REVIEW DESIGN-BASED STEREOLOGY IN NEUROSCIENCE C. SCHMITZ a,b * AND P. R. HOF c a Department of Psychiatry and Neuropsychology, Division of Cellular Neuroscience, University of Maastricht, P.O. Box 616, NL-6200 MD Maastricht, Netherlands b European Graduate School of Neuroscience, Maastricht, Netherlands c Department of Neuroscience and Department of Geriatrics and Adult Development, Mount Sinai School of Medicine, New York, NY 10029, USA Abstract—Quantitative morphology of the CNS has re- cently undergone major developments. In particular, sev- eral new approaches, known as design-based stereologic methods, have become available and have been success- fully applied to neuromorphological research. However, much confusion and uncertainty remains about the mean- ing, implications, and advantages of these design-based stereologic methods. The objective of this review is to provide some clarification. It does not comprise a full de- scription of all stereologic methods available. Rather, it is written by users for users, provides the reader with a guided tour through the relevant literature. It has been the experience of the authors that most neuroscientists poten- tially interested in design-based stereology need to ana- lyze volumes of brain regions, numbers of cells (neurons, glial cells) within these brain regions, mean volumes (nu- clear, perikaryal) of these cells, length densities of linear biological structures such as vessels and nerve fibers within brain regions, and the cytoarchitecture of brain re- gions (i.e. the spatial distribution of cells within a region of interest). Therefore, a comprehensive introduction to de- sign-based stereologic methods for estimating these pa- rameters is provided. It is demonstrated that results ob- tained with design-based stereology are representative for the entire brain region of interest, and are independent of the size, shape, spatial orientation, and spatial distribution of the cells to be investigated. Also, it is shown that bias (i.e. systematic error) in results obtained with design- based stereology can be limited to a minimum, and that it is possible to assess the variability of these results. These characteristics establish the advantages of design-based stereologic methods in quantitative neuromorphology. © 2004 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: brain, morphometry, neuron counts, stereology. Contents Section 1: Basic information about design-based stereology in the literature 814 Section 2: Considerations for specimen preparation for design-based stereologic analysis 815 Section 3: Laboratory equipment for design-based stereologic analyses 815 Section 4: Potential bias in results of design-based stereologic analyses 816 Section 5: Parameters that can be assessed by design-based stereology 817 5.1: Considerations about delineation of brain regions. 817 5.2: Volume of a brain region 817 5.3: Number of neurons within a given tissue volume. 818 5.4: Number of neurons within a given brain region 819 5.5: Mean cellular/nuclear volume 819 5.6: Surface area 820 5.7: Fiber length 822 5.8: Three-dimensional cytoarchitecture 822 Section 6: Variability of estimates obtained with design-based stereology 823 Section 7: Presentation of results obtained with design-based stereology 825 Section 8: The future of design-based stereology in neuroscience 827 Concluding remarks 828 Acknowledgments 828 References 828 Key questions in quantitative neuromorphology are why do we want to be able to generate numbers from microscope images and what questions can we ask and answer with numbers that we cannot ask and answer with photomicro- graphs? Histologic sections are used to define the normal or abnormal appearance of the brain tissue. In this regard histologic descriptions often use terms like “large,” “small,” “many,” “few,” “absent” or “present.” These terms are very helpful and often sufficient to describe features of sections, particularly when there are distinct differences between pathologic and physiologic conditions. However, they are not good enough to test for statistically significant changes in appearance resulting from a disease or an experimental treatment, particularly if differences between pathologic and physiologic conditions are discrete. In this case, one needs to attach numbers to the more or less subjective terms used in descriptions. Fortunately, almost all terms that are subjective can be associated with numbers, and design-based stereology is the state-of-the-art methodol- ogy for doing so. Originally, the word “stereology” was coined to de- scribe a set of methods that provide a three-dimensional interpretation of structures based on observations made *Correspondence to: C. Schmitz, Department of Psychiatry and Neu- ropsychology, Division of Cellular Neuroscience, University of Maas- tricht, P.O. Box 616, NL-6200 MD Maastricht, Netherlands. Tel: 31- 43-38-84108; fax: 31-43-367-1096. E-mail address: [email protected] (C. Schmitz). Abbreviations: BrdU, bromodeoxyuridine; BROI, brain region of inter- est; CE, coefficient of error; 2D, 2-dimensional; 3D, 3-dimensional; GLI, gray level index; LBS, linear biological structure; N V , neuron density within a given brain region; V Ref , reference space of a given brain region. Neuroscience 130 (2005) 813– 831 0306-4522/05$30.000.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2004.08.050 813

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R
D
Ca
NMb
c
DU
Acemfmispswgetlgcbwgisrttto(bics©r
K
*rt4EAeGdb
Neuroscience 130 (2005) 813–831
0d
EVIEW
ESIGN-BASED STEREOLOGY IN NEUROSCIENCE
AR
Kwingoh“hppnitanttdo
s
. SCHMITZa,b* AND P. R. HOFc
Department of Psychiatry and Neuropsychology, Division of Cellulareuroscience, University of Maastricht, P.O. Box 616, NL-6200 MDaastricht, Netherlands
European Graduate School of Neuroscience, Maastricht, Netherlands
Department of Neuroscience and Department of Geriatrics and Adultevelopment, Mount Sinai School of Medicine, New York, NY 10029,SA
bstract—Quantitative morphology of the CNS has re-ently undergone major developments. In particular, sev-ral new approaches, known as design-based stereologicethods, have become available and have been success-
ully applied to neuromorphological research. However,uch confusion and uncertainty remains about the mean-
ng, implications, and advantages of these design-basedtereologic methods. The objective of this review is torovide some clarification. It does not comprise a full de-cription of all stereologic methods available. Rather, it isritten by users for users, provides the reader with auided tour through the relevant literature. It has been thexperience of the authors that most neuroscientists poten-ially interested in design-based stereology need to ana-yze volumes of brain regions, numbers of cells (neurons,lial cells) within these brain regions, mean volumes (nu-lear, perikaryal) of these cells, length densities of lineariological structures such as vessels and nerve fibersithin brain regions, and the cytoarchitecture of brain re-ions (i.e. the spatial distribution of cells within a region of
nterest). Therefore, a comprehensive introduction to de-ign-based stereologic methods for estimating these pa-ameters is provided. It is demonstrated that results ob-ained with design-based stereology are representative forhe entire brain region of interest, and are independent ofhe size, shape, spatial orientation, and spatial distributionf the cells to be investigated. Also, it is shown that biasi.e. systematic error) in results obtained with design-ased stereology can be limited to a minimum, and that it
s possible to assess the variability of these results. Theseharacteristics establish the advantages of design-basedtereologic methods in quantitative neuromorphology.
2004 IBRO. Published by Elsevier Ltd. All rightseserved.
ey words: brain, morphometry, neuron counts, stereology.
Correspondence to: C. Schmitz, Department of Psychiatry and Neu-opsychology, Division of Cellular Neuroscience, University of Maas-richt, P.O. Box 616, NL-6200 MD Maastricht, Netherlands. Tel: �31-3-38-84108; fax: �31-43-367-1096.-mail address: [email protected] (C. Schmitz).bbreviations: BrdU, bromodeoxyuridine; BROI, brain region of inter-st; CE, coefficient of error; 2D, 2-dimensional; 3D, 3-dimensional;LI, gray level index; LBS, linear biological structure; NV, neuron
iensity within a given brain region; VRef, reference space of a givenrain region.
306-4522/05$30.00�0.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.08.050
813
ContentsSection 1: Basic information about design-based stereology in
the literature 814Section 2: Considerations for specimen preparation for
design-based stereologic analysis 815Section 3: Laboratory equipment for design-based stereologic
analyses 815Section 4: Potential bias in results of design-based
stereologic analyses 816Section 5: Parameters that can be assessed by design-based
stereology 8175.1: Considerations about delineation of brain regions. 8175.2: Volume of a brain region 8175.3: Number of neurons within a given tissue volume. 8185.4: Number of neurons within a given brain region 8195.5: Mean cellular/nuclear volume 8195.6: Surface area 8205.7: Fiber length 8225.8: Three-dimensional cytoarchitecture 822
Section 6: Variability of estimates obtained with design-basedstereology 823
Section 7: Presentation of results obtained with design-basedstereology 825
Section 8: The future of design-based stereology inneuroscience 827
Concluding remarks 828cknowledgments 828eferences 828
ey questions in quantitative neuromorphology are why doe want to be able to generate numbers from microscope
mages and what questions can we ask and answer withumbers that we cannot ask and answer with photomicro-raphs? Histologic sections are used to define the normalr abnormal appearance of the brain tissue. In this regardistologic descriptions often use terms like “large,” “small,”many,” “few,” “absent” or “present.” These terms are veryelpful and often sufficient to describe features of sections,articularly when there are distinct differences betweenathologic and physiologic conditions. However, they areot good enough to test for statistically significant changes
n appearance resulting from a disease or an experimentalreatment, particularly if differences between pathologicnd physiologic conditions are discrete. In this case, oneeeds to attach numbers to the more or less subjectiveerms used in descriptions. Fortunately, almost all termshat are subjective can be associated with numbers, andesign-based stereology is the state-of-the-art methodol-gy for doing so.
Originally, the word “stereology” was coined to de-cribe a set of methods that provide a three-dimensional
nterpretation of structures based on observations madeved.

ofHstcdtta(ame1
tpdcvg(gglllanlwp
wdtostwuri(1
dRtctletvcps
gt
Ss
Mdiawo(aRqb(a“(c“l(
Pcdfh(mcpdtbJedc
NpbMm(tlatpHatoc
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831814
n two-dimensional sections (Weibel, 1979, 1980, 1992;or an overview on the history of neuromorphometry seeaug, 1986). However, in the current use of design-basedtereology many methods (so-called probes) make use ofhree-dimensional sections. The term “design-based” indi-ates that the probes and the sampling schemes thatefine the newer methods in stereology are “designed,”
hat is, defined a priori, in such a manner that one need notake into consideration the size, shape, spatial orientation,nd spatial distribution of the cells to be investigatedWest, 2002). The elimination of the need for informationbout the geometry of the cells to be investigated results inore robust data because potential sources of systematicrrors in the calculations are eliminated (Gundersen et al.,988; West, 1993, 2002).
Design-based stereology can be divided into analyses ofhe global and local characteristics of tissues, the most im-ortant of which are volume, number, connectivity, spatialistribution, and length of linear biological structures. Theseharacteristics can be expressed as absolute values (e.g. theolume of the rat cerebellar granule cell layer, the number ofranule cells in the rat cerebellum, etc.) or as relative valuese.g. the volume fraction of the rat cerebellum occupied by theranule cell layer, the density of granule cells within theranule cell layer of the rat cerebellum, etc.). Both global and
ocal characteristics can be analyzed by a variety of stereo-ogic probes. For example, the volume of the granule cellayer is a global characteristic of the cerebellum that can benalyzed with Cavalieri’s principle and point counting tech-iques; the mean nuclear volume of the granule cells is a
ocal characteristic of the cerebellum that can be analyzedith the so-called Nucleator, the Rotator, and the Point Sam-led Intercepts method.
Design-based stereology can nowadays be viewed as aell-established methodology to reveal certain features ofevelopment, repair, natural aging, and normal anatomy of
he brain which could not be detected otherwise. (A continu-usly updated list of currently more than 1100 papers de-cribing stereologic investigations of the brain can be found inhe Stereology Literature Database [Enterprise Biology Soft-are Project; Medina, WA, USA].) Furthermore, the rigorousse of design-based stereologic methods allows one to di-ectly compare results of different quantitative histologic stud-es on the same topic, as has been noted in the literatureCoggeshall and Lekan, 1996; Coleman, 1997; Harrison,999).
