MOLECULAR AND CELLULAR BIOLOGY, JUlY 1987, p. 2530-2537 Vol. 7, No. 7 0270-7306/87/072530-08$02.00/0 Copyright © 1987, American Society for Microbiology RNA Processing and Expression of an Intron-Encoded Protein in Yeast Mitochondria: Role of a Conserved Dodecamer Sequence HONG ZHU, IAN G. MACREADIE,t AND RONALD A. BUTOW* Department of Biochemistry, The University of Texas Health Science Center at Dallas, Dallas, Texas 75235 Received 4 February 1987/Accepted 18 April 1987 The 3' ends of most Saccharomyces cerevisiae mitochondrial mRNAs terminate at a conserved dodecamer sequence, 5'-AAUAAUAUUCUU-3', of unknown function. We have studied the consequences of mutations within a dodecamer found in an 1,143-base-pair optional intron of the mitochondrial large (21S) rRNA gene on RNA processing. The dodecamer is situated at the 3' end of an expressed open reading frame (ORF) within that intron, and the mutations are two adjacent transversions that extend the intron ORF by 51 nucleotides. The strain harboring these mutations, L5-10-1, is defective in biased intron transmission in crosses to strains that lack the intron, as are other mutants which contain nucleotide changes within the ORF (I. G. Macreadie, R. M. Scott, A. R. Zinn, and R. A. Butow, Cell 41:395-402, 1985). However, unlike these other mutants, wild-type strains, or petites which retain the intron allele, L5-10-1 is defective in processing at the intron dodecamer. In addition, L5-10-1 lacks a prominent 2.7-kilobase RNA containing both intron and exon sequences and at least two of four RNAs that correspond to various forms of the excised intron. We propose that these RNAs, missing in L5-10-1 but present in al other strains examined, arise in part by processing at the intron dodecamer. In addition, in all strains examined, we have detected a novel processing activity in which precursor 21S rRNA transcripts are cleaved in the upstream exon, about 1,500 nucleotides from the 5' end of the RNA. This activity, together with 3' intron dodecamer cleavage, probably accounts for the 2.7-kilobase RNA species, a candidate for the mRNA for the intron-encoded protein. Mature transcripts of the mitochondrial genome of Sac- charomyces cerevisiae are generated by diverse pathways. Some genes contain a highly conserved nonanucleotide sequence at their 5' end that functions as an initiator for mitochondrial RNA polymerase (7, 29). Other genes are cotranscribed into polycistronic precursors that are subse- quently processed to yield the mature RNAs (6, 9, 23, 37). At the 3' end of almost all yeast mitochondrial protein structural genes is a highly conserved dodecamer sequence, 5'- AATAATATTCTT-3', which is located either between genes within a single transcription unit or at the 3' end of a transcription unit. In either case, the putative mRNAs for these genes terminate within the dodecamer sequence, most likely as a result of processing (30, 33). The role of this 12-mer in mRNA activity or stability is unknown. A conserved dodecamer is also found within an optional 1,143-base-pair (bp) intron in the 21S rRNA gene but in an unusual location; it is at the 3' end of a freestanding open reading frame (ORF) (13) located entirely within the intron, and the termination codon for that ORF is part of the dodecamer sequence. The 21S rRNA intron is unique among optional mitochondrial introns, because in genetic crosses between strains that have it (wX) and those lacking it (w-), almost all of the w- alleles are converted to w+ (14, 40). Recent experiments show that expression of the intron ORF, which we have called fiti, is required for this conversion. First, in crosses between o+ and o- strains, the biased transmission of the w+ intron and its flanking markers occurs only when one or both of the parents are capable of mitochondrial protein synthesis; no conversion is apparent * Corresponding author. t Present address: Commonwealth Scientific and Industrial Re- search Organization Division of Protein Chemistry, Parkville, Victoria 3052, Australia. when crosses are carried out between petites whose mito- chondrial genomes contain these alleles (19, 41). Second, mutations within the fiti gene that would alter or block synthesis of the product also eliminate conversion (19, 24). Third, correlating precisely with these data is the presence of a double-strand break, which appears transiently during mating between + and o- strains at or near the intron insertion site in o- DNA (40, 41). A recent result shows, moreover, that the fit] gene, appropriately engineered for expression in Escherichia coli, expresses a double-strand endonuclease activity with specificity for cleavage of o- DNA (8). These data leave little doubt that the fiti gene is expressed and that its product is required for the conversion of o- to o . The presence of a functional protein-coding sequence within the w+ intron raises the question of how an mRNA is generated among the products of rRNA maturation. Forma- tion of the 21S rRNA in w+ strains involves a series of posttranscriptional events that include 3' processing of tran- scripts extending into downstream tRNA genes (23, 27), splicing (1, 2, 32), and cleavage within the conserved dodec- amer (30, 32). Since the fiti protein is encoded by a free- standing gene within an intron, its mRNA could be the excised intron, a derivative of it, or some RNA molecule containing both intron and exon sequences. These possibil- ities differ from the way in which most other introns in the yeast mitochondrial genome containing long ORFs are ex- pressed; these intron ORFs, many of which encode maturase proteins required for splicing, are in frame with their up- stream exon (21, 25, 35). Because of this arrangement, the splicing event destroys the message for that exon-intron fusion protein. Further, except for the fiti gene and intron aISP of the oxi3 gene, none of the other introns with ORFs contain a conserved dodecamer within the intron itself. 2530 on May 5, 2016 by guest http://mcb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MOLECULAR AND CELLULAR BIOLOGY, JUlY 1987, p. 2530-2537 Vol. 7, No. 70270-7306/87/072530-08$02.00/0Copyright © 1987, American Society for Microbiology
RNA Processing and Expression of an Intron-Encoded Protein inYeast Mitochondria: Role of a Conserved Dodecamer Sequence
HONG ZHU, IAN G. MACREADIE,t AND RONALD A. BUTOW*Department of Biochemistry, The University of Texas Health Science Center at Dallas, Dallas, Texas 75235
Received 4 February 1987/Accepted 18 April 1987
The 3' ends of most Saccharomyces cerevisiae mitochondrial mRNAs terminate at a conserved dodecamersequence, 5'-AAUAAUAUUCUU-3', of unknown function. We have studied the consequences of mutationswithin a dodecamer found in an 1,143-base-pair optional intron of the mitochondrial large (21S) rRNA gene onRNA processing. The dodecamer is situated at the 3' end of an expressed open reading frame (ORF) within thatintron, and the mutations are two adjacent transversions that extend the intron ORF by 51 nucleotides. Thestrain harboring these mutations, L5-10-1, is defective in biased intron transmission in crosses to strains thatlack the intron, as are other mutants which contain nucleotide changes within the ORF (I. G. Macreadie,R. M. Scott, A. R. Zinn, and R. A. Butow, Cell 41:395-402, 1985). However, unlike these other mutants,wild-type strains, or petites which retain the intron allele, L5-10-1 is defective in processing at the introndodecamer. In addition, L5-10-1 lacks a prominent 2.7-kilobase RNA containing both intron and exonsequences and at least two of four RNAs that correspond to various forms of the excised intron. We proposethat these RNAs, missing in L5-10-1 but present in al other strains examined, arise in part by processing atthe intron dodecamer. In addition, in all strains examined, we have detected a novel processing activity inwhich precursor 21S rRNA transcripts are cleaved in the upstream exon, about 1,500 nucleotides from the 5'end of the RNA. This activity, together with 3' intron dodecamer cleavage, probably accounts for the2.7-kilobase RNA species, a candidate for the mRNA for the intron-encoded protein.
