RNA therapeutics targeting osteoclast-mediated excessive bone resorption ☆ Yuwei Wang a , David W. Grainger a, b, ⁎ a Department of Pharmaceutics and Pharmaceutical Chemistry, University of Utah, Salt Lake City, UT 84112-5820, USA b Department of Bioengineering, University of Utah, Salt Lake City, UT 84112-5820, USA abstract article info Article history: Received 4 August 2011 Accepted 5 September 2011 Available online xxxx Keywords: siRNA Osteoclast Molecular target Bone resorption Osteoporosis Gene therapy RNAi RNA interference (RNAi) is a sequence-specific post-transcriptional gene silencing technique developed with dramatically increasing utility for both scientific and therapeutic purposes. Short interfering RNA (siRNA) is currently exploited to regulate protein expression relevant to many therapeutic applications, and commonly used as a tool for elucidating disease-associated genes. Osteoporosis and their associated osteoporotic fragil- ity fractures in both men and women are rapidly becoming a global healthcare crisis as average life expectan- cy increases worldwide. New therapeutics are needed for this increasing patient population. This review describes the diversity of molecular targets suitable for RNAi-based gene knock down in osteoclasts to control osteoclast-mediated excessive bone resorption. We identify strategies for developing targeted siRNA delivery and efficient gene silencing, and describe opportunities and challenges of introducing siRNA as a therapeutic approach to hard and connective tissue disorders. © 2011 Elsevier B.V. All rights reserved. Contents 1. Introduction: the need for improved therapeutics for osteoporosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 2. RNA interference and siRNA delivery as a new therapeutic approach. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 3. RNAi applications in new osteoporosis therapies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 4. Osteoclastogenesis and osteoclastic bone resorption . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 5. Targets for siRNA in osteoporosis and bone metabolism. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 5.1. Osteoclastogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 5.2. Bone resorption targets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 5.3. Osteoclast viability targets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 5.4. Other potential osteoporosis targets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 6. siRNA delivery to bone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 6.1. Challenges and strategies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 6.2. Non-specific siRNA-cell bioactivities and off-target side effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 7. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 Acknowledgments. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0 Advanced Drug Delivery Reviews xxx (2011) xxx–xxx Abbreviations: 3BP2, c-Abl SH3 domain-binding protein-2; ADAM, a disintegrin and metalloproteinases; Ago, Argonaute; AP-1, activator protein-1; Ckb, brain-type creatine ki- nase; CNV, choroidal neovascularization; CPA, cation-proton antiporter; CPC, calcium phosphate bone cements; DC-STAMP, dendritic cell-specific transmembrane protein; EGFR, epidermal growth factor receptor; FPPS, farnesyl pyrophosphate synthase; GGPP, geranylgeranyl pyrophosphate; H + -ATPase, H + -adenosine triphosphatase; ICAM-1, intercellular adhesion molecule-1; IKK, IκB kinase; IP3R1, inositol-1,4,5-triphosphate receptor-1; IP3R1, inositol-1,4,5-triphosphate receptor-1; KCCs, K + /Cl − co-transporters; M-CSF, macro- phage colony-stimulating factor; MDM2, murine double minute 2; N-BP, nitrogen-containing bisphosphonate; NCX, Na + /Ca 2+ ion exchanger; NFATc1, nuclear factor of activated T cells; NF-κB, nuclear factor kappa B; OCIF, osteoclastogenesis inhibitory factor; OC-STAMP, osteoclast stimulatory transmembrane protein; OGR1, ovarian cancer G-protein- coupled receptor 1; OPG, osteoprotegerin; PBMC, blood mononuclear cells; PIP 2 , phosphatidylinositol 4,5-bisphosphate; PTK, protein tyrosine kinase; PTP-oc, osteoclast-specific transmembrane protein-tyrosine phosphatase; PTP-PEST, protein-tyrosine phosphatase-proline, glutamic acid, serine, threonine amino acid sequence; RA, rheumatoid arthritis; RANK, receptor activator of NF-κB; RANKL, RANK ligand; RGS, regulators of G-protein signaling; RISC, RNA-induced silencing complexes; RIZ1, retinoblastoma protein-interacting zinc finger 1; RNAi, RNA interference; shRNA, short hairpin RNA; siRNA, small interfering RNA; TLR3, toll-like receptor 3; TRAF6, TNF receptor-associated factor 6; TRAIL, TNF-re- lated apoptosis-inducing ligand; TRAP, tartrate resistant acid phosphatase; TREM2, triggering receptor expressed in myeloid cells-1; WASP, Wiscott-Aldrich syndrome protein. ☆ This review is part of the Advanced Drug Delivery Reviews theme issue on “Targeted delivery of therapeutics to bone and connective tissues”. ⁎ Corresponding author at: Department of Pharmaceutics and Pharmaceutical Chemistry, University of Utah, Salt Lake City, UT 84112-5820, USA. Tel.: + 1 801 581 3715; fax: + 1 801 581 3674. E-mail addresses: [email protected] (Y. Wang), [email protected] (D.W. Grainger). ADR-12185; No of Pages 17 0169-409X/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.addr.2011.09.002 Contents lists available at SciVerse ScienceDirect Advanced Drug Delivery Reviews journal homepage: www.elsevier.com/locate/addr Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics targeting osteoclast-mediated excessive bone resorption , Adv. Drug Deliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
ADR-12185; No of Pages 17
Contents lists available at SciVerse ScienceDirect
Advanced Drug Delivery Reviews
j ourna l homepage: www.e lsev ie r .com/ locate /addr
RNA therapeutics targeting osteoclast-mediated excessive bone resorption☆
Yuwei Wang a, David W. Grainger a,b,⁎a Department of Pharmaceutics and Pharmaceutical Chemistry, University of Utah, Salt Lake City, UT 84112-5820, USAb Department of Bioengineering, University of Utah, Salt Lake City, UT 84112-5820, USA
Abbreviations: 3BP2, c-Abl SH3 domain-binding protnase; CNV, choroidal neovascularization; CPA, cation-prepidermal growth factor receptor; FPPS, farnesyl pyrophadhesion molecule-1; IKK, IκB kinase; IP3R1, inositol-1phage colony-stimulating factor; MDM2, murine doubleT cells; NF-κB, nuclear factor kappa B; OCIF, osteoclastcoupled receptor 1; OPG, osteoprotegerin; PBMC, bloodtransmembrane protein-tyrosine phosphatase; PTP-PESRANK, receptor activator of NF-κB; RANKL, RANK ligandzinc finger 1; RNAi, RNA interference; shRNA, short hairlated apoptosis-inducing ligand; TRAP, tartrate resistan☆ This review is part of the Advanced Drug Delivery Re⁎ Corresponding author at: Department of Pharmaceu
801 581 3674.E-mail addresses: [email protected] (Y. W
0169-409X/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.addr.2011.09.002
Please cite this article as: Y. Wang, D.W. GDeliv. Rev. (2011), doi:10.1016/j.addr.2011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 August 2011Accepted 5 September 2011Available online xxxx
Keywords:siRNAOsteoclastMolecular targetBone resorptionOsteoporosisGene therapyRNAi
RNA interference (RNAi) is a sequence-specific post-transcriptional gene silencing technique developed withdramatically increasing utility for both scientific and therapeutic purposes. Short interfering RNA (siRNA) iscurrently exploited to regulate protein expression relevant to many therapeutic applications, and commonlyused as a tool for elucidating disease-associated genes. Osteoporosis and their associated osteoporotic fragil-ity fractures in both men and women are rapidly becoming a global healthcare crisis as average life expectan-cy increases worldwide. New therapeutics are needed for this increasing patient population. This reviewdescribes the diversity of molecular targets suitable for RNAi-based gene knock down in osteoclasts to controlosteoclast-mediated excessive bone resorption. We identify strategies for developing targeted siRNA deliveryand efficient gene silencing, and describe opportunities and challenges of introducing siRNA as a therapeuticapproach to hard and connective tissue disorders.
ein-2; ADAM, a disintegrin and metalloproteinases; Ago,oton antiporter; CPC, calcium phosphate bone cements;osphate synthase; GGPP, geranylgeranyl pyrophosphate,4,5-triphosphate receptor-1; IP3R1, inositol-1,4,5-triphominute 2; N-BP, nitrogen-containing bisphosphonate; Nogenesis inhibitory factor; OC-STAMP, osteoclast stimumononuclear cells; PIP2, phosphatidylinositol 4,5-bisphT, protein-tyrosine phosphatase-proline, glutamic acid,; RGS, regulators of G-protein signaling; RISC, RNA-indupin RNA; siRNA, small interfering RNA; TLR3, toll-like ret acid phosphatase; TREM2, triggering receptor expresseviews theme issue on “Targeted delivery of therapeuticstics and Pharmaceutical Chemistry, University of Utah, Sa
ang), [email protected] (D.W. Grainger).
l rights reserved.
rainger, RNA therapeutics targeting osteocla.09.002
© 2011 Elsevier B.V. All rights reserved.
Contents
1. Introduction: the need for improved therapeutics for osteoporosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02. RNA interference and siRNA delivery as a new therapeutic approach. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 03. RNAi applications in new osteoporosis therapies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04. Osteoclastogenesis and osteoclastic bone resorption . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05. Targets for siRNA in osteoporosis and bone metabolism. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
5.1. Osteoclastogenesis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.2. Bone resorption targets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.3. Osteoclast viability targets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 05.4. Other potential osteoporosis targets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
6. siRNA delivery to bone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.1. Challenges and strategies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06.2. Non-specific siRNA-cell bioactivities and off-target side effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
7. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Acknowledgments. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
Argonaute; AP-1, activator protein-1; Ckb, brain-type creatine ki-DC-STAMP, dendritic cell-specific transmembrane protein; EGFR,; H+-ATPase, H+-adenosine triphosphatase; ICAM-1, intercellularsphate receptor-1; KCCs, K+/Cl− co-transporters; M-CSF, macro-CX, Na+/Ca2+ ion exchanger; NFATc1, nuclear factor of activatedlatory transmembrane protein; OGR1, ovarian cancer G-protein-osphate; PTK, protein tyrosine kinase; PTP-oc, osteoclast-specificserine, threonine amino acid sequence; RA, rheumatoid arthritis;ced silencing complexes; RIZ1, retinoblastoma protein-interactingceptor 3; TRAF6, TNF receptor-associated factor 6; TRAIL, TNF-re-d in myeloid cells-1; WASP, Wiscott-Aldrich syndrome protein.to bone and connective tissues”.lt Lake City, UT 84112-5820, USA. Tel.: +1 801 581 3715; fax: +1
st-mediated excessive bone resorption , Adv. Drug

2 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
1. Introduction: the need for improved therapeutics for osteoporosis
Current treatments for osteoporosis can be divided into two cate-gories: antiresorptive modulators and anabolic therapies. Estrogen,estrogen receptor modulators, calcitonin and bisphosphonates fallinto the first category. Only one anabolic agent–teriparatide–is clini-cally approved, administrated by daily injection [1]. The limited num-ber of options in antiresorptive therapy reflects inadequate efficacy,severe side effects, high dosing frequency or low patient compliance[1–6]: estrogen treatment exhibits known side effects on breast anduterus inappropriate for long-term use; long-term calcitonin treat-ment can induce tolerance since more than half of the patients pro-duce circulating antibodies to calcitonin [7]. Bisphosphonates, themost common current osteoporosis treatment, exhibit general sideeffects including gastrointestinal irritation, bone/joint pain and jawosteonecrosis [8–10]. The latter two adverse effects can persist andeven spread to new areas of unaffected bone after bisphosphonatetherapy is discontinued. Because of bisphosphonate's long half-life,e.g., alendronate's half-life in bone is 10.9 years [11], drug can re-main active in bone for long periods after drug therapy is discontin-ued. Severe suppression of bone turnover has also been noted[12,13]. Long-term bisphosphonate therapy increases fracture risk,including atypical fractures first reported in 2005 [14,15]. Recently,oral bisphosphonates were associated with 23 cases of esophagealcancer [16]. Therefore, maximum treatment duration for bispho-sphonates is suggested to be five years [17]. Bisphosphonates areconsequently counter-indicated in the context of fragility fracturehealing. This enduring drug bioactivity presents substantial chal-lenges for treatment of osteoporosis. Moreover, clinical studies ofsystemic bisphosphonate administration in combination with im-plants for 6 months or 2 years showed significant bone loss in theperi-implant area within three months post-operation [18,19]. Sim-ilarly, in total hip arthroplasty, up to 14% bone loss was reported dur-ing the first three months post-surgery [20].
Human clinical experience indicates that improved therapeuticstrategies are needed in the context of osteoclastic bone resorptiontherapies. In 2010, denosumab (Prolia™, Amgen), a fully humanizedmonoclonal antibody targeting receptor activator of nuclear factor-kappa B ligand (RANKL), was approved by the FDA as a twice-yearlysubcutaneous injection for treating postmenopausal osteoporosis.This antibody therapeutic appears well-tolerated and superior to thecommon bisphosphonate drug, alendronate, in preserving bone min-eral density in several clinical trials to date [21,22]. Additional newtherapeutics are actively under investigation to meet the require-ments of this fast-growing patient population worldwide.
2. RNA interference and siRNA delivery as a new therapeuticapproach
RNA interference (RNAi) is a sequence-specific post-transcriptionalgene silencing tool [23]. The process of gene-specific silencing throughdestruction of itsmRNA transcript can be triggered by endogenous or ex-ogenous small interfering RNAs (siRNAs) [24]. Long double-strandedmRNAs derived from endogenous gene transcription or transfectedtransgene plasmids present in the cytoplasm can trigger the cleavage ac-tivity of the intracellular enzyme, Dicer, to cut mRNA into 19-nucleotidepairs with two nucleotide overhangs at both 3′-ends, called small inter-fering RNA (siRNA) [25]. These double-stranded siRNA pieces then in-corporate into RNA-induced silencing complexes (RISCs) which have acatalytic core comprising Argonaute (Ago) family proteins [26,27].Ago-2 then cleaves and removes the siRNA sense strand and thereafterRISC becomes activated with the remaining antisense siRNA strand.The antisense siRNA strand guides RISC activity in the cytoplasm tocleave its targeted complementary mRNA molecules, resulting indown-regulation of the targeted gene and corresponding protein ex-pression. The mechanism of RNAi operation in mammalian cells is
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
shown in Fig. 1. First introduced in 2001, RNAi has been exploited forits highly specific mechanism of mRNA transcript targeting, and as a tar-get screening and validation tool for cell signaling studies of many types.
RNAi is used for investigating and elucidating mammalian gene func-tion as an alternative to knockout techniques. Specifically, siRNA alsoshows great potential for future targeted therapeutics for gene-associateddiseases. Gene silencing using siRNA has several advantages intrinsic toRNAi, such as its high specificity, intrinsic biological response [28] andmore efficient and specific silencing effects with lower dosing require-ments, compared to antisense-based gene silencing [29].
However, as polyanions, synthetic or in situ transcribed siRNAs do notreadily or reliably entermammalian cells. Themain challenge in develop-ing siRNA therapies, like other nucleic acid therapeutics, is to deliver themspecifically into targeted tissues or cells. Viral and non-viral vectors havebeen employed to address siRNA cell transfection inefficiency, and non-viral delivery is typically achieved by cationic lipoplexing reagents[24,30]. Viral vector-based delivery is consistently associated with vec-tor-based short hairpin RNA (shRNA) production systems, a DNA-basedstrategy to encode and obtain host-synthesized shRNAs in situ. TheseshRNAs can be further intracellularly processed into siRNA by Dicer.Both methods have their advantages and disadvantages. Non-viral deliv-ery uses siRNA directly to generate potent silencing effects; therefore, it issimple and controllable. However, single-dose siRNA silencing effects aretransient (up to five days in dividing cells) [24], and lipid-based siRNAde-livery complexes can be removed from circulation by the liver rapidly,and lack tissue/cell specificity. The viral vector-based shRNA strategyhas the potential of being able to provide stable, enduring gene silencing.Gene therapy can in principle continuously generate siRNA. The majorbottleneck of the viral vector is its well-known safety issues [24]. Never-theless, while non-viral delivery avoids the pitfalls of viral vector delivery,including high viral toxicity, possible carcinogenicity, proven immunoge-nicity, and significant cost limitations [31], it is extremely inefficient intargeting, transfection and expression. Because of the substantial chal-lengeswith reliable systemic siRNA delivery and targeting, almost all cur-rent clinical foci for siRNA-based therapeutics are based on local or topicalsiRNA therapeutics. Successful siRNA delivery approaches currently in-clude ocular, respiratory, central nervous system, dermal and vaginal de-liverywhere local dosing accesses target cell populations directly [32–36].One largely unexplored delivery route is via implantable combination de-vices facilitating local siRNA delivery directly from medical implants toadjacent tissue sites [37].
3. RNAi applications in new osteoporosis therapies
As a nucleic acid therapeutic precedent, DNA-based gene therapyhas developed rapidly for musculoskeletal applications in the lasttwo decades. The therapeutic approach has been introduced to vari-ous disease categories: osteogenesis imperfecta [38], lysosomal stor-age disorders [39], rheumatoid arthritis [40,41], osteoarthritis [42],and osteoporosis [43–45]. Specific to osteoporosis, gene transferstrategies deliver genetic material, either using intravenous injectionof viral vectors carrying osteoprotegerin (OPG) cDNA [44,45] or localinjection of interleukin-1 receptor antagonist cDNA-transduced cells[43]. Due to desirable short-term transgene expression without theneed to closely regulate transgene expression, DNA-based gene ther-apy has recently produced progress in musculoskeletal tissue healing.In a rat critical size defect model in femurs, BMP-2 cDNA-transducedcells seeded into collagenous scaffolds showed better healing com-pared with use of recombinant BMP-2 protein directly [46]. The feasi-bility of intralesional injection of viruses carrying cDNA encodingosteoinductive genes has been demonstrated in both rabbit and ratsegmental defect models [47,48]. Gene transfer strategies have beendeveloped for many applications for musculoskeletal healing, suchas spine fusion, articular cartilage and meniscus, intervertebral disc,ligament and tendon [49]. As DNA-based transgene therapies contin-ue to demonstrate the potential for treating musculoskeletal diseases,
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug
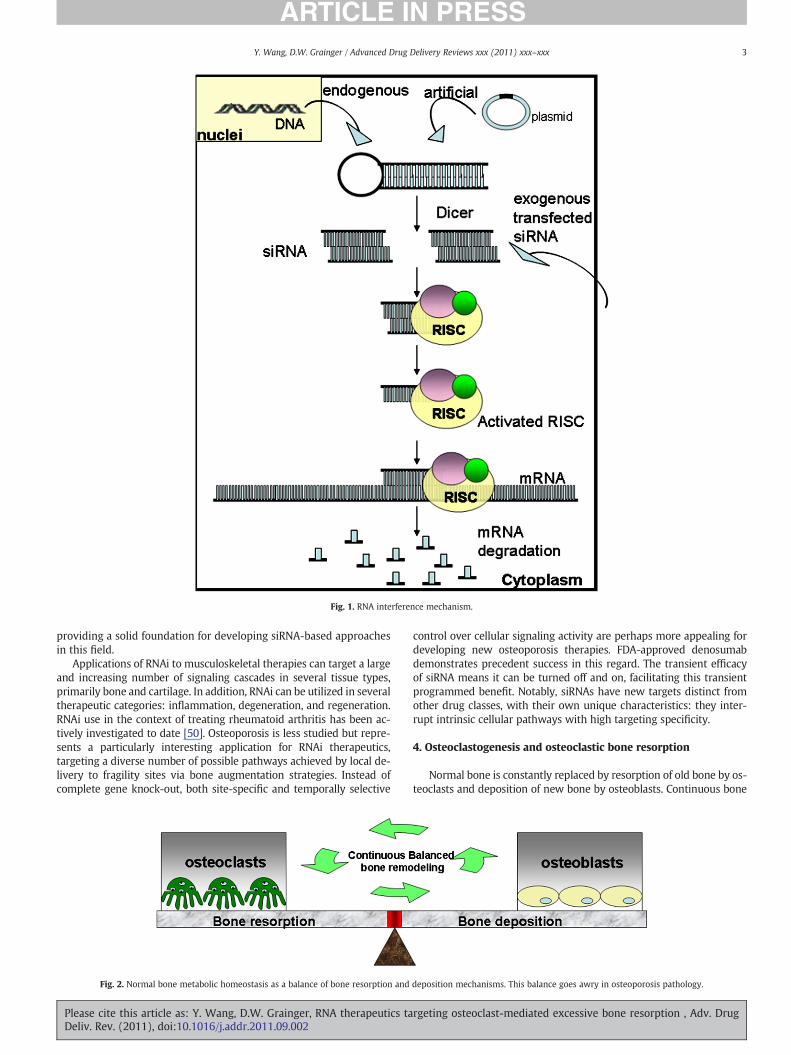
Fig. 1. RNA interference mechanism.
3Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
providing a solid foundation for developing siRNA-based approachesin this field.
Applications of RNAi to musculoskeletal therapies can target a largeand increasing number of signaling cascades in several tissue types,primarily bone and cartilage. In addition, RNAi can be utilized in severaltherapeutic categories: inflammation, degeneration, and regeneration.RNAi use in the context of treating rheumatoid arthritis has been ac-tively investigated to date [50]. Osteoporosis is less studied but repre-sents a particularly interesting application for RNAi therapeutics,targeting a diverse number of possible pathways achieved by local de-livery to fragility sites via bone augmentation strategies. Instead ofcomplete gene knock-out, both site-specific and temporally selective
Fig. 2. Normal bone metabolic homeostasis as a balance of bone resorption and
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
control over cellular signaling activity are perhaps more appealing fordeveloping new osteoporosis therapies. FDA-approved denosumabdemonstrates precedent success in this regard. The transient efficacyof siRNA means it can be turned off and on, facilitating this transientprogrammed benefit. Notably, siRNAs have new targets distinct fromother drug classes, with their own unique characteristics: they inter-rupt intrinsic cellular pathways with high targeting specificity.
4. Osteoclastogenesis and osteoclastic bone resorption
Normal bone is constantly replaced by resorption of old bone by os-teoclasts and deposition of new bone by osteoblasts. Continuous bone
deposition mechanisms. This balance goes awry in osteoporosis pathology.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