It is not the intention of this review to provide a fullescription of all design-based stereologic probes available.ather, it aims to provide the reader with a guided tour
hrough the relevant literature. It contains eight sections thatan be read independently of each other. Among them, sec-ion 5 addresses the central question of what can be ana-yzed with design-based stereology, considering the experi-nce of the authors that most neuroscientists potentially in-
erested in design-based stereology need to analyzeolumes of brain regions, numbers of cells (neurons, glialells) within these brain regions, mean volumes (nuclear,erikaryal) of these cells, length densities of linear biological
tructures such as vessels and nerve fibers within brain re- cions, and the cytoarchitecture of brain regions (i.e. the spa-ial distribution of cells within a region of interest).
ection 1: Basic information about design-basedtereology in the literature
ore than 20 reviews and eight comprehensive booksedicated to design-based stereology and its applications
n neurosciences are available. Among the reviews, someddress more basic concepts of design-based stereology,hereas others deal with specific details of this methodol-gy. Within the first category, the papers by Gundersen1986, 1992), Cruz-Orive and Weibel (1990), Coggeshallnd Lekan (1996), Mayhew and Gundersen (1996) andoyet (1991) provide introductions to the basic concepts ofuantification, probes and sampling, whereas the papersy Gundersen et al. (1988), Mayhew (1992), Hyman et al.1998) and particularly West (1993, 1999, 2002) are morepplication oriented. Comparisons between earlierassumption-based” or “model based” stereologic methodsdue to their reliance on a model of cell geometry fororrection formulas) and the more recently developeddesign-based” (assumption-free or model-free) stereo-ogic methods are given in the papers by Coggeshall1999), Geuna (2000) and West (2001).
Among the specialized reviews, the papers by Dorph-etersen et al. (2001) and von Bartheld (2002) are dedi-ated to the strengths and limitations of profile- andisector-based cell counting methods, with a particularocus on the problem of tissue deformation as a result ofistologic preparations. The paper by Calhoun and Mouton2001) addresses the history and problems of length esti-ations, and the paper by Glaser and Glaser (2000) the
urrent advantages in computer-based stereology. Theaper by Oorschot et al. (1991) describes the usability ofesign-based stereology in quantitative analyses of cul-ured explants of the nervous system. Finally, the papersy Mayhew (1996), Petersen (1999), Kubinova andanacek (2001) and Kubinova et al. (2004) present strat-gies and illustrations of approaches to implementingesign-based stereologic procedures in electron micros-opy and confocal microscopy.
The books written by Howard and Reed (1998) and byurcombe et al. (1999) provide relatively short but com-rehensive overviews of stereologic methodology and cane recommended in particular to newcomers in the field.outon’s (2002) book is a more in-depth description of theany stereologic probes; the books by Vedel Jensen
1998) and by Russ and Dehoff (2000) are more dedicatedo the statistical and mathematical foundations of stereo-ogic methods. Those particularly interested in stereologyt the electron microscopy level may find useful informa-ion in the book by Reith and Mayhew (1988). Finally, theioneering books by Weibel and Elias (1967), Elias andyde (1983) and Weibel (1979, 1980) must be mentioneds the original text dealing with these issues. However,hese books do not cover the recent methodologic devel-pments for measuring global and local volumes, forounting cells, and for analyzing the spatial distribution of
ells within a given tissue or brain region.
Sf
Dmgbithalaosubsb
ptgppas5talassapsvWapopp12G(ucand
ibcos
ltdrtbmissseectgrabspcasdd
Ss
Rtwiul5Nasamop
t(Co(SIrTm3ssl
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 815
ection 2: Considerations for specimen preparationor design-based stereologic analysis
esign-based stereologic methods have been developed toake statements about structures such as an identified or-an, a definable brain region, a population of cells, or lineariological structures within a tissue. In the case of the brain,
f such statements are to be valid for an entire brain region,hen the analyzed sample of sections and microscopic fieldsas to be representative of it. This requires that one hasccess to the entire brain region, that all cells or linear bio-
ogical structures within this region are recognizable by anppropriate (sensitive and specific) staining, and that all partsf the brain region have the same chance to contribute to theample. Furthermore, if these statements are supposed to benbiased (i.e. without systematic error), the estimates muste independent of the size, shape, spatial orientation, andpatial distribution of the cells or linear biological structures toe investigated (see section 4 for details).
It is an advantage of design-based stereology that allrobes described in section 5 have been developed taking
hese prerequisites into account. Access to the entire re-ion of interest is achieved by systematic-random sam-ling of sections from exhaustive section series encom-assing the entire region (Gundersen, 1986; Gundersennd Jensen, 1987). Selection of microscopic fields in aystematic-random manner (explained in detail in section) guarantees that all parts of the region of interest havehe same chance to contribute to the sample (Gundersennd Jensen, 1987). Independence of design-based stereo-
ogic estimates from the size, shape, spatial orientation,nd spatial distribution of the neurons or linear biologicaltructures under study is achieved by the three-dimen-ional design of almost all probes described in section 5,s well as by the development of special sections (“isotro-ic uniform random” [Miles and Davy, 1976] or “vertical”ections [Baddeley et al., 1986]), or the analysis of thinirtual sections within thick brain sections (Sterio, 1984;illiams and Rakic, 1988; West, 1993). In any case, the
pplication of these new 3-dimensional (3D) stereologicrobes requires the use of thick (i.e. 3D) sections insteadf thin, 2-dimensional (2D) sections. In several studies,rotocols have been provided and tested to appropriatelyrepare such thick (3D) sections (Hatton and von Bartheld,999; Heinsen et al., 2000; Messina et al., 2000; Perl et al.,000; Schmitz et al., 2000; Dorph-Petersen et al., 2001;ardella et al., 2003). Obviously, under certain conditions
such as stereologic analyses in electron microscopy) these of thin (2D) sections cannot be circumvented. In thisase, however, certain rules have to be considered suchs the use of the so-called physical disector for countingeurons instead of the optical disector (see section 5.3 foretails).
Thus, design-based stereologic analyses cannot benitiated right away on existing sections, if they have noteen prepared adequately for design-based stereology. Toircumvent this problem, one could argue that the analysisf only one or a few sections per investigated brain is
ufficient, if they are “representative” sections and se- sected, for example, by clear anatomical landmarks. Fur-hermore, one could argue that the analysis of neuronensities (which does not require access to the entire brainegion) is as useful as the analysis of the correspondingotal numbers of neurons (requiring access to the entirerain region). However, alterations in neuron densitiesay considerably differ from alterations in the correspond-
ng total numbers of neurons, and results obtained fromystematically and randomly sampled sections may con-iderably differ from corresponding results obtained usingingle “representative” sections (for a particularly clearxample of this issue see the companion study by Schmitzt al., 2004). Finally, one could argue that at least partialorrection for the influence of size, shape, spatial orienta-ion, and spatial distribution of the neurons to be investi-ated using approaches such as the Abercrombie’s cor-ection for estimates of numbers of cells (based on thessumption that cells or cell nuclei are spheres; Abercrom-ie, 1946) may circumvent the need for using thick (3D)ections (Hedreen, 1998). In practice, however, it is rarelyossible to figure out how well the application of suchorrection methods really adjusted for bias in quantitativenalyses based on the size, shape, spatial orientation, andpatial distribution of the neurons to be investigated (foretails see, e.g. Mouton, 2002). With the correct use ofesign-based stereology this is not any longer an issue.
ection 3: Laboratory equipment for design-basedtereologic analyses
ecent developments in design-based stereology includeechniques and analyses that have become possible onlyith the introduction of computer-interfaced microscopes and
maging instrumentation, in particular, estimates of local vol-mes with the Optical Rotator (section 5.5), estimates of the
ength of linear biological structures with Space Balls (section.7), and investigations on the spatial distribution of cells withearest-Neighbor analysis (section 5.8). Furthermore, thevailability of semiautomated, computer-based stereologyystems has substantially reduced both the observer’s effortnd potential errors associated with the use of the otherethods described in section 5. Moreover, these systemsffer the combination of computer-based anatomical map-ing and rigorous stereologic estimates.
Currently the commercially available stereology sys-ems include (in alphabetical order): AIS stereology systemImaging Research, St. Catharines, Ontario, Canada),AST (Visiopharm, Hørsholm, Denmark), Digital Stereol-gy (Kinetic Imaging, Bromborough, UK), StereologerSystems Planning and Analysis, Alexandria, VA, USA),tereology Toolkit Plug-in for NOVA PRIME (Bioquant
mage Analysis Corporation, Nashville, TN, USA) and Ste-eoInvestigator (MicroBrightField, Williston, VT, USA).hese systems integrate a three-axis motor-driven speci-en stage with a computer in order to acquire data fromD structures, and implement (to various degrees) thetereologic probes described in section 5. The motorizedtage is used to map brain regions and objects that are
arger than a single microscopic field, to rapidly access
pecific locations throughout the entire region of interest
ra2wmstOy
mastac1asrmfa
upatsbswvitpstiosfifio
bmparcc
Ss
Lnbw
wwpg(bnwstsd
dtafaaotbmdtorrunlcpsmizp1bttu1tervsipaDpptfis
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831816
egardless of optical magnification, and to perform system-tic random sampling (for details see Glaser and Glaser,000). Furthermore, the microscope is usually equippedith a z axis position encoder (sometimes referred to as aicrocator), that measures accurately the actual focal po-
ition of the microscope stage. This is particularly impor-ant when using 3D probes like the Optical Disector, theptical Rotator, Space Balls and Nearest-Neighbor anal-sis (see section 5).
The tissue specimen is usually viewed on a computeronitor via a high-resolution analog CCD video camera ormodern digital camera with more than 10 frames per
econd, allowing one to focus through the tissue in realime (see Glaser and Glaser, 2000, for details). In addition,s stereologic applications now extend to confocal micros-opy (Petersen, 1999), electron microscopy (Mayhew,996), computed tomography (Pakkenberg et al., 1989)nd magnetic resonance imaging (Roberts et al., 1993),ome commercial stereology systems provide image fileeaders that are capable of accepting 3D confocal andagnetic resonance imaging image sets, as well as the file
ormats generated by a variety of electron microscopesnd flatbed scanners.
When using any of these visualization methods, theser views the tissue specimen with the stereologicrobe’s geometric overlay superimposed upon it. For ex-mple, when counting neurons with the Optical Fractiona-
or, the system generates an unbiased counting frame (seeection 5.3 for details). When investigating lengths of lineariological structures or Nearest-Neighbor analyses theoftware generates virtual spheres, whose intersectionsith the current focal plane are represented by circles ofarying diameter, as the focal plane of the microscope lens
s moved through the thick tissue sections (details in sec-ion 5.7). Then the user applies the counting rules of therobe by mouse-clicking on the sites of interaction of thepecimen with the probe’s overlay. Depending on the sys-em used, the resulting data may be presented and storedn different ways. Some systems save only the raw databtained, such as the number of counted cells. Otherystems save all of the acquired data in an object-orientedle containing the 3D locations of all points entered into thele and their relationship to one another, i.e. as a contourr a set of points of a particular type.
In summary, with the advent of advanced computerased systems, stereology became a practical laboratoryethod. Although some stereologic analyses can still beerformed with simple microscopes and minimal computerssistance, the advantages provided by design-based ste-eology are best obtained when it is integrated intoomputer-based microscopy systems that optimize dataollection, storage and analysis.
ection 4: Potential bias in results of design-basedtereologic analyses
et us assume a stereologic estimate of the total number ofeurons within a certain brain region. If this estimate coulde repeated ad infinitum and the mean of the estimates
ould equal the (unknown) true total number of neurons aithin this region, the estimates would be unbiased (i.e.ithout systematic error; for details see West, 2002). Im-ortantly, the use of design-based stereology does notuarantee unbiasedness of the corresponding estimatesGuillery and Herrup, 1997). Rather, the use of design-ased stereologic methods facilitates estimates of totalumbers of neurons, cell volumes, object lengths etc.,ithout having to take into consideration the size, shape,patial orientation, and spatial distribution of the structureso be investigated (West, 2002). To minimize bias (i.e.ystematic error) in stereologic estimates, a couple of ad-itional requirements are necessary.
