Resonancia Magnética Nuclear 5.9 RMN Bidimensional Un experimento multipulso contiene etapas de preparación, evolución y detección. Los espectros que se obtiene en estos experimentos dependen de la naturaleza de la preparación y de la longitud del periodo de evolución. Lo que ocurre durante el período de evolución no es detectado directamente, pero puede estimarse realizando múltiples experimentos con periodos de evolución de diferente duración. De esta forma puede obtenerse información sobre dos conjuntos separados de parámetros espectrales: aquel que influye sobre la magnetización durante el periodo de evolución y el que actúa durante la adquisición propiamente dicha. En RMN-2D se utiliza una segunda transfomada de Fourier para convertir la dependencia temporal durante la evolución en una segunda frecuencia, lo que permite diseñar experimentos de diversa índole, muy ricos en información. La idea original sobre RMN-Bidimensional (RMN-2D) se debe a Jeener (1971), aunque fue llevada a la práctica por Ernst y colaboradores en 1976. La RMN-2D corresponde a la generación de espectros de RMN con dos frecuencias como variables independientes. Estos espectros RMN-2D se generan registrando FID (tB 2 B) al final de una secuencia de pulsos con variación sistemática de otro parámetro, más comúnmente el tiempo de evolución tB 1 B entre un par de pulsos. Los FIDs sucesivos se adquieren por incremento de los tiempos de evolución y se almacenan como filas en una matriz de datos, tal como se muestra en la Figura 5.121, donde se comparan las técnicas 1D y 2D. El espectro 2D se obtiene por una doble transformación de Fourier (2D-TF). Esta doble transformación consiste en TF unidimensionales a todas las filas y columnas de la matriz de datos. Una 2D-TF conduce al espectro 2D que muestran picos mezclas de componentes de absorción y dispersión que pueden muchas veces corregirse por ciclado de fases y manipulación de los datos, resultando en espectros con fases absortivas puras. Dependiendo de la secuencia de pulsos, se obtienen correlaciones muy útiles entre diferentes picos de los espectros 1D o se simplifican espectros 1D muy superpuestos por dispersión en las nuevas dimensiones. Los experimentos de más de dos dimensiones (RMN-3D y -4D) se generan introduciendo tiempos de evolución adicionales y se aplican principalmente en el campo de las biomoléculas, donde la complejidad estructural y la superposición espectral las hacen necesarias. 191

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Resonancia Magnética Nuclear
5.9 RMN Bidimensional Un experimento multipulso contiene etapas de preparación, evolución y detección. Los
espectros que se obtiene en estos experimentos dependen de la naturaleza de la
preparación y de la longitud del periodo de evolución. Lo que ocurre durante el período
de evolución no es detectado directamente, pero puede estimarse realizando múltiples
experimentos con periodos de evolución de diferente duración. De esta forma puede
obtenerse información sobre dos conjuntos separados de parámetros espectrales: aquel
que influye sobre la magnetización durante el periodo de evolución y el que actúa durante
la adquisición propiamente dicha. En RMN-2D se utiliza una segunda transfomada de
Fourier para convertir la dependencia temporal durante la evolución en una segunda
frecuencia, lo que permite diseñar experimentos de diversa índole, muy ricos en
información. La idea original sobre RMN-Bidimensional (RMN-2D) se debe a Jeener
(1971), aunque fue llevada a la práctica por Ernst y colaboradores en 1976.
La RMN-2D corresponde a la generación de espectros de RMN con dos frecuencias
como variables independientes. Estos espectros RMN-2D se generan registrando FID (tB2B)
al final de una secuencia de pulsos con variación sistemática de otro parámetro, más
comúnmente el tiempo de evolución tB1B entre un par de pulsos. Los FIDs sucesivos se
adquieren por incremento de los tiempos de evolución y se almacenan como filas en una
matriz de datos, tal como se muestra en la Figura 5.121, donde se comparan las técnicas
1D y 2D. El espectro 2D se obtiene por una doble transformación de Fourier (2D-TF).
Esta doble transformación consiste en TF unidimensionales a todas las filas y columnas
de la matriz de datos. Una 2D-TF conduce al espectro 2D que muestran picos mezclas
de componentes de absorción y dispersión que pueden muchas veces corregirse por
ciclado de fases y manipulación de los datos, resultando en espectros con fases absortivas
puras. Dependiendo de la secuencia de pulsos, se obtienen correlaciones muy útiles entre
diferentes picos de los espectros 1D o se simplifican espectros 1D muy superpuestos por
dispersión en las nuevas dimensiones.
Los experimentos de más de dos dimensiones (RMN-3D y -4D) se generan introduciendo
tiempos de evolución adicionales y se aplican principalmente en el campo de las
biomoléculas, donde la complejidad estructural y la superposición espectral las hacen
necesarias.
191

Resonancia Magnética Nuclear
Figura 5.121 RMN-1D y -2D
Existen diferentes métodos, basados en la técnica del eco de espines o en la
transferencia de polarización, que permiten editar un espectro 1D del espín heteronuclear
X, y que suministran información sobre el número de hidrógenos unidos al heteroátomo
(APT, INEPT, DEPT). Sin embargo, estos métodos no aportan evidencias directas sobre
qué protones están unidos a cada heteronúcleo (átomo de carbono) en una molécula. Los
experimentos de correlación heteronuclear nos permiten resolver este problema.
Comenzaremos el análisis de las técnicas de RMN-2D con el experimento de correlación
de desplazamientos químicos heteronuclear (HETCOR), en el que en una dimensión
tenemos la información sobre un tipo de espín, usualmente P
13PC, y en la otra sobre los
protones acoplados. Cada señal o pico en el diagrama de contornos nos informa sobre los
desplazamientos químicos de un núcleo de carbono y de los protones con los que está
directamente enlazado. Tal espectro permite una asignación de las señales de P
13PC si se
conocen los desplazamientos químicos de los protones o viceversa. También podemos
asignar las señales protónicas en caso que la superposición sea muy fuerte en el espectro
monodimensional, si en el espectro de P
13PC, como es habitual, se tiene una buena
dispersión de señales. Este experimento puede considerarse como un análogo
192

Resonancia Magnética Nuclear
bidimensional del INEPT, presentando el mismo incremento en sensibilidad derivado de
la transferencia de polarización desde los protones a los núcleos de P
13PC, los cuales poseen
menor razón magnetogírica.
Utilizaremos el modelo vectorial, aplicado anteriormente en los experimentos
multipulsos, que nos permite tener una idea de las bases físicas de algunas de las
secuencias más comunes. Es importante señalar que este modelo no es aplicable a la
descripción de la mayoría de los experimentos de RMN bidimensional basados en el
acoplamiento escalar, pero si es aplicable al experimento HETCOR. Un tratamiento más
riguroso y general, aplicable en sistemas con acoplamiento escalar débil, es el formalismo
del operador producto. Su complejidad no nos permite incluirlo en este texto.
5.9.1 Experimento de correlación heteronuclear directa HC-COSY (Heteronuclear
correlation, HETCOR)
Este experimento bidimensional (Freeman, Morris, Bax, 1978, 1981) puede ser
interpretado sobre la base de las secuencias antes vistas de eco de espines y tranferencia
de polarización. La secuencia de pulsos se muestra en la Figura 5.122.
Figura 5.122 Secuencia de pulsos del experimento HCCOSY
Como se observa en la Figura 5.122 el experimento consta de 4 etapas bien definidas:
preparación, evolución, mezcla y detección. Solo la etapa de mezcla resulta nueva
respecto a los experimentos multipulsos vistos con anterioridad.
En la preparación el sistema recupera el equilibrio y se crea coherencia protónica por el
pulso inicial. El período de evolución consta en este caso de un eco de espines donde
se desarrolla la coherencia protónica de acuerdo con los desplazamientos químicos
respectivos. En la etapa de mezcla se crea coherencia de protones en antifase (ΔB1B = 1/2J),
que el doble pulso simultáneo de 90P
0P transforma en coherencia en antifase de carbono por
193

Resonancia Magnética Nuclear
transferencia de polarización desde los protones. Durante la segunda mitad del período de
mezcla se crea coherencia de P
13PC en fase, que es la que detectamos en la etapa final con
desacoplamiento protónico simultáneo.
Analicemos, mediante el modelo vectorial de la magnetización, el desarrollo del
experimento para un grupo CH (P
13PC-P
1PH); que puede generalizarse a grupos CHB2B y CHB3B.
Preparación: durante el periodo de preparación la magnetización longitudinal de los
protones en equilibrio (1) se convierte en coherencia protónica cuyos dos componentes
correspondientes a las proyecciones de espín α y β del carbono inmediato comienzan el
periodo de evolución (2).
Figura 5.123 Experimento HCCOSY: magnetización transversal o
coherencia durante la evolución (modelo vectorial)
Evolución (Puntos 2-5, Figura 5.123): la evolución es un periodo de eco de espines. La
coherencia protónica evolucionará de acuerdo a sus desplazamientos químicos. La
aplicación del pulso de 180P
0P (P
13PC) reenfoca el acoplamiento heteronuclear: la posición
de la magnetización de cada protón al final del periodo de evolución, dada por el ángulo
φ (5), dependerá sólo de su desplazamiento químico (modulado débilmente por el
acoplamiento homonuclear interprotónico) y del tiempo tB1B.
194

Resonancia Magnética Nuclear
Figura 5.124 Experimento HCCOSY: coherencia durante el período de mezcla (modelo vectorial). Hasta 7 se representa la magnetización protónica. A partir de 8 se muestra la magnetización de P
13PC.
Mezcla (Puntos 5-9, Figura 5.124): El periodo de mezcla comienza con intervalo fijo ΔB1B
= 1/2J (J constante de acoplamiento heteronuclear directa C-H) para lograr poner en
antifase los componentes del doblete protónico debido al acoplamiento con el P
13PC (6,
representación en 2 y 3 dimensiones). El doble pulso de 90P
0P de protones y de P
13PC produce
la transferencia de polarización. Sus efectos se pueden analizar en forma secuencial. El
pulso de 90P
0P protónico convierte la coherencia en antifase en magnetización longitudinal
con uno de los componentes invertido (7). El pulso de 90P
0P de P
13PC genera entonces
transferencia de polarización del protón al P
13PC y coherencia en antifase de este último(8).
La eficiencia de esta transferencia dependerá de la posición de la coherencia protónica
en antifase al aplicar los pulsos de 90P
0 Py por lo tanto de lo ocurrido durante el período de
evolución. El intervalo final permite poner en fase los componentes de la coherencia de
P
13PC (9).
Detección: la magnetización transversal de P
13PC es registrada durante este periodo con
desacoplamiento protónico simultáneo. Por lo tanto sólo evolucionará el desplazamiento
químico de P
13PC.
195