4 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
turnover results in the adult human skeleton being completely replacedevery 10 years [51]. This balance of bone turnover is tightly regulated inhealthy individuals as shown in Fig. 2. Osteoclasts residing at or near thebone surface are multinucleated phagocytic cells formed by the fusionof monocyte macrophage precursor cells [52]. They are responsible forresorbing bone, working together with osteoblasts (bone producingcells), and playing a central role in normal bone remodeling. Two cyto-kines – receptor activator of nuclear factor kappa B (NF-κB) (RANK) li-gand (RANKL, also called TRANCE/OPGL/ODF) and macrophage colony-stimulating factor (M-CSF) [53,54] – are essential to this process [55–57] as shown in Fig. 3. Previously published evidence shows that osteo-clast formation can be stimulated in vitro using co-cultures of bonemarrow cells and stromal cells/osteoblasts expressing those two cyto-kines [58–60]. M-CSF interacting with its c-fms receptor provides sig-nals necessary for precursor cell survival and proliferation by receptorbinding on early osteoclast precursor cells [61]. RANKL is the key cyto-kine for promoting osteoclastogenesis. Interactions between solubleRANKL and RANK receptor on the surface of osteoclast precursors areessential for expression of osteoclast-specific genes [62], bone resorp-tion and survival of mature osteoclasts [63]. Furthermore, osteoclasto-genesis is negatively regulated by osteoprotegerin (OPG, orosteoclastogenesis inhibitory factor, OCIF), also expressed by stromalcells and osteoblasts [64]. OPG is a decoy receptor that competes withRANK for binding RANKL [65]. Osteoclast production can be blockedby over-expression of OPG, resulting in osteopetrosis in mice [64,66].In this RANKL/RANK/OPG regulatory axis, positive regulator RANKLand negative regulator OPG are coordinated through interaction withRANK to regulate normal bone formation and degradation.
RANK is therefore a central factor in this bone metabolic regulato-ry pathway. It is central to much of the osteoclast functional pheno-type and to bone metabolic balance. Therefore, RANK was chosen asthe knock down target to suppress osteoclast-mediated bone resorp-tion [67]. Selective knockdown of RANK in mouse bone marrow cellscan significantly block formation of tartrate resistant acid phospha-tase (TRAP)-positive cells with RANKL and M-CSF in vitro. Cell–cellfusion can be inhibited in RANK siRNA-transfected osteoclasts, consis-tent with the fact that RANKL stimulation is critical for cell fusion toosteoclasts [68]. Successful transfections have been seen in matureosteoclasts and osteoclast-mediated bone resorption was dramatical-ly inhibited by RANK siRNA using the pit formation assay in vitro [67].
Activation of RANK by RANKL not only induces osteoclast differenti-ation and survival, but also leads to activation of bone resorption byma-ture osteoclasts. Once activated, differentiated osteoclasts move to thetarget matrix and attach to the bone surface. After attachment, an isolat-ed extracellular microenvironment, called the sealing zone, formed be-tween the interface of the underlying bone with osteoclastmembranes, is actively generated by osteoclasts [69]. Osteoclasts polar-ize themselves to form a ruffled membrane which is their resorptive or-ganelle adjacent to the bone surface. Osteoclasts then acidify anddissolve the underlying bonematrix by pumping hydrogen ionsmediat-ed by a vacuolar H+-adenosine triphosphatase (H+-ATPase) through
Fig. 3. Cellular mechanisms of osteoclastogenesis involving osteoblast/macrophage precursor
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
the ruffled membrane into the sealing zone. Osteoclast intracellular pHis maintained by HCO3
−/Cl− exchange across cell surfaces other thanthe ruffled membrane area [70]. These ion transporting events result inan acidic pH of ~4.5 only within the resorptive microenvironment [71].The mineral phase of bone is first digested under this acidic milieu, fol-lowed by degradation of the collagen-rich demineralized organic com-ponent of bone by released hydrolytic enzymes [72]. Bone matrixdegradation proteins are taken up by osteoclasts through the ruffledmembrane from the resorption lacuna and released through the basolat-eral membrane into the extracellular space [73]. After local bone resorp-tion is complete at this site, osteoclasts detach from the bonematrix andmigrate to a new site to begin the next functional cycle.
Because of this central role in bone processing, osteoclasts are nat-urally a major target for most studies seeking to suppress bone re-sorption. In this review, siRNA targets for osteoporosis are sortedinto three categories: osteoclastogenesis, osteoclastic bone resorptionactivity and osteoclast survival, including other potential associatedtargets identified to date. In addition, progress in developing efficienttarget silencing, delivery issues, and emerging opportunities andchallenges to exploit siRNA for therapeutic purposes are discussed.
5. Targets for siRNA in osteoporosis and bone metabolism
5.1. Osteoclastogenesis
Transcription factors essential for osteoclastogenesis, includingNF-κB downstream of RANK [74], mediate osteoclast differentiationand inflammatory osteolysis [75]. Osteoclast differentiation failure isreported to be caused by mutant NF-κB in mice [76]. Its upstream regu-lator, IκB kinase (IKK) complex, contains IKKα and IKKβ as catalytic sub-units and IKKγ as a regulatory subunit [77,78]. IKKα and IKKβ arerequired for normal bone homeostasis. IKKβ is essential for osteoclasto-genesis and osteoclast survival since deletion of IKKβ impairs osteoclastdifferentiation in vitro and in vivo [79,80]. IKK alpha(−/−) mice did notshow overall skeletal defects, but IKK alpha(−/−) hematopoietic cellsfailed to differentiate into multinucleated osteoclasts [81]. Nonetheless,the role of IKKγ is unclear. Recently, transient transfection of siRNA to in-hibit IKKγ in bone marrow-derived osteoclast precursors was reportedusing a retroviral delivery approach [82]. When IKKγ was knockeddown, formation of multinucleated osteoclasts was reduced 83% com-paredwith controls. In addition, the role of IKKγwas confirmed throughobservation that RANKL-induced osteolysis was impeded by preventingoligomerization of IKKγ monomers using peptides in mice [82]. Thestudy suggested that IKKγ is essential for osteoclastogenesis, and thus,all three IKK subunits could be potential targets for inhibiting osteoclas-togenesis and osteolysis using siRNA.
Interactions between RANKL and RANK are essential for osteoclas-togenesis. RANK has three TNF receptor-associated factor 6 (TRAF6)binding sites and recruits TRAF6 to activate the transcription factorsNF-κB and activator protein-1 (AP-1) [83,84]. Nuclear factor of acti-vated T cells (NFATc1) has been demonstrated as the transcription
s cell interactions, mediated by RANK, RANKL, and OPG among other signaling partners.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

5Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
factor most strongly induced by NF-κB activation [85]. NFATc1 is alsoknown to be important in immune response and also plays an essen-tial role in osteoclastogenesis [85,86]. Therefore, siRNA targetingNFATc1 in both RAW264.7 cells and RAW-induced osteoclasts underLPS stimulation was examined [87]. Specific knockdown of NFATc1led to reduced formation of mature osteoclasts and significantly de-creased osteoclast-specific gene expression, including TRAP and ca-thepsin K, both responsible for effective osteoclastic bone resorption.
Once RANK is activated by its ligand, several signaling cascades areinduced during osteoclast formation or activation, resulting in activa-tion of several transcription factors mediated by protein kinases [88],such as c-Fos, NF-κB, c-Jun, NFATc1 [84]. Transcription factor c-Junhas recently shown its essential role in osteoclastogenesis [89]. Inter-action between RANK and RANKL can stimulate JNK and activate c-Jun which complexes with c-Fos downstream to form AP-1 [84].Moreover, c-Jun was reported to regulate osteoclastogenic activitiesof NFATc-1: dominant-negative c-Jun transgenic mice were unableto form osteoclasts due to arrested activation and expression of theNFAT family proteins [90]. Recently, using a proteomic approach, amember of a family of growth regulatory genes, schlafen2 (Slfn2),was found to be highly induced by RANKL activation and capable ofregulating c-Jun activation [91]. Using RNA silencing technology,knockdown of Slfn2 resulted in significantly reduced expression ofc-Jun and NFATc1 and 50% less TRAP-positive multinucleated cellscompared with control. Furthermore, over-expression of Slfn2 en-hanced c-Jun phosphorylation. Slfn2 therefore functions downstreamof RANK/RANKL signaling and upstream of c-Jun and NFATc1 inosteoclastogenic regulation.
Dendritic cell-specific transmembrane protein (DC-STAMP), aseven subunit-transmembrane protein originally found in dendriticcells [92] is highly expressed in osteoclasts but not in macrophages[93]. Expression levels of DC-STAMP can be rapidly up-regulatedunder RANKL-mediated osteoclastogenesis in osteoclast precur-sors [94]. Using siRNA to specifically knock down its expression inRAW-D cells completely abrogated the formation of multinucleatedosteoclast-like cells stimulated with RANKL and TNF-α in vitro [94].Consistent with this, osteoclastogenesis can also be enhanced byover-expression of DC-STAMP and vice versa [94]. In addition, DC-STAMP has been further proven essential for both osteoclast and for-eign body giant cell fusion [93]. No multinuclear osteoclasts wereobserved in bone sections from DC-STAMP-deficient mice, and theirmononuclear cells were unable to differentiate to multinuclear osteo-clasts under stimulation with RANKL and M-CSF in vitro, remainingmononuclear, TRAP-positive cells [93]. These cells are able to resorbbone, but with much lower efficiency, exhibiting the pathogenesisof moderate osteopetrosis [93]. Consistent with these results, bonemineral density and bone volume per tissue volume in DC-STAMP(−/−) mice increased compared with wild type controls [93]. DC-STAMP is therefore concluded to be indispensable for osteoclastmultinucleation.
A novel gene bearing significant similarity to the DC-STAMP fam-ily, called osteoclast stimulatory transmembrane protein (OC-STAMP), was identified in both primary bone marrow cells andRAW264.7 cells in 2007 [95]. It is strongly up-regulated in responseto RANKL stimulation as a multiple transmembrane protein in osteo-clasts. Blocking OC-STAMP expression by either RNA interference orOC-STAMP antibodies showed similar results as blocked DC-STAMP:mononuclear, TRAP-positive cells [95]. The results suggest that bothDC-STAMP and OC-STAMP are responsible for cell fusion and eachof them cannot compensate for the other since knock down of onlyone of them impaired osteoclast multinucleation.
A study aimed at protecting biomaterials by inhibiting macro-phage fusion at implant sites showed that using siRNA targetingRac1 can limit fusion without limiting phagocytosis [96]. As thestudy attempted to find the fusion target shared by foreign bodygiant cells formed by monocyte fusion and also by osteoclasts,
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
osteoclast formation theoretically should be inhibited as well. Furtherstudy is required to confirm the effect of Rac1 silencing on monocytefusion to osteoclasts.
A membrane glycoprotein, CD9, one of the tetraspanins, has beenreported to play an important role in cell fusion during osteoclastogenesis[97]. CD9 has been reported to be involved in cell–cell fusion and cellmo-tility in different cells [98,99]. In RAW264.7 cells, CD9 locates to the cellmembrane and its expression is up-regulated by RANKL stimulation[97]. Targeted inhibition of CD9 using siRNA is able to significantly inhibitthe formation of RAW264.7 differentiated osteoclasts by inhibiting cellfusion in cultures. Over-expression of CD9, interestingly, can promotecell fusion in RAW264.7 cultures without RANKL stimulation, but fusedcells are TRAP-negative. In the same study, CD9 is confirmed to beexpressed in osteoclasts in vivo by immunohistochemical analysis of tis-sue sections from mouse femoral bone.
During the osteoclastogenesis cell–cell fusion process, sialic acidwhich is involved in a number of biologic responses, was hypothesizedto play a role in this process [100]. Takahata et al. [101] found thatalpha(2,6)-linked-sialic acid degraded during osteoclast differentiationand desialylated cells only could form mononuclear TRAP+ cells withnormal expression of osteoclast markers. Using siRNA to knock downalpha(2,6)-sialyltransferase produced significant inhibition of osteoclastmultinucleation, which suggests a role for alpha (2,6)-linked-sialic acidin cell–cell fusion processes during osteoclastogenesis.
A novel gene, NHA-oc/NHA2, is significantly upregulated in RANKL-stimulated osteoclast precursors [102]. As the murine orthologue of thehuman gene HsNHA2 [103], encoding a cation-proton antiporter (CPA)localized on themitochondria, it is proposed to regulate proton concen-tration in osteoclast mitochondria. NHA-oc/NHA2 selectively expressedin differentiated osteoclasts regulates Na+-dependent mitochondrialpH changes and mitochondrial passive swelling. Importantly, NHA-oc/NHA2 siRNA inhibits TRAP-positive multinucleated cell formationand resorption activity under RANKL stimulation. This reduction par-tially resulted from apoptosis induced from the loss of inhibition of cas-pase-9 activation by NHA-oc/NHA2. Therefore, NHA-oc/NHA2 isintegrally involved in osteoclast formation, resorption and survival.
Ovarian cancer G-protein-coupled receptor 1 (OGR1) is a histi-dine-enriched proton-sensor. The OGR1 gene was found to be 7-foldup-regulated in the long bone of CSF-1-null osteopetrotic rats afterCSF-1 injections compared to untreated mutants [104]. Expressionof OGR1 can also be induced in RAW264.7 cells under RANKL stimu-lation during osteoclast differentiation. Specifically knocking downOGR1 expression by siRNA (≥1 μg/ml) produced more than 50% inhi-bition of osteoclast formation in both mouse bone marrow mononu-clear and RAW264.7 cells without significant cell death.Concomitant with confirming OGR1's role in regulating osteoclasto-genesis, the expression and function of the regulators of G-proteinsignaling (RGS) proteins of osteoclastogenic RANKL signaling cas-cades were investigated. RGS proteins have been suggested to physi-ologically regulate G-protein cycles and G protein signaling inhematopoietic cells [105,106]. In this protein family, RGS18 specifi-cally expressed in hematopoietic cells, showed consistent decreasesin mRNA expression levels during RANKL stimulated osteoclastogen-esis [107,108]. Target-specific knockdown of RGS18 in RAW264.7cells using siRNA resulted in enhanced osteoclast formation underRANKL induction. Additionally, antibodies against OGR1 with Zn2+
ion addition (antagonist of OGR1) [109] significantly reversed the ef-fects of RGS18 siRNA. These observations suggest that the ability ofRGS18 to suppress osteoclastogenesis depends on the OGR1 signalingpathway and inhibition of OGR1-mediated cell signaling [108].
Another RGS member, RGS12, is reported to be significantly upre-gulated after RANKL stimulation, with its expression being dose-de-pendent on RANKL [110]. Using vector-based RNAi technology,stable RGS12-silenced RAW264.7 cells showed 18.7 times lowernumbers of TRAP-positive multinucleated cells under RANKL stimula-tion compared with control groups. The mechanism of completely
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

6 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
blocking osteoclast differentiation was suggested to lie in regulationof intracellular Ca2+ oscillations in the NFAT2 pathway during differ-entiation. Such Ca2+ oscillations can be completely blocked and theexpression of NFAT2 simultaneously significantly inhibited in RGS12-silenced cells [110].
Triggering receptor expressed in myeloid cells-2 (TREM2) is pro-duced in myeloid cells in bone marrow and locates to these cell sur-faces. Interactions between TREM2 and DAP12 transmit signals toactivate the cells. Osteoclast precursor cells harvested fromDAP12 (−/−) mice only form mononuclear TRAP-positive cellsunder stimulation with RANKL and M-CSF. These cells exhibit only50% ability to resorb bone compared with wild type cells [111].siRNA against TREM2-transfected RAW264.7 cells largely reducesnumbers of TRAP-positive cells, and most were small (b10 nuclei)or mononucleated cells. Thus, TREM2/DAP12 signaling plays an im-portant role in osteoclast differentiation under RANKL stimulation[112]. The same study blocked TREM2 in murine primary cell-inducedosteoclasts using antibodies and showed decreased bone resorption,providing evidence that the TREM2/DAP12 signal also regulates oste-oclast function.
Extended space flight can cause severe bone loss in astronauts dueto microgravity. Compared with normal gravity, microgravity can ac-celerate osteoclastogenesis about 2-fold [113]. Gene expression pro-filing of RAW264.7 cells under microgravity conditions has shownincreased expression of the calcium-binding protein A8/calgranulinA (S100A8). siRNA knockdown of S100A8 in RAW264.7 cells signifi-cantly suppressed osteoclastogenesis under microgravity conditions.S100A8 could be a therapeutic target for preventing bone loss duringextended space flights.
Retinoblastoma protein-interacting zinc finger 1 (RIZ1) protein alsoparticipates in RANKL-induced osteoclastogenesis [114], binding withboth retinoblastoma protein and estrogen receptors and reportedly in-volved in osteosarcoma [115–117]. RIZ1 expression increases underRANKL induction at 24 h, and RIZ1 siRNA-transfected RAW264.7 cellsshowed significantly inhibited NFATc1 activation 3 days post-RANKLandM-CSF treatment, but with no significant influence on TRAF6 expres-sion. Thus, RIZ1 is suggested to positively regulate NFAT-1 activity at thelast stages of osteoclastogenesis. RAW264.7 cells with reduced RIZ1 ex-pression exhibited substantially reduced ability to form TRAP-positivemulti-nucleated osteoclast-like cells under RANKL and M-CSF stimula-tion. Recently, c-Abl SH3 domain-binding protein-2 (3BP2) has been rec-ognized as a key regulator of RANKL-induced osteoclastogenesis [118].3BP2 regulates several immunoreceptor signaling pathways in immunecells, such as T, B, and NK cells, and was reported to promote activationof NFAT in T and B cells [119–123]. Knockdown of 3BP2 in RAW264.7cells decreased expression of NFATc1 and completely suppressedRANKL-induced TRAP-positive multinucleate cell formation [118].
The family of disintegrin and metalloproteinases (ADAM) peptidesare cell surface proteins involved in cell adhesion and cell fusion [124].ADAM8 is a transmembrane glycoprotein expressed in monocytes andsignificantly up-regulated during osteoclastogenesis [125,126]. Recently,ADAM8 has been shown to promote osteoclastogenesis: vitamin 1,25-(OH)D3-stimulated osteoclast formation was inhibited by ADAM8 anti-sense treatment [126]. Its direct effect on osteoclastogenesis was also in-vestigated using siRNA [127]. Silencing ADAM8 in RAW264.7 cellsdecreased osteoclast formation and cell size compared with controlsand modestly decreased osteoclast marker mRNA levels when stimulat-ed with RANKL. In contrast, transfection of ADAM8 into RAW264.7 cul-tures increased osteoclast marker mRNA expression levels andincreased the number of TRAP-positive cells. The study also showedthat ADAM8 was highly expressed in rheumatoid arthritis (RA) pannusmacrophages and multinucleated cells adjacent to eroded cartilage.Therefore, ADAM8 was a recommended target for suppressing RA pro-gression by preventing osteoclast formation.
Epidermal growth factor receptor (EGFR) has been the focus of in-creasing attention in osteoclastic bone resorption, shown to promote
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
bone resorption in vitro [128]. Its inhibition in vitro resulted in sup-pression of bone marrow stromal cell induced osteoclast differentia-tion [129]. Yi et al. [130] investigated EGFR function in osteoclastformation and survival, and found that EGFR expression was highlyup-regulated by RANKL. Knockdown of EGFR in bone marrow mono-cytes using lentivirus expressing shRNA completely suppressedosteoclastogenesis with RANKL and M-CSF. In addition, EGFR siRNAblocked NFATc1 expression completely under RANKL stimulation.This study also showed that EGFR co-immunoprecipitated withRANK and Gab2 which mediates the RANK signaling cascade. Thesedata imply that EGFR may couple with RANK via the signaling medi-ator Gab2 to regulate RANK-stimulated downstream pathways, play-ing an important role in osteoclastogenesis.
Multinucleated osteoclasts are formed by fusion of monocytemacro-phages. Therefore, the role of integrins important for trafficking mono-cytes in osteoclastogenesis has been assessed using siRNA. Only twointegrin pairs are found on pre-osteoclast monocytes: CD11a/CD18(LFA-1), and CD11b/CD18 (Mac-1) which is considered the premiermarker of pre-osteoclasts [131,132]. Antibodies against CD11b andCD18 to blockMac-1 both in RAW264.7 and primary cell culture can sig-nificantly inhibit osteoclastogenesis, but not antibodies against CD11a[133]. Specific knockdown of CD11b using siRNA in RAW264.7 cells,yielding a 50% decrease in CD11b expression, resulted in ~50% reductionin osteoclast area. This inhibition was suggested to occur in the earlystages of cell differentiation since it was accompanied by a three-fold in-crease in the mRNA expression level of NFATc1, a major regulator ofosteoclastogenesis [133]. Taken together, integrin Mac-1 is suggestedto play an important role in early stage osteoclast differentiation viapre-osteoclast cell–cell interactions.
A new member of the TNF family that induces cell apoptosis, calledTNF-related apoptosis-inducing ligand (TRAIL), shares a 25% aminoacid homology with RANKL [134] and is reported to inhibit osteoclasto-genesis in both human peripheral bloodmononuclear cells (PBMC) andRAW264.7 cells, possibly by inhibiting the p38/MAPK pathway [135]. Itis also involved in human osteoclast apoptosis [136]. More recently,TRAIL has been found to inhibit the accumulation of RANKL-dependentp27Kip1 in PBMC [137]. p27Kip1 is a cyclin-dependent kinase inhibitorshown to be progressively upregulated to play an important role inme-diating RANKL-induced murine osteoclastogenesis [138,139]. Additionof TRAIL into pre-osteoclast cultures incubated with RANKL and M-CSF resulted in the 6-fold decrease in formation of TRAP-positive cellsand significant reduction in the expression of p27Kip1 [137]. UsingsiRNA to specifically knock down p27Kip1 in PBMC culture dramaticallyreduces formation of TRAP-positive cells induced by RANKL andM-CSF,consistent with the results of p27Kip1-deficient mice [139]. Taken to-gether, p27Kip1 function alone appears essential for RANKL-mediatedosteoclastogenesis.
5.2. Bone resorption targets
After osteoclasts mobilize the bone mineral phase, several hydro-lytic enzymes degrade the organic bone component. The principalprotease involved in this process is cathepsin K [72,140], a lysosomalprotease mainly expressed in osteoclasts and released from the ruf-fled membrane to the resorption lacunae in the sealed zone. Cathep-sin K mRNA and protein expression levels are stimulated by RANKL ina time- and dose-dependent manner, and increase with osteoclastdifferentiation and activation [141–143]. Cathepsin K efficiently de-grades type I and II collagen, and gene mutation can cause osteope-trosis and exhibit features of pycnodysostosis [140,144,145]. Specificdown-regulation of cathepsin K mRNA expression decreased boththe number and area of bone pits more than 50% without significantlychanging cell viability [146]. The unchanged osteoclast number maybenefit maintenance of bone homeostasis since osteoclast and osteo-blast activities are closely correlated, and disruption of this communi-cation may cause other issues [147]. In mature osteoclasts, cathepsin
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

7Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
K expression is increased by RANKL, both in vitro and in vivo [141].RANK–RANKL binding activates at least five distinct downstream sig-naling pathways [148]. TRAF6 acts as a critical adaptor in bindingwith the cytoplasmic domain of RANK; its mutations cause osteope-trosis [149,150]. Activation of transcription factor AP-1 has been sug-gested to result from TRAF6 signaling [143]. AP-1 is a heterodimericprotein composed of Jun (c-Jun, JunB or JunD) and Fos [151]. Trans-fecting RAW264.7 cells with either the dominant negative form ofc-fos or siRNA against either c-jun or junB down-regulated RANKL-mediated cathepsin K gene expression. Both transfections inhibitedCTSK promoter activity, suggesting that AP-1 stimulated the cathep-sin K promoter [143]. In addition, c-Fos is known to play a criticalrole in osteoclastogenesis: over-expression of c-Fos rescues RANKL-induced osteoclast formation previously blocked by N,N-dimethyl-d-erythro-sphingosine treatment [152].
H+-ATPase in the osteoclast ruffled membrane is responsible forreducing and maintaining the low pH in the sealed resorption lacunaduring the process of osteoclast-mediated extracellular acidification.The ATPases have multiple subunits. One subunit isoform located inthe ruffled membrane, subunit a3, is reported to be highly expressedand essential to osteoclast resorption function. Defects in this subunitof the vacuolar proton pump induce severe osteopetrosis [153,154].Hu et al. [155] showed successful carrier-free a3 siRNA transfectionof primary rat osteoclast cultures, reporting that actin ring structurescharacteristic of actively resorbing osteoclasts were reduced to 20% ofcontrols after siRNA transfection, but re-emerge after halting trans-fections. Bone resorption pits were significantly reduced 48 h aftertransfections and type I collagen C-terminal cross-linked telopeptidesin the culture media were decreased more than 50% compared to con-trols or non-targeting siRNA treated groups. H+-ATPase knockdownis similar with cathepsin K knockdown: specific gene down-regula-tion reduces osteoclastic resorption activity without inducing celldeath.
H+-ATPases have two domains: V1 located on the cytoplasm sideand V0 bound within the membrane [156]. ATP-hydrolytic domain V1has 8 individual subunits, A–H [157,158]. Subunit C in murine H+-ATPase has three isoforms: Atp6v1c1 (C1), Atp6v1c2a (C2a), andAtp6v1c2b (C2b) [159]. Only C1 is highly expressed in osteoclastscompared to the other two, and is highly up-regulated after RANKLand M-CSF stimulation during differentiation [160]. Feng et al. [160]depleted C1 expression in murine bone marrow macrophages withno significant difference in numbers of TRAP-positive multinucleatedcells compared with untreated cultures. However, osteoclastic abilityto actively produce H+ to acidify the extracellular sealing zone envi-ronment was largely impaired and areas of resorption pits on dentinslices by siRNA-treated osteoclasts were 0.5–0.75% of control cultures.Co-immunoprecipitation studies showed that C1 interacted with an-other essential ruffled border H+-ATPase subunit a3 and co-localizeswith microtubules, but distinct from a3 deficiency [62], C1 depletionwas able to impair F-actin ring formation [160]. C1 was therefore sug-gested to be a regulator of F-actin sealing ring formation during oste-oclast activation. Taken together, C1 knockdown does not affectosteoclastogenesis, but inhibits osteoclastic bone resorption.
Electrostatic neutralization of themembrane potential gradient dur-ing osteoclast acid secretion is maintained by Cl− ion channels on theosteoclast's ruffled border [161]. ClC-7 Cl− channels are highlyexpressed in osteoclasts and localize to the ruffled border membrane.Disruption of ClC-7 in both humans andmice produced severe osteope-trosis [161]. With the exception of ClC-7, ClC-3 in the CLC family ofchannels was identified in osteoclasts as the functional Cl− ion-specificchannel in intracellular organelles, such as endosomes and lysosomes[162]. Both organelle acidity and bone resorption activity are reducedusing siRNA against ClC-3. Okamoto et al. concluded that observed oste-oclastic bone resorption relied on such internal organelle acidification,since ClC-3 only locates to intracellular organelles, influencing this or-ganelle acidity exclusively. However, bone resorption activity remains
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
in the absence of ClC-3 activity in osteoclasts, attributed to the role ofClC-7 or other redundant Cl− ion channels [162,163]. Since severeosteopetrosis is observed in ClC-7 chloride channel-deficient mice[161], the functional role of K+/Cl− co-transporters (KCCs) in mouseosteoclast-mediated bone resorption has been a focus. KCCs exist inmany tissues and cells, transporting K+ and Cl− ions each driven bytheir respective individual chemical gradients [164,165]. A previousstudy showed that both ClC-7 and KCC1 mRNA were expressed in mu-rine osteoclasts, with KCC1 locating to the cell membrane [163]. TheKCC inhibitor, R(+)-butylindazone (DIOA) was able to increase the in-tracellular concentration of both Cl− and H+ in resorbing osteoclasts.Thus, inhibiting KCCs can suppress Cl− secretion in resorbing osteo-clasts, which eventually produces reduced H+ pumping activity. Trans-fecting bone marrow cell-induced osteoclasts with siRNA against KCC1dramatically reduces of the area of bone resorption pits compared withcontrols and these results are similar, but slightly less potent, to thosefrom ClC-7 inhibition [163].
The Na+/Ca2+ ion exchanger (NCX) is a bi-directional membranetransporter regulating Ca2+ homeostasis in many tissues on cell plas-ma membrane [166]. The NCX family comprises homologous pro-teins: NCX 1, NCX 2, and NCX 3 [167,168]. In murine osteoclasts,three splice variants of NCX1 and NCX3, called NCX1.3, NCX1.41 andNCX3.2, were detected [169]. NCX mediates both Ca2+ efflux and in-flux and is predicted to locate to the ruffled border and control Ca2+
influx in resorbing osteoclasts [169]. SiRNA delivered against eachNCX1.3, NCX1.41 and NCX3.2 at 100 nM in mouse bone marrowcell-induced osteoclasts all significantly reduced the area of bone re-sorption pits per osteoclast approximately 50% [169]. These data sup-port the essential role of these three NCX in Ca2+ transport and theirimportant role in regulating bone resorption.
The c-Src gene, encoding a cytosolic protein tyrosine kinase (PTK)[170], is also essential for osteoclast activity. Lack of c-Src expressionin mice resulted in osteopetrosis characterized by inactive osteoclasts[171]. Previous work indicates that c-Src is functional as an adaptor inosteoclasts, regulating cell attachment and migration by recruitingessential signaling proteins [172]. Strong c-Src PTK activity has beenfound in ruffled borders in active osteoclasts [173]. Osteoclast-medi-ated bone resorption can be abolished both in vivo and in vitro byblocking c-Src PTK activity in osteoclasts [174]. Knockdown of Src ex-pression in RAW264.7 cells reduces both size and number of actinrings, and decreased cell spreading and fusion rates [175]. c-Src PTKactivity is activated by dephosphorylation, and a structurally uniqueosteoclast-specific transmembrane protein-tyrosine phosphatase(PTP-oc) in osteoclasts is known [176]. RAW/C4 osteoclast-like pre-cursor cells transfected with PTP-oc siRNA in serum-containingmedia and showed RANKL-mediated suppression of osteoclast differ-entiation. Both osteoclast number and size were significantly reducedby decreasing endogenous PTP-oc mRNA levels [177]. In addition, ap-optosis was induced significantly by PTP-oc siRNA compared to con-trol siRNA. Enhanced apoptosis from such knock down also suggestsits role in osteoclasts. Additionally, PTP-oc is also indicated to be in-volved in regulating RANKL-mediated osteoclastic differentiation[178]. A homologous recombination strategy was used to inhibitPTP-oc function in RAW264.7 cell culture. PTP-oc knockout cellscould not form TRAP-positive multinucleated osteoclast-like cellsunder RANKL treatment after 7 days. Therefore, PTP-oc is a targetnot only on mature osteoclasts to inhibit bone resorption, but alsoon osteoclast precursors to limit differentiation into osteoclasts.
The osteoclast sealing zone requires tight attachment between os-teoclasts and the bone surface for successful bone resorption [179]and this depends upon formation of a dense belt-like actin ring sur-rounding the ruffled membrane [180]. These osteoclast actin ringshave high concentrations of actin filaments assembled locally into dy-namic structures as podosomes in bone attachment sites. With its sig-nificant prerequisite role in osteoclastic bone resorption, actin ringformation is a target for inhibiting osteoclasts and has been
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

8 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
investigated in many studies. Gelsolin is an important actin regulator,necessary for podosome formation but not necessary for actin ringformation, which means that gelsolin-null osteoclasts are able to re-sorb bone, but with reduced potency [181–183]. However, Wiscott-Aldrich syndrome protein (WASP), critical for podosome assemblyexists in the actin ring of gelsolin-null osteoclasts [182]. WASP inter-acts with phosphatidylinositol 4,5-bisphosphate (PIP2) and Cdc42 inresponse to integrin ανβ3 signaling, enhancing the sealing zone for-mation and bone resorption [182]. Mice without WASP expressionexhibited defects in formation of podosomes and actin rings in thesealing zone and bone formation [184]. Attenuation of WASP usingsiRNA demonstrated the absence of podosome formation of actinrings and reduced bone resorption. WASP is activated by the coordi-nated binding of Cdc42 and PIP2, and full activation stimulates theactin-nucleating function of the actin-related protein (Arp)2/3 com-plex by associating with Arp2/3 [185–188]. The Arp2/3 complex initi-ates actin nucleation and crosslinking, and has been reported to be 3-fold upregulated in RAW264.7 cells in response to RANKL [189,190].Knock down of Arp2 in RANKL-stimulated RAW264.7 cells usingsiRNA generated an average 70% protein reduction 30 h post-trans-fection [190]. Fewer podosome-like structures appeared and com-pared with control, less than 1% actin rings were observed afterknockdown, proving the vital role for Arp2/3 in actin ring formationand its potential to be a new target for therapeutic agents. Actin nu-cleation requires phosphorylation of WASP's C-terminal VCA domainduring WASP binding to Arp2/3 complex after integrating a numberof signals [191–193]. The PTP-PEST (protein-tyrosine phosphatase-proline, glutamic acid, serine, threonine amino acid sequence) hasbeen shown to be involved in WASP and Src phosphorylation and de-phosphorylation [194]. This regulates phosphorylation of proteins as-sociated with WASP and enhances interactions between WASP, actinmonomers, and the Arp2/3 complex by increasing the WASP confor-mational stability [194]. At the same time, it regulates Src activitythrough (de)phosphorylation [195]. Reducing PTP-PEST expressionlevels using siRNA in osteoclasts eliminated actin rings and signifi-cantly inhibited formation of the sealing zone, formation of WASP–cortactin–Arp2 complexes, [195] and bone resorption [194]. This indi-cates a direct role of PTP-PEST in the formation of cell-sealing zoneswhile interacting with WASP.
Simultaneously with WASP, cortactin, a c-Src substrate, binds withArp2/3 at its N-terminal acidic domain to promote Arp2/3-inducedactin assembly and stabilize actin filaments by binding to their repeat re-gions [196]. Cortactinwas not found in hematopoietic cell precursors, butis induced in differentiated osteoclasts. Depletion of cortactin in primarymouse osteoclasts using siRNA resulted in the absence of podosomes,sealing rings and loss of bone-resorbing ability, suggesting an indispens-able role for cortactin in osteoclastic bone resorption. Cortactin's mecha-nism for regulating sealing ring formation has been further studied [197].Instead of inhibiting the initial actin aggregate phase, knockdown of cor-tactin using siRNA suppressed the subsequent sealing zone maturationprocess.
Osteoclast ability to attach to bone surfaces is essential for suc-cessful bone resorption. An intracellular calcium channel, inositol-1,4,5-triphosphate receptor-1 (IP3R1) is involved in the regulationof reversible osteoclast attachment [198]. IP3R1 can bind an endoso-mal isoform of IP3R-associated cGMP-dependent kinase substrate(IRAG). Using siRNA against IRAG in human osteoclasts in vitro re-duced cell spreading diameters and displayed distributed podosomes[199]. Loss of podosome ring structures explained reduced cell adhe-sion. Knockdown of the orphan nuclear receptor ERRα in RAW264.7cells [200] generated similar effects as IRAG. Expression of osteopon-tin and the β3 integrin subunit is down-regulated after silencingERRα. Transfected cells detach from the substrate easily and no podo-some belts are observed. Though cell adhesion and migration is im-paired, their differentiation and precursor cell proliferation are notinfluenced by ERRα knockdown.
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
Given the essential role of actin assembly in osteoclastic bone re-sorption, the motor protein family of myosins responsible for actin-based motility was examined in osteoclasts [201,202]. Myosin X(Myo10) maintains low expression levels in most vertebrate tissues[203], binding actin and microtubules [204]. Assembly of podosomesand sealing zones in osteoclasts depends on a complete intact micro-tubule system [205,206]. RNAi-mediated inhibition of Myo10 in bothRAW264.7 and mouse bone marrow-derived osteoclasts reduced thesizes of sealing zones and cell perimeters, but did not influence cellfusion [202].
Brain-type creatine kinase (Ckb) exhibits high up-regulation uponosteoclast maturation as witnessed with proteomic technology [207].Increased expression occurs in both mice bone marrow cells andRAW264.7 cells exposed to RANKL and M-CSF. Ckb regulates ATP dis-tribution and supply in cells, required in osteoclast actin formationand maintenance [208,209]. Down-regulation of Ckb in cell culturesproduced prominent reduction in areas of resorption pits on dentinslices [207]. Reduced resorption ability was suggested to be partiallydue to impaired actin ring structures observed in transfected osteo-clasts. In addition, Ckb was also shown to affect V-ATPase activity[207], a major proton pump essential for osteoclastic resorption. Nor-mal osteoclasts can recover their intracellular pH under challengewith strong acid over about 400 s. However, the lack of such intercel-lular pH recovery observed when inhibiting Ckb, infers that Ckb influ-ences osteoclast bone resorption by affecting V-ATPase. In vitro cellculture data were confirmed in vivo through reduced bone surfaceerosion in ovariectomized Ckb (−/−) mice. Interestingly, no signifi-cant differences in numbers of osteoclasts were observed betweenCkb (−/−) and wild type mice [207]. Therefore, Ckb is a target for al-tering osteoclast activity, not osteoclastogenesis.
5.3. Osteoclast viability targets
In addition to targeting osteoclastogenesis and osteoclastic bone re-sorption, targeted induction of osteoclast apoptosis is also an efficientstrategy to suppress excessive bone loss. EGFR is a target for regulatingosteoclast differentiation (mentioned in Section 5.1). Inhibition of EGFRexpression was also reported to cause apoptosis of mature osteoclaststhrough a caspase-9/caspase-3-dependent pathway by inhibiting theactivation of P13K-Ake/PKB [130]. Taken together, siRNA against EGFRcannot only inhibit osteoclast differentiation, but also suppress matureosteoclast survival.
RANK siRNA can be similarly characterized, as RANKL/RANK interac-tion is responsible for osteoclast differentiation, bone resorption andmature osteoclast survival as well. Therefore, mature osteoclasts trans-fected with RANK siRNA resulted in cell death [67]. Taken together,transfection of either osteoclast precursors or mature osteoclasts withRANK siRNA can reduce osteoclast-mediated bone resorption by inhi-biting osteoclast differentiation, resorption activity and osteoclastsurvival.
Bisphosphonates, the most commonly used pharmacological ap-proach for targeting osteoporosis currently, have several side effectswhich have clouded their therapeutic efficacy [9,10,15]. As an alterna-tive, siRNA against alendronate's (a nitrogen-containing bisphospho-nate, N-BP) known molecular target [210], farnesyl pyrophosphatesynthase (FPPS), was investigated to inhibit osteoclasts and promoteosteoblast activity simultaneously. FPPS plays an important role inthe mevalonate pathway which produces lipids essential for cell sur-vival [211]. Additionally, N-BPs are reported to induce human osteo-blast differentiation and mineralization in culture by inhibiting themevalonate pathway [212]. siRNA targeting FPPS significantly sup-pressed osteoclast cell viability with a single transfection and signifi-cantly increased osteoblast differentiation with effects sustained5 days post-transfection in osteoblasts. However, this treatmentdoes not significantly change osteoblast proliferation and mineraliza-tion compared to controls [213]. Therefore, selective knock down of
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

9Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
FPPS expression has the potential to inhibit osteoclasts while at thesame time promoting osteoblastic activity.
Other siRNA targets described above influence osteoclast survivalas well. NHA-oc/NHA2, mentioned as a target for osteoclastogenesis,inhibits osteoclast resorption activity partially because reducedNHA-oc/NHA2 express induces apoptosis by loss of inhibition of cas-pase-9 activation [102]; significant osteoclast death can also be in-duced by OGR1 siRNA transfection (Section 5.1); siRNA againststructurally unique osteoclast-specific transmembrane PTP-oc in-duces significant apoptosis in RAW/C4 osteoclast-like precursor cells(Section 5.2). In addition, many other general cell apoptosis signalsare reported. Using siRNA to target the proteins regulating thesepathways, cell-specific delivery is essential to avoid undesired sideeffects.
5.4. Other potential osteoporosis targets
Recognition of bone by osteoclasts is regulated by cell integrin recep-tors [214]. Integrins, as calcium-dependent, heterodimeric transmem-brane protein receptors, mediate cell attachment with extracellularmatrix or with other cells. Four different integrins are actively expressedin osteoclasts [215]. Among them,ανβ3 is highly expressed in osteoclastsand exhibits an important role in facilitating osteoclast attachment tobone and subsequent bone resorptive processes [216]. Osteoclast-medi-ated bone resorption was shown to be significantly inhibited by anti-ανβ3 antibody treatment in vitro [217] and increased skeletal mass, ab-sence of actin rings and abnormal osteoclast ruffled membranes wereall observed in osteoclasts in ανβ3-deficient mice [216]. Integrin ανβ3recognizes the extracellular RGD peptide sequence and therefore RGDpeptide has been used to label and target ανβ3-positive cells in vivo[218]. In addition, inhibition of ανβ3 using a soluble RGD mimic as a re-ceptor antagonist reduced osteoclastic bone resorption both in vitro andin vivo, suggesting that ανβ3 blockade prevents rapid bone loss causedby estrogen withdrawal [219]. In a study seeking to identify human pe-ripheral blood monocyte subsets differentiating into osteoclasts, integ-rin-β3 subunit siRNA was transfected into human CD16+ monocytes[220]. This transfection significantly reduced TRAP-positive multinu-cleated cells in a dose-dependent manner. Silencing the β3 subunit inmyeloma cells from human patients suppresses bone resorption activity(i.e., osteoclast-like activity) of these cells in vitro [221]. These data reflectthe importance of β3 integrin subunit in RANKL-induced osteoclastogen-esis. Therefore, ανβ3 could be an attractive target to attenuate or modu-late osteoclastic bone resorption.
The soluble cell signaling protein, p53, is well known to protectcells from malignant tumor transformation by coordinating severalintracellular signaling networks [222]. The ubiquitin ligase, murinedouble minute 2 (MDM2), regulates p53 activation by controllingp53 half-life [223]. Recently, a small molecule inhibitor, Nutlin, isreported to competitively bind MDM2 to interrupt p53-MDM2 inter-action and activate the p53 pathway [224,225]. In order to investigatethe effect of Nutlin-3 on pre-osteoclastic precursor cell survival, pro-liferation and differentiation, human peripheral blood mononuclearcell pre-osteoclasts were transfected with p53 siRNA [226]. Resultsshowed that Nutlin-3 anti-osteoclastogenesis activity was greatlycompromised by p53 siRNA, indicating that the inhibitory effects ofNutlin-3 on osteoclast precursors require the p53 activation pathway.Using siRNA technology to interrupt p53-MDM2 interactions maytherefore have therapeutic implications for controlling excessive os-teoclastic activity.
OGR1 was mentioned (Section 5.1) as a possible target for control-ling osteoclastogenesis. This seven-pass transmembrane proteinbinds protons and exhibits pH-sensing activity [109]. Given that in-creased extracellular proton concentrations led to nuclear transloca-tion of NFATc1, a downstream mediator of RANKL stimulation inosteoclasts [227], OGR1's role, while still unclear, could be linked tobone pH homeostasis working as a proton sensor in response to
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
external acidosis [109]. Therefore, further studies investigatingOGR1 function in bone resorption sites are necessary to validateOGR1 as a target for osteoclast-mediated excessive bone resorption.
FPPS has been used as an siRNA target to inhibit osteoclast viability,mimicking the pharmacology of N-BPs (Section 5.3). However, depletionof intracellular geranylgeranyl pyrophosphate (GGPP) has also beenreported to produce analogous phenotypic effects of bisphosphonatetreatment on both osteoclasts and osteoblasts [228,229]. Therefore,siRNA inhibition of GGPP synthase may induce apoptosis in osteoclastswhile simultaneously promoting osteoblastic activities. Using siRNAagainst GGPP synthase could also potentially suppress excessive boneloss by stimulating osteoblastic bone production.
As osteoclasts form from cell fusion, cell-to-cell contact is critical forosteoclastogenesis. Cell adhesion molecules such as intercellular adhe-sionmolecule-1 (ICAM-1), whose expression is regulated by intracellularsignaling through NF-κB and JNK, are involved in cell–cell contact be-tween osteoclast precursors and osteoblasts/stromal cells, or amongoste-oclast precursors [230,231]. ICAM-1 binds to integrin pairs, LFA-1(CD11a/CD18), blocking interactions between ICAM-1 and LFA-1, inhibit-ing osteoclast formation [230]. Hidetaka et al. reported that siRNA knockdownof LFA-1 inRAW264.7 cells hadno effects on osteoclastogenesis. Al-ternatively, ICAM-1 could be an siRNA target to inhibit osteoclasts asICAM-1 is not only expressed on osteoclast precursors andmature osteo-clasts, but also on osteoblasts [232,233]. Therefore, cell-specific targeteddelivery will be essential for the specific bioactivity and siRNA selectivityto osteoclasts as desired.
6. siRNA delivery to bone
6.1. Challenges and strategies
RNAi as a novel therapeutic approach has considerable potential tosilence abnormal genes, especially for gene targets not effectively tar-geted by conventional therapeutics (i.e., by small molecules, proteinsand antibodies). Themajor obstacle for developing siRNA for therapeu-tics is also its targeted delivery with clinically acceptable formulationsand reliable routes of administration. This is particularly true of siRNAdelivery to bone with its intrinsically poor drug penetration and vascu-lar perfusion. Bone is primarily composed of three cell types: osteo-clasts, osteoblasts, and osteocytes. Bone's unique extracellular matrixis strongly mineralized with calcium phosphate, forming ~70 wt.% apa-tite, a major reservoir of the body's calciummost actively involved withphysiological calcium and phosphorus homeostasis. Increasingmolecu-lar and cellular understanding of bone biology has produced continuousreports of new potential therapeutic targets for various bone patholo-gies. Nonetheless, despite new targets identified as osteoclast-specific,targeting methods are not yet reliably tissue-based specifically withrequisite bone-specificity necessary for these therapeutic agents. Seri-ous complications from cross-reactivity with other non-target tissuesor poor efficacy due to lowdrug target tissue concentration require a re-liable targeting strategy. Many attempts to improve the targeting ofdrugs to bone mineral fraction (apatite) include drug conjugationwith bone-targeting ligands, such as tetracycline [234], estradiol, andbisphosphonate [235,236]. Bisphosphonates, especially N-BPs, as anti-resorptive drugs having intrinsic high affinity to the bone apatite sur-face, are very attractive for delivery of conjugated non-specific bonetherapeutic agents. However, their exploitation as bone-targetingagents must consider that free bisphosphonate itself also will increaseanti-resorptive activity.
SiRNAs are well known for their very short circulating half-lives invivo [237]. Unmodified naked siRNA cannot be directly administered di-rectly in vivo, exhibiting only a 6 min half-life after systemic administra-tion to rats due to degradation by serum nucleases [237]. Many studiesseek to overcome the challenges of siRNA delivery by the chemical mod-ification of siRNAnucleotides, lipid/liposomal/polymer-based complexes,and collagen (atelocollagen)-based [238,239] formulations for siRNA, as
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

10 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
well as viral vectors and polymer-complexed siRNA carriers [240]. Thesemethods eachhave their advantages anddisadvantages, but non-viral de-livery is considered amore clinically relevant deliverymechanism, avoid-ing the known problems of current viral vector delivery: high viraltoxicity, possible carcinogenicity, proven immunogenicity, and significantcost limitations [31]. Delivery of siRNA targeted specifically to bone is notan entirely new story. To date, in vivo siRNA studies in humans have fo-cused on rheumatoid arthritis (RA) by local delivery (intra-articular in-jection [241,242], electroporated siRNA [243], topical cream [34]), or bysystemic delivery, each with modest effects [244]. Delivery of siRNA tobone osteoclasts in vivo, either targeted to or addressing, is not yetreported.
To improve siRNA stability in vivo, various chemical modificationshave been investigated. SiRNA stability against nuclease degradationcan be improved by introducing a phosphorothioate (P_S) backbonelinkage or modifying the ribose 2′-hydroxyl position as 2′-OMe or 2′-F,as well as a 4′-thioribose modification [245]. All show significant im-provements over unmodified siRNA [246–248]. Combination of 2′- and4′-thioribose modification results in substantially improved siRNA bioac-tivity and plasma stability [249]. This improved siRNA plasma stability fa-cilitates more reliable systemic dosing and versatile delivery options. Inaddition, specific siRNA chemical modifications can potentially be usedto improve conjugation and targeting aswell. Bone-targeting ligands, an-tibodies [250] or small molecule drugs (vida supra) can be conjugatedwith siRNA molecules with various chemistries for systemic delivery ofsiRNA-based bone disease therapeutics. For example, bisphosphonates,can be conjugated to ribose hydroxyl groups in siRNA sugars. Modifica-tion of the ribose 2′-OH at the 3′ end of the guide siRNA strand appearseffective for improving siRNA activity [240]. In this case, bisphosphonateperforms both as a drug and also as a targeting moiety to deliver siRNAspecifically to bone.
Using tissue-specific targeting methods, both locally and systemi-cally delivered siRNA can be more reliably dosed by judicious combi-nation of other “homing” molecules. The concept of siRNA targetingto improve both systemic and local efficacy is shown in Fig. 4. Estab-lished drug delivery strategies can be utilized for both targeted sys-temic and local siRNA delivery, including siRNA conjugation to cellmembrane-targeting ligands, antibodies, water-soluble polymers[251], or cell-penetrating peptides. These methods seek to improveamount of siRNA delivered specifically to targeted bone cells, and tolimit drug off-target effects in cells other than osteoclasts.
Fig. 4. The concept of therapeutic siRNA targeting delivery to
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
RANK expressed on osteoclast precursors as well as on mature oste-oclasts serves as a popular molecular target for osteoporosis therapies.Antibodies against RANK have been tested for effective osteoclast target-ing by conjugationwith calcitonin, an anti-resorptive protein drug [252].The conjugate showed efficacy inhibiting osteoclast formation and boneresorption. The strategy, using receptor ligand binding peptides as thetargeting moiety, provides a general approach to generating an osteo-clast-targeting platform for specific drug delivery to osteoclasts. Otherosteoclast-specific proteins can also be utilized for this purpose, suchas cathepsin K and NHA-oc/NHA2. In addition, many calcium-bindingproteins, recently reviewed [253], including acidic proline-rich salivaryproteins, osteocalcin, sialoprotein, and osteopontin, have the potentialas novel bone-targeting moieties [253]. The hexapeptide (Asp)6 exhib-ited high affinity to bone and was used as bone-targeting moiety conju-gated with estradiol in ovariectomizedmice. The conjugates maintainedbone density similarly to unconjugated estradiol, but without displayingincreased liver and uterus weight [254,255].
In addition to specific chemical modifications, protection of siRNAin plasma is also provided by formulating siRNA into nano/macro-sized particles for delivery, as recently reviewed [256–258]. Mostdrug delivery strategies to date primarily target the liver, either delib-erately or non-discriminately, a natural result of first-pass metabo-lism and highly efficient liver-based drug and colloidal clearance.This is an advantage for liver targeted therapies: for example, hydro-dynamic tail-vein injection of Fas siRNA successfully protected micefrom liver failure and fibrosis [259]. However, the liver and other re-ticuloendothelial (MPS/RES) system tissues efficiently filter mostdrug carrier particles (i.e., N90%) despite active targeting to other tar-get tissue sites. To avoid this massive dose removal, locally deliveredsiRNA-particle formulations can be used in bone-targeted therapeu-tics. Specifically to target osteoclast or osteoclast precursors, siRNAdelivery can exploit drug carrier particle size-selective cell-specificuptake to numerous phagocyte target cells in bone tissue using localdelivery. In the context of osteoporosis therapy, targeted cells arepre-osteoclastic monocytes and osteoclasts— all naturally phagocyticcells. Phagocytes efficiently take up particles with diameters up to10 μm [260]. However, particles with diameters greater than300 nm are too large to penetrate non-phagocytic cell membranes(i.e., osteoblasts, fibroblasts) either passively or by endocytosis or pi-nocytosis [261]. Therefore, siRNA-loaded microparticles with con-trolled sizes could be exploited for selective internalization by
improves siRNA bioavailability and therapeutic efficacy.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

11Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
osteoclastic phagocytic mechanisms and to target siRNA to these cellsin new osteoporotic therapeutic strategies without a conjugated tar-geting moiety. These particles could be delivered locally to bonesites in clinical cases, especially in bone augmentation and implantsurgeries. Additionally, for some bone molecular drug targets appro-priate both for osteoclasts and osteoblasts, siRNA-containing nano-particles capable of being taken up by both cell types might beeffective for increasing bone mass by inhibiting osteoclasts while si-multaneously stimulating osteoblasts. Particle opsonisation by hostplasma proteins, commonly a problem for drug carriers, may con-found target cell phagocytosis since opsonisation (particularly, non-specific adsorption of immunoglobulins and complement onto theparticles) may activate macrophages and T-cells and cause inflamma-tion in surrounding tissues and damage to healthy cells [262].
Currently, local siRNA administration ismore efficient than its system-ic delivery for short-term therapeutic purposes [263]. Nonetheless, localdelivery must also consider target specificity. For example, intra-articularinjection of siRNA for RA is not an effective route of administration fortransfecting osteoclasts located near or on bone surfaces atmost commonosteoporotic bone sites, notably pelvis, vertebrae andwrist [264]. Tomax-imize drug availability to specific bone sites and obtain a reliable, repro-ducible and predictable pharmacokinetic profile, bioactive materials,including porous chitosan/collagen scaffolds [265] and injectable FDA-ap-proved calcium phosphate bone cements (CPC) are being developed asdrug carriers for local bone delivery. Polymethylmethacrylate (PMMA)bone cements used clinically for implant fixation, kyphoplasty [266],and bone augmentation [267], exhibit very poor control over entrappeddose release kinetics, often leaving up to 40% of the drug trapped inPMMA [268,269]. Injectable CPC exhibits several unique properties forbone drug delivery, including peri-operative preparation and drug load-ing, liquid–solid setting in situ, intrinsic osteoconductivity (degradableby osteoclasts), osteoinductivity (infiltrated by osteoblasts and replacedwith new bone), and acceptable biocompatibility [270–272]. CPC's self-setting ability at ambient or body temperature within the bone cavity[273] enables injectable formulation, largely expanding its therapeuticutility [274,275]. To date, a large body of evidence supports the feasibilityand value of using CPC as a local drug carrier in bone augmentation. Phar-macologically active molecules are readily dispersed or distributedthroughout CPC prior to setting and surgical placement, providing a sus-tainable and controlled drug release medium. Several conventionalsmall molecule and protein-based drugs have been delivered using CPCin bone, including antibiotics to decrease post-surgical infections [276–279], anticancer drugs to reduce tumorogenesis [280,281] and growthfactors to promote bone healing [282,283]. Therefore, despite its limitedmechanical properties, CPC could also be used to deliver siRNA to boneas a local delivery matrix.
A degradable cationic hydrogel comprising of gelatin and chitosanhas been used for local delivery of an antisense oligonucleotide target-ing murine TNF-α to suppress endotoxin-induced osteolysis in mousecalvaria [284]. Cationic polymeric chitosan complexes with anionic nu-cleotides and also facilitates cell transfection. Osteoclast numbers andbone resorption were significantly suppressed 4 weeks post-implanta-tion. The same idea can also be extended to siRNA local delivery to bone.
The intrinsically low pH in osteoclast resorption lacuna and in thecell lysosome is attractive as a basis for targeting drugs conjugated tocarriers containing acid-cleavable hydrazone bonds [285]. Additional-ly, various acid-sensitive, cathepsin K-specific, and MMPs-specificpeptide linkages (reviewed recently [253]) also provide disease-and environmentally specific bone-targeting strategies appropriatefor targeted siRNA bone delivery.
6.2. Non-specific siRNA-cell bioactivities and off-target side effects
The first clinical trials involving siRNA were applied by local intraoc-ular injection in patients with choroidal neovascularization (CNV)[33,286]. CNV is an age-related eye disorder characterized by the
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
choroidal vessel invasion to the retina beneath retinal pigmented epithe-lium. Naked siRNA targeting VEGFA or VEGFR1 showed significant sup-pression in a laser-injury-induced CNV mice model [33,286]. However,in 2008, new data showed that this therapeutic suppression is a generalsiRNA-class effect, independent of siRNA sequence [25]. The observedtherapy was based on interactions between cell-surface (but not endo-somal) toll-like receptor 3 (TLR3), a cell membrane double-strandedviral RNA sensor found on numerous cell types [287], and siRNA, regard-less of RNA sequences, target, and internalization. However, to bindTLR3, siRNA of 21 nucleotides or longer was required to form a 2:1TLR3-RNA conjugate. TLR3 plays an indispensable role in the TRIF-NF-κB cell signaling cascade where TRIF is the TLR3 adaptor protein[288]. This important discovery has revealed generic properties of siR-NAs on vascular tissues, and improved the global understanding ofsiRNA general bioactivities, both desired and potentially undesired,through non-specific signaling pathways.
In this regard, primary human choroidal endothelial cells showeddecreased survival in serum-containing media cultured with serum-stable 2′O-methyl-Lus-siRNA (without transfection reagents) [25].Such extracellular siRNA-induced cytotoxicity might affect other or-gans, including bone. Osteoclast precursors but not mature osteo-clasts express TLR3 [59]. Poly(I:C)dsRNA, which acts as a TLR3ligand to stimulate cell surface TLR3, produced strong inhibition of os-teoclast differentiation in both mouse bone marrow cultures andhuman peripheral blood monocytes at 1 μg/ml in a dose-dependentmanner under induction by M-CSF and RANKL. TLR stimulation in-duces the production of various proinflammatory cytokines such asTNF-α [289] and prompts high levels of phagocytic activity in osteo-clast precursors [59], indicating an enhanced immune response afterTLR stimulation in osteoclast precursors. Taken together, both the un-anticipated vascular and immune-general siRNA-class effects shouldbe carefully considered for siRNA therapies, especially when deliv-ered systemically.
7. Conclusions
Gene silencing using siRNA has many potential therapeutic applica-tions due to several advantages intrinsic to RNAi, such as its high targetspecificity and intrinsic biological response [28]. Effective siRNA deliv-ery and selective targeting to desired tissue sites remain problematic.siRNA delivery to bone has developed relatively slowly compared toits delivery to other tissues. In this regard, osteoporosis is an increasing-ly challenging problem globally with a need for improved therapeutics.Many new cellular targets in osteoclasts are continually reported andattractive for siRNA knock down. Efficient siRNA delivery methodsand selective targeting to bone connective tissue and osteoclasts mustbe developed, improved and optimized. siRNA's efficacy, bioavailability,and therapeutic duration in the context of osteoclast-mediated exces-sive bone resorption are largely unexplored, but this strategy has attrac-tive options for development of new therapeutics.
Acknowledgments
The authors acknowledge support from NIH grants EB000894 andEB011879.
References
[1] C. Deal, J. Gideon, Recombinant human PTH 1–34 (Forteo): an anabolic drug forosteoporosis, Cleve. Clin. J. Med. 70 (2003) 585–586 (589–590, 592–584passim).
[2] G.A. Rodan, T.J. Martin, Therapeutic approaches to bone diseases, Science 289(2000) 1508–1514.
[3] J.E. Rossouw, G.L. Anderson, R.L. Prentice, et al., Risks and benefits of estrogenplus progestin in healthy postmenopausal women: principal results from thewomen's health initiative randomized controlled trial, JAMA 288 (2002)321–333.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