First, it is well known that different fixation and embed-ing protocols can result in very different shrinkage of theissue (Bauchot, 1967; Kretschmann et al., 1982; Questernd Schröder, 1997; Dorph-Petersen et al., 2001). There-
ore estimates of the volume of brain regions or cells (butlso length estimates, investigations on neuron densitynd analyses of the spatial distribution of cells) will dependn the protocols used. In consequence, comparisons be-ween groups should be restricted to material which haseen processed under identical conditions. Note that esti-ates of total numbers of neurons (when carried out asescribed in sections 5.3 and 5.4) are not affected byissue shrinkage during fixation and embedding. Second,ne needs access to the entire brain region, the entireegion must be recognizable, and all parts of the brainegion under study must have the same chance to contrib-te to the stereologic sample. Third, the estimates musteither be affected by loss of structures at the upper or the
ower surface of the sections when hit by the knife duringutting the tissue, nor by incomplete staining of the tissue,articularly in the middle of the section thickness (seeection 5.3 for details). Fourth, particularly the use ofethacrylate and paraffin as embedding media may cause
nhomogeneous compression of tissue sections along theaxis during cutting of the tissue, resulting in differences inarticle densities along this axis (Hatton and von Bartheld,999; von Bartheld, 2002; Gardella et al., 2003). This maye a potential source of bias, particularly in estimates ofotal numbers of neurons and analyses of the spatial dis-ribution of cells. Cryostat sections have been proven to benaffected by this problem (Hatton and von Bartheld,999; Gardella et al., 2003). However, the real impact of
his problem might be much less than anticipated: in fact,stimates of the total number of pyramidal cells within theat hippocampus obtained on methacrylate sections areery similar to corresponding estimates obtained on cryo-tat sections (for details see Schmitz et al., 2000). Fifth,
rrespective of the embedding medium and the cuttingrocedure one has to consider shrinkage of the sectionslong the z axis (West et al., 1991; Messina et al., 2000;orph-Petersen et al., 2001). This shrinkage may be sur-risingly high (Schmitz et al., 2000) and may vary from oneart of a section to another. Measurements of the sectionhickness (preferably at each investigated microscopiceld) with the z axis position encoder of a stereologyystem (see section 3 for details) can give reasonably
ccurate readings of the actual section thickness, provided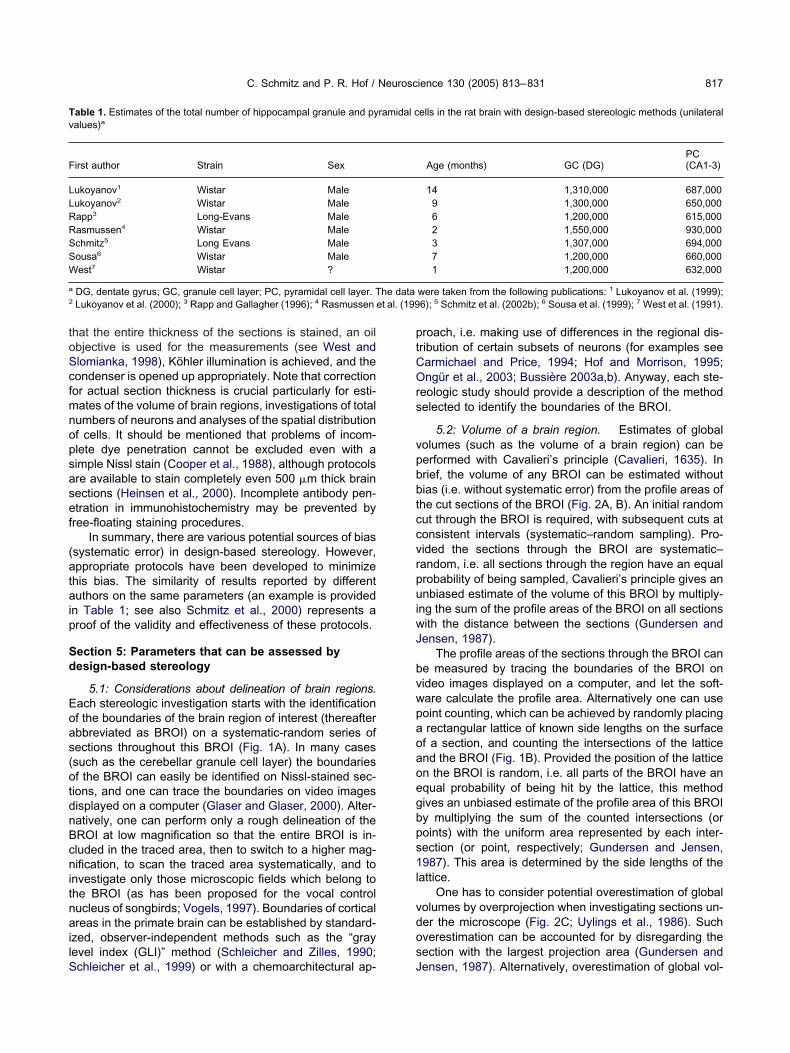
toScfmnopsasef
(ataip
Sd
Eoas(otdnBcnitnailS
ptCOrs
vpbbtccvrpuiwJ
bvwpaoaoegbps1l
vdos
Tv
F
LLRRSSW
a
2 et al. (199
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 817
hat the entire thickness of the sections is stained, an oilbjective is used for the measurements (see West andlomianka, 1998), Köhler illumination is achieved, and theondenser is opened up appropriately. Note that correctionor actual section thickness is crucial particularly for esti-ates of the volume of brain regions, investigations of totalumbers of neurons and analyses of the spatial distributionf cells. It should be mentioned that problems of incom-lete dye penetration cannot be excluded even with aimple Nissl stain (Cooper et al., 1988), although protocolsre available to stain completely even 500 �m thick brainections (Heinsen et al., 2000). Incomplete antibody pen-tration in immunohistochemistry may be prevented byree-floating staining procedures.
In summary, there are various potential sources of biassystematic error) in design-based stereology. However,ppropriate protocols have been developed to minimize
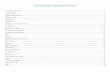
his bias. The similarity of results reported by differentuthors on the same parameters (an example is provided
n Table 1; see also Schmitz et al., 2000) represents aroof of the validity and effectiveness of these protocols.
ection 5: Parameters that can be assessed byesign-based stereology
5.1: Considerations about delineation of brain regions.ach stereologic investigation starts with the identificationf the boundaries of the brain region of interest (thereafterbbreviated as BROI) on a systematic-random series ofections throughout this BROI (Fig. 1A). In many casessuch as the cerebellar granule cell layer) the boundariesf the BROI can easily be identified on Nissl-stained sec-ions, and one can trace the boundaries on video imagesisplayed on a computer (Glaser and Glaser, 2000). Alter-atively, one can perform only a rough delineation of theROI at low magnification so that the entire BROI is in-luded in the traced area, then to switch to a higher mag-ification, to scan the traced area systematically, and to
nvestigate only those microscopic fields which belong tohe BROI (as has been proposed for the vocal controlucleus of songbirds; Vogels, 1997). Boundaries of corticalreas in the primate brain can be established by standard-
zed, observer-independent methods such as the “grayevel index (GLI)” method (Schleicher and Zilles, 1990;
able 1. Estimates of the total number of hippocampal granule and pyalues)a
irst author Strain Sex
ukoyanov1 Wistar Maleukoyanov2 Wistar Maleapp3 Long-Evans Maleasmussen4 Wistar Malechmitz5 Long Evans Maleousa6 Wistar Maleest7 Wistar ?
DG, dentate gyrus; GC, granule cell layer; PC, pyramidal cell layer. TLukoyanov et al. (2000); 3 Rapp and Gallagher (1996); 4 Rasmussen
chleicher et al., 1999) or with a chemoarchitectural ap- J
roach, i.e. making use of differences in the regional dis-ribution of certain subsets of neurons (for examples seearmichael and Price, 1994; Hof and Morrison, 1995;ngür et al., 2003; Bussière 2003a,b). Anyway, each ste-
eologic study should provide a description of the methodelected to identify the boundaries of the BROI.
5.2: Volume of a brain region. Estimates of globalolumes (such as the volume of a brain region) can beerformed with Cavalieri’s principle (Cavalieri, 1635). Inrief, the volume of any BROI can be estimated withoutias (i.e. without systematic error) from the profile areas ofhe cut sections of the BROI (Fig. 2A, B). An initial randomut through the BROI is required, with subsequent cuts atonsistent intervals (systematic–random sampling). Pro-ided the sections through the BROI are systematic–andom, i.e. all sections through the region have an equalrobability of being sampled, Cavalieri’s principle gives annbiased estimate of the volume of this BROI by multiply-
ng the sum of the profile areas of the BROI on all sectionsith the distance between the sections (Gundersen andensen, 1987).
The profile areas of the sections through the BROI cane measured by tracing the boundaries of the BROI onideo images displayed on a computer, and let the soft-are calculate the profile area. Alternatively one can useoint counting, which can be achieved by randomly placingrectangular lattice of known side lengths on the surface
f a section, and counting the intersections of the latticend the BROI (Fig. 1B). Provided the position of the latticen the BROI is random, i.e. all parts of the BROI have anqual probability of being hit by the lattice, this methodives an unbiased estimate of the profile area of this BROIy multiplying the sum of the counted intersections (oroints) with the uniform area represented by each inter-ection (or point, respectively; Gundersen and Jensen,987). This area is determined by the side lengths of the
attice.One has to consider potential overestimation of global
olumes by overprojection when investigating sections un-er the microscope (Fig. 2C; Uylings et al., 1986). Suchverestimation can be accounted for by disregarding theection with the largest projection area (Gundersen and
ells in the rat brain with design-based stereologic methods (unilateral
Age (months) GC (DG)PC(CA1-3)
14 1,310,000 687,0009 1,300,000 650,0006 1,200,000 615,0002 1,550,000 930,0003 1,307,000 694,0007 1,200,000 660,0001 1,200,000 632,000
were taken from the following publications: 1 Lukoyanov et al. (1999);6); 5 Schmitz et al. (2002b); 6 Sousa et al. (1999); 7 West et al. (1991).
ramidal c
he data
ensen, 1987). Alternatively, overestimation of global vol-

ueirnto
Cnedco3fbsrsm2
cs“isaFsyt
alotpeuapAt
(dtanrsscww(nn
Ftslsisection: sections no. 7, 17, . . .). (B) Estimate of the projection area
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831818
mes can almost be eliminated by cutting the BROI into anxhaustive series of sections of uniform thickness, select-
ng a systematic-random series of these sections with aandom start for analysis (e.g. every 10th section: sectionso. 7, 17, . . .), and performing the analysis using objec-ives with short depth of focus, with Köhler illumination, andpen condenser.
5.3: Number of neurons within a given tissue volume.utting brain tissue into sections also results in cutting theeurons (or glial cells, respectively) within this tissue. How-ver, the number of neuron fragments in the sectionsiffers from the number of neurons within the tissue. As aonsequence estimates of neuron numbers based solelyn counts of neuron fragments in sections are biased (Fig.). Numerous correction procedures have been developedor solving this problem such as Abercrombie’s formula,ased on the assumption that cells or cell nuclei arepheres (Abercrombie, 1946). In practice, however, it isarely possible to figure out how well the application ofuch correction methods really adjusted for bias in esti-ates of numbers of neurons (for details see, e.g. Mouton,002).
In design-based stereology this problem is solved byounting neurons with so-called unbiased virtual countingpaces (Schmitz and Hof, 2000; often also referred to asoptical disector” [for example see West, 1993] or “count-ng box” [Williams and Rakic, 1988]), consisting of a virtualpace within the section thickness and a so-called unbi-sed counting frame (Gundersen, 1977; Fig. 1D, E andig. 4). Alternatively, neurons can be counted with theo-called disector (Sterio, 1984) or physical disector (anal-sis of two focal planes within a thick [3D] section, or of twohin [2D] adjacent sections; Fig. 5).
A potential source of bias in neuron counts with unbi-sed virtual counting spaces are so-called lost caps, i.e.
oss of nucleoli or neurons at the upper or the lower surfacef sections when hit by the knife during sectioning of theissue (Andersen and Gundersen, 1999). Lost caps can berevented using adequate histologic techniques (Schmitzt al., 2000) and by introducing a “guard zone” at both thepper and the lower surface of the sections (Fig. 1D). Thislso prevents potential bias in neuron number estimatesroduced by uneven or wavy surfaces of the sections.nother potential source of bias is incomplete staining of
he tissue, particularly in the middle of the section thick-
i.e. cross-sectional area) of the cerebellar granule cell layer by ran-omly placing a rectangular lattice with side lengths sla-x and sla-y on
he surface of a section, and by counting the intersections of the latticend the granule cell layer (arrows). (C–E) Estimate of the number ofeurons within the cerebellar granule cell layer by randomly placing aectangular lattice with side lengths sln-x and sln-y on the surface of aection, determining the positions of unbiased counting spaces (grayquares in C; black squares in D) with base area a (so-called unbiasedounting frame; shown in E) and height h at a depth d within a sectionith thickness t. Neurons are counted if they are coming into focusithin h and if they are found either within the unbiased counting frame
neuron no. 2 in E) or hitting only the inclusion lines (dashed lines and
euron no. 3 in E) but not the exclusion lines (solid lines and neuronig. 1. Essential steps to estimate the volume of a brain region andhe number of neurons within this brain region with design-basedtereologic methods. The procedures are shown for the granule cellayer of the rat cerebellum as practical illustration. (A) Selection of aystematic random series of sections through the entire region ofnterest with a different random start for each brain (e.g. every 10th
o. 1 in E) of an unbiased counting frame.