Resonancia Magnética Nuclear
La señal detectada (caída de inducción libre, FID) es función no sólo del tiempo real de
adquisición tB2B, sino también de la duración del periodo de evolución tB1B. Si
secuencialmente realizamos un tándem de experimentos con valores variables de tB1B
tendremos una matriz bidimensional de datos S(tB1B,tB2B) correspondiente a la digitalización
de los FIDs (tB2B) para diferentes valores de tB1B. Una primera transformación de Fourier
sobre tB2B suministra un conjunto de espectros de P
13PC con señales cuya intensidad depende
de la duración del intervalo tB1B. Ésto se debe a que la eficiencia de la transferencia de
polarización está modulada por la posición que ocupe el doblete protónico en antifase (6)
lo que es función del desplazamiento químico protónico, y la duración del periodo de
evolución tB1B. Una segunda transformación de Fourier, ahora con respecto a tB1B, nos
revelará las frecuencias de las coherencias activas durante el período de evolución, es
decir los desplazamientos químicos de los protones acoplados directamente a cada núcleo
de carbono:
S(tB1B, tB2B) TF(tB2B) S(tB1B, δBCB) TF(tB1B) S(δBHB, δBCB)
Existe una diferencia significativa entre los períodos ΔB1B y ΔB2B. El primero se introduce
para poner en antifase los componentes del doblete protónico y por lo tanto dependerá
sólo de la magnitud de la constante de acoplamiento directa P
13PC-P
1PH, que en la mayoría de
los casos toma valores entre 125 y 160 Hz. Es usual adoptar un valor de compromiso de
135 Hz, que corresponde a un valor de 1/2J de 3.7ms. Durante el período ΔB2B deseamos
llevar los componentes del multiplete de P
13PC, inicialmente en antifase, a encontrarse en
fase. Este proceso depende ahora fuertemente del número de protones unidos a dicho
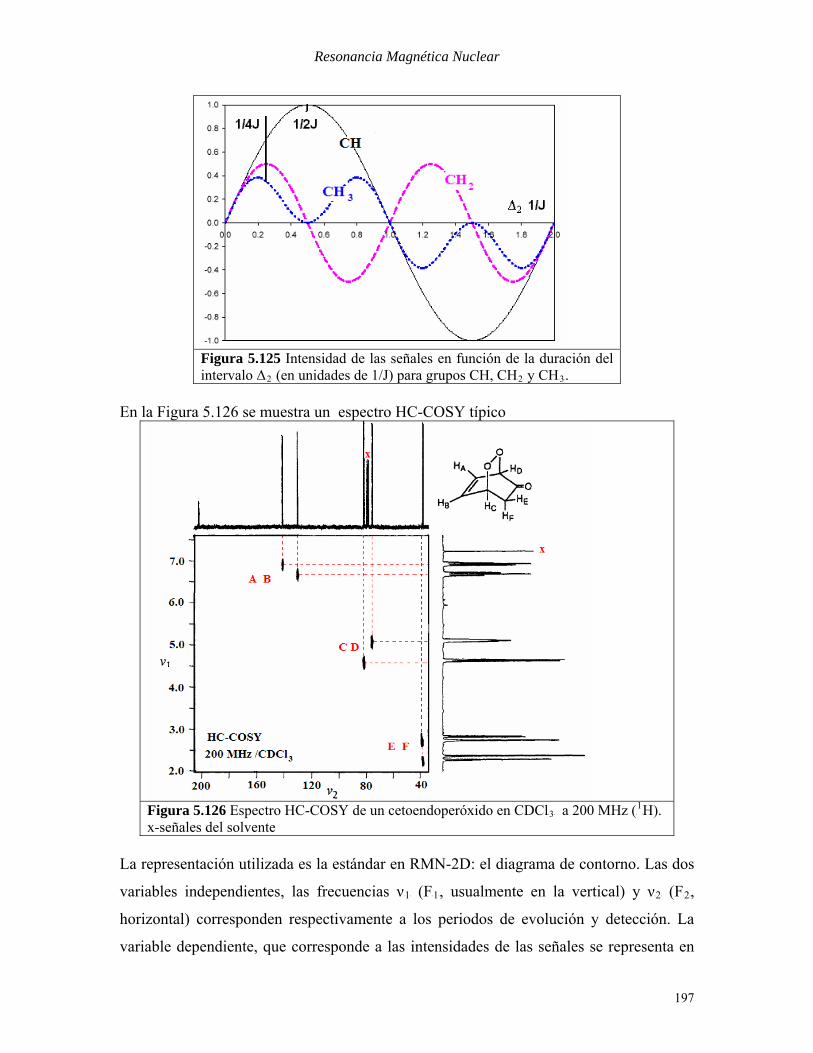
carbono, como se ha visto anteriormente en la técnica INEPT. En la Figura 5.125 se
muestra la situación para grupos CH, CHB2B y CHB3B en función de la duración del período
ΔB2B. Se adopta usualmente un valor de 1/4J (1.85ms), para el cual el nivel de reenfoque es
aceptablemente alto para todos los carbonos hidrogenados.
196
Resonancia Magnética Nuclear
Figura 5.125 Intensidad de las señales en función de la duración del intervalo ΔB2B (en unidades de 1/J) para grupos CH, CHB2B y CHB3B.
En la Figura 5.126 se muestra un espectro HC-COSY típico
Figura 5.126 Espectro HC-COSY de un cetoendoperóxido en CDClB3B a 200 MHz (P
1PH).
x-señales del solvente La representación utilizada es la estándar en RMN-2D: el diagrama de contorno. Las dos
variables independientes, las frecuencias νB1B (FB1B, usualmente en la vertical) y νB2B (FB2B,
horizontal) corresponden respectivamente a los periodos de evolución y detección. La
variable dependiente, que corresponde a las intensidades de las señales se representa en
197

Resonancia Magnética Nuclear
forma análoga a un mapa de relieve con curvas de nivel. Se muestran los espectros
monodimensionales P
1PH y de P
13PC como referencia a lo largo de FB1B(δBHB) y FB2B (δBCB)
respectivamente.
El espectro HC COSY mostrado nos permite correlacionar las señales de los carbonos
con las de los protones directamente enlazados. La escala FB2B, que corresponde a las
frecuencias del período de detección, muestra los desplazamientos químicos de P
13PC. La
escala FB1B, que corresponde a las frecuencias del período de evolución muestra los
desplazamientos químicos de los protones.
Así las señales A y B vinculan a los protones olefínicos con los carbonos
correspondientes (5.6ppm/120.0ppm). Las señales C y D vinculan a los protones
alifáticos de los CH puentes unidos al grupo epoxi con los carbonos correspondientes en
las cercanías de 80 ppm. Finalmente las señales E y F corresponden a los protones
disterotópicos del grupo CHB2B que correlacionan con el mismo carbono cerca de 40 ppm.
5.9.2 Experimentos de correlación heteronuclear directa a larga distancia: LR-
HETCOR y COLOC.
Aunque el experimento HETCOR resulta muy útil en la elucidación de estructuras, su
limitación fundamental es su carácter local. Sólo logramos correlacionar protones y
carbonos unidos directamente. Una variante de este experimento permite correlacionar
protones y carbonos separados por 2 o 3 enlaces, los que presentan pequeños
acoplamientos geminales y vecinales generalmente inferiores a 10Hz. Se impone
modificar la duración de los periodos de mezcla ΔB1B y ΔB2B, alargándolos para permitir la
creación de componentes protónicos en antifase respecto a los acoplamientos indirectos y
posterior reenfoque de los componentes de P
13PC asociados dichos acoplamientos
indirectos. Tomando un valor de compromiso para los acoplamientos a dos y tres enlaces
C-H de 7 Hz, el valor del intervalo ΔB1B debe extenderse hasta unos 71ms. El experimento
muestra ahora picos cruzados entre protones y carbonos separados 2 o 3 enlaces además
de la presencia eventual de los picos cruzados originalmente presentes en el experimento
HETCOR, que indican la conectividad a un enlace. Se ha utilizado esta técnica en el
pasado para correlacionar protones de grupos metilos con carbonos cuaternarios vecinos.
198

Resonancia Magnética Nuclear
El incremento de los periodos de mezcla en el experimento LR-HETCOR (Long Range-
HETCOR) alarga considerablemente la secuencia de pulsos reduciendo drásticamente la
sensibilidad por la acción de los procesos de relajación. Se logra compensar parcialmente
esta pérdida de sensibilidad haciendo el tiempo de evolución constante (constant time
experiment) e incluyéndolo dentro del período de polarización INEPT. El efecto neto es
que el pulso de 180P
0P es el que se desplaza en función del tiempo de evolución, de forma
tal que al final de la etapa la coherencia protónica adquiere carácter de antifase
respecto al acoplamiento con el P
13PC. Puede entonces tener lugar de inmediato la
transferencia de polarización INEPT (doble pulso de 90P
0P aplicado a ambos núcleos). Esta
modificación denominada COLOC (Correlation Spectroscopy via Long Range
Couplings, Kessler, Griesinger 1984, 1987), mostrada en la Figura 5.127, reduce la
duración de la secuencia de pulsos y el experimento es más sensible por reducción de
los efectos de la relajación.
Figura 5.127 Secuencia de pulsos del COLOC. El intervalo de tiempo entre los 2 pulsos protónicos de 90P
0P es constante. Valores típicos son ΔB1B = 25ms y
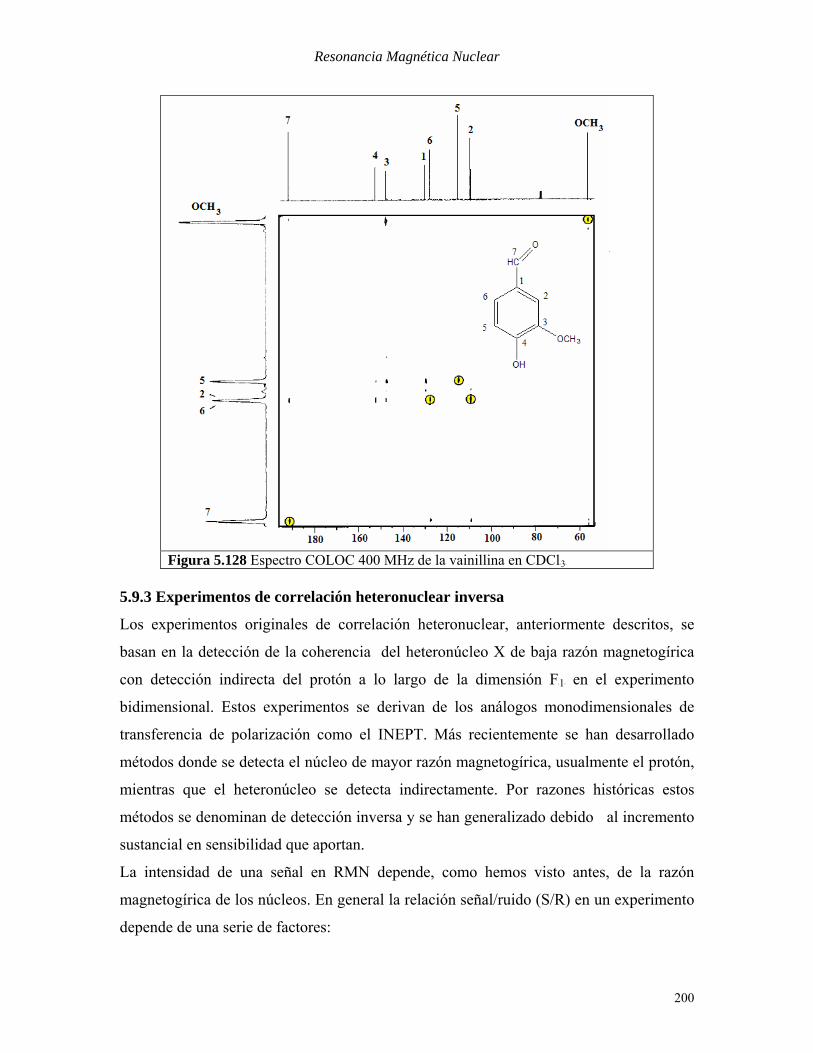
ΔB2B=33ms (optimizadas para un acoplamiento indirecto de 10 Hz) En la Figura 5.128 se muestra el espectro COLOC de la vainillina. Las correlaciones
directas, que aunque atenuadas aparecen en los espectros COLOC, aparecen indicadas
con un círculo. Obsérvese las 2 correlaciones observadas para el protón aldehídico 7 que
corresponden a los carbonos 2 y 3. Asimismo el carbono cuaternario 3 puede asignarse
fácilmente pues es el único que correlaciona con los protones del grupo metoxilo. Ya
hemos visto que en sistemas aromáticos los acoplamientos P
2PJBCH Bson débiles y los picos
cruzados correspondientes pueden estar ausentes de los espectros COLOC (HB5B muestra
picos cruzados intensos con los carbonos vecinales CB1B y CB3B, otro más débil con el
carbono geminal CB4B y no se observa con el otro carbono geminal CB6B)
199
Resonancia Magnética Nuclear
Figura 5.128 Espectro COLOC 400 MHz de la vainillina en CDClB3B
5.9.3 Experimentos de correlación heteronuclear inversa
Los experimentos originales de correlación heteronuclear, anteriormente descritos, se
basan en la detección de la coherencia del heteronúcleo X de baja razón magnetogírica
con detección indirecta del protón a lo largo de la dimensión FB1B en el experimento
bidimensional. Estos experimentos se derivan de los análogos monodimensionales de
transferencia de polarización como el INEPT. Más recientemente se han desarrollado
métodos donde se detecta el núcleo de mayor razón magnetogírica, usualmente el protón,
mientras que el heteronúcleo se detecta indirectamente. Por razones históricas estos
métodos se denominan de detección inversa y se han generalizado debido al incremento
sustancial en sensibilidad que aportan.
La intensidad de una señal en RMN depende, como hemos visto antes, de la razón
magnetogírica de los núcleos. En general la relación señal/ruido (S/R) en un experimento
depende de una serie de factores:
200

Resonancia Magnética Nuclear
-número de moléculas en el volumen activo de la sonda (concentración) N
-abundancia natural de los núcleos observados en el experimento A
-temperatura de la muestra T
-intensidad del campo magnético estático BB0B
-razones magnetogíricas de los núcleos inicialmente excitados y observados γBexcB γBobs
-tiempo de relajación transversal efectivo (define ancho de banda) TB2PB
*P
-número de FIDs acumulados n
21*
22
32
1
01 nTBNAT
RS
obsexcγγ−∝ [5.103]
Al diseñar un experimento bidimensional buscando la mayor sensibilidad para una
muestra concreta en un equipo determinado en un mínimo de tiempo, los factores que
podemos modificar son γBexcB y γBobsB: debemos tratar de que los núcleos excitados y
observados sean los de mayor razón magnetogírica.
Puede pensarse en cuatro combinaciones para experimentos de correlación
heteronuclear XH de acuerdo a la naturaleza de los núcleos excitados y detectados, las
que se muestran en la Figura 5.129.
Figura 5.129 Esquemas para generar espectros 2D de correlación heteronuclear
P, E, M, D-períodos de preparación, evolución, mezcla y detección
201

