Exp Brain Res (1993) 95:261-270 Experimental BrainResearch Springer-Verlag 1993 Reticulospinal actions on primary afferent depolarization of cutaneous and muscle afferents in the isolated frog neuraxis H. Gonzfilez, I. Jim6nez, P. Rudomin Laboratory of Neural Control, Department of Physiology,Biophysicsand Neurosciences, Centro de Investigacidny de Estudios Avanzados del I.P.N., Apartado Postai 14-740, M~xico D.F. 07000, M~xico Received: 23 October 1992 / Accepted: 12 February 1993 Abstract. The effects of the brainstem reticular formation on the intraspinal excitability of low threshold cutaneous and muscle afferents were studied in the frog neuraxis isolated together with the right hindlimb nerves. Stimula- tion of low threshold fibers (less than two times threshold) in cutaneous nerves produced short latency, negative field potentials in the ipsilateral dorsal neuropil (200-400 gm depth) that reversed to positivity at deeper regions (500~700 gm). Stimulation of low threshold fibers (less than two times threshold) in muscle nerves pro- duced, instead, negative responses that acquired their maximum amplitude in the ventral neuropil (700-900 gm depth). These electrophysiological findings suggest, in agreement with observations in the cat, that low threshold cutaneous and muscle afferents end at different sites in the spinal cord. Intraspinal microstimulation ap- plied within the dorsal neuropil produced antidromic re- sponses in low threshold cutaneous afferents that were increased in size following stimulation of the dorsal or ventral roots, as well as of the brainstem reticular forma- tion. This increase in excitability is interpreted as being due to primary afferent depolarization (PAD) of the in- traspinal terminals of cutaneous fibers. Antidromic re- sponses recorded in muscle nerves following intraspinal stimulation within the ventral neuropil were also in- creased following conditioning stimulation of adjacent dorsal or ventral roots. However, stimulation of the bul- bar reticular formation produced practically no changes in the antidromic responses, but was able to inhibit the PAD of low threshold muscle afferents elicited by stimu- lation of the dorsal or ventral roots. It is suggested that the PAD of low threshold cutaneous and muscle afferents is mediated by independent sets of interneurons. Reticu- lospinal fibers would have excitatory connections with the interneurons mediating the PAD of cutaneous fibers and inhibitory connections with the interneurons mediat- ing the PAD of muscle afferents. Although our results provide no direct information on whether the reticu- lospinal depression of the PAD elicited in low threshold Correspondence to: P. Rudomin muscle afferents is due to inhibition along the pathways producing PAD of muscle spindle or of tendon organ afferents, it seems likely by analogy with what has been seen in the cat spinal cord - that these inhibitory actions are mostly restricted to the pathways producing PAD in the terminal arborizations of muscle spindle afferents. These results emphasize the specificity of the descending control of the synaptic efficacy of low threshold cuta- neous and muscle afferents which could be of importance for motor performance. Key words: Bulbo-spinal inhibition - Inhibition of PAD - Spinal cord - Brainstem - Frog Introduction Information on the supraspinal control of synaptic effec- tiveness of muscle and skin afferents has been mostly derived from studies made in mammals (for review, see Rudomin 1990 a, b). In the cat, stimulation of the motor cortex and of the bulbar reticular formation produces primary afferent depolarization (PAD) of tendon organ and of skin afferents, but inhibits the PAD of muscle spindle afferents. Recent studies performed in humans have indicated that during the execution of a voluntary movement there is an increase in the synaptic effective- ness of muscle spindle afferents that innervate the con- tracting muscles, and a decrease in the synaptic effective- ness of muscle spindle afferents innervating non-con- tracting homonymous muscles (Hultborn et al. 1987). These findings support the existence of a highly elaborat- ed central control of the afferent input that may play an important role in the execution of specific motor tasks. Although there are strong similarities in the functional organization of the pathways that produce PAD in mam- mals and amphibia, there are also important differences which need to be considered in current models of motor behavior. For example, in the cat, stimulation of motor axons has no presynaptic actions on afferent fibers, but

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Exp Brain Res (1993) 95:261-270
Experimental Brain Research �9 Springer-Verlag 1993
Reticulospinal actions on primary afferent depolarization of cutaneous and muscle afferents in the isolated frog neuraxis H. Gonzfilez, I. Jim6nez, P. Rudomin
Laboratory of Neural Control, Department of Physiology, Biophysics and Neurosciences, Centro de Investigacidn y de Estudios Avanzados del I.P.N., Apartado Postai 14-740, M~xico D.F. 07000, M~xico
Received: 23 October 1992 / Accepted: 12 February 1993
Abstract. The effects of the brainstem reticular formation on the intraspinal excitability of low threshold cutaneous and muscle afferents were studied in the frog neuraxis isolated together with the right hindlimb nerves. Stimula- tion of low threshold fibers (less than two times threshold) in cutaneous nerves produced short latency, negative field potentials in the ipsilateral dorsal neuropil (200-400 gm depth) that reversed to positivity at deeper regions (500~700 gm). Stimulation of low threshold fibers (less than two times threshold) in muscle nerves pro- duced, instead, negative responses that acquired their maximum amplitude in the ventral neuropil (700-900 gm depth). These electrophysiological findings suggest, in agreement with observations in the cat, that low threshold cutaneous and muscle afferents end at different sites in the spinal cord. Intraspinal microstimulation ap- plied within the dorsal neuropil produced antidromic re- sponses in low threshold cutaneous afferents that were increased in size following stimulation of the dorsal or ventral roots, as well as of the brainstem reticular forma- tion. This increase in excitability is interpreted as being due to primary afferent depolarization (PAD) of the in- traspinal terminals of cutaneous fibers. Antidromic re- sponses recorded in muscle nerves following intraspinal stimulation within the ventral neuropil were also in- creased following conditioning stimulation of adjacent dorsal or ventral roots. However, stimulation of the bul- bar reticular formation produced practically no changes in the antidromic responses, but was able to inhibit the PAD of low threshold muscle afferents elicited by stimu- lation of the dorsal or ventral roots. It is suggested that the PAD of low threshold cutaneous and muscle afferents is mediated by independent sets of interneurons. Reticu- lospinal fibers would have excitatory connections with the interneurons mediating the PAD of cutaneous fibers and inhibitory connections with the interneurons mediat- ing the PAD of muscle afferents. Although our results provide no direct information on whether the reticu- lospinal depression of the PAD elicited in low threshold
Correspondence to: P. Rudomin
muscle afferents is due to inhibition along the pathways producing PAD of muscle spindle or of tendon organ afferents, it seems likely by analogy with what has been seen in the cat spinal cord - that these inhibitory actions are mostly restricted to the pathways producing PAD in the terminal arborizations of muscle spindle afferents. These results emphasize the specificity of the descending control of the synaptic efficacy of low threshold cuta- neous and muscle afferents which could be of importance for motor performance.
