Ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO): A long-lived protein in the deep ocean Mo ´ nica V. Orellana, a,* and Dennis A. Hansell b a Institute for Systems Biology, Seattle, Washington b University of Miami, Rosenstiel School of Marine and Atmospheric Science, Miami, Florida Abstract We demonstrate that the distribution of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) in the deep North Pacific is a unique tracer for the accumulation of biochemically identifiable organic residue of the export flux. RuBisCO is found both dissolved and assembled in microgels in a dynamic gel-to-dissolved-to-gel continuum that may protect RuBisCO from degradation in the water column. High concentrations are located below biologically productive equatorial and subarctic systems, and low concentrations are associated with the subtropical gyre. RuBisCO tracks the advective transport of export products along deep circulation pathways of the ocean interior, serving as a quantifiable biochemical tracer of modern organic carbon exported to and resident in the deep ocean. The fate of most organic matter exported by sinking to the deep ocean interior is mineralization, which contributes to the elevated concentrations of inorganic carbon and nutrients found there. These mineralization products are returned to the ocean surface with the vertical circulation of the ocean, either to be re-assimilated into biomass and eventually returned to depth to complete the cycle or, in the case of dissolved inorganic carbon, to be exchanged with the atmosphere as CO 2 . A different and poorly understood fate of the exported material is to accumulate in the suspended particulate, gel, and dissolved phases as organic matter that is highly resistant to biological decay. In its most refractory form, the carbon is locked in the ocean in a stable state, unavailable for exchange with the atmosphere for periods longer than the circulation time of the ocean (Williams and Druffel 1987). The accumulated organic matter holds molecular information on the ocean’s major biogeochem- ical processes that has proven difficult to extract (Hansell et al. 2009); most of the organic material undergoes chemical and/or structural modifications that disguise much of the information originally held in its biochemical composition. Our limited ability to trace major biogeo- chemical processes and their pools has slowed assessment of the roles these processes play in the ocean carbon and nitrogen cycles. 15 N-nuclear magnetic resonance spectroscopy shows that an important subset of the nitrogen (N) containing refractory material in the ocean’s interior appears in the form of amides that resist chemical hydrolysis and biological degradation for reasons that are not well understood (Aluwihare et al. 2005). Amides are found in living organisms as proteins and N-acetyl amino sugars (e.g., chitin, peptidoglycan). In surface waters, half of the materials containing amides are proteins; however, in deep oceanic waters, nearly all amides are present in the form of proteins (Aluwihare et al. 2005). Proteins are valuable as biomarkers because their amino acid sequence provides a direct link to the coding deoxyribonucleic acid (DNA) of an organism. Proteins represent the catalytic potential and reactivity of an organism and, collectively, of an ecosystem, providing a direct link between the ocean’s biology and chemistry. Thus, the identification and quantification of biogeochemically relevant proteins in the water column provides a powerful approach to understanding the cycling of major elements such as carbon (C) and N. Genomics, as well as mass spectrometry–based proteo- mics analyses, demonstrate that the proteins sequestered in the particulate organic matter (POM) pool are phylogenet- ically and functionally diverse (Yooseph et al. 2007). However, detailed chemical characterization of marine proteins has been difficult, in part because of technological limitations (Mitchell 2010); this is particularly true for proteins present in the dissolved organic matter pool (DOM). Current analytical methods underestimate the protein content of seawater DOM because of inefficient recovery (Aluwihare and Meador 2008) or because the proteins have suffered physicochemical changes that prevent accurate quantification (Walker et al. 2011). Modifications include glucosylation, formation of liposome-like particles, encapsulation and/or absorption of peptides to organic material (Tomaszewski et al. 2011), and partitioning of different types of proteins (hydrophobic or hydrophilic) into microgel networks (Orellana et al. 2011). Hydrophobic amino acids (AAs, such as leucine, isoleucine, phenylalanine, and cysteine) create hydrophobic pockets that are important in the processes of assembly and aggregation of polymer microgels (Orellana et al. 2011; Verdugo 2012); gelation of polymers and biopolymers driven by hydrophobic effects is well characterized in aquatic media (Verdugo 2012). Poly- cationic proteins, peptides, and peptidic chains containing hydrophobic domains will preferentially assemble into marine microgels, which can anneal to form bigger aggregates (Orellana et al. 2011; Verdugo 2012). Enzyme- linked immunosorbent assay (ELISA) offers a quantitative tool with which to assess key protein pools with important * Corresponding author: [email protected] Limnol. Oceanogr., 57(3), 2012, 826–834 E 2012, by the Association for the Sciences of Limnology and Oceanography, Inc. doi:10.4319/lo.2012.57.3.0826 826

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO): A long-lived protein in
the deep ocean
Monica V. Orellana,a,* and Dennis A. Hansell b
a Institute for Systems Biology, Seattle, WashingtonbUniversity of Miami, Rosenstiel School of Marine and Atmospheric Science, Miami, Florida
Abstract
We demonstrate that the distribution of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) in thedeep North Pacific is a unique tracer for the accumulation of biochemically identifiable organic residue of theexport flux. RuBisCO is found both dissolved and assembled in microgels in a dynamic gel-to-dissolved-to-gelcontinuum that may protect RuBisCO from degradation in the water column. High concentrations are locatedbelow biologically productive equatorial and subarctic systems, and low concentrations are associated with thesubtropical gyre. RuBisCO tracks the advective transport of export products along deep circulation pathways ofthe ocean interior, serving as a quantifiable biochemical tracer of modern organic carbon exported to and residentin the deep ocean.
The fate of most organic matter exported by sinking tothe deep ocean interior is mineralization, which contributesto the elevated concentrations of inorganic carbon andnutrients found there. These mineralization products arereturned to the ocean surface with the vertical circulation ofthe ocean, either to be re-assimilated into biomass andeventually returned to depth to complete the cycle or, in thecase of dissolved inorganic carbon, to be exchanged withthe atmosphere as CO2.
A different and poorly understood fate of the exportedmaterial is to accumulate in the suspended particulate, gel,and dissolved phases as organic matter that is highlyresistant to biological decay. In its most refractory form,the carbon is locked in the ocean in a stable state,unavailable for exchange with the atmosphere for periodslonger than the circulation time of the ocean (Williams andDruffel 1987). The accumulated organic matter holdsmolecular information on the ocean’s major biogeochem-ical processes that has proven difficult to extract (Hansellet al. 2009); most of the organic material undergoeschemical and/or structural modifications that disguisemuch of the information originally held in its biochemicalcomposition. Our limited ability to trace major biogeo-chemical processes and their pools has slowed assessmentof the roles these processes play in the ocean carbon andnitrogen cycles.
15N-nuclear magnetic resonance spectroscopy shows thatan important subset of the nitrogen (N) containingrefractory material in the ocean’s interior appears in theform of amides that resist chemical hydrolysis andbiological degradation for reasons that are not wellunderstood (Aluwihare et al. 2005). Amides are found inliving organisms as proteins and N-acetyl amino sugars(e.g., chitin, peptidoglycan). In surface waters, half of thematerials containing amides are proteins; however, in deepoceanic waters, nearly all amides are present in the form ofproteins (Aluwihare et al. 2005). Proteins are valuable as
biomarkers because their amino acid sequence provides adirect link to the coding deoxyribonucleic acid (DNA) ofan organism. Proteins represent the catalytic potential andreactivity of an organism and, collectively, of an ecosystem,providing a direct link between the ocean’s biology andchemistry. Thus, the identification and quantification ofbiogeochemically relevant proteins in the water columnprovides a powerful approach to understanding the cyclingof major elements such as carbon (C) and N.