12 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
[4] I. Lambrinoudaki, G. Christodoulakos, D. Botsis, Bisphosphonates, Ann. N. Y.Acad. Sci. 1092 (2006) 397–402.
[5] B.W. Walsh, L.H. Kuller, R.A. Wild, S. Paul, M. Farmer, J.B. Lawrence, A.S. Shah,P.W. Anderson, Effects of raloxifene on serum lipids and coagulation factors inhealthy postmenopausal women, JAMA 279 (1998) 1445–1451.
[6] Drugs for prevention and treatment of postmenopausal osteoporosis, Treat.Guidel. Med. Lett. 3 (2005) 69–74.
[7] S. Wada, N. Udagawa, N. Nagata, T.J. Martin, D.M. Findlay, Physiological levels ofcalcitonin regulate the mouse osteoclast calcitonin receptor by a protein kinasealpha-mediated mechanism, Endocrinology 137 (1996) 312–320.
[8] T.B. Dodson, W.C. Guralnick, R.B. Donoff, L.B. Kaban, Massachusetts General Hos-pital/Harvard Medical School MD oral and maxillofacial surgery program: a 30-year review, J. Oral Maxillofac. Surg. 62 (2004) 62–65.
[9] R.E. Marx, Pamidronate (Aredia) and zoledronate (Zometa) induced avascularnecrosis of the jaws: a growing epidemic, J. Oral Maxillofac. Surg. 61 (2003)1115–1117.
[10] S.L. Ruggiero, B. Mehrotra, T.J. Rosenberg, S.L. Engroff, Osteonecrosis of the jawsassociated with the use of bisphosphonates: a review of 63 cases, J. Oral Maxil-lofac. Surg. 62 (2004) 527–534.
[11] S.A. Khan, J.A. Kanis, S. Vasikaran, W.F. Kline, B.K. Matuszewski, E.V. McCloskey,M.N. Beneton, B.J. Gertz, D.G. Sciberras, S.D. Holland, J. Orgee, G.M. Coombes, S.R.Rogers, A.G. Porras, Elimination and biochemical responses to intravenous alen-dronate in postmenopausal osteoporosis, J. Bone Miner. Res. 12 (1997)1700–1707.
[12] K.H. Yang, J.H. Won, H.K. Yoon, J.H. Ryu, K.S. Choo, J.S. Kim, High concentrationsof pamidronate in bone weaken the mechanical properties of intact femora in arat model, Yonsei Med. J. 48 (2007) 653–658.
[13] A. Sebba, Osteoporosis: how long should we treat? Curr. Opin. Endocrinol. Dia-betes Obes. 15 (2008) 502–507.
[14] D.E. Sellmeyer, Atypical fractures as a potential complication of long-termbisphosphonate therapy, JAMA 304 (2010) 1480–1484.
[15] C.V. Odvina, J.E. Zerwekh, D.S. Rao, N. Maalouf, F.A. Gottschalk, C.Y. Pak, Severelysuppressed bone turnover: a potential complication of alendronate therapy,J. Clin. Endocrinol. Metab. 90 (2005) 1294–1301.
[16] D.K. Wysowski, Reports of esophageal cancer with oral bisphosphonate use, N.Engl. J. Med. 360 (2009) 89–90.
[17] S.M. Ott, Long-term safety of bisphosphonates, J. Clin. Endocrinol. Metab. 90(2005) 1897–1899.
[18] P.K. Venesmaa, H.P. Kroger, H.J. Miettinen, J.S. Jurvelin, O.T. Suomalainen, E.M.Alhav, Alendronate reduces periprosthetic bone loss after uncemented primarytotal hip arthroplasty: a prospective randomized study, J. Bone Miner. Res. 16(2001) 2126–2131.
[19] A. Nehme, G. Maalouf, J.L. Tricoire, G. Giordano, P. Chiron, J. Puget, Effect of alen-dronate on periprosthetic bone loss after cemented primary total hip arthro-plasty: a prospective randomized study, Rev. Chir. Orthop. Reparatrice Appar.Mot. 89 (2003) 593–598.
[20] P.K. Venesmaa, H.P. Kroger, H.J. Miettinen, J.S. Jurvelin, O.T. Suomalainen, E.M.Alhava, Monitoring of periprosthetic BMD after uncemented total hip arthro-plasty with dual-energy X-ray absorptiometry—a 3-year follow-up study, J.Bone Miner. Res. 16 (2001) 1056–1061.
[21] J.P. Brown, R.L. Prince, C. Deal, R.R. Recker, D.P. Kiel, L.H. de Gregorio, P. Hadji,L.C. Hofbauer, J.M. Alvaro-Gracia, H. Wang, M. Austin, R.B. Wagman, R. New-mark, C. Libanati, J. San Martin, H.G. Bone, Comparison of the effect of denosu-mab and alendronate on BMD and biochemical markers of bone turnover inpostmenopausal women with low bone mass: a randomized, blinded, phase 3trial, J. Bone Miner. Res. 24 (2009) 153–161.
[22] D.L. Kendler, C. Roux, C.L. Benhamou, J.P. Brown, M. Lillestol, S. Siddhanti, H.S.Man, J. San Martin, H.G. Bone, Effects of denosumab on bone mineral densityand bone turnover in postmenopausal women transitioning from alendronatetherapy, J. Bone Miner. Res. 25 (2010) 72–81.
[23] A. Fire, S. Xu, M.K. Montgomery, S.A. Kostas, S.E. Driver, C.C. Mello, Potent andspecific genetic interference by double-stranded RNA in Caenorhabditis elegans,Nature 391 (1998) 806–811.
[24] L. Aagaard, J.J. Rossi, RNAi therapeutics: principles, prospects and challenges,Adv. Drug Deliv. Rev. 59 (2007) 75–86.
[25] M.E. Kleinman, K. Yamada, A. Takeda, V. Chandrasekaran, M. Nozaki, J.Z. Baffi, R.J.Albuquerque, S. Yamasaki, M. Itaya, Y. Pan, B. Appukuttan, D. Gibbs, Z. Yang, K.Kariko, B.K. Ambati, T.A. Wilgus, L.A. DiPietro, E. Sakurai, K. Zhang, J.R. Smith,E.W. Taylor, J. Ambati, Sequence- and target-independent angiogenesis suppres-sion by siRNA via TLR3, Nature 452 (2008) 591–597.
[26] S.M. Hammond, E. Bernstein, D. Beach, G.J. Hannon, An RNA-directed nucleasemediates post-transcriptional gene silencing in Drosophila cells, Nature 404(2000) 293–296.
[27] M.A. Beer, S. Tavazoie, Predicting gene expression from sequence, Cell 117(2004) 185–198.
[28] C.D. Novina, P.A. Sharp, The RNAi revolution, Nature 430 (2004) 161–164.[29] T.V. Achenbach, B. Brunner, K. Heermeier, Oligonucleotide-based knockdown
technologies: antisense versus RNA interference, Chembiochem 4 (2003)928–935.
[30] L.D. Kumar, A.R. Clarke, Gene manipulation through the use of small interfering RNA(siRNA): from in vitro to in vivo applications, Adv. Drug Deliv. Rev. 59 (2007) 87–100.
[31] Q.Z. Wang, Y.H. Lv, Y. Diao, R. Xu, The design of vectors for RNAi delivery system,Curr. Pharm. Des. 14 (2008) 1327–1340.
[32] K.A. Woodrow, Y. Cu, C.J. Booth, J.K. Saucier-Sawyer, M.J. Wood, W.M. Saltzman,Intravaginal gene silencing using biodegradable polymer nanoparticles denselyloaded with small-interfering RNA, Nat. Mater. 8 (2009) 526–533.
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
[33] S.J. Reich, J. Fosnot, A. Kuroki, W. Tang, X. Yang, A.M. Maguire, J. Bennett, M.J.Tolentino, Small interfering RNA (siRNA) targeting VEGF effectively inhibits oc-ular neovascularization in a mouse model, Mol. Vis. 9 (2003) 210–216.
[34] M. Takanashi, K. Oikawa, K. Sudo, M. Tanaka, K. Fujita, A. Ishikawa, S. Nakae, R.L.Kaspar, M. Matsuzaki, M. Kudo, M. Kuroda, Therapeutic silencing of an endoge-nous gene by siRNA cream in an arthritis model mouse, Gene Ther. 16 (2009)982–989.
[35] D. Massaro, G.D. Massaro, L.B. Clerch, Noninvasive delivery of small inhibitoryRNA and other reagents to pulmonary alveoli in mice, Am. J. Physiol. Lung Cell.Mol. Physiol. 287 (2004) L1066–L1070.
[36] D.R. Thakker, F. Natt, D. Husken, R. Maier, M. Muller, H. van der Putten, D. Hoyer,J.F. Cryan, Neurochemical and behavioral consequences of widespread geneknockdown in the adult mouse brain by using nonviral RNA interference, Proc.Natl. Acad. Sci. U.S.A. 101 (2004) 17270–17275.
[37] P. Wu, D.W. Grainger, Drug/device combinations for local drug therapies and in-fection prophylaxis, Biomaterials 27 (2006) 2450–2467.
[38] C. Niyibizi, P. Smith, Z. Mi, C.L. Phillips, P. Robbins, Transfer of proalpha2(I) cDNAinto cells of a murine model of human osteogenesis imperfecta restores synthe-sis of type I collagen comprised of alpha1(I) and alpha2(I) heterotrimers in vitroand in vivo, J. Cell. Biochem. 83 (2001) 84–91.
[39] J.A. Barranger, E.O. Rice, J. Dunigan, C. Sansieri, N. Takiyama, M. Beeler, J. Lancia,S. Lucot, S. Scheirer-Fochler, T. Mohney, W. Swaney, A. Bahnson, E. Ball, Gau-cher's disease: studies of gene transfer to haematopoietic cells, Baillieres Clin.Haematol. 10 (1997) 765–778.
[40] C.H. Evans, S.C. Ghivizzani, J.H. Herndon, M.C. Wasko, J. Reinecke, P. Wehling,P.D. Robbins, Clinical trials in the gene therapy of arthritis, Clin. Orthop. Relat.Res. (2000) S300–S307.
[41] C.H. Evans, P.D. Robbins, S.C. Ghivizzani, J.H. Herndon, R. Kang, A.B. Bahnson, J.A.Barranger, E.M. Elders, S. Gay, M.M. Tomaino, M.C.Wasko, S.C.Watkins, T.L.White-side, J.C. Glorioso, M.T. Lotze, T.M.Wright, Clinical trial to assess the safety, feasibil-ity, and efficacy of transferring a potentially anti-arthritic cytokine gene to humanjoints with rheumatoid arthritis, Hum. Gene Ther. 7 (1996) 1261–1280.
[42] C.H. Evans, Gene therapies for osteoarthritis, Curr. Rheumatol. Rep. 6 (2004)31–40.
[43] A.W. Baltzer, J.D. Whalen, P. Wooley, C. Latterman, L.M. Truchan, P.D. Robbins,C.H. Evans, Gene therapy for osteoporosis: evaluation in a murine ovariectomymodel, Gene Ther. 8 (2001) 1770–1776.
[44] B. Bolon, C. Carter, M. Daris, S. Morony, C. Capparelli, A. Hsieh, M. Mao, P. Koste-nuik, C.R. Dunstan, D.L. Lacey, J.Z. Sheng, Adenoviral delivery of osteoprotegerinameliorates bone resorption in a mouse ovariectomy model of osteoporosis,Mol. Ther. 3 (2001) 197–205.
[45] P.J. Kostenuik, B. Bolon, S. Morony, M. Daris, Z. Geng, C. Carter, J. Sheng, Genetherapy with human recombinant osteoprotegerin reverses established osteo-penia in ovariectomized mice, Bone 34 (2004) 656–664.
[46] J.R. Lieberman, A. Daluiski, S. Stevenson, L. Wu, P. McAllister, Y.P. Lee, J.M. Kabo,G.A. Finerman, A.J. Berk, O.N. Witte, The effect of regional gene therapywith bone morphogenetic protein-2-producing bone-marrow cells on therepair of segmental femoral defects in rats, J. Bone Joint Surg. Am. 81 (1999)905–917.
[47] A.W. Baltzer, C. Lattermann, J.D. Whalen, P. Wooley, K. Weiss, M. Grimm, S.C.Ghivizzani, P.D. Robbins, C.H. Evans, Genetic enhancement of fracture repair:healing of an experimental segmental defect by adenoviral transfer of theBMP-2 gene, Gene Ther. 7 (2000) 734–739.
[48] O. Betz, M. Vrahas, A. Baltzer, J.R. Lieberman, P.D. Robbins, C.H. Evans, Genetransfer approaches to enhancing bone healing, in: J.R. Lieberman, G.E.Friedlaender (Eds.), Bone regeneration and repair: Biology and clinical Applica-tions, Humana Press, Totowa, NJ, 2005, pp. 157–168.
[49] C.H. Evans, S.C. Ghivizzani, J.H. Herndon, P.D. Robbins, Gene therapy for the treatmentof musculoskeletal diseases, J. Am. Acad. Orthop. Surg. 13 (2005) 230–242.
[50] G. Courties, J. Presumey, I. Duroux-Richard, C. Jorgensen, F. Apparailly, RNA in-terference-based gene therapy for successful treatment of rheumatoid arthritis,Expert Opin. Biol. Ther. 9 (2009) 535–538.
[51] S.S.Y. Anil, K. Jain, Bone health— an investment, Indian J. Orthop. 43 (2009) 223–224.[52] K. Vaananen, Mechanism of osteoclast mediated bone resorption—rationale for
the design of new therapeutics, Adv. Drug Deliv. Rev. 57 (2005) 959–971.[53] H. Yasuda, N. Shima, N. Nakagawa, K. Yamaguchi, M. Kinosaki, S. Mochizuki, A.
Tomoyasu, K. Yano, M. Goto, A. Murakami, E. Tsuda, T. Morinaga, K. Higashio,N. Udagawa, N. Takahashi, T. Suda, Osteoclast differentiation factor is a ligandfor osteoprotegerin/osteoclastogenesis—inhibitory factor and is identical toTRANCE/RANKL, Proc. Natl. Acad. Sci. U.S.A. 95 (1998) 3597–3602.
[54] D.L. Lacey, E. Timms, H.L. Tan, M.J. Kelley, C.R. Dunstan, T. Burgess, R. Elliott, A.Colombero, G. Elliott, S. Scully, H. Hsu, J. Sullivan, N. Hawkins, E. Davy, C. Cappar-elli, A. Eli, Y.X. Qian, S. Kaufman, I. Sarosi, V. Shalhoub, G. Senaldi, J. Guo, J. Dela-ney, W.J. Boyle, Osteoprotegerin ligand is a cytokine that regulates osteoclastdifferentiation and activation, Cell 93 (1998) 165–176.
[55] N. Takahashi, H. Yamana, S. Yoshiki, G.D. Roodman, G.R. Mundy, S.J. Jones, A.Boyde, T. Suda, Osteoclast-like cell formation and its regulation by osteotropichormones in mouse bone marrow cultures, Endocrinology 122 (1988)1373–1382.
[56] N. Amizuka, N. Takahashi, N. Udagawa, T. Suda, H. Ozawa, An ultrastructuralstudy of cell–cell contact between mouse spleen cells and calvaria-derived oste-oblastic cells in a co-culture system for osteoclast formation, Acta Histochem.Cytochem. 30 (1997) 351–362.
[57] V. Nicolin, G. Baldini, R. Bareggi, M. Zweyer, G. Zauli, M. Vaccarezza, P. Narducci,Morphological features of osteoclasts derived from a co-culture system, J. Mol.Histol. 37 (2006) 171–177.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

13Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
[58] L.M. Chamberlain, M.L. Godek, M. Gonzalez-Juarrero, D.W. Grainger, Phenotypicnon-equivalence of murine (monocyte-) macrophage cells in biomaterial andinflammatory models, J. Biomed. Mater. Res. A 88 (2009) 858–871.
[59] M. Takami, N. Kim, J. Rho, Y. Choi, Stimulation by toll-like receptors inhibits os-teoclast differentiation, J. Immunol. 169 (2002) 1516–1523.
[60] Y. Wang, D. Lebowitz, C. Sun, H. Thang, M.D. Grynpas, M. Glogauer, Identifyingthe relative contributions of Rac1 and Rac2 to osteoclastogenesis, J. BoneMiner. Res. 23 (2008) 260–270.
[61] D.A. Corral, M. Amling, M. Priemel, E. Loyer, S. Fuchs, P. Ducy, R. Baron, G.Karsenty, Dissociation between bone resorption and bone formation in osteope-nic transgenic mice, Proc. Natl. Acad. Sci. U.S.A. 95 (1998) 13835–13840.
[62] W.C. Dougall, M. Glaccum, K. Charrier, K. Rohrbach, K. Brasel, T. De Smedt, E.Daro, J. Smith, M.E. Tometsko, C.R. Maliszewski, A. Armstrong, V. Shen, S. Bain,D. Cosman, D. Anderson, P.J. Morrissey, J.J. Peschon, J. Schuh, RANK is essentialfor osteoclast and lymph node development, Genes Dev. 13 (1999) 2412–2424.
[63] D.L. Lacey, H.L. Tan, J. Lu, S. Kaufman, G. Van, W. Qiu, A. Rattan, S. Scully, F.Fletcher, T. Juan, M. Kelley, T.L. Burgess, W.J. Boyle, A.J. Polverino, Osteoprote-gerin ligand modulates murine osteoclast survival in vitro and in vivo, Am. J.Pathol. 157 (2000) 435–448.
[64] W.S. Simonet, D.L. Lacey, C.R. Dunstan, M. Kelley, M.S. Chang, R. Luthy, H.Q.Nguyen, S. Wooden, L. Bennett, T. Boone, G. Shimamoto, M. DeRose, R. Elliott,A. Colombero, H.L. Tan, G. Trail, J. Sullivan, E. Davy, N. Bucay, L. Renshaw-Gegg,T.M. Hughes, D. Hill, W. Pattison, P. Campbell, S. Sander, G. Van, J. Tarpley, P.Derby, R. Lee, W.J. Boyle, Osteoprotegerin: a novel secreted protein involved inthe regulation of bone density, Cell 89 (1997) 309–319.
[65] H. Yasuda, N. Shima, N. Nakagawa, S.I. Mochizuki, K. Yano, N. Fujise, Y. Sato, M.Goto, K. Yamaguchi, M. Kuriyama, T. Kanno, A. Murakami, E. Tsuda, T. Morinaga,K. Higashio, Identity of osteoclastogenesis inhibitory factor (OCIF) and osteopro-tegerin (OPG): a mechanism by which OPG/OCIF inhibits osteoclastogenesis invitro, Endocrinology 139 (1998) 1329–1337.
[66] E.F. Wagner, G. Karsenty, Genetic control of skeletal development, Curr. Opin.Genet. Dev. 11 (2001) 527–532.
[67] Y. Wang, D.W. Grainger, siRNA knock-down of RANK signaling to control osteo-clast-mediated bone resorption, Pharm. Res. 27 (2010) 1273–1284.
[68] R. Iwasaki, K. Ninomiya, K. Miyamoto, T. Suzuki, Y. Sato, H. Kawana, T.Nakagawa, T. Suda, T. Miyamoto, Cell fusion in osteoclasts plays a critical rolein controlling bone mass and osteoblastic activity, Biochem. Biophys. Res. Com-mun. 377 (2008) 899–904.
[69] S. Mundlos, F. Otto, C. Mundlos, J.B. Mulliken, A.S. Aylsworth, S. Albright, D.Lindhout, W.G. Cole, W. Henn, J.H. Knoll, M.J. Owen, R. Mertelsmann, B.U.Zabel, B.R. Olsen, Mutations involving the transcription factor CBFA1 cause clei-docranial dysplasia, Cell 89 (1997) 773–779.
[70] A. Teti, H.C. Blair, S.L. Teitelbaum, A.J. Kahn, C. Koziol, J. Konsek, A. Zambonin-Zallone, P.H. Schlesinger, Cytoplasmic pH regulation and chloride/bicarbonateexchange in avian osteoclasts, J. Clin. Invest. 83 (1989) 227–233.
[71] J.G. Emery, P. McDonnell, M.B. Burke, K.C. Deen, S. Lyn, C. Silverman, E. Dul, E.R.Appelbaum, C. Eichman, R. DiPrinzio, R.A. Dodds, I.E. James, M. Rosenberg, J.C.Lee, P.R. Young, Osteoprotegerin is a receptor for the cytotoxic ligand TRAIL, J.Biol. Chem. 273 (1998) 14363–14367.
[72] M. Gowen, F. Lazner, R. Dodds, R. Kapadia, J. Feild, M. Tavaria, I. Bertoncello, F.Drake, S. Zavarselk, I. Tellis, P. Hertzog, C. Debouck, I. Kola, Cathepsin K knockoutmice develop osteopetrosis due to a deficit in matrix degradation but not de-mineralization, J. Bone Miner. Res. 14 (1999) 1654–1663.
[73] S.A. Nesbitt, M.A. Horton, Trafficking of matrix collagens through bone-resorb-ing osteoclasts, Science 276 (1997) 266–269.
[74] B.G. Darnay, V. Haridas, J. Ni, P.A. Moore, B.B. Aggarwal, Characterization of theintracellular domain of receptor activator of NF-kappaB (RANK). Interactionwith tumor necrosis factor receptor-associated factors and activation of NF-kap-pab and c-Jun N-terminal kinase, J. Biol. Chem. 273 (1998) 20551–20555.
[75] S. Dai, T. Hirayama, S. Abbas, Y. Abu-Amer, The IkappaB kinase (IKK) inhibitor,NEMO-binding domain peptide, blocks osteoclastogenesis and bone erosion ininflammatory arthritis, J. Biol. Chem. 279 (2004) 37219–37222.
[76] V. Iotsova, J. Caamano, J. Loy, Y. Yang, A. Lewin, R. Bravo, Osteopetrosis in micelacking NF-kappaB1 and NF-kappaB2, Nat. Med. 3 (1997) 1285–1289.
[77] J.A. DiDonato, M. Hayakawa, D.M. Rothwarf, E. Zandi, M. Karin, A cytokine-re-sponsive IkappaB kinase that activates the transcription factor NF-kappaB, Na-ture 388 (1997) 548–554.
[78] F. Mercurio, H. Zhu, B.W. Murray, A. Shevchenko, B.L. Bennett, J. Li, D.B. Young,M. Barbosa, M. Mann, A. Manning, A. Rao, IKK-1 and IKK-2: cytokine-activatedIkappaB kinases essential for NF-kappaB activation, Science 278 (1997)860–866.
[79] M.G. Ruocco, S. Maeda, J.M. Park, T. Lawrence, L.C. Hsu, Y. Cao, G. Schett, E.F.Wagner, M. Karin, I{kappa}B kinase (IKK){beta}, but not IKK{alpha}, is a criticalmediator of osteoclast survival and is required for inflammation-induced boneloss, J. Exp. Med. 201 (2005) 1677–1687.
[80] J.E. Otero, S. Dai, D. Foglia, M. Alhawagri, J. Vacher, M. Pasparakis, Y. Abu-Amer,Defective osteoclastogenesis by IKKbeta-null precursors is a result of receptoractivator of NF-kappaB ligand (RANKL)-induced JNK-dependent apoptosis andimpaired differentiation, J. Biol. Chem. 283 (2008) 24546–24553.
[81] M.L. Chaisson, D.G. Branstetter, J.M. Derry, A.P. Armstrong, M.E. Tometsko, K.Takeda, S. Akira, W.C. Dougall, Osteoclast differentiation is impaired in the ab-sence of inhibitor of kappa B kinase alpha, J. Biol. Chem. 279 (2004)54841–54848.
[82] I. Darwech, J. Otero, M. Alhawagri, S. Dai, Y. Abu-Amer, Impediment of NEMOoligomerization inhibits osteoclastogenesis and osteolysis, J. Cell. Biochem. 108(2009) 1337–1345.
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
[83] J. Gohda, T. Akiyama, T. Koga, H. Takayanagi, S. Tanaka, J. Inoue, RANK-mediatedamplification of TRAF6 signaling leads to NFATc1 induction during osteoclasto-genesis, EMBO J. 24 (2005) 790–799.
[84] W.J. Boyle, W.S. Simonet, D.L. Lacey, Osteoclast differentiation and activation,Nature 423 (2003) 337–342.
[85] H. Takayanagi, The role of NFAT in osteoclast formation, Ann. N. Y. Acad. Sci.1116 (2007) 227–237.
[86] N. Ishida, K. Hayashi, M. Hoshijima, T. Ogawa, S. Koga, Y. Miyatake, M.Kumegawa, T. Kimura, T. Takeya, Large scale gene expression analysis of osteo-clastogenesis in vitro and elucidation of NFAT2 as a key regulator, J. Biol. Chem.277 (2002) 41147–41156.
[87] F.S. Fahid, J. Jiang, Q. Zhu, C. Zhang, E. Filbert, K.E. Safavi, L.S. Spangberg, Applica-tion of small interfering RNA for inhibition of lipopolysaccharide-induced oste-oclast formation and cytokine stimulation, J. Endod. 34 (2008) 563–569.
[88] J.P. David, K. Sabapathy, O. Hoffmann, M.H. Idarraga, E.F. Wagner, JNK1 modu-lates osteoclastogenesis through both c-Jun phosphorylation-dependent and-independent mechanisms, J. Cell Sci. 115 (2002) 4317–4325.
[89] L. Kenner, A. Hoebertz, T. Beil, N. Keon, F. Karreth, R. Eferl, H. Scheuch, A.Szremska, M. Amling, M. Schorpp-Kistner, P. Angel, E.F. Wagner, Mice lackingJunB are osteopenic due to cell-autonomous osteoblast and osteoclast defects,J. Cell Biol. 164 (2004) 613–623.
[90] F. Ikeda, R. Nishimura, T. Matsubara, S. Tanaka, J. Inoue, S.V. Reddy, K. Hata, K.Yamashita, T. Hiraga, T. Watanabe, T. Kukita, K. Yoshioka, A. Rao, T. Yoneda, Crit-ical roles of c-Jun signaling in regulation of NFAT family and RANKL-regulatedosteoclast differentiation, J. Clin. Invest. 114 (2004) 475–484.
[91] N.K. Lee, H.K. Choi, H.J. Yoo, J. Shin, S.Y. Lee, RANKL-induced schlafen2 is a pos-itive regulator of osteoclastogenesis, Cell. Signal. 20 (2008) 2302–2308.
[92] F.C. Hartgers, J.L. Vissers, M.W. Looman, C. van Zoelen, C. Huffine, C.G. Figdor, G.J.Adema, DC-STAMP, a novel multimembrane-spanning molecule preferentiallyexpressed by dendritic cells, Eur. J. Immunol. 30 (2000) 3585–3590.
[93] M. Yagi, T. Miyamoto, Y. Sawatani, K. Iwamoto, N. Hosogane, N. Fujita, K. Morita,K. Ninomiya, T. Suzuki, K. Miyamoto, Y. Oike, M. Takeya, Y. Toyama, T. Suda, DC-STAMP is essential for cell–cell fusion in osteoclasts and foreign body giant cells,J. Exp. Med. 202 (2005) 345–351.
[94] T. Kukita, N. Wada, A. Kukita, T. Kakimoto, F. Sandra, K. Toh, K. Nagata, T. Iijima, M.Horiuchi, H. Matsusaki, K. Hieshima, O. Yoshie, H. Nomiyama, RANKL-induced DC-STAMP is essential for osteoclastogenesis, J. Exp. Med. 200 (2004) 941–946.
[95] M. Yang, M.J. Birnbaum, C.A. MacKay, A. Mason-Savas, B. Thompson, P.R. Odgren,Osteoclast stimulatory transmembrane protein (OC-STAMP), a novel protein in-duced by RANKL that promotes osteoclast differentiation, J. Cell. Physiol. 215(2008) 497–505.
[96] S.M. Jay, E. Skokos, F. Laiwalla, M.M. Krady, T.R. Kyriakides, Foreign body giantcell formation is preceded by lamellipodia formation and can be attenuated byinhibition of Rac1 activation, Am. J. Pathol. 171 (2007) 632–640.
[97] M. Ishii, K. Iwai, M. Koike, S. Ohshima, E. Kudo-Tanaka, T. Ishii, T. Mima, Y.Katada, K. Miyatake, Y. Uchiyama, Y. Saeki, RANKL-induced expression of tetra-spanin CD9 in lipid raft membrane microdomain is essential for cell fusion dur-ing osteoclastogenesis, J. Bone Miner. Res. 21 (2006) 965–976.
[98] I. Tachibana, M.E. Hemler, Role of transmembrane 4 superfamily (TM4SF) pro-teins CD9 and CD81 in muscle cell fusion and myotube maintenance, J. CellBiol. 146 (1999) 893–904.
[99] K. Kaji, S. Oda, T. Shikano, T. Ohnuki, Y. Uematsu, J. Sakagami, N. Tada, S. Miya-zaki, A. Kudo, The gamete fusion process is defective in eggs of Cd9-deficientmice, Nat. Genet. 24 (2000) 279–282.
[100] S.M. Crean, J.P. Meneski, T.G. Hullinger, M.J. Reilly, E.H. DeBoever, R.S. Taichman,N-linked sialyated sugar receptors support haematopoietic cell-osteoblast adhe-sions, Br. J. Haematol. 124 (2004) 534–546.
[101] M. Takahata, N. Iwasaki, H. Nakagawa, Y. Abe, T. Watanabe, M. Ito, T. Majima, A.Minami, Sialylation of cell surface glycoconjugates is essential for osteoclasto-genesis, Bone 41 (2007) 77–86.
[102] R.A. Battaglino, L. Pham, L.R. Morse, M. Vokes, A. Sharma, P.R. Odgren, M. Yang,H. Sasaki, P. Stashenko, NHA-oc/NHA2: a mitochondrial cation-proton antipor-ter selectively expressed in osteoclasts, Bone 42 (2008) 180–192.
[103] C.L. Brett, M. Donowitz, R. Rao, Evolutionary origins of eukaryotic sodium/pro-ton exchangers, Am. J. Physiol. Cell Physiol. 288 (2005) C223–C239.
[104] M. Yang, G. Mailhot, M.J. Birnbaum, C.A. MacKay, A. Mason-Savas, P.R. Odgren,Expression of and role for ovarian cancer G-protein-coupled receptor 1(OGR1) during osteoclastogenesis, J. Biol. Chem. 281 (2006) 23598–23605.
[105] J.H. Kehrl, Heterotrimeric G protein signaling: roles in immune function andfine-tuning by RGS proteins, Immunity 8 (1998) 1–10.
[106] C. Beadling, K.M. Druey, G. Richter, J.H. Kehrl, K.A. Smith, Regulators of G proteinsignaling exhibit distinct patterns of gene expression and target G protein spec-ificity in human lymphocytes, J. Immunol. 162 (1999) 2677–2682.
[107] D. Yowe, N. Weich, M. Prabhudas, L. Poisson, P. Errada, R. Kapeller, K. Yu, L.Faron, M. Shen, J. Cleary, T.M. Wilkie, C. Gutierrez-Ramos, M.R. Hodge, RGS18is a myeloerythroid lineage-specific regulator of G-protein-signalling moleculehighly expressed in megakaryocytes, Biochem. J. 359 (2001) 109–118.
[108] K. Iwai, M. Koike, S. Ohshima, K. Miyatake, Y. Uchiyama, Y. Saeki, M. Ishii,RGS18 acts as a negative regulator of osteoclastogenesis by modulating theacid-sensing OGR1/NFAT signaling pathway, J. Bone Miner. Res. 22 (2007)1612–1620.
[109] M.G. Ludwig, M. Vanek, D. Guerini, J.A. Gasser, C.E. Jones, U. Junker, H. Hofstetter,R.M. Wolf, K. Seuwen, Proton-sensing G-protein-coupled receptors, Nature 425(2003) 93–98.
[110] S. Yang, Y.P. Li, RGS12 is essential for RANKL-evoked signaling for terminal dif-ferentiation of osteoclasts in vitro, J. Bone Miner. Res. 22 (2007) 45–54.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

14 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
[111] M.B. Humphrey, K. Ogasawara, W. Yao, S.C. Spusta, M.R. Daws, N.E. Lane, L.L.Lanier, M.C. Nakamura, The signaling adapter protein DAP12 regulates multi-nucleation during osteoclast development, J. Bone Miner. Res. 19 (2004)224–234.
[112] M.B. Humphrey, M.R. Daws, S.C. Spusta, E.C. Niemi, J.A. Torchia, L.L. Lanier, W.E.Seaman, M.C. Nakamura, TREM2, a DAP12-associated receptor, regulates osteo-clast differentiation and function, J. Bone Miner. Res. 21 (2006) 237–245.
[113] S. Yuvaraj, J.J. Blanchard, G. Daughtridge, R.J. Kolb, S. Shanmugarajan, T.A.Bateman, S.V. Reddy, Microarray profile of gene expression during osteoclastdifferentiation in modeled microgravity, J. Cell. Biochem 111 (2010)1179–1187.
[114] A.S. Noman, N. Koide, E.K.I. Iftakhar, J. Dagvadorj, G. Tumurkhuu, Y. Naiki, T.Komatsu, T. Yoshida, T. Yokochi, Retinoblastoma protein-interacting zinc finger1 (RIZ1) participates in RANKL-induced osteoclast formation via regulation ofNFATc1 expression, Immunol. Lett. 131 (2010) 166–169.
[115] S. Huang, G. Shao, L. Liu, The PR domain of the Rb-binding zinc finger protein RIZ1 isa protein binding interface and is related to the SET domain functioning in chroma-tin-mediated gene expression, J. Biol. Chem. 273 (1998) 15933–15939.
[116] T. Carling, K.C. Kim, X.H. Yang, J. Gu, X.K. Zhang, S. Huang, A histone methyl-transferase is required for maximal response to female sex hormones, Mol.Cell. Biol. 24 (2004) 7032–7042.
[117] C. Abbondanza, F. de Nigris, C. De Rosa, R. Rossiello, G.A. Puca, C. Napoli, Silenc-ing of YY1 downregulates RIZ1 promoter in human osteosarcoma, Oncol. Res. 17(2008) 33–41.
[118] A. GuezGuez, V. Prod'homme, X. Mouska, A. Baudot, C. Blin-Wakkach, R.Rottapel, M. Deckert, 3BP2 Adapter protein is required for receptor activator ofNFkappaB ligand (RANKL)-induced osteoclast differentiation of RAW264.7cells, J. Biol. Chem. 285 (2010) 20952–20963.
[119] S. Le Bras, C.Moon, I. Foucault, J.P. Breittmayer,M.Deckert, Abl-SH3 binding protein2, 3BP2, interacts with CIN85 and HIP-55, FEBS Lett. 581 (2007) 967–974.
[120] U. Shukla, T. Hatani, K. Nakashima, K. Ogi, K. Sada, Tyrosine phosphorylation of3BP2 regulates B cell receptor-mediated activation of NFAT, J. Biol. Chem. 284(2009) 33719–33728.
[121] D. Jevremovic, D.D. Billadeau, R.A. Schoon, C.J. Dick, P.J. Leibson, Regulation of NKcell-mediated cytotoxicity by the adaptor protein 3BP2, J. Immunol. 166 (2001)7219–7228.
[122] I. Foucault, S. Le Bras, C. Charvet, C. Moon, A. Altman, M. Deckert, The adaptor pro-tein 3BP2 associates with VAV guanine nucleotide exchange factors to regulateNFAT activation by the B-cell antigen receptor, Blood 105 (2005) 1106–1113.
[123] M. Deckert, S. Tartare-Deckert, J. Hernandez, R. Rottapel, A. Altman, Adaptorfunction for the Syk kinases-interacting protein 3BP2 in IL-2 gene activation, Im-munity 9 (1998) 595–605.
[124] J. Schlondorff, C.P. Blobel, Metalloprotease-disintegrins: modular proteins capa-ble of promoting cell–cell interactions and triggering signals by protein-ectodo-main shedding, J. Cell Sci. 112 (Pt 21) (1999) 3603–3617.
[125] S. Yoshida, M. Setoguchi, Y. Higuchi, S. Akizuki, S. Yamamoto, Molecular cloningof cDNA encoding MS2 antigen, a novel cell surface antigen strongly expressedin murine monocytic lineage, Int. Immunol. 2 (1990) 585–591.
[126] S.J. Choi, J.H. Han, G.D. Roodman, ADAM8: a novel osteoclast stimulating factor, J.Bone Miner. Res. 16 (2001) 814–822.
[127] M. Ainola, T.F. Li, J. Mandelin, M. Hukkanen, S.J. Choi, J. Salo, Y.T. Konttinen, In-volvement of a disintegrin and a metalloproteinase 8 (ADAM8) in osteoclasto-genesis and pathological bone destruction, Ann. Rheum. Dis. 68 (2009)427–434.
[128] L.G. Raisz, H.A. Simmons, A.L. Sandberg, E. Canalis, Direct stimulation of bone re-sorption by epidermal growth factor, Endocrinology 107 (1980) 270–273.
[129] N. Normanno, A. De Luca, D. Aldinucci, M.R. Maiello, M. Mancino, A. D'Antonio, R.De Filippi, A. Pinto, Gefitinib inhibits the ability of human bone marrow stromalcells to induce osteoclast differentiation: implications for the pathogenesis andtreatment of bone metastasis, Endocr. Relat. Cancer 12 (2005) 471–482.
[130] T. Yi, H.L. Lee, J.H. Cha, S.I. Ko, H.J. Kim, H.I. Shin, K.M. Woo, H.M. Ryoo, G.S. Kim,J.H. Baek, Epidermal growth factor receptor regulates osteoclast differentiationand survival through cross-talking with RANK signaling, J. Cell. Physiol. 217(2008) 409–422.
[131] H.C. Blair, M. Zaidi, Osteoclastic differentiation and function regulated by oldand new pathways, Rev. Endocr. Metab. Disord. 7 (2006) 23–32.
[132] L. Xing, E.M. Schwarz, B.F. Boyce, Osteoclast precursors, RANKL/RANK, and im-munology, Immunol. Rev. 208 (2005) 19–29.
[133] H. Hayashi, K. Nakahama, T. Sato, T. Tuchiya, Y. Asakawa, T. Maemura, M.Tanaka, M. Morita, I. Morita, The role of Mac-1 (CD11b/CD18) in osteoclast dif-ferentiation induced by receptor activator of nuclear factor-kappaB ligand, FEBSLett. 582 (2008) 3243–3248.
[134] S.R. Wiley, K. Schooley, P.J. Smolak, W.S. Din, C.P. Huang, J.K. Nicholl, G.R.Sutherland, T.D. Smith, C. Rauch, C.A. Smith, et al., Identification and character-ization of a new member of the TNF family that induces apoptosis, Immunity3 (1995) 673–682.
[135] G. Zauli, E. Rimondi, V. Nicolin, E. Melloni, C. Celeghini, P. Secchiero, TNF-relatedapoptosis-inducing ligand (TRAIL) blocks osteoclastic differentiation induced byRANKL plus M-CSF, Blood 104 (2004) 2044–2050.
[136] S. Roux, P. Lambert-Comeau, C. Saint-Pierre, M. Lepine, B. Sawan, J.L. Parent,Death receptors, Fas and TRAIL receptors, are involved in human osteoclast ap-optosis, Biochem. Biophys. Res. Commun. 333 (2005) 42–50.
[137] G. Zauli, E. Rimondi, S. Stea, F. Baruffaldi, M. Stebel, C. Zerbinati, F. Corallini, P.Secchiero, TRAIL inhibits osteoclastic differentiation by counteracting RANKL-dependent p27Kip1 accumulation in pre-osteoclast precursors, J. Cell. Physiol.214 (2008) 117–125.
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
[138] N. Okahashi, Y. Murase, T. Koseki, T. Sato, K. Yamato, T. Nishihara, Osteoclast dif-ferentiation is associated with transient upregulation of cyclin-dependent ki-nase inhibitors p21(WAF1/CIP1) and p27(KIP1), J. Cell. Biochem. 80 (2001)339–345.
[139] U. Sankar, K. Patel, T.J. Rosol, M.C. Ostrowski, RANKL coordinates cell cycle with-drawal and differentiation in osteoclasts through the cyclin-dependent kinaseinhibitors p27KIP1 and p21CIP1, J. Bone Miner. Res. 19 (2004) 1339–1348.
[140] P. Saftig, E. Hunziker, O. Wehmeyer, S. Jones, A. Boyde, W. Rommerskirch, J.D.Moritz, P. Schu, K. von Figura, Impaired osteoclastic bone resorption leads toosteopetrosis in cathepsin-K-deficient mice, Proc. Natl. Acad. Sci. U.S.A. 95(1998) 13453–13458.
[141] S. Corisdeo, M. Gyda, M. Zaidi, B.S. Moonga, B.R. Troen, New insights into theregulation of cathepsin K gene expression by osteoprotegerin ligand, Biochem.Biophys. Res. Commun. 285 (2001) 335–339.
[142] V. Shalhoub, J. Faust, W.J. Boyle, C.R. Dunstan, M. Kelley, S. Kaufman, S. Scully, G.Van, D.L. Lacey, Osteoprotegerin and osteoprotegerin ligand effects on osteoclastformation from human peripheral blood mononuclear cell precursors, J. Cell.Biochem. 72 (1999) 251–261.
[143] M. Pang, A.F. Martinez, I. Fernandez, W. Balkan, B.R. Troen, AP-1 stimulates thecathepsin K promoter in RAW 264.7 cells, Gene 403 (2007) 151–158.
[144] W. Kafienah, D. Bromme, D.J. Buttle, L.J. Croucher, A.P. Hollander, Human ca-thepsin K cleaves native type I and II collagens at the N-terminal end of the tri-ple helix, Biochem. J. 331 (Pt 3) (1998) 727–732.
[145] M.R. Johnson, M.H. Polymeropoulos, H.L. Vos, R.I. Ortiz de Luna, C.A. Franco-mano, A nonsense mutation in the cathepsin K gene observed in a family withpycnodysostosis, Genome Res. 6 (1996) 1050–1055.
[146] C.I. Selinger, C.J. Day, N.A. Morrison, Optimized transfection of diced siRNA intomature primary human osteoclasts: inhibition of cathepsin K mediated bone re-sorption by siRNA, J. Cell. Biochem. 96 (2005) 996–1002.
[147] T.C. Phan, J. Xu, M.H. Zheng, Interaction between osteoblast and osteoclast: im-pact in bone disease, Histol. Histopathol. 19 (2004) 1325–1344.
[148] Z.H. Lee, H.H. Kim, Signal transduction by receptor activator of nuclear factorkappa B in osteoclasts, Biochem. Biophys. Res. Commun. 305 (2003) 211–214.
[149] M.A. Lomaga, W.C. Yeh, I. Sarosi, G.S. Duncan, C. Furlonger, A. Ho, S. Morony, C.Capparelli, G. Van, S. Kaufman, A. van der Heiden, A. Itie, A. Wakeham, W.Khoo, T. Sasaki, Z. Cao, J.M. Penninger, C.J. Paige, D.L. Lacey, C.R. Dunstan, W.J.Boyle, D.V. Goeddel, T.W. Mak, TRAF6 deficiency results in osteopetrosis and de-fective interleukin-1, CD40, and LPS signaling, Genes Dev. 13 (1999) 1015–1024.
[150] N. Kobayashi, Y. Kadono, A. Naito, K. Matsumoto, T. Yamamoto, S. Tanaka, J.Inoue, Segregation of TRAF6-mediated signaling pathways clarifies its role inosteoclastogenesis, EMBO J. 20 (2001) 1271–1280.
[151] P. Angel, M. Karin, The role of Jun, Fos and the AP-1 complex in cell-proliferationand transformation, Biochim. Biophys. Acta 1072 (1991) 129–157.
[152] H.J. Kim, Y. Lee, E.J. Chang, H.M. Kim, S.P. Hong, Z.H. Lee, J. Ryu, H.H. Kim, Sup-pression of osteoclastogenesis by N, N-dimethyl-D-erythro-sphingosine: asphingosine kinase inhibition-independent action, Mol. Pharmacol. 72 (2007)418–428.
[153] Y.P. Li, W. Chen, Y. Liang, E. Li, P. Stashenko, Atp6i-deficient mice exhibit severeosteopetrosis due to loss of osteoclast-mediated extracellular acidification, Nat.Genet. 23 (1999) 447–451.
[154] A. Frattini, P.J. Orchard, C. Sobacchi, S. Giliani, M. Abinun, J.P. Mattsson, D.J.Keeling, A.K. Andersson, P. Wallbrandt, L. Zecca, L.D. Notarangelo, P. Vezzoni,A. Villa, Defects in TCIRG1 subunit of the vacuolar proton pump are responsiblefor a subset of human autosomal recessive osteopetrosis, Nat. Genet. 25 (2000)343–346.
[155] Y. Hu, J. Nyman, P. Muhonen, H.K. Vaananen, T. Laitala-Leinonen, Inhibition ofthe osteoclast V-ATPase by small interfering RNAs, FEBS Lett. 579 (2005)4937–4942.
[156] J. Xu, T. Cheng, H.T. Feng, N.J. Pavlos, M.H. Zheng, Structure and function of V-ATPases in osteoclasts: potential therapeutic targets for the treatment of osteo-lysis, Histol. Histopathol. 22 (2007) 443–454.
[157] M. Forgac, Structure and properties of the vacuolar (H+)-ATPases, J. Biol. Chem.274 (1999) 12951–12954.
[158] T. Xu, E. Vasilyeva, M. Forgac, Subunit interactions in the clathrin-coated vesiclevacuolar (H(+))-ATPase complex, J. Biol. Chem. 274 (1999) 28909–28915.
[159] G.H. Sun-Wada, T. Yoshimizu, Y. Imai-Senga, Y. Wada, M. Futai, Diversity ofmouse proton-translocating ATPase: presence of multiple isoforms of the C, dand G subunits, Gene 302 (2003) 147–153.
[160] S. Feng, L. Deng, W. Chen, J. Shao, G. Xu, Y.P. Li, Atp6v1c1 is an essential compo-nent of the osteoclast proton pump and in F-actin ring formation in osteoclasts,Biochem. J. 417 (2009) 195–203.
[161] U. Kornak, D. Kasper, M.R. Bosl, E. Kaiser, M. Schweizer, A. Schulz, W. Friedrich,G. Delling, T.J. Jentsch, Loss of the ClC-7 chloride channel leads to osteopetrosisin mice and man, Cell 104 (2001) 205–215.
[162] F. Okamoto, H. Kajiya, K. Toh, S. Uchida, M. Yoshikawa, S. Sasaki, M.A. Kido, T.Tanaka, K. Okabe, Intracellular ClC-3 chloride channels promote bone resorptionin vitro through organelle acidification in mouse osteoclasts, Am. J. Physiol. CellPhysiol. 294 (2008) C693–C701.
[163] H. Kajiya, F. Okamoto, J.P. Li, A. Nakao, K. Okabe, Expression of mouse osteoclastK–Cl Co-transporter-1 and its role during bone resorption, J. Bone Miner. Res. 21(2006) 984–992.
[164] P.K. Lauf, J. Bauer, N.C. Adragna, H. Fujise, A.M. Zade-Oppen, K.H. Ryu, E. Delpire,Erythrocyte K–Cl cotransport: properties and regulation, Am. J. Physiol. 263(1992) C917–C932.
[165] L. Reuss, Basolateral KCl co-transport in a NaCl-absorbing epithelium, Nature305 (1983) 723–726.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

15Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
[166] K.D. Philipson, D.A. Nicoll, M. Ottolia, B.D. Quednau, H. Reuter, S. John, Z. Qiu, TheNa+/Ca2+ exchange molecule: an overview, Ann. N. Y. Acad. Sci. 976 (2002)1–10.
[167] K.D. Philipson, S. Longoni, R. Ward, Purification of the cardiac Na+–Ca2+ ex-change protein, Biochim. Biophys. Acta 945 (1988) 298–306.
[168] D.A. Nicoll, L.V. Hryshko, S. Matsuoka, J.S. Frank, K.D. Philipson, Mutation ofamino acid residues in the putative transmembrane segments of the cardiac sar-colemmal Na+–Ca2+ exchanger, J. Biol. Chem. 271 (1996) 13385–13391.
[169] J.P. Li, H. Kajiya, F. Okamoto, A. Nakao, T. Iwamoto, K. Okabe, Three Na+/Ca2+ exchanger (NCX) variants are expressed in mouse osteoclasts and mediatecalcium transport during bone resorption, Endocrinology 148 (2007)2116–2125.
[170] P. Kefalas, T.R. Brown, P.M. Brickell, Signalling by the p60c-src family of protein-tyrosine kinases, Int. J. Biochem. Cell Biol. 27 (1995) 551–563.
[171] P. Soriano, C. Montgomery, R. Geske, A. Bradley, Targeted disruption of the c-srcproto-oncogene leads to osteopetrosis in mice, Cell 64 (1991) 693–702.
[172] P.L. Schwartzberg, L. Xing, O. Hoffmann, C.A. Lowell, L. Garrett, B.F. Boyce, H.E.Varmus, Rescue of osteoclast function by transgenic expression of kinase-defi-cient Src in src−/− mutant mice, Genes Dev. 11 (1997) 2835–2844.
[173] S. Tanaka, N. Takahashi, N. Udagawa, T. Sasaki, Y. Fukui, T. Kurokawa, T. Suda,Osteoclasts express high levels of p60c-src, preferentially on ruffled bordermembranes, FEBS Lett. 313 (1992) 85–89.
[174] B.F. Boyce, T. Yoneda, C. Lowe, P. Soriano, G.R. Mundy, Requirement of pp 60c-src expression for osteoclasts to form ruffled borders and resorb bone in mice,J. Clin. Invest. 90 (1992) 1622–1627.
[175] S.E. Winograd-Katz, M.C. Brunner, N. Mirlas, B. Geiger, Analysis of the signalingpathways regulating Src-dependent remodeling of the actin cytoskeleton, Eur. J.Cell Biol. 90 (2011) 143–156.
[176] L.W.Wu, D.J. Baylink, K.H. Lau, Molecular cloning and expression of a unique rab-bit osteoclastic phosphotyrosyl phosphatase, Biochem. J. 316 (Pt 2) (1996)515–523.
[177] M. Amoui, M.H. Sheng, S.T. Chen, D.J. Baylink, K.H. Lau, A transmembrane osteo-clastic protein-tyrosine phosphatase regulates osteoclast activity in part by pro-moting osteoclast survival through c-Src-dependent activation of NFkappaB andJNK2, Arch. Biochem. Biophys. 463 (2007) 47–59.
[178] J.H. Yang, M. Amoui, K.H. Lau, Targeted deletion of the osteoclast protein-tyro-sine phosphatase (PTP-oc) promoter prevents RANKL-mediated osteoclastic dif-ferentiation of RAW264.7 cells, FEBS Lett. 581 (2007) 2503–2508.
[179] S.L. Teitelbaum, Bone resorption by osteoclasts, Science 289 (2000)1504–1508.
[180] P. Lakkakorpi, J. Tuukkanen, T. Hentunen, K. Jarvelin, K. Vaananen, Organizationof osteoclast microfilaments during the attachment to bone surface in vitro, J.Bone Miner. Res. 4 (1989) 817–825.
[181] R.S. Biswas, D. Baker, K.A. Hruska, M.A. Chellaiah, Polyphosphoinositides-depen-dent regulation of the osteoclast actin cytoskeleton and bone resorption, BMCCell Biol. 5 (2004) 19.
[182] M.A. Chellaiah, Regulation of actin ring formation by rho GTPases in osteoclasts,J. Biol. Chem. 280 (2005) 32930–32943.
[183] M. Chellaiah, N. Kizer, M. Silva, U. Alvarez, D. Kwiatkowski, K.A. Hruska, Gelsolindeficiency blocks podosome assembly and produces increased bone mass andstrength, J. Cell Biol. 148 (2000) 665–678.
[184] Y. Calle, G.E. Jones, C. Jagger, K. Fuller, M.P. Blundell, J. Chow, T. Chambers, A.J.Thrasher, WASp deficiency in mice results in failure to form osteoclast sealingzones and defects in bone resorption, Blood 103 (2004) 3552–3561.
[185] R. Rohatgi, L. Ma, H. Miki, M. Lopez, T. Kirchhausen, T. Takenawa, M.W.Kirschner, The interaction between N-WASP and the Arp2/3 complex linksCdc42-dependent signals to actin assembly, Cell 97 (1999) 221–231.
[186] R.H. Insall, L.M. Machesky, Regulation of WASP: PIP2 pipped by Toca-1? Cell 118(2004) 140–141.
[187] M. Glogauer, J. Hartwig, T. Stossel, Two pathways through Cdc42 couple the N-formyl receptor to actin nucleation in permeabilized human neutrophils, J. CellBiol. 150 (2000) 785–796.
[188] A.L. Rozelle, L.M. Machesky, M. Yamamoto, M.H. Driessens, R.H. Insall, M.G. Roth,K. Luby-Phelps, G. Marriott, A. Hall, H.L. Yin, Phosphatidylinositol 4,5-bispho-sphate induces actin-based movement of raft-enriched vesicles throughWASP-Arp2/3, Curr. Biol. 10 (2000) 311–320.
[189] L.M. Machesky, K.L. Gould, The Arp2/3 complex: a multifunctional actin organiz-er, Curr. Opin. Cell Biol. 11 (1999) 117–121.
[190] I.R. Hurst, J. Zuo, J. Jiang, L.S. Holliday, Actin-related protein 2/3 complex is re-quired for actin ring formation, J. Bone Miner. Res. 19 (2004) 499–506.
[191] E. Torres, M.K. Rosen, Protein-tyrosine kinase and GTPase signals cooperate tophosphorylate and activate Wiskott-Aldrich syndrome protein (WASP)/neuro-nal WASP, J. Biol. Chem. 281 (2006) 3513–3520.
[192] X. Wu, S. Suetsugu, L.A. Cooper, T. Takenawa, J.L. Guan, Focal adhesion kinaseregulation of N-WASP subcellular localization and function, J. Biol. Chem. 279(2004) 9565–9576.
[193] G.O. Cory, R. Cramer, L. Blanchoin, A.J. Ridley, Phosphorylation of the WASP-VCAdomain increases its affinity for the Arp2/3 complex and enhances actin poly-merization by WASP, Mol. Cell 11 (2003) 1229–1239.
[194] M.A. Chellaiah, D. Kuppuswamy, L. Lasky, S. Linder, Phosphorylation of a Wis-cott-Aldrich syndrome protein-associated signal complex is critical in osteoclastbone resorption, J. Biol. Chem. 282 (2007) 10104–10116.
[195] M.A. Chellaiah, M.D. Schaller, Activation of Src kinase by protein-tyrosine phos-phatase-PEST in osteoclasts: comparative analysis of the effects of bisphospho-nate and protein-tyrosine phosphatase inhibitor on Src activation in vitro, J.Cell. Physiol. 220 (2009) 382–393.
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
[196] A.M. Weaver, A.V. Karginov, A.W. Kinley, S.A. Weed, Y. Li, J.T. Parsons, J.A.Cooper, Cortactin promotes and stabilizes Arp2/3-induced actin filament net-work formation, Curr. Biol. 11 (2001) 370–374.
[197] T. Ma, K. Sadashivaiah, N. Madayiputhiya, M.A. Chellaiah, Regulation of sealingring formation by L-plastin and cortactin in osteoclasts, J. Biol. Chem. 285(2011) 29911–29924.
[198] J. Cui, S.J. Matkovich, N. deSouza, S. Li, N. Rosemblit, A.R. Marks, Regulation of thetype 1 inositol 1,4,5-trisphosphate receptor by phosphorylation at tyrosine 353,J. Biol. Chem. 279 (2004) 16311–16316.
[199] B.B. Yaroslavskiy, I. Turkova, Y. Wang, L.J. Robinson, H.C. Blair, Functional osteo-clast attachment requires inositol-1,4,5-trisphosphate receptor-associatedcGMP-dependent kinase substrate, Lab. Invest. 90 (2010) 1533–1542.
[200] E. Bonnelye, F. Saltel, A. Chabadel, R.A. Zirngibl, J.E. Aubin, P. Jurdic, Involvementof the orphan nuclear estrogen receptor-related receptor alpha in osteoclast ad-hesion and transmigration, J. Mol. Endocrinol. 45 (2010) 365–377.
[201] M.J. Tyska, D.M. Warshaw, The myosin power stroke, Cell Motil. Cytoskeleton 51(2002) 1–15.
[202] B.K. McMichael, R.E. Cheney, B.S. Lee, Myosin X regulates sealing zone pattern-ing in osteoclasts through linkage of podosomes and microtubules, J. Biol.Chem. 285 (2010) 9506–9515.
[203] J.S. Berg, B.H. Derfler, C.M. Pennisi, D.P. Corey, R.E. Cheney, Myosin-X, a novelmyosin with pleckstrin homology domains, associates with regions of dynamicactin, J. Cell Sci. 113 (Pt 19) (2000) 3439–3451.
[204] K.L. Weber, A.M. Sokac, J.S. Berg, R.E. Cheney, W.M. Bement, A microtubule-binding myosin required for nuclear anchoring and spindle assembly, Nature431 (2004) 325–329.
[205] S. Okumura, T. Mizoguchi, N. Sato, M. Yamaki, Y. Kobayashi, H. Yamauchi, H.Ozawa, N. Udagawa, N. Takahashi, Coordination of microtubules and the actincytoskeleton is important in osteoclast function, but calcitonin disrupts sealingzones without affecting microtubule networks, Bone 39 (2006) 684–693.
[206] P. Jurdic, F. Saltel, A. Chabadel, O. Destaing, Podosome and sealing zone: specific-ity of the osteoclast model, Eur. J. Cell Biol. 85 (2006) 195–202.
[207] E.J. Chang, J. Ha, F. Oerlemans, Y.J. Lee, S.W. Lee, J. Ryu, H.J. Kim, Y. Lee, H.M. Kim,J.Y. Choi, J.Y. Kim, C.S. Shin, Y.K. Pak, S. Tanaka, B. Wieringa, Z.H. Lee, H.H. Kim,Brain-type creatine kinase has a crucial role in osteoclast-mediated bone resorp-tion, Nat. Med. 14 (2008) 966–972.
[208] M. Wyss, R. Kaddurah-Daouk, Creatine and creatinine metabolism, Physiol. Rev.80 (2000) 1107–1213.
[209] D. Zhang, N. Udagawa, I. Nakamura, H. Murakami, S. Saito, K. Yamasaki, Y. Shiba-saki, N. Morii, S. Narumiya, N. Takahashi, et al., The small GTP-binding protein,rho p21, is involved in bone resorption by regulating cytoskeletal organizationin osteoclasts, J. Cell Sci. 108 (Pt 6) (1995) 2285–2292.
[210] E. van Beek, E. Pieterman, L. Cohen, C. Lowik, S. Papapoulos, Farnesyl pyrophos-phate synthase is the molecular target of nitrogen-containing bisphosphonates,Biochem. Biophys. Res. Commun. 264 (1999) 108–111.
[211] J.C. Lacal, Regulation of proliferation and apoptosis by Ras and Rho GTPasesthrough specific phospholipid-dependent signaling, FEBS Lett. 410 (1997)73–77.
[212] G.G. Reinholz, B. Getz, E.S. Sanders, M.Y. Karpeisky, N. Padyukova, S.N. Mikhai-lov, J.N. Ingle, T.C. Spelsberg, Distinct mechanisms of bisphosphonate action be-tween osteoblasts and breast cancer cells: identity of a potent newbisphosphonate analogue, Breast Cancer Res. Treat. 71 (2002) 257–268.
[213] Y. Wang, A. Panasiuk, D.W. Grainger, Small interfering RNA knocks down themolecular target of alendronate, farnesyl pyrophosphate synthase, in osteoclastand osteoblast cultures, Mol. Pharm. 8 (2011) 1016–1024.
[214] G. Shankar, I. Davison, M.H. Helfrich, W.T. Mason, M.A. Horton, Integrin recep-tor-mediated mobilisation of intranuclear calcium in rat osteoclasts, J. Cell Sci.105 (Pt 1) (1993) 61–68.
[215] S. Nesbitt, A. Nesbit, M. Helfrich, M. Horton, Biochemical characterization ofhuman osteoclast integrins. Osteoclasts express alpha v beta 3, alpha 2 beta 1,and alpha v beta 1 integrins, J. Biol. Chem. 268 (1993) 16737–16745.
[216] K.P. McHugh, K. Hodivala-Dilke, M.H. Zheng, N. Namba, J. Lam, D. Novack, X. Feng,F.P. Ross, R.O. Hynes, S.L. Teitelbaum,Mice lacking beta3 integrins are osteoscleroticbecause of dysfunctional osteoclasts, J. Clin. Invest. 105 (2000) 433–440.
[217] M.A. Horton, M.L. Taylor, T.R. Arnett, M.H. Helfrich, Arg-Gly-Asp (RGD) peptidesand the anti-vitronectin receptor antibody 23C6 inhibit dentine resorption andcell spreading by osteoclasts, Exp. Cell Res. 195 (1991) 368–375.
[218] H.D. Han, L.S. Mangala, J.W. Lee, M.M. Shahzad, H.S. Kim, D. Shen, E.J. Nam, E.M.Mora, R.L. Stone, C. Lu, S.J. Lee, J.W. Roh, A.M. Nick, G. Lopez-Berestein, A.K. Sood,Targeted gene silencing using RGD-labeled chitosan nanoparticles, Clin. CancerRes. 16 (2010) 3910–3922.
[219] V.W. Engleman, G.A. Nickols, F.P. Ross, M.A. Horton, D.W. Griggs, S.L. Settle, P.G.Ruminski, S.L. Teitelbaum, A peptidomimetic antagonist of the alpha(v)beta3integrin inhibits bone resorption in vitro and prevents osteoporosis in vivo, J.Clin. Invest. 99 (1997) 2284–2292.
[220] Y. Komano, T. Nanki, K. Hayashida, K. Taniguchi, N. Miyasaka, Identification of ahuman peripheral blood monocyte subset that differentiates into osteoclasts,Arthritis Res. Ther. 8 (2006) R152.
[221] M. Tucci, R. De Palma, L. Lombardi, G. Rodolico, L. Berrino, F. Dammacco, F.Silvestris, beta(3) Integrin subunit mediates the bone-resorbing functionexerted by cultured myeloma plasma cells, Cancer Res. 69 (2009)6738–6746.
[222] S.H. Kim, C.R. Dass, p53-targeted cancer pharmacotherapy: move towards smallmolecule compounds, J. Pharm. Pharmacol. 63 (2011) 603–610.
[223] D. Michael, M. Oren, The p53-Mdm2 module and the ubiquitin system, Semin.Cancer Biol. 13 (2003) 49–58.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

