1995, 69(12):7888. J. Virol. M S Parcells, A S Anderson and T W Morgan genes. virus mutant lacking six unique short region Retention of oncogenicity by a Marek's disease http://jvi.asm.org/content/69/12/7888 Updated information and services can be found at: These include: CONTENT ALERTS more» cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new articles http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on February 5, 2014 by guest http://jvi.asm.org/ Downloaded from on February 5, 2014 by guest http://jvi.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1995, 69(12):7888. J. Virol.
M S Parcells, A S Anderson and T W Morgan genes.virus mutant lacking six unique short region Retention of oncogenicity by a Marek's disease
http://jvi.asm.org/content/69/12/7888Updated information and services can be found at:
These include:
CONTENT ALERTS more»cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new articles
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

JOURNAL OF VIROLOGY, Dec. 1995, p. 7888–7898 Vol. 69, No. 120022-538X/95/$04.0010Copyright q 1995, American Society for Microbiology
Retention of Oncogenicity by a Marek’s Disease Virus MutantLacking Six Unique Short Region Genes†MARK S. PARCELLS, AMY S. ANDERSON, AND ROBIN W. MORGAN*
Department of Animal Science and Agricultural Biochemistry, College of Agricultural Sciences,University of Delaware, Newark, Delaware 19717-1303
Received 25 July 1995/Accepted 21 September 1995
We previously reported the construction of Marek’s disease virus (MDV) strains having mutations in variousgenes that map to the unique short (US) region of the viral genome (J. L. Cantello, A. S. Anderson, A.Francesconi, and R. W. Morgan, J. Virol. 65:1584–1588, 1991; M. S. Parcells, A. S. Anderson, and R. W.Morgan, Virus Genes 9:5–13, 1994; M. S. Parcells, A. S. Anderson, and R. W. Morgan, J. Virol. 68:8239–8253,1994). These strains were constructed by using a high-passage-level serotype 1 MDV strain which grew well inchicken embryo fibroblasts. Despite the growth of the parent and mutant viruses in cell culture, in vivo studieswere limited by poor growth of these strains in chickens. One of the mutants studied lacked 4.5 kbp of USregion DNA and contained the lacZ gene of Escherichia coli inserted at the site of the deletion. The deletionremoved MDV homologs to the US1, US2, and US10 genes of herpes simplex virus type 1 as well as threeMDV-specific open reading frames. We now report the construction of a mutant MDV containing a similardeletion in the US region of the highly oncogenic RB1B strain. This mutant, RB1BD4.5lac, had a growthimpairment in established chicken embryo fibroblasts similar to that described previously for MDVs lackinga functional US1 gene. In chickens, RB1BD4.5lac showed decreased early cytolytic infection, mortality, tumorincidence, and horizontal transmission. Several lymphoblastoid cell lines were established from RB1BD4.5lac-induced tumors, and virus reactivated from these cell lines was LacZ1. These results indicate that the deletedgenes are nonessential for the transformation of chicken T cells or for the establishment and maintenance oflatency. On the basis of the growth impairment observed for RB1BD4.5lac in cell culture and in vivo, weconclude that deletion of these genes affects the lytic replication of MDV. This is the first MDV mutantconstructed in the RB1B oncogenic strain, and the methodology described herein provides for the directexamination of MDV-encoded determinants of oncogenicity.
Marek’s disease virus (MDV) is a transforming herpesvirusthat rapidly induces T-cell lymphomas in susceptible chickens(7). Although it is classified as a gammaherpesvirus on thebasis of its biological properties, the genomic organization andavailable DNA sequence data for MDV indicate that it is moreclosely related to the alphaherpesviruses (5, 6, 32, 34, 35, 45).The mechanism of MDV-mediated transformation of T cellsremains unknown. Oncogenic MDV strains do not replicatewell in cell culture and become attenuated rapidly during serialpassage (10). Attenuation is accompanied by alterations in thesequence and expression of the MDV genome (3, 4, 13, 21, 41,44), loss of horizontal transmission (10), and a decreased abil-ity to infect lymphocytes in vivo and in vitro (39).Previous studies of MDV transformation have focused on
the identification of (i) regions of the MDV genome thatundergo alteration during attenuation, (ii) viral gene productsexpressed in MDV-induced tumors and tumor-derived lym-phoblastoid cell lines (LBCLs), (iii) viral gene products be-lieved to be expressed exclusively by serotype 1 MDV strains,and (iv) sites of MDV integration into the host genome. Theseapproaches have led to the identification of a number of MDVgenes and regions having possible significance for transforma-tion. The repeats (terminal and inverted, respectively) flankingthe unique long (UL) region of the MDV genome (TRL andIRL) encode a family of RNAs that become disrupted duringattenuation (3, 4). This region also contains the meq gene,
which encodes a leucine-zipper-containing protein expressedin LBCLs (18). Genes encoding phosphoproteins pp38 andpp24 initiate in these repeats (2, 9, 20, 46). These phospho-proteins are expressed in cells lytically infected with MDVserotype 1 strains as well as in LBCLs (9). cDNAs generatedfrom transcripts expressed from these regions in LBCLs en-code small open reading frames (15, 30, 31). Antisera gener-ated against two of these open reading frames have detected a14-kDa protein expressed in lytically infected chicken embryofibroblasts (CEF) as well as in an LBCL (15). Antiserum raisedagainst another open reading frame detected a 7-kDa proteinexpressed in lytically infected CEF and in LBCLs (31). Therepeats flanking the unique short (US) region (IRS and TRS)are also implicated in transformation. In particular, a cDNAexpressed from IRS and TRS encodes an open reading framethat could specify a 94-amino-acid protein (27). Antiserumraised against this open reading frame stained RB1B-infectedchicken kidney cells but not LBCLs. Finally, the examinationof MDV integration sites in chromosomes of LBCLs has iden-tified numerous sites of insertion, but none have been associ-ated with alterations in either cellular or viral gene expression(11). Despite the contributions of this body of research to thestudy of MDV, the role of MDV-encoded gene products intransformation remains unknown.We recently described the characterization of mutant atten-
uated MDVs containing insertions and deletions in the USregion (8, 28, 29). Of these mutants, only those lacking afunctional US1 gene have a discernible phenotype in cell cul-ture, namely, a growth impairment in established CEF (29).This growth impairment correlated with decreased plaque sizeand numbers of PFU obtained per plaque. Although MDV is
* Corresponding author. Phone: (302) 831-2524.† Published as Miscellaneous Paper No. 1565 of the Delaware Ag-
ricultural Experiment Station.