Genomics, as well as mass spectrometry–based proteo-mics analyses, demonstrate that the proteins sequestered inthe particulate organic matter (POM) pool are phylogenet-ically and functionally diverse (Yooseph et al. 2007).However, detailed chemical characterization of marineproteins has been difficult, in part because of technologicallimitations (Mitchell 2010); this is particularly true forproteins present in the dissolved organic matter pool(DOM). Current analytical methods underestimate theprotein content of seawater DOM because of inefficientrecovery (Aluwihare and Meador 2008) or because theproteins have suffered physicochemical changes that preventaccurate quantification (Walker et al. 2011). Modificationsinclude glucosylation, formation of liposome-like particles,encapsulation and/or absorption of peptides to organicmaterial (Tomaszewski et al. 2011), and partitioning ofdifferent types of proteins (hydrophobic or hydrophilic) intomicrogel networks (Orellana et al. 2011). Hydrophobicamino acids (AAs, such as leucine, isoleucine, phenylalanine,and cysteine) create hydrophobic pockets that are importantin the processes of assembly and aggregation of polymermicrogels (Orellana et al. 2011; Verdugo 2012); gelation ofpolymers and biopolymers driven by hydrophobic effects iswell characterized in aquatic media (Verdugo 2012). Poly-cationic proteins, peptides, and peptidic chains containinghydrophobic domains will preferentially assemble intomarine microgels, which can anneal to form biggeraggregates (Orellana et al. 2011; Verdugo 2012). Enzyme-linked immunosorbent assay (ELISA) offers a quantitativetool with which to assess key protein pools with important* Corresponding author: [email protected]
Limnol. Oceanogr., 57(3), 2012, 826–834
E 2012, by the Association for the Sciences of Limnology and Oceanography, Inc.doi:10.4319/lo.2012.57.3.0826
826

catalytic capacities and allocations that would permitunderstanding of key sources and transformations of DOM.
An especially important protein is ribulose-1,5-bis-phosphate carboxylase/oxygenase (RuBisCO), the mostabundant protein on Earth (Ellis 1979). RuBisCO is thekey enzyme (EC 4.1.1.39) catalyzing the first and major stepof carbon fixation in vascular and aquatic plants, algae,cyanobacteria, and phototropic and chemoautotrophicbacteria. In a paramount role, RuBisCO converts CO2 toorganic carbon in the Calvin–Benson–Bassham pathway andis responsible for 95% of marine photosynthesis. Marinemetagenomic libraries show that the marine RuBisCOinventory is phylogenetically highly diverse (Yooseph et al.2007). The enzyme’s large-subunit gene expression has beenused to investigate the distribution and activity of marinephytoplankton (Corredor et al. 2004). Although the regula-tion of RuBisCO production and activity is a highly complexprocess, the maximal rates of photosynthesis and biomassaccumulation depend largely on the enzyme’s concentration(Corredor et al. 2004). Additionally, as photosynthetic cellsallocate important amounts of nitrogen to RuBisCO, itserves as a major nitrogen storage molecule (Ellis 1979).
Using an immunological probe, RuBisCO was foundassembled in marine polymer microgels (Chin et al. 1998).Microgels are three-dimensional polymer networks heldtogether by Ca2+ ionic bonds (Chin et al. 1998; Orellanaet al. 2011) ranging from colloidal (100–200 nm) tomicrometer size (6–8 mm; Chin et al. 1998), thereforestanding at the interface between DOM and POM(Verdugo 2012). Because the thermodynamic yield ofmicrogel assembly is , 10% (Chin et al. 1998) or higher(30% in the high Arctic [Orellana et al. 2011]), polymer gelsmay account for at least 6.6 3 1016 g C in the globalorganic dissolved organic carbon (DOC) pool (estimated tobe 6.6 3 1017 g C (Hansell et al. 2009). Thus, microgelsmight be an important mechanism for transferring proteinsand DOM to the deep ocean.
One characteristic property of polymer microgels is theirability to undergo reversible volume phase transition fromswollen and hydrated to a dense condensed network. Phasetransitions occur in response to environmental parameterssuch as temperature and ionic strength (Tanaka et al. 1980;Verdugo 2012), pH (Chin et al. 1998; Orellana et al. 2011),and important climate-relevant substances, such as di-methylsulfoniopropionate (DMSP) and dimethylsulfide(DMS) (Orellana et al. 2011). The condensed phase mayrender gel-contained proteins refractory to bacterialdegradation (Orellana et al. 2011; Verdugo 2012).
The goal of this work is to understand the distribution ofbiomarker proteins in the ocean so as to develop insightsinto the export, accumulation, and transport of modernorganic carbon in the deep ocean. Here we evaluate theconcentration and distribution of RuBisCO as a modelprotein and tracer in the Pacific Ocean. We assess (1) thedistribution of RuBisCO, (2) the presence of RuBisCO inassembled microgels in the deep ocean, and (3) thephysicochemical changes that might control the bioavail-ability of RuBisCO and other proteins in the water columnand explain the resistance of marine proteins to decay in thewater column (Aluwihare et al. 2005).
Methods
Sampling and analyses—Water samples were taken at 0.5uintervals at , 36 depths from the surface to the bottom into12-liter Niskin bottles during the Climate Variability andPredictability (CLIVAR) cruise P16N in February and March2006 (Fig. 1). All samples were analyzed for silicic acidconcentrations, and about half of those (alternating stations)were analyzed for DOC and total dissolved nitrogen (TDN).Data for these variables were taken from the P16N datacollection held at http://cdiac.ornl.gov/oceans/home.html(Feely et al. 2009). Of the samples analyzed for DOC,, 800 from the upper 4000 m were also analyzed forRuBisCO concentrations (after having been stored frozen).DOC, TDN, and RuBisCO samples from , 270-m depthwere filtered prior to analyses using pre-combusted GF/Ffilters (nominal pore size of 0.7 mm) held in in-linepolycarbonate filter holders. Samples from greater depthswere analyzed whole (unfiltered) to avoid contamination.