nstus
Inptrplspganaupep
bdn(wouoabbawlstpp
woVtooceH
p(ctI
Flbn(oaoabc(arrows). Methods to prevent this are described in the text.
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 819
ess. Thus, the height of the unbiased virtual countingpaces should always be adjusted appropriately to thehickness of the sections showing adequate staining, fig-red out in a small pilot study on the sections prior totereologic analysis (for example see Jinno et al., 1998).
5.4: Number of neurons within a given brain region.n most experimental settings it is not possible to count alleurons within the BROI. Rather, one has to select aroper sample of microscopic fields to be investigated, ando derive an estimated total number of neurons within theegion of interest from the number of neurons in the sam-le and the sampling probability. The solution to this prob-
em in design-based stereology is shown (for a singleection) in Fig. 1C. Unbiased virtual counting spaces arelaced in a systematic (spacing) and random (position ofrid on section) manner within a series of systematicallynd randomly sampled sections throughout the BROI, andeurons are counted according to the criteria discussedbove at the selected microscopic fields. Note that anique identifier needs to be defined for each neuron (i.e.,rofile, nucleus, nucleolus, top of the nucleus, etc.), pref-rably a punctate one (i.e., the so-called characteristicoint; König et al., 1991; see also Fig. 4).
Estimated total numbers of neurons within a BROI cane obtained by the “VRef�NV” method (West and Gun-ersen, 1990), i.e. by calculating the product of the meaneuron density within the unbiased virtual counting spacesNV) and the global volume of the BROI (VRef; estimatedith Cavalieri’s principle; see section 5.2). Alternatively,ne can multiply the number of neurons counted within allnbiased virtual counting spaces with the reciprocal valuef the sampling probability (“Fractionator” method; West etl., 1991). The sampling probability depends on the num-er of investigated sections compared with the total num-er of sections (the “section sampling fraction”), the baserea of the unbiased virtual counting spaces comparedith the product of the side lengths of the rectangular
attice used for placing the counting frames within theections (the “area sampling fraction”), and the height ofhe unbiased virtual counting spaces (h in Fig. 1D) com-ared with the average section thickness after histologicrocessing (the “thickness sampling fraction”).
Estimating the number of neurons within a given BROIith the Fractionator method is from an economical pointf view more efficient than doing the same with the
Ref�NV method. This is due to the fact that the Fractiona-or method does not require estimates of the global volumef the BROI. Interestingly, estimated numbers of neuronsbtained with the Fractionator method are also more pre-ise (i.e. statistically more efficient) than correspondingstimates obtained with the VRef�NV method (Schmitz andof, 2000).
5.5: Mean cellular/nuclear volume. Alterations inerikaryal or nuclear volumes of neurons or glial cellsso-called local volumes) under pathologic or experimentalonditions can be regarded as, non-specific in situ indica-ors of alterations in cell metabolism (Arnold et al., 1995;
ig. 2. Procedure to estimate the volume of a brain region with Cava-ieri’s principle. (A) Schematic drawing of an oblate brain region. (B) Therain region is cut into an exhaustive series of sections of uniform thick-ess, and the top profile area of each (or every nth) section is measuredarrows). An unbiased estimate of the volume of this brain region isbtained by multiplying the sum of the profile areas of the brain region onll sections with the uniform section thickness. Note that the first sectionf this brain region, shown on top of (B), does not show a top profile areand is therefore not taken into account for the analysis. (C) A potentialias in estimates of global volumes with Cavalieri’s principle can beaused by overprojection when investigating sections at the microscope
nsausti et al., 1997; Bussière et al., 2003b). Among other

at11ohtw“Js
t(aaNHtolem
i1msFeSRam
ttbtespsab
Fantlafo wo in (C)o ts of neu
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831820
pproaches, the “Nucleator” method (Gundersen, 1988),he “Rotator” method (Vedel Jensen and Gundersen,993) and the “Optical Rotator” method (Tandrup et al.,997) have been developed to obtain unbiased estimatesf number-weighted mean local volumes (i.e. each neuronas the same probability of being selected for investiga-ion; see summary in Fig. 6). Alternatively, volume-eighted mean local volumes can be estimated with the
Point Sampled Intercepts” method (Gundersen andensen, 1985). Here the probability of a neuron beingelected for investigation depends on its individual size.
Importantly, these methods require the use of certainypes of sections, the so-called isotropic uniform randomMiles and Davy, 1976) or vertical sections (Baddeley etl., 1986). It is quite possible to prepare such sections fromgiven BROI (Baddeley et al., 1986; Mattfeldt et al., 1990;yengaard and Gundersen, 1992; Schmitz et al., 1999).owever, in isotropic uniform random or vertical sections
he plane of section is unknown, resulting in potential lossf orientation within the sections. To circumvent this prob-
em, the Optical Rotator method was developed (Tandrupt al., 1997), which works well on sections showing only a
ig. 3. Occurrence of bias (systematic error) in counting neurons by ind density of neurons (gray elements), as well as the same spatial diseurons differ in size, whereas in (C) and (D) they differ in their spatiahrough the tissue. Asterisks indicate neuron fragments which are deower surface of the section are marked by arrows. Counting all neuronnd one in (D). In contrast, counting only those neuron fractions which cractions in (A), one in (B), two in (C), and zero in (D). Furthermore, cof the section would result in three counted fractions in (A), two in (B), tf neuron numbers within a given tissue volume based solely on coun
inimum shrinkage in the z axis (e.g. sections embedded C
n methacrylate; Sousa et al., 1999; Lukoyanov et al.,999; Messina et al., 2000). Unfortunately, embedding inethacrylate is not possible in most applications in neuro-
cience since it makes immunohistochemistry impossible.rozen or vibratome sections, however, can show consid-rable shrinkage in the z axis (Messina et al., 2000;chmitz et al., 2000), preventing the use of the Opticalotator. This problem has to be individually solved in eachpplication (see Schmitz et al. 1999, for an example in theouse hippocampus and cerebellum).
5.6: Surface area. Gender-related differences in cor-ical surface area (Luders et al., 2004) or age-related al-erations in the surface area of brain capillaries (that cane interpreted as alterations in the exchange surface be-ween blood and nerve tissue; Villena et al., 2003) arexamples of parameters that can be estimated with de-ign-based stereologic methods. The methodological ap-roach is based on counting intersections between theurface of interest and a spatial grid, consisting of system-tic probes of parallel test lines (resembling nails of a fakired piercing the surface, hence termed “Fakir probes”;
sections of tissue. In all examples shown there is the same numberof these neurons when focusing on their midpoints. In (A) and (B) the
ion. The gray bar at the bottom of each example represents a sectionthe upper surface of the section, whereas fragments detected at thets would result in five counted fractions in (A), three in (B), four in (C),tected at the upper surface of the section would result in three countedly those neuron fractions which can be detected at the lower surface
, and one in (D). This illustrates the problems resulting from estimatesron fragments in the sections.
nspectingtribution
l orientattected at
fragmenan be deunting on
ruz-Orive, 1993). Fakir probes are available for the use of
Ffemcm�coftuniewwwo
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 821
ig. 4. Procedure to investigate the number of neurons within a given tissue volume with unbiased virtual counting spaces. The procedure is shownor pyramidal neurons in layer III of an adult mouse neocortex as an example. After perfusion fixation, the mouse brain was frozen and cut into anxhaustive series of 30 �m-thick frontal sections. Immunohistochemical detection of NeuN was carried out according to standard protocols using aonoclonal mouse anti-neuronal nuclei antibody (MAB377; Chemicon, Temecula, CA, USA; 1:100) and a secondary donkey anti-mouse IgG antibody
onjugated to Alexa Fluor 488 (Molecular Probes, Eugene, OR, USA; 1:100). The figure shows the upper surface of a section as well as the sameicroscopic field at 17 consecutive focal planes below the upper surface, with a distance of 1 �m between the focal planes. Between �3 �m and14 �m an unbiased counting frame is shown at each focal plane (exclusion lines in red, inclusion lines in yellow), representing an unbiased virtual
ounting space with a height of 12 �m. The midpoint of the nucleolus was used as the sampling unit (i.e. the “characteristic point”; König et al., 1991)f the neurons. At several focal planes neurons were found where the midpoint of the nucleolus came into focus (arrows). However, only the neuronsound at �9 �m, �11 �m and �14 �m were counted. The neuron found at �2 �m was positioned above the unbiased virtual counting space, andhe neuron found at �15 �m was positioned below the counting space. The midpoint of the nucleolus of the neuron found at �3 �m was outside thenbiased counting frame. Note that if one is counting nucleoli (or cell tops, etc.) and the nucleoli are outside the counting frame, the neuron shouldot be counted even if the cell body profile is partly inside the counting frame (as being the case for the neuron found at �3 �m). Vice versa, if one
s counting nucleoli (or cell tops, etc.) and the nucleoli are inside the counting frame and not touching the exclusion lines, the neuron should be countedven if the cell body profile does touch the exclusion lines (as being the case for the neurons found at �9 �m and at �14 �m). Photomicrographsere produced by digital photography using a Nikon DXM 1200F digital camera (Nikon, Tokyo, Japan) and ACT-1 software (Nikon); the final figuresere constructed using Corel Photo-Paint v. 11 and Corel Draw v. 11 (Corel, Ottawa, Canada). Only minor adjustments of contrast and brightnessere made, which in no case altered the appearance of the original materials. Scale bar�20 �m. Section contributed by B. P. F. Rutten, University
f Maastricht, Netherlands.
“ts1s1oaita
tc(sst
sslwtuT
baatLibveo
urttGtLtcs
satrsoc(uf
tbdlboi1o2mi
bNmbt1rcaom
Ftnfibb4fmnaennmfhapliccahbtl
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831822
isotropic uniform random” sections (see section 5.5, Spa-ial Grid; Sandau, 1987), the use of “vertical” sections (seeection 5.5, Vertical Spatial Grid; Cruz-Orive and Howard,995) and the use of (thick) sections with any plane ofection (Isotropic Fakir method; Kubinova and Janacek,998). As shown in Fig. 7 for the Isotropic Fakir method,rthogonal triplets of perpendicular test lines are system-tically and randomly placed within thick sections contain-
ng the surface of interest. Then, the intersections betweenhe surface and the test lines are counted, and the surfacerea is calculated from the total number of intersections.
For the sake of completeness it should be mentionedhat surface areas of local entities (such as single cells)an also be investigated with the Optical Rotator methodKiêu and Jensen, 1993; Tandrup et al., 1997; see alsoection 5.5). However, the Fakir methods can be univer-ally applied so that the surface areas of capillaries, ven-ricles and other hollow bodies can also be estimated.