7888
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

an attractive system for the analysis of viral gene function inthe natural host, the usefulness of previously reported MDVmutants has been severely limited by their poor growth inchickens. This report describes the first mutant MDV con-structed in the highly oncogenic RB1B strain of MDV. Thismutant, RB1BD4.5lac, contains a deletion identical to that ofGAD4.8lac (29).
MATERIALS AND METHODS
Cells and viruses. MDV was propagated in secondary CEF with growth me-dium consisting of M199 medium (Life Technologies, Inc. [LT], Grand Island,N.Y.) supplemented with 3% calf serum (LT), 100 IU of penicillin G per ml, and100 mg of dihydrostreptomycin per ml (23). Passage level 7 of RB1B (RB1Bp7),a highly pathogenic strain of MDV (38), was obtained from Intervet Inc., Mills-boro, Del., and passed twice in CEF for the preparation of RB1B-infected cellDNA. This RB1Bp9 DNA stock was used for mutant construction.Plasmid constructions. Cloning was done by standard methods (37), and
restriction endonuclease digestions were carried out according to the recommen-dations of the suppliers. Plasmid pMD190 was used to generate the RB1Bdeletion mutant (RB1BD4.5lac) and has been described previously (29). Briefly,a 12.2-kbp HindIII-BamHI subfragment of the BamHI-A fragment from the GAstrain of MDV (12) was cloned into pUC18 to generate pMD163. PlasmidpMD163 was digested with PstI (LT), and the 10.1-kbp vector-containing frag-ment was purified by preparative agarose gel electrophoresis and religated togenerate pMD164. Plasmid pMD164 DNA was digested with PstI and bluntended by using the Klenow enzyme (LT). A lacZ cassette, derived from pCH110(Pharmacia, Inc., Piscataway, N.J.) and modified for insertion mutagenesis (43),was blunt ended and ligated into pMD164 to generate pMD190. PlasmidpMD190 DNA was purified by using the Qiagen maxi-preparation procedure(Qiagen Corp., Chatsworth, Calif.).RB1BD4.5lac MDV strain construction. The RB1B deletion mutant was con-
structed by using a modified version of a method used to construct recombinantattenuated MDV strains (8) (Fig. 1). Ten micrograms of RB1B-infected CEFDNA was cotransfected with 0.5 mg of pMD190 DNA into freshly plated CEFwith calcium phosphate (23). One week posttransfection, monolayers were har-vested and diluted to 20 ml with complete medium, and 200 ml was added to eachwell of a 96-well microtiter dish. This dish was used as a template, in that 100 mlfrom each well was added to duplicate 96-well microtiter dishes that had beenseeded previously with 100 ml of CEF (;105 cells per ml). One week postplating,one microtiter dish was fixed with 95% ethanol (EtOH) and subjected to immu-nofluorescence analysis (IFA) with anti-b-galactosidase antiserum (5-Prime-3-Prime, Boulder, Colo.) followed by goat anti-rabbit immunoglobulin G fluores-cein isothiocyanate-conjugated antiserum (Sigma Chemical Co., St. Louis, Mo.).Wells from the unfixed microtiter dish corresponding to those containing fluo-rescent plaques on the fixed microtiter dish, as well as numerous surroundingwells, were harvested by the addition of 50 ml of 0.5% trypsin per well; this wasfollowed by the addition of 50 ml of calf serum (LT) per well. These wells wereplated, in duplicate, onto fresh CEF. After 6 days, one of each duplicate dish wasstained with Bluo-gal as previously described (8). After 10 additional rounds ofpicking and staining plaques, all detectable plaques were b-galactosidase posi-tive. When this procedure was used, one recombinant was plaque purified by atotal passage level of 24. This strain was designated RB1BD4.5lac. The RB1Bparent virus strain was also passaged in culture an equivalent number of times sothat any changes in oncogenicity and pathogenicity attributable merely to serialpassage in culture could be identified. This equal-passage-level parent was des-ignated RB1Bp24.Southern hybridizations. For Southern blot analysis, 10 mg of DNA from
MDV-infected or uninfected CEF was digested with EcoRI, and Southern blotswere prepared by standard methods (37). The MDV-specific probes used weresubfragments of the BamHI-A fragment of MDV strain GA (12) (Fig. 2A). ThelacZ-specific probe was a 2.1-kbp BglI fragment contained within the codingsequence of the lacZ gene (29). The probes were gel purified and biotin labelledwith the Random Images (United States Biochemical Corp., Cleveland, Ohio)nonisotopic random-primed DNA labelling kit.Northern (RNA) hybridization. Total RNA was purified by the guanidinium
isothiocyanate method for RNA purification followed by cesium chloride stepgradient centrifugation (1). RNA preparations were treated with RNase-freeRQ1 DNase (Promega Biotec, Madison, Wis.) and quantitated by A260. ForNorthern blot analysis, 10 mg of total RNA from each sample was separated ona 1.2% agarose-formaldehyde gel and transferred to nitrocellulose by standardmethods (1). For Northern blot analysis, probes were labelled with [a-32P]dCTPand the RadPrime random-primed DNA labelling kit (LT).Growth curves. Twelve identical 60-mm-diameter tissue culture dishes of CEF
