Rice SIZ1, a SUMO E3 ligase, controls spikelet fertility through regulation of anther dehiscence Saminathan Thangasamy 1,2,3 , Cian-Ling Guo 3 , Ming-Hsiang Chuang 3 , Ming-Hsing Lai 4 , Jychian Chen 1,5,6 and Guang-Yuh Jauh 1,3,7 1 Molecular and Biological Agricultural Sciences, Taiwan International Graduate Program, National Chung-Hsing University – Academia Sinica, Taipei, Taiwan; 2 Graduate Institute of Biotechnology, National Chung-Hsing University, Taichung, Taiwan; 3 Institute of Plant and Microbial Biology, Academia Sinica, Taipei, Taiwan; 4 Crop Science Division, Taiwan Agricultural Research Institute, Wufeng, Taichung, Taiwan; 5 Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan; 6 Department of Life Sciences, National Chung-Hsing University, Taichung, Taiwan; 7 Biotechnology Center, National Chung-Hsing University, Taichung, Taiwan Author for correspondence: Guang-Yuh Jauh Tel: +886 2 27893173 Email: [email protected] Received: 8 June 2010 Accepted: 1 October 2010 New Phytologist (2011) 189: 869–882 doi: 10.1111/j.1469-8137.2010.03538.x Key words: anther dehiscence, endothecium, rice (Oryza sativa), SIZ1, spikelet fertility, SUMO E3 ligase. Summary • Sumoylation, a post-translational modification, has important functions in both animals and plants. However, the biological function of the SUMO E3 ligase, SIZ1, in rice (Oryza sativa) is still under investigation. • In this study, we employed two different genetic approaches, the use of siz1 T-DNA mutant and SIZ1-RNAi transgenic plants, to characterize the function of rice SIZ1. • Genetic results revealed the co-segregation of single T-DNA insertional recessive mutation with the observed phenotypes in siz1. In addition to showing reduced plant height, tiller number and seed set percentage, both the siz1 mutant and SIZ1-RNAi transgenic plants showed obvious defects in anther dehiscence, but not pollen viability. The anther indehiscence in siz1 was probably a result of defects in endothecium development before anthesis. Interestingly, rice orthologs of AtIRX and ZmMADS2, which are essential for endothecium development during anther dehiscence, were significantly down-regulated in siz1. Compared with the wild-type, the sumoylation profile of high-molecular-weight proteins in mature spikelets was reduced significantly in siz1 and the SIZ1-RNAi line with notably reduced SIZ1 expression. The nuclear localization signal located in the SIZ1 C-terminus was sufficient for its nuclear targeting in bombarded onion epidermis. • The results suggest the functional role of SIZ1, a SUMO E3 ligase, in regulating rice anther dehiscence. Introduction Rice (Oryza sativa) is one of the most important food crops for more than one-half of the world’s population. The grains of most crops are the major food resources for most animals, including humans, and their production depends on successful sexual reproduction. The stamen is the male reproductive organ; pollen is one of the major routes of gene flow in nature through cross-pollination and is the major concern in the spread of genetically modified plants (Ma, 2005). The microspore initiates and undergoes a series of differentiation programs in the anther to result in the male gametophyte, pollen grain, for double fertilization. A comprehensive understanding of the functions of the genes and mechanisms involved in anther and pollen development will greatly benefit our knowledge of basic plant development, agriculture and the environment. Male sterility caused by various defects, including anther indehiscence, has adverse effects on agricultural productivity and diminishes significantly the crop yield. The study of male sterility is an excellent way to understand the molecular mechanisms essential for the complicated sexual reproduc- tion process. The final step of anther development is anther dehiscence, the opening of the anther wall, to release mature pollen grains for pollination, fertilization and seed produc- tion. For successful anther dehiscence, the indispensable New Phytologist Research ȑ 2010 The Authors New Phytologist ȑ 2010 New Phytologist Trust New Phytologist (2011) 189: 869–882 869 www.newphytologist.com

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Rice SIZ1, a SUMO E3 ligase, controls spikelet fertilitythrough regulation of anther dehiscence
Saminathan Thangasamy1,2,3, Cian-Ling Guo3, Ming-Hsiang Chuang3, Ming-Hsing Lai4, Jychian Chen1,5,6 and
Guang-Yuh Jauh1,3,7
1Molecular and Biological Agricultural Sciences, Taiwan International Graduate Program, National Chung-Hsing University – Academia Sinica, Taipei,
Taiwan; 2Graduate Institute of Biotechnology, National Chung-Hsing University, Taichung, Taiwan; 3Institute of Plant and Microbial Biology, Academia
Sinica, Taipei, Taiwan; 4Crop Science Division, Taiwan Agricultural Research Institute, Wufeng, Taichung, Taiwan; 5Institute of Molecular Biology,
Academia Sinica, Taipei, Taiwan; 6Department of Life Sciences, National Chung-Hsing University, Taichung, Taiwan; 7Biotechnology Center, National
Chung-Hsing University, Taichung, Taiwan
Author for correspondence:Guang-Yuh Jauh
Tel: +886 2 27893173
Email: [email protected]
Received: 8 June 2010Accepted: 1 October 2010
New Phytologist (2011) 189: 869–882doi: 10.1111/j.1469-8137.2010.03538.x
Key words: anther dehiscence,endothecium, rice (Oryza sativa), SIZ1,
spikelet fertility, SUMO E3 ligase.
Summary
• Sumoylation, a post-translational modification, has important functions in both
animals and plants. However, the biological function of the SUMO E3 ligase, SIZ1,
in rice (Oryza sativa) is still under investigation.
• In this study, we employed two different genetic approaches, the use of siz1
T-DNA mutant and SIZ1-RNAi transgenic plants, to characterize the function of
rice SIZ1.
• Genetic results revealed the co-segregation of single T-DNA insertional recessive
mutation with the observed phenotypes in siz1. In addition to showing reduced
plant height, tiller number and seed set percentage, both the siz1 mutant and
SIZ1-RNAi transgenic plants showed obvious defects in anther dehiscence, but not
pollen viability. The anther indehiscence in siz1 was probably a result of defects in
endothecium development before anthesis. Interestingly, rice orthologs of AtIRX
and ZmMADS2, which are essential for endothecium development during anther
dehiscence, were significantly down-regulated in siz1. Compared with the
wild-type, the sumoylation profile of high-molecular-weight proteins in mature
spikelets was reduced significantly in siz1 and the SIZ1-RNAi line with notably
reduced SIZ1 expression. The nuclear localization signal located in the SIZ1
C-terminus was sufficient for its nuclear targeting in bombarded onion epidermis.
• The results suggest the functional role of SIZ1, a SUMO E3 ligase, in regulating
rice anther dehiscence.
Introduction
Rice (Oryza sativa) is one of the most important food cropsfor more than one-half of the world’s population. Thegrains of most crops are the major food resources for mostanimals, including humans, and their production dependson successful sexual reproduction. The stamen is the malereproductive organ; pollen is one of the major routes ofgene flow in nature through cross-pollination and is themajor concern in the spread of genetically modified plants(Ma, 2005). The microspore initiates and undergoes a seriesof differentiation programs in the anther to result in themale gametophyte, pollen grain, for double fertilization. A
comprehensive understanding of the functions of the genesand mechanisms involved in anther and pollen developmentwill greatly benefit our knowledge of basic plant development,agriculture and the environment.
Male sterility caused by various defects, including antherindehiscence, has adverse effects on agricultural productivityand diminishes significantly the crop yield. The study ofmale sterility is an excellent way to understand the molecularmechanisms essential for the complicated sexual reproduc-tion process. The final step of anther development is antherdehiscence, the opening of the anther wall, to release maturepollen grains for pollination, fertilization and seed produc-tion. For successful anther dehiscence, the indispensable
NewPhytologist Research
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882 869www.newphytologist.com

processes are the timing of dehiscence, formation of second-ary wall thickening, degeneration of various anther tissues,changes in carbohydrate metabolism and movement ofwater out of the anther (Goldberg et al., 1993; Scott et al.,2004; Ma, 2005). The anther dehiscence process is initiatedafter the formation of tetrads and involves three uniquetissues: endothecium, septum and stomium. The stomiumtissue, constituting a single layer of specialized cells, is thelast breakage site for anther dehiscence (Keijzer, 1987). Inaddition to anatomical observations, recent genetic studieshave highlighted the molecular functions of several genesregulating anther dehiscence, such as receptor-like proteinkinase (Mizuno et al., 2007) and polygalacturonase (Gorguetet al., 2009), that are essential for stomium opening andmale fertility.
Plant hormones play important roles in anther dehis-cence. For example, the delayed dehiscence1 mutant, defectivein the enzyme 12-oxophytodienoate reductase in jasmonicacid (JA) biosynthesis, shows abnormal stomium degenera-tion and timing of anther dehiscence (Sanders et al., 2000).DEFECTIVE IN ANTHER DEHISCENCE1 (DAD1)encodes the chloroplastic phospholipase A1, which partici-pates in JA biosynthesis; its corresponding mutant exhibitsdefects in flower opening, anther dehiscence and pollenmaturation (Ishiguro et al., 2001). Mutation of another JAbiosynthesis pathway gene, ALLENE OXIDE SYNTHASE(AOS), also reveals delayed anther dehiscence and, conse-quently, causes male sterility (Park et al., 2002). In tobacco,ethylene regulates the timing of anther dehiscence insteadof affecting ovule and anther development. Tobacco plantscarrying an Arabidopsis mutated etr1-1 allele show delayeddegeneration and dehydration of stomium cells, which alterthe synchronization for pollination (Rieu et al., 2003). Twoethylene receptors from petunia, PhERS1 and PhETR2,regulate the synchronization of anther dehiscence withflower opening (Wang & Kumar, 2007). Auxin coordinatesfilament elongation, pollen maturation and anther dehis-cence (Cecchetti et al., 2008). The herbicide glyphosateinhibits anther dehiscence in cotton by inducing changes inauxin-mediated microtubule orientation from longitudinalto transverse and secondary wall modifications in endo-thecium cells (Yasuor et al., 2006). Recent studies have alsosuggested that other internal and external factors affectanther dehiscence, including high temperature (Matsui et al.,2001) and the availability and ⁄ or movement of water in rice(Bots et al., 2005), mineral nutrient potassium in barley(Rehman & Yun, 2006) and the accumulation of flavonoidsin Arabidopsis (Thompson et al., 2009).
