol 23:222–226
Klattenhoff C, Theurkauf W (2008) Biogenesis and germlinefunctions of piRNAs. Development 135:3–9
Klenov MS, Lavrov SA, Stolyarenko AD, Ryazansky SS, Aravin AA,Tuschl T, Gvozdev VA (2007) Repeat-associated siRNAs causechromatin silencing of retrotransposons in the Drosophilamelanogaster germline. Nucleic Acids Res 35(16):5430–5438
Knight SW, Bass BL (2001) A role for the RNase III enzyme DCR-1 inRNA interference and germ line development in Caenorhabditiselegans. Science 293(5538):2269–2271
Knight SW, Bass BL (2002) The role of RNA editing by ADARs inRNAi. Mol Cell 10:809–817
Lai EC (2002) Micro RNAs are complementary to 3′ UTR sequencemotifs that mediate negative post-transcriptional regulation. NatGenetics 30:363–364
Lee RC, Feinbaum RL, Ambros V (1993) The C. elegans hetero-chronic gene lin-4 encodes small RNAs with antisense comple-mentarity to lin-14. Cell 75(5):843–854
Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J, Lee J, Provost P, RadmarkO, Kim S, Kim VN (2003) The nuclear RNase III Droshainitiates microRNA processing. Nature 425(6956):415–419
Funct Integr Genomics

Lee RC, Hammell CM, Ambros V (2006) Interacting endogenous andexogenous RNAi pathways in Caenorhabditis elegans. RNA 12(4):589–597
Lewis BP, Burge CB, Bartel DP (2005) Conserved seed pairing, oftenflanked by adenosines, indicates that thousands of human genesare microRNA targets. Cell 120:15–20
Li WX, Ding SW (2001) Viral suppressors of RNA silencing. CurrOpin Biotechnol 12:150–154
Li F, Ding SW (2006) Virus counterdefense: diverse strategies forevading the RNA-silencing immunity. Annu Rev Microbiol60:503–531
Li HW, Lucy AP, Guo HS, Li WX, Ji LH, Wong SM (1999) Stronghost resistance targeted against a viral suppressor of the plantgene silencing defense mechanism. EMBO J 18:2683–2691
Li HW, Li WX, Ding SW (2002) Induction and suppression of RNAsilencing by an animal virus. Science 296:1319–1321
Lim LP, Glasner ME, Yekta S, Burge CB, Bartel DP (2003) VertebratemicroRNA genes. Science 299(5612):1540
Lin SL, Miller JD, Ying SY (2006) Intronic microRNA (miRNA). JBiomed Biotechnol 2006(4):26818
Lipardi C, Wei Q, Paterson BM (2001) RNAi as random degradativePCR: siRNA primers convert mRNA into dsRNAs that aredegraded to generate new siRNAs. Cell 107:297–307
Lippman Z, Martienssen R (2004) The role of RNA interference inheterochromatic silencing. Nature 431:364–370
Lu R, Maduro M, Li F, Li HW, Broitman-Maduro G, Li WX, Ding SW(2005) Animal virus replication and RNAi-mediated antiviralsilencing in Caenorhabditis elegans. Nature 436(7053):1040–1043
Lu R, Yigit E, Li W-X, Ding S-W (2009) An RIG-I-like RNA helicasemediates antiviral RNAi downstream of viral siRNA biogenesisin Caenorhabditis elegans. PLoS Pathog 5(2):e1000286
Luco RF, Pan Q, Tominaga K, Blencowe BJ (2010) Regulation ofalternative splicing by histone modifications. Science 327:996–1000
Ma JB, Yuan YR, Meister G, Pei Y, Tuschl T, Patel DJ (2005)Structural basis for 5′-end-specific recognition of guide RNA bythe A. fulgidus Piwi protein. Nature 434:666–670
Maine EM, Hauth J, Ratliff T, Vought VE, She X, Kelly WG (2005)EGO-1, a putative RNA-dependent RNA polymerase, is requiredfor heterochromatin assembly on unpaired DNA during C.elegans meiosis. Curr Biol 15:1972–1978
Maniar JM, Fire AZ (2011) EGO-1, a C. elegans RdRP, modulatesgene expression via production of mRNA-templated shortantisense RNAs. Curr Biol 21(6):450–459
Martens H, Novotny J, Oberstrass J, Steck TL, Postlethwait P, NellenW (2002) RNAi in Dictyostelium: the role of RNA-directedRNA polymerases and double-stranded RNase. Mol Biol Cell13:445–453