were inoculated with RB1Bp24- or RB1BD4.5lac-infected CEF at approximately200 PFU per dish. At 1, 2, 3, 5, and 7 days postinoculation, monolayers wereharvested from duplicate dishes and inoculated onto fresh CEF. Titer disheswere counted after 6 days, and the mean number of plaques was determined.Titer dishes were fixed with 95% EtOH, and plaques were counted by IFA with
polyclonal chicken antiserum to MDV; this was followed by incubation withfluorescein isothiocyanate-conjugated rabbit anti-chicken immunoglobulin G an-tiserum (Sigma) as previously described (29). Growth curves were done withCEF monolayers incubated at either 37 or 418C, the latter being the bodytemperature of the chicken.Virus growth in vivo. One-day-old specific-pathogen-free single-comb White
Leghorn chickens were obtained from SPAFAS (Norwich, Conn.), wingbanded,randomly assigned to experimental groups, and housed in Montair Andersengloveport isolation cabinets. The chickens were inoculated intra-abdominallywith various doses of RB1Bp9-, RB1Bp24-, or RB1BD4.5lac-infected CEF orwith uninfected CEF. For experiment 1, 30 chickens were inoculated per treat-ment group. To evaluate early cytolytic infection, virus reisolations were at-tempted from spleen cells and peripheral blood leukocytes (PBLs) at 4, 7, and 11days postinoculation. At 14 days postinoculation, 10 additional one-day-oldchickens were added to each isolator to assess horizontal transmission of theviruses. Virus reisolations from spleen cells and PBLs of contact-exposed chick-ens were attempted at 14 days postplacement. For experiment 2, 25 chickenswere inoculated with uninfected CEF and 35 chickens were inoculated for theother treatment groups. Virus reisolations from the primary inoculates wereattempted at 7, 14, 21, and 28 days postinoculation. At 14 days postinoculation,10 additional one-day-old chickens were added to the control group and 15one-day-old chickens were added to the other treatment groups. Virus reisola-tions from contact-exposed chickens were attempted at 7 weeks postplacement.For each experiment, inoculates and contacts were observed daily for 9 and 7weeks, respectively, for signs of Marek’s disease. Tissue samples were taken forhistological analysis from representative chickens.Virus reisolations. For virus reisolations from spleen cells, spleens were re-
moved and pooled from three chickens per treatment group, rinsed in incom-plete medium consisting of M199 (LT) supplemented with 100 IU of penicillin Gper ml and 100 mg of dihydrostreptomycin per ml, and homogenized on ice witha Ten Broeck tissue grinder. Splenocytes were washed three times in incompletemedium and resuspended at concentrations of either 23 107 or 43 107 cells perml in complete medium (incomplete medium containing 3% heat-inactivated calf
FIG. 1. Diagram of steps in the construction of RB1BD4.5lac.
VOL. 69, 1995 MAREK’S DISEASE VIRUS MUTANT RETAINS ONCOGENICITY 7889
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
serum; LT). For virus reisolations, 1 3 106 or 2 3 106 viable cells were platedonto fresh CEF in triplicate, and plaques were counted 6 days later by IFA.For virus reisolations from PBLs, blood was drawn from three chickens per
treatment group into one heparinized syringe, adjusted to a volume of 8 ml withincomplete medium, and placed into a conical 15-ml polypropylene tube. PBLswere purified by underlaying 3 ml of Ficoll Hypaque 1119 (Sigma) and centri-fuging at 700 3 g for 30 min at room temperature. PBLs were removed, washedtwice with 10 ml of complete medium, and adjusted to a concentration of 23 107
cells per ml. For virus reisolations, 1 3 106 or 2 3 106 viable PBLs were platedonto fresh CEF in triplicate, and plaques were counted 6 days later by IFA.Cell line establishment. Tumors were removed from RB1Bp9- or RB1BD4.5lac-
infected chickens, washed with incomplete media, homogenized with a TenBroeck tissue grinder, and filtered through sterile cheesecloth. Lymphocyteswere purified from the tumor homogenate with Ficoll Hypaque 1119 (Sigma).Cells were washed and resuspended in LM Hahn medium (40) and plated in12-well dishes at concentrations ranging from 1 3 107 to 5 3 105 cells per ml.Cells were split 1:1 to 1:3 in LM Hahn medium approximately every 4 to 5 dayswhen a drop in the medium pH was apparent until the cell lines became estab-lished. The cells were split 1:5 to 1:10 thereafter as needed. Cell lines establishedfrom RB1BD4.5lac-induced tumors have been designated MDCC-UD01, -UD02
and -UD03 (UD01, UD02, and UD03, respectively) and were derived fromRB1BD4.5lac-induced ovarian, spleen, and liver tumors from separate chickens,respectively. A cell line derived from an RB1Bp9-induced kidney tumor wasdesignated MDCC-UD14 (UD14).Virus reactivation from MDV cell lines. Virus was recovered from lympho-
blastoid cell lines by cocultivation of 13 105 to 13 106 lymphoblastoid cells withsecondary CEF plated on 60-mm-diameter dishes. Seven days after cocultivation,the dishes were fixed with 95% EtOH and plaques were counted by IFA withrabbit anti-b-galactosidase polyclonal antibodies (5-Prime-3-Prime) or rabbitanti-MDV-US1 protein (graciously provided by Peter Brunovskis, Case WesternReserve University, and Leland Velicer, Michigan State University) as the pri-mary antiserum.