Key words: Bulbo-spinal inhibition - Inhibition of PAD - Spinal cord - Brainstem - Frog
Introduction
Information on the supraspinal control of synaptic effec- tiveness of muscle and skin afferents has been mostly derived from studies made in mammals (for review, see Rudomin 1990 a, b). In the cat, stimulation of the motor cortex and of the bulbar reticular formation produces primary afferent depolarization (PAD) of tendon organ and of skin afferents, but inhibits the PAD of muscle spindle afferents. Recent studies performed in humans have indicated that during the execution of a voluntary movement there is an increase in the synaptic effective- ness of muscle spindle afferents that innervate the con- tracting muscles, and a decrease in the synaptic effective- ness of muscle spindle afferents innervating non-con- tracting homonymous muscles (Hultborn et al. 1987). These findings support the existence of a highly elaborat- ed central control of the afferent input that may play an important role in the execution of specific motor tasks. Although there are strong similarities in the functional organization of the pathways that produce PAD in mam- mals and amphibia, there are also important differences which need to be considered in current models of motor behavior. For example, in the cat, stimulation of motor axons has no presynaptic actions on afferent fibers, but

262
produces postsynaptic inhibition of motoneurons via Renshaw cells (see Davidoff and Hackman 1984). How- ever, in the frog, PAD (and presynaptic inhibition) is elic- ited by stimulation of sensory nerves, as well as by stimu- lation of motor axons (Grinnell 1966). Although the PAD elicited by sensory and motor axons in the frog spinal cord appears to be mediated by partially indepen- dent pathways (see Galindo and Rudomin 1978), it is not known if they are subjected to the same or to a different supraspinal control.
We have shown recently that, in the isolated neuraxis of the frog, conditioning stimulation of the brainstem reticular formation (RF) with intensities below those re- quired to produce a dorsal root potential (DRP) reduces the amplitude of the DRPs elicited either by or thodromic stimulation of muscle and cutaneous afferents or by an- tidromic stimulation of motoneuron axons (Gonz~ilez et al. 1992a). By analogy with observations in the cat, we proposed that reticulospinal fibers produce PAD in cuta- neous fibers and inhibit transmission along the pathways producing PAD of muscle spindle afferents. However, analysis of the changes of DRPs produced by interaction of segmental and descending inputs does not allow dis- closure of the target fibers that are being affected. This can be investigated by examining the intraspinal ex- citability changes produced in specific sets of afferent fibers (Wall 1958; Eccles et al. 1962) or by recording the PAD of single, functionally identified afferent fibers (Jim~nez et al. 1988). Our initial approach to this problem was to analyze the effects produced by stimulation of the brain-stem RF on the intraspinal excitability of popula- tions of low threshold cutaneous and muscle afferents.
The results obtained support our previous proposal that, in the frog, RF stimulation facilitates the PAD that is produced in low threshold cutaneous afferents by or- thodromic stimulation of dorsal roots, as well as by an- tidromic stimulation of the ventral roots. On the other
hand, the PAD that is produced in low threshold muscle afferents that terminate in the ventral neuropil (assumed to arise from muscle spindles) is inhibited by RF stimula- tion. These findings stress the similarities in the action of reticulospinal pathways on the synaptic efficacy of low threshold cutaneous and muscle afferents in the cat and frog spinal cord, and suggest that supraspinal control of the synaptic effectiveness of afferent fibers may play an important role in motor performance. A preliminary ac- count of these findings has been published (Gonzfilez et al. 1992b).
Materials and methods
The experiments were made on the isolated neuraxis of the frog (Rana palmipens). This preparation included rostral structures up to the cerebellum and the hindlimb nerves of the right side (Fig. 1). The dissection was made in a chamber placed on crushed ice and filled with cooled Ringer solution saturated with 02. The Ringer compo- sition was (mM): NaC1 100; KC1 2.0; CaC12 1.5; NaHCO 3 20 and glucose 5.5. The pH was adjusted to 7.42.
The neuraxis was exposed by removing the dorsal bone of the vertebrae. All ventral roots and all dorsal roots, with the exception of the right 9th and 10th, were also sectioned. The dorsal columns were cut at the level of the brachial plexus to eliminate possible antidromic activation of afferent fibers following stimulation of the brainstem. The following nerves in the right hindlimb were dissect- ed: gluteus (Glut), gracilis (Grac), superficial peroneus (SP), sural (SU), tibialis (Tib) and abductor magnus (Am, see Carpenter and Rudomin 1973).
After the dissection was completed, the preparation was trans- ferred to a recording chamber and maintained with oxygenated Ringer solution at 14~ Patches of pia were removed from the dorsal surface of the lumbar spinal cord to facilitate insertion of the micropipettes. Peripheral sensory nerves and ventral or dorsal roots were stimulated using bipolar suction electrodes. The brainstem was stimulated by means of a pair of fine tungsten electrodes insert- ed into a double-barrelled glass micropipette. The protruding tung- sten tips were sharpened by electrolysis, leaving uninsulated seg- ments of 10-15 gm diameter and 50-100 gm long separated by
A1
S3
j.s"
. ~ " _ 2 - 1 _
$1
Fig. 1. Diagram of the method. A1, amplifier used to record intraspinal field potentials; A2, amplifier used to record antidromic responses from peripheral nerves; $I, electrodes to stimulate the homosegmen- tal ventral root (VR); $2, electrodes to stimulate a dorsal root (DR) in an adjacent segment; $3, bipolar electrode used to stimulate the bulbar reticular for- mation (RF); S 4, intraspinal stimulus for excitability testing; Grac, gracilis; Tib, tibialis
263
A G r a c i l i s
B T i b i a l i s
i
o i
I
I 100
r ! i
.4-- r- I I
- i ~ 0 i n
-1 .0 " C
mV
- 0 , 5 -
C u t a n e o u s A f f e r e n t s
A~--- - q k~A...A ~.A 7 Gracilis
Superficial ~ Peroneus
o o !