Resonancia Magnética Nuclear
El esquema 1 con excitación y detección de los núcleos de menor γBXB , y por lo tanto de menor
sensibilidad, se toma como referencia de intensidades.
En el esquema 2, que corresponde al HETCOR, se excita inicialmente el protón cuya coherencia
se deja evolucionar y durante el periodo de mezcla se transfiere la coherencia al heteronúcleo que
es el detectado directamente. En dependencia del valor de γBXB se obtienen incrementos en
sensibilidad respecto a los valores de referencia, que son mostrados a la derecha de la Figura
5.126 (γB1HB ~ 2.5 γB31PB ~4 γB13CB ~ 10 γB15NB).
Al esquema 4 le corresponden la mayores sensibilidades relativas. Aquí se excita inicialmente al
protón, se crea coherencia de X que evoluciona durante tB1B y finalmente durante la mezcla se
vuelve a coherencia protónica que es la detectada directamente. Estos experimentos de detección
inversa, que estudiaremos a continuación, representan ganancias sustanciales de sensibilidad
respecto al HETCOR que para las correlaciones P
1PH-P
13PC y P
1PH-P
15PN son de 8 y 30 veces. Aunque
estos valores teóricos de incremento de sensibilidad no sean alcanzados completamente en la
práctica, aun un incremento moderado, digamos sólo un factor de 4 para las correlaciones P
13PC-P
1PH
representan una reducción del tiempo de adquisición en 16 veces. Así si un experimento
HETCOR requiere por las condiciones de la muestra de un tiempo de adquisición del orden de
las horas, puede obtenerse la información equivalente por los métodos inversos en cuestión de
minutos. Como ventaja adicional de los experimentos de correlación inversa se tiene que en ellos
en FB1B, donde se tiene menor resolución, se despliegan los desplazamientos del heteronúcleo, que
normalmente están mucho menos superpuestos que los protónicos. Los experimentos de
correlación inversa se han impuesto y son los que se utilizan en la actualidad para las
correlaciones X-P
1PH.
Existen 2 técnicas ampliamente utilizadas para la correlación heteronuclear inversa a un enlace,
que se conocen por sus siglas en inglés: la correlación heteronuclear múltiple cuántica HMQC
(Heteronuclear Multiple Quantum Correlation) y la correlación heteronuclear a simple cuanto
HSQC (Heteronuclear Single Quantum Correlation). La información que suministran ambas
técnicas es esencialmente la misma, difiriendo sólo en detalles. El experimento HMQC es menos
sensible a desajuste en los parámetros de registro, mientras que el HSQC presenta una mayor
resolución, ventaja que es explotada particularmente en espectros muy cargados de señales como
son los de los biopolímeros. El experimento HSQC es más versátil, existiendo numerosas
variantes del mismo, lo que lo ha popularizado en la comunidad química. Ambas técnicas, como
métodos inversos, requieren una supresión efectiva de las señales que no provienen de los
protones unidos al heteronúcleo activo (P
12PC-H y P
14PN-H). Esta supresión puede lograrse por
202

Resonancia Magnética Nuclear
diferentes métodos. El mas utilizado y efectivo es el los gradientes de campo pulsados (PFGs-
Pulsed Field Gradients).
5.9.4 HMQC
Este experimento de correlación heteronuclear inverso fue desarrollado por Bax en 1986. El
HMQC permite correlacionar protones y carbonos unidos directamente a través de un enlace con
elevada sensibilidad. La secuencia del HMQC se muestra en la Figura 5.130. La programación de
fases de pulsos y receptor mostrada es para eliminar las señales provenientes de los protones
unidos a P
12PC.
Figura 5.130 Secuencia de pulsos y CTC del HMQC
La secuencia básica del HMQC es relativamente simple conteniendo sólo 4 pulsos de RF.
Ilustraremos su funcionamiento para un grupo CH, tal como hicimos en el HETCOR.
Preparación.- la secuencia comienza con un pulso de 90P
0P(P
1PH) que genera coherencia protónica
seguida de la evolución de la magnetización protónica en un intervalo Δ = 1/2P
1PJBCHB (~3.3ms) para
poner dicha coherencia en antifase con respecto al acoplamiento con el carbono vecino. Como en
el INEPT está magnetización en antifase puede transferirse a la pareja en acoplamiento mediante
un pulso. El pulso de 90P
0P(P
13PC) transfiere coherencia al carbono-13 creando lo que se conoce
como coherencia a múltiple cuanto (MQC), donde hay magnetización transversal de ambos
núcleos.
203

Resonancia Magnética Nuclear
Evolución.- las coherencias a múltiple cuanto no son detectables directamente en RMN (una
adquisición inmediata no registraría señal alguna) y evolucionan con los desplazamientos
químicos de ambos núcleos. Para estas coherencias el acoplamiento mutuo no se desarrolla. La
aplicación de un pulso de 180P
0P (P
1PH) en el centro de esta etapa elimina la evolución del
desplazamiento químico protónico. Luego durante tB1B evoluciona sólo el desplazamiento químico
de P
13PC y los acoplamientos interprotónicos (JBHHB)
Mezcla.- El pulso final de 90P
0P(P
13PC) da inicio al periodo de mezcla. Este pulso reconvierte la
MQC en coherencia protónica en antifase. Un nuevo intervalo Δ = 1/2P
1PJBCHB lleva a la coherencia
protónica en fase.
Detección.- La coherencia protónica es detectada durante tB2B evolucionando de acuerdo con los
desplazamientos químicos protónicos y las constantes de acoplamiento homonucleares JBHHB. El
acoplamiento con el P
13PC es eliminado por desacoplamiento.
Tenemos entonces:
Evolución durante tB1B: δ (P
13PC*) + P
nPJBHHB
Detección durante tB2B: δ (P
1PH-unidos a P
13PC*) + P
nPJBHHB
El espectro 2D HMQC mostrará picos o señales de cruce entre núcleos de carbonos y protones
separados por un enlace. En la dimensión FB1B podremos evaluar el desplazamiento químico del
P
13PC con la estructura fina de los acoplamientos H-H, usualmente no resueltos, mientras que en FB2B
tendremos los desplazamientos químicos de los protones unidos a cada carbono con estructura
fina correspondiente a los acoplamientos interprotónicos.
En la parte inferior de la Figura 5.130 se muestran los llamados caminos de transferencia de
coherencia (CTC). Estos CTC permiten visualizar de forma simplificada la evolución de las
coherencias durante una secuencia. Los órdenes de coherencia p para cada núcleo definen la
magnetización presente en las diferentes etapas del experimento. Sólo los pulsos de RF pueden
modificar los órdenes de coherencia. A las magnetizaciones longitudinales (equilibrio)
corresponde p = 0, a las magnetizaciones transversales o coherencias a simple cuanto generadas
por la acción de pulsos de 90P
0 Pcorresponde p = ±1. Los pulsos de 180P
0P sólo pueden invertir
coherencias. Las coherencias a múltiple cuanto se generan por aplicación de pulsos de 90P
0P a
sistemas fuera del equilibrio. Sólo es posible la detección de coherencia a simple cuanto. En la
detección por cuadratura sólo se puede detectar una de las coherencias a simple cuanto (por
convenio se utiliza p = -1)
Veamos el experimento HMQC de acuerdo a los CTC. Inicialmente en el equilibrio tenemos p =
0 para ambos núcleos. El pulso inicial de 90P
0P protónico genera coherencia a simple cuanto con p
= ±1, en el diagrama sólo se indica el camino con p = +1 que es el que finalmente conduce a la
204

Resonancia Magnética Nuclear
detección. La aplicación del pulso de 90P
0P P
13PC crea coherencias con p = ±1 para este núcleo y
globalmente coherencias a múltiple cuanto para el sistema CH. Estas coherencias son mezclas de
coherencias a cero y doble cuanto (Σp = 0, ±2) de acuerdo con los signos relativos de las
coherencias protónicas y de P
13PC.Estas coherencias múltiples están presentes durante toda la
evolución, pero la evolución con el desplazamiento químico protónico es eliminada por el pulso
de 180P
0P que invierte la coherencia protónica en el medio del intervalo. El pulso final de P
13PC
elimina la coherencia de este núcleo quedando solo la coherencia protónica que es detectada.
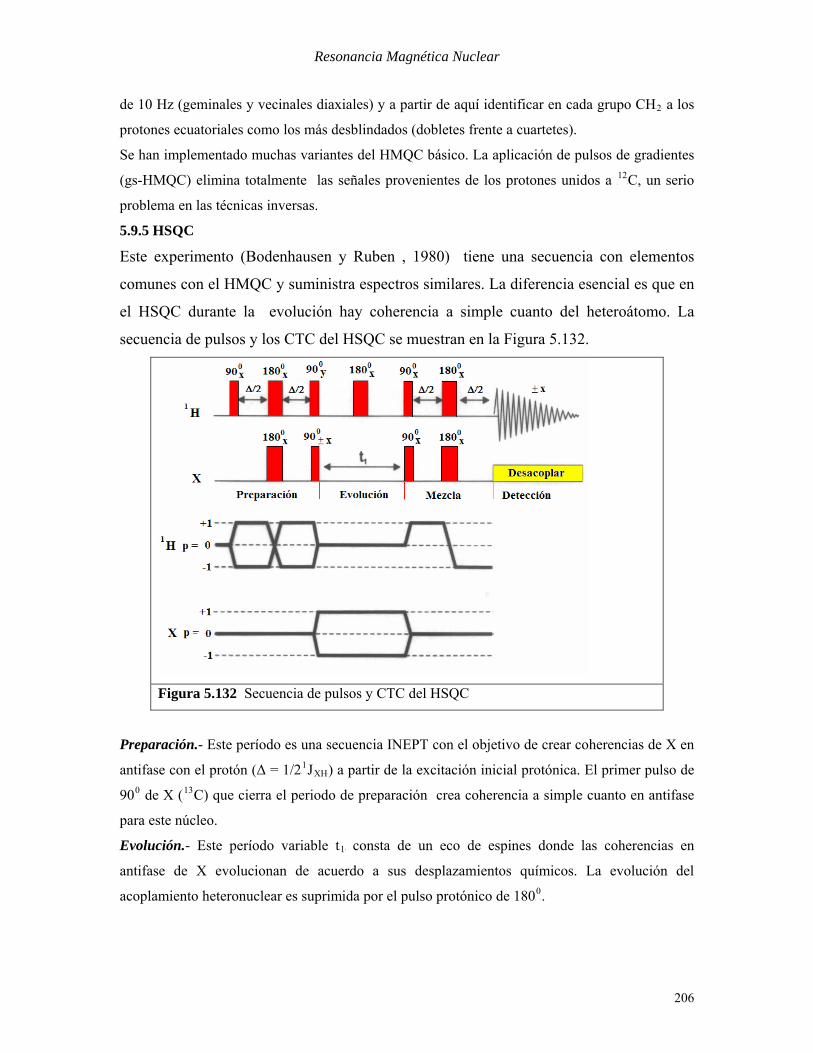
En la Figura 5.131 se muestra el espectro HMQC del mentol.
Figura 5.131 Espectro HMQC del mentol
En la Figura 5.131 se incluyen como referencias los espectros 1D RMN-P
13PC (FB1B) y RMN-P
1PH
(FB2B). Si conocemos las asignaciones de los carbonos, este experimento nos permite asignar el
espectro protónico. La señal del carbono más desblindado (CB1B) correlaciona con el protón más
desblindado en δ = 3.3 ppm. Asimismo, los carbonos unidos a protones diaterotópicos (CB3B, CB4PB
Py
CB6B) correlacionan con 2 señales protónicas cada uno. La resolución espectral en FB2B es baja, pero
permite observar estructura fina en los picos de cruce correspondientes a acoplamientos del orden
205
Resonancia Magnética Nuclear
de 10 Hz (geminales y vecinales diaxiales) y a partir de aquí identificar en cada grupo CHB2B a los
protones ecuatoriales como los más desblindados (dobletes frente a cuartetes).
Se han implementado muchas variantes del HMQC básico. La aplicación de pulsos de gradientes
(gs-HMQC) elimina totalmente las señales provenientes de los protones unidos a P
12PC, un serio
problema en las técnicas inversas.
5.9.5 HSQC
Este experimento (Bodenhausen y Ruben , 1980) tiene una secuencia con elementos
comunes con el HMQC y suministra espectros similares. La diferencia esencial es que en
el HSQC durante la evolución hay coherencia a simple cuanto del heteroátomo. La
secuencia de pulsos y los CTC del HSQC se muestran en la Figura 5.132.
Figura 5.132 Secuencia de pulsos y CTC del HSQC
Preparación.- Este período es una secuencia INEPT con el objetivo de crear coherencias de X en
antifase con el protón (Δ = 1/2P
1PJBXHB) a partir de la excitación inicial protónica. El primer pulso de
90P
0P de X (P
13PC) que cierra el periodo de preparación crea coherencia a simple cuanto en antifase
para este núcleo.
Evolución.- Este período variable tB1B consta de un eco de espines donde las coherencias en
antifase de X evolucionan de acuerdo a sus desplazamientos químicos. La evolución del
acoplamiento heteronuclear es suprimida por el pulso protónico de 180P
0P.
206