RESULTS
Construction of RB1BD4.5lac. After the initial IFA screen-ing of the EtOH-fixed microtiter dish, seven wells containedfluorescent plaques. We harvested the corresponding wells on
2.8 5.3E EEE 1.8
EE E 7.81.5
lacZ
E E2.6
E E2.6
pMD100pMD161
pMD193
P P
1 2 US1 US10 3 US2 US3 4 US6
C
A
C P M C P M C P M C P M
2.61.81.5
2.8
7.85.34.3
7.8
2.1
2.6 2.6
1.7
0.9
6.2
3.73.32.62.0
3.6
pMD193
pMD193
pMD161
pMD161 pMD100
lacZ
lacZ
B1.8 C P M R 2.8 C P M R 4.3 C P M R lac C P M R
pMD100
B
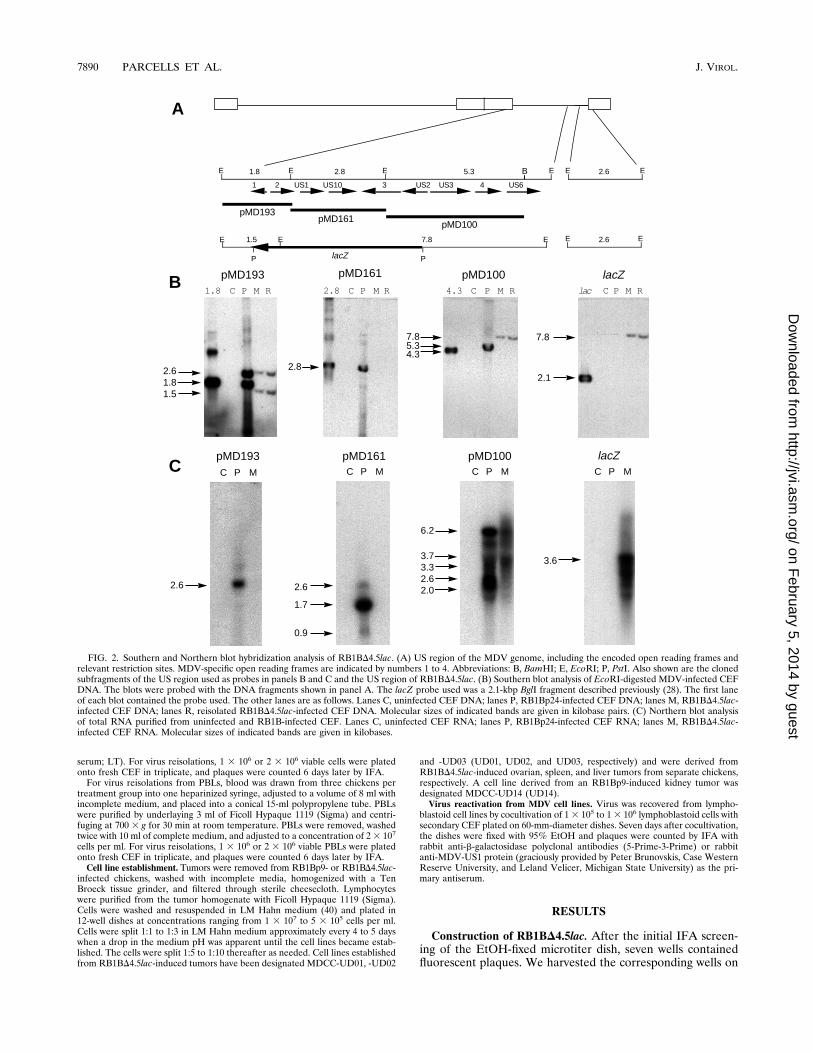
FIG. 2. Southern and Northern blot hybridization analysis of RB1BD4.5lac. (A) US region of the MDV genome, including the encoded open reading frames andrelevant restriction sites. MDV-specific open reading frames are indicated by numbers 1 to 4. Abbreviations: B, BamHI; E, EcoRI; P, PstI. Also shown are the clonedsubfragments of the US region used as probes in panels B and C and the US region of RB1BD4.5lac. (B) Southern blot analysis of EcoRI-digested MDV-infected CEFDNA. The blots were probed with the DNA fragments shown in panel A. The lacZ probe used was a 2.1-kbp BglI fragment described previously (28). The first laneof each blot contained the probe used. The other lanes are as follows. Lanes C, uninfected CEF DNA; lanes P, RB1Bp24-infected CEF DNA; lanes M, RB1BD4.5lac-infected CEF DNA; lanes R, reisolated RB1BD4.5lac-infected CEF DNA. Molecular sizes of indicated bands are given in kilobase pairs. (C) Northern blot analysisof total RNA purified from uninfected and RB1B-infected CEF. Lanes C, uninfected CEF RNA; lanes P, RB1Bp24-infected CEF RNA; lanes M, RB1BD4.5lac-infected CEF RNA. Molecular sizes of indicated bands are given in kilobases.