I
i
i I
I I
i
-o +0.5 200 ~ ~ M u s c l e A f f e r e n f s
" -6 -1 .0 "1 O__O_O/c~ ~, l>Tibialis , ~ / ~ O A b d u c t o r 300 mVJ / I>~ ~0.,.0 Magnus : - 0.51
soo ~ , ,~ 0 " - - -
' -o 0 Cutaneous I I
600 ----4 ;
I
~oo ~ \1 ~vj
Depth I I
( / a m ) ' 10 ms
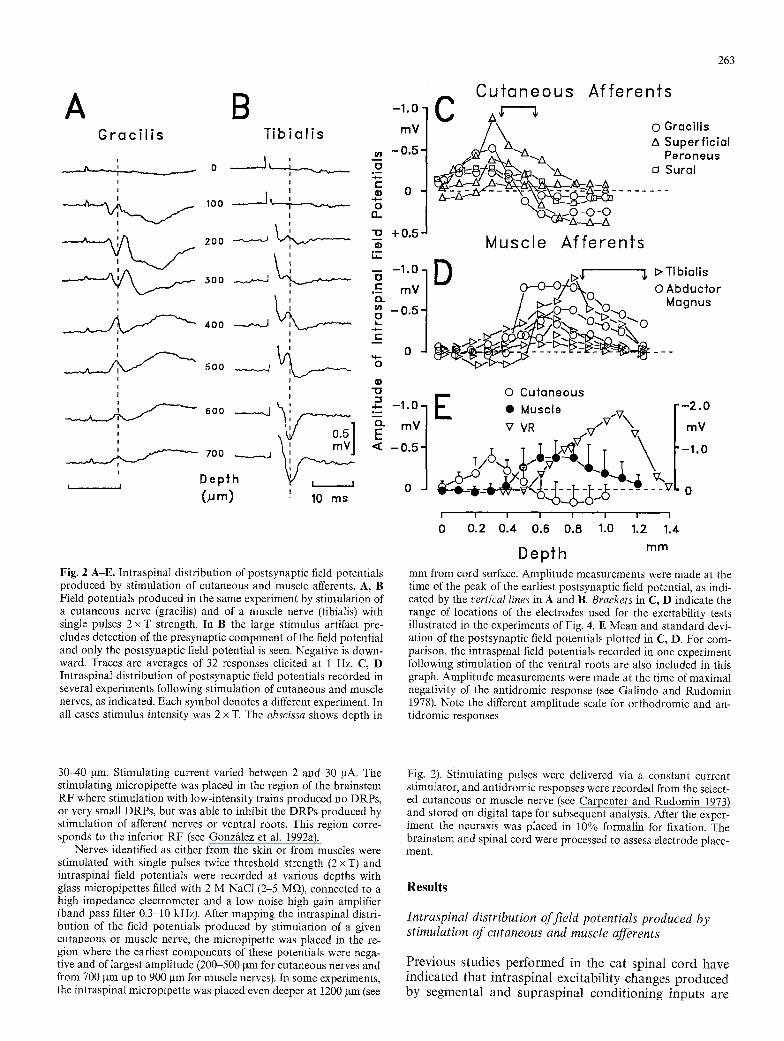
Fig. 2 A-E. Intraspinal distribution of postsynaptic fieId potentials produced by stimulation of cutaneous and muscle afferents. A, B Field potentials produced in the same experiment by stimulation of a cutaneous nerve (gracilis) and of a muscle nerve (tibialis) with single pulses 2 x T strength. In B the large stimulus artifact pre- cludes detection of the presynaptic component of the field potential and only the postsynaptic field potential is seen. Negative is down- ward. Traces are averages of 32 responses elicited at 1 Hz. C, D Intraspinal distribution of postsynaptic field potentials recorded in several experiments following stimulation of cutaneous and muscle nerves, as indicated. Each symbol denotes a different experiment. In all cases stimulus intensity was 2 x T. The abscissa shows depth in
-I -1 .0 E
~- mV v VR d V / v mV - 0 . 5 -1 .0
- [ l I I I l I i I
0 0.2 0.4 0.6 0.8 1.0 1.2 1.4
D e p t h mm
mm from cord surface. Amplitude measurements were made at the time of the peak of the earliest postsynaptic field potential, as indi- cated by the vertical lines in A and B. Brackets in C, D indicate the range of locations of the electrodes used for the excitability tests illustrated in the experiments of Fig. 4. E Mean and standard devi- ation of the postsynaptic field potentials plotted in C, D. For com- parison, the intraspinal field potentials recorded in one experiment following stimulation of the ventral roots are also included in this graph. Amplitude measurements were made at the time of maximal negativity of the antidromic response (see Galindo and Rudomin 1978). Note the different amplitude scale for orthodromic and an- tidromic responses
30-40 pm. Stimulating current varied between 2 and 30 gA. The stimulating micropipette was placed in the region of the brainstem RF where stimulation with low-intensity trains produced no DRPs, or very small DRPs, but was able to inhibit the DRPs produced by stimulation of afferent nerves or ventral roots. This region corre- sponds to the inferior RF (see Gonzfilez et al. 1992a).
Nerves identified as either from the skin or from muscles were stimulated with single pulses twice threshold strength (2 x T) and intraspinal field potentials were recorded at various depths with glass micropipettes filled with 2 M NaCI (2-5 M~), connected to a high impedance electrometer and a low noise high gain amplifier (band pass filter 0.3-10 kHz). After mapping the intraspinal distri- bution of the field potentials produced by stimulation of a given cutaneous or muscle nerve, the micropipette was placed in the re- gion where the earliest components of these potentials were nega- tive and of largest amplitude (200-500 ~tm for cutaneous nerves and from 700 gm up to 900 gm for muscle nerves), In some experiments, the intraspinal micropipette was placed even deeper at 1200 gm (see
Fig. 2). Stimulating pulses were delivered via a constant current stimulator, and antidromic responses were recorded from the select- ed cutaneous or muscle nerve (see Carpenter and Rudomin 1973) and stored on digital tape for subsequent analysis. After the exper- iment the neuraxis was placed in 10% formalin for fixation. The brainstem and spinal cord were processed to assess electrode place- ment.
Results
Intraspinal distribution of field potentials produced by stimulation of cutaneous and muscle afferents
Prev ious s tudies p e r f o r m e d in the cat sp ina l co rd have i n d i c a t e d t ha t i n t r a s p i n a l exc i tab i l i ty changes p r o d u c e d by s egm en ta l a n d s u p r a s p i n a l c o n d i t i o n i n g i n p u t s are

264
more readily detected when the excitability testing mi- croelectrode is placed in the region of the largest, focal postsynaptic potentials produced by stimulation of the afferent fibers (Rudomin et al. 1983); i.e. in the region where a significant number of afferent fibers synapse with second order neurons (Coombs et al. 1956). According to Szekely (1976), Frank and Westerfield (1982), Jhaveri and Frank (1983) and Szekely and Antal (1984), in the spinal cord of the frog the small afferent fibers enter medially and terminate in the substantia gelatinosa. The coarser afferent fibers enter more laterally, run deeper and end in a triangular shaped area at the base of the dorsal horn (dorsal neuropil) and in the intermediate gray matter (ventral neuropil). Afferent fibers of cutaneous origin end in the dorsal neuropil, and afferent fibers of muscular origin end in the ventral neuropil (Jhaveri and Frank 1983). Szekely has suggested that the axons that leave the ventral neuropil and invade the ventral horn to synapse with motoneurons arise from muscle spindles and are equivalent to the Ia fibers of mammals (Szekely 1976; Szekely et al. 1982).