Several transcription factors are involved in anther dehis-cence. For example, ANTHER INDEHISCENCE1 (AID1)encodes a MYB protein, and its mutation results in partialto complete spikelet sterility because of defective pollendevelopment, septum degradation and stomium breakagein rice (Zhu et al., 2004). Arabidopsis male sterile35, a
MYB26 mutant, produces viable pollen, but also malesterility, as a result of nondehiscent anthers (Steiner-Langeet al., 2003), because MYB26 regulates secondary wallthickening in the endothecium and subsequent dehiscenceby modulating the expression of several IRXs and NSTs(Yang et al., 2007). NAC (NO APICAL MERISTEM,ARABIDOPSIS TRANSCRIPTION ACTIVATION FACTORAND CUP-SHAPED COTYLEDON), secondary wallthickening promoting factor 1 (NST1) and NST2 regulatethe thickening of the secondary wall that is necessaryfor anther dehiscence (Mitsuda et al., 2005). In maize,ZmMADS2, highly expressed in endothecium and con-nective tissues before dehiscence and in mature pollen afterdehiscence, accumulates apoptotic bodies during antherdehiscence and is required for anther dehiscence andpollen maturation (Schreiber et al., 2004). Accumulatingstudies have revealed important transcriptional regulationin anther dehiscence; however, the role of post-transla-tional modification, such as sumoylation, in this process isunclear.
The process of sumoylation conjugates Small Ubiquitin-related MOdifier (SUMO) to substrate proteins throughreversible post-translational modification, and is highlyregulated by the environment in yeast and animal systems(Johnson, 2004). Similar to ubiquitination, sumoylationalso involves a cascade of biochemical processes with theenzymes SUMO E1 activating enzyme, E2 conjugatingenzyme and E3 ligase enzyme (Hay, 2005). In metazoansand yeast, sumoylated proteins play vital roles in diversecellular events, including cell cycle progression and mitosis,DNA repair, chromatin stability, cell division, nucleartargeting, innate immunity and transcriptional regulation(Johnson, 2004; Gill, 2005). Two members of the ProteinInhibitor of Activated STAT (PIAS) group of SUMO E3ligases (SIZs) are in yeast (SIZ1, SIZ2), one member is inArabidopsis (AtSIZ1) and two members are in rice (OsSIZ1and OsSIZ2) (Miura et al., 2007a). In Arabidopsis, sumoy-lation by AtSIZ1 is implicated in the regulation of phos-phate starvation by modifying the transcription factorPHR1 to positively regulate the expression of the down-stream genes AtIPS1 and AtRNS1 (Miura et al., 2005), inlow-temperature stress tolerance by altering ICE1 transcrip-tion factor (Miura et al., 2007b), and in the process of plantgrowth and development through hormone signaling.In addition, sumoylation participates in salicylic acid(SA)-independent basal thermotolerance (Yoo et al., 2006),SA-mediated innate immunity (Lee et al., 2007), SA-mediated floral repression by changing FLC chromatinstructure (Jin et al., 2008), negative regulation of abscisicacid (ABA) signaling by sumoylating ABI5 (Miura et al.,2009) and drought responses (Catala et al., 2007). All char-acterized SUMO E3 ligases contain conserved domainsindispensable for DNA binding, sumoylation and nuclearlocalization. Arabidopsis AtSIZ1 is expressed in all plant
870 Research
NewPhytologist
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com

tissues, and its mutant, atsiz1-3, produces reduced growthand low tolerance to drought with significant down-regu-lation of many genes participating in the brassinosteroidbiosynthesis ⁄ signaling and drought tolerance ⁄ JA signalingpathways, respectively (Catala et al., 2007). In addition,AtSIZ1 regulates cell growth and plant development inArabidopsis by regulating SA accumulation (Miura et al.,2010). Recently, OsSIZ1 and OsSIZ2 have been suggestedto function as SUMO E3 ligases to induce protein sumoyla-tion under different stress conditions, including low and hightemperature, NaCl and ABA (Park et al., 2010); nevertheless,their biological functions in rice are still under investigation.
Sumoylation plays diverse functions in yeast, metazoans,animals and Arabidopsis; however, the role of SUMO E3ligase in rice is still unclear. In this study, we investigatedthe functional characterization of a single recessive riceSUMO E3 ligase mutant, siz1, regulating spikelet sterility,plant height and tillering phenotypes. Interestingly, similarphenomena were found in SIZ1-RNAi transgenic lineswith significantly down-regulated expression of SIZ1.Comprehensive morphological investigation of siz1 andSIZ1-RNAi anthers revealed that the spikelet sterility wascaused by defective anther dehiscence instead of pollenviability. Moreover, the expression of a few genes involvedin secondary wall thickening in the endothecium duringanther dehiscence in other species and their rice orthologswere down-regulated in the siz1 mutant. We discuss thepossible mechanisms of SIZ1 in the regulation of antherdehiscence.
Materials and Methods
Plant materials and growth conditions
Seeds of rice (Oryza sativa L. cv Japonica) cv Tainung67(wild-type, Wt), siz1 and SIZ1-RNAi were germinated, and3-wk-old seedlings were transplanted under geneticallymodified organism-free field conditions at the TaiwanAgricultural Research Institute, Taichung, and under glass-house conditions (30�C and 20�C during the day andnight, respectively, with a 14 h light : 10 h dark cycle) atAcademia Sinica in Taipei.
Generation of SIZ1-RNAi transgenic plants
The inverted repeat region of SIZ1 (Fig. 4a and SupportingInformation Fig. S1a) was amplified by the use of primersets (Table S1) and cloned into the pANDA RNAi-expressionvector (Miki & Shimamoto, 2004) with a gateway cloning kit(Invitrogen, Carlsbad, CA, USA). Agrobacterium-mediatedtransformation of rice wild-type calli was performed asdescribed by Sallaud et al. (2003). Plantlets regenerated fromtransformed calli were selected for hygromycin resistance, andregenerated transgenic plants were moved to an isolated field
for phenotypic observation. The transgenic seedlings were con-firmed by genotyping and the expression of SIZ1. The locusnumber for SIZ1 from the TIGR version 6.1 rice genomeannotation database (http://rice.plantbiology.msu.edu/) isOs05g03430.
DNA extraction, genotyping and DNA blotting
Genomic DNA was isolated from rice seedlings of thewild-type, T2 segregants and siz1 by the cetyl trimethyl-ammonium bromide (CTAB) method (Murray & Thompson,1980). Genotyping of T2 segregants involved the primersets in Table S1. For Southern blot analysis, genomic DNA(20 lg) from each sample was digested with differentrestriction enzymes, separated on 0.8% agarose gel andtransferred onto GeneScreen Plus (PerkinElmer, Zaventem,Belgium) as described in Ho et al. (2000). The b-glucuron-idase (GUS) region from T-DNA (pTAG8; the vector usedto generate the T-DNA mutant population) and Tos17sequences from wild-type genomic DNA were PCR ampli-fied with primers (Table S1) and labeled with a32P-dCTPusing the Amersham Rediprime II random prime labelingsystem (GE Healthcare, Amersham, UK). Prehybridization,hybridization and washing steps followed the manufac-turer’s protocol (Biochain Institute, Hayward, CA, USA).Images of membranes were developed after exposure toBioMax MS film (Carestream Health, Inc. Rochester, NY,USA) at )70�C for 6 d.
RNA extraction, RNA blotting, semi-quantitative andreal-time reverse transcription-polymerase chainreaction (RT-PCR)
Total RNA was isolated from different tissues and stagesof wild-type, siz1 and SIZ1-RNAi plants with the use ofTRIzol reagent, according to the manufacturer’s protocol(Invitrogen). Ten micrograms of total RNA were separatedon 1.2% agarose gel with formaldehyde and transferredinto GeneScreen Plus (PerkinElmer) as described previously(Ho et al., 2000). The 3¢-cDNA of SIZ1 was PCR-amplified with the primer sets (Table S1), and probepreparation, prehybridization, hybridization and washingsteps were performed as described previously. For semi-quantitative RT-PCR, total RNA was treated with RNase-free rDNase I (USB Corp., Cleveland, OH, USA). In total,3 lg of RNA from spikelets was employed to preparecDNA with the use of ThermoScript reverse transcriptase(Invitrogen) at 65�C for 1.5 h, followed by RNaseHtreatment and RT-PCR with the corresponding primer sets(Table S1). SIZ1–GUS fusion transcripts were obtainedfrom total RNA of siz1 leaves using RT-PCR with theprimers in Table S1.
Real-time RT-PCR was conducted with a 7500 Fast Real-Time PCR system (Applied Biosystems, Foster City, CA,
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
NewPhytologist Research 871

USA) and SYBR Green (Applied Biosystems) according tothe manufacturer’s instructions. Each real-time experimentwith different primer pairs and ⁄ or different templates wasrepeated three times, and the expression levels of selectedgenes were normalized to the constitutive expression level ofElongation Factor 1 (EF1). Finally, the averaged relative geneexpression levels were calculated by the 2)DDCT (Livak &Schmittgen, 2001) method.