Resonancia Magnética Nuclear
Mezcla.- Consta de una secuencia retro-INEPT. Aquí las coherencias X en antifase con el protón
se reconvierten por transferencia de polarización en coherencias protónicas en antifase con X y al
final de la etapa quedan como coherencias protónicas en fase.
Detección.- Las coherencias protónicas evolucionan de acuerdo a sus desplazamientos químicos y
acoplamientos homonucleares P
nPJBHHB eliminándose el acoplamiento heteronuclear con X mediante
el desacoplador.
Tenemos entonces:
Evolución durante tB1B: δ (P
13PC*)
Detección durante tB2B: δ (P
1PH-unidos a P
13PC*) + P
nPJBHHB
La diferencia fundamental con el HMQC radica en que en la dimensión FB1B están ausentes los
acoplamientos homonucleares, por lo que los anchos de línea en esta dimensión están
apreciablemente reducidos en el HSQC. Cuando se desea una buena resolución en FB1B por la
presencia de un gran número de señales, el experimento a escoger es el HSQC. Así, el espectro
patrón para caracterizar una proteína marcada con P
15PN es el P
1PH-P
15PN-HSQC, que debe mostrar un
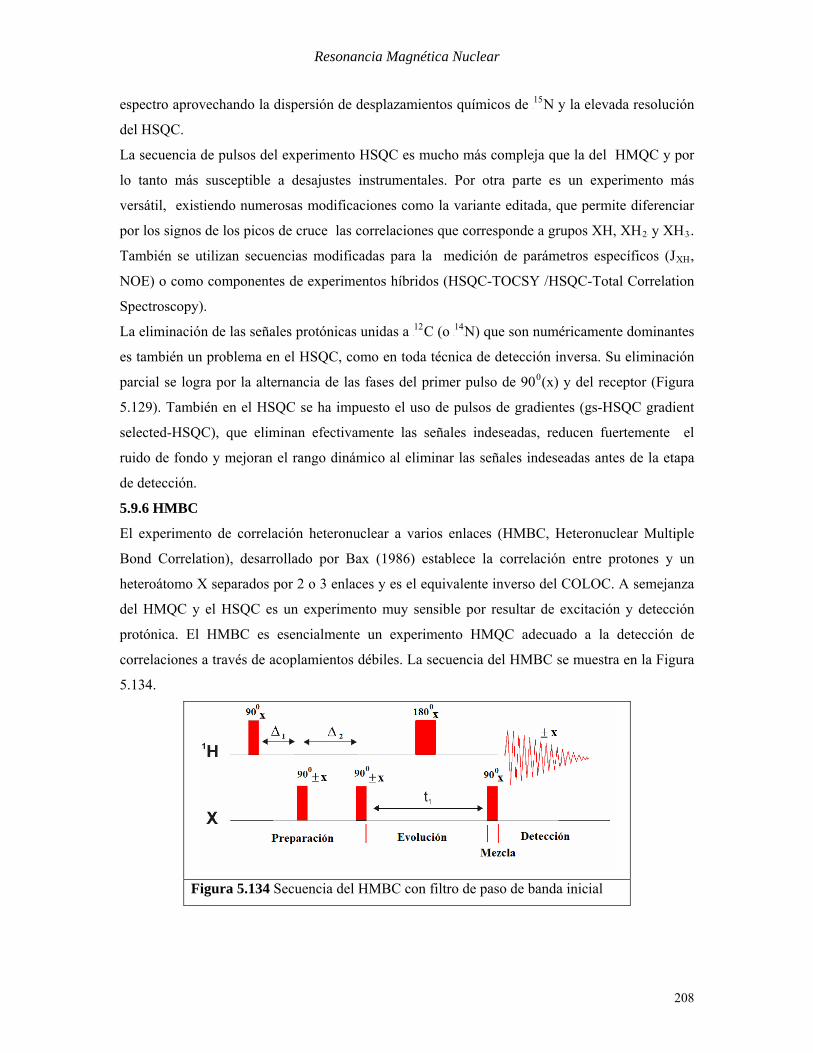
número de picos equivalentes al de residuos aminoacídicos en la estructura. En la Figura 5.133 se
muestra el espectro P
1PH-P
15PN-HSQC de una proteína.
Figura 5.133 Espectro (sección) P
1PH-P
15PN-HSQC 600 MHz de
la fracción soluble de un citocromo c (100 residuos AA)
Cada uno de los picos de cruce en el espectro anterior corresponde al grupo N-H de un residuo
aminoacídico. Obsérvese el gran número de señales que pueden distinguirse en esta parte del
207
Resonancia Magnética Nuclear
espectro aprovechando la dispersión de desplazamientos químicos de P
15PN y la elevada resolución
del HSQC.
La secuencia de pulsos del experimento HSQC es mucho más compleja que la del HMQC y por
lo tanto más susceptible a desajustes instrumentales. Por otra parte es un experimento más
versátil, existiendo numerosas modificaciones como la variante editada, que permite diferenciar
por los signos de los picos de cruce las correlaciones que corresponde a grupos XH, XHB2B y XHB3B.
También se utilizan secuencias modificadas para la medición de parámetros específicos (JBXHB,
NOE) o como componentes de experimentos híbridos (HSQC-TOCSY /HSQC-Total Correlation
Spectroscopy).
La eliminación de las señales protónicas unidas a P
12PC (o P
14PN) que son numéricamente dominantes
es también un problema en el HSQC, como en toda técnica de detección inversa. Su eliminación
parcial se logra por la alternancia de las fases del primer pulso de 90P
0P(x) y del receptor (Figura
5.129). También en el HSQC se ha impuesto el uso de pulsos de gradientes (gs-HSQC gradient
selected-HSQC), que eliminan efectivamente las señales indeseadas, reducen fuertemente el
ruido de fondo y mejoran el rango dinámico al eliminar las señales indeseadas antes de la etapa
de detección.
5.9.6 HMBC
El experimento de correlación heteronuclear a varios enlaces (HMBC, Heteronuclear Multiple
Bond Correlation), desarrollado por Bax (1986) establece la correlación entre protones y un
heteroátomo X separados por 2 o 3 enlaces y es el equivalente inverso del COLOC. A semejanza
del HMQC y el HSQC es un experimento muy sensible por resultar de excitación y detección
protónica. El HMBC es esencialmente un experimento HMQC adecuado a la detección de
correlaciones a través de acoplamientos débiles. La secuencia del HMBC se muestra en la Figura
5.134.
Figura 5.134 Secuencia del HMBC con filtro de paso de banda inicial
208

Resonancia Magnética Nuclear
Preparación.- La secuencia es esencialmente la misma del HMQC, sólo se introduce un pulso de
90P
0P (X) inicial adicional que actúa como filtro para eliminar la señal proveniente de
acoplamiento directo (ΔB1B= 1/2P
1PJBXH B~ 3.3 ms). Se debe esperar el tiempo suficiente para poner en
antifase la coherencia protónica (Δ = ΔB1B + ΔB2B = 1/P
nPJBXHB ~ 100 ms) optimizándose la creación de
coherencia a múltiple cuanto para P
nPJBXHB mediante el segundo pulso de 90P
0P (X).
Evolución.- La MQC evoluciona durante tB1B.
Mezcla.- La MQC es reconvertida en coherencia protónica por el pulso final X. El intervalo de
reenfoque Δ al final del período de mezcla en HMQC es eliminado aquí por razones de
sensibilidad: es demasiado largo y la relajación amortiguaría fuertemente las señales.
Detección.- Comúnmente la detección puede o no acompañarse de desacoplamiento de X. Se
prefiere no desacoplar a X para que las señales residuales indeseadas que corresponden a los
acoplamientos P
1PJBXHB aparezcan como dobletes en FB2B lo que facilita su identificación.
En una medida aun mayor que sus homólogos la técnica HMBC se beneficia grandemente con la
introducción de los pulsos de gradiente, que reducen considerablemente el ruido de fondo de los
espectros.
Las técnicas de correlación heteronuclear a 2 o 3 enlaces permiten obtener información
valiosísima en la determinación estructural. La intensidad de los picos de cruce en el espectro
depende tanto de la magnitud de los acoplamientos como del intervalo Δ seleccionado. Estos
acoplamientos son comúnmente de 2-7 Hz, a lo que corresponde un Δ óptimo de unos 100ms.
El debilitamiento de las señales por relajación durante el tiempo entre la excitación y la detección
hace recomendable reducir significativamente este intervalo y valores de Δ de compromiso del
orden de 60-80 ms (óptimas para acoplamientos de 4-10 Hz) son recomendables para los trabajos
de rutina. Sólo en moléculas pequeñas con relajación poco eficiente pueden elevarse los valores
de Δ en busca de conectividades a través de acoplamientos más pequeños. En la Figura 5.135 se
muestra el espectro HMBC de un compuesto bicíclico.
209

Resonancia Magnética Nuclear
Figura 5.135 Espectro HMBC sin desacoplar durante la detección
En la Figura 5.135 se muestran las conectividades del protón HB2B que muestra picos de cruce con
CB9B (geminal) y CB4B, CB5B y CB8B (vecinales). Se observa además el acoplamiento directo con CB2B que
puede distinguirse de los anteriores por el doblete resultante del acoplamiento P
1PJBCHB en FB2B (doble
flecha). También se indican otros picos de cruce producto de acoplamientos directos.
La riqueza de la información que suministra el HMBC lo convierte en un experimento muy
utilizado en ensamblar una estructura desconocida a partir de fragmentos conocidos. Mientras que
con los espectros HMQC o HSQC sólo logramos conectividades locales a través de un enlace, el
HMBC permite conectar átomos más distantes, inclusive aquellos que carecen de protones y para
los cuales las técnicas de correlación homonuclear no funcionan. En la Figura 5.136 se muestra
el espectro HMBC del α-pineno.
210
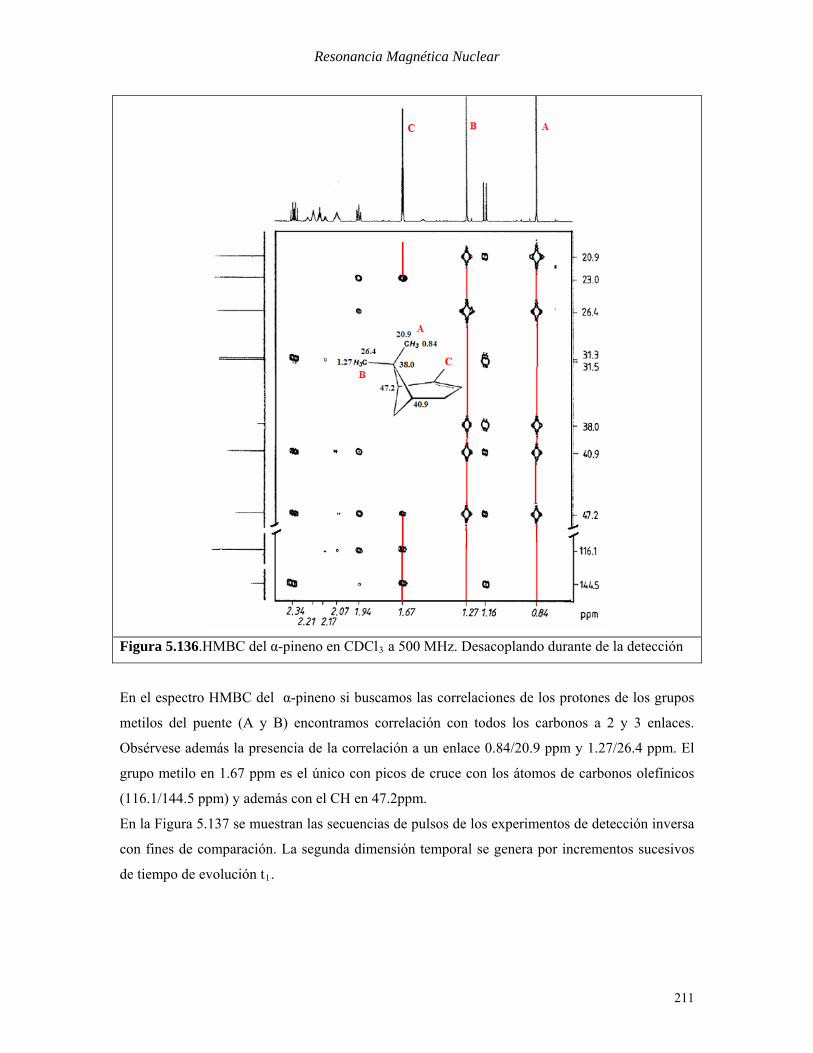
Resonancia Magnética Nuclear
Figura 5.136.HMBC del α-pineno en CDClB3B a 500 MHz. Desacoplando durante de la detección
En el espectro HMBC del α-pineno si buscamos las correlaciones de los protones de los grupos
metilos del puente (A y B) encontramos correlación con todos los carbonos a 2 y 3 enlaces.
Obsérvese además la presencia de la correlación a un enlace 0.84/20.9 ppm y 1.27/26.4 ppm. El
grupo metilo en 1.67 ppm es el único con picos de cruce con los átomos de carbonos olefínicos
(116.1/144.5 ppm) y además con el CH en 47.2ppm.
En la Figura 5.137 se muestran las secuencias de pulsos de los experimentos de detección inversa
con fines de comparación. La segunda dimensión temporal se genera por incrementos sucesivos
de tiempo de evolución tB1B.
211