Meister G, Tuschl T (2004) Mechanisms of gene silencing by double-stranded RNA. Nature 431:343–349
Nakamura M, Ando R, Nakazawa T, Yudazono T, Tsutsumi N,Hatanaka N, Ohgake T, Hanaoka F, Eki T (2007) Dicer-relateddrh-3 gene functions in germ-line development by maintenanceof chromosomal integrity in Caenorhabditis elegans. Genes Cells12(9):997–1010
Naqvi AR, Islam MN, Choudhury NR, Haq QMR (2009) Thefascinating world of RNA interference. Int J Biol Sci 5(2):97–117
Noble SL, Allen BL, Goh LK, Nordick K, Evans TC (2008) MaternalmRNAs are regulated by diverse P body-related mRNP granulesduring early Caenorhabditis elegans development. J Cell Biol182(3):559–572
Obbard DJ, Gordon KHJ, Buck AH, Jiggins FM (2009) The evolutionof RNAi as a defence against viruses and transposable elements.Phil Trans R Soc B 364:99–115
Ohara T, Sakaguchi Y, Suzuki T, Ueda H, Miyauchi K, Suzuki T (2007)The 3′ termini of mouse Piwi-interacting RNAs are 2′-O-methylated.Nat Struct Mol Biol 14:349–350
Okamura K, Lai EC (2010) A deathly DNase activity for dicer. DevCell 18:692–694
Okamura K, Hagen JW, Duan H, Tyler DM, Lai EC (2007) Themirtron pathway generates microRNA-class regulatory RNAs inDrosophila. Cell 130(1):1–12
Okamura K, Chung W-J, Ruby JG, Guo H, Bartel DP, Lai EC (2008)The Drosophila hairpin RNA pathway generates endogenousshort interfering RNAs. Nature 453:803–806
Ouellet J, Roy R (2007) The lin-35/Rb and RNAi pathways cooperate toregulate a key cell cycle transition inC. elegans. BMC Dev Biol 7:38
Pak J, Fire A (2007) Distinct populations of primary and secondaryeffectors during RNAi in C. elegans. Science 315(5809):241–244
Parker JS, Barford D (2006) Argonaute: a scaffold for the function ofshort regulatory RNAs. Trends Biochem Sci 31:622–630
Parker JS, Roe SM, Barford D (2005) Structural insights into mRNArecognition from a PIWI domain-siRNA guide complex. Nature434:663–666
Peters AH et al (2003) Partitioning and plasticity of repressive histonemethylation states in mammalian chromatin. Mol Cell 12:1577–1589
Rajewsky N (2006) MicroRNA target predictions in animals. NatGenet 38:S8–S13
Rana TM (2007) Illuminating the silence: understanding the structureand function of small RNAs. Nat Rev Mol Cell Biol 8:23–36
Richards EJ, Elgin SC (2002) Epigenetic codes for heterochromatinformation and silencing: rounding up the usual suspects. Cell108:489–500
Rij RPv, Saleh MC, Berry B, Foo C, Houk A, Antoniewski C, AndinoR (2006) The RNA silencing endonuclease Argonaute 2 mediatesspecific antiviral immunity in Drosophila melanogaster. GenesDev 20:2985–2995
Robert VJ, Sijen T, van Wolfswinkel J, Plasterk RH (2005) Chromatinand RNAi factors protect the C. elegans germline againstrepetitive sequences. Genes Dev 19(7):782–787
Rocheleau CE, Cullison K, Huang K, Bernstein Y, Spilker AC,Sundaram MV (2008) The Caenorhabditis elegans ekl (enhancerof ksr-1 lethality) genes include putative components of agermline small RNA pathway. Genetics 178(3):1431–1443
Rodriguez A, Griffith-Jones S, Ashurst JL, Bradley A (2004)Identification of mammalian microRNA host genes and tran-scription units. Genome Res 14(104):1902–1910
Ruby JG, Jan C, Player C, Axtell MJ, Lee Wet al (2006) Large-scalesequencing reveals 21U-RNAs and additional microRNAs andendogenous siRNAs in C. elegans. Cell 127:1193–1207
Ruby JG, Jan CH, Bartel DP (2007) Intronic microRNA precursorsthat bypass Drosha processing. Nature 448(7149):83–86
Saito K, Sakaguchi Y, Suzuki T, Suzuki T, Siomi H, Siomi MC (2007)Pimet, the Drosophila homolog of HEN1, mediates 2′-O-methylation of Piwi-interacting RNAs at their 3′ ends. GenesDev 21:1603–1608