Figure 2 shows the intraspinal distribution of the field potentials recorded in one experiment following stimula- tion of a predominantly cutaneous nerve (gracilis, panel A) and of a predominantly muscle nerve (tibialis, panel B). Stimulus strength was set to twice the intensity re- quired to produce a detectable potential (2 x T). The in- traspinal field potentials produced by stimulation of the cutaneous nerve were negative in the most dorsal regions of the spinal cord. They were of maximal amplitude be- tween 200 and 300 gm from the cord dorsum and re- versed in sign deeper in the cord (between 400 and 500 gin). These potentials usually have several components. In the experiment illustrated in Fig. 2A, the earliest com- ponent had an onset latency of about 3.4 ms. This com- ponent probably represents the potential field generated by monosynaptic activation of second order neurons (Holemans et al. 1966). The onset latency of the earliest components produced by stimulation of cutaneous nerves in all experiments ranged between 2.8 and 3.4 ms (n = 6). Considering the conduction distances involved in these recordings, it seems likely that the fastest afferent fibers involved in the generation of these responses had conduction velocities between 17.7 and 21.4 m/s. These values are in agreement with available measurements (see Spray 1976).
Stimulation of the tibial nerve produces a fast presy- naptic and slow postsynaptic field potential in the ventral neuropil (Glusman and Rudomin 1974). Figure 2B shows the postsynaptic field potential recorded at various depths. This potential was positive between 200 and 500 gm from the cord dorsum and reversed to negative be- tween 600 and 700 gin. This potential had an onset laten- cy of 3.0 ms. The onset latencies of the negative responses produced in other experiments by stimulation of muscle nerves (tibialis and abductor magnus) varied between 2.5 and 3.3 ms (n = 7), suggesting monosynaptic activation of second order neurons (see Glusman and Rudomin 1974).
Figure 2C, D shows the intraspinal distribution of field potentials recorded in individual experiments fol- lowing stimulation of low threshold cutaneous and mus-
cle afferents. It may be seen that these field potentials had a different spatial distribution in the spinal cord. These differences are emphasized in the plot of Fig. 2E that shows the means and standard deviations of the in- traspinal potential fields depicted in Fig. 2C, D. For com- parison, Fig. 2E also shows the intraspinal field poten- tials recorded in one experiment following antidromic stimulation of a ventral rootlet. These potentials were negative and largest between 900 and 1200 gm depth and result from antidromic activation of motoneurons (see Glusman and Rudomin 1974; Galindo and Rudomin 1978).
The above observations agree reasonably well with the available anatomical data pertaining to the sites of termination of large muscle and cutaneous afferents in the spinal cord of the frog, and provide good landmarks to place the microelectrode for testing the intraspinal ex- citability of the afferent fibers. In fact, even when record- ing from mixed nerves, careful electrode placement in the regions where the field potentials were largest ensured that antidromic responses were mostly, if not totally, of cutaneous or muscle afferents, respectively.
Excitability changes of the intraspinal terminals of low threshold cutaneous afferents
Dorsal root and RF conditioning. As initially reported by Carpenter and Rudomin (1973), intraspinal microstimu- lation applied in the dorsal horn produces antidromic responses in cutaneous nerves. In the experiment illus- trated in Fig. 3 A and B, the stimulating micropipette was placed 400 gm from the cord dorsum and antidromic responses were recorded from the gracilis nerve. A pulse of 6 gA produced the antidromic responses shown in trace 1 of Fig. 3A. These responses had an onset latency of 3 ms. Since the conduction distance was of 51 mm, the calculated conduction velocity of the fastest fibers would be of the order of 17 m/s. Trace 2 in Fig. 3A shows that the amplitude of the antidromic response was increased to 117%, when the adjacent dorsal root was stimulated with a single pulse 1.6 times threshold intensity (x T) applied 18 ms before the test pulse. The increase in the amplitude of the antidromic response is due to the addi- tional firing of afferent fibers that show a subthreshold depolarization following the conditioning stimulation of the dorsal root; i.e. to primary afferent depolarization (PAD, see Wall 1958 and Carpenter and Rudomin 1973). As shown by the open and filled circles in Fig. 3B, condi- tioning stimulation of the dorsal root facilitated the an- tidromic responses produced by intraspinal stimulation throughout a wide range of intensities (from 3.3 up to 10 gA) that activated fibers with intraspinal thresholds up to 3 x T; i.e. of the fastest and most excitable myelinated afferents.
Stimulation of the bulbar reticular formation with a train of four shocks, 200 Hz and 15 gA, applied 80 ms before the test pulse also increased the antidromic re- sponses elicited in the gracilis nerve. In the example illus- trated in Fig. 3A trace 3, the increase was of 135%. As illustrated by the filled triangles in Fig. 3B, stimulation of

265
C U T A N E O U S A F F E R E N T S
A Gracilis
Affl L
DReAff2 L
RF-*Aff =~= 3
RF+DR..-)Af f + 100
5 m s
la.i
E I . V ) 0 CJ
Z
B
100"
(V.)
50.
O' :.3.3 1.0
6.6 (,uA) 9.9 2.0 (xT) 3.0
INTRASPINAL STIMULATION
C Superficial Peroneus
Aff .... 1 -J I /~F-
VR--*Aff T 2 ----JV,/~/,, ~
RF-*Aff .....
3 --JL f - - m
R F+VR~Af f -] 10D
jaV 4. ---Jp, ],,~__.~
5 m s
D
100' Ld
o.. i,(/1 O s 50
<N Z
~. .'"":"'"'"'A ....... i ........ '
I f
6:z 9:4 Is 2.0 3,0 (xT) 4.0
INTRASPINAL STIMULATION
Fig. 3 A-D. Brainstem conditioning stimu- lation produces PAD of cutaneous affer- ents and facilitates the PAD produced by segmental inputs. A Trace 1 shows the an- tidromic responses (aft) recorded in the gracilis nerve following single pulse stimu- lation (6 gA) of the dorsal neuropil (400 Hm depth, see insert in panel B). Trace 2 shows that conditioning stimulation of the adjacent dorsal root with a single pulse 1.6 x T applied 18 ms before the test pulse increases the antidromic response pro- duced by the same intraspinal stimulus. Trace 3 shows the increase in the an- tidromic response produced with condi- tioning stimulation of the RF (four shocks, 200 Hz, 15 gA applied 80 ms before the test pulse). Trace 4 shows the changes in the antidromic response produced by con- current DR and RF stimulation. Hatched horizontal bar indicates the amplitude of the unconditioned test responses depicted in trace l. Black horizontal bar shows the amplitude of the antidromic response that was facilitated by DR conditioning stimu- lation (response shown in trace 2). All traces are averages of 32 responses elicited at I Hz. B Effects of DR and RF stimula- tion on the area of the antidromic respons- es produced by intraspinal stimuli of differ- ent strengths. Curves obtained from data partly illustrated in A. C, D Same format as A and B, respectively. Data from anoth- er experiment. Test antidromic responses were recorded from the superficial per- oneus nerve following a single pulse (8 HA strength) applied at 500 gm depth. Condi- tioning stimulation of the VR was a single pulse 2.4 • T applied 25 ms before the test pulse. The RF was stimulated with a train of four shocks, 200 Hz, 20 gA applied 90 ms before the test pulse
the reticular formation increased the area of the an- tidromic responses throughout the whole explored range. In this experiment the effects of the RF were slightly greater or equal to those produced by dorsal root stimu- lation.