7890 PARCELLS ET AL. J. VIROL.
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
the unfixed sister dish and plated the contents onto duplicate60-mm-diameter dishes of CEF. Since MDV is cell associatedand trypsinization does not separate individual cells com-pletely, we worried that the distribution of infected cells acrossthe duplicate microtiter plates might not be equivalent. There-fore, we also harvested 14 surrounding wells and plated thecontents onto duplicate 60-mm-diameter dishes of CEF. All ofthe remaining wells were harvested and plated onto 24-welldishes of CEF. As we suspected, some recombinant MDVswere detected in cultures derived from wells that had notcorresponded to those wells containing fluorescent plaques onthe fixed plate. In any event, the presence of fluorescentplaques on the fixed microtiter dish allowed us to conclude thatrecombinants had been generated and that they were likely tobe present on the unfixed plate. One mutant, designatedRB1BD4.5lac, was obtained. The deletion contained in thismutant is identical to that of GAD4.8lac (29). The publishedDNA sequence of this region (5, 34, 35) indicates that thedeleted region is 4.5 kbp rather than the 4.8 kbp previouslyestimated from electrophoretic mobility.Analysis of the RB1BD4.5lac genome. Southern blot hybrid-
ization of EcoRI-digested DNA from uninfected CEF or CEFinfected with RB1Bp24, RB1BD4.5lac, or reisolated RB1BD4.5lac indicated that insertion of the lacZ cassette at the site ofthe 4.5-kbp deletion occurred as expected and that the recom-binant virus stock was free of detectable parent virus (Fig. 2).In addition, hybridization patterns were essentially identical tothose previously reported for the GAD4.8lac mutant (29). A1.8-kbp DNA probe (pMD193) detected two bands in MDV-infected cell DNA of 2.6 and 1.8 kbp in the case of RB1Bp24and 2.6 and 1.5 kbp in the case of the mutant virus DNAs (Fig.2B). The 2.6-kbp band was shown previously to encode theUS-TRS junction fragment which bears some sequence identityto the 1.8-kbp IRS-US junction fragment (29). The 1.5-kbpfragment results from the deletion of a portion of this region inthe mutant genome and the insertion of an EcoRI site which ispresent in the lacZ cassette. The