Resonancia Magnética Nuclear
Figura 5.137 Experimentos de correlación heteronuclear con detección inversa.
5.9.7 Experimento de correlación homonuclear (Correlation Spectroscopy, COSY)
Este experimento mediante el cual se correlacionan núcleos a través del acoplamiento escalar fue
históricamente el primero de los experimentos bidimensionales (Jeener 1971, Ernst 1976). La
técnica COSY permite el mapeo de los núcleos (protones) que tienen acoplamientos escalares en
una molécula. La secuencia de pulsos del COSY, mostrada en la Figura 5.138 es muy simple: se
aplican 2 pulsos de 90P
0P separados por un intervalo variable de tiempo que es el periodo de
evolución y al final del segundo pulso se adquiere el FID.
Figura 5.138 Secuencia de pulsos del COSY
El período de mezcla del COSY se reduce al segundo pulso de 90P
0P. Este pulso transfiere
magnetización de un núcleo a otro núcleo que esté acoplado con él. Este proceso se conoce como
transferencia de coherencia y ha sido tratado anteriormente en el caso heteronuclear como
transferencia de polarización. Para heteronúcleos esta transferencia puede describirse mediante
212

Resonancia Magnética Nuclear
el modelo vectorial porque los pulsos protónicos y del heteronúcleo actúan independientemente.
En el caso homonuclear el problema es mucho más complejo porque todos los espines
experimentan la acción de los pulsos simultáneamente. No obstante los procesos de transferencia
son análogos al caso heteronuclear y la transferencia de coherencias tiene lugar entre los espines
protónicos acoplados.
Consideremos el experimento COSY en un sistema de 2 protones débilmente acoplados (sistema
AX) con una constante de acoplamiento JBAXB y desplazamientos químicos νBAB y νBBB. La
magnetización asociada con A comenzará a precesar de acuerdo a νBAB durante tB1B. El segundo pulso
transfiere una parte de la magnetización (coherencia) de A hacia X y la otra parte permanece
como magnetización (coherencia) de A. Aquella coherencia de A que permanece después del
segundo pulso evolucionará de acuerdo con νBAB durante tB2B. El pico correspondiente en el espectro
bidimensional aparecerá a (νBAB, νBAB). Este pico que aparece a las mismas frecuencias en ambas
dimensiones se denomina pico diagonal. La coherencia original de A que resulta transferida a X
por el pulso de mezcla, evolucionará durante tB2B como νBXB y el pico correspondiente aparecerá en el
espectro 2D a (νBAB, νBXB). Este pico es denominado pico de cruce, se encuentra fuera de la diagonal
y nos indica que los núcleos A y X están acoplados. En efecto, para que ocurra transferencia de
coherencia, la coherencia de A tiene que haber desarrollado carácter de antifase respecto a X y
esto sólo puede ocurrir si ambos núcleos están acoplados. Este análisis puede repetirse para una
excitación inicial de X que conduce al pico diagonal (νBXB, νBXB) y al pico cruzado (νBXB, νBAB) al otro
lado de la diagonal respecto a (νBAB, νBXB). Una representación esquemática del espectro COSY del
sistema AX se muestra en la Figura 5.139.
Figura 5.139 Esquema del espectro COSY para sistema AX
La información en un espectro COSY está en los picos de cruce fuera de la diagonal. Estos picos
indican el acoplamiento escalar entre dos espines y por lo tanto que un reducido número de
enlaces separan a los núcleos correspondientes. La eficiencia del pulso de mezcla en transferir
coherencia depende, como hemos visto, de la presencia de componentes en antifase en el
213
Resonancia Magnética Nuclear
momento de su aplicación. El periodo de evolución debe ser suficientemente largo para permitir
la formación de estos componentes en antifase (φ = 2πJtB1B). Si la constante de acoplamiento es
muy pequeña, el tiempo requerido para su desarrollo puede ser muy largo respecto a la acción de
los procesos de relajación y los picos de cruce correspondientes no son detectables. No obstante,
en muchas ocasiones, pueden detectarse picos de cruce en el COSY correspondientes a
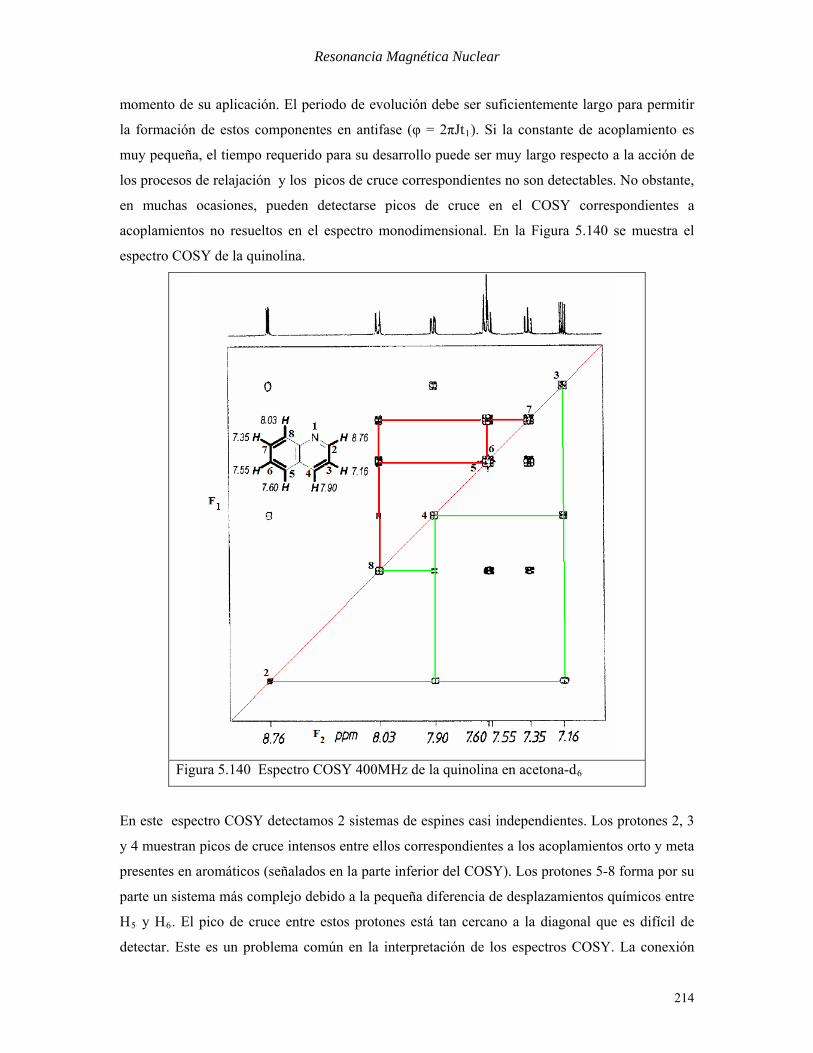
acoplamientos no resueltos en el espectro monodimensional. En la Figura 5.140 se muestra el
espectro COSY de la quinolina.
Figura 5.140 Espectro COSY 400MHz de la quinolina en acetona-dB6B
En este espectro COSY detectamos 2 sistemas de espines casi independientes. Los protones 2, 3
y 4 muestran picos de cruce intensos entre ellos correspondientes a los acoplamientos orto y meta
presentes en aromáticos (señalados en la parte inferior del COSY). Los protones 5-8 forma por su
parte un sistema más complejo debido a la pequeña diferencia de desplazamientos químicos entre
HB5B y HB6B. El pico de cruce entre estos protones está tan cercano a la diagonal que es difícil de
detectar. Este es un problema común en la interpretación de los espectros COSY. La conexión
214

Resonancia Magnética Nuclear
entre ambos sistemas de espines se evidencia por el débil pico de cruce entre HB4B y HB8B (7.90-8.03
ppm) correspondiente al acoplamiento en zigzag a larga distancia P
5PJ no resuelto en el espectro
1D.
En el COSY original no pueden separarse las componentes absortivas de las dispersivas de las
señales por correcciones de fase como es común en RMN-1D. El espectro COSY se presenta en
modo magnitud, por lo que las señales aparecen muy ensanchadas (mezclas de absorción con
dispersión). Se han desarrollado variantes del COSY, como el COSY con doble filtro cuántico
(DQF-COSY Double Quantum Filter-COSY) que permiten presentaciones en modo absortivo
con mucha mejor resolución y picos diagonales considerablemente reducidos (que no aportan
información y pueden impedir la observación de picos cruzados cercanos a la diagonal).
La aplicación de pulsos de gradientes permite eliminar o reducir las programaciones de fase de
los experimentos y acortar considerablemente los tiempos de registro (gs-COSY, gs-COSY
sensible a la fase). La obtención de un gs-COSY es cuestión de unos pocos minutos. En todas
estas variantes la interpretación del espectro es idéntica a la del COSY original.
5.9.8 Experimento de correlación homonuclear total (Total Correlation Spectroscopy,
TOCSY).
La característica más notable del COSY es la obtención de correlación directa entre 2 protones
acoplados, que inmediatamente sugiere que los mismos están en una relación geminal o vecinal
en la molécula. Podemos ver al experimento COSY como equivalente al conjunto de todos los
experimentos de desacoplamiento de espines imaginables para una molécula. El experimento de
correlación total TOCSY también suministra correlaciones entre espines directamente acoplados,
pero incluye además correlaciones entre todos los espines de un sistema, aun los que no estén
directamente acoplados. Por ejemplo, en una cadena de protones acoplados A-B-C-D, en el
experimento COSY observaríamos pico cruzado de A con su única pareja de acoplamiento B. En
el experimento TOCSY observaríamos picos cruzados de A con B, C y D si los acoplamientos B-
C y C-D son apreciables. Podemos correlacionar en principio mediante el TOCSY todos los
núcleos de un sistema de espines y de aquí su nombre de espectroscopia de correlación total. La
habilidad del TOCSY para detectar transferencia de coherencia a través de una cadena es muy útil
sobre todo en biomoléculas complejas donde están presentes sistemas de espines aislados, en
unidades discretas. En las proteínas los protones de los residuos aminoacídicos constituyen
sistemas de espines independientes, no hay acoplamientos interprotónicos a través del enlace
peptídico. En los polisacáridos, los protones de las unidades de azúcares forman sistemas de
espines independientes, no hay acoplamientos interprotónicos a través del enlace glicosídico.
215