16 Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
[224] L.T. Vassilev, B.T. Vu, B. Graves, D. Carvajal, F. Podlaski, Z. Filipovic, N. Kong, U.Kammlott, C. Lukacs, C. Klein, N. Fotouhi, E.A. Liu, In vivo activation of thep53 pathway by small-molecule antagonists of MDM2, Science 303 (2004)844–848.
[225] D. Carvajal, C. Tovar, H. Yang, B.T. Vu, D.C. Heimbrook, L.T. Vassilev, Activation ofp53 by MDM2 antagonists can protect proliferating cells from mitotic inhibitors,Cancer Res. 65 (2005) 1918–1924.
[226] G. Zauli, E. Rimondi, F. Corallini, R. Fadda, S. Capitani, P. Secchiero, MDM2 antag-onist Nutlin-3 suppresses the proliferation and differentiation of human pre-os-teoclasts through a p53-dependent pathway, J. Bone Miner. Res. 22 (2007)1621–1630.
[227] S.V. Komarova, A. Pereverzev, J.W. Shum, S.M. Sims, S.J. Dixon, Convergent sig-naling by acidosis and receptor activator of NF-kappaB ligand (RANKL) on thecalcium/calcineurin/NFAT pathway in osteoclasts, Proc. Natl. Acad. Sci. U.S.A.102 (2005) 2643–2648.
[228] F.P. Coxon, M.H. Helfrich, R. Van't Hof, S. Sebti, S.H. Ralston, A. Hamilton, M.J.Rogers, Protein geranylgeranylation is required for osteoclast formation, func-tion, and survival: inhibition by bisphosphonates and GGTI-298, J. Bone Miner.Res. 15 (2000) 1467–1476.
[229] J.E. Fisher, M.J. Rogers, J.M. Halasy, S.P. Luckman, D.E. Hughes, P.J. Masarachia, G.Wesolowski, R.G. Russell, G.A. Rodan, A.A. Reszka, Alendronate mechanism ofaction: geranylgeraniol, an intermediate in the mevalonate pathway, preventsinhibition of osteoclast formation, bone resorption, and kinase activation invitro, Proc. Natl. Acad. Sci. U.S.A. 96 (1999) 133–138.
[230] H. Harada, T. Kukita, A. Kukita, Y. Iwamoto, T. Iijima, Involvement of lymphocytefunction-associated antigen-1 and intercellular adhesion molecule-1 in osteo-clastogenesis: a possible role in direct interaction between osteoclast precur-sors, Endocrinology 139 (1998) 3967–3975.
[231] N. Tani-Ishii, J.M. Penninger, G. Matsumoto, T. Teranaka, T. Umemoto, The role ofLFA-1 in osteoclast development induced by co-cultures of mouse bone marrowcells and MC3T3-G2/PA6 cells, J. Periodontal Res. 37 (2002) 184–191.
[232] B.R. Wong, J. Rho, J. Arron, E. Robinson, J. Orlinick, M. Chao, S. Kalachikov, E.Cayani, F.S. Bartlett 3rd, W.N. Frankel, S.Y. Lee, Y. Choi, TRANCE is a novel ligandof the tumor necrosis factor receptor family that activates c-Jun N-terminal ki-nase in T cells, J. Biol. Chem. 272 (1997) 25190–25194.
[233] Y. Okada, I. Morimoto, K. Ura, K. Watanabe, S. Eto, M. Kumegawa, L. Raisz, C.Pilbeam, Y. Tanaka, Cell-to-cell adhesion via intercellular adhesion molecule-1and leukocyte function-associated antigen-1 pathway is involved in 1alpha,25(OH)2D3, PTH and IL-1alpha-induced osteoclast differentiation and bone re-sorption, Endocr. J. 49 (2002) 483–495.
[234] W.M. Pierce Jr., L.C. Waite, Bone-targeted carbonic anhydrase inhibitors: effect of aproinhibitor on bone resorption in vitro, Proc. Soc. Exp. Biol. Med. 186 (1987) 96–102.
[235] F. Hosain, R.P. Spencer, H.M. Couthon, G.L. Sturtz, Targeted delivery of antineo-plastic agent to bone: biodistribution studies of technetium-99m-labeled gem-bisphosphonate conjugate of methotrexate, J. Nucl. Med. 37 (1996) 105–107.
[236] H. Uludag, J. Yang, Targeting systemically administered proteins to bone bybisphosphonate conjugation, Biotechnol. Prog. 18 (2002) 604–611.
[237] J. Soutschek, A. Akinc, B. Bramlage, K. Charisse, R. Constien, M. Donoghue, S.Elbashir, A. Geick, P. Hadwiger, J. Harborth, M. John, V. Kesavan, G. Lavine, R.K.Pandey, T. Racie, K.G. Rajeev, I. Rohl, I. Toudjarska, G. Wang, S. Wuschko, D. Bum-crot, V. Koteliansky, S. Limmer, M. Manoharan, H.P. Vornlocher, Therapeutic si-lencing of an endogenous gene by systemic administration of modified siRNAs,Nature 432 (2004) 173–178.
[238] Y. Minakuchi, F. Takeshita, N. Kosaka, H. Sasaki, Y. Yamamoto, M. Kouno, K.Honma, S. Nagahara, K. Hanai, A. Sano, T. Kato, M. Terada, T. Ochiya, Atelocolla-gen-mediated synthetic small interfering RNA delivery for effective gene silenc-ing in vitro and in vivo, Nucleic Acids Res. 32 (2004) e109.
[239] F. Takeshita, Y. Minakuchi, S. Nagahara, K. Honma, H. Sasaki, K. Hirai, T. Teratani,N. Namatame, Y. Yamamoto, K. Hanai, T. Kato, A. Sano, T. Ochiya, Efficient deliv-ery of small interfering RNA to bone-metastatic tumors by using atelocollagen invivo, Proc. Natl. Acad. Sci. U.S.A. 102 (2005) 12177–12182.
[240] D. Bumcrot, M. Manoharan, V. Koteliansky, D.W. Sah, RNAi therapeutics: a po-tential new class of pharmaceutical drugs, Nat. Chem. Biol. 2 (2006) 711–719.
[241] T. Tomita, E. Takeuchi, N. Tomita, R. Morishita, M. Kaneko, K. Yamamoto, T.Nakase, H. Seki, K. Kato, Y. Kaneda, T. Ochi, Suppressed severity of collagen-in-duced arthritis by in vivo transfection of nuclear factor kappaB decoy oligodeox-ynucleotides as a gene therapy, Arthritis Rheum. 42 (1999) 2532–2542.
[242] J.A. Roman-Blas, S.A. Jimenez, NF-kappaB as a potential therapeutic target in oste-oarthritis and rheumatoid arthritis, Osteoarthritis Cartilage 14 (2006) 839–848.
[243] R.M. Schiffelers, J. Xu, G. Storm, M.C. Woodle, P.V. Scaria, Effects of treatmentwith small interfering RNA on joint inflammation in mice with collagen-inducedarthritis, Arthritis Rheum. 52 (2005) 1314–1318.
[244] M. Khoury, P. Louis-Plence, V. Escriou, D. Noel, C. Largeau, C. Cantos, D.Scherman, C. Jorgensen, F. Apparailly, Efficient new cationic liposome formula-tion for systemic delivery of small interfering RNA silencing tumor necrosis fac-tor alpha in experimental arthritis, Arthritis Rheum. 54 (2006) 1867–1877.
[245] S. Hoshika, N. Minakawa, A. Matsuda, Synthesis and physical and physiologicalproperties of 4′-thioRNA: application to post-modification of RNA aptamer to-ward NF-kappaB, Nucleic Acids Res. 32 (2004) 3815–3825.
[246] C.R. Allerson, N. Sioufi, R. Jarres, T.P. Prakash, N. Naik, A. Berdeja, L. Wanders, R.H.Griffey, E.E. Swayze, B. Bhat, Fully 2′-modified oligonucleotide duplexes withimproved in vitro potency and stability compared to unmodified small interfer-ing RNA, J. Med. Chem. 48 (2005) 901–904.
[247] B.J. Li, Q. Tang, D. Cheng, C. Qin, F.Y. Xie, Q.Wei, J. Xu, Y. Liu, B.J. Zheng,M.C.Woodle,N. Zhong, P.Y. Lu, Using siRNA in prophylactic and therapeutic regimens againstSARS coronavirus in Rhesus macaque, Nat. Med. 11 (2005) 944–951.
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
[248] S. Choung, Y.J. Kim, S. Kim, H.O. Park, Y.C. Choi, Chemical modification of siRNAsto improve serum stability without loss of efficacy, Biochem. Biophys. Res. Com-mun. 342 (2006) 919–927.
[249] P. Dande, T.P. Prakash, N. Sioufi, H. Gaus, R. Jarres, A. Berdeja, E.E. Swayze, R.H.Griffey, B. Bhat, Improving RNA interference in mammalian cells by 4′-thio-modified small interfering RNA (siRNA): effect on siRNA activity and nucleasestability when used in combination with 2′-O-alkyl modifications, J. Med.Chem. 49 (2006) 1624–1634.
[250] E. Song, P. Zhu, S.K. Lee, D. Chowdhury, S. Kussman, D.M. Dykxhoorn, Y. Feng, D.Palliser, D.B. Weiner, P. Shankar, W.A. Marasco, J. Lieberman, Antibody mediatedin vivo delivery of small interfering RNAs via cell-surface receptors, Nat. Bio-technol. 23 (2005) 709–717.
[251] D. Wang, S. Miller, M. Sima, P. Kopeckova, J. Kopecek, Synthesis and evaluationof water-soluble polymeric bone-targeted drug delivery systems, Bioconjug.Chem. 14 (2003) 853–859.
[252] M. Newa, K.H. Bhandari, L. Tang, R. Kalvapalle, M. Suresh, M.R. Doschak, Anti-body-mediated “universal” osteoclast targeting platform using calcitonin as amodel drug, Pharm. Res. 28 (2011) 1131–1143.
[253] D. Wang, S.C. Miller, P. Kopeckova, J. Kopecek, Bone-targeting macromoleculartherapeutics, Adv. Drug Deliv. Rev. 57 (2005) 1049–1076.
[254] K. Yokogawa, K. Miya, T. Sekido, Y. Higashi, M. Nomura, R. Fujisawa, K. Morito, Y.Masamune, Y. Waki, S. Kasugai, K. Miyamoto, Selective delivery of estradiol tobone by aspartic acid oligopeptide and its effects on ovariectomized mice, Endo-crinology 142 (2001) 1228–1233.
[255] S. Kasugai, R. Fujisawa, Y. Waki, K. Miyamoto, K. Ohya, Selective drug deliverysystem to bone: small peptide (Asp)6 conjugation, J. Bone Miner. Res. 15(2000) 936–943.
[256] M. Aouadi, G.J. Tesz, S.M. Nicoloro, M. Wang, M. Chouinard, E. Soto, G.R. Ostroff,M.P. Czech, Orally delivered siRNA targeting macrophage Map4k4 suppressessystemic inflammation, Nature 458 (2009) 1180–1184.
[257] S.Y. Wu, N.A. McMillan, Lipidic systems for in vivo siRNA delivery, AAPS J. 11(2009) 639–652.
[258] M.D. Krebs, E. Alsberg, Localized, targeted, and sustained siRNA delivery, Chem-istry 17 (2011) 3054–3062.
[259] E. Song, S.K. Lee, J. Wang, N. Ince, N. Ouyang, J. Min, J. Chen, P. Shankar, J. Lieber-man, RNA interference targeting Fas protects mice from fulminant hepatitis, Nat.Med. 9 (2003) 347–351.
[260] Y. Tabata, Y. Ikada, Phagocytosis of polymer microspheres by macrophages, Adv.Polym. Sci. 94 (1990) 110–141.
[261] M. Marsh, H.T. McMahon, The structural era of endocytosis, Science 285 (1999)215–220.
[262] L. Kobzik, S. Huang, J.D. Paulauskis, J.J. Godleski, Particle opsonization and lungmacrophage cytokine response. In vitro and in vivo analysis, J. Immunol. 151(1993) 2753–2759.
[263] D. Cejka, D. Losert, V. Wacheck, Short interfering RNA (siRNA): tool or therapeu-tic? Clin. Sci. (Lond.) 110 (2006) 47–58.
[264] M.S. Taljanovic, M.D. Jones, J.T. Ruth, J.B. Benjamin, J.E. Sheppard, T.B. Hunter,Fracture fixation, Radiographics 23 (2003) 1569–1590.
[265] Y. Zhang, J. Song, B. Shi, Y. Wang, X. Chen, C. Huang, X. Yang, D. Xu, X. Cheng, X.Chen, Combination of scaffold and adenovirus vectors expressing bone morpho-genetic protein-7 for alveolar bone regeneration at dental implant defects, Bio-materials 28 (2007) 4635–4642.
[266] A. Perry, A. Mahar, J. Massie, N. Arrieta, S. Garfin, C. Kim, Biomechanical evalua-tion of kyphoplasty with calcium sulfate cement in a cadaveric osteoporotic ver-tebral compression fracture model, Spine J. 5 (2005) 489–493.
[267] L.L. Dreinhöfer Karsten, Maalouf Ghassan, Bone augmentation in osteoporosis —an update on vertebroplasty and kyphoplasty, Eur. Musculoskelet. Rev. 3 (2008)14–16.
[268] J. Jackson, C. Duncan, C. Mugabe, H. Burt, The use of bone cement for the local-ized, controlled release of the antibiotics vancomycin, linezolid, or fusidic acid:effect of additives on drug release rates and mechanical strength, Drug Deliv.and Transl. Res. 1 (2011) 121–131.
[269] D. Neut, O.S. Kluin, J. Thompson, H.C. van der Mei, H.J. Busscher, Gentamicin re-lease from commercially-available gentamicin-loaded PMMA bone cements in aprosthesis-related interfacial gap model and their antibacterial efficacy, BMCMusculoskelet. Disord. 11 (2010) 258.
[270] W.J. Habraken, J.G. Wolke, Injectable PLGA microsphere/calcium phosphate ce-ments: physical properties and degradation characteristics, J. Biomater. Sci.Polym. Ed. 17 (2006) 1057–1074.
[271] M.P. Ginebra, T. Traykova, J.A. Planell, Calcium phosphate cements: competitivedrug carriers for the musculoskeletal system? Biomaterials 27 (2006) 2171–2177.
[272] F. Wu, J. Su, J. Wei, H. Guo, C. Liu, Injectable bioactive calcium–magnesium phos-phate cement for bone regeneration, Biomed. Mater. 3 (2008) 44105.
[273] M. Bohner, U. Gbureck, J.E. Barralet, Technological issues for the development ofmore efficient calcium phosphate bone cements: a critical assessment, Biomate-rials 26 (2005) 6423–6429.
[274] A. Tofighi, S. Mounic, P. Chakravarthy, C. Rey, D. Lee, Setting reactions involvedin injectable cements based on amorphous calcium phosphate, Key Eng.Mater. 192–195 (2000) 769–772.
[275] B.R. Constantz, I.C. Ison, M.T. Fulmer, R.D. Poser, S.T. Smith, M. VanWagoner, J.Ross, S.A. Goldstein, J.B. Jupiter, D.I. Rosenthal, Skeletal repair by in situ forma-tion of the mineral phase of bone, Science 267 (1995) 1796–1799.
[276] M. Takechi, Y. Miyamoto, K. Ishikawa, M. Nagayama, M. Kon, K. Asaoka, K.Suzuki, Effects of added antibiotics on the basic properties of anti-washout-type fast-setting calcium phosphate cement, J. Biomed. Mater. Res. 39 (1998)308–316.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug

17Y. Wang, D.W. Grainger / Advanced Drug Delivery Reviews xxx (2011) xxx–xxx
[277] A. Ratier, I.R. Gibson, S.M. Best, M. Freche, J.L. Lacout, F. Rodriguez, Setting char-acteristics and mechanical behaviour of a calcium phosphate bone cement con-taining tetracycline, Biomaterials 22 (2001) 897–901.
[278] M.T. Ethell, R.A. Bennett, M.P. Brown, K. Merritt, J.S. Davidson, T. Tran, In vitroelution of gentamicin, amikacin, and ceftiofur from polymethylmethacrylateand hydroxyapatite cement, Vet. Surg. 29 (2000) 375–382.
[279] M. Takechi, Y. Miyamoto, Y. Momota, T. Yuasa, S. Tatehara, M. Nagayama, K. Ishi-kawa, K. Suzuki, The in vitro antibiotic release from anti-washout apatite ce-ment using chitosan, J. Mater. Sci. Mater. Med. 13 (2002) 973–978.
[280] M. Otsuka, Y. Matsuda, Y. Suwa, J.L. Fox, W.I. Higuchi, A novel skeletal drug de-livery system using a self-setting calcium phosphate cement. 5. Drug release be-havior from a heterogeneous drug-loaded cement containing an anticancerdrug, J. Pharm. Sci. 83 (1994) 1565–1568.
[281] M. Otsuka, Y. Matsuda, J.L. Fox, W.I. Higuchi, A novel skeletal drug delivery sys-tem using self-setting calcium phosphate cement. 9: effects of the mixing solu-tion volume on anticancer drug release from homogeneous drug-loadedcement, J. Pharm. Sci. 84 (1995) 733–736.
[282] E.J. Blom, J. Klein-Nulend, C.P. Klein, K. Kurashina, M.A. van Waas, E.H. Burger,Transforming growth factor-beta1 incorporated during setting in calcium phos-phate cement stimulates bone cell differentiation in vitro, J. Biomed. Mater. Res.50 (2000) 67–74.
[283] R.B. Edwards III, H.J. Seeherman, J.J. Bogdanske, J. Devitt, R. Vanderby, M.D. Mar-kee, Percutaneous injection of recombinant human bone morphogenetic
Please cite this article as: Y. Wang, D.W. Grainger, RNA therapeutics taDeliv. Rev. (2011), doi:10.1016/j.addr.2011.09.002
protein-2 in a calcium phosphate paste accelerates healing of a canine tibialosteotomy, J. Bone Joint Surg. Am. 86 (2004) 1425–1438.
[284] L. Dong, Z. Huang, X. Cai, J. Xiang, Y.A. Zhu, R. Wang, J. Chen, J. Zhang, Localizeddelivery of antisense oligonucleotides by cationic hydrogel suppresses TNF-alpha expression and endotoxin-induced osteolysis, Pharm. Res. 28 (2011)1349–1356.
[285] R.J. Christie, D.W. Grainger, Design strategies to improve soluble macromolecu-lar delivery constructs, Adv. Drug Deliv. Rev. 55 (2003) 421–437.
[286] J. Shen, R. Samul, R.L. Silva, H. Akiyama, H. Liu, Y. Saishin, S.F. Hackett, S. Zinnen,K. Kossen, K. Fosnaugh, C. Vargeese, A. Gomez, K. Bouhana, R. Aitchison, P. Pavco,P.A. Campochiaro, Suppression of ocular neovascularization with siRNA target-ing VEGF receptor 1, Gene Ther. 13 (2006) 225–234.
[287] L. Alexopoulou, A.C. Holt, R. Medzhitov, R.A. Flavell, Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3, Nature 413(2001) 732–738.
[288] M. Yamamoto, S. Sato, H. Hemmi, K. Hoshino, T. Kaisho, H. Sanjo, O. Takeuchi, M.Sugiyama, M. Okabe, K. Takeda, S. Akira, Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway, Science 301 (2003) 640–643.
[289] A. Aderem, Role of Toll-like receptors in inflammatory response in macrophages,Crit. Care Med. 29 (2001) S16–S18.
rgeting osteoclast-mediated excessive bone resorption , Adv. Drug
Related Documents