Mature transcripts of the mitochondrial genome of Sac-charomyces cerevisiae are generated by diverse pathways.Some genes contain a highly conserved nonanucleotidesequence at their 5' end that functions as an initiator formitochondrial RNA polymerase (7, 29). Other genes arecotranscribed into polycistronic precursors that are subse-quently processed to yield the mature RNAs (6, 9, 23, 37). Atthe 3' end of almost all yeast mitochondrial protein structuralgenes is a highly conserved dodecamer sequence, 5'-AATAATATTCTT-3', which is located either betweengenes within a single transcription unit or at the 3' end of atranscription unit. In either case, the putative mRNAs forthese genes terminate within the dodecamer sequence, mostlikely as a result of processing (30, 33). The role of this12-mer in mRNA activity or stability is unknown.A conserved dodecamer is also found within an optional
1,143-base-pair (bp) intron in the 21S rRNA gene but in anunusual location; it is at the 3' end of a freestanding openreading frame (ORF) (13) located entirely within the intron,and the termination codon for that ORF is part of thedodecamer sequence. The 21S rRNA intron is unique amongoptional mitochondrial introns, because in genetic crossesbetween strains that have it (wX) and those lacking it (w-),almost all of the w- alleles are converted to w+ (14, 40).Recent experiments show that expression of the intron ORF,which we have called fiti, is required for this conversion.First, in crosses between o+ and o- strains, the biasedtransmission of the w+ intron and its flanking markers occursonly when one or both of the parents are capable ofmitochondrial protein synthesis; no conversion is apparent
* Corresponding author.t Present address: Commonwealth Scientific and Industrial Re-
search Organization Division of Protein Chemistry, Parkville,Victoria 3052, Australia.
when crosses are carried out between petites whose mito-chondrial genomes contain these alleles (19, 41). Second,mutations within the fiti gene that would alter or blocksynthesis of the product also eliminate conversion (19, 24).Third, correlating precisely with these data is the presence ofa double-strand break, which appears transiently duringmating between + and o- strains at or near the introninsertion site in o- DNA (40, 41). A recent result shows,moreover, that the fit] gene, appropriately engineered forexpression in Escherichia coli, expresses a double-strandendonuclease activity with specificity for cleavage of o-DNA (8). These data leave little doubt that the fiti gene isexpressed and that its product is required for the conversionof o- to o .The presence of a functional protein-coding sequence
within the w+ intron raises the question of how an mRNA isgenerated among the products of rRNA maturation. Forma-tion of the 21S rRNA in w+ strains involves a series ofposttranscriptional events that include 3' processing of tran-scripts extending into downstream tRNA genes (23, 27),splicing (1, 2, 32), and cleavage within the conserved dodec-amer (30, 32). Since the fiti protein is encoded by a free-standing gene within an intron, its mRNA could be theexcised intron, a derivative of it, or some RNA moleculecontaining both intron and exon sequences. These possibil-ities differ from the way in which most other introns in theyeast mitochondrial genome containing long ORFs are ex-pressed; these intron ORFs, many ofwhich encode maturaseproteins required for splicing, are in frame with their up-stream exon (21, 25, 35). Because of this arrangement, thesplicing event destroys the message for that exon-intronfusion protein. Further, except for the fiti gene and intronaISP of the oxi3 gene, none of the other introns with ORFscontain a conserved dodecamer within the intron itself.
2530
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

YEAST MITOCHONDRIAL CONSERVED DODECAMER SEQUENCE
In our collection of fitl gene mutants that fail to transmitthe w+ intron at high frequency or to support the cleavage ofw DNA in crosses (24), one was of particular interest, sinceit contained two base changes within the dodecamer motif.In this report, we analyze the consequences of these muta-tions on the pattern of processing of w+ 21S rRNA tran-scripts. In characterizing these processing pathways, wehave also uncovered a novel 5' exon cleavage that suggeststhe possibility of alternate forms of the fiti mRNA.
MATERIALS AND METHODS
Strains. The yeast strains used in this study are as follows:L5230 (a met tyr leu trp p+ w') and afiti mutant derivative,L5-10-1, as well as other fitl mutants of L5230 that havebeen characterized by Macreadie et al. (24), Fll (a his trp p-w), and IL8-8C (a his trp p+ w').