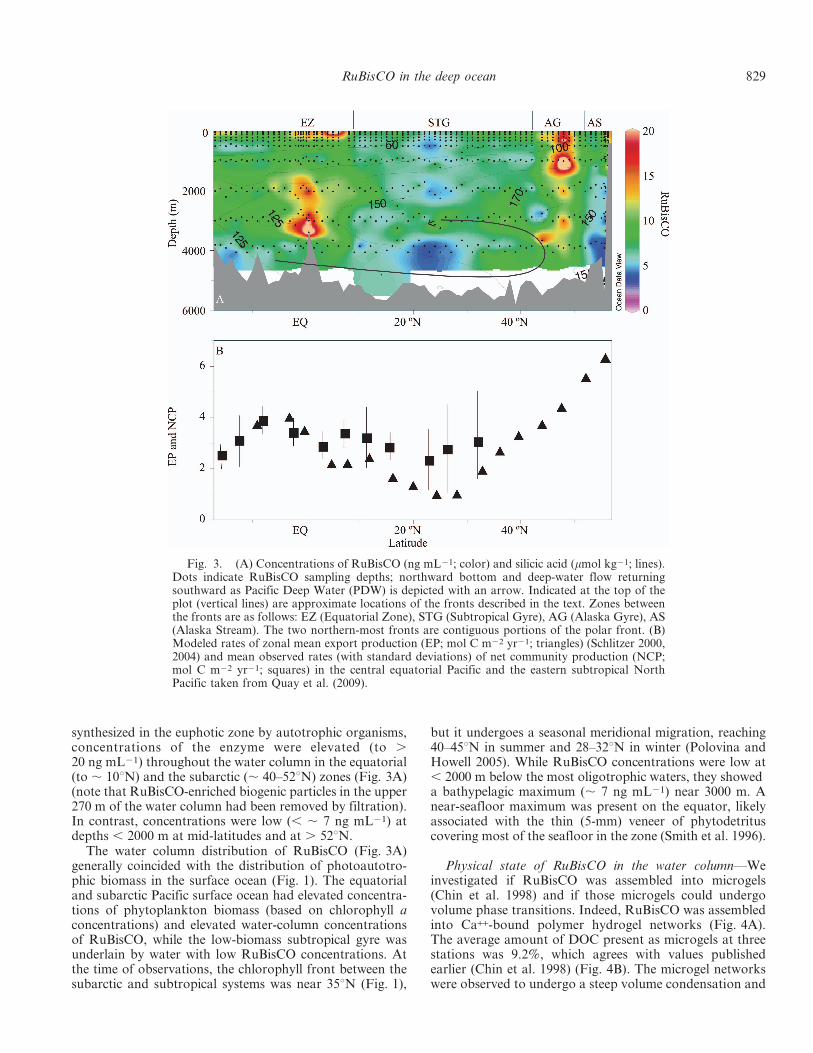
ELISA assays—We developed an indirect ELISA forRuBisCO (Orellana and Perry 1992) using an affinity-purified rabbit anti-RbcL (Agrisera AS03 037 developedagainst a conjugated synthetic peptide conserved acrossRbcL protein sequences including RuBisCO forms I [L8S8]and form II [L2-8]) and secondary goat anti-rabbit biotin-conjugated antibody (Pierce 31807). The resulting immuno-complex was detected with a poly-horseradish peroxidase(HRP) streptavidin and a fluorogenic peroxidase substratesolution (Pierce 15169) measured at 420 nm. The assay wasoptimized for antigen and antibody concentrations and forincubation periods by checkerboard titration to give amaximum signal range (0–100 ng mL21 RuBisCO). Acalibration curve with RbcL protein solutions in triplicatewas included in each assay plate (Fig. 2), with a standarddeviation , 2%. All samples were run in triplicate. The anti-rabbit anti-RbcL serum (Agrisera AS03 037) cross-reactednot only with forms I and II (including diatoms, dinoflagel-lates, cyanobacteria, photosynthetic bacteria, and archaea)but also with RuBisCO large subunit of unknown originextracted from seawater (Fig. 2, insert) and polymericmaterial assembled into microgels, that were analyzed byimmunofluorescence assays (Orellana et al. 2011).
Extraction of RuBisCO from water samples—To testcross-reactivity of the protein(s) present in seawater (ofunknown phytoplanktonic or bacterial origin) with theantibody probe we extracted RuBisCO and other proteinsfrom unconcentrated seawater samples from the PacificOcean (CLIVAR line P16N) with a high recovery immu-noaffinity column, according to Orellana and Perry (1992),with modifications. Seawater proteins were analyzed tocheck for cross-reactivity by western blot (Orellana andPerry 1992) (Fig. 2, insert). In short, proteins extracted fromseawater were separated by denaturing sodium dodecylsulfate–polyacrylamide gel electrophoresis, electrophoreti-cally transferred onto a nitrocellulose membrane, and thendetected by chemiluminescence (NovexH ECL enhancedchemiluminescent immunodetection kit, Invitrogen) usingthe biosensor that cross-reacts with RuBisCO forms I and II.
RuBisCO in the deep ocean 827
Microgel measurements—We evaluated microgel charac-teristics and properties in order to determine if the particlespresent in deep-water samples are indeed microgelsaccording to Chin et al. (1998) and Orellana et al. (2011).Samples were stained with chlortetracycline (lex 5 374 nm,lem 5 560 nm, 50 mmol L21, pH 8.0), which labels particles
containing bound Ca2+ as their polymers are stabilized byCa2+ ionic bonds, and dispersed following calcium chela-tion by ethylenediamine tetraacetic acid (pH 8). Microgelswere observed and photographed on a Delta Visionconfocal microscope and counted with a calibrated InFluxflow cytometer according to Orellana et al. (2007). Gelswere also quantified by fluorometry (Orellana et al. 2011)and the pH determined by spectrophotometry (Dicksonet al. 2007).
Polymer gel reversible volume phase transition—Swellingand condensation transition of the microgels was per-formed at 20uC and was induced by changing the pH from8.05 to 2 with HCl. Changes in gel volume were recorded aschanges in forward scatter and concentration of microgelsmeasured at a given size by an InFlux (Cytopeia) flowcytometer calibrated with Polysciences 1-mm beads (Or-ellana et al. 2007). Microgels undergo volume phasetransition to sizes below the limit of detection of theinstrument (300 nm); however, as soon as the pH isincreased the microgels can be recounted because theseRuBisCO microgels swell at higher pH. The reversibility ofthe process was tested for periods of a week.
Bacterial abundance—Bacteria and archaea in the watercolumn were stained with SYBRH green and counted withan InFlux flow cytometer (Cytopeia) calibrated withPolysciences 1-mm–size beads (Orellana et al. 2007).
Results
RuBisCO distribution—RuBisCO was assayed on samplescollected in the Pacific Ocean (Fig. 1). While RuBisCO is
Fig. 2. RuBisCO ELISA calibration curve (ng mL21). RFU,relative fluorescence units. Each datum corresponds to theaverage and standard deviation (typically , 2%) of three samples.Insert: MW, molecular weight markers; LSU, large subunit of theRuBisCO enzyme in the sample showing no cross-reactivity of theprobe with other proteins.