Schot DH, Cureton DK, Whelan SP, Hunter CP (2005) An antiviralrole for the RNA interference machinery in Caenorhabditiselegans. PNAS 102(51):18420–18424
Schramke V, Allshire R (2003) Hairpin RNAs and retrotransposonLTRs effect RNAi and chromatin-based gene silencing. Science301:1069–1074
Schwarz DS, Hutvagner G, Du T, Xu Z, Aronin N, Zamore PD (2003)Asymmetry in the assembly of the RNAi enzyme complex. Cell115:199–208
Segers GC, Zhang XM, Deng FY, Sun QH, Nuss DL (2007) Evidencethat RNA silencing functions as an antiviral defense mechanismin fungi. Proc Natl Acad Sci USA 104:12,902–12,906
Shabalina SA, Koonin EV (2008) Origins and evolution of eukaryoticRNA interference. Trends Ecol Evol 23(10):578–587
Sijen T, Plasterk RH (2003) Transposon silencing in the Caenorhab-ditis elegans germ line by natural RNAi. Nature 426(6964):310–314
Funct Integr Genomics

Sijen T, Fleenor J, Simmer F, Thijssen KL, Parrish S, Timmons L,Plasterk RHA, Fire A (2001) On the role of RNA amplificationin dsRNA-triggered gene silencing. Cell 107:465–476
Sijen T, Steiner FA, Thijssen KL, Plasterk RHA (2007) SecondarysiRNAs result from unprimed RNA synthesis and form a distinctclass. Science 315:244–247
Simmer F, Tijsterman M, Parrish S, Koushika SP, Nonet ML, Fire A,Ahringer J, Plasterk RH (2002) Loss of the putative RNA-directed RNA polymerase RRF-3 makes C. elegans hypersensi-tive to RNAi. Curr Biol 12(15):1317–1319
Siolas D, Lerner C, Burchard J, Ge W, Linsley PS, Paddison PJ,Hannon GJ, Cleary MA (2005) Synthetic shRNAs as potentRNAi triggers. Nat Biotechnol 23:227–231
Siomi H, Siomi MC (2009) On the road to reading the RNA-interference code. Nature 457:396–404
Smardon A, Spoerke JM, Stacey SC, Klein ME, Mackin N, MaineEM (2000) EGO-1 is related to RNA-directed RNA polymeraseand functions in germ-line development and RNA interference inC. elegans. Curr Biol 10(4):169–178
Song JJ, Joshua-Tor L (2006) Argonaute and RNA-getting into thegroove. Curr Opin Struct Biol 16:5–11
Steiner FA, Okihara KL, Hoogstrate SW, Sijen T, Ketting RF (2009)RDE-1 slicer activity is required only for passenger-strandcleavage during RNAi in Caenorhabditis elegans. Nat StructuralMol Biol 16(2):207–211
Strome S, Powers J, Dunn M, Reese K, Malone CJ, White J, SeydouxG, Saxton W (2001) Spindle dynamics and the role of gamma-tubulin in early Caenorhabditis elegans embryos. Mol Biol Cell12:1751–1764
Tabara H, Sarkissian M, Kelly WG, Fleenor J, Grishok A, Timmons L,Fire A, Mello CC (1999) The rde-1 gene, RNA interference, andtransposon silencing in C. elegans. Cell 99(2):123–132
Tabara H, Yigit E, Siomi H, Mello CC (2002) The dsRNA bindingprotein RDE-4 interacts with RDE-1, DCR-1, and a DExH-boxhelicase to direct RNAi in C. elegans. Cell 109(7):861–871
Tijsterman M, Okihara KL, Thijssen K, Plasterk RH (2002a) PPW-1, aPAZ/PIWI protein required for efficient germline RNAi, isdefective in a natural isolate of C. elegans. Curr Biol 12(17):1535–1540
Tijsterman M, Ketting RF, Okihara KL, Sijen T, Plasterk RH(2002b) RNA helicase MUT-14-dependent gene silencingtriggered in C. elegans by short antisense RNAs. Science 295(5555):694–697
Tijsterman M, May RC, Simmer F, Okihara KL, Plasterk RH (2004)Genes required for systemic RNA interference in Caenorhabditiselegans. Curr Biol 14(2):111–116
Tilgner H, Guigó R (2010) From chromatin to splicing RNA-processing as a total artwork. Epigenetics 5(3):1–5
Tolia NH, Joshua-Tor L (2007) Slicer and the Argonautes. Nat ChemBiol 3:36–43
Tops BB, Tabara H, Sijen T, Simmer F, Mello CC, Plasterk RH, Ketting RF(2005) RDE-2 interacts with MUT-7 to mediate RNA interference inCaenorhabditis elegans. Nucleic Acids Res 33(1):347–355