Isolation of mitochondrial RNA. Cells were grown at 28°Cto early stationary phase in YPD medium (1% yeast extract,1% Bacto-Peptone, 2% glucose), harvested by centrifugationin a Beckman JAlO rotor at 5,000 rpm for 5 min, andsuspended at 100 m/liter in buffer containing 100 mM Tris(pH 9.3), 20 mM EDTA, and 5% mercaptoethanol. Afterincubation at 28°C for 15 min, the cells were collected bycentrifugation and washed with a solution containing 0.6 Msorbitol, 0.3 M mannitol, 20 mM K2HPO4, 20 mM citric acid,and 1 mM EDTA (pH 5.8). The cells were then incubatedwith Zymolyase 20T at 28°C for 1 h to convert them tospheroplasts. The spheroplasts were washed in ice-coldbuffer A (0.6 M sorbitol, 150 mM KCl, 2 mM EDTA, 20 mMTris chloride [pH 7.4]) and then resuspended in 2.5 ml ofbuffer A per gram (wet weight) of cells. The spheroplastswere broken by two 30-s bursts of homogenization at highspeed in a Sorvall Omnimixer. After two centrifugations at4,000 rpm for 10 min each to remove debris, mitochondriawere pelleted from the supernatant by centrifugation at11,000 rpm for 10 min in a Beckman JA20 rotor. Themitochondrial pellets were resuspended in an Eppendorftube to a final concentration of 4 ml/liter of buffer Acontaining 32% (vol/vol) Percoll and were centrifuged in anEppendorf centrifuge for 20 min. The brown mitochondrialband was collected and diluted with buffer A. The solutionsin the following steps were all treated with diethylpyrocar-bonate. The mitochondria were pelleted and lysed in a
solution containing 0.3 M NaOAc, 10 mM EDTA, 50 mMTris chloride (pH 7.5), 2% sodium dodecyl sulfate, and 0.1%mercaptoethanol. The lysates were extracted three timeswith phenol, once with phenol-chloroform (1:1), once withchloroform, and three times with diethyl ether. RNA wasprecipitated by the addition of 3 volumes of ethanol at -20°Covernight and then precipitated twice more with ethanol at-70°C for 1 h from a solution containing 0.3 M NaOAc, 10mM Tris chloride (pH 7.4), and 1 mM EDTA.Northern blots. RNA pellets were suspended in electro-
phoresis buffer (40 mM Tris phosphate [pH 7.4], 36 mMNaH2PO4, 1 mM EDTA) containing 20o sucrose. RNAswere fractionated by electrophoresis through either 1.5%agarose-6 M urea gels at 3 V/cm for 18 h or 6% acrylamide-6M urea gels at 25 V/cm for 18 h. For both types of gels, 10 to20,ug of RNA was loaded in each lane. The gels were
equilibrated in three changes of lx TEA (40 mM Tris [pH7.4], 20 mM NaOAC, 1 mM EDTA [pH 7.4]) for 1 h beforeelectrophoretic transfer to a Zeta-Probe membrane. Transferwas carried out inlx TEA at4°C at 1.5 A for at least 4 h.After transfer, the membrane was air dried and then baked ina vacuum oven at 80°C for 2 h. The membrane was
prehybridized for 4 h at 42°C in a solution containing 30%formamide, 5x SCC (lx SSC is 0.15 M NaCl plus 0.015 Msodium citrate), 50 mM Na3PO4 (pH 6.5), 1.0% sodiumdodecyl sulfate, 0.1% Ficoll, 0.1% polyvinylpyrrolidone,0.1% bovine serum albumin, and 1 mg of calf thymus DNAper ml. The blots were then hybridized with radioactiveprobe at 105 cpm/ml of hybridization solution for 20 h at 42°Cand then washed four times at 42°C in lx SSC-1% sodiumdodecyl sulfate. Blots were exposed to Kodak X-Omat film.
S1 nuclease mapping. RNA was electroeluted from gels aspreviously described (36). The 588-bp HindIII-SalI and4.4-kb EcoRV fragments were end labeled with 32P by fillingin with Klenow fragment and T4 polynucleotide kinase,respectively. The 588-bp HindIII-SalI fragment was thendenatured by heating at 90°C for 2 min in a solution contain-ing 50 mM NaOH, 10 mM EDTA, and 10% sucrose. Afterquick cooling on ice, the denatured fragment was separatedby electrophoresis on a 5% acrylamide gel at 4°C. Theseparated strands were located by autoradiography andelectroeluted from the gel. The 4.4-kb EcoRV fragment wascut out from a preparative agarose gel, submerged in 0.3 MNaOH at room temperature for 30 min, rinsed with ice-coldwater, and then loaded into slots in a 1% agarose gel. Thestrands were separated by electrophoresis on agarose gels asdescribed by Maniatis et al. (26), located by staining withethidium bromide, and eluted as described above. Thetemplate strand was identified by dot-blot hybridization ofeach strand to total mitochondrial RNA. For S1 nucleaseanalysis, single-stranded probes and RNA were precipitatedtogether with ethanol. The precipitate was dried and dis-solved in 25 pl of Si hybridization solution (60% formamide,400 mM NaCl, 10 mM PIPES [piperazine-N,N'-bis(2-ethanesulfonic acid] [pH 6.4], 10 mM EDTA). The mixturewas first heated at 65°C for 5 min and transferred to 42°C.Hybridization was carried out at 42°C for 16 h. The hybrid-ization mixture was then diluted with 200,ul ofS1 reactionbuffer (280 mM NaCl, 30 mM NaOAc [pH 4.6], 4.5 mMZnOAc, 40,ug of heat-denatured calf thymus DNA per ml).Digestion was initiated by the addition of 200 U of S1nuclease and continued for 1 h at 37°C. Reactions wereterminated by the addition of 25 pAl of 2.7 M NH4OAc-0.05 MEDTA, and the nucleic acids were precipitated with 3volumes of ethanol. The resultant pellet was washed with70% ethanol and taken up with formamide plus 0.03%(wt/vol) bromophenol blue, 0.03% xylene cyanol, 1 mMEDTA, and 5% H20 (vol/vol). Samples were heated at 90°Cfor 10 min and chilled on ice. Analysis of protected frag-ments was carried out by electrophoresis on 5 or 7%acrylamide-8 M urea gels, which were then dried andexposed to Kodak X-Omat film.
RESULTS
Dodecamer mutations. By mutagenizing mating mixturesofw+ and w- cells for short periods of time with Mn2+, wewere able to direct mutations to the fit] gene, probablybecause of the active replicative conversion of w- to w+ (24).Most of these mutations were combinations of frameshiftand missense changes, and in crosses to w- strains, allresulted in the elimination of the biased output of the w+allele. One mutant we obtained, called L5-10-1, also failed toshow a bias in intron transmission, although no base changeswere found within thefitl-coding region; instead, there weretwo adjacent transversions at the 3' end of the gene whichchanged the wild-type TAA stop codon to ATA, a codonwhich is translated as methionine in yeast mitochondria (17).
VOL. 7, 1987 2531
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

2532 ZHU ET AL.
t
Hindm Hindm
EcoR3E ATG s 1 ntAATAATATTCTT------TAA
A1AT (L5-10-1)
I I I I *
0.4 Q8 1.2 1.6 2.0 2.4 2.8 3.2 3.6 4.0 44Kiloboses
FIG. 1. Map of w4 rRNA gene showing location of conserveddodecamer and mutations within that sequence and upstream exonprocessing site (arrow). The 1,143-bp intron is indicated by the box,and the coding sequence of ftl] is shown by the hatched area. nt,Nucleotide.
If the fiti gene in L5-10-1 were translated, the resultantprotein would be extended at its carboxyl terminus by 17amino acids. These mutations were located within the con-served dodecarner (Fig. 1).21S rRNA transcripts in wild-type and fit] mutants. To
determine if any of the mutants affect processing of 21SrRNA precursors, we first compared the amounts of mature21S rRNA in different fiti mutants and in the wild type by
3co
J0 J0
VI'
ID r- if _to, 0 ff) toa) ro 0 CD (0
i -I -I
21S rRNA-_ _w _ I w
EXON
PROBE
kb
44 -_ _* -*I
2.7 --
1.1 S _
_ _ _ _ _0v....