Concurrent stimulation of the RF and D R increased the antidromic responses to 157% of the control value (Fig. 3A trace 4). This effect was slightly above the value obtained by algebraic addition of the effects produced by separate activation of both pathways. A more than linear summat ion of the effects produced by concurrent D R and RF stimulation was observed for antidromic re- sponses that were 2 0 7 0 % of maximal size (evoked by intraspinal stimuli smaller than 2 x T; see Fig. 3B). This suggests convergence of afferent and reticulospinal exci- tatory inputs on common interneurons along the path- ways producing PAD of low threshold cutaneous affer- ents.
Figure 4A summarizes the results obtained from four different experiments where we tested the intraspinal ex- citability changes produced in low threshold cutaneous fibers by D R and RF stimulation. To allow comparison
between data obtained from different experiments, we calculated the ratio of stimulus strengths necessary to produce conditioned responses of 25% and 50% of the area of the unconditioned responses. These values were selected because they are in the region of max imum sensi- tivity of the input-output curves and well below the satu- ration level. For example, in the experiment shown in Fig. 3B, stimuli of 1.85 x T were required to produce uncondi- tioned responses that were 50% of maximal area. Follow- ing dorsal root conditioning, the stimuli required to pro- duce a response 50% of the max imum were reduced to 1.72 x T. The ratio between the two stimulus strengths (control/conditioned) gives an indication of the excitabil- i ty change produced by D R conditioning which was, in this case, 1.08; i.e. conditioning stimulation of the dorsal root increased the excitability by 8%. In those three ex- periments in which RF and of D R stimulation produced a relatively small excitability increase (range 4-14%), concurrent stimulation of the RF and the D R produced effects that were larger than the sum of those produced by separate stimulation of each pathway (open triangles, cir- cles and diamonds in Fig. 4A). The more than linear

266
2 . 0
Ld (,3 Z
"r- ( J
>...
,,_I
F-
X lal
1 . 0 "
CUTANEOUS AFFERENTS
A B
#
RF D'R RF RF DR RF RF VR RF RF VR RF + + "t- 4-
DR DR VR VR
25N 5 0 ~ 25N 5 0 g
z.o M CLE AFFERENTS
C D
_5 ~ 1.5 ~ ~ i
lal
1.0 . . . . . . . . . . . . . . . . . . . . . . . . . . .
RF DR RF RF DR RF RF VR RF RF VR RF + + + 4- DR DR VR VR
2 5 ~ 5 0 ~ 2 5 ~ 5 0 ~
Fig. 4 A-D. Summary of the intraspinal excitability changes of cutaneous and muscle afferents produced by segmental and de- scending inputs. A, B Cutaneous afferents; C, D Muscle afferents. Excitability changes were calculated from the ratio of the stimulus strengths (expressed as multiples of minimal threshold) required to produce control and conditioned antidromic responses of the same amplitude (set to 25% and 50% of maximum size, as indicated). Ratios larger than 1 indicate an increase in excitability, that is, in PAD, Note opposite actions of RF conditioning on PAD of cuta- neous and muscle afferents. In these experiments, DR and VR stim- ulation was one pulse 1.5-2.0 x T applied 35-50 ms before the ex- citability testing pulse. RE stimulation were trains of four to six pulses at 100-200 Hz, 10-40 gA applied 70-100 ms before the excitability testing. Location of RF electrodes was 0.5-1 mm rostral to the obex, 0.4 to 0.7 mm lateral, 0.6-1.2 mm depth. Range of intraspinal location of the excitability testing micropipettes is shown by the horizontal brackets in Fig. 2C, D
addition of the effects suggests facilitatory interactions between inputs converging onto common elements. In the fourth experiment (open squares), separate stimula- tion of the RF and of the DR produced a larger excitabil- ity increase, and the effects produced by simultaneous stimulation of both pathways were smaller than those obtained by adding the effects produced by each input. The less than linear interaction between both inputs
could be due to occlusion or to inhibition (see Baldissera et al. 1981).
Ventral root and RF conditioning. In the cat spinal cord, antidromic stimulation of ventral roots produces recur- rent postsynaptic inhibition of motoneurons but no PAD (Renshaw 1946; see Davidoff and Hackman 1984). How- ever, in the frog, antidromic stimulation of the ventral roots does produce dorsal root potentials and presynap- tic inhibition (Grinnell 1966; Carpenter and Rudomin 1973; Glusman and Rudomin 1974; Shapovalov and Shiriaev 1980). It was established by Carpenter and Rudomin (1973) that these DRPs resulted from PAD generated in cutaneous as well as in muscle afferents. We have shown previously that RF stimulation inhibits the DRPs elicited by ventral root stimulation (Gonz/dez et al. 1992a), but the type of afferent fibers whose PAD was inhibited by this supraspinal input was not determined.
The data depicted in Fig. 3C-D illustrate the effects produced by VR and RF stimulation on the PAD of afferent fibers in the superficial peroneus nerve. Available evidence indicates that the afferent fibers in this nerve carry information from skin afferents (Gaupp 1897; Car- penter and Rudomin 1973). To further ensure that the excitability tests were indeed restricted to cutaneous af- ferents, the intraspinal microstimulation was applied in the dorsal horn rather superficially (500 gm), as indicated by the insert in Fig. 3D.
Trace 1 in Fig. 3C illustrates the antidromic response produced by a single pulse of 8 pA. Trace 2 shows the increase in the antidromic response produced by a 2.4 x T conditioning pulse applied to the homosegmental ventral root 25 ms before the test pulse. It may be seen that the amplitude of the antidromic response increased to 136% relative to the unconditioned test response. Conditioning stimulation of the RF also produced PAD. A train of four shocks at 200 Hz and of 20 gA applied 90 ms before the test pulse increased the antidromic re- sponse to 112% of the control value (Fig. 3C trace 3). Concurrent stimulation of the homosegmental ventral root and of the RF led to a marked increase in the an- tidromic responses (155%), slightly above that which would be obtained by adding the effects produced by separate stimulation of these two pathways (Fig. 3C trace 4).
The graph in Fig. 3D illustrates the effects produced by RF and VR conditioning throughout the whole range of antidromic responses. It may be seen that with the selected VR and RF stimuli, both conditioning proce- dures produced about the same increase in the an- tidromic responses (filled circles and filled triangles) and that concurrent stimulation of both pathways produced a larger effect (open triangles). That is, during concurrent VR and RF stimulation there was a reduction in the strength of the intraspinal stimulus required to produce an antidromic response of a constant size.