adjacent 2.8-kbp probe(pMD161) detected a 2.8-kbp fragment in the parent virusDNA which was deleted from the mutant (Fig. 2B). The down-stream 4.3-kbp EcoRI-BamHI probe (pMD100) detected a5.3-kbp fragment in the parent and a 7.8-kbp fragment in themutant virus DNAs (Fig. 2B). This 7.8-kbp band results fromthe deletion of two EcoRI sites in the mutant and the insertionof one EcoRI site present in the lacZ cassette. A lacZ-specificprobe detected the 7.8-kbp band specific for the mutant virusDNAs (Fig. 2B).Mutagenesis-associated changes in MDV gene expression.
To examine changes in MDV gene expression associated withmutagenesis, Northern blot hybridization with total RNA pu-rified from uninfected and RB1Bp24- and RB1BD4.5lac-in-fected CEF was done (Fig. 2C). The probes used were identicalto those used for Southern hybridization (Fig. 2A), and theNorthern hybridization pattern was like that previously re-ported for the GAD4.8lac mutant (29). Probe pMD193 de-tected a 2.6-kb transcript in RB1Bp24-infected but notRB1BD4.5lac-infected CEF RNA (Fig. 2C). The adjacentprobe, pMD161, detected the 2.6- as well as the 1.7- and 0.9-kbtranscripts in RB1Bp24- but not RB1BD4.5lac-infected CEFRNA (Fig. 2C). The pMD100 probe detected several tran-scripts present in both RB1Bp24- and RB1BD4.5lac-infectedCEF RNA (Fig. 2C), indicating that the region downstream ofthe deletion was actively transcribed. The lacZ-specific probedetected an abundant 3.6-kb transcript in RB1BD4.5lac- butnot RB1Bp24-infected CEF RNA. These data indicated thatthe deletion dramatically affected MDV transcription in theregion. In addition, the pattern of immunofluorescence stain-
ing of RB1Bp24 and RB1BD4.5lac with a rabbit anti-US1 poly-clonal antiserum (provided by Peter Brunovskis, Case WesternReserve University, and Leland Velicer, Michigan State Uni-versity) was identical to that previously reported for GAatt85and GAD4.8lac, respectively (data not shown) (29). These re-sults further demonstrated that the recombinant virus stockwas free of detectable parent virus.RB1BD4.5lac growth in cell culture. RB1BD4.5lac showed a
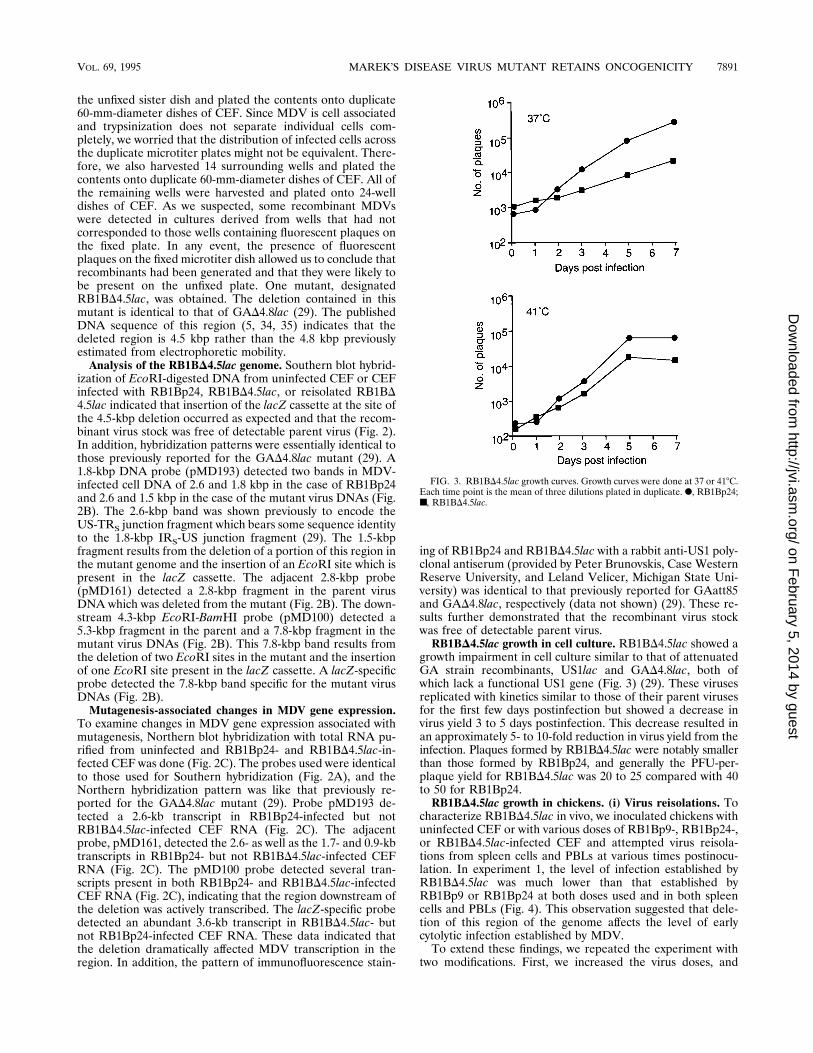