INTRONPROBE
AL Ia ,S1
FIG. 2. 21S rRNA transcripts in wild-type (W.T.) and fit] mu-tants. Mitochondrial RNA was isolated from each of the strainsshown and fractionated on 1.5% agarose-6 M urea gels. Afterelectrophoretic transfer to Zeta-Probe, one blot (top) was hybridizedwith a 3.3-kp HpaII DNA fragment corresponding to the entire 21SrRNA gene from an w- strain; the other blot (bottom) was hybrid-ized with a 495-bp HindlIl fragment located entirely within the1.1-kb intron (see Fig. 1). Each lane contains 15 ,ug of totalmitochondrial RNA.
FIG. 3. w+ intron RNAs in p- Fll. Northern analysis wascarried out as described in the legend to Fig. 2 on total mitochondrialRNA from L5230, L5-10-1, and Fil. The 495-bp HindIII intronfragment was used as the probe.
Northern blot analysis. All of the mutant strains containingbase changes within thefiti gene, including L5-10-1, mnade amature 21S rRNA in amounts comparable to that of the wildtype (Fig. 2, top). This result is not surprising because, first,all of the mutants can grow on nonfermentable carbonsources, and second, expression of the fiti gene is notrequired for splicing of the 1,143-bp intron in w+ strains.Moreover, we can also conclude that the mutations do notaffect splicing in some indirect way. When the same RNAblots were probed with a 495-bp HindIII intron probe (Fig. 2,bottom), a spectrum of intron-containing RNAs was ob-served, ranging in size from a 4.4-kb transcript containing allof the 21S rRNA exon and intron sequences to transcripts inthe 1.1-kb region of the gel. The overall pattern of intron-containing transcripts is similar to those described by Bos etal. (2). Of all of the RNA species detected with the intronprobe, the one consistent difference we have noted amongstrains when RNAs are analyzed on agarose-urea gels is theabsence of a band in L5-10-1 with an apparent size of 2.7 kb;this RNA was present in the wild type and at comparablelevels in all of the other fiti mutants we have examined.Hence, the absence of the 2.7-kb RNA in L5-10-1 is notlikely to be due simply to the inability of this strain to makea functional fiti protein, since other mutants that have thisRNA contain termination and missense mutations within thefiti reading frame (24). In fact, production of the 2.7-kb RNAdid not require any mitochondrial protein synthesis, sincethat RNA was found in a petite strain (Fil) whose mitochon-drial genome has the entire w+ 21S rRNA gene (Fig. 3, andbelow). Because restoration of biased + intron transmis-sion and double-strand cleavage activity can be completelyrestored to L5-10-1 in crosses with petites whose genomesretain just the wild-type + allele (24), it is also unlikely thatthe L5-10-1 phenotype is due to mutations outside of the 21SrRNA region.
3' dodecamer processing. Previous Si nuclease protectionexperiments established that the 1,143-bp w+ intron can becleaved at the conserved dodecamer (30, 32). Thus, onepossible consequence of the two base changes in L5-10-1 isan alteration in dodecamer processing. Putting aside for themoment whether altered processing could account for theinability of this mutant to effect intron transmission incrosses, the most obvious difference between L5-10-1 andthe other fiti mutants is the absence of the 2.7-kb RNAspecies seen on 1.5% agarose-urea gels. Tabak et al. (32)suggested alternative processing pathways for the excised
o
CJ I-LO Ln- r
_j Lj 1
kb
-2.7
MOL. CELL. BIOL.
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from
YEAST MITOCHONDRIAL CONSERVED DODJtCAMER SEQUENCE
+ RNA4 SI
-RNA L5-10-1 L5230I I+Sl -SI 2.7 1.1 2.7 1.1
__b
v 9
- 622
- 527
-407
-309
Hind m
5 '{ '3
!581525385
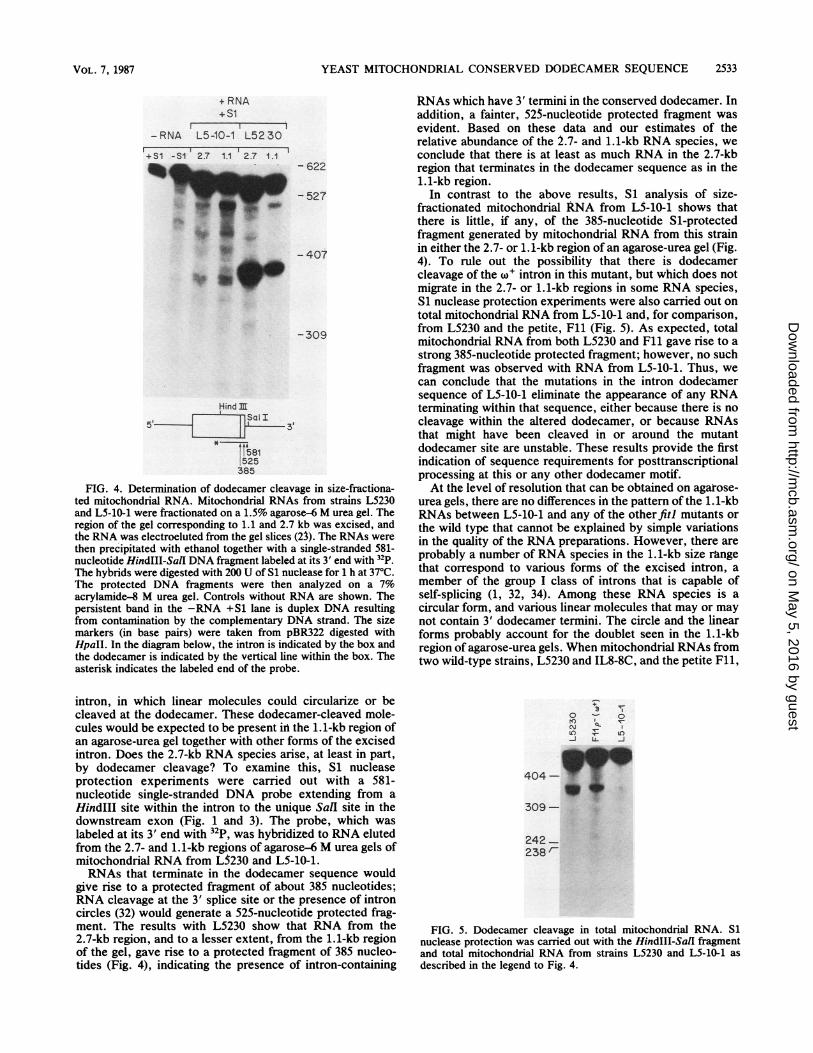
FIG. 4. Determination of dodecamer cleavage in size-fractiona-ted mitochondrial RNA. Mitochondrial RNAs from strains L5230and L5-10-1 were fractionated on a 1.5% agarose-6 M urea gel. Theregion of the gel corresponding to 1.1 and 2.7 kb was excised, andthe RNA was electroeluted from the gel slices (23). The RNAs werethen precipitated with ethanol together with a single-stranded 581-nucleotide HindIII-SalI DNA fragment labeled at its 3' end with 32p.The hybrids were digested with 200 U of Si nuclease for 1 h at 37°C.The protected DNA fragments were then analyzed on a 7%acrylamide-8 M urea gel. Controls without RNA are shown. Thepersistent band in the -RNA +S1 lane is duplex DNA resultingfrom contamination by the complementary DNA strand. The sizemarkers (in base pairs) were taken from pBR322 digested withHpaII. In the diagram below, the intron is indicated by the box andthe dodecamer is indicated by the vertical line within the box. Theasterisk indicates the labeled end of the probe.