Protein extraction and analysis of protein sumoylation
Total proteins from wild-type, siz1 and several RNAi lineplants were isolated from maturing spikelets before anthesisas described previously (Chaikam & Karlson, 2008). Forsumoylation assay, 30 lg of total proteins from eachplant, separated by 4–15% gradient sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE), were usedfor western blot analysis. The membranes were probed withanti-SUMO1 antibody (Ab5316; Abcam, Cambridge, MA,USA; Kurepa et al., 2003), followed by incubation withhorseradish peroxidase-conjugated goat anti-rabbit IgG sec-ondary antibody (Thermo Fisher Scientific Inc., Waltham,MA, USA). The signals were detected using an enhancedchemiluminescence-plus detection kit (GE HealthcareBiosciences, Piscataway, NJ, USA).
Subcellular localization of SIZ1
The open reading frame of the SIZ1 C-terminus was sub-cloned in front of enhanced green fluorescent protein(eGFP) with the use of primers (Table S1) containing restric-tion sites, and was driven by the cauliflower mosaic virus35S promoter (CaMV35S). As a positive control for nucleus,cDNA of Ethylene Response Factor 4 (ERF4 ) was clonedin front of monomeric red fluorescent protein (mRFP;L. Chang and G. Y. Jauh, unpublished). The in-frame fusionconstructs were bombarded into onion epidermal cells asdescribed previously (Wang et al., 2008). Bombarded onionepidermal cells were incubated in the dark for 16 h at roomtemperature, and fluorescent signals were observed using anLSM 510 Meta confocal laser-scanning microscope (Zeiss,Thornwood, NY, USA). All images were recorded at 1024 ·1024 pixels by line two averaged scanning.
Preparation of rice anthers for pollen fertility andmicroscopic analyses
To observe the dehiscence of anthers, mature panicles werecollected during flowering and were examined under a dis-section microscope (Zeiss Stemi 2000-C Stero Microscope,Carl Zeiss MicroImaging, Thornwood, NY, USA). Forpollen fertility analysis, anthers from spikelets just beforeand 2 h after anthesis were collected. The pollen grains werethen analyzed for fertility in iodine–potassium iodide (I2–KI)
solution (Chen et al., 2007). For anther morphology analy-sis, fresh anthers from wild-type, siz1 and SIZ1-RNAi plantswere collected 2 h after anthesis and loaded on a stub. Thesamples were frozen by liquid nitrogen slush, and then trans-ferred to a sample preparation chamber at )160�C. After5 min, the temperature was raised to )130�C, and the sam-ples were fractured. The samples were then etched for10 min at )85�C. After coating at )130�C, the samples weretransferred to a scanning electron microscopy chamber andobserved at )160�C using a cryo-scanning electron micro-scope (FEI Quanta 200 SEM ⁄ Quorum Cryo SystemPP2000TR FEI, Hillsboro, OR, USA). To analyze antherdehiscence, anthers before and after anthesis were collected,fixed and sectioned as suggested in the protocol (An Ellis,2006). After staining with toluidine blue, the cross-sections(0.9 lm) were observed by light microscopy (BX51;Olympus, Tokyo, Japan) and images were recorded by cool-ing CCD (DP70; Olympus) through ·4 and ·10 (UPlanFI;Olympus) lenses.
Protein alignment and phylogenetic tree analysis
BlastP was used to search all SIZ1 orthologs in the NationalCenter for Biotechnology Information and the TIGR version6.1 rice genome annotation databases. Sequences of plant-specific SIZ proteins were then aligned using ClustalW2(Larkin et al., 2007). A phylogenetic tree was constructedwith the neighbour-joining algorithm (Saitou & Nei, 1987),as instructed in MEGA 4.0 (Kumar et al., 2008).
Results
Isolation, identification and molecular analysis of siz1mutant
Successful reproductive growth is important for crop pro-ductivity. To identify the genes essential for rice reproduc-tion and grain yield, T-DNA activation mutants obtainedfrom the Taiwan Rice Insertional Mutants (TRIM)Database (Hsing et al., 2007) were screened with the criteriaof abnormal spikelet development and sterility. One of themutants with a reduced plant height and lower seed produc-tion rate drew our attention (Fig. 1). One-quarter of the 19segregated T2 plants showed reduced plant height, increasedtiller number (Fig. 1a–c) and significantly reduced numbersof total spikelets and filled seeds (seed set percentage)(Fig. 1d,e). In addition, the panicles of the mutant did notcompletely emerge out of the leaf sheath (Fig. 1b). Manyempty seeds (sterile spikelets) found in panicles, showing awhite color without filling (Fig. 1d), may account for thelower seed production rate of the mutant. Flanking sequenceanalysis from the TRIM database indicated that the 15thof the 17 exons of SIZ1 encoding the rice SUMO E3 ligasewas disrupted by T-DNA insertion (Figs 2a, S2). We then
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
872 Research
NewPhytologist
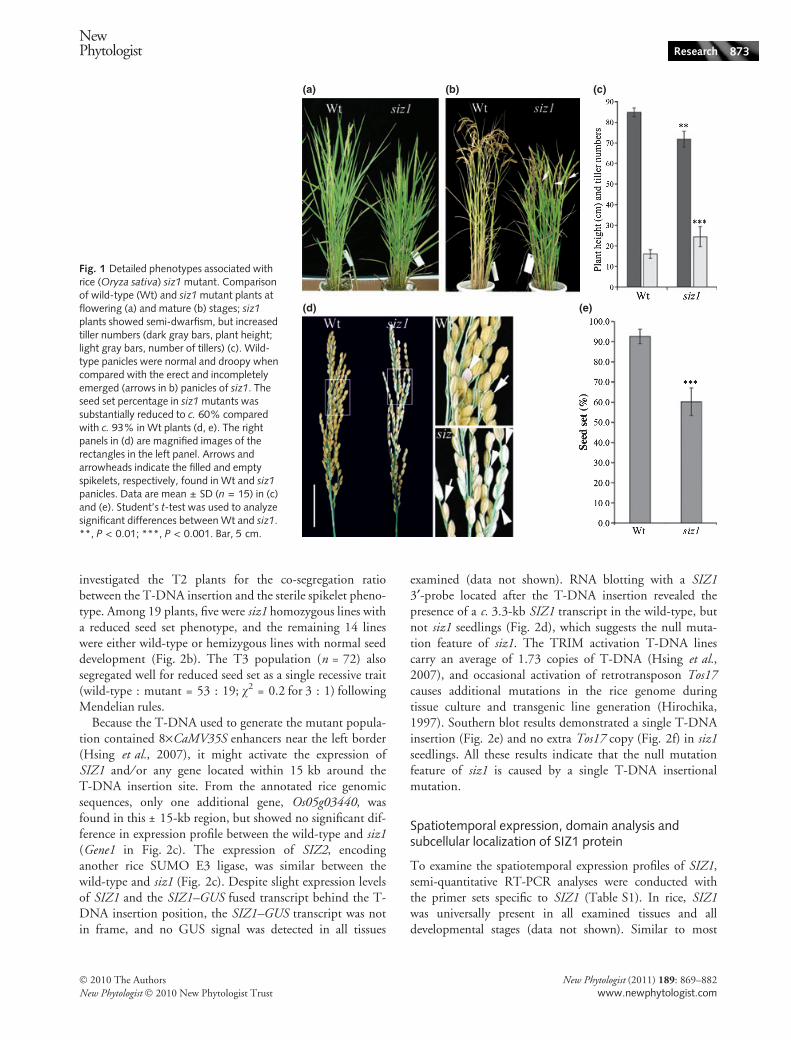
investigated the T2 plants for the co-segregation ratiobetween the T-DNA insertion and the sterile spikelet pheno-type. Among 19 plants, five were siz1 homozygous lines witha reduced seed set phenotype, and the remaining 14 lineswere either wild-type or hemizygous lines with normal seeddevelopment (Fig. 2b). The T3 population (n = 72) alsosegregated well for reduced seed set as a single recessive trait(wild-type : mutant = 53 : 19; v2 = 0.2 for 3 : 1) followingMendelian rules.
Because the T-DNA used to generate the mutant popula-tion contained 8·CaMV35S enhancers near the left border(Hsing et al., 2007), it might activate the expression ofSIZ1 and ⁄ or any gene located within 15 kb around theT-DNA insertion site. From the annotated rice genomicsequences, only one additional gene, Os05g03440, wasfound in this ± 15-kb region, but showed no significant dif-ference in expression profile between the wild-type and siz1(Gene1 in Fig. 2c). The expression of SIZ2, encodinganother rice SUMO E3 ligase, was similar between thewild-type and siz1 (Fig. 2c). Despite slight expression levelsof SIZ1 and the SIZ1–GUS fused transcript behind the T-DNA insertion position, the SIZ1–GUS transcript was notin frame, and no GUS signal was detected in all tissues
examined (data not shown). RNA blotting with a SIZ13¢-probe located after the T-DNA insertion revealed thepresence of a c. 3.3-kb SIZ1 transcript in the wild-type, butnot siz1 seedlings (Fig. 2d), which suggests the null muta-tion feature of siz1. The TRIM activation T-DNA linescarry an average of 1.73 copies of T-DNA (Hsing et al.,2007), and occasional activation of retrotransposon Tos17causes additional mutations in the rice genome duringtissue culture and transgenic line generation (Hirochika,1997). Southern blot results demonstrated a single T-DNAinsertion (Fig. 2e) and no extra Tos17 copy (Fig. 2f) in siz1seedlings. All these results indicate that the null mutationfeature of siz1 is caused by a single T-DNA insertionalmutation.