Figure 4B shows the results obtained in four experi- ments where we tested the effects of separate and concur- rent RF and VR stimulation on the excitability of low threshold cutaneous afferents. In one case (filled dia- monds), the effects produced by concurrent stimulation

of the RF and VR were less than the algebraic sum of the effects produced by the activation of each pathway, sug- gesting occlusion. In two cases (filled squares and filled circles), the interactions obtained with responses 50% of max imum were facilitatory, and in one case (filled trian- gles) the interactions were facilitatory for responses 25% of max imum and less than linear for responses 50% of maximum.
The above results support the existence of reticu- lospinal pathways that are able to depolarize low threshold cutaneous afferents and converge with the pathways producing PAD following stimulation of the ventral roots. The PAD of cutaneous fibers produced by the antidromic activation of the motor axons has proba- bly some relation with the finding of Stehouwer and Farel (1981) of PAD that is related to motor activity during episodes of fictive swimming in the tadpole.
Intraspinal excitability changes of low threshold muscle afferents
Dorsal root and RF conditioning. Analysis of the effects of conditioning stimulation produced by segmental and de- scending pathways on the PAD of muscle afferents was
267
made using the same experimental protocol as that uti- lized for the analysis of cutaneous fibers. The only differ- ence was that, in this case, the intraspinal stimulus was applied deeper in the spinal cord and the antidromic re- sponses were recorded from a muscle nerve. In the exam- ple depicted in Fig. 5A, B, the stimulating micropipette was placed at 1200 gm below the dorsal surface, already within the motor pool, and the antidromic responses were recorded from the tibialis nerve. This nerve contains fibers mostly of muscular origin, al though it may have some cutaneous afferents as well. However, stimulation at this depth ensures that antidromic responses are essen- tially from muscle afferents (see Fig. 2D, E). As shown in Fig. 5A traces 1 and 2, stimulation of the adjacent D R with a pulse 1.5 x T applied 10 ms before the test stimulus increased the amplitude of the antidromic spike to 143%. This confirms the initial findings of Carpenter and Rudomin (1973). Unlike the situation observed in cuta- neous fibers, conditioning stimulation of the RF (four shocks at 200 Hz 20 ~tA, applied 60 ms before the test pulse) had practically no effect on the antidromic re- sponses of the muscle afferents (Fig. 5A trace 3). Howev- er, RF stimulation was able to reduce the amplitude of the facilitated antidromic response (to 126%, see Fig. 5A
MUSCLE AFFERENTS
A T i b i a l i s
A f f i ~ 1 - 100,
,,, (~)
T DR-*Aff E 2 ~--{"-'~,JV,./.-"~ ,.mo tJ
. <~ 50. RF~Aff i - ~ , ~ '"
3 �9 < a F- Z
RF+D R-*AIf "1"" 100 ~: 4. . . . . . . / k , / ~ ] ~v 0
5 m s
B
l~m ~ ~/~ ..-'"'""A" �9
4:0 8:0 12'.0 (;JA) 16:0 1.0 2.0 ,:3.0 ('xT) 4 .0
INTRASPINAL STIMULATION
C T i b i a l i s
Af f _ _ ~ " 1 ~ ,.,
VR-~AI f ,,m
IM O
RF "'" ~ Z
RF+VR Aff -/F ]2.0o
5 m s
D
100- (~)
50-
0 4.0 1.0
. . ,A
slo O,A) 12:o z.o (xT) 3.0
INTRASPINAL STIMULATION
Fig. 5 A-D. Brainstem conditioning stimu- lation inhibits the PAD elicited in muscle afferents by dorsal and ventral root stimu- lation. Same format as in Fig. 3. A An- tidromic responses produced by intraspinal microstimulation with single pulses of con- stant strength (10 pA) applied to the ven- tral neuropil at 1200 pm depth. Trace 1 control response; Trace 2 conditioning stimulation of the adjacent dorsal root (a single pulse 1.5 x T applied 10 ms before the test stimulus); Trace 3 conditioning stimulation of the RF (four shocks at 200 Hz, 20 pA applied 60 ms before the test pulse); Trace 4 concurrent RF and DR stimulation. Note that RF stimulation had practically no effect on the antidromic re- sponses, but was able to reduce the facili- tation produced by DR stimulation. B Ef- fects of DR and RF stimulation on an- tidromic responses produced by different strengths of intraspinal microstimulation. C, D Same format as A and B, but data from a different experiment. Antidromic re- sponses in the tibial nerve were produced with intraspinal pulses of 6.4 pA (1.8 x T) applied at 900 ~tm depth. VR stimulus was a single pulse 2.5 xT applied 35 ms before the test stimulus. RF stimulus was four shocks at 200 Hz, 20 gA applied 60 ms be- fore the test pulse

268
trace 4). As shown in Fig. 5B, the inhibitory effect of the RF was observed throughout the whole range of an- tidromic responses up to 4 x T.
Figure 4C summarizes the results obtained in five dif- ferent experiments where we tested the effects of DR and RF conditioning on the antidromic responses of muscle afferents in the tibialis nerve. This figure shows very clearly that low threshold muscle afferents were depolar- ized by DR stimulation and very little or not at all by RF stimulation. Nevertheless, conditioning stimulation of the RF produced a marked reduction in the excitability increase produced by DR stimulation. This depression was evident for responses that were 25% or 50% of max- imum, and appears not to depend on the magnitude of the excitability increase produced by DR stimulation. Hence, it seems very likely that the depression produced by RF stimulation was due to inhibition of the PAD.
Ventral root and RF conditioning. As shown by Carpenter and Rudomin (1973), antidromic stimulation of the ven- tral roots also produces PAD of muscle afferents. Trace 1 in Fig. 5C shows the antidromic spike recorded from the tibialis nerve following an intraspinal stimulus of 6.4 gA applied at 900 gm depth. Trace 2 shows that conditioning stimulation of the homosegmental ventral root (2.5 x T strength applied 35 ms before the test pulse) increased the antidromic response to 166% relative to control. Unlike with VR stimulation, RF stimulation (four pulses at 200 Hz, 18 gA applied 80 ms before the test pulse) had prac- tically no effect on the antidromic spike recorded from the tibialis nerve (trace 3). However, as shown in trace 4, RF stimulation reduced very effectively the facilitation produced by VR conditioning (from 166% to 110%). The inhibitory action of the RF was observed over the whole range ofintraspinal stimuli up to 3 x T strength (Fig. 5D).
Figure 4D summarizes the results obtained from five different experiments in which RF stimulation produced practically no PAD, but was nevertheless able to inhibit the PAD produced by VR stimulation. It should be noted that in two experiments VR stimulation produced a large increase in the intraspinal excitability of the muscle affer- ents, yet there was appreciable inhibition by RF condi- tioning stimulation.