intron, in which linear molecules could circularize or becleaved at the dodecamer. These dodecamer-cleaved mole-cules would be expected to be present in the 1.1-kb region ofan agarose-urea gel together with other forms of the excisedintron. Does the 2.7-kb RNA species arise, at least in part,by dodecamer cleavage? To examine this, Si nucleaseprotection experiments were carried out with a 581-nucleotide single-stranded DNA probe extending from aHindIII site within the intron to the unique Sall site in thedownstream exon (Fig. 1 and 3). The probe, which waslabeled at its 3' end with 32p, was hybridized to RNA elutedfrom the 2.7- and 1.1-kb regions of agarose-6 M urea gels ofmitochondrial RNA from L5230 and L5-10-1.RNAs that terminate in the dodecamer sequence would
give rise to a protected fragment of about 385 nucleotides;RNA cleavage at the 3' splice site or the presence of introncircles (32) would generate a 525-nucleotide protected frag-ment. The results with L5230 show that RNA from the2.7-kb region, and to a lesser extent, from the 1.1-kb regionof the gel, gave rise to a protected fragment of 385 nucleo-tides (Fig. 4), indicating the presence of intron-containing
RNAs which have 3' termini in the conserved dodecamer. Inaddition, a fainter, 525-nucleotide protected fragment wasevident. Based on these data and our estimates of therelative abundance of the 2.7- and 1.1-kb RNA species, weconclude that there is at least as much RNA in the 2.7-kbregion that terminates in the dodecamer sequence as in the1.1-kb region.
In contrast to the above results, Si analysis of size-fractionated mitochondrial RNA from L5-10-1 shows thatthere is little, if any, of the 385-nucleotide Sl-protectedfragment generated by mitochondrial RNA from this strainin either the 2.7- or 1.1-kb region of an agarose-urea gel (Fig.4). To rule out the possibility that there is dodecamercleavage of the w+ intron in this mutant, but which does notmigrate in the 2.7- or 1.1-kb regions in some RNA species,Si nuclease protection experiments were also carried out ontotal mitochondrial RNA from L5-10-1 and, for comparison,from L5230 and the petite, Fll (Fig. 5). As expected, totalmitochondrial RNA from both L5230 and Fll gave rise to astrong 385-nucleotide protected fragment; however, no suchfragment was observed with RNA from L5-10-1. Thus, wecan conclude that the mutations in the intron dodecamersequence of L5-10-1 eliminate the appearance of any RNAterminating within that sequence, either because there is nocleavage within the altered dodecamer, or because RNAsthat might have been cleaved in or around the mutantdodecamer site are unstable. These results provide the firstindication of sequence requirements for posttranscriptionalprocessing at this or any other dodecamer motif.At the level of resolution that can be obtained on agarose-
urea gels, there are no differences in the pattern of the 1.1-kbRNAs between L5-10-1 and any of the otherfitl mutants orthe wild type that cannot be explained by simple variationsin the quality of the RNA preparations. However, there areprobably a number of RNA species in the 1.1-kb size rangethat correspond to various forms of the excised intron, amember of the group I class of introns that is capable ofself-splicing (1, 32, 34). Among these RNA species is acircular form, and various linear molecules that may or maynot contain 3' dodecamer termini. The circle and the linearforms probably account for the doublet seen in the 1.1-kbregion of agarose-urea gels. When mitochondrial RNAs fromtwo wild-type strains, L5230 and IL8-8C, and the petite Fl,
3 i
o o'- o
-J
4044-
309-
242_238r
FIG. 5. Dodecamer cleavage in total mitochondrial RNA. 51nuclease protection was carred out with the HindIII-SalI fragmentand total mitochondrial RNA from strains L5230 and L5-10-1 asdescribed in the legend to Fig. 4.
VOL. 7, 1987 2533
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

2534 ZHU ET AL.
were fractionated on a 6% acrylamide gel, four distinctintron RNA species were detected in the 1.1-kb region (Fig.6). In contrast, at least two of these RNAs were absent inL5-10-1. These missing species are good candidates for thoselinear forms of the excised intron that have been cleaved atthe conserved dodecamer, but additional experiments will berequired to prove this point.
5' exon processing. If the 3' end of the 2.7-kb transcriptterminates within the dodecamer, then its 5' end should fallwithin the upstream exon, roughly 1.5 kb from the start oftranscription. Our initial attempts to locate the putative 5'cleavage site by Si nuclease protection experiments withupstream exon probes were inconclusive, probably becausethe vast excess of the mature 21S rRNA and its degradationproducts obscured the signal. To obviate this problem, wehave attempted to map the 5' exon cleavage site by S1nuclease protection with a 4.4-kb EcoRV probe derived froma cloned 21S rRNA allele of the p+ parent of the petite Fll.The 5' labeled end of this probe fell within the intron andextended upstream into the 5' flanking region of the 21SrRNA gene (Fig. 1); hence, only those RNAs containingboth intron and exon sequences would be detected. Theprobe was hybridized to total mitochondrial RNA fromL5-10-1 and Fll, as well as to size-fractionated RNA fromFll. We used the petite strain in these experiments ratherthan L5230, first, because that wild-type strain containssequence polymorphisms in the upstream exon which differfrom those of L5-10-1 (resulting in spurious signals in the Siprotection analysis [Hong Zhu, unpublished observations]),and second, because the 2.7-kb RNA species is relativelymore abundant in the petite than in wild-type strains. Theresults of the Si analysis show a major 1.23-kb protectedfragment in all strains examined. (Fig. 7.) This fragment wassome 500 nucleotides shorter than the size predicted on thebasis of our estimate of 2.7 kb for the RNA species missingin L5-10-1. However, since yeast mitochondrial RNAs shownotoriously anomalous migration behavior on agarose-ureagels (22), this apparent discrepancy is not surprising. Thevirtual identity between L5-10-1 and Fli in the pattern ofEcoRV-protected fragments suggests that 5' exon processingis the same as that in cells which process RNAs at thewild-type intron dodecamer.