Spatiotemporal expression, domain analysis andsubcellular localization of SIZ1 protein
To examine the spatiotemporal expression profiles of SIZ1,semi-quantitative RT-PCR analyses were conducted withthe primer sets specific to SIZ1 (Table S1). In rice, SIZ1was universally present in all examined tissues and alldevelopmental stages (data not shown). Similar to most
(a)
(d) (e)
(b) (c)
Fig. 1 Detailed phenotypes associated withrice (Oryza sativa) siz1 mutant. Comparisonof wild-type (Wt) and siz1 mutant plants atflowering (a) and mature (b) stages; siz1
plants showed semi-dwarfism, but increasedtiller numbers (dark gray bars, plant height;light gray bars, number of tillers) (c). Wild-type panicles were normal and droopy whencompared with the erect and incompletelyemerged (arrows in b) panicles of siz1. Theseed set percentage in siz1 mutants wassubstantially reduced to c. 60% comparedwith c. 93% in Wt plants (d, e). The rightpanels in (d) are magnified images of therectangles in the left panel. Arrows andarrowheads indicate the filled and emptyspikelets, respectively, found in Wt and siz1panicles. Data are mean ± SD (n = 15) in (c)and (e). Student’s t-test was used to analyzesignificant differences between Wt and siz1.**, P < 0.01; ***, P < 0.001. Bar, 5 cm.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
NewPhytologist Research 873

of the characterized and annotated SIZ ⁄ PIAS (SAP andMIZ ⁄ Protein Inhibitor of Activated STAT) SUMO E3ligases from all organisms, rice SIZ1 also contains SAP,PINIT, SP-RING, a SUMO binding motif (hhhSXSaaa),a C-terminal nuclear localization signal (NLS) and theplant-specific PHD domain (plant homeodomain withC4HC3-type Zn-finger) (Fig. S2a,b). PINIT (Pro-Ile-Asn-Ile-Thr) and SP-RING, containing a zinc-finger (C2HC3),are essential for SUMO E3 ligase activity, and SAP (scaffoldattachment factors SAF-A ⁄ B, Acinus, PIAS, a helix–extended loop–helix) forms a helix–extended loop–helixstructure probably involved in DNA binding (Aravind &Koonin, 2000). Amino acid comparison revealed that allplant SIZs possess all consensus domains (Fig. S2b). ABlastP search analysis with SIZ1 amino acid sequences usedas a query was employed to generate a rooted phylogenetictree to illustrate the relationship of rice SIZs to their plantorthologs. As shown in Fig. S2c, rice SIZ1 and SIZ2 arepositioned in different clades, and AtSIZ1 is closer to riceSIZ1 than to SIZ2. Sequence similarity analysis also con-
firmed that rice SIZ1 is closely related to AtSIZ1 (Fig. S2d).The amino acid sequences of SIZ1 and SIZ2 showed highsimilarity to sorghum loci Sb09g002225 and Sb08g000380,with 65.0% and 57.1% identity, respectively (data notshown). This is probably because both rice and sorghumoriginate from the grass family and have high synteny forsharing similar panicle architecture.
In Arabidopsis, AtSIZ1 protein is primarily localized inthe nucleus, and PHR1, a MYB transcriptional activator, isits direct target in response to phosphate deficiency (Miuraet al., 2005). Therefore, the subcellular localization isimportant to reveal the potential functions of rice SIZs.Recently, Park et al. (2010) showed the nuclear localizationof OsSIZ1 and OsSIZ2 in rice protoplasts. To verifywhether NLS in the SIZ1 C-terminus is responsible for itsnuclear targeting, the NLS-containing C-terminal SIZ1 wasfused to the N-terminus of eGFP for particle bombard-ment-mediated transient assay in onion epidermis (Fig. 3a).The mRFP-tagged Ethylene Response Factor 4 (P35S::ERF4-mRFP:T35S), a known transcription factor (Ohta et al.,
(a)
(b)
(d) (e) (f)
(c)
Fig. 2 Null mutation feature of siz1 was caused by a single recessive T-DNA insertion. (a) Schematic representation of the genomic organizationof the SIZ1 gene, position of T-DNA insertion and primer sets used for genotyping and gene expression analyses. Solid boxes represent exons,and gray boxes indicate 5¢- and 3¢-untranslated regions. The T-DNA vector contains promoter-less GUS (GUS) in the right border and eightcopies of 35S enhancers (8·35S) in the left border. The primer sets used for genotyping and semi-quantitative reverse transcription-polymerasechain reaction (RT-PCR) are indicated as P1, P2 and P3, and 1F, 1R, 2F, 2R and GUS-R, respectively. (b) Genotyping of T2 seedlings with primersets indicated to reveal the wild-type (W), heterozygous (H) and homozygous (M) progenies. T-DNA F and R are the forward and reverseprimers designed within the T-DNA region, respectively. (c) Semi-quantitative RT-PCR analysis of gene expression revealed that T-DNAinsertion truncated the expression of SIZ1 in siz1. 8·35S residing in T-DNA had no effect on the expression of the neighboring gene Gene1
(Os05g03440). The expression levels of SIZ2 (Os03g50980) were similar in both wild-type and siz1. (d) Northern blot with the 3¢-proberevealed the absence of the expected SIZ1 transcript (3.3 kb) in the siz1 mutant. Elongation factor 1-a (EF1) and ribosomal RNAs were used asloading controls in semi-quantitative RT-PCR (c) and northern blot analysis (d), respectively. (e) Southern blot analysis of wild-type and siz1
genomic DNA digested with EcoRI (RI) and XbaI revealed single T-DNA insertion in the siz1 mutant with GUS used as a probe. (f) Southern blotanalysis of retrotransposon Tos17 copy numbers in wild-type (Wt) and T2 segregants (W, wild-type; H, heterozygous mutant; M, homozygousmutant) genomic DNAs digested with XbaI, followed by probing with Tos17 probe. Asterisks indicate three copies of Tos17.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
874 Research
NewPhytologist
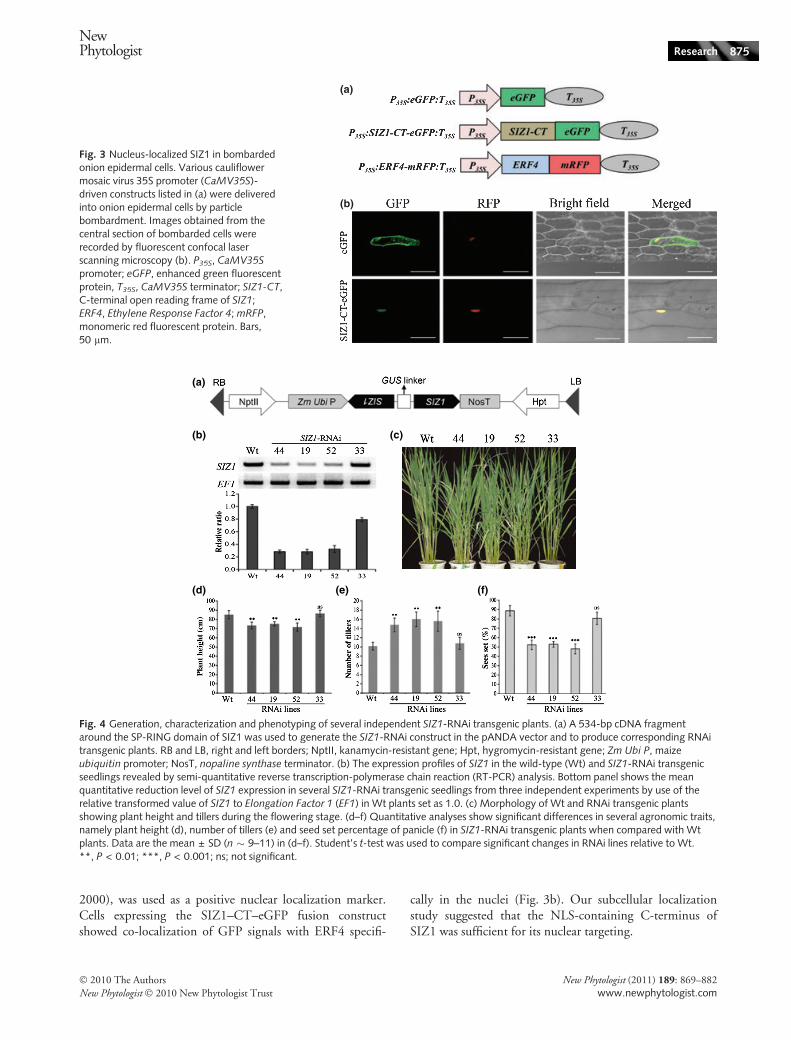
2000), was used as a positive nuclear localization marker.Cells expressing the SIZ1–CT–eGFP fusion constructshowed co-localization of GFP signals with ERF4 specifi-
cally in the nuclei (Fig. 3b). Our subcellular localizationstudy suggested that the NLS-containing C-terminus ofSIZ1 was sufficient for its nuclear targeting.
(a)
(b)
Fig. 3 Nucleus-localized SIZ1 in bombardedonion epidermal cells. Various cauliflowermosaic virus 35S promoter (CaMV35S)-driven constructs listed in (a) were deliveredinto onion epidermal cells by particlebombardment. Images obtained from thecentral section of bombarded cells wererecorded by fluorescent confocal laserscanning microscopy (b). P35S, CaMV35S
promoter; eGFP, enhanced green fluorescentprotein, T35S, CaMV35S terminator; SIZ1-CT,C-terminal open reading frame of SIZ1;ERF4, Ethylene Response Factor 4; mRFP,monomeric red fluorescent protein. Bars,50 lm.