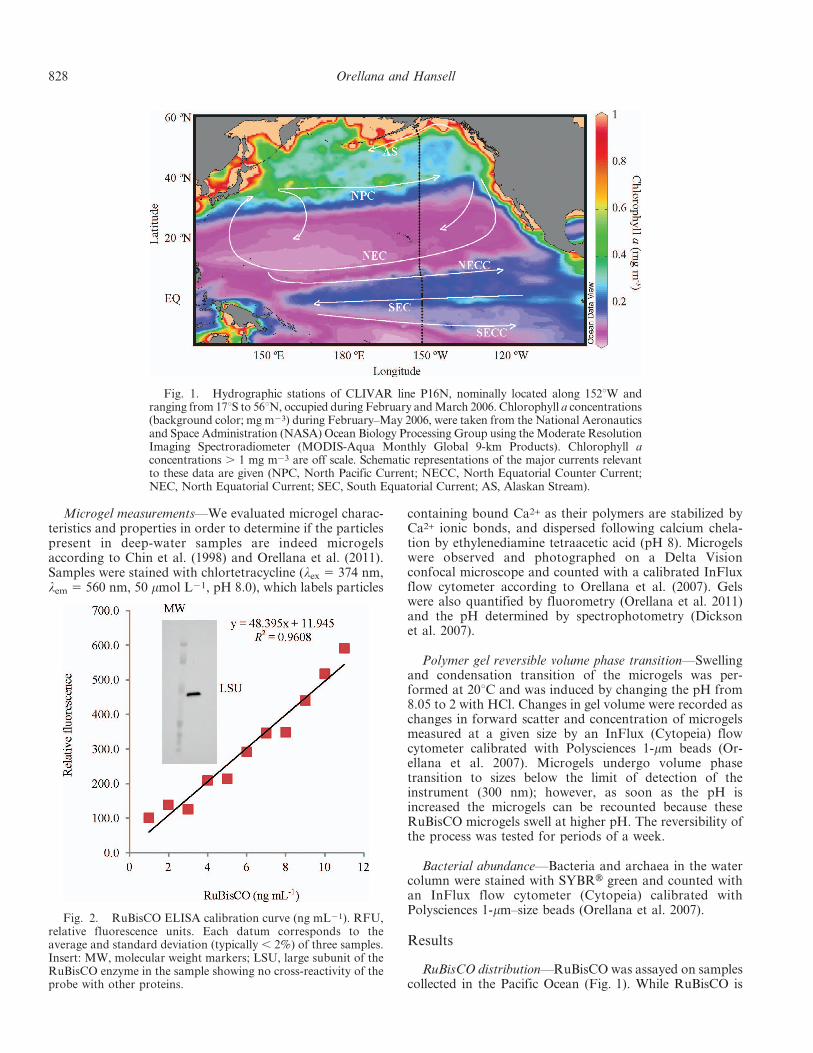
Fig. 1. Hydrographic stations of CLIVAR line P16N, nominally located along 152uW andranging from 17uS to 56uN, occupied during February and March 2006. Chlorophyll a concentrations(background color; mg m23) during February–May 2006, were taken from the National Aeronauticsand Space Administration (NASA) Ocean Biology Processing Group using the Moderate ResolutionImaging Spectroradiometer (MODIS-Aqua Monthly Global 9-km Products). Chlorophyll aconcentrations . 1 mg m23 are off scale. Schematic representations of the major currents relevantto these data are given (NPC, North Pacific Current; NECC, North Equatorial Counter Current;NEC, North Equatorial Current; SEC, South Equatorial Current; AS, Alaskan Stream).
828 Orellana and Hansell
synthesized in the euphotic zone by autotrophic organisms,concentrations of the enzyme were elevated (to .20 ng mL21) throughout the water column in the equatorial(to , 10uN) and the subarctic (, 40–52uN) zones (Fig. 3A)(note that RuBisCO-enriched biogenic particles in the upper270 m of the water column had been removed by filtration).In contrast, concentrations were low (, , 7 ng mL21) atdepths , 2000 m at mid-latitudes and at . 52uN.
The water column distribution of RuBisCO (Fig. 3A)generally coincided with the distribution of photoautotro-phic biomass in the surface ocean (Fig. 1). The equatorialand subarctic Pacific surface ocean had elevated concentra-tions of phytoplankton biomass (based on chlorophyll aconcentrations) and elevated water-column concentrationsof RuBisCO, while the low-biomass subtropical gyre wasunderlain by water with low RuBisCO concentrations. Atthe time of observations, the chlorophyll front between thesubarctic and subtropical systems was near 35uN (Fig. 1),
but it undergoes a seasonal meridional migration, reaching40–45uN in summer and 28–32uN in winter (Polovina andHowell 2005). While RuBisCO concentrations were low at, 2000 m below the most oligotrophic waters, they showeda bathypelagic maximum (, 7 ng mL21) near 3000 m. Anear-seafloor maximum was present on the equator, likelyassociated with the thin (5-mm) veneer of phytodetrituscovering most of the seafloor in the zone (Smith et al. 1996).
Physical state of RuBisCO in the water column—Weinvestigated if RuBisCO was assembled into microgels(Chin et al. 1998) and if those microgels could undergovolume phase transitions. Indeed, RuBisCO was assembledinto Ca++-bound polymer hydrogel networks (Fig. 4A).The average amount of DOC present as microgels at threestations was 9.2%, which agrees with values publishedearlier (Chin et al. 1998) (Fig. 4B). The microgel networkswere observed to undergo a steep volume condensation and
Fig. 3. (A) Concentrations of RuBisCO (ng mL21; color) and silicic acid (mmol kg21; lines).Dots indicate RuBisCO sampling depths; northward bottom and deep-water flow returningsouthward as Pacific Deep Water (PDW) is depicted with an arrow. Indicated at the top of theplot (vertical lines) are approximate locations of the fronts described in the text. Zones betweenthe fronts are as follows: EZ (Equatorial Zone), STG (Subtropical Gyre), AG (Alaska Gyre), AS(Alaska Stream). The two northern-most fronts are contiguous portions of the polar front. (B)Modeled rates of zonal mean export production (EP; mol C m22 yr21; triangles) (Schlitzer 2000,2004) and mean observed rates (with standard deviations) of net community production (NCP;mol C m22 yr21; squares) in the central equatorial Pacific and the eastern subtropical NorthPacific taken from Quay et al. (2009).
RuBisCO in the deep ocean 829
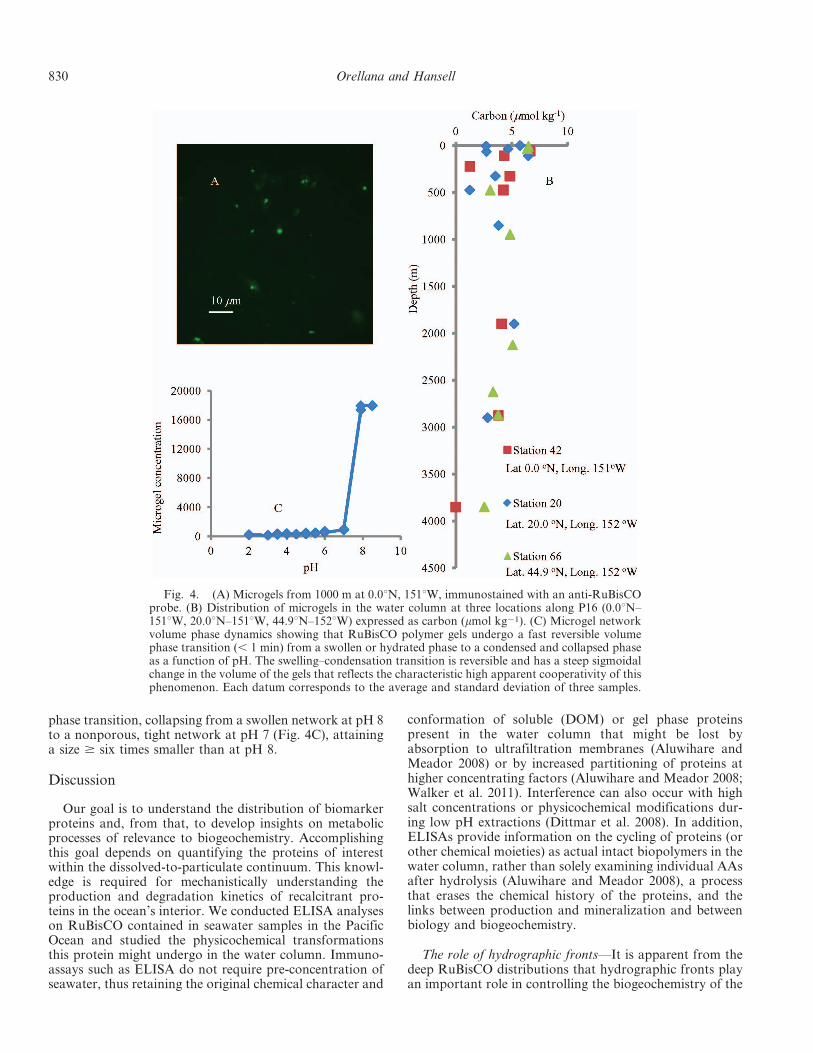
phase transition, collapsing from a swollen network at pH 8to a nonporous, tight network at pH 7 (Fig. 4C), attaininga size $ six times smaller than at pH 8.