DISCUSSION
The present work establishes the importance of sequenceconservation for RNA processing within the conserveddodecamer motif, 5'-AAUAAUAUUCUU-3'. This se-quence is found at the 3' end of most protein structural geneson the yeast mitochondrial genome, including two unas-
o u
LC)
kb LL H
0
-j
1.07-1.0-
FIG. 6. Northern analysis of excised + intron RNAs. Mito-chondrial RNAs from Fll, L5230, IL8-8C, and L5-lO-1 werefractionated on a 6% acrylamide-8 M urea gel and transferred toZeta-Probe. The blot was hybridized with the 495-bp HindIII intronfragment.
0
LC)
F11p-w+ J - RNA
kb 27 T T +SI -Si
44-2.8 -
1.35-
2. -
I A..
aI.
:
s...
0.82 -
LcoRVEcoR c
5 3
FIG. 7. 5' exon processing in total and size-fractionated mito-chondrial RNA. Sl nuclease protection was carried out as describedin the legend to Fig. 3 on total (T) and 2.7-kb size-fractionatedmitochondrial RNA from strains Fll and L5-10-1. The DNA probewas a single-stranded 4.4-kb EcoRV fragment (Fig. 1) labeled at its5' end with 32p. The protected DNA products were analyzed on a5% acrylamide-8 M urea gel. The size markers (in kilobase pairs)were taken from X DNA digested with HindIII and 4X174 digestedwith HaeIII.
signed reading frames and two reading frames within introns(30, 33, 37). One of those intron reading frames is fit), thefreestanding gene within the 1,143-bp intron of the w+ alleleof the 21S rRNA gene. We have confirmed previous obser-vations that wild-type cells and petites which retain the +'allele can process w+ 21S rRNA transcripts at the introndodecamer site (30, 32).The fitl mutant, L5-10-1, is the first characterized strain
containing mutations of a dodecamer sequence. These mu-tations consist of two adjacent base changes that convert thefit) TAA stop codon, located within the dodecamer, to ATA.This results in an extension of the fit) reading frame by 17amino acids. Like other mutants we have studied containingframeshift and base changes within this gene (24), L5-10-1 isunable to support --to-w+ conversion in crosses and dou-ble-strand cleavage activity ofw- DNA in zygotes. The mostsignificant difference between L5-10-1 and the other fit)mutants, however, is that this mutant is defective in dodec-amer processing. Since the otherfitl mutants and the petitestrains whose mitochondrial genome contains the entire w+21S rRNA gene can still process RNAs at the dodecamer, itis clear that expression of the intron-encoded protein cannotbe required for this activity.The unique absence of the 2.7-kb RNA in L5-10-1 suggests
that this RNA species arises, at least in part, from processingat the intron dodecamer. Support for this conclusion comesfrom S1 nuclease protection experiments with size-frac-tionated mitochondrial RNA in which we find that in wild-type cells, the abundance of dodecamer-processed RNA inthe 2.7-kb region is at least as great as that in the 1.1-kb RNAfraction. Tabak et al. (32) showed that the RNAs in the
MOL. CELL. BIOL.
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

YEAST MITOCHONDRIAL CONSERVED DODECAMER SEQUENCE
1.1-kb region contain various forms of the excised w+ intron.From Si nuclease protection experiments, they suggestedthat among these forms was a linear species with a 3'dodecamer terminus, although they were unable to detectthe predicted downstream fragment with a 5' dodecamerend. Possibly then, the RNAs with dodecamer termini foundin the 1.1-kb fraction might be due entirely to the degrada-tion of larger RNA molecules, for example, from the 2.7-kbsize class. However, the absence in L5-10-1 of at least two offour 1.1-kb intron forms present in the wild type and in an wpetite would argue against this interpretation, since splicingappears normal in this mutant, and the only processingdefect we detect is the absence of dodecamer cleavage. Wewould therefore conclude that for wild-type w+ alleles,dodecamer processing can occur before, as well as after,intron excision.We have also identified an unusual processing event in
which precursor 21S rRNA transcripts containing theunspliced intron can be cleaved within the upstream exon,about 1,500 nucleotides from the 5' end of the 21S rRNA.This activity, together with 3' dodecamer processing, mostlikely accounts for the 2.7-kb RNA seen in all w+ cellsexcept L5-10-1. That specific 5' exon cleavage is still de-tected in L5-10-1 indicates that this novel activity is inde-pendent of dodecamer processing. It also suggests that theabsence of the 2.7-kb RNA in L5-10-1 is not due to theinstability of some RNA with the same 5' end as the 2.7-kbspecies but which lacks a 3' dodecamer terminus.RNA processing represents an important control of mito-
chondrial gene expression in yeast, accounting in one way oranother for the generation of virtually all mRNAs. Theintegrity of 5' nontranslated regions for the expression ofcertain mRNAs has now been established. For example, theputative varl mRNA, a 16S RNA species, is derived by 5'processing from a 19S precursor (15, 37). Both RNAs havethe same 3' dodecamer terminus, but the 19S RNA, whichaccumulates in certain varl mutants, is not translated (15,38). A number of nuclear genes have now been identifiedwhose products appear to be targeted to the 5' end ofmitochondrial RNAs, in one case, for proper 5' end proc-essing of cytochrome b transcripts (11), and in other cases,for translation of cytochrome b (12) and cytochrome oxidasesubunit III (5, 10, 28) mRNAs. The products of these nucleargenes are believed to interact in some way with specific 5'sequences, but detailed mechanisms of how they functionhave not yet been established.By contrast, the importance of 3' flanking sequences, and
in particular the conserved dodecamer, in the control ofmitochondrial gene espression, has not yet been determined.The most obvious possibilities for the function of a 3'dodecamer terminus are message stability, translatability, orboth. These would be similar to functions that have beensuggested for 3' polyadenylation of eukaryotic cytoplasmicmRNAs (3), although presently, polyadenylation seems tocorrelate best with message stability (18, 39). It is nowapparent that polyadenylation is only one of several struc-tural features that affect message stability (4). For example,some mRNAs may be protected from 3' exonucleolyticattack by stem-loop structure (16). In others, the presence ofAU-rich sequences in the 3' nontranslated region appears todestabilize the message (31). It is therefore plausible that a 3'dodecamer terminus could also affect the stability of yeastmitochondrial mRNAs. However, it is also possible that thedodecamer plays no role in either message stability ortranslatability. In either case, an alternative explanation forthe inability of L5-10-1 to support biased intron transmission
AAUAAUAUUCUU
, [}
5' cleavage assembly withL subunit proteins
21S rRNA
03' cleavage
1.1-kb intron
{Fm 2.7 kb
FIG. 8. Model for alternate processing of a mitochondrial largerRNA precursor from a wild-type w' strain. The top of the figure isa precursor w+ large rRNA transcript showing the 1.1-kb intron(open box), the intron ORF (hatched area), the location of thedodecamer sequence, and the upstream exon cleavage site. Theorder of 3' and 5' processing as shown has not been established.