Resonancia Magnética Nuclear
En el TOCSY los componentes de los multipletes en los picos de cruce están en fase mientras
que en el COSY estos componentes están en antifase. En condiciones de baja resolución, los
componentes en antifase del COSY se superponen y tienden a anularse mutuamente, mientras que
en el TOCSY se refuerzan, por lo que esta resulta una técnica más sensible.
La secuencia del experimento TOCSY se muestra en la Figura 5.141. La secuencia de pulsos del
TOCSY es similar a la del COSY excepto que el pulso de mezcla del COSY es sustituido por una
etapa de mezclado con bloqueo de espines, cuyo objetivo es permitir la transferencia de
coherencias a lo largo del sistema de espines.
Figura 5.141 Secuencia de pulsos del TOCSY y tren de pulsos para bloqueo de espines
Para entender el funcionamiento del período de bloqueo de espines en el TOCSY veamos lo que
ocurriría para un período de evolución tB1B = 0. El pulso inicial crea magnetización en el eje x.
Inmediatamente se aplica el bloqueo de espines que está compuesto por un tren de pulsos de
180P
0Px separados por cortos intervalos de tiempo 2δ. Podemos considerar a todo el período de
mezcla como una sucesión de ecos de espines (δ-180-δ). Como conocemos, en estos ecos se
reenfocan los desplazamientos químicos pero no los acoplamientos homonucleares. Es decir, en
todo el período de mezcla τBmB queda congelada la evolución por desplazamiento químico, los
espines experimentan el mismo campo magnético efectivo y los vectores correspondientes
permanecen fijos en el eje y del sistema de coordenadas rotatorio, de ahí el nombre de bloqueo
de espines. Por contraste los acoplamientos homonucleares evolucionan a través de la toda la
secuencia de ecos de espines. Durante el período de mezcla los espines de un sistema se
comportan como equivalentes (no hay diferencias de desplazamientos químicos efectivos) y
acoplados por lo que constituyen un sistema de no primer orden donde los espines individuales
pierden su identidad (mezcla isotrópica). Esta situación permite el intercambio de coherencias
entre todos los espines del sistema.
La secuencia del TOCSY puede interpretarse entonces como:
216
Resonancia Magnética Nuclear
Preparación.- creación de coherencias.
Evolución.- evolución de las señales, quedando marcadas por sus desplazamientos químicos. (δBHB
+ JBHHB)
Mezcla.- durante el bloqueo de espines se produce transferencia de coherencia a otros núcleos de
su sistema de espines, cuya efectividad depende de los valores de las constantes de acoplamiento
y el tiempo de mezcla.
Detección.- se detectarán las señales provenientes de un núcleo que no transfirió coherencia a
otro (pico diagonal) o que recibió coherencia de otros miembros del sistema de espines (picos de
cruce). Evolucionan (δBHB + JBHHB).
La apariencia del espectro TOCSY es similar al COSY, con picos simétricos alrededor de la
diagonal, pero con mucha mayor abundancia de picos de cruce.
El intercambio de coherencia tiene carácter oscilante y su efectividad depende del tiempo de
mezcla y de los valores de las constantes de acoplamiento a lo largo del sistema de espines. Para
tiempos de mezcla cortos (< 20 ms) la transferencia de coherencia sólo alcanza a los núcleos
directamente acoplados y el TOCSY registrado en estas condiciones se asemeja al COSY. A
tiempos de mezcla mayores es posible la transferencia de coherencia entre núcleos no acoplados
directamente a través de intermediarios. En casos favorables pueden encontrarse picos de cruce
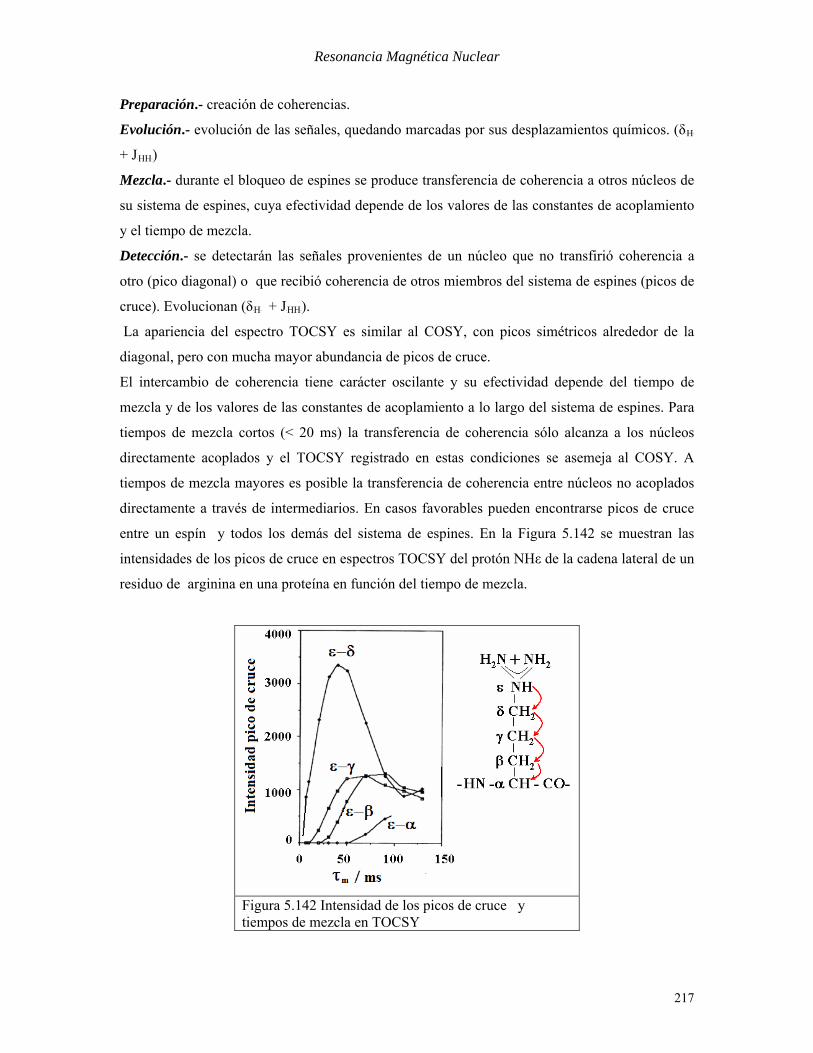
entre un espín y todos los demás del sistema de espines. En la Figura 5.142 se muestran las
intensidades de los picos de cruce en espectros TOCSY del protón NHε de la cadena lateral de un
residuo de arginina en una proteína en función del tiempo de mezcla.
Figura 5.142 Intensidad de los picos de cruce y tiempos de mezcla en TOCSY
217

Resonancia Magnética Nuclear
Para tiempos de mezcla cortos (τBmB< 0.1/JBAXB) sólo hay transferencia efectiva al protón vecinal (ε -
δ). Sólo a partir de 80 ms resulta apreciable el pico de cruce con el protón más remoto del sistema
de espines (ε – α).
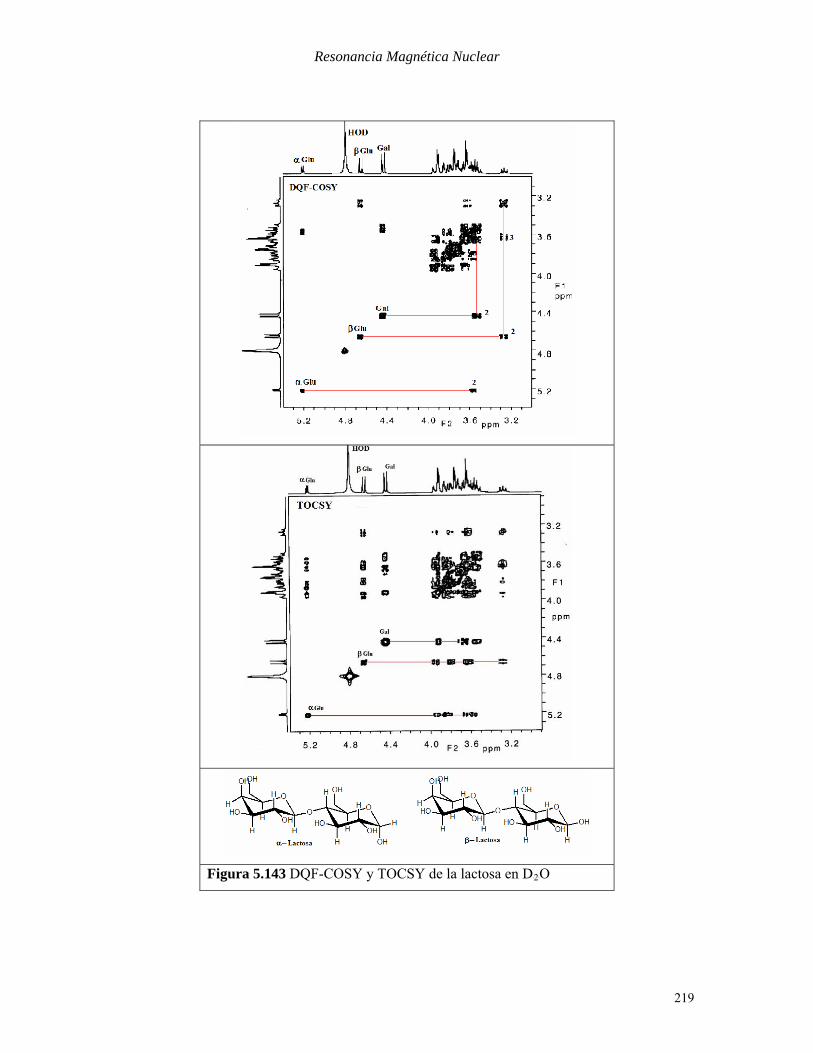
En la Figura 5.143 se muestran los espectros COSY y TOCSY de la lactosa (mezcla de anómeros
β y α). Los tres protones anoméricos aparecen apreciablemente desblindados por encima de 4.4
ppm. El resto de los protones de la unidades de glucosa (Glu) y galactosa (Gal) forman un bloque
compacto en la zona de 3.5-4.0 ppm. Los picos de cruce del espectro COSY nos permiten
identificar claramente donde están los protones en las posiciones 2, pero más allá resultan
difíciles las asignaciones por la gran superposición de señales. En el espectro TOCSY sin
embargo, a partir de los protones anoméricos, podemos identificar las posiciones de los protones
de los diferentes azúcares mediante los picos de cruce de los tres protones anoméricos.
Para asignar los protones dentro de cada sistema de espines pueden realizarse experimentos
TOCSY con diferentes tiempos de mezcla. A tiempos de mezcla crecientes van apareciendo las
señales de los protones cada vez más alejados del protón anomérico. Una alternativa es obtener
espectros TOCSY en su variante monodimensional. Estos espectros se obtienen eliminando el
período de evolución y sustituyendo al pulso inicial de excitación por un pulso selectivo aplicado
a la frecuencia del protón cuyo sistema de espines quiere identificarse. En la Figura 5.144 se
muestra los espectros 1D-TOCSY con diferentes tiempos de mezcla. En este caso se aplica el
pulso selectivo a la frecuencia del protón anomérico de la unidad de β –glucosa. Con tiempos de
mezcla crecientes se puede observar la aparición sucesiva de los protones a lo largo de la cadena
de acoplamiento. Para observar los protones en posición 6 hay que esperar un tiempo muy largo,
donde ya la relajación ha comenzado a debilitar el grueso de las señales.
218
Resonancia Magnética Nuclear
Figura 5.143 DQF-COSY y TOCSY de la lactosa en DB2BO
219
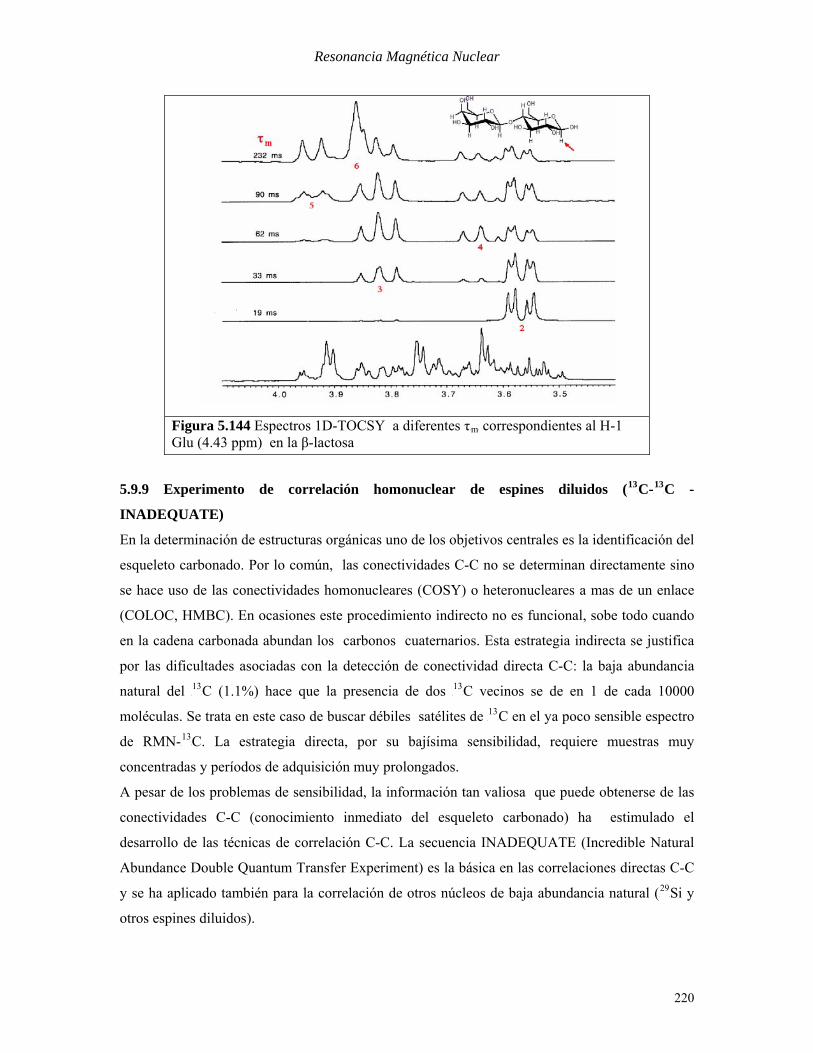
Resonancia Magnética Nuclear
Figura 5.144 Espectros 1D-TOCSY a diferentes τBmB correspondientes al H-1 Glu (4.43 ppm) en la β-lactosa
5.9.9 Experimento de correlación homonuclear de espines diluidos (P
13PC-P
13PC -
INADEQUATE)
En la determinación de estructuras orgánicas uno de los objetivos centrales es la identificación del
esqueleto carbonado. Por lo común, las conectividades C-C no se determinan directamente sino
se hace uso de las conectividades homonucleares (COSY) o heteronucleares a mas de un enlace
(COLOC, HMBC). En ocasiones este procedimiento indirecto no es funcional, sobe todo cuando
en la cadena carbonada abundan los carbonos cuaternarios. Esta estrategia indirecta se justifica
por las dificultades asociadas con la detección de conectividad directa C-C: la baja abundancia
natural del P
13PC (1.1%) hace que la presencia de dos P
13PC vecinos se de en 1 de cada 10000
moléculas. Se trata en este caso de buscar débiles satélites de P
13PC en el ya poco sensible espectro
de RMN-P
13PC. La estrategia directa, por su bajísima sensibilidad, requiere muestras muy
concentradas y períodos de adquisición muy prolongados.
A pesar de los problemas de sensibilidad, la información tan valiosa que puede obtenerse de las
conectividades C-C (conocimiento inmediato del esqueleto carbonado) ha estimulado el
desarrollo de las técnicas de correlación C-C. La secuencia INADEQUATE (Incredible Natural
Abundance Double Quantum Transfer Experiment) es la básica en las correlaciones directas C-C
y se ha aplicado también para la correlación de otros núcleos de baja abundancia natural (P
29PSi y
otros espines diluidos).
220