Turner JM, Mahadevaiah SK, Fernandez-Capetillo O, Nussenzweig A,Xu X, Deng CX, Burgoyne PS (2005) Silencing of unsynapsedmeiotic chromosomes in the mouse. Nat Genet 37:41–47
Vaistij FE (2002) Spreading of RNA targeting and DNA methylationin RNA silencing requires transcription of the target gene and aputative RNA-dependent RNA polymerase. Plant Cell 14:857–867
Vasale JJ, Gu W, Thivierge C, Batista PJ, Claycomb JM, YoungmanEM, Duchaine TF, Melloa CC, Darryl Conte J (2010) Sequentialrounds of RNA-dependent RNA transcription drive endogenoussmall-RNA biogenesis in the ERGO-1/ Argonaute pathway.PNAS 107(8):3582–3587
Vastenhouw NL, Plasterk RH (2004) RNAi protects the Caenorhabditiselegans germline against transposition. Trends Genet 20(7):314–319
Vastenhouw NL, Fischer SEJ, VrJP R, Thijssen KL, Fraser AG,Kamath RS, Ahringer J, Plasterk RHA (2003) A genome-widescreen identifies 27 genes involved in transposon silencing in C.elegans. Curr Biol 13:1311–1316
Vastenhouw NL, Brunschwig K, Okihara KL, Muller F, Tijsterman M,Plasterk RH (2006) Gene expression: long-term gene silencingby RNAi. Nature 442:882
Vasudevan S, Steitz J (2007) AU-rich-element-mediated upregulationof translation by FXR1 and Argonaute 2. Cell 128(6):1105–1118
Vasudevan S, Tong Y, Steitz JA (2007) Switching from repression toactivation: microRNAs can up-regulate translation. Science 318(5858):1931–1934
Volpe TA, Kidner C, Hall IM, Teng G (2002) Regulation ofheterochromatic silencing and histone H3 lysine-9 methylationby RNAi. Science 297:1833–1837
Vought VE, Ohmachi M, Lee M-H, Maine EM (2005) EGO-1, aputative RNA-directed RNA polymerase, promotes germlineproliferation in parallel with GLP-1/notch signaling andregulates the spatial organization of nuclear pore complexesand germline P granules in Caenorhabditis elegans. Genetics170:1121–1132
Wakiyama M, Takimoto K, Ohara O, Yokoyama S (2007) Let-7microRNA mediated mRNA deadenylation and translationalrepression in a mammalian cell-free system. Genes Dev 21(15):1857–1862
Wang M-B, Metzlaff M (2005) RNA silencing and antiviral defense inplants. Curr Opin Plant Biol 8:216–222
Wang XH, Aliyari R, Li WX, Li HW, Kim K, Carthew R, Atkinson P,Ding SW (2006) RNA interference directs innate immunityagainst viruses in adult Drosophila. Science 312:452–454
Wang Y, Medvid R, Melton C, Jaenisch R, Blelloch R (2007) DGCR8is essential for microRNA biogenesis and silencing of embryonicstem cell self-renewal. Nat Genet 39(3):380–385
Wassenegger M (2005) The role of the RNAi machinery inheterochromatin formation. Cell 122:13–16
Wilkins C, Dishongh R, Moore SC, Whitt MA, Chow M, Machaca K(2005) RNA interference is an antiviral defence mechanism inCaenorhabditis elegans. Nature 436(7053):1044–1047
Winston WM, Molodowitch C, Hunter CP (2002) Systemic RNAi inC. elegans requires the putative transmembrane protein SID-1.Science 295(5564):2456–2459
Xie Z, Kasschau KD, Carrington JC (2003) Negative feedbackregulation of Dicer-like1 (DCL1) in Arabidopsis by microRNA-guided mRNA degradation. Curr Biol 13(9):784–789
Yigit E, Batista PJ, Bei Y, Pang KM, Chen CC, Tolia NH, Joshua-TorL, Mitani S, Simard MJ, Mello CC (2006) Analysis of the C.elegans Argonaute family reveals that distinct Argonautes actsequentially during RNAi. Cell 127(4):747–757
Zambon RA, Vakharian VN, Wu LP (2006) RNAi is an antiviral immuneresponse against a dsRNA virus in Drosophila melanogaster. CellMicrobiol 8:880–889
Zaratiegui M, Irvine DV, Martienssen RA (2007) Noncoding RNAsand gene silencing. Cell 128:763–776
Funct Integr Genomics
Related Documents