Discussion
The data presented here, together with data from other investigators (Szekely et al. 1982; Jhavery and Frank 1983; Szekely and Antal 1984) provide reasonably firm evidence that in the frog spinal cord afferent fibers from cutaneous nerves project more dorsally than afferent fibers from muscle nerves. In this sense, the differential projection of cutaneous and muscle afferents is similar to that seen in the cat, and probably reflects general princi- ples of organization common to all vertebrates (Jhavery and Frank 1983). It thus seems reasonable to assume that the responses produced in cutaneous or in mixed nerves by stimulation of the dorsal neuropil in the frog can be reliably attributed to antidromic responses of cutaneous afferents. Since these afferents had conduction velocities
between 17.7 and 21.4 m/s, they were most likely touch afferents (see Spray 1976).
Our results further indicate that stimulation of the brainstem RF produces PAD of low threshold cutaneous afferents. This PAD adds non-linearly with the PAD pro- duced in the same population of afferents by stimulation of an adjacent dorsal root or ventral root. Thus, it may be concluded, in agreement with our previous proposal (Gonz/tlez et al. 1992a) that reticulospinal fibers have a facilitatory action on the pathways that produce PAD of low threshold cutaneous fibers. In this regard, the action of reticulospinal fibers observed in the frog spinal cord is similar to that observed in the cat, where activation of reticulospinal, rubrospinal and raphespinal fibers pro- duces PAD of cutaneous afferents (Martin et al. 1979; Jim6nez et al. 1987). However, it must be stressed that, in the frog spinal cord, PAD of cutaneous fibers is produced not only by stimulation of dorsal roots or sensory nerves but also by stimulation of the ventral roots. The PAD produced by antidromic stimulation of motor axons seems to be produced by activation of at least two in- terneurons; one that is activated by cholinergic synapses from motor axon collaterals and another that has GABAergic synaptic contacts with afferent fibers (for dis- cussion, see Gonz/tlez et al. 1992a). In this sense they would be different from Renshaw interneurons in the cat spinal cord, which make GABAergic and glycinergic synapses with motoneurons, but have no action on affer- ent fibers (Cullheim and Kellerth 1981). The interaction between the PAD produced in cutaneous afferents by segmental and reticulospinal inputs documented in the present series of observations is best explained by assum- ing that the GABAergic interneurons that produce PAD receive convergent excitatory synaptic inputs from affer- ent fibers and motor axon collaterals and also from retic- ulospinal fibers, as proposed previously (see Fig. 10, Gon- zfilez et al. 1992a).
In this context it is interesting to note that, in the frog with the intact neuraxis, synaptic responses elicited by repetitive stimulation of cutaneous afferents decrement quite rapidly. Simpson (1976) proposed that this decre- ment results from descending inhibition, because it is not observed in the spinal preparation. It is possible that the descending inhibition is due, at least in part, to activity in reticulospinal pathways that produce a tonic PAD (and presynaptic inhibition) of the cutaneous afferent fibers.
Pertaining to the muscle afferents, our results indicate quite clearly that reticulospinal inputs produce a rather small or no PAD in fibers projecting to the ventral neu- ropil. Instead, they produce a powerful depression of the PAD elicited by dorsal root or ventral root stimulation. This depression can be ascribed to inhibition exerted on the interneurons that mediate the PAD rather than to occlusion, because the reticulospinal inputs alone pro- duce no PAD. Inhibition exerted on the input side can also be excluded in those cases in which the PAD has been produced by stimulation of the ventral roots, be- cause RF stimulation does not decrease impulse trans- mission at the intraspinal axonal branches of motoneu- rons (see Gonz/dez et al. 1992a).
Based on what has been observed in the cat spinal

269
cord (for review, see Rudomin 1990a, b), it is tempting to suggest that reticulospinal inputs inhibit the PAD elicit- ed in muscle spindle afferents and produce PAD in ten- don organ afferents (see also Gonzfilez et al. 1992a). If this were the case, it would be necessary to assume that the proport ion of muscle spindle afferents that were an- tidromically activated under the present experimental conditions exceeded by far the proport ion of antidromi- cally activated tendon organ afferents. Hence, the inhibi- tion of the PAD elicited in muscle spindle afferents would not be masked by the PAD elicited in tendon organ after- ents. Alternatively, it is possible that reticulospinal inputs inhibit the PAD elicited at the intraspinal terminals of the muscle spindle afferents, as well as in the terminals of the tendon organ afferents. If so, this would be at vari- ance with what has been observed in the cat (Rudomin et al. 1983, 1986; Jim6nez et al. 1988). At the present time, it is difficult to decide between these two possibilities be- cause although activity of tendon organs in response to stretch and muscle contraction has been studied in the sartorius muscle of the frog (Ito 1968), no studies appear to be available on the intraspinal distribution of muscle spindle and tendon organ afferents or of their relative proport ion in the various muscle nerves (see Simpson 1976 and Proske 1981). Another question which remains unsolved is whether or not antidromic responses pro- duced with intraspinal stimuli above 2-3 x T are also from muscle spindle and tendon organ afferents or if there is a group of muscle afferent fibers equivalent to that of group II muscle afferent fibers in the cat.
There has been considerable discussion of the relative importance of cutaneous versus muscle spindle afferents in the generation of spinal reflexes in the frog. Stretch reflexes in the frog hindlimb muscles appear to be very weak, or absent, whereas stimulation of the skin - even of relatively small regions produces strong reflex activa- tion of flexor muscles. This has led to the view that cuta- neous afferents are significantly more effective than mus- cle spindle afferents in producing reflex activation ofmo- toneurons (for review see Simpson 1976). The present series of investigations provides direct evidence that acti- vation of reticulospinal fibers has opposed actions on the PAD (and presumably also on the synaptic effectiveness) of low threshold cutaneous and muscle spindle afferents. The differential action of reticulospinal fibers on the pathways mediating the PAD of cutaneous and muscle afferents could play an important role in motor control by setting the balance between information arising from skin receptors and from muscle spindle afferents required for the execution of specific motor tasks. Reticulospinal fibers also have monosynaptic connections with mo- toneurons. Following their activation (for example dur- ing jumping), there would be increased presynaptic inhi- bition of the inputs provided by cutaneous afferents, so they would have fewer possibilities to interfere with the descending activation of the motoneurons. At the same time, there would be increased inhibition of the PAD elicited in muscle afferents either by sensory or motor axon activation; i.e. information transmitted by muscle spindles concerning muscle length would be emphasized because of presynaptic disinhibition. Clearly, this is an
oversimplified view of the complex interactions that take place between sensory and descending inputs to the spinal cord during performance of specific motor tasks or during postural adjustments. However, it serves its pur- pose by stressing the existence, in lower vertebrates as well, of mechanisms of descending control of the synaptic efficacy of muscle and cutaneous afferents that may be relevant for the control of movement. In fact, recent evi- dence stresses the role of sensory input in setting the equilibrium point of the forces elicited in the hindlimb of the spinal frog by intraspinal microstimulation (Loeb et al. 1991), and it is not unlikely that the descending con- trol of the synaptic efficacy of afferent fibers plays a rele- vant part in this setting.