5.7: Fiber length. Quantification of linear biologicaltructures (LBS) has important applications in neuro-cience. For example, it may be relevant to know theength of neurotransmitter-specific axonal innervationithin a given BROI. Earlier methods to estimate LBS on
issue sections (Smith and Guttman, 1953) required these of isotropic uniform random sections (see section 5.5).
ig. 5. Procedure to investigate the number of neurons within a givenissue volume with the disector. The procedure is shown for pyramidaleurons in layer III of an adult mouse neocortex as an example. Thegure shows the same microscopic field at two different focal planeselow the upper surface of the section, with a distance of 6 �metween the focal planes (for details on histologic procedures, see Fig.). At �4 �m the look-up frame is shown, and at �10 �m the referencerame (exclusion lines in red, inclusion lines in yellow). The neuronsarked with arrows and asterisks are seen in the reference frame butot in the look-up frame. However, only the neuron marked with anrrow is counted, because the neurons marked with asterisks hit thexclusion lines of the reference frame (note that in this case the entireeuronal profile is used as counting unit, not the nucleoli of theeurons). It should be noted that when using the disector, one has toake sure that the distance in the z-direction between the look-up
rame and the reference frame is smaller than the smallest z axiseight of the neurons under study, taking into account any kind of zxis compression or shrinkage of the tissue as a result of histologicrocessing. Otherwise, neurons could be positioned between the
ook-up frame and the reference frame and would not be registered,ntroducing bias to the neuron counts, or the profiles of two neuronsould be seen at the same position resulting in a neuron not beingounted. As a rule of thumb the distance between the “look-up frame”nd the “reference frame” should be one third of the smallest z axiseight of the neurons under study, figured out by a small pilot studyefore starting the counting procedure itself. Scale bar�20 �m. Sec-
ion contributed by B. P. F. Rutten, University of Maastricht, Nether-ands.
o circumvent this problem, the “Space Balls” method has a
een developed (Calhoun and Mouton, 2001; Mouton etl., 2002). As shown in Fig. 8, spheres (or hemispheres)re systematically and randomly placed within the sectionshrough the BROI. Then, the average length density of theBS under study is obtained from the total number of
ntersections between the spheres and the LBS. By com-ining this length density estimate with an estimate of theolume of the BROI (as explained in section 5.2), anstimate of the total length of the LBS within the BROI isbtained.
It should be mentioned that these estimates are notnbiased in a strict sense. Potential sources of bias are theatio of the diameter of the space balls and the diameter ofhe fibers investigated, and the z axis shrinkage of theissue due to histologic processing (details can be found inundersen, 2002). It is actually not possible to correct for
his shrinkage, unlike with 3D reconstructions. Thus, if theBS are not isotropic (i.e. if they have a preferential direc-ion), the length estimates will be different if sections areut in different planes (see Gundersen, 2002, for practicalolutions to minimize bias in investigations of LBS).
Another possibility to quantify LBS with design-basedtereology is the use of “isotropic virtual planes” (Larsen etl., 1998). In this case, the LBS under study contained inhick (3D) sections are investigated with software-andomized isotropic virtual planes in volume probes inystematically sampled microscope fields. A disadvantagef this technique may be the fact that the analysis has to bearried out on virtual planes within the thick sectionswhich can be tedious and cumbersome), whereas bysing the space balls the plane of analysis is always theocal plane of the microscopic fields.
5.8: Three-dimensional cytoarchitecture. Discrete al-erations in the normal cytoarchitecture of the adult cere-ral cortex attributable to disturbed neuronal migrationuring development have usually been investigated by
abeling precursors of neurons in the brain in utero withromodeoxyuridine (BrdU). Then, the distribution patternsf BrdU-labeled cells have been analyzed postnatally by
mmunohistochemistry (for example see Fushiki et al.,997). However, this approach suffers from several meth-dological limitations (see discussion in Schmitz et al.,002a). Most importantly, it cannot be applied to the hu-an brain, as it is impossible to label cells in the develop-
ng human cerebral cortex with BrdU.To circumvent these limitations, a novel 3D design-
ased stereologic method was recently developed, namedearest Neighbor analysis (Schmitz et al., 2002a). Thisethod, which essential steps are shown in Fig. 9, isased on the “nearest-neighbor distance distribution func-ion” analysis founded in theoretical statistics (Diggle,983). It makes it possible to determine whether the neu-ons within a given BROI exhibit spatial randomness, alustered distribution, or a more dispersed distribution. Thepplication of Nearest Neighbor analysis requires the usef thick sections encompassing the entire BROI (approxi-ately 100–150 �m thick sections for the mouse brain,
nd up to 750 �m thick sections for the human brain; any
palo(tt
n(HsoUah
Sd
Tfmwpv
itewaedoa“
musttttt(drltttlnnppTbmo
Fawo(Ttmtnnnuclear boundary. (B) Estimates of nuclear volumes with the Rotator
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 823
lane of section), as well as a staining method that guar-ntees staining throughout the section thickness (e.g. gal-
ocyanin). Nearest Neighbor analysis comprises estimatesf both the volume of the BROI using Cavalieri’s principlesee section 5.2) and of the total number of neurons withinhis BROI using the optical fractionator method (see sec-ion 5.4).
Another approach to study the spatial distribution ofeurons within a BROI is the so-called Voronoi tessellationDuyckaerts et al., 1994; Duyckaerts and Godefroy, 2000;of et al., 2003). This approach focuses on the region ofpace that any cell within a BROI occupies, i.e. the regionf space that is closer to that cell than to any other.nfortunately, it has not been possible so far to adaptlgorithms that have been published for 3D tessellations toistologic specimens.
ection 6: Variability of estimates obtained withesign-based stereology
he results of quantitative histologic investigations per-ormed with design-based stereologic methods are esti-ates rather than exact measurements. Thus, the resultsill vary if the same stereologic estimate were to be inde-endently repeated. This implies that information about theariability of stereologic estimates is an important topic.
If the same stereologic estimate would be repeated adnfinitum, one could calculate the coefficient of variation ofhese estimates (note that the mean of such ad infinitumstimates must equal the true value under study; other-ise, the estimates would not be unbiased). In practicalpplications of design-based stereology, however, this co-fficient of variation is unknown. It is one of the mostesirable advantages of design-based stereology overther quantitative histologic techniques that predictionsbout this coefficient of variation (usually presented ascoefficient of error,” CE) are possible.
ethod. Again, the midpoint of the nucleolus is selected as samplingnit, and the analysis is carried out as follows. First, an axis along thehortest aspect of the sampled nucleus is generated on the sectionhrough the midpoint of the nucleolus(‘i’ in Fig. 6B). Then the top andhe bottom position of the nuclear profile are marked with respect tohe defined axis (’ii’; light dashed lines in Fig. 6B) in order to measurehe height of the nuclear profile (‘h’; thick solid line in Fig. 6B). Then,hree parallel test lines are generated perpendicular to the defined axis’iii’; light solid lines in Fig. 6B). The position of the test lines on theefined axis is uniformly random, i.e. the position of the first one isandomly chosen in an interval of h/3, with the distance between theines being h/3. The lengths of the intercepts of the test lines betweenhe defined axis and the nuclear boundary (li; thick dashed line) arehen measured. From these measurements an unbiased estimate ofhe nuclear volume can be obtained (Jensen and Gundersen, 1993). li,ength of the intercept of a test line between a defined axis and theuclear boundary; m, midpoint of the nucleolus. (C) Estimates ofuclear volumes with the Point Sampled Intercepts method. All nuclearrofiles are considered for analysis which are found in a given focallane. A test system of parallel lines is randomly placed on the section.hen, the length of the intercept of the test line between the nuclearoundaries (l0) is measured for each sampled nucleus. From these
easurements an unbiased estimate of the nuclear volume can beig. 6. Procedure to estimate the mean nuclear volume of neurons. Inll examples schematic drawings of two neuronal profiles are shownhich were selected by an unbiased counting frame. For the neuronn the right the nucleolus is found within the selected focal planepoint). (A) Estimates of nuclear volumes with the Nucleator method.he midpoint of the nucleolus is selected as sampling unit. The dis-
ance from the midpoint of the nucleolus to the nuclear boundary (l) iseasured in four mutually orthogonal directions. From the average of
he third powers of these measurements an unbiased estimate of theuclear volume can be obtained (Gundersen, 1988). m, midpoint of theucleolus; lm, distance between the midpoint of the nucleolus and the
btained (Gundersen and Jensen, 1985).

g(tTsbt1trcF4
po1taMtwGipscoosa
easswssdh(gsefpsrmmmdttatn
F(wtoaascifbeJinto sections (as shown in A).
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831824
For estimates of the total number of neurons within aiven brain region obtained with the Fractionator methodthereafter Fractionator estimates), 16 different procedureso predict the CE have been described (summarized inable 1 in Schmitz and Hof, 2000). Extensive computerimulations have shown that several of these methods cane applied to predict precisely the CE of Fractionator es-imates (for details see Schmitz, 1998; Glaser and Wilson,998, 1999; Schmitz and Hof, 2000). As a rule of thumb,
he CE of a Fractionator estimate approximately equals theeciprocal value of the square root of the number ofounted neurons (Schmitz, 1998; Schmitz and Hof, 2000).or example, if a Fractionator estimate results in counting00 neurons, the CE is predicted as 1⁄�400�0.05.
Three different procedures have been proposed toredict the CE of estimated total numbers of neuronsbtained with the VRef�NV method (summarized in Tablein Schmitz and Hof, 2000). However, computer simula-
ions have shown that these procedures do not result indequate predictions of CE (Schmitz and Hof, 2000).oreover, four different procedures have been described
o predict the CE of estimates of global volumes obtainedith Cavalieri’s principle (thereafter Cavalieri estimates;undersen and Jensen, 1987; Roberts et al., 1994; Gein-
sman et al., 1996; Garcia-Fiñana et al., 2003). Pilot com-uter simulations have shown that the procedure de-cribed in Garcia-Fiñana et al. (2003) is likely the besthoice for predicting the CE of Cavalieri estimates basedn point counting (C. Schmitz and P. R. Hof, unpublishedbservations). For the remaining methods described inection 5, proven procedures to predict precisely the CEre currently not available.
An unsolved problem in design-based stereology is thestablishment of criteria for sampling requirements tochieve a specific variability in the estimates. In this re-pect it has been a long-standing rule in design-basedtereology that sufficient precision of estimates is obtainedhen in analyses of a sample of cases, the mean of thequared CEs of the individual estimates is smaller than thequared coefficient of variation of the estimates (Gun-ersen and Østerby, 1981). Recent computer simulations,owever, have shown the limitation of this approachSchmitz and Hof, 2000). In fact, there is currently noeneral answer to the question how to design stereologicampling protocols so that both sufficient precision of thestimates is achieved while the work is efficient by per-orming as few measurements as possible to achieve thisrecision. This is essentially due to the fact that differentcientific questions to be addressed by design-based ste-eology require different levels of precision in the esti-ates. For example, a correlation analysis usually requiresore precise estimates than the comparison of two groupeans with a Student’s t-test. It is always advisable toesign an individual sampling scheme for each new study,aking into account all potential factors that will likely affecthe precision of estimates such as potential biologic vari-bility, the number of animals or brains to be investigated,he number of sections to be analyzed, the number of
ig. 7. Procedure to estimate the surface area of biological objects.A) A schematic drawing of an object (e.g. a brain region) is shownhich is hit by perpendicular test lines. For the sake of clarity, only two
est lines each in directions X, Y and Z are shown. For a real estimatef the surface area of this object, a systematically and randomly placedrray of test lines in directions X, Y and Z each would have to be used,s shown in (B) for direction Z. (C) Schematic drawing of two thickections from the object shown in (A). In this case each sectionontains only a part of the surface of the object, and only a fewntersections of the test lines and the surface area of the object areound in a given section. However, the total number of intersectionsetween the test lines and the surface area of the object provides anstimate of the surface area (as explained in detail in Kubinova andanacek, 1998), as is the case when the test lines hit the object not cut
eurons to be counted, etc.

Sd
Iri
pt
s
Fhs(stsaaswti ibuted by
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 825
ection 7: Presentation of results obtained withesign-based stereology
nvestigators and readers, as well as editorial boards andeviewers have all been concerned by the issue of what
ig. 8. Procedure to estimate the total length of a LBS within a givenilus of an adult mouse hippocampus. After perfusion fixation, the brections. For myelin staining a silver impregnation modified from ZilleDG, dentate gyrus; CA3, region CA3). The square indicates the regionurface of the section as well as the same microscopic field at 13 conshe focal planes. Between �1 �m and �11 �m the intersections of ahown as circles. This space ball was centered at a depth of �6 �m and the myelinated fibers in focus at the point of intersection were found the size of the sphere, the length density of myelinated fibers at thepace balls in a systematic–random manner throughout the entire hippoithin this region. Finally, by combining length density with an estimat
otal length of myelinated fibers within the mouse hippocampal hilus isn (A), and 10 �m in the high-power photomicrographs. Section contr
nformation and discussion are essential in a stereologic i
aper. Whereas there is no general answer to this ques-ion, the following recommendations might be useful.