Discussion
Our goal is to understand the distribution of biomarkerproteins and, from that, to develop insights on metabolicprocesses of relevance to biogeochemistry. Accomplishingthis goal depends on quantifying the proteins of interestwithin the dissolved-to-particulate continuum. This knowl-edge is required for mechanistically understanding theproduction and degradation kinetics of recalcitrant pro-teins in the ocean’s interior. We conducted ELISA analyseson RuBisCO contained in seawater samples in the PacificOcean and studied the physicochemical transformationsthis protein might undergo in the water column. Immuno-assays such as ELISA do not require pre-concentration ofseawater, thus retaining the original chemical character and
conformation of soluble (DOM) or gel phase proteinspresent in the water column that might be lost byabsorption to ultrafiltration membranes (Aluwihare andMeador 2008) or by increased partitioning of proteins athigher concentrating factors (Aluwihare and Meador 2008;Walker et al. 2011). Interference can also occur with highsalt concentrations or physicochemical modifications dur-ing low pH extractions (Dittmar et al. 2008). In addition,ELISAs provide information on the cycling of proteins (orother chemical moieties) as actual intact biopolymers in thewater column, rather than solely examining individual AAsafter hydrolysis (Aluwihare and Meador 2008), a processthat erases the chemical history of the proteins, and thelinks between production and mineralization and betweenbiology and biogeochemistry.
The role of hydrographic fronts—It is apparent from thedeep RuBisCO distributions that hydrographic fronts playan important role in controlling the biogeochemistry of the
Fig. 4. (A) Microgels from 1000 m at 0.0uN, 151uW, immunostained with an anti-RuBisCOprobe. (B) Distribution of microgels in the water column at three locations along P16 (0.0uN–151uW, 20.0uN–151uW, 44.9uN–152uW) expressed as carbon (mmol kg21). (C) Microgel networkvolume phase dynamics showing that RuBisCO polymer gels undergo a fast reversible volumephase transition (, 1 min) from a swollen or hydrated phase to a condensed and collapsed phaseas a function of pH. The swelling–condensation transition is reversible and has a steep sigmoidalchange in the volume of the gels that reflects the characteristic high apparent cooperativity of thisphenomenon. Each datum corresponds to the average and standard deviation of three samples.
830 Orellana and Hansell

deep underlying water masses. Fronts between major oceancurrents are important delineators of marine biogeograph-ical regimes (Belkin et al. 2009). Depending on thecirculation dynamics at the fronts, they can be sites ofnutrient upwelling with intense biological activity ofphytoplankton and increased secondary production, highautotrophic production of biogenic sinking particles, andenhanced concentration and vertical export of largephytoplankton such as diatoms (Kemp et al. 2006). Thefront near 8–10uN, separating the eastward-flowing NorthEquatorial Counter Current (south of the front) from thewestward-flowing North Equatorial Current (Fig. 1),coincided with a strong front in the deep RuBisCOdistribution (Fig. 3A), with RuBisCO enriched in theequatorial zone. The North Pacific subpolar front near40uN, separating the subtropical gyre and the cyclonicsubarctic, Alaska Gyre, also coincided with a discontinuityin the subsurface distribution of RuBisCO, with highervalues underlying the Alaska Gyre. The Alaska Stream, atthe northern border of the subarctic gyre, was underlain bylow concentrations of the protein. It is evident thathydrographic fronts affect the distribution of recentlyproduced organic matter thousands of meters below theocean surface.
Mechanisms for RuBisCO distributions—Basin-wide ex-port production in the North Pacific sums to 2–2.5 Pg Cyr21 (Feely et al. 2004; Schlitzer 2004), with highest fluxesoriginating in nutrient-enriched surface waters. The spatialpattern of export production in the North Pacific (Fig. 3B)generally tracks the water-column distributions of Ru-BisCO. For example, elevated rates of carbon export in theequatorial and subarctic systems track elevated concentra-tions of RuBisCO at depth. This relationship indicatescausality: A proposed mechanism is that particles enrichedwith autotrophic biomass are exported with their RuBisCOcontent to the abyss. As the sinking particles disaggregateand solubilize, RuBisCO is transferred to a dynamic gel-to-dissolved-to-gel continuum state (Verdugo et al. 2004),undergoing unknown biophysical and/or physicochemicaltransformations that leave RuBisCO resistant to rapiddecay. Diatoms and coccolithophorids are likely to have animportant role here since they are primary agents for rapid,deep export of organic matter in the equatorial (Honjo
et al. 2008) and subarctic Pacific (Kawahawa 2007; Honjoet al. 2008). The biosensor used in these assays cross-reactswith RuBisCO forms I and II, including diatoms andcoccolithophorids, further indicating that the deep distri-butions result from their export. Mass spectrometry aminoacid sequencing of RuBisCO extracted from seawater willbe necessary to confirm the link.
An alternative mechanism is that RuBisCO is produced insitu by chemoautotrophs. Swan et al. (2011) demonstratedthe presence of chemolithoautotrophic bacteria containingRuBisCO and sulfur-oxidizing genes associated with parti-cles in deep waters. The microgels present in the deep Pacificare a possible source of reduced sulfur, a possible substratefor the autotrophic bacteria, since they contain a highconcentration of proteins. Organically bonded sulfur is acomponent of all proteins (i.e., in the AAs cysteine andmethionine), and RuBisCO is not an exception; microgelscould also contain DMS and DMSP (Orellana et al. 2010).The Pacific Ocean ammonia-oxidizing Crenearcheota areunlikely sources of deep RuBisCO. While the expression ofammonia monooxygenase genes indicates that they areactive in the epipelagic and mesopelagic waters (Church etal. 2010), in the bathypelagic zone these organisms might bechemoorgano-heterotrophs or even completely heterotro-phic as a result of the lack of substrate (Baltar et al. 2010).