(a)
(b)
(d) (e) (f)
(c)
Fig. 4 Generation, characterization and phenotyping of several independent SIZ1-RNAi transgenic plants. (a) A 534-bp cDNA fragmentaround the SP-RING domain of SIZ1 was used to generate the SIZ1-RNAi construct in the pANDA vector and to produce corresponding RNAitransgenic plants. RB and LB, right and left borders; NptII, kanamycin-resistant gene; Hpt, hygromycin-resistant gene; Zm Ubi P, maizeubiquitin promoter; NosT, nopaline synthase terminator. (b) The expression profiles of SIZ1 in the wild-type (Wt) and SIZ1-RNAi transgenicseedlings revealed by semi-quantitative reverse transcription-polymerase chain reaction (RT-PCR) analysis. Bottom panel shows the meanquantitative reduction level of SIZ1 expression in several SIZ1-RNAi transgenic seedlings from three independent experiments by use of therelative transformed value of SIZ1 to Elongation Factor 1 (EF1) in Wt plants set as 1.0. (c) Morphology of Wt and RNAi transgenic plantsshowing plant height and tillers during the flowering stage. (d–f) Quantitative analyses show significant differences in several agronomic traits,namely plant height (d), number of tillers (e) and seed set percentage of panicle (f) in SIZ1-RNAi transgenic plants when compared with Wtplants. Data are the mean ± SD (n � 9–11) in (d–f). Student’s t-test was used to compare significant changes in RNAi lines relative to Wt.**, P < 0.01; ***, P < 0.001; ns; not significant.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
NewPhytologist Research 875

Spikelet sterility in siz1 and SIZ1-RNAi independentlines was caused by defective anther dehiscence, butnot impaired pollen fertility
To functionally confirm that the sterile spikelet found insiz1 was truly caused by T-DNA insertion, we used an RNAiapproach to knock down the expression of SIZ1. A 534-bpcDNA fragment of SIZ1 around the SP-RING domain waschosen to generate SIZ1-RNAi transgenic plants (Figs 4a,S1, S2a). Semi-quantitative RT-PCR of more than 10 SIZ1-RNAi independent transgenic lines revealed substantialdown-regulation of SIZ1; examples are lines 44, 19 and 52shown in Fig. 4b. Phenotyping results indicated that allSIZ1-down-regulated RNAi lines exhibited reduced plantheight (Fig. 4c,d), increased tiller number (Fig. 4c,e) andreduced seed set percentages (Fig. 4f), which werecomparable with that of the siz1 mutant during the tilleringand maturity phases. The severity of the observed pheno-types in these SIZ1-RNAi independent lines was associatedwith the magnitude of the reduction in SIZ1 transcripts. InSIZ1-RNAi line 33, the expression of SIZ1 was not notablyaffected, and the line showed phenotypes similar to that ofthe wild-type. Although the RNAi target sequence is some-how unique to SIZ1, a BLAST search of the Rice GenomeAnnotation Project showed that the sequence still contains59% homology to SIZ2. However, the SIZ2 expressionprofile revealed slight down-regulation of SIZ2 found insome transgenic plants, such as line 44 (Fig. S3), whichshowed phenotypes similar to those in other SIZ1-RNAiindependent lines. These results demonstrate that the defec-tive phenotypes observed in these SIZ1-RNAi independentlines, and in siz1, were strongly associated with the degree ofdown-regulated SIZ1 transcript levels.
After anther dehiscence, pollen grains released from asingle anther are sufficient to fertilize the ovule for seed pro-duction in a self-pollinated crop such as rice. Normally, riceanthers reach the top of the spikelet before dehiscence andthen release pollen through dehiscence, and pollen grainsare shed over the stigma of the pistil just before spikeletopening to ensure successful self-pollination. Anthesis andfilament elongation were normal in the wild-type, siz1 andSIZ1-RNAi mutant plants (Fig. 5a,b). In the wild-type, theanthers dehisced completely and shed pollen before thespikelet began to close; however, c. 70% of siz1 and c. 60%of SIZ1-RNAi spikelets showed various degrees of defects inanther dehiscence even after the spikelets had closed, and avariation in anther indehiscence was observed from oneanther to six anthers of a single spikelet in siz1 and RNAiknock-down plants (Fig. 5b). To investigate the dehiscenceevents, scanning electron microscopy was used to comparethe morphological variation of anthers among the wild-type, siz1 and SIZ1-RNAi plants just after anthesis.Wild-type anthers showed normal dehiscence to releasefertile pollen grains, whereas > 50% of siz1 and SIZ1-RNAi
anthers were completely indehiscent during or even afteranthesis (Fig. 5c).
Our phenotypic results suggest that the spikelet sterilityfound in siz1 and SIZ1-RNAi plants is mainly caused byanther indehiscence. However, we could not rule out thepossibility that spikelet sterility was caused by pollensterility. Therefore, we examined pollen fertility by I2–KIstaining before spikelet opening and after spikelet closing.Wild-type and siz1 pollen grains were fertile and remainedinside the anthers before spikelet opening (Fig. 5d).Nevertheless, when compared with wild-type anthers, siz1anthers retained fertile pollen grains after anthesis (Fig. 5e)because of anther indehiscence. To further investigate thetissues showing failed anther dehiscence, we investigatedtransverse sections of anthers collected immediately beforeand after anthesis from wild-type and siz1 plants. Becausethe opening of rice theca initiates from the stomium of theapical part and near the anther wall of the basal part of thelarge locule towards the central theca (Matsui et al., 1999),we examined the top portions of the opening anthers.When compared with the wild-type anther, which showedcomplete dehiscence at the apical part of the theca (leftpanel in Fig. 5f), the siz1 anther showed only partial dehis-cence (middle panel in Fig. 5f) or complete indehiscence(right panel in Fig. 5f) for the same regions. In completelyindehiscent siz1 anthers, the endothecium and septumtissues near the stomium were still intact and left a smallcavity and tight association of the epidermal cells of twoadjacent locules after anthesis (inset in right panel inFig. 5f). These defects may be unable to support stomiumtissue opening during the final step and thus cause incom-plete anther dehiscence. All these results suggest thatSIZ1plays a significant role in regulating the developmentof endothecium tissues during anther dehiscence.
Sumoylation assay and expression of genes involved inanther dehiscence
To explore the possible biological function of SIZ1 ⁄sumoylation in anther dehiscence, we examined the sumoy-lation patterns of protein profiles from spikelets of wild-type, siz1 and two RNAi lines: RNAi-33 and RNAi-52. Forthe wild-type and RNAi-33 line with slightly reduced SIZ1expression (Fig. 4b), intense protein sumoylation profileswere observed. However, siz1 and the RNAi-52 line show-ing a reduced SIZ expression level revealed a significantreduction of sumoylation profiles in higher molecularweight proteins (Fig. 6a). This result shows the convincingcorrelation between the expression levels of SIZ1 andsumoylation profiles in the spikelets, and implies that SIZ1probably has an important function in anther dehiscence.
Next, to explore the possible molecular mechanism ofanther indehiscence in siz1, we examined the expressionprofiles of the genes potentially participating in anther
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
876 Research
NewPhytologist

dehiscence in both wild-type and siz1 plants. About twodozen genes are involved in anther dehiscence, and 12 of 23genes (orthologs of OsAID1, AtIRX1, AtNST2, ZmMADS2,AtCOI1, AtAOS and PhERS1) were found in rice after asearch of the rice genome sequences. Typical results of threebiological repeats of semiquantitative RT-PCR and real-time PCR revealed the expression of Os2g08420 (orthologof AtIRX1) and Os2g36924 (ortholog of ZmMADS2) to bedown-regulated by c. 50% in siz1 (Fig. 6b,c). This partialreduction of their expression may contribute to the partialspikelet sterility in siz1. These results suggest that SIZ1-mediated sumoylation of some regulatory players may
regulate the expression of the downstream genes participat-ing in rice anther dehiscence.
Discussion
Successful reproductive growth is important for crop pro-ductivity, and various defects in reproductive organdevelopment, such as male sterility, cause adverse cropyields (Ma, 2005). The identification and functional char-acterization of diverse mutants and their correspondinggenes with defective anther development will reveal theregulatory mechanisms of male gametophyte development.