is that because of the 17-amino-acid C-terminal extension,the fit] protein in that strain is nonfunctional or unstable.Inspection of the extended sequence in L5-10-1 shows thatsuch a protein would have an extremely hydrophobic C-terminal tail, and this could affect its activity or conceivablycreate a membrane anchor which removes the protein fromits site of action. Presently, distinguishing among thesevarious possibilities for the fit] dodecamer is particularlydifficult because it has not been possible to detect the fit)protein directly, even with very sensitive labeling and gelprocedures (I. Macreadie, unpublished observations). Asimilar problem has been encountered in the detection ofother intron-encoded proteins (maturases) in wild-type yeastmitochondria. However, there is a reasonable explanationfor why the abundance of maturases would be low; unlikethe fit) protein, which is encoded by a freestanding gene,maturases are produced by translation of in-frame exon-intron fusions (21, 25, 35), and thus the functional maturasemRNA is continuously being destroyed by splicing. Anadditional problem, although not peculiar to fit], is the lackof a reliable system for the in vitro translation of definedmitochondrial mRNAs.The 2.7-kb RNA, which we suggest is generated by 5'
cleavage of 21S precursors in the upstream exon and cleav-age within the intron at the conserved dodecamer, accountsquantitatively for at least as much dodecamer-cleaved RNAas the excised intron species do. If, in a wild-type back-ground, a determinant of message function is the presence ofa 3' dodecamer terminus, the 2.7-kb species or some form ofthe excised, dodecamer-cleaved intron could serve as thefitlmRNA. One plausible model for the expression of the fit)gene is depicted in Fig. 8. An unspliced 21S rRNA precursorin w+ strains could be processed both at the dodecamer andin the upstream exon, releasing the 2.7-kb RNA. The deci-sion whether to splice or to produce the 2.7-kb species wouldbe determined by the accessibility of the substrate forsplicing or processing, that is, whether the precursor isassembled into a putative preribosomal subunit or is proc-essed at the dodecamer and upstream exon sites. Althoughthere is no direct evidence that in yeast, splicing of w+transcripts occurs in a complex between ribosomal proteinsand precursor RNA, we have indicated this route by analogy
VOL. 7, 1987 2535
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

2536 ZHU ET AL.
with large rRNA splicing in Neurospora mitochondria (20).Whether such a pathway represents some form of control ofexpression of the fiti gene remains to be established.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grantsGM22525 and GM35510 from the National Institutes of Health anda grant from The Robert A. Welch Foundation.We thank Marie Rotondi for her help in the preparation of the
manuscript.
LITERATURE CITED1. Arnberg, A. C., G. Van der Horst, and H. F. Tabak. 1986.
Formation of lariats and circles and self-splicing of the precur-sor to the large ribosomal RNA of yeast mitochondria. Cell44:235-242.
2. Bos, J. L., K. A. Osinga, G. Van der Horst, N. G. Hecht, H. F.Tabak, G. B. Van Ommen, and P. Borst. 1980. Splice pointsequence and transcripts of the intervening sequence in themitochondrial 21S ribosomal RNA gene of yeast. Cell 20:207-214.
3. Brawerman, G. 1981. The role of the poly(A) sequence inmRNA. Crit. Rev. Biochem. 10:1-38.
4. Brawerman, G. '1987. Determinants of messenger RNA stabil-ity. Cell 48:5-6.
5. Cabral, F., and G. Schatz. 1978. Identification of cytochrome coxidase subunits in nuclear yeast mutants lacking the functionalenzyme. J. Biol. Chem. 253:4396-4401.
6. Christianson, T., J. C. Edward, D. M. Mueller, and M.Rabinowitz. 1983. Identification of a single transcriptional initi-ation site for the glutamic tRNA and COB genes in yeastmitochondria. Proc. Natl. Acad. Sci. USA 80:5564-5568.
7. Christianson, T., and M. Rabinowitz. 1983. Identification ofmultiple transcriptional initiation sites on the yeast mitochon-drial genome by in vitro capping with guanylyltransferase. J.Biol. Chem. 258: 14025-14033.
8. Colleaux, L., L. d'Auriol, M. Betermier, G. Cottarel, A.Jacquier, F. Galibert, and B. Dujon. 1986. Universal codeequivalent of a yeast mitochondrial intron reading frame isexpressed into E. coli as a specific double-strand endonuclease.Cell 44:521-533.
9. Coruzzi, G., S. G. Bonitz, B. E. Thalenfeld, and A. Tzagoloff.1981. Assembly of the mitochondrial membrane system. Anal-ysis of the nucleotide sequence and transcripts in the oxil regionof yeast mitochondrial DNA. J. Biol. Chem. 256:12780-12787.
10. Costanzo, M. C., and T. D. Fox. 1986. Product of Sac-charomyces cerevisiae nuclear gene PET494 activates transla-tion of a specific mitochondrial mRNA. Mol. Cell. Biol.6:3694-3703.
11. Diekmann, C. L., T. J. Koerner, and A. Tzagoloff. 1984.Assembly of the mitochondrial membrane system. CBP6, ayeast nuclear gene involved in 5' end processing of cytochromeb pre-mRNA. J. Biol. Chem. 259:4722-4731.
12. Diekmann, C. L., and A. Tzagoloff. 1985. Assembly of themitochondrial membrane system. CBP6, a yeast nuclear genenecessary for synthesis of cytochrome b. J. Biol. Chem.260:1513-1520.
13. Dujon, B. 1980. Sequence of the intron and flanking exons of themitochondrial 21S rRNA gene of yeast strains having differentalleles at the X and rib-1 loci. Cell 20:185-197.
14. Dujon, B., P. P. Slonimski, and L. Weill. 1974. Mitochondrialgenetics. IX. A model for recombination and segregation ofmitochondrial genomes in Saccharomyces cerevisiae. Genetics78:415-437.