The deep-water samples (. 270 m) analyzed herecontained , 105 mL21 microbes (bacteria and or archaea)(Table 1), similar to previously reported abundances in thePacific (Nagata et al. 2010). Some of these organisms arechemoautotrophs holding their own allocation of RuBisCO(Swan et al. 2011), so their contributions to the protein’stotal concentrations can be estimated. Microbes hold, 2.1 fg N per cell (Fukuda et al. 1998); assuming that, 5% of the total N in the cells is present as RuBisCO, thenthe contribution from these organisms would reach 0.01 ngN mL21 at 105 cells mL21. Thus, , 0.1% of the observedRuBisCO might be from the protein present in theseorganisms. From this calculation, it is apparent thatRuBisCO in the water column is primarily present in thedissolved and gel phase (Verdugo et al. 2004) and less in theresident chemoautotrophs.
Tracking deep interior currents—RuBisCO concentra-tions were low at , 2000 m below the subtropical gyre, butthey showed a bathypelagic maximum (, 7 ng mL21) near
Fig. 5. Distributions of DOC (mmol C kg21; color) and silicicacid (mmol Si kg21; lines) along P16N in the North Pacific. Notenon-linearity in the color scale.
Table 1. Bacterial abundance at 44.9uN, 152uW and00.uN, 151uW.
Latitude Depth (m) Abundance
44.9uN 325.2 0.4833105
474.2 1.1773105
951.2 0.4473105
1900.3 0.7153105
2876.3 0.1083105
3850.1 0.8363105
00.0uN 324.8 0.5223105
474.9 0.1863105
850.8 0.3433105
1899.9 6.973105
2900.5 3.963105
3750.1 4.493105
RuBisCO in the deep ocean 831

3000 m. To understand this feature, the concentrations ofsilicic acid were plotted along with a two-dimensionaldepiction of bottom-water and deep-water flow (Fig. 3A).Silicic acid is enriched in the deep North Pacific as a resultof benthic and water-column dissolution of biogenic opalintroduced largely as diatoms exported from the surface(Kawahawa 2007). A bottom current (Lower CircumpolarDeep Water) with modest silicic acid concentrations (,125 mmol kg21) moves northward across the equator, thengains buoyancy in the North Pacific, to return south atmid-depths as Pacific Deep Water (PDW).
The PDW entrains silicic acid–enriched waters, carryingthe nutrient south within a core near 3000 m (Fig. 3A; notethe 150–mmol kg21 isoline). The apparent enrichment ofRuBisCO within this water mass indicates a similarentrainment and transport of the protein within thePDW. Given the role of deep circulation in determiningthe distribution of silicic acid (Fig. 3A), the elevatedconcentrations of RuBisCO deep beneath the subtropicalgyre are likewise due to transport of this protein with thedeep currents. If this inference is correct, then this proteinserves as a tracer of deep ocean flow and of export productsspreading throughout the ocean interior. The time elapsedfor deep transport from the subarctic region south acrossthe subtropics to near 20uN is not known, but a direct,meridional transport from 40uN to 20uN would require 1–2 yr of transit time at 5 cm s21. Given that these mid-basindeep waters flow zonally within gyre circulations (Wijffelset al. 1998), the lifetime for the enzyme may be decades,which in terms of refractory DOM cycling is short(Williams and Druffel 1987), but this lifetime is long interms of the protein degradation that can occur in minutes(Cooper and Hausman 2009). This phenomenon points tothe complexity of the DOM in which constituent molecules(e.g., dissolved monosaccharides) have residence times thatdiffer from the bulk radiocarbon DOC (3 yr; Repeta andAluwihare 2006).
RuBisCO physicochemical dynamics in the water column—RuBisCO in the deep ocean assembles into microgels(Fig. 4A) as its hydrophobic surfaces promote the process(Orellana et al. 2011). These biopolymeric networks canundergo volume phase transition from expanded-and-swollen to condensed-and-collapsed (Fig. 4C). The pH (7)at which this protein undergoes volume phase transition ishigher than for DOM (pH 4.5; Chin et al. 1998). Volumephase transition of microgels containing RuBisCO resultsin nonporous polymeric networks analogous to a stone-likeparticle smaller than 300 nm (flow cytometry measurement;Fig. 4C), preventing further degradation. As a solubleenzyme within chloroplasts (pH 8), RuBisCO might beexpected to be consumed or degraded by protozoa orzooplankton rather quickly. However, a pH change in thevacuoles of protozoa (pH 7 to pH 3; Laybourn-Parry 1984)or in the guts of zooplankton (pH 5 6.86–7.19 for Calanushelgolandicus; Pond et al. 1995) would induce physical–chemical alterations and microgel volume phase transitionfrom swollen and hydrated to condensed and collapsed.This process could prevent RuBisCO degradation bydigestive enzymes, as when RuBisCO did not suffer
degradation when incubated at pH , 7 for up to a week(Fig. 4C). This process would remove recently producedautotrophic biomolecules from further degradation in thewater column. Volume phase transition driven by hydro-phobic interactions can also be induced by temperaturechanges, with hydrographically relevant critical points of5uC and 24–30uC (Verdugo 2012). Furthermore, volumephase transitions can be induced by very low concentra-tions (nmol L21) of DMS and DMSP (Orellana et al. 2011).Concentrations of these chemical species are relatively high(6 to 3 nmol L21) in the equatorial Pacific Ocean and arepresent throughout the year (Bates and Quinn 1997).Moreover, during long-term transit in the water column,DOM–RuBisCO interactions might induce in situ post-translational modifications of the protein, including glyco-sylation (Mirzaei et al. 2008).
Bulk contribution of RuBisCO to the deep DOM pool—While the chemical composition of DON in the ocean hasnot been fully elucidated (Aluwihare et al. 2005; Aluwihareand Meador 2008), recent findings indicate that DONpreservation exists by virtue of a non-discriminating andrapid shunt that effectively removes recently formedautotrophic biomolecules from further recycling(McCarthy et al. 2004). Microgel phase transition maythe mechanism for shunting molecules to a protected state.By this mechanism, the marine DON pool, then, even in theabyssal ocean, would retain an unaltered signature of itsautotrophic synthesis. The concentrations of RuBisCOreported here indicate that this enzyme is an importantcomponent of the recently produced, unaltered DON pool.At the mean North Pacific RuBisCO concentration of8.9 mg m23, and with C and N contents of 47–49% and 13–15%, respectively, this enzyme contributes 0.4 mmol kg21 ofmodern C to the deep DOC pool (, 1% of the mean DOCconcentration at . 1000 m) and 0.075 mmol N kg21 to theDON pool (, 2% of mean DON concentration at .1000 m). While this latter value is surprising, as it isequivalent to the total AAs measured in the deep Pacific(Kaiser and Benner 2009), the AA recoveries from seawatercan be low and may thus underestimate the actual proteinand AA-dissolved pools. Only a fraction of AAs arehydrolyzable (Benner 2002), probably as a result ofproblems of solubility, degradation, and peptide posttrans-lational modifications during handling (Mirzaei et al.2008), highlighting the importance of understanding thephysicochemical dynamics of proteins in seawater.
If the enzyme’s concentrations in the North Pacific arerepresentative of those in other major ocean basins, theglobal ocean inventory of RuBisCO carbon is , 5.75 Pg, or0.85–1% of the 662 Pg C in the global ocean DOC inventory(Hansell et al. 2009). This small addition of modern carbonin RuBisCO will have minimal effect on the 14C compositionof deep DOC, although the sum of inputs from otherbiochemicals introduced by similar mechanisms is unknown.