Resonancia Magnética Nuclear
El problema más grave de esta técnica viene dado por la interferencia causada por las señales
dominantes que carecen de conectividades homonucleares y por lo tanto no pueden suministrar la
información buscada. En el INADEQUATE la acción de un doble filtro cuántico elimina las
señales que corresponden a especies con un solo P
13PC. La secuencia del experimento y los CTC del
INDEQUATE se muestran en la Figura 5.145. El espectro de correlación bidimensional se genera
permitiendo a las coherencias a doble cuanto (sólo presentes en especies con 2 P
13PC acoplados)
evolucionar durante tB1B seguida de una reconversión a coherencia a simple cuanto de P
13PC que se
detecta durante tB2B.
Figura 5.145 Secuencia de pulsos y CTC del experimento INADEQUATE
Preparación.- Se crea coherencia a simple cuanto por el pulso inicial. La generación de
coherencia a doble cuanto requiere de una disposición antifase de los vectores de acoplamiento
que se desarrollan durante el período Δ = 1/2P
1PJBCCB. Ésto se logra mediante un eco de espines que
hace este proceso independiente de los desplazamientos químicos. Finalmente el segundo pulso
de 90P
0P crea las coherencias a doble cuanto.
Evolución.- El período variable tB1B corresponde a las coherencias a doble cuanto evolucionando a
la suma de los desplazamientos químicos de los núcleos de carbonos acoplados. Así, para dos
carbonos A y X acoplados, la frecuencia del pico de cruce en FB1B viene dada por (νBAB + νBBB).
Mezcla.- La reconversión a coherencias a simple cuanto de A o X (νBAB , νBXB) se logra mediante el
pulso final de mezcla que da inicio al período de detección.
Detección.- Durante este período evolucionan las coherencias de núcleos de carbono a simple
cuanto y los acoplamientos P
1PJBCCB.
221

Resonancia Magnética Nuclear
Durante toda la secuencia se irradian los protones en forma no selectiva para eliminar la
evolución de los acoplamientos heteronucleares y generar efectos NOE para aumentar la
sensibilidad.
La interpretación de los espectros 2D-INADEQUATE es algo más complicada que la de otros
espectros 2D. En la dimensión FB1B, que corresponde a la evolución de las coherencias a doble
cuanto, los picos cruzados aparecen a la suma de las frecuencias relativas de ambos espines en el
sistema de coordenadas rotatorio, sin que exista un espectro 1D de referencia. Como a cada
coherencia DQ corresponden 2 coherencias a simple cuanto, moviéndonos en la horizontal
podemos leer en FB2B las frecuencias de los espines acoplados y que dan lugar a dicha coherencia
a DQ. En la Figura 5.146 se muestra el espectro 2D-INADEQUATE del n-butanol.
Figura 5.146 Espectro 2D-Inadequate del n-butanol
En el espectro tenemos la dimensión FB1B que corresponde al DQ (νBAB + νBXB), en esta escala no
tenemos representación espectral. Los picos de cruce aparecen en FB2B a las frecuencias de
resonancia de los carbonos asociados al DQ. Para interpretar el espectro buscamos en FB2B un pico
de cruce de un núcleo de carbono que se pueda asignar inequívocamente. En este caso la señal
del CB1B debe corresponder a la más desblindada (62 ppm). El DQ vincula a este pico de cruce con
otro del núcleo de carbono en δ = 35 ppm, que puede asignarse entonces al CB2B. Como CB2B está
unido directamente a dos núcleos de carbono, debe tener otro pico de cruce que se identifica
222

Resonancia Magnética Nuclear
fácilmente y lo vincula con el CB3B (18 ppm). Finalmente a partir de CB3B se identifica el CB4B (13
ppm). En FB2B los picos de cruce muestran una estructura fina de dobletes (acoplamiento P
1PJBCCB). La
interpretación se facilita por la existencia de una seudodiagonal que une los centros de las líneas
horizontales que ligan cada par de picos de cruce.
5.9.10 Experimentos de correlación dipolar
Todos los experimentos bidimensionales anteriormente descritos detectan conectividades
entre espines que están acoplados escalarmente o que forman parte de un mismo sistema
de espines. Comenzaremos a tratar ahora los experimentos bidimensionales que nos
permiten encontrar las conectividades entre núcleos que tienen acoplamiento dipolar, es
decir aquellos que están vecinos espacialmente y tienen contactos NOE. El experimento
básico de este tipo es el 2D-NOESY (Nuclear Overhauser Enhancement Spectroscopy).
Para moléculas en régimen de movilidad intermedio ( 1≈cωτ ), donde la magnitud del
NOE es muy pequeña, se puede utilizar con éxito el experimento ROESY (Rotating
frame Overhauser Enhancement Spectroscopy) para la detección de vecindad espacial
entre espines.
5.9.10.1 Experimento NOESY
El experimento NOESY permite la detección de NOEs transitorios. La secuencia de
pulsos y los caminos de transferencia de coherencia del NOESY se muestran en la Figura
5.147.
Figura 5.147 Secuencia de pulsos y CTC del NOESY
La secuencia del NOESY está relacionada estrechamente con la del COSY. La diferencia
mas significativa está en el tiempo de mezcla durante el cual se desarrolla el efecto NOE.
Afortunadamente el experimento NOESY puede interpretarse haciendo uso del modelo
223

Resonancia Magnética Nuclear
vectorial (Figura 5.148), toda vez que aquí los acoplamientos escalares no juegan ningún
papel. Después de la excitación inicial (Figura 5.148-2) y durante la evolución (tB1B) los
vectores de magnetización están en el plano xy (coherencias) y evolucionarán en dicho
plano de acuerdo con sus desplazamientos químicos (Figura 5.148-3, υ - offset respecto a
la RF). El segundo pulso de 90P
0P lleva componentes de la magnetización al eje –z (Figura
5.148-4), creando la inversión de poblaciones requerida para observar NOE transitorios
que se desarrollan durante el tiempo de mezcla. La fracción de magnetización remanente
en el plano xy daría lugar al COSY si se adquiriera inmediatamente. Al final del tiempo
de mezcla las magnetizaciones resultantes en el eje z son convertidas en coherencias por
el pulso final de 90P
0P y detectadas durante tB2B. El período de evolución sirve para marcar
los desplazamientos químicos. Si partimos de la excitación de un núcleo S, este
evolucionará a su frecuencia durante tB1B y su magnetización, invertida por el segundo
pulso de 90P
0P, se puede transferir a un núcleo vecino I si hay relajación cruzada (σBISB) entre
los mismos. Esta magnetización transferida se detecta a la frecuencia de I durante tB2B por
lo que observamos un pico de cruce (FB1 B- υBSB, FB2B - υBIB) que indica el contacto NOE entre
los mismos. La magnetización no transferida durante el período de mezcla da lugar a los
picos diagonales.
La secuencia es repetida para los diferentes valores de los tiempos de evolución.
Generalmente el período de preparación incluye un intervalo de recuperación de las
magnetizaciones previo al pulso inicial de 1-3 s.
Figura 5.148 Modelo vectorial del experimento NOESY
La secuencia NOESY requiere una programación de fases de pulsos y receptor (no
mostrada en la Figura 5.147) para eliminar coherencias indeseadas. Como en el NOESY
durante el período de mezcla sólo interesa la magnetización longitudinal (p = 0),
cualquier coherencia a simple o doble cuanto indeseada puede eliminarse de forma
relativamente sencilla. Una alternativa a la programación de fases es el uso de un pulso
224