Acknowledgements. We would like to thank C. Le6n and A. Rivera for technical assistance, and J. Gonzitlez for computer program- ming. The work was partly supported by NIH grant NS 09196 and CONACyT grant 41739.
References
Baldissera F, Hultborn H, Illert M (1981) Integration in spinal neuronal systems. In: Brookhart JM, Mountcastle VB, Brooks VB, Geiger SR (eds) Handbook of physiology, sect 1. The ner- vous system, vol 2. The American Physiological Society, Bethes- da, Md., pp 509-595
Carpenter D, Rudomin P (1973) The organization of primary affer- ent depolarization in the isolated spinal cord of the frog. J Phys- iol (Lond) 229:471-493
Coombs JS, Eccles JC, Landgren S (1956) Spinal cord potentials generated by impulses in muscle and cutaneous afferent fibers. J Neurophysiol 19:452-467
Cullheim S, Kellerth J (1981) Two kinds of recurrent inhibition of the cat spinal alpha motoneurons as differentiated pharmaco- logically. J Physiol (Lond) 312:209-224
Davidoff RA, Hackman JC (1984) Spinal inhibition. In: Davidoff RA (ed) Handbook of the spinal cord, vols 2, 3, Anatomy and physiology. Dekker, New York, pp 385-459
Eccles JC, Magni F, Willis WD (1962) Depolarization of central terminals of group I afferent fibers from muscle. J Physiol (Lond) 160:62-93
Frank E, Westerfield M (1982) Synaptic organization of sensory and motor neurones innervating triceps brachii muscles in the bull- frog. J Physiol (Lond) 324:479-494
Galindo J, Rudomin P (1978) The effects of gallamine on field and dorsal root potentials produced by antidromic stimulation of motor fibers in the frog spinal cord. Exp Brain Res 32:135-150
Gaupp E (1897) Ecker and Wiedersheim's Anatomie des Frosches, 2. Aufl, Lehre vom Nervensystem. Vieweg, Braunschweig, p 548
Glusman S, Rudomin P (1974) Presynaptic modulation of synaptic effectiveness of afferent and ventrolateral tract fibers in the frog spinal cord. Exp Neurol 45:474-490
Gonzfilez H, Jim6nez I, Rudomin P (1992a) Bulbospinal inhibition of PAD elicited by stimulation of afferent and motor axons in the isolated frog spinal cord and brainstem. Exp Brain Res 88:106-116
Gonz~lez H, Jim6nez I, Rudomin P (1992b) Differential effects of reticulo-spinal stimulation on the intraspinal threshold of cuta- neous and muscle afferents in the isolated frog neuraxis. Soc Neurosci Abstr 18:514
Grinnell AD (1966) A study of the interaction between motoneu- rones in the frog spinal cord. J Physiol (Lond) 182:612-648
Holemans KC, Meij HS, Meyer BJ (1966) The existence of a monosynaptic reflex arc in the spinal cord of the frog. Exp Neurol 14:175-186

270
Hultborn H, Meunier S, Pierrot-Deseilligni E, Shindo M (1987) Changes in presynaptic inhibition of Ia fibers at the onset of voluntary contraction in man. J Physiol (Lond) 389:757-772
Ito F (1968) Functional properties of tendon receptors in the frog. Jpn J Physiol 18:576-589
Jhavery S, Frank E (1983) Central projections of the brachial nerve in bullfrog: muscle and cutaneous afferents project to different regions of the spinal cord. J Comp Neurol 221:304-312
Jim6nez I, Rudomin P, Solodkin M (1987) Mechanisms involved in the depolarization of cutaneous afferents produced by segmental and descending inputs in the cat spinal cord. Exp Brain Res 69:195-207
Jim6nez I, Rudomin P, Solodkin M (1988) PAD patterns of physio- logically identified afferent fibers from the medial gastrocnemius muscle. Exp Brain Res 71:643-657
Loeb EP, Gizter SF, Bizzi E, Mussa-Ivaldi FA (1991) Effects of dorsal root cut on forces evoked by spinal stimulation in spinal- ized frogs. Soc Neurosci Abstr 17:1576
Martin RF, Haber LH, Willis WD (1979) Primary afferent depolar- ization of identified cutaneous fibers following stimulation in medial brain stem. J Neurophysiol 42:779-790
Proske U (1981) The Golgi tendon organ: properties of the receptor and reflex action of impulses arising from tendon organs. Int Rev Physiol 25:127-171
Renshaw B (1946) Central effects of centripetal impulses in axons of spinal ventral roots. J Neurophysiol 9:191-204
Rudomin P (1990a) Presynaptic control of synaptic effectiveness of muscle spindle and tendon organ afferents in the mammalian spinal cord. In: Binder MD, Mendell LM (eds) The segmental motor system. Oxford University Press, New York, pp 349-380
Rudomin P (1990b) Presynaptic inhibition of muscle spindle and tendon organ afferents in mammalian spinal cord. Trends Neu- rosci 13 : 499-505
Rudomin P, Jim6nez I, Solodkin M, Duefias S (1983) Sites of action of segmental and descending control of transmission on path- ways mediating PAD of Ia and Ib afferent fibers in the cat spinal cord. J Neurophysiol 50:743-769
Rudomin P, Solodkin M, Jim~nez I (1986) PAD and PAH response patterns of group Ia and Ib fibers to cutaneous and descending inputs in the cat spinal cord. J Neurophysiol 56:987-1006
Shapovalov AI, Shiriaev BI (1980) Recurrent interactions between individual motoneurones and dorsal root fibres in the frog. Exp Brain Res 38:115-116
Simpson JI (1976) Functional synaptology of the spinal cord. In: Llinas R, Precht W (eds) Frog neurobiology. Springer, Berlin, pp 728-749
Spray DC (1976) Pain and temperature receptors in anurans. In: Llinas R, Precht W (eds) Frog neurobiology. Springer, Berlin, pp 607-628
Stehouwer D, Farel P (1981) Sensory interactions with a central motor program in anuran larvae. Brain Res 218:131-140
Szekely G (1976) The morphology of motoneurons and dorsal root fibers in the frog spinal cord. Brain Res 103:275-290
Szekely G, Antal M (1984) Segregation of muscle and cutaneous afferent fiber terminals in the brachial spinal cord of the frog. J Hirnforsch 25:671-675
Szekely G, Matersz R, Baker R, Antal M (1982) The termination of cutaneous nerve in dorsal horn of the spinal cord in normal and skin rotated frogs. Exp Brain Res 45:19-28
Wall P (1958) Excitability changes in afferent fibre terminations and their relation to slow potentials. J Physiol (Lond) 142:1-21
Related Documents