The materials and methods of stereologic studieshould provide a description of the histologic process-
his example illustrates the total length of myelinated fibers within thembedded in paraffin and cut into a series of 13 �m-thick horizontal
was applied. Panel (A) shows an overview on the hippocampal hilusthe high-power photomicrographs were taken. These show the uppercal planes below the upper surface, with a distance of 1 �m between“space ball”) and the focal plane at the investigated focal depth are
radius of 5 �m. Except for �6 �m, intersections between the sphereshown focal planes (arrows). From the total number of intersectionsted microscopic field can be obtained (Mouton et al., 2002). By placingilus, one can estimate the average length density of myelinated fibers
olume of the region (as shown in Fig. 2), an unbiased estimate of thed. Photomicrographs were produced as in Fig. 4. Scale bar�200 �mS. J. P. Haas, University of Rostock, Germany.
tissue. Tain was es (1985)at which
ecutive fosphere (and had and at allinvestigacampal h
e of the vobtaine
ng, with consideration of the specific requirements

ts
l
Fl(go(p(a(tnarNtnrbficlfOc
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831826
o be met (outlined in section 2) and the potential
ig. 9. Procedure to analyze the nearest-neighbor distance distributioateral nucleus of the right amygdala in a brain from a 55-year-oldMicroBrightField). The brain was embedded in gelatin and was cut inallocyanin. To prevent any damage to the cortical surface, the gelatinf the lateral nucleus (lat), the basal nucleus (bas) and the accessory baarrow) at a depth of 60 �m below the upper surface of the section wrocedure; the asterisk shows the Focus Position Meter display indicaPN; yellow dot) by mouse-clicking on its nucleolus. Note that the termsprocess related to neurogenesis; these terms were taken from the de
D) Identification of another neuron (arrow) in the same microscopic fiehe focal plane of the PN), which may be the nearest-neighbor to theeuron 1” (ON1; white dot) by mouse-clicking on its nucleolus. S1, circlnd a sphere centered on the nucleolus of the PN, with the 3D distanadius. (F) Identification of another three neurons (X, Y and Z) in the samote that neurons Y and Z were lying inside the circle representing the
he nucleolus of these neurons and the PN was smaller than the distanearest-neighbor to the PN. (G) Categorizing neuron Z as “offsprinepresenting the intersection of the focal plane at the investigated focaetween the nucleolus of the PN and the nucleolus of ON2 as the spheld (i.e. to the focal plane at which ON1 was found). Note that ON1 waonfirming that ON1 was not the nearest-neighbor of the PN since thearger than the 3D distance between the nucleolus of the PN and the nound inside the sphere centered on the nucleolus of the PN with the 3DN as the sphere’s radius. The radius of this sphere was the nearontributed by Dr. H. Heinsen, University of Würzburg, Germany.
ources of bias (summarized in section 4). The stereo- t
ogic probes which are used should briefly be men-n for neurons in a given brain region. The procedure is shown for therenic patient as an example, using the StereoInvestigator softwareplete series of 600 �m-thick frontal sections, which were stained withremoved during histologic processing (asterisk in A). (A) Delineationus (ab) of the amygdala. Scale bar�3.5 mm. (B) Sampling of a neuronbiased virtual counting space (see Fig. 4 for details of the samplingsoftware). (C) Categorizing the sampled neuron as a “parent neuron”neuron” and “offspring neurons” (below) are not intended to representof nearest-neighbor analysis in the statistical literature (Diggle, 1983).pth of 53 �m below the upper surface of the section (i.e. 7 �m above
d parent neuron. (E) Categorizing the identified neuron as “offspringnting the intersection of the focal plane at the investigated focal depth
en the nucleolus of the PN and the nucleolus of ON1 as the sphere’sscopic field at a depth of 45 �m below the upper surface of the section.tersection of S1 at this focal depth. Accordingly, the distance betweenen the nucleolus of ON1 and the PN, indicating that ON1 was not the2” (ON2; white dot) by mouse-clicking on its nucleolus. S2, circled a sphere centered on the nucleolus of the PN, with the 3D distanceius. (H) Focusing back to a depth of 53 �m in the same microscopictside the circle representing the plane section of S2 at this focal depth,nce between the nucleolus of the PN and the nucleolus of ON1 wasof ON2. The steps D–H were repeated until no new neuron could bebetween the nucleolus of the PN and the nucleolus of last categorized
bor distance of the selected PN. Scale bar�30 �m (B–H). Section
n functioschizophto a com
was notsal nucleith an un
tor of the“parent
scriptionld at a deselecte
e represece betwe
e microplane ince betwe
g neuronl depth anere’s rad
s lying ou3D distaucleolusdistance
est-neigh
ioned, together with a reference to the corresponding
lmpstsp
welts(ottsldctetgeteAsp(
Sn
Octreadbsfb
d(mlwmscba
isiofoo
T
V
MMMSMMMM
C
a
iothfisRm
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 827
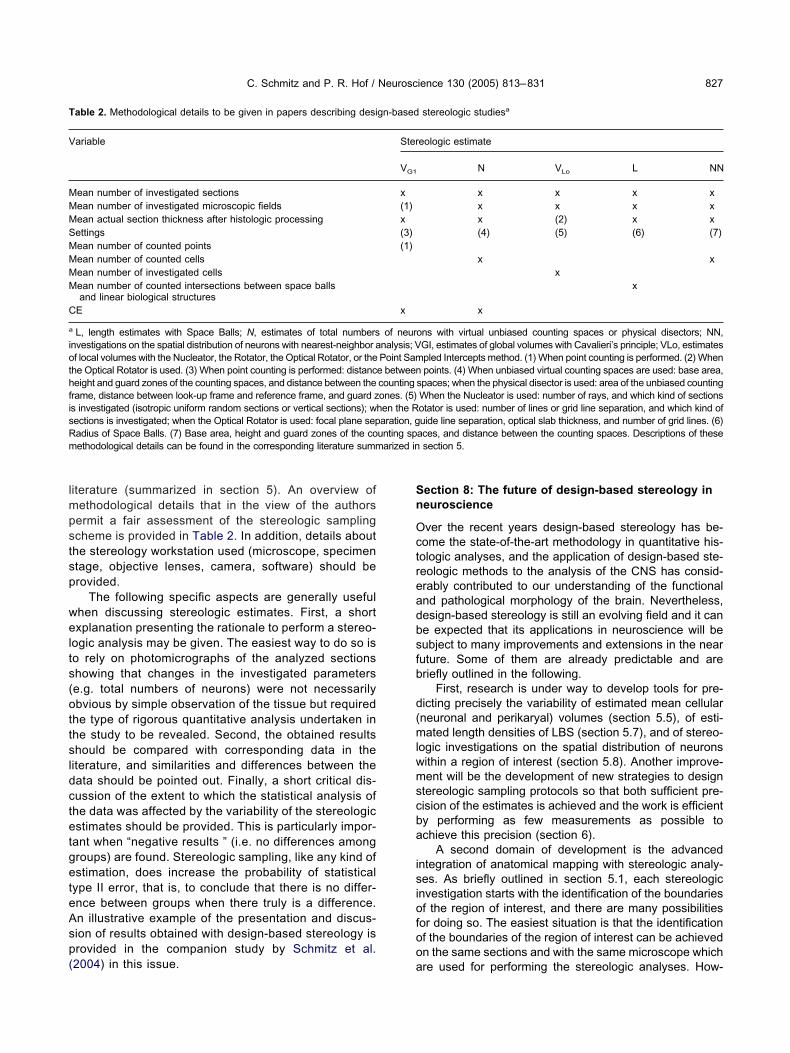
iterature (summarized in section 5). An overview ofethodological details that in the view of the authorsermit a fair assessment of the stereologic samplingcheme is provided in Table 2. In addition, details abouthe stereology workstation used (microscope, specimentage, objective lenses, camera, software) should berovided.
The following specific aspects are generally usefulhen discussing stereologic estimates. First, a shortxplanation presenting the rationale to perform a stereo-
ogic analysis may be given. The easiest way to do so iso rely on photomicrographs of the analyzed sectionshowing that changes in the investigated parameterse.g. total numbers of neurons) were not necessarilybvious by simple observation of the tissue but required
he type of rigorous quantitative analysis undertaken inhe study to be revealed. Second, the obtained resultshould be compared with corresponding data in the
iterature, and similarities and differences between theata should be pointed out. Finally, a short critical dis-ussion of the extent to which the statistical analysis ofhe data was affected by the variability of the stereologicstimates should be provided. This is particularly impor-
ant when “negative results ” (i.e. no differences amongroups) are found. Stereologic sampling, like any kind ofstimation, does increase the probability of statistical
ype II error, that is, to conclude that there is no differ-nce between groups when there truly is a difference.n illustrative example of the presentation and discus-ion of results obtained with design-based stereology isrovided in the companion study by Schmitz et al.
able 2. Methodological details to be given in papers describing desi
ariable
ean number of investigated sectionsean number of investigated microscopic fieldsean actual section thickness after histologic processingettingsean number of counted pointsean number of counted cellsean number of investigated cellsean number of counted intersections between space ballsand linear biological structures
E
L, length estimates with Space Balls; N, estimates of total numbersnvestigations on the spatial distribution of neurons with nearest-neighbor af local volumes with the Nucleator, the Rotator, the Optical Rotator, or thehe Optical Rotator is used. (3) When point counting is performed: distanceight and guard zones of the counting spaces, and distance between the
rame, distance between look-up frame and reference frame, and guard zs investigated (isotropic uniform random sections or vertical sections); wections is investigated; when the Optical Rotator is used: focal plane sepadius of Space Balls. (7) Base area, height and guard zones of the coethodological details can be found in the corresponding literature summ
2004) in this issue. a
ection 8: The future of design-based stereology ineuroscience
ver the recent years design-based stereology has be-ome the state-of-the-art methodology in quantitative his-ologic analyses, and the application of design-based ste-eologic methods to the analysis of the CNS has consid-rably contributed to our understanding of the functionalnd pathological morphology of the brain. Nevertheless,esign-based stereology is still an evolving field and it cane expected that its applications in neuroscience will beubject to many improvements and extensions in the nearuture. Some of them are already predictable and areriefly outlined in the following.
First, research is under way to develop tools for pre-icting precisely the variability of estimated mean cellularneuronal and perikaryal) volumes (section 5.5), of esti-ated length densities of LBS (section 5.7), and of stereo-
ogic investigations on the spatial distribution of neuronsithin a region of interest (section 5.8). Another improve-ent will be the development of new strategies to design
tereologic sampling protocols so that both sufficient pre-ision of the estimates is achieved and the work is efficienty performing as few measurements as possible tochieve this precision (section 6).
A second domain of development is the advancedntegration of anatomical mapping with stereologic analy-es. As briefly outlined in section 5.1, each stereologic
nvestigation starts with the identification of the boundariesf the region of interest, and there are many possibilitiesor doing so. The easiest situation is that the identificationf the boundaries of the region of interest can be achievedn the same sections and with the same microscope which
stereologic studiesa
eologic estimate
N VLo L NN
x x x xx x x xx (2) x x(4) (5) (6) (7)
x xx
x
x
ons with virtual unbiased counting spaces or physical disectors; NN,GI, estimates of global volumes with Cavalieri’s principle; VLo, estimatespled Intercepts method. (1) When point counting is performed. (2) Whenpoints. (4) When unbiased virtual counting spaces are used: base area,
spaces; when the physical disector is used: area of the unbiased countingWhen the Nucleator is used: number of rays, and which kind of sectionsotator is used: number of lines or grid line separation, and which kind ofuide line separation, optical slab thickness, and number of grid lines. (6)
aces, and distance between the counting spaces. Descriptions of thesesection 5.
gn-based
Ster
VG1
x(1)x(3)(1)
x
of neurnalysis; VPoint Same betweencountingones. (5)hen the Raration, gunting sp
arized in
re used for performing the stereologic analyses. How-

eiaenaamcsasibdssts
fcrapotsstJcebmdoo
C
Mbqitfbaaldntqquadp
ptsse
ApSosOAi
A
A
A
B
B
B
B
C
C
C
C
C
C
C
C
C
C
D
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831828
ver, in many situations this is not the case. For example,f boundaries of the region of interest are established with
chemoarchitectural approach (i.e. making use of differ-nces in the regional distribution of certain subsets ofeurons) it can become necessary to identify the bound-ries of the region of interest and to perform the stereologicnalyses on adjacent, differently stained sections. Further-ore, identification of the boundaries of cortical and sub-
ortical areas in the human brain can require the use ofpecial equipment to apply the GLI method (Schleicher etl., 2000; see section 5.1 for details) or a stereomicro-cope (Heinsen et al., 1999). Once appropriate softwarenterfaces will have been developed, the boundaries of therain regions of interest identified with the GLI method canirectly be used by the commercially available stereologyystems (see section 3). Another advance will be to fittereomicroscopes with transparent graphic tablets andransfer the delineations directly to the software of thetereology systems.