(a)
(b)
(c)
(d)
(f)
(e)
Fig. 5 Anther indehiscence, but not impairedpollen fertility, causes spikelet sterility foundin siz1 and SIZ1-RNAi independent plants.(a) Appearance of dehiscent anthers in wild-type (Wt), siz1 and SIZ1-RNAi plants duringflowering. Spikelets of the panicles withanthesis are shown. Except for antherdehiscence, panicle morphology was normalin all plants. (b) Magnified images ofcorresponding plants marked by a square in(a) showing various degrees of antherdehiscence. Arrowheads and arrows in (a,b)mark dehiscent and indehiscent anthers,respectively. (c) Scanning electronmicrographs of anthers collected afteranthesis show anther indehiscence in T-DNA(siz1) and SIZ1-RNAi transgenic plants.Arrowheads indicate anther dehiscenceduring anthesis. Pollen grains from anthers ofboth Wt and the siz1 mutant examined byI2–KI staining before (d) and after (e)anthesis are fertile. (f) Transverse sections ofWt and siz1 anthers after anthesis. Wt anthershows complete dehiscence and sheds allpollen grains (left panel), whereas siz1
anthers show partial dehiscence (middlepanel) or complete indehiscence (right panel)in the top and central theca, respectively.Arrowheads indicate the intact endotheciumtissue in the siz1 theca. Inset shows thedetails around the cavity marked by a squareline in siz1 and the arrowhead indicates thefailure of endothecium to promotedehiscence before stomium tissue opening.C, cavity; EP, epidermis; EN, endothecium; L,locule; P, pollen grain; SP, septum; St,stomium. Bars: (a) c. 0.8 cm; (b) c. 1.2 mm;(c) 500 lm; (f) 50 lm.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
NewPhytologist Research 877

In this study, an activation-tagged rice mutant siz1 wasfunctionally characterized for its unique role in regulatingspikelet fertility. SIZ1 is a universally expressed gene andregulates both vegetative and reproductive growth anddevelopment, but has adverse effects on spikelet fertility(Figs 1, 4). Results from tight phenotypic segregation withT-DNA insertion, as well as northern and Southern blotanalyses (Fig. 2), clearly revealed siz1 as a recessive mutant,with SIZ1 disrupted by a single T-DNA insertion. SIZ1contains all the conserved domains found in most plantSIZs (Fig. S2), localizes in the nucleus (Fig. 3; Park et al.,2010) and participates in the sumoylation process (Fig. 6a).Importantly, SIZ1 plays a unique and critical role in regulat-ing anther dehiscence during rice anthesis and fertilizationprocesses (Fig. 5). The function of SIZ1 in anther dehis-cence was supported by spikelet sterility in several SIZ1-RNAi transgenic lines: the down-regulation of SIZ1 tran-scripts was closely associated with moderately reduced seedset percentage in several SIZ1-RNAi lines (Fig. 4). All these
results show that SIZ1 is involved in regulating rice antherdehiscence.
Sumoylation is emerging as one of the major post-tran-scriptional modification processes in plants by the PIAS(SIZs) group SUMO E3 ligase (Miura et al., 2007a).Arabidopsis AtSIZ1 is the best-characterized plant SUMOE3 ligase and plays critical roles under stress conditions(Miura et al., 2005, 2007b; Yoo et al., 2006), regulatingplant growth (Catala et al., 2007), cell proliferation (Catalaet al., 2007; Huang et al., 2009; Miura et al., 2010) andFLC-mediated flowering (Jin et al., 2008). Most eukaryoticSUMO E3 ligases share multiple domains SAP, PINIT andSP-RING for SUMO binding, together with a putativeNLS for nuclear localization, but plant SUMO E3 ligasescontain an extra PHD domain, a plant-specific zinc-fingerhomeodomain, which is probably involved in chromatinremodeling (Bienz, 2006) and phosphoinositide receptoractivity (Gozani et al., 2003). A study of yeast PIAS-typeULL1 ⁄ Siz1 revealed that N-terminal PINIT and SP-RING
(a)
(c)
(b)
Fig. 6 Down-regulation of spikelet sumoylation profiles and expression of several orthologous genes involved in rice (Oryza sativa) antherdehiscence in siz1 plants. (a) Total proteins extracted from maturing spikelets of wild-type (Wt), siz1, RNAi-33 and RNAi-52 lines wereanalyzed by immunoblotting with anti-SUMO1 polyclonal antibody to show the profiles of SUMO–protein conjugates. The large subunit (LS)of Rubisco (55 kDa) and free SUMO, revealed by Coomassie brilliant blue (CBB) staining and immunoblotting, respectively, were used asloading controls. Each lane contains 30 lg of total protein. (b) Twelve rice orthologs of known genes shown to be involved in antherdehiscence in different plant species were identified, and their expression profiles in Wt and siz1 plants were examined by semi-quantitativereverse transcription-polymerase chain reaction (RT-PCR) analysis. The locus numbers of individual genes were obtained from the TIGRversion 6.1 rice genome annotation database (http://rice.plantbiology.msu.edu/). Elongation Factor 1 (EF1) was selected as the loadingcontrol. The number of cycles in RT-PCR for individual genes in the experiment differed. The relative expression ratio of individual genes insiz1 was calculated by comparison with the expression in Wt. Arrows indicate the genes moderately down-regulated and the asterisk indicatesa nonspecific PCR product. (c) Real-time quantitative PCR analysis (n = 3) was used to evaluate the expression of four genes marked byarrows in (b), and endogenous EF1 was used as a reference. Wt, light gray bars; siz1, dark gray bars. Error bars, ± SD.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
878 Research
NewPhytologist

domains were required for SUMO ligase activity, whereasthe SAP domain was involved in nuclear localization(Takahashi & Kikuchi, 2005). In addition, the N-terminalULL1–GFP fusion protein was localized in the nucleus andthe cytoplasm, which implies the necessary roles of ULL1 inboth locations. In budding yeast, cytoplasmic proteinsDdc48, Vps72, Arc35 and Arc40, which localize in thecytoplasm, were confirmed to be sumoylated in vivo(Wohlschlegel et al., 2004). As shown in Fig. 3, the NLSresiding in its C-terminus of OsSIZ1 is sufficient for itsnuclear localization, although the functions of otherdomains need to be studied. Recently, structural studies ofyeast and rice SIZs revealed the presence of four helix bun-dles within SAP domains exhibiting DNA binding ability(Suzuki et al., 2009). Its nuclear localization raises the pos-sibility that it may function as a transcriptional coregulatorfor a transcriptional regulatory complex, as found in mam-malian cells (Schmidt & Muller, 2003). Further detailedstudy of the individual domains of SIZ1 would generatemore information about their specific functions in rice. Thepartial spikelet sterility found in siz1 and SIZ1-RNAi inde-pendent lines indicates that SIZ1 is important, but notdefinitely essential, for anther dehiscence. The partial effectmay be the result of the presence of an additional functionalSUMO E3 ligase, SIZ2, which can moderately complementthe lost function of SIZ1 in the siz1 mutant. Although theidentities of the potential target proteins in rice are lacking,our data suggest that SIZ1 may participate in anther dehis-cence through post-translational modification of several keytranscription factors (to be described further).
Anther dehiscence is a critical step in releasing maturepollen grains through the functions of three novel anthertissues: endothecium, septum and stomium. Anther dehis-cence is initiated after the formation of tetrads, when acavity gradually forms because of the dissociation of thestomium from the broken-down septum (Zhu et al., 2004).In the wild-type rice anther, the theca opens on the stomi-um from the apical part and near the anther wall of thebasal part of the large locule (Matsui et al., 1999). Duringanther dehiscence, the degeneration of the septum forms abilocular anther, which is followed by the enlargement ofendothecial and connective cells and strengthening of theendothecium wall for correct dehiscence. Finally, with thedriving force from the turgor pressure of swollen pollengrains and the inward bending of the locules, the stomium,a single layer of specialized cells defining the breakdown sitefor anther opening, splits to release pollen grains (Keijzer,1987; Dawson et al., 1999; Matsui et al., 1999; Sanderset al., 1999; Zhu et al., 2004). However, in siz1, with itscompletely indehiscent anther, the endothecium andseptum near the stomium were still intact after anthesis andthe stomium was still linked to the epidermal cells of twoadjacent locules; the cavity was slightly formed after thedegeneration of the septum in siz1 (Fig. 5f). These defects
may be unable to support the stomium tissue openingduring the final step of anther dehiscence. In rice, rapidswelling of pollen grains just before dehiscence generates asignificant mechanical force to rupture the septum (Matsuiet al., 1999). For example, the rice aid1 mutant showsinsufficient pressure, one of the reasons for anther indehis-cence, because of the reduced size of pollen grains (Zhuet al., 2004). Nevertheless, this insufficient pressure frompollen grains is probably not the case in siz1, because mostof the anthers from both wild-type and siz1 plants wereplump and the pollen grains were fertile (Fig. 5). A detailedseries of anatomical investigations of anther development inboth wild-type and siz1 plants will provide realistic explana-tions for anther indehiscence in siz1.
Many genes have been found to be involved in antherdevelopment (Goldberg et al., 1993), but the regulationand mechanism of anther dehiscence in rice are unclear.Recent genetic studies from plants highlighted > 20 genesparticipating in anther dehiscence, and 12 of their orthologswere found in the rice genome (Fig. 6). When comparedwith the wild-type, orthologs of AtIRX (Os2g08420) andZmMADS2 (Os2g36924) were down-regulated in siz1(Fig. 6). Remarkably, maize ZmMADS2, also highlyexpressed in anther endothecium and connective tissuesbefore dehiscence and in mature pollen after dehiscence, isessential for anther dehiscence and pollen maturation(Schreiber et al., 2004). In addition, large amounts ofZmMADS2 protein accumulate in degenerating endothe-cial and connective cells. Both AtIRX and AtNST1 areinvolved in secondary wall thickening in the anther andsubsequent dehiscence (Mitsuda et al., 2005), and theirexpression is up-regulated by AtMYB26 during secondarywall thickening (Yang et al., 2007). Exploring the existenceof putative MYB binding sites in their promoter regions isof importance. From cis-element prediction (Prestridge,1991; Higo et al., 1999), the promoter regions ofOs2g08420 and Os2g36924 contain putative MYB bindingsites. All these results suggest that defects in the develop-ment of endothecium and connective tissues regulated bythe above genes may cause anther indehiscence in the siz1mutant, and imply that an unknown MYB factor may bethe candidate of SIZ1. A few MYB-class transcription fac-tors, such as OsAID1 (MYB) and AtMYB26, regulateanther dehiscence in rice and Arabidopsis. Phenotypes ofaid1 anthers share high similarity with the siz1 dehiscentanther. From the above results, we hypothesize that theMYB transcription factors, such as AID1 or rice orthologsof AtMYB26, may be the targets of SIZ1 for post-transla-tional modification, which is essential for their function inregulating the expression of the downstream genes involvedin secondary wall thickening in the endothecium and con-nective tissues during rice anther dehiscence. As SIZ1 is anuclear protein, SIZ1 may function as a SUMO E3 ligaseto indirectly sumoylate the upstream putative transcrip-
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
NewPhytologist Research 879

tional factor. Or, it may function as a transcriptional coreg-ulator, as found in mammalian cells, to directly sumoylateother components of this transcriptional complex and up-regulate the expression of AtIRX (Os2g08420) andZmMADS2 (Os2g36924) orthologs before anther dehis-cence. Nevertheless, additional genetic and biochemicalstudies are necessary to illuminate the cellular and molecu-lar mechanisms of these genes and their sumoylation sitesrecognized by SIZ1 for anther dehiscence. Interestingly,Arabidopsis atsiz1 also showed a reduced number of siliquesin addition to slow growth and development (Catala et al.,2007); however, the anther development and dehiscence inthe atsiz1 mutant were not fully characterized. It will beinteresting to explore whether SIZ1 plays a universal func-tion in the regulation of anther dehiscence in both monocotand dicot plants.