15. Farreily, F., H. P. Zassenhaus, and R. A. Butow. 1982. Charac-terization of transcripts from the varl region on mitochondrialDNA of Saccharomyces cerevisiae. J. Biol. Chem. 257:6581-6587.
16. Georgiev, O., and M. L. Birnstiel. 1985. The conservedCAAGAAAGA spacer sequence is an essential element for the
formation of 3' termini of the sea urchin H3 histone mRNA byRNA processing. EMBO J. 4:481-489.
17. Hudspeth, M. E. S., W. M. Ainley, D. S. Shummard, R. A.Butow, and L. I. Grossman. 1982. Location and structure of thevarl gene on yeast mitochondrial DNA: nucleotide sequence ofthe 40.0 allele. Cell 30:617-626.
18. Huez, G., C. Bruck, and Y. Cleuter. 1981. Translational stabilityof native and deadenylylated rabbit globin mRNA injected intoHeLa cells. Proc. Natl. Acad. Sci. USA 78:908-911.
19. Jacquier, A., and B. Dujon. 1985. An intron-encoded protein isactive in a gene conversion process that spreads an intron intoa mitochondrial gene. Cell 41:383-394.
20. Lapolia, R. J., and A. M. Lambowitz. 1979. Binding of mito-chondrial ribosomal proteins to a mitochondrial ribosomal pre-cursor RNA containing a 2.3 kilobase intron. J. Biol. Chem.254:11746-11750.
21. Lazowska, J., C. Jacq, and P. P. Slonimski. 1980. Sequence ofintrons and flanking exons in wild-type and box3 mutants ofcytochrome b reveals an interlaced splicing protein. Cell22:333-348.
22. Locker, J. 1979. Analytical and preparative electrophoresis ofRNA in agarose-urea. Anal. Biochem. 98:358-367.
23. Locker, J., and M. Rabinowitz. 1981. Transcription in yeastmitochondria: analysis of the 21S rRNA region and its tran-scripts. Plasmid 6:302-314.
24. Macreadie, I. G., R. M. Scott, A. R. Zinn, and R. A. Butow.1985. Transposition of an intron in yeast mitochondrial requiresa protein encoded by that intron. Cell 41:395-402.
25. Mahler, H. R., D. K. Hanson, M. R. Lamb, P. S. Perlman, P. Q.Anziano, K. R. Glaus, and M. L. Haldi. 1982. Regulatory inter-actions between mitochondrial genomes: expressed introns-their function and regulation, p. 185-199. In P. Slonimski, P.Borst, and G. Attardi, (ed), Mitochondrial genes. Cold SpringHarbor Laboratory, Cold Spring Harbor, New York.
26. Maniatis, T., E. F. Fritsch, and J. F. Sambrook. 1983. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, New York.
27. Merten, S., R. M. Synenki, J. Locker, T. Christianson, and M.Rabinowitz. 1980. Processing of precursors of 21S ribosomalRNA from yeast mitochondria. Proc. Natl. Acad. Sci. USA77:1417-1421.
28. Muller, P. P., M. K. Reif, S. Zonghou, C. Sengstag, T. L.Mason, and T. D. Fox. 1984. A nuclear mutation that post-transcriptionally blocks accumulation of a yeast mitochondrialgene product can be suppressed by a mitochondrial gene rear-rangement. J. Mol. Biol. 175:431-452.
29. Osinga, K. A., M. De Haan, T. Christianson, and H. F. Tabak.1982. A nonanucleotide sequence involved in promotion ofribosomal RNA synthesis and RNA priming ofDNA replicationin yeast mitochondria. Nucleic Acids Res. 10:7993-8007.
30. Osinga, K. A., E. De Vries, G. Van der Horst, and H. F. Tabak.1984. Processing of yeast mitochondrial messenger RNAs at aconserved dodecamer sequence. EMBO J 3:829-834.
31. Shaw, G., and R. Kamen. 1986. A conserved AU sequence fromthe 3' untranslated region of GM-CSF mRNA mediates selec-tive mRNA degradation. Cell 46:659-667.
32. Tabak, H. F., G. Van der Horst, K. A. Osinga, and A. C.Arnberg. 1984. Splicing of large ribosomal precursor RNA andprocessing of intron RNA in yeast mitochondria. Cell39:623-629.
33. Thalenfeld, B. E., S. G. Bonitz, F. G. Nobrega, G. Macino, andA. Tzagoloff. 1983. olil transcripts in wild-type and in a cyto-plasmic "petite" mutant of yeast. J. Biol. Chem.258:14065-14068.
34. Van der Horst, G., and H. F. Tabak. 1985. Self-splicing of yeastmitochondrial ribosomal and messenger RNA precursors. Cell40:759-766.
35. Weiss-Brummer, B., G. Rodel, R. J. Schweyen, and F.Kaudewitz. 1982. Expression of the split gene cob in yeast:evidence for a precursor of a "maturase" protein translatedfrom intron 4 and preceding exons. Cell 29:527-536.
36. Zassenhaus, H. P., R. A. Butow, and Y. P. Hannon. 1982. Rapidelectroelution of nucleic acids from agarose and acrylamide
MOL. CELL. BIOL.
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from

YEAST MITOCHONDRIAL CONSERVED DODECAMER SEQUENCE
gels. Anal. Biochem. 125:125-130.37. snhaus, H. P., N. C. Martin, and R. A. Butow. 1984. Origins
of transcripts of the yeast mitochondrial varl gene. J. Biol.Chem. 259:6019-6027.
38. Zassenhaus, H. P., and P. S. Perlman. 1982. Respiration-deficient mutants in the A+T-rich region on yeast mitochondrialDNA containing the varl gene. Current Genet. 6:179-188.
39. Zeevi, H., J. R. Nevins, and J. E. Darneil, Jr. 1983. Newlyformed mRNA lacking polyadenylic acid enters the cytoplasm
and the polyribosomes but has a short half-life in the absence ofpolyadenylic acid. Mol. Cell. Biol. 2:517-525.
40. Zinn, A. R., and R. A. Butow. 1984. Kinetics and intermediatesof yeast mitochondrial DNA recombination. Cold Spring Har-bor Symp. Quant. Biol. 49:115-121.
41. Zinn, A. R., and R. A. Butow. 1985. Nonreciprocal exchangebetween alleles of the yeast mitochondrial 21S rRNA gene:kinetics and the involvement of a double-strand break. Cell40:887-895.
VOL. 7, 1987 2537
on May 5, 2016 by guest
http://mcb.asm
.org/D
ownloaded from
Related Documents