Third, quantitative neuromorphology will greatly benefitrom further integration of design-based stereology withonfocal microscopy and electron microscopy. The opticalesolution of confocal microscopy and electron microscopys well as the ability to collect registered series of focallanes is ideally suited for the three-dimensional samplingf design-based stereology (see section 5). First attempts
o integrate the collection of confocal and electron micro-copic images with the implementation of design-basedtereology has been undertaken (Mayhew, 1996; Pe-ersen, 1999; Glaser and Glaser, 2000; Kubinova andanacek, 2001; Howell et al., 2002). As for light micros-opy, it is to be expected that this field will considerablyxpand with the recognition that the integration of design-ased stereology with confocal microscopy and electronicroscopy is fundamental in revealing certain features ofevelopment, repair, aging, pathology and normal anat-my of the brain that would not have been detectedtherwise.
oncluding remarks
any important questions in neuroscience cannot simplye answered with a simple qualitative analysis. Wheneveruantitative morphologic approaches are used, the follow-
ng issues should be considered: are the results represen-ative for the entire BROI? Are the results independentrom the size, shape, spatial orientation, and spatial distri-ution of the cells to be investigated? Is bias (i.e. system-tic error) in the results kept to a minimum? Is it possible tossess the variability of the results? In numerous pub-
ished studies reporting quantitative data not obtained withesign-based stereologic methods, these questions can-ot be adequately answered. In some cases, such quan-itative data can even be more misleading than purelyualitative statements, because they are based on inade-uate methods and are prone to overinterpretation. These of design-based stereologic methods permits one tonswer all these questions positively. Furthermore,esign-based stereology comprises rigorous emphasis on
rospective experimental design and the requirement forroper sampling, presumably the most important contribu-ions of this methodology to neuroscience. Hopefully theame principles will be increasingly applied to other fieldsuch as 3D neuronal reconstruction, quantitative (immuno)lectron microscopy, and neurophysiology.
cknowledgments—We thank our many colleagues whorompted us to write this review. We would like to thank thetanley Medical Research Institute and the US-National Alliancef Autism Research (to C.S. and P.R.H.), the Alzheimer For-chung Initiative e.V., the Internationale Stichting Alzheimernderzoek and the Hersenstichting (to C.S.), and NIH grantsG02219, AG05138, MH58911, and MH66392 (to P.R.H.). P.R.H.
s the Regenstreif Professor of Neuroscience.
REFERENCES
bercrombie M (1946) Estimation of nuclear population from mic-rotome sections. Anat Rec 94:239–247.
ndersen BB, Gundersen HJG (1999) Pronounced loss of cell nuclei andanisotropic deformation of thick sections. J Microsc 196:69–73.
rnold SE, Franz BR, Gur RC, Gur RE, Shapiro RM, Moberg PJ,Trojanowski JQ (1995) Smaller neuron size in schizophrenia inhippocampal subfields that mediate cortical-hippocampal interac-tions. Am J Psychiatry 152:738–748.
addeley AJ, Gundersen HJG, Cruz-Orive LM (1986) Estimation ofsurface area from vertical sections. J Microsc 142:259–276.
auchot R (1967) Les modifications du poids encephalique au coursde la fixation [Modifications of brain weight in the course of fixa-tion]. J Hirnforsch 9:253–283.
ussière T, Giannakopoulos P, Bouras C, Perl DP, Morrison JH, HofPR (2003a) Progressive degeneration of nonphosphorylated neu-rofilament protein-enriched pyramidal neurons predicts cognitiveimpairment in Alzheimer’s disease: stereologic analysis of prefron-tal cortex area 9. J Comp Neurol 463:281–302.
ussière T, Gold G, Kovari E, Giannakopoulos P, Bouras C, Perl DP,Morrison JH, Hof PR (2003b) Stereologic analysis of neurofibrillarytangle formation in prefrontal cortex area 9 in aging and Alzhei-mer’s disease. Neuroscience 117:577–592.
alhoun ME, Mouton PR (2001) Length measurement: new develop-ments in neurostereology and 3D imagery. J Chem Neuroanat21:61–69.
armichael ST, Price JL (1994) Architectonic subdivision of the orbitaland medial prefrontal cortex in the macaque monkey. J CompNeurol 346:366–402.
avalieri B (1635) Geometria indivisibilibus continuorum. Bononiae:Typis Clementis Ferronij (reprinted 1966 as Geometria Degli Indi-visibili. Torino: Unione Tipografico-Editrice Torinese).
oggeshall RE (1999) Assaying structural changes after nerve dam-age, an essay on quantitative morphology. Pain Suppl 6:S21–S25.
oggeshall RE, Lekan HA (1996) Methods for determining numbers ofcells and synapses: a case for more uniform standards of review.J Comp Neurol 364:6–15.
oleman PD (1997) Research uses of neuropathological data. Neu-robiol Aging 18:S97–S98.
ooper JD, Payne JN, Horobin RW (1988) Accurate counting ofneurons in frozen sections: some necessary precautions. J Anat157:13–21.
ruz-Orive, LM (1993) Systematic sampling in stereology. Bull InternStatist Inst Proc 49th Session, Florence 55:451–468.
ruz-Orive LM, Weibel ER (1990) Recent stereological methods forcell biology: a brief survey. Am J Physiol 258:L148–L156.
ruz-Orive LM, Howard CV (1995) Estimation of individual featuresurface area with the vertical spatial grid. J Microsc 178:146–151.
iggle PJ (1983) Statistical analysis of spatial point patterns. New
York: Academic Press.
D
D
D
EF
G
G
G
G
G
G
G
G
G
G
GG
G
G
G
G
G
H
H
H
H
H
H
H
H
H
H
H
I
J
K
K
K
K
K
K
L
L
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 829
orph-Petersen KA, Nyengaard JR, Gundersen HJG (2001) Tissueshrinkage and unbiased stereological estimation of particle num-ber and size. J Microsc 204:232–246.
uyckaerts C, Godefroy G (2000) Voronoi tessellation to study thenumerical density and the spatial distribution of neurones. J ChemNeuroanat 20:83–92.
uyckaerts C, Godefroy G, Hauw JJ (1994) Evaluation of neuronalnumerical density by Dirichlet tessellation. J Neurosci Methods51:47–69.
lias H, Hyde MD (1983) Guide to practical stereology. Basel: Karger.ushiki S, Hyodo-Taguchi Y, Kinoshita C, Ishikawa Y, Hirobe T (1997)
Short- and long-term effects of low-dose prenatal X-irradiation inmouse cerebral cortex, with special reference to neuronal migra-tion. Acta Neuropathol 93:443–449.
ardella D, Hatton WJ, Rind HB, Rosen GD, von Bartheld CS (2003)Differential tissue shrinkage and compression in the z-axis: impli-cations for optical disector counting in vibratome-, plastic- andcryosections. J Neurosci Methods 124:45–59.
arcia-Fiñana M, Cruz-Orive LM, Mackay CE, Pakkenberg B, RobertsN (2003) Comparison of MR imaging against physical sectioning toestimate the volume of human cerebral compartments. Neuroim-age 18:505–516.
einisman Y, Gundersen HJG, van der Zee E, West MJ (1996)Unbiased stereological estimation of the total number of synapsesin a brain region. J Neurocytol 25:805–819.
euna S (2000) Appreciating the difference between design-basedand model-based sampling strategies in quantitative morphologyof the nervous system. J Comp Neurol 427:333–339.
laser EM, Wilson PD (1998) The coefficient of error of optical frac-tionator population size estimates: a computer simulation compar-ing three estimators. J Microsc 192:163–171.
laser EM, Wilson PD (1999) A comparison of three estimators of thecoefficient of error of optical fractionator cell count estimates. ActaStereol 18:15–25.
laser J, Glaser EM (2000) Stereology, morphometry, and mapping:the whole is greater than the sum of its parts. J Chem Neuroanat20:115–126.
uillery RW, Herrup K (1997) Quantification without pontification:choosing a method for counting objects in sectioned tissues.J Comp Neurol 386:2–7.
undersen HJG (1977) Notes on the estimation of the numericaldensity of arbitrary particles: the edge effect. J Microsc111:�219–233.
undersen HJG (1986) Stereology of arbitrary particles. J Microsc143:3–45.
undersen HJG (1988) The nucleator. J Microsc 151:3–21.undersen HJG (1992) Stereology: the fast lane between neuroanat-
omy and brain function, or still only a tightrope? Acta Neurol ScandSuppl 137:8–13.
undersen HJG (2002) Stereological estimation of tubular length. JMicrosc 207:155–160.
undersen HJG, Østerby R (1981) Optimizing sampling efficiency ofstereological studies in biology: or “Do more less well!” J Microsc121:65–73.
undersen HJG, Jensen EB (1985) Stereological estimation of thevolume-weighted mean volume of arbitrary particles observed onrandom sections. J Microsc 138:127–142.
undersen HJG, Jensen EB (1987) The efficiency of systematic sam-pling and its prediction. J Microsc 147:229–263.
undersen HJG, Bagger A, Bendtsen TF, Evans SM, Korbo L, Mar-cussen N, Møller A, Nielsen K, Nyengaard JR, Pakkenberg B,Sørensen FB, Vesterby A, West MJ (1988) The new stereologicaltools: disector, fractionator, nucleator and point sampled interceptsand their use in pathological research and diagnosis. Acta PatholMicrobiol Immunol Scand 96:857–881.
arrison PJ (1999) The neuropathology of schizophrenia: a critical
review of the data and their interpretation. Brain 122:593–624.atton WJ, von Bartheld CS (1999) Analysis of cell death in thetrochlear nucleus of the chick embryo: calibration of the opticaldisector counting method reveals systematic bias. J Comp Neurol409:169–186.
aug H (1986) History of neuromorphometry. J Neurosci Methods18:1–17.
edreen JC (1998) What was wrong with the Abercrombie and em-pirical cell counting methods? A review. Anat Rec 250:373–380.
einsen H, Rüb U, Bauer M, Ulmar G, Bethke B, Schuler M, Böcker F,Eisenmenger W, Götz M, Korr H, Schmitz C (1999) Nerve cell lossin the thalamic mediodorsal nucleus in Huntington’s disease. ActaNeuropathol 97:613–622.
einsen H, Arzberger T, Schmitz C (2000) Celloidin mounting (em-bedding without infiltration): a new, simple and reliable method forproducing serial sections of high thickness through complete hu-man brains and its application to stereological and immunohisto-chemical investigations. J Chem Neuroanat 20:49–59.
of PR, Morrison JH (1995) Neurofilament protein defines regionalpatterns of cortical organization in the macaque monkey visualsystem: a quantitative immunohistochemical analysis. J CompNeurol 352:161–186.
of PR, Haroutunian V, Friedrich VL Jr, Byne W, Buitron C, Perl DP,Davis KL (2003) Loss and altered spatial distribution of oligoden-drocytes in the superior frontal gyrus in schizophrenia. Biol Psy-chiatry 53:1075–1085.
oward CV, Reed MG (1998) Unbiased stereology: three-dimensionalmeasurement in microscopy. Berlin: Springer.
owell K, Hopkins N, Mcloughlin P (2002) Combined confocal micros-copy and stereology: a highly efficient and unbiased approach toquantitative structural measurement in tissues. Exp Physiol87:747–756.
yman BT, Gomez-Isla T, Irizarry MC (1998) Stereology: a practicalprimer for neuropathology. J Neuropathol Exp Neurol 57:305–310.
nsausti AM, Gaztelu JM, Gonzalo LM, Romero-Vives M, BarrenecheaC, Felipo V, Insausti R (1997) Diet induced hyperammonemiadecreases neuronal nuclear size in rat entorhinal cortex. NeurosciLett 231:179–181.
inno S, Aika Y, Fukuda T, Kosaka T (1998) Quantitative analysis ofGABAergic neurons in the mouse hippocampus, with optical disectorusing confocal laser scanning microscopy. Brain Res 814:55–70.
iêu K, Jensen EBV (1993) Stereological estimation based on isotro-pic slices through fixed points. J Microsc 170:45–51.