Because RuBisCO, or at least a certain percentage of it,resides in the continuum between the gel and soluble phases, wecompare its distribution with the distribution of DOC in thePacific Ocean. The generally vertical alignments in thedistribution of RuBisCO (Fig. 3A) contrast with the generally
832 Orellana and Hansell

horizontal alignment of DOC (Fig. 5). DOC found in the deepNorth Pacific is mostly introduced with bottom-waterformation in the Southern Ocean, which flows northward asnear-bottom flow, as described above. Concentrations changemodestly while en route (Hansell et al. 2009), decreasing from, 40 mmol C kg21 near bottom on the equator to 35–38 mmolC kg21 in the southward return flow of the PDW (traced byelevated silicic acid concentrations in Fig. 3). RuBisCO (andpresumably other solubilized byproducts of the particulateexport flux) is added to this deep field of DOC. The true (gross)loss of the DOC introduced with bottom-water formation maybe concealed then by DOM added through solubilization ofsinking particles. Addition of DOM by solubilization is anunquantified process, but one that RuBisCO indicates isoccurring under highly productive systems.
This work demonstrates the value of RuBisCO as a tracer ofmarine dissolved and gel-phase organic matter dynamics in thedeepest ocean layers, showing for the first time an importantsingle protein in this pool and demonstrating a means by whichthis protein is protected from degradation. Previous analyticalwork has commonly been limited to assessing total combinedsingle AAs, thus erasing the chemical history and identity of theproteins and, most importantly, the link between biology andphylogenetics in the ocean surface and ocean biogeochemistry.Further work must now be directed at assessing thedistributions and turnover of functional proteins importantto other major, climate-sensitive biogeochemical processes(e.g., carbon fixation by photo- and chemoautotrophy,nitrogen fixation, denitrification).
AcknowledgmentsWe thank Amanda Pease and Aimee Desaki for technical
support, N. S. Baliga and colleagues at the Institute for SystemsBiology (ISB) and ISB for providing a stimulating environment,and two anonymous reviewers for their helpful comments. Thiswork was supported by the U.S. National Science Foundation,Division of Ocean Sciences (OCE) grants OCE-0452563 (toM.V.O.) and OCE-0752972 (to D.A.H.).
References
ALUWIHARE, L. I., AND T. MEADOR. 2008. Chemical compositionof marine dissolved organic nitrogen. Elsevier.
———, D. J. REPETA, S. PANTOJA, AND C. G. JOHNSON. 2005. Twochemically distinct pools of organic nitrogen accumulate inthe ocean. Science 308: 1007–1010, doi:10.1126/science.1108925
BALTAR, F., J. ARıSTEGUI, E. SINTES, J. M. GASOL, T. REINTHALER,AND G. J. HERNDL. 2010. Significance of non-sinkingparticulate organic carbon and dark CO2 fixation toheterotrophic carbon demand in the mesopelagic northeastAtlantic. Geophys. Res. Lett 37: L09602, doi:10.1029/2010GL043105
BATES, T. S., AND P. K. QUINN. 1997. Dimethylsulfide (DMS) inthe equatorial Pacific Ocean (1982 to 1996): Evidence of aclimate feedback? Geophys. Res. Lett. 24: 861–864,doi:10.1029/97GL00784
BELKIN, I. M., P. C. CORNILLON, AND K. SHERMAN. 2009. Fronts inlarge marine ecosystems. Prog. Oceanogr. 81: 223–236,doi:10.1016/j.pocean.2009.04.015
BENNER, R. 2002. Chemical composition and reactivity, p. 59–90.In D. HANSELL and C. CARLSON [eds.], Biogeochemistry ofmarine dissolved organic matter. Academic Press.
CHIN, W.-C., M. V. ORELLANA, AND P. VERDUGO. 1998.Spontaneous assembly of marine dissolved organic matterinto polymer gels. Nature 391: 568–572, doi:10.1038/35345
CHURCH, M. J., B. WAI, D. M. KARL, AND E. F. DELONG. 2010.Abundances of crenarchaeal amoA genes and transcripts inthe Pacific Ocean. Environ. Microbiol. 12: 679–688,doi:10.1111/j.1462-2920.2009.02108.x
COOPER, G. M., AND R. E. HAUSMAN. 2009. The cell: A molecularapproach. ACM Press & Sinauer Associates.
CORREDOR, J. E., AND OTHERS. 2004. Geochemical rate-RNAintegration study: Ribulose-1,5-bisphosphate carboxylase/oxygenase gene transcription and photosynthetic capacity ofplanktonic photoautotrophs. Appl. Environ. Microbiol. 70:5459–5468, doi:10.1128/AEM.70.9.5459-5468.2004
DICKSON, A. G., C. L. SABINE, AND J. R. E. CHRISTIAN. 2007.Guide to best practices for ocean CO2 measurements. PICESSpecial Publication.
DITTMAR, T., B. KOCH, N. HERTKORN, AND G. KATTNER. 2008. Asimple and efficient method for the solid-phase extraction ofdissolved organic matter (SPE-DOM) from seawater. Limnol.Oceanogr.: Methods 6: 230–235, doi:10.4319/lom.2008.6.230
ELLIS, R. J. 1979. The most abundant protein in the world. TrendsBiochem. Sci. 4: 241–244.
FEELY, R., C. SABINE, R. SCHLITZER, J. BULLISTER, S. MECKING,AND D. GREELEY. 2004. Oxygen utilization and organiccarbon remineralization in the upper water column of thePacific Ocean. J. Oceanogr. 60: 45–52, doi:10.1023/B:JOCE.0000038317.01279.aa
FEELY, R. A., AND OTHERS. 2009. Carbon dioxide, hydrographic,and chemical data obtained during the R/Vs Roger Revelleand Thomas G. Thompson repeat hydrography cruises in thePacific Ocean: CLIVAR CO2 Sections P16S_2005 (6 January–19 February 2005) and P16N_2006 (13 February–30 March,2006).Carbon Dioxide Information Analysis Center, OakRidge National Laboratory, U.S. Department of Energy.ORNL/CDIAC-155 NDP-090, http://cdiac.ornl.gov/ftp/ndp090/ndp090.pdf, p. 46.
FUKUDA, R., H. OGAWA, T. NAGATA, AND I. KOIKE. 1998. Directdetermination of carbon and nitrogen contents of naturalbacterial assemblages in marine environments. Appl. Environ.Microbiol. 64: 3352–3358.
HANSELL, D. A., C. A. CARLSON, D. J. REPETA, AND R. SCHLITZER.2009. Dissolved organic matter in the ocean. A controversystimulates new insights. Oceanography 22: 52–61,doi:10.5670/oceanog.2009.109
HONJO, S., S. J. MANGANINI, R. A. KRISHFIELD, AND R. FRANCOIS.2008. Particulate organic carbon fluxes to the ocean interiorand factors controlling the biological pump: A synthesis ofglobal sediment trap programs since 1983. Prog. Oceanogr.76: 217–285, doi:10.1016/j.pocean.2007.11.003
KAISER, K., AND R. BENNER. 2009. Biochemical composition andsize distribution of organic matter at the Pacific and Atlantictime-series stations. Mar. Chem. 113: 63–77, doi:10.1016/j.marchem.2008.12.004
KAWAHAWA, H. 2007. Settling particles in the central NorthPacific. Elsevier Oceanogr. Ser. 73: 107–133.