In conclusion, by using T-DNA activation mutants withdefective spikelet development and sterility, we have identi-fied a rice gene, SIZ1, encoding a nucleus-localized riceSUMO E3 ligase, and have provided the first preliminaryevidence of its role in controlling anther dehiscence in rice,particularly the failure of the degeneration of the endothe-cium and septum near the stomium during the final step ofanther dehiscence.
Acknowledgements
We greatly appreciate the contributions of Drs Yue-IeHsing, Chyr-Guan Chern, Ming-Jen Fan and Su-May Yuin generating the TRIM database. We also thank Dr KoShimamoto (Nara Institute of Science and Technology,Japan) for sharing the pANDA-RNAi vector, Dr Shu-Hsing Wu (Institute of Plant and Microbial Biology,Academia Sinica) for her valuable comments on the manu-script, and Drs Su-May Yu (Institute of Molecular Biology,Academia Sinica) and Wann-Neng Jane (Cell Biology CoreFacility, Institute of Plant and Microbial Biology, AcademiaSinica) for assistance in rice transgenic and scanning elec-tron microscopy experiments, respectively. We also thankMs. Lin-Yun Kuang (Transgenic Plant Laboratory, Instituteof Plant and Microbial Biology, Academia Sinica) for assis-tance in particle bombardment. We appreciate Ms LauraSmales for her excellent editing work. This work wassupported by research grants from Academia Sinica(Taiwan), the National Science and Technology Programfor Agricultural Biotechnology (NSTP ⁄ AB, Taiwan), theNational Science Council (98-2313-B-001-001-MY3,Taiwan) and the Li Foundation (USA) to G.Y. Jauh.
References
An Ellis E. 2006. Solutions to the problem of substitution of ERL 4221
for vinyl cyclohexene dioxide in spurr low viscosity embedding
formulations. Microscopy Today 14: 32–33.
Aravind L, Koonin EV. 2000. SAP – a putative DNA-binding motif
involved in chromosomal organization. Trends in Biochemical Sciences25: 112–114.
Bienz M. 2006. The PHD finger, a nuclear protein-interaction domain.
Trends in Biochemical Sciences 31: 35–40.
Bots M, Vergeldt F, Wolters-Arts M, Weterings K, van As H, Mariani C.
2005. Aquaporins of the PIP2 class are required for efficient anther
dehiscence in tobacco. Plant Physiology 137: 1049–1056.
Catala R, Ouyang J, Abreu IA, Hu Y, Seo H, Zhang X, Chua NH. 2007.
The Arabidopsis E3 SUMO ligase SIZ1 regulates plant growth and
drought responses. The Plant Cell 19: 2952–2966.
Cecchetti V, Altamura MM, Falasca G, Costantino P, Cardarelli M. 2008.
Auxin regulates Arabidopsis anther dehiscence, pollen maturation, and
filament elongation. The Plant Cell 20: 1760–1774.
Chaikam V, Karlson D. 2008. Functional characterization of two cold
shock domain proteins from Oryza sativa. Plant Cell and Environment31: 995–1006.
Chen RZ, Zhao X, Shao Z, Wei Z, Wang YY, Zhu LL, Zhao J, Sun MX,
He RF, He GC. 2007. Rice UDP-glucose pyrophosphorylase1 is
essential for pollen callose deposition and its cosuppression results in a
new type of thermosensitive genic male sterility. The Plant Cell 19:
847–861.
Dawson J, Sozen E, Vizir I, Van Waeyenberge S, Wilson ZA, Mulligan
BJ. 1999. Characterization and genetic mapping of a mutation (ms35)
which prevents anther dehiscence in Arabidopsis thaliana by affecting
secondary wall thickening in the endothecium. New Phytologist 144:
213–222.
Gill G. 2005. Something about SUMO inhibits transcription. CurrentOpinion in Genetics and Development 15: 536–541.
Goldberg RB, Beals TP, Sanders PM. 1993. Anther development: basic
principles and practical applications. The Plant Cell 5: 1217–1229.
Gorguet B, Schipper D, van Lammeren A, Visser RGF, van Heusden
AW. 2009. ps-2, the gene responsible for functional sterility in tomato,
due to non-dehiscent anthers, is the result of a mutation in a novel
polygalacturonase gene. Theoretical and Applied Genetics 118:
1199–1209.
Gozani O, Karuman P, Jones DR, Ivanov D, Cha J, Lugovskoy AA, Baird
CL, Zhu H, Field SJ, Lessnick SL et al. 2003. The PHD finger of the
chromatin-associated protein ING2 functions as a nuclear
phosphoinositide receptor. Cell 114: 99–111.
Hay RT. 2005. SUMO: a history of modification. Molecular Cell 18: 1–12.
Higo K, Ugawa Y, Iwamoto M, Korenaga T. 1999. Plant cis-acting
regulatory DNA elements (PLACE) database: 1999. Nucleic AcidsResearch 27: 297–300.
Hirochika H. 1997. Retrotransposons of rice: their regulation and use for
genome analysis. Plant Molecular Biology 35: 231–240.
Ho SL, Tong WF, Yu SM. 2000. Multiple mode regulation of a cysteine
proteinase gene expression in rice. Plant Physiology 122: 57–66.
Hsing YI, Chern CG, Fan MJ, Lu PC, Chen KT, Lo SF, Sun PK, Ho SL,
Lee KW, Wang YC et al. 2007. A rice gene activation ⁄ knockout
mutant resource for high throughput functional genomics. PlantMolecular Biology 63: 351–364.
Huang LX, Yang SG, Zhang SC, Liu M, Lai JB, Qi YL, Shi SF, Wang JX,
Wang YQ, Xie Q et al. 2009. The Arabidopsis SUMO E3 ligase
AtMMS21, a homologue of NSE2 ⁄ MMS21, regulates cell proliferation
in the root. Plant Journal 60: 666–678.
Ishiguro S, Kawai-Oda A, Ueda J, Nishida I, Okada K. 2001. The
DEFECTIVE IN ANTHER DEHISCENCE1 gene encodes a novel
phospholipase A1 catalyzing the initial step of jasmonic acid
biosynthesis, which synchronizes pollen maturation, anther dehiscence,
and flower opening in Arabidopsis. The Plant Cell 13: 2191–2209.
Jin JB, Jin YH, Lee J, Miura K, Yoo CY, Kim WY, Van Oosten M, Hyun
Y, Somers DE, Lee I et al. 2008. The SUMO E3 ligase, AtSIZ1,
regulates flowering by controlling a salicylic acid-mediated floral
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
880 Research
NewPhytologist

promotion pathway and through affects on FLC chromatin structure.
Plant Journal 53: 530–540.
Johnson ES. 2004. Protein modification by SUMO. Annual Review ofBiochemistry 73: 355–382.
Keijzer CJ. 1987. The processes of anther dehiscence and pollen dispersal.
1. The opening mechanism of longitudinally dehiscing anthers. NewPhytologist 105: 487–498.
Kumar S, Nei M, Dudley J, Tamura K. 2008. MEGA: a biologist-centric
software for evolutionary analysis of DNA and protein sequences.
Briefings in Bioinformatics 9: 299–306.
Kurepa J, Walker JM, Smalle J, Gosink MM, Davis SJ, Durham TL,
Sung DY, Vierstra RD. 2003. The Small Ubiquitin-like MOdifier
(SUMO) protein modification system in Arabidopsis – accumulation of
SUMO1 and -2 conjugates is increased by stress. Journal of BiologicalChemistry 278: 6862–6872.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA,
McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R et al.2007. Clustal W and Clustal X version 2.0. Bioinformatics 23:
2947–2948.
Lee J, Nam J, Park HC, Na G, Miura K, Jin JB, Yoo CY, Baek D, Kim
DH, Jeong JC et al. 2007. Salicylic acid-mediated innate immunity in
Arabidopsis is regulated by SIZ1 SUMO E3 ligase. Plant Journal 49:
79–90.
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data
using real-time quantitative PCR and the 2)DDCt method. Methods 25:
402–408.
Ma H. 2005. Molecular genetic analyses of microsporogenesis and
microgametogenesis in flowering plants. Annual Review of Plant Biology56: 393–434.
Matsui T, Omasa K, Horie T. 1999. Mechanism of anther dehiscence in
rice (Oryza sativa L.). Annals of Botany 84: 501–506.
Matsui T, Omasa K, Horie T. 2001. The difference in sterility due to
high temperatures during the flowering period among japonica-rice
varieties. Plant Production Science 4: 90–93.