önig D, Carvajal-Gonzales S, Downs AM, Vassy J, Rigaut JP (1991)Modelling and analysis of 3-D arrangements of particles by pointprocesses with examples of application to biological data obtainedby confocal scanning light microscopy. J Microsc 161:405–433.
retschmann HJ, Tafesse U, Herrmann A (1982) Different volumechanges of cerebral cortex and white matter during histologicalpreparation. Microsc Acta 86:13–24.
ubinova L, Janacek J (1998) Estimating surface area by the isotropicfakir method from thick slices cut in an arbitrary direction. J Microsc191:201–211.
ubinova L, Janacek J (2001) Confocal microscopy and stereology:estimating volume, number, surface area and length by virtual testprobes applied to three-dimensional images. Microsc Res Tech53:425–435.
ubinova L, Janacek J, Karen P, Radochova B, Difato F, Krekule I(2004) Confocal stereology and image analysis: methods for esti-mating geometrical characteristics of cells and tissues from three-dimensional confocal images. Physiol Res 53(Suppl 1):S47–55.
arsen JO, Gundersen HJG, Nielsen J (1998) Global spatial samplingwith isotropic virtual planes: estimators of length density and totallength in thick, arbitrarily oriented sections. J Microsc191:238–248.
uders E, Narr KL, Thompson PM, Rex DE, Jancke L, Steinmetz H,Toga AW (2004) Gender differences in cortical complexity. Nat
Neurosci, online publication (doi:10.1038/nn1277).
L
L
M
M
M
M
M
M
M
M
N
N
O
O
P
P
P
Q
R
R
R
R
R
R
R
S
S
S
S
S
S
S
S
S
S
S
S
S
S
T
U
V
V
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831830
ukoyanov NV, Madeira MD, Paula-Barbosa MM (1999) Behavioraland neuroanatomical consequences of chronic ethanol intake andwithdrawal. Physiol Behav 66:337–346.
ukoyanov NV, Brandao F, Cadete-Leite A, Madeira MD, Paula-Bar-bosa MM (2000) Synaptic reorganization in the hippocampal for-mation of alcohol-fed rats may compensate for functional deficitsrelated to neuronal loss. Alcohol 20:139–148.
attfeldt T, Mall G, Gharehbaghi H (1990) Estimation of surface areaand length with the orientator. J Microsc 159:301–317.
ayhew TM (1992) A review of recent advances in stereology forquantifying neural structure. J Neurocytol 21:313–328.
ayhew TM (1996) How to count synapses unbiasedly and efficientlyat the ultrastructural level: proposal for a standard sampling andcounting protocol. J Neurocytol 25:793–804.
ayhew TM, Gundersen HJG (1996) If you assume, you can make anass out of u and me: a decade of the disector for stereologicalcounting of particles in 3D space. J Anat 188:1–15.
essina A, Sangster CL, Morrison WA, Galea MP (2000) Require-ments for obtaining unbiased estimates of neuronal numbers infrozen sections. J Neurosci Methods 97:133–137.
iles RE, Davy PJ (1976) Precise and general conditions for thevalidity of a comprehensive set of stereological fundamental for-mulae. J Microsc 107:211–226.
outon PR (2002) Principles and practices of unbiased stereology: anintroduction for bioscientists. Baltimore: Johns Hopkins UniversityPress.
outon PR, Gokhale AM, Ward NL, West MJ (2002) Stereologicallength estimation using spherical probes. J Microsc 206:54–64.
urcombe V, Wreford N, Bertram JF (1999) Stereological methods forbiology. Boston: Chapman & Hall.
yengaard JR, Gundersen HJG (1992) The disector: a simple anddirect method for generating isotropic, uniform random sectionsfrom small specimens. J Microsc 165:427–431.
ngür D, Ferry AT, Price JL (2003) Architectonic subdivision of thehuman orbital and medial prefrontal cortex. J Comp Neurol460:425–449.
orschot DE, Peterson DA, Jones DG (1991) Neurite growth from,and neuronal survival within, cultured explants of the nervoussystem: a critical review of morphometric and stereological meth-ods, and suggestions for the future. Prog Neurobiol 37:525–546.
akkenberg B, Boesen J, Albeck M, Gjerris F (1989) Unbiased andefficient estimation of total ventricular volume of the brain obtainedfrom CT-scans by a stereological method. Neuroradiology31:413–417.
erl DP, Good PF, Bussière T, Morrison JH, Erwin JM, Hof PR (2000)Practical approaches to stereology in the setting of aging- anddisease-related brain banks. J Chem Neuroanat 20:7–19.
etersen DA (1999) Quantitative histology using confocal microscopy:implementation of unbiased stereology procedures. Methods18:493–507.
uester R, Schröder R (1997) The shrinkage of the human brain stemduring formalin fixation and embedding in paraffin. J NeurosciMethods 75:81–89.
app P, Gallagher M (1996) Preserved neuron number in the hip-pocampus of aged rats with spatial learning deficits. Proc NatlAcad Sci USA 93:9926–9930.
asmussen T, Schliemann T, Sørensen JC, Zimmer J, West MJ(1996) Memory impaired aged rats: no loss of principal and sub-icular neurons. Neurobiol Aging 17:143–147.
eith A, Mayhew TM (1988) Stereology and morphometry in electronmicroscopy: some problems and their solutions (an ultrastructuralpathology publication). London: Taylor and Francis.
oberts N, Cruz-Orive LM, Reid NM, Brodie DA, Bourne M, EdwardsRH (1993) Unbiased estimation of human body composition by theCavalieri method using magnetic resonance imaging. J Microsc
171:239–253.oberts N, Garden AS, Cruz-Orive LM, Whitehouse GH, Edwards RH(1994) Estimation of fetal volume by magnetic resonance imagingand stereology. Br J Radiol 67:1067–1077.
oyet JP (1991) Stereology: a method for analyzing images. ProgNeurobiol 37:433–474.
uss JC, Dehoff RT (2000) Practical stereology, 2nd ed. New York:Plenum Publishers.
andau K (1987) How to estimate the area of a surface using a spatialgrid. Acta Stereol 6:31–36.
chleicher A, Zilles K (1990) A quantitative approach tocytoarchitectonics: analysis of structural inhomogeneities in ner-vous tissue using an image analyser. J Microsc 157:367–381.
chleicher A, Amunts K, Geyer S, Morosan P, Zilles K (1999) Observ-er-independent method for microstructural parcellation of cerebralcortex: a quantitative approach to cytoarchitectonics. Neuroimage9:165–177.
chleicher A, Amunts K, Geyer S, Kowalski T, Schormann T, Pal-omero-Gallagher N, Zilles K (2000) A stereological approach tohuman cortical architecture: identification and delineation of corti-cal areas. J Chem Neuroanat 20:31–47.
chmitz C (1998) Variation of fractionator estimates and its predic-tion. Anat Embryol 198:371–397.
chmitz C, Hof PR (2000) Recommendations for straightforward andrigorous methods of counting neurons based on a computer sim-ulation approach. J Chem Neuroanat 20:93–114.
chmitz C, Materne S, Korr H (1999) Cell-type-specific differences inage-related changes of DNA repair in the mouse brain: molecularbasis for a new approach to understand the selective neuronalvulnerability in Alzheimer’s disease. J Alz Dis 1:387–407.
chmitz C, Dafotakis M, Heinsen H, Mugrauer K, Niesel A, PopkenGJ, Stephan M, Van de Berg WDJ, von Hoersten S, Korr H(2000) Use of cryostat sections from snap-frozen nervous tissuefor combining stereological estimates with histological, cellular,or molecular analyses on adjacent sections. J Chem Neuroanat20:21–29.
chmitz C, Grolms N, Hof PR, Boehringer R, Glaser J, Korr H (2002a)Altered spatial arrangement of layer V pyramidal cells in the mousebrain following prenatal low-dose X-irradiation: a stereologicalstudy using a novel three-dimensional analysis method to estimatethe nearest neighbor distance distributions of cells in thick sec-tions. Cereb Cortex 12:954–960.
chmitz C, Rhodes ME, Bludau M, Kaplan S, Ong P, Ueffing I, VehoffJ, Korr H, Frye CA (2002b) Depression: reduced number of granulecells in the hippocampus of female, but not male, rats due toprenatal restraint stress. Mol Psychiatry 7:810–813.
chmitz C, Born M, Dolezel P, Hof PR, Korr H (2004) Prenatal pro-tracted gamma irradiation with extremely low dose rate over sev-eral days induces massive neuronal loss in rat cerebellum andhippocampus. Neuroscience, in press.
mith CS, Guttman L (1953) Measurement of internal boundaries inthree dimensional structures by random sectioning. Trans AIME197:81–92.
ousa N, Paula-Barbosa MM, Almeida OF (1999) Ligand and subfieldspecificity of corticoid-induced neuronal loss in the rat hippocampalformation. Neuroscience 89:1079–1087.
terio DC (1984) The unbiased estimation of number and size ofarbitrary particles using the disector. J Microsc 134:127–136.
andrup T, Gundersen HJG, Jensen EB (1997) The optical rotator. JMicrosc 186:108–120.
ylings HB, van Eden CG, Hofman MA (1986) Morphometry of size/volume variables and comparison of their bivariate relations in thenervous system under different conditions. J Neurosci Methods18:19–37.
edel Jensen EB (1998) Local stereology. London: World ScientificPublishing.
edel Jensen EB, Gundersen HJG (1993) The rotator. J Microsc
170:35–44.
V
V
v
W
W
W
W
W
W
W
W
W
W
W
W
Z
C. Schmitz and P. R. Hof / Neuroscience 130 (2005) 813–831 831
illena A, Vidal L, Diaz F, Perez De Vargas I (2003) Stereologicalchanges in the capillary network of the aging dorsal lateral genic-ulate nucleus. Anat Rec 274A:857–861.
ogels O (1997) How should brain nuclei be delineated? They don’tneed to be. Trends Neurosci 17:343–344.
on Bartheld C (2002) Counting particles in tissue sections: choices ofmethods and importance of calibration to minimize biases. HistolHistopathol 17:639–648.
eibel ER (1979) Stereological methods, Vol. 1: practical methods forbiological morphometry. London: Academic Press.
eibel ER (1980) Stereological methods, Vol. 2: theoretical founda-tions. London: Academic Press.
eibel ER (1992) Stereology in perspective: a mature scienceevolves. Acta Stereol 11(Suppl I):1–13.
eibel ER, Elias H (1967) Quantitative methods in morphology. Berlin:Springer.
est MJ (1993) New stereological methods for counting neurons.Neurobiol Aging 14:275–285.
est MJ (1999) Stereological methods for estimating the total number
of neurons and synapses: issues of precision and bias. TrendsNeurosci 22:51–61.
est MJ (2001) Design based stereological methods for estimatingthe total number of objects in histological material. Folia Morphol60:11–19.
est MJ (2002) Design-based stereological methods for countingneurons. Prog Brain Res 135:43–51.
est MJ, Gundersen HJG (1990) Unbiased stereological estimation ofthe number of neurons in the human hippocampus. J Comp Neurol296:1–22.
est MJ, Slomianka L (1998) Total number of neurons in the layers ofthe human entorhinal cortex [erratum]. Hippocampus 8:426
est MJ, Slomianka L, Gundersen HJG (1991) Unbiased stereologicalestimation of the total number of neurons in the subdivisions of the rathippocampus using the optical fractionator. Anat Rec 231:482–497.
illiams RW, Rakic P (1988) Three-dimensional counting: an accurateand direct method to estimate numbers of cells in sectioned ma-terial. J Comp Neurol 278:344–352.
illes K (1985) The cortex of the rat: a stereotaxic atlas, pp 69–89.
Berlin: Springer.(Accepted 30 August 2004)(Available online 11 November 2004)
Related Documents