KEMP, A. E. S., R. B. PEARCE, I. GRIGOROV, J. RANCE, C. B.LANGE, P. QUILTY, AND I. SALTER. 2006. Production of giantmarine diatoms and their export at oceanic frontal zones:Implications for Si and C flux from stratified oceans. Glob.Biogeochem. Cycles 20: GB4S04, doi:10.1029/2006GB002698
LAYBOURN-PARRY, J. 1984. Protozoan plankton ecology. Chap-man and Hall.
MCCARTHY, M. D., R. BENNER, C. LEE, J. I. HEDGES, AND M. L.FOGEL. 2004. Amino acid carbon isotopic fractionationpatterns in oceanic dissolved organic matter: An unaltered
RuBisCO in the deep ocean 833

photoautotrophic source for dissolved organic nitrogen in theocean? Mar . Chem. 92: 123–134 , do i :10 .1016 /j.marchem.2004.06.021
MIRZAEI, H., J. K. MCBEE, J. WATTS, AND R. AEBERSOLD. 2008.Comparative evaluation of current peptide productionplatforms used in absolute quantification in proteomics.Mol. Cell. Proteomics 7: 813–823.
MITCHELL, P. 2010. Proteomic retrenches. Nat. Biotechnol. 28:665–670, doi:10.1038/nbt0710-665
NAGATA, T., AND OTHERS. 2010. Emerging concepts on microbialprocesses in the bathypelagic ocean—ecology, biogeochemis-try, and genomics. Deep-Sea Res. II 57: 1519–1536,doi:10.1016/j.dsr2.2010.02.019
ORELLANA, M. V., P. A. MATRAI, M. JANER, AND C. RAUSCHEN-
BERG. 2010. DMSP storage in Phaeocystis secretory vesicles. J.Phycol. 47: 112–117, doi:10.1111/j.1529-8817.2010.00936.x
———, ———, C. LECK, C. D. RAUSCHENBERG, A. M. LEE, AND
E. COZ. 2011. Marine microgels as a source of cloudcondensation nuclei in the high Arctic. Proc. Natl. Acad.Sci. USA 108: 13612–13617, doi:10.1073/pnas.1102457108
———, AND M. J. PERRY. 1992. An immunoprobe to measureRubisco concentrations and maximal photosynthetic rates ofindividual phytoplankton cells. Limnol. Oceanogr. 37:478–490, doi:10.4319/lo.1992.37.3.0478
———, T. W. PETERSEN, A. H. DIERCKS, S. DONOHOE, P.VERDUGO, AND G. VAN DEN ENGH. 2007. Marine microgels:Optical and proteomic fingerprints. Mar. Chem. 105:229–239, doi:10.1016/j.marchem.2007.02.002
POLOVINA, J. J., AND E. A. HOWELL. 2005. Ecosystem indicatorsderived from satellite remotely sensed oceanographic data forthe North Pacific. ICES. J. Mar. Sci. 62: 319–327,doi:10.1016/j.icesjms.2004.07.031
POND, D. W., R. P. HARRIS, AND C. BROWNLEE. 1995. Amicroinjection technique using a pH-sensitive dye to deter-mine the gut pH of Calanus helgolandicus. Mar. Biol. 123:75–79, doi:10.1007/BF00350325
QUAY, P. D., J. STUTSMAN, R. A. FEELY, AND L. W. JURANEK. 2009.Net community production rates across the subtropical andequatorial Pacific Ocean estimated from d13C;13C disequilib-rium. Glob. Biogeochem. Cycles 23: GB2006, doi:10.1029/2008GB003193
REPETA, D. J., AND L. I. ALUWIHARE. 2006. Radiocarbon analysisof neutral sugars in high-molecular-weight dissolved organiccarbon: Implications for organic carbon cycling. Limnol.Oceanogr. 51: 1045–1053, doi:10.4319/lo.2006.51.2.1045
SCHLITZER, R. 2000. Applying the adjoint method for globalbiogeochemical modeling, p. 107–124. In P. Kasibhatla andothers [eds.], Inverse methods in global biogeochemical cycles.AGU.
———. 2004. Export production in the equatorial and NorthPacific derived from dissolved oxygen, nutrient and carbondata. J. Oceanogr. 60: 53–62, doi:10.1023/B:JOCE.0000038318.38916.e6
SMITH, C. R., D. J. HOOVER, S. E. DOAN, R. H. POPE, D. J.DEMASTER, F. C. DOBBS, AND M. A. ALTABET. 1996.Phytodetritus at the abyssal seafloor across 10u of latitudein the central equatorial Pacific. Deep-Sea Res. II 43:1309–1338.
SWAN, B. K., AND OTHERS. 2011. Potential for chemolithoauto-trophy among ubiquitous bacteria lineages in the darkocean. Science 333: 1296–1300, doi:10.1126/science.1203690
TANAKA, T., D. FILLMORE, S.-T. SUN, I. NISHIO, G. SWISLOW, AND
A. SHAH. 1980. Phase transitions in ionic gels. Phys. Rev. Lett.45: 1636–1639, doi:10.1103/PhysRevLett.45.1636
TOMASZEWSKI, J. E., R. P. SCHWARZENBACH, AND M. SANDER. 2011.Protein encapsulation by humic substances. Environ. Sci.Technol. 45: 6003–6010, doi:10.1021/es200663h
VERDUGO, P. 2012. Marine microgels. Ann. Rev. Mar. Sci. 4:9.1–9.25, doi:10.1146/annurev-marine-120709-142759
———, A. L. ALLDREDGE, F. AZAM, D. L. KIRCHMAN, U. PASSOW,AND P. SANTSCHI. 2004. The oceanic gel phase: A bridge in theDOM–POM continuum. Mar. Chem. 92: 67–85, doi:10.1016/j.marchem.2004.06.017
WALKER, B. D., S. R. BEAUPRE, T. P. GUILDERSON, E. R. M.DRUFFEL, AND M. D. MCCARTHY. 2011. Large-volumeultrafiltration for the study of radiocarbon signatures andsize vs. age relationships in marine dissolved organic matter.Geochim. Cosmochim. Acta 75: 5187–5202, doi:10.1016/j.gca.2011.06.015
WIJFFELS, S. E., M. M. HALL, T. JOYCE, D. J. TORRES, P. HACKER,AND E. FIRING. 1998. Multiple deep gyres of the westernNorth Pacific: A WOCE section along 149uE. J. Geophys.Res. 103: 12985–13009, doi:10.1029/98JC01016
WILLIAMS, P. M., AND E. R. M. DRUFFEL. 1987. Radiocarbon indissolved organic matter in the central North Pacific Ocean.Nature 330: 246–248, doi:10.1038/330246a0
YOOSEPH, S., G. SUTTON, D. RUSCH, A. HALPERN, AND S.WILLIAMSON. 2007. The Sorcerer II Global Ocean Samplingexpedition: Expanding the universe of protein families. PLoSBiol 5: e16, doi:10.1371/journal.pbio.0050016
Associate editor: Robert R. Bidigare
Received: 05 September 2011Accepted: 03 January 2012
Amended: 21 February 2012
834 Orellana and Hansell
Related Documents