growth impairment in cell culture similar to that of attenuatedGA strain recombinants, US1lac and GAD4.8lac, both ofwhich lack a functional US1 gene (Fig. 3) (29). These virusesreplicated with kinetics similar to those of their parent virusesfor the first few days postinfection but showed a decrease invirus yield 3 to 5 days postinfection. This decrease resulted inan approximately 5- to 10-fold reduction in virus yield from theinfection. Plaques formed by RB1BD4.5lac were notably smallerthan those formed by RB1Bp24, and generally the PFU-per-plaque yield for RB1BD4.5lac was 20 to 25 compared with 40to 50 for RB1Bp24.RB1BD4.5lac growth in chickens. (i) Virus reisolations. To
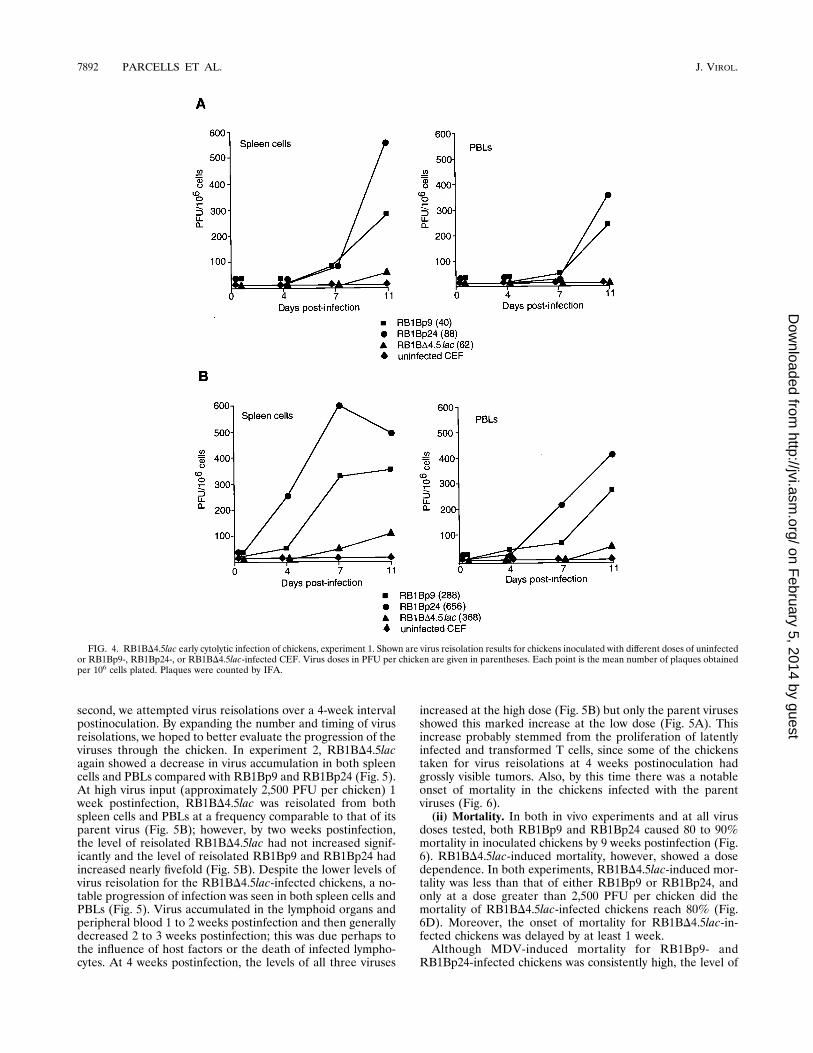
characterize RB1BD4.5lac in vivo, we inoculated chickens withuninfected CEF or with various doses of RB1Bp9-, RB1Bp24-,or RB1BD4.5lac-infected CEF and attempted virus reisola-tions from spleen cells and PBLs at various times postinocu-lation. In experiment 1, the level of infection established byRB1BD4.5lac was much lower than that established byRB1Bp9 or RB1Bp24 at both doses used and in both spleencells and PBLs (Fig. 4). This observation suggested that dele-tion of this region of the genome affects the level of earlycytolytic infection established by MDV.To extend these findings, we repeated the experiment with
two modifications. First, we increased the virus doses, and
FIG. 3. RB1BD4.5lac growth curves. Growth curves were done at 37 or 418C.Each time point is the mean of three dilutions plated in duplicate. F, RB1Bp24;■, RB1BD4.5lac.
VOL. 69, 1995 MAREK’S DISEASE VIRUS MUTANT RETAINS ONCOGENICITY 7891
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
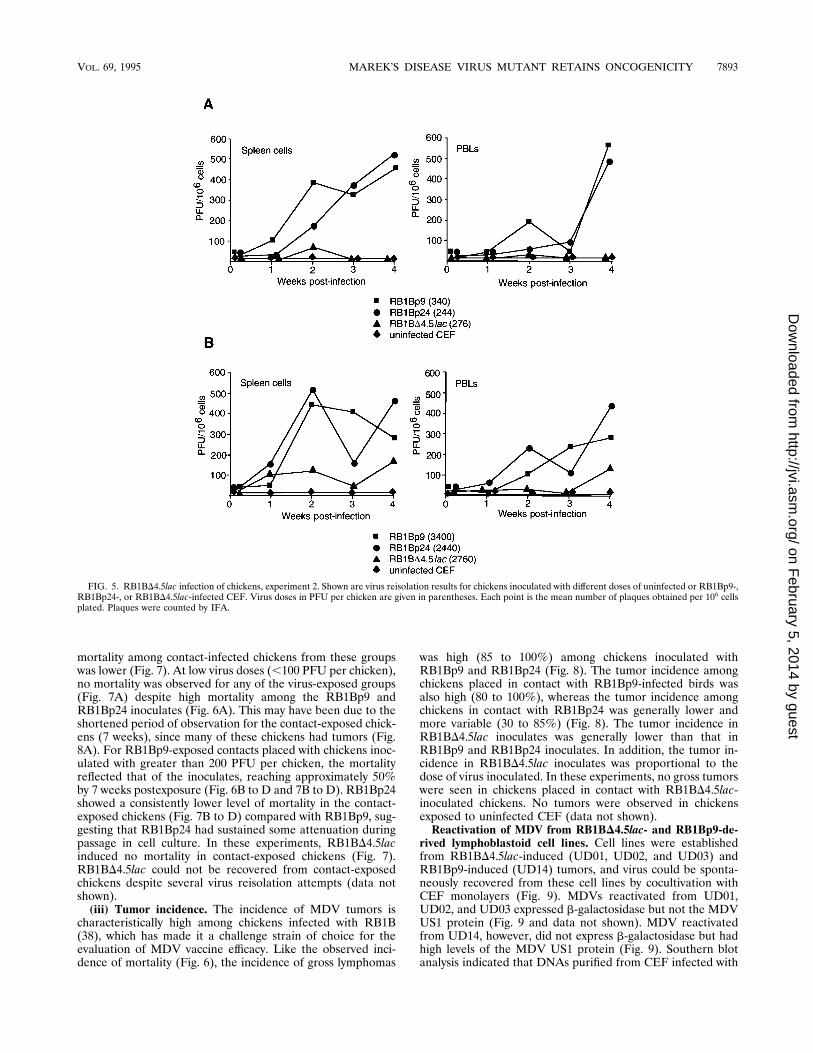
second, we attempted virus reisolations over a 4-week intervalpostinoculation. By expanding the number and timing of virusreisolations, we hoped to better evaluate the progression of theviruses through the chicken. In experiment 2, RB1BD4.5lacagain showed a decrease in virus accumulation in both spleencells and PBLs compared with RB1Bp9 and RB1Bp24 (Fig. 5).At high virus input (approximately 2,500 PFU per chicken) 1week postinfection, RB1BD4.5lac was reisolated from bothspleen cells and PBLs at a frequency comparable to that of itsparent virus (Fig. 5B); however, by two weeks postinfection,the