Miki D, Shimamoto K. 2004. Simple RNAi vectors for stable and
transient suppression of gene function in rice. Plant and Cell Physiology45: 490–495.
Mitsuda N, Seki M, Shinozaki K, Ohme-Takagi M. 2005. The NAC
transcription factors NST1 and NST2 of Arabidopsis regulate secondary
wall thickenings and are required for anther dehiscence. The Plant Cell17: 2993–3006.
Miura K, Jin JB, Hasegawa PM. 2007a. Sumoylation, a post-translational
regulatory-process in plants. Current Opinion in Plant Biology 10:
495–502.
Miura K, Jin JB, Lee J, Yoo CY, Stirm V, Miura T, Ashworth EN,
Bressan RA, Yun DJ, Hasegawa PM. 2007b. SIZ1-mediated
sumoylation of ICE1 controls CBF3 ⁄ DREB1A expression and freezing
tolerance in Arabidopsis. The Plant Cell 19: 1403–1414.
Miura K, Lee J, Jin JB, Yoo CY, Miura T, Hasegawa PM. 2009.
Sumoylation of ABI5 by the Arabidopsis SUMO E3 ligase SIZ1
negatively regulates abscisic acid signaling. Proceedings of the NationalAcademy of Sciences, USA 106: 5418–5423.
Miura K, Lee J, Miura T, Hasegawa PM. 2010. SIZ1 controls cell growth
and plant development in Arabidopsis through salicylic acid. Plant andCell Physiology 51: 103–113.
Miura K, Rus A, Sharkhuu A, Yokoi S, Karthikeyan AS, Raghothama
KG, Baek D, Koo YD, Jin JB, Bressan RA et al. 2005. The
Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency
responses. Proceedings of the National Academy of Sciences, USA 102:
7760–7765.
Mizuno S, Osakabe Y, Maruyama K, Ito T, Osakabe K, Sato T,
Shinozaki K, Yamaguchi-Shinozaki K. 2007. Receptor-like protein kinase2 (RPK 2) is a novel factor controlling anther development in
Arabidopsis thaliana. Plant Journal 50: 751–766.
Murray MG, Thompson WF. 1980. Rapid isolation of high molecular
weight plant DNA. Nucleic Acids Research 8: 4321–4325.
Ohta M, Ohme-Takagi M, Shinshi H. 2000. Three ethylene-responsive
transcription factors in tobacco with distinct transactivation functions.
Plant Journal 22: 29–38.
Park JH, Halitschke R, Kim HB, Baldwin IT, Feldmann KA, Feyereisen
R. 2002. A knock-out mutation in allene oxide synthase results in male
sterility and defective wound signal transduction in Arabidopsis due to a
block in jasmonic acid biosynthesis. Plant Journal 31: 1–12.
Park HC, Kim H, Koo SC, Park HJ, Cheong MS, Hong H, Baek D,
Chung WS, Kim DH, Bressan RA et al. 2010. Functional
characterization of the SIZ ⁄ PIAS-type SUMO E3 ligases, OsSIZ1 and
OsSIZ2 in rice. Plant, Cell & Environment. 33: 1923–1934.
Prestridge DS. 1991. SIGNAL SCAN: a computer program that scans
DNA sequences for eukaryotic transcriptional elements. ComputerApplications in the Biosciences 7: 203–206.
Rehman S, Yun SJ. 2006. Developmental regulation of K accumulation in
pollen, anthers, and papillae: are anther dehiscence, papillae hydration,
and pollen swelling leading to pollination and fertilization in barley
(Hordeum vulgare L.) regulated by changes in K concentration? Journalof Experimental Botany 57: 1315–1321.
Rieu I, Wolters-Arts M, Derksen J, Mariani C, Weterings K. 2003.
Ethylene regulates the timing of anther dehiscence in tobacco. Planta217: 131–137.
Saitou N, Nei M. 1987. The neighbor-joining method – a new method
for reconstructing phylogenetic trees. Molecular Biology and Evolution 4:
406–425.
Sallaud C, Meynard D, van Boxtel J, Gay C, Bes M, Brizard JP,
Larmande P, Ortega D, Raynal M, Portefaix M et al. 2003. Highly
efficient production and characterization of T-DNA plants for rice
(Oryza sativa L.) functional genomics. Theoretical and Applied Genetics106: 1396–1408.
Sanders PM, Bui AQ, Weterings K, McIntire KN, Hsu YC, Lee PY,
Truong MT, Beals TP, Goldberg RB. 1999. Anther developmental
defects in Arabidopsis thaliana male-sterile mutants. Sexual PlantReproduction 11: 297–322.
Sanders PM, Lee PY, Biesgen C, Boone JD, Beals TP, Weiler EW,
Goldberg RB. 2000. The Arabidopsis DELAYED DEHISCENCE1 gene
encodes an enzyme in the jasmonic acid synthesis pathway. The PlantCell 12: 1041–1061.
Schmidt D, Muller S. 2003. PIAS ⁄ SUMO: new partners in
transcriptional regulation. Cellular and Molecular Life Sciences 60:
2561–2574.
Schreiber DN, Bantin J, Dresselhaus T. 2004. The MADS box
transcription factor ZmMADS2 is required for anther and pollen
maturation in maize and accumulates in apoptotic bodies during anther
dehiscence. Plant Physiology 134: 1069–1079.
Scott RJ, Spielman M, Dickinson HG. 2004. Stamen structure and
function. The Plant Cell 16: S46–S60.
Steiner-Lange S, Unte US, Eckstein L, Yang CY, Wilson ZA, Schmelzer
E, Dekker K, Saedler H. 2003. Disruption of Arabidopsis thalianaMYB26 results in male sterility due to non-dehiscent anthers. PlantJournal 34: 519–528.
Suzuki R, Shindo H, Tase A, Kikuchi Y, Shimizu M, Yamazaki T. 2009.
Solution structures and DNA binding properties of the N-terminal SAP
domains of SUMO E3 ligases from Saccharomyces cerevisiae and Oryzasativa. Proteins 75: 336–347.
Takahashi Y, Kikuchi Y. 2005. Yeast PIAS-type ULL1 ⁄ SIZ1 is composed
of sumo ligase and regulatory domains. Journal of Biological Chemistry280: 35822–35828.
Thompson EP, Wilkins C, Demidchik V, Davies JM, Glover BJ. 2009.
An Arabidopsis flavonoid transporter is required for anther
dehiscence and pollen development. Journal of Experimental Botany 61:
439–451.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
NewPhytologist Research 881

Wang Y, Kumar PP. 2007. Characterization of two ethylene receptors
PhERS1 and PhETR2 from petunia: PhETR2 regulates timing of
anther dehiscence. Journal of Experimental Botany 58: 533–544.
Wang HJ, Wan AR, Jauh GY. 2008. An actin-binding protein, LlLIM1,
mediates calcium and hydrogen regulation of actin dynamics in pollen
tubes. Plant Physiology 147: 1619–1636.
Wohlschlegel JA, Johnson ES, Reed SI, Yates JR, 3rd. 2004. Global
analysis of protein sumoylation in Saccharomyces cerevisiae. Journal ofBiological Chemistry 279: 45662–45668.
Yang C, Xu Z, Song J, Conner K, Vizcay Barrena G, Wilson ZA. 2007.
Arabidopsis MYB26 ⁄ MALE STERILE35 regulates secondary thickening
in the endothecium and is essential for anther dehiscence. The Plant Cell19: 534–548.
Yasuor H, Abu-Abied M, Belausov E, Madmony A, Sadot E, Riov J,
Rubin B. 2006. Glyphosate-induced anther indehiscence in cotton is
partially temperature dependent and involves cytoskeleton and
secondary wall modifications and auxin accumulation. Plant Physiology141: 1306–1315.
Yoo CY, Miura K, Jin JB, Lee J, Park HC, Salt DE, Yun DJ, Bressan RA,
Hasegawa PM. 2006. SIZ1 small ubiquitin-like modifier E3 ligase
facilitates basal thermotolerance in Arabidopsis independent of salicylic
acid. Plant Physiology 142: 1548–1558.
Zhu QH, Ramm K, Shivakkumar R, Dennis ES, Upadhyaya NM. 2004.
The ANTHER INDEHISCENCE1 gene encoding a single MYB domain
protein is involved in anther development in rice. Plant Physiology 135:
1514–1525.
Supporting Information
Additional supporting information may be found in theonline version of this article.
Fig. S1 Amino acid sequence with the specific domains ofrice SIZ1.
Fig. S2 Schematic diagram of SIZ protein domains; aminoacid comparison of SIZ1 protein with its orthologs in otherplant species.
Fig. S3 Expression profile of SIZ2 in wild-type and severalSIZ1-RNAi transgenic lines.
Table S1 Primers used in this study
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting informationsupplied by the authors. Any queries (other than missingmaterial) should be directed to the New Phytologist CentralOffice.
� 2010 The Authors
New Phytologist � 2010 New Phytologist Trust
New Phytologist (2011) 189: 869–882
www.newphytologist.com
882 Research
NewPhytologist
Related Documents


![SIZ1 Small Ubiquitin-Like Modifier E3 Ligase …...SIZ1 Small Ubiquitin-Like Modifier E3 Ligase Facilitates Basal Thermotolerance in Arabidopsis Independent of Salicylic Acid1[W][OA]](https://static.cupdf.com/doc/110x72/5f808b34f08f5c13890b6672/siz1-small-ubiquitin-like-modiier-e3-ligase-siz1-small-ubiquitin-like-modiier.jpg)