Resonancia Magnética Nuclear
de gradientes durante el período de mezcla. La inhomogeneidad asociada al pulso de
gradientes en z (inhomogeneidad a lo largo de z, campos BB0B efectivos que varían
linealmente) desfasa y anula toda coherencia SQ o DQ durante este tiempo pero no
afecta la magnetización longitudinal, que es la de interés en el NOESY. Las
interferencias más difíciles de eliminar son las coherencias a cero cuanto existentes
durante el período de mezcla que sean convertidas por el pulso final en coherencias a
simple cuanto detectables. El procedimiento más común para eliminar estas interferencias
es variar ligeramente en forma aleatoria el tiempo de mezcla de un incremento de tB1B a
otro. La magnetización de interés para el NOESY (magnetización longitudinal) es poco
sensible a pequeños cambios en τBmB, mientras que las coherencias a cero cuanto (oscilarán
de acuerdo al offset y serán muy sensible aún a cambios moderados de τBmB, anulándose
efectivamente por este procedimiento.
Los espectros NOESY presentan una apariencia similar al COSY, con simetría respecto
a la diagonal. Se prefiere su presentación en formato sensible a la fase, donde pueden
ajustarse todas las señales como absortivas positivas o negativas. Lo importante es el
signo relativo de las señales. Si ajustamos las señales en la diagonal como positivas, el
signo de los picos de cruce depende del origen del pico y de la movilidad de la red, como
se muestra en la Tabla 5.29.
Tabla 5.29 Signo de los picos de cruce en NOESY Signo diagonal Origen pico cruce Signo pico de cruce
NOE positivo (alta movilidad) Negativo NOE negativo (baja movilidad) Positivo
Positivo (convenio)
Intercambio químico Positivo
Dos espines en intercambio químico lento (señales independientes) pueden intercambiar
magnetización y dar lugar a picos de cruce en el NOESY. En moléculas pequeñas con NOEs
positivos los picos de cruce por contacto NOE y por intercambio químico pueden ser distinguidos
por su signo. Sin embargo en macromoléculas ambos picos de cruce tienen el mismo signo y no
pueden ser diferenciados.
Un parámetro importante a definir al diseñar un experimento NOESY es el tiempo de mezcla (ver
Tabla 5.30). Este debe por una parte ser lo suficientemente largo para permitir la creación del
NOE, pero por otra su duración esta limitada por la relajación longitudinal que tiende a debilitar
todas las señales. Si se quieren determinar distancias intenucleares se debe trabajar en la zona
lineal inicial de desarrollo del NOE, evitándose además la aparición de picos de cruce falsos por
225
Resonancia Magnética Nuclear
difusión de espines. En la Tabla 5.30 se indican los valores de tiempo de mezcla recomendables
para los experimentos NOESY.
Tabla 5.30 Tiempos de mezcla para NOESY Régimen de movilidad NOE Características Tiempos de mezcla Moléculas pequeñas + TB1 B↑B BσBISB↓ ~ TB1B 500-2000
ms Macromoléculas - TB1 B↓ σBISB↑ ~0.5 TB1 B100-300 ms Moléculas intermedias 0 NOESY no viable, usar ROESY
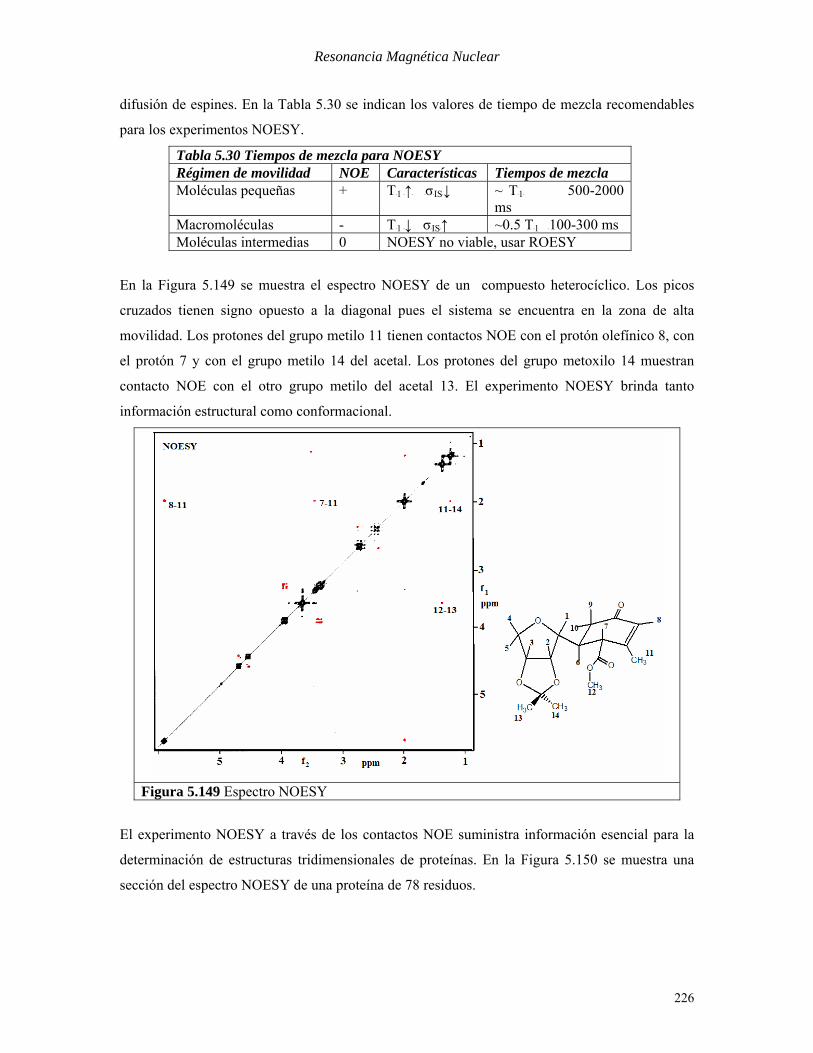
En la Figura 5.149 se muestra el espectro NOESY de un compuesto heterocíclico. Los picos
cruzados tienen signo opuesto a la diagonal pues el sistema se encuentra en la zona de alta
movilidad. Los protones del grupo metilo 11 tienen contactos NOE con el protón olefínico 8, con
el protón 7 y con el grupo metilo 14 del acetal. Los protones del grupo metoxilo 14 muestran
contacto NOE con el otro grupo metilo del acetal 13. El experimento NOESY brinda tanto
información estructural como conformacional.
Figura 5.149 Espectro NOESY
El experimento NOESY a través de los contactos NOE suministra información esencial para la
determinación de estructuras tridimensionales de proteínas. En la Figura 5.150 se muestra una
sección del espectro NOESY de una proteína de 78 residuos.
226
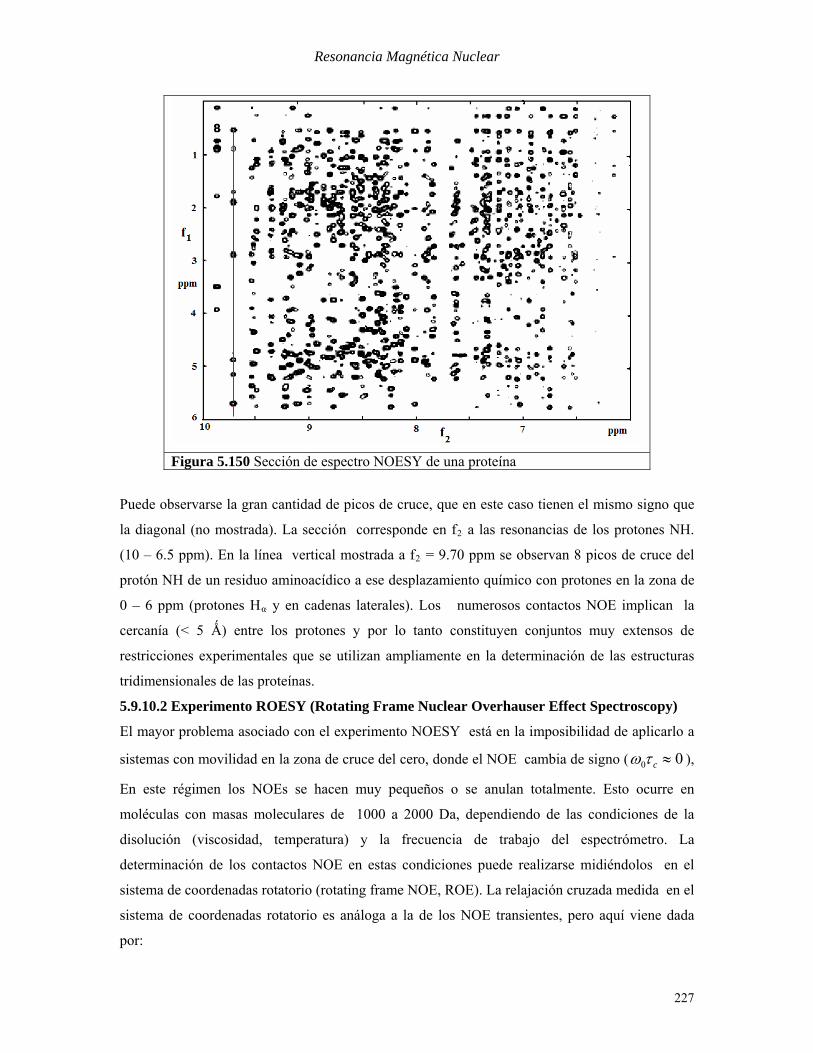
Resonancia Magnética Nuclear
Figura 5.150 Sección de espectro NOESY de una proteína
Puede observarse la gran cantidad de picos de cruce, que en este caso tienen el mismo signo que
la diagonal (no mostrada). La sección corresponde en fB2B a las resonancias de los protones NH.
(10 – 6.5 ppm). En la línea vertical mostrada a fB2B = 9.70 ppm se observan 8 picos de cruce del
protón NH de un residuo aminoacídico a ese desplazamiento químico con protones en la zona de
0 – 6 ppm (protones HBαB y en cadenas laterales). Los numerosos contactos NOE implican la
cercanía (< 5 Ǻ) entre los protones y por lo tanto constituyen conjuntos muy extensos de
restricciones experimentales que se utilizan ampliamente en la determinación de las estructuras
tridimensionales de las proteínas.
5.9.10.2 Experimento ROESY (Rotating Frame Nuclear Overhauser Effect Spectroscopy)
El mayor problema asociado con el experimento NOESY está en la imposibilidad de aplicarlo a
sistemas con movilidad en la zona de cruce del cero, donde el NOE cambia de signo ( 00 ≈cτω ),
En este régimen los NOEs se hacen muy pequeños o se anulan totalmente. Esto ocurre en
moléculas con masas moleculares de 1000 a 2000 Da, dependiendo de las condiciones de la
disolución (viscosidad, temperatura) y la frecuencia de trabajo del espectrómetro. La
determinación de los contactos NOE en estas condiciones puede realizarse midiéndolos en el
sistema de coordenadas rotatorio (rotating frame NOE, ROE). La relajación cruzada medida en el
sistema de coordenadas rotatorio es análoga a la de los NOE transientes, pero aquí viene dada
por:
227
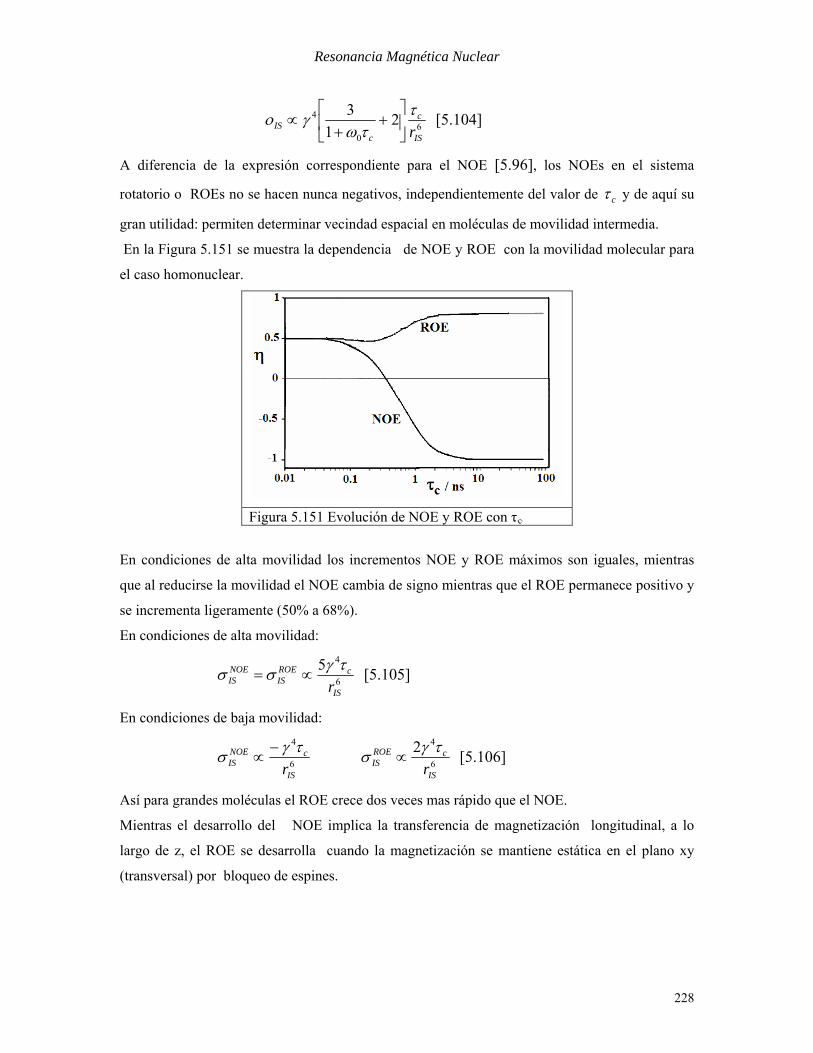
Resonancia Magnética Nuclear
60
4 21
3
IS
c
cIS r
ττω
γο ⎥⎦
⎤⎢⎣
⎡+
+∝ [5.104]
A diferencia de la expresión correspondiente para el NOE [5.96], los NOEs en el sistema
rotatorio o ROEs no se hacen nunca negativos, independientemente del valor de cτ y de aquí su
gran utilidad: permiten determinar vecindad espacial en moléculas de movilidad intermedia.
En la Figura 5.151 se muestra la dependencia de NOE y ROE con la movilidad molecular para
el caso homonuclear.
Figura 5.151 Evolución de NOE y ROE con τBcB
En condiciones de alta movilidad los incrementos NOE y ROE máximos son iguales, mientras
que al reducirse la movilidad el NOE cambia de signo mientras que el ROE permanece positivo y
se incrementa ligeramente (50% a 68%).
En condiciones de alta movilidad:
6
45
IS
cROEIS
NOEIS r
τγσσ ∝= [5.105]
En condiciones de baja movilidad:
6
4
IS
cNOEIS r
τγσ −∝ 6
42
IS
cROEIS r
τγσ ∝ [5.106]
Así para grandes moléculas el ROE crece dos veces mas rápido que el NOE.
Mientras el desarrollo del NOE implica la transferencia de magnetización longitudinal, a lo
largo de z, el ROE se desarrolla cuando la magnetización se mantiene estática en el plano xy
(transversal) por bloqueo de espines.
228

Resonancia Magnética Nuclear
La secuencia de pulsos del experimento ROESY (Figura 5.152) es muy similar a la del
experimento TOCSY. En el ROESY la potencia del campo estático BB1B para el bloqueo de
espines es considerablemente más débil que el tren de pulsos del TOCSY.
Figura 5.152 Secuencia de pulsos del ROESY
La selección de los tiempos de mezcla es análoga a la del NOESY. Valores del orden de 600 ms o
menores son adecuados para moléculas de masas moleculares moderadas. En macromoléculas
deben utilizarse los valores recomendados para el NOESY (100-300 ms) o menores teniendo en
cuenta que el ROE crece dos veces mas rápido que el NOE.
La acción del bloqueo de espines es mantener congelada la disposición que los vectores de
magnetización adoptan durante el período de evolución. Esta disposición se perdería por la
acción de la dispersión de desplazamientos químicos en ausencia del bloqueo de espines. El
bloqueo permite el desarrollo de los ROEs por relajación cruzada en el plano transversal. Aquí la
relajación está caracterizada por una constante de tiempo TB1ρB (TB1ρB~ TB2B). Al utilizar el bloqueo de
espines estamos sustituyendo el campo estático BB0B del NOE por el campo BB1B de la RF, mucho
más débil. Mientras que γBB0B corresponde a frecuencias del orden de los cientos de MHz, el valor
de γBB1B corresponde a los kHz (frecuencias en el sistema rotatorio). De aquí que la relación
11 <<cτω se cumple para todos los valores accesibles de τBcB y las moléculas, independientemente
de su tamaño, se comportan como si estuvieran en el régimen de agudeza extrema. Los ROE son
pues siempre positivos. Por lo tanto en el espectro ROESY los picos de cruce originados por
contactos ROE siempre tienen signo opuesto a la diagonal. Aparte de los signos de las señales, la
apariencia e interpretación de los espectros ROESY es semejante a los obtenidos por el
experimento NOESY.
Los espectros ROESY tienden a presentar más artefactos que los espectros NOESY, lo que
puede conducir a interpretaciones erróneas. Como se señaló antes, la secuencia de pulsos del
ROESY es muy similar a la del TOCSY aunque en el ROESY el bloqueo de espines se emplea
para optimizar la transferencia de magnetización por interacciones dipolares en lugar de la
transferencia coherente de magnetización a través de los acoplamientos escalares que caracteriza
al TOCSY. Así como en el NOESY se pueden presentar artefactos tipo COSY, en el ROESY los
229

Resonancia Magnética Nuclear
más frecuentes son de tipo TOCSY, es decir pueden observarse picos de cruce espúreos entre
espines del mismo sistema, pero que carecen de un real contacto ROE. Como se mencionó
anteriormente, el campo de bloqueo de espines en ROESY es apreciablemente más débil que el
TOCSY. Esto reduce la intensidad de los picos espúreos tipo TOCSY, que por otra parte tienen el
mismo signo que los picos diagonales y pueden identificarse como tales en condiciones
favorables. La precaución de desplazar la frecuencia de BB1B del centro del rango espectral
contribuye a minimizar los artefactos TOCSY en el espectro ROESY. Los riesgos mayores de
artefactos se presentan entre espines de desplazamiento químico muy semejante y picos de cruce
cercanos a la diagonal.
230
Related Documents