level of reisolated RB1BD4.5lac had not increased signif-icantly and the level of reisolated RB1Bp9 and RB1Bp24 hadincreased nearly fivefold (Fig. 5B). Despite the lower levels ofvirus reisolation for the RB1BD4.5lac-infected chickens, a no-table progression of infection was seen in both spleen cells andPBLs (Fig. 5). Virus accumulated in the lymphoid organs andperipheral blood 1 to 2 weeks postinfection and then generallydecreased 2 to 3 weeks postinfection; this was due perhaps tothe influence of host factors or the death of infected lympho-cytes. At 4 weeks postinfection, the levels of all three viruses
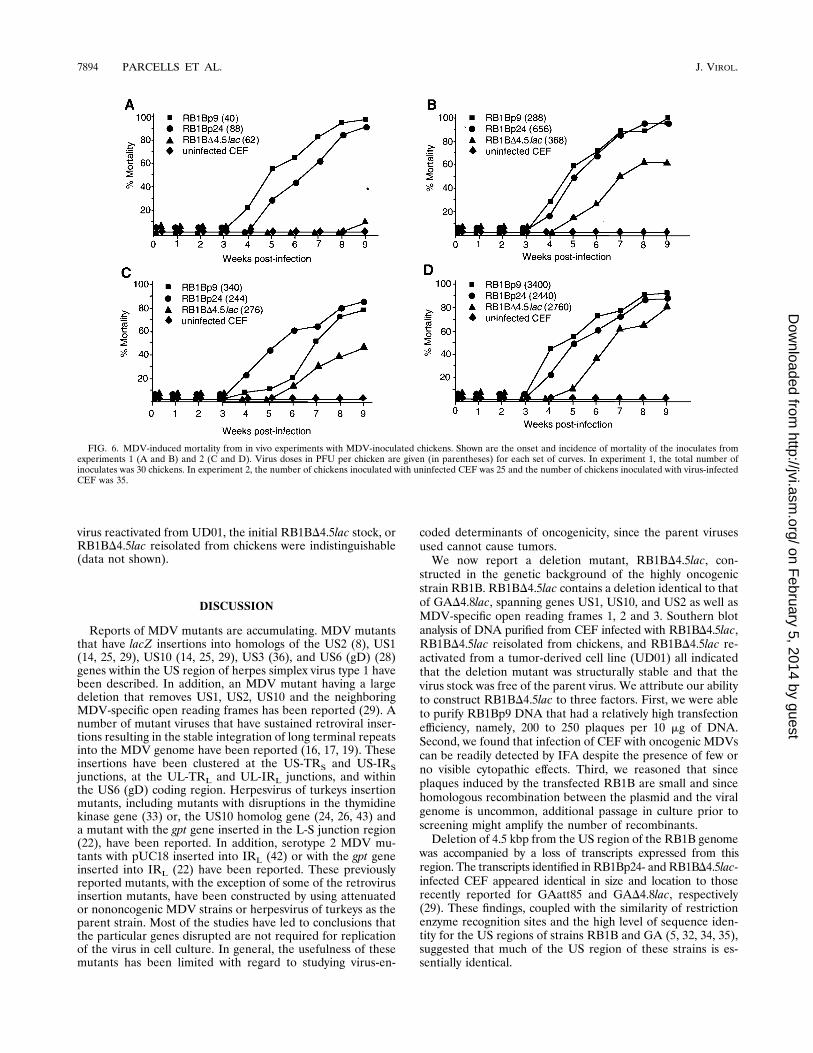
increased at the high dose (Fig. 5B) but only the parent virusesshowed this marked increase at the low dose (Fig. 5A). Thisincrease probably stemmed from the proliferation of latentlyinfected and transformed T cells, since some of the chickenstaken for virus reisolations at 4 weeks postinoculation hadgrossly visible tumors. Also, by this time there was a notableonset of mortality in the chickens infected with the parentviruses (Fig. 6).(ii) Mortality. In both in vivo experiments and at all virus
doses tested, both RB1Bp9 and RB1Bp24 caused 80 to 90%mortality in inoculated chickens by 9 weeks postinfection (Fig.6). RB1BD4.5lac-induced mortality, however, showed a dosedependence. In both experiments, RB1BD4.5lac-induced mor-tality was less than that of either RB1Bp9 or RB1Bp24, andonly at a dose greater than 2,500 PFU per chicken did themortality of RB1BD4.5lac-infected chickens reach 80% (Fig.6D). Moreover, the onset of mortality for RB1BD4.5lac-in-fected chickens was delayed by at least 1 week.Although MDV-induced mortality for RB1Bp9- and
RB1Bp24-infected chickens was consistently high, the level of
FIG. 4. RB1BD4.5lac early cytolytic infection of chickens, experiment 1. Shown are virus reisolation results for chickens inoculated with different doses of uninfectedor RB1Bp9-, RB1Bp24-, or RB1BD4.5lac-infected CEF. Virus doses in PFU per chicken are given in parentheses. Each point is the mean number of plaques obtainedper 106 cells plated. Plaques were counted by IFA.
7892 PARCELLS ET AL. J. VIROL.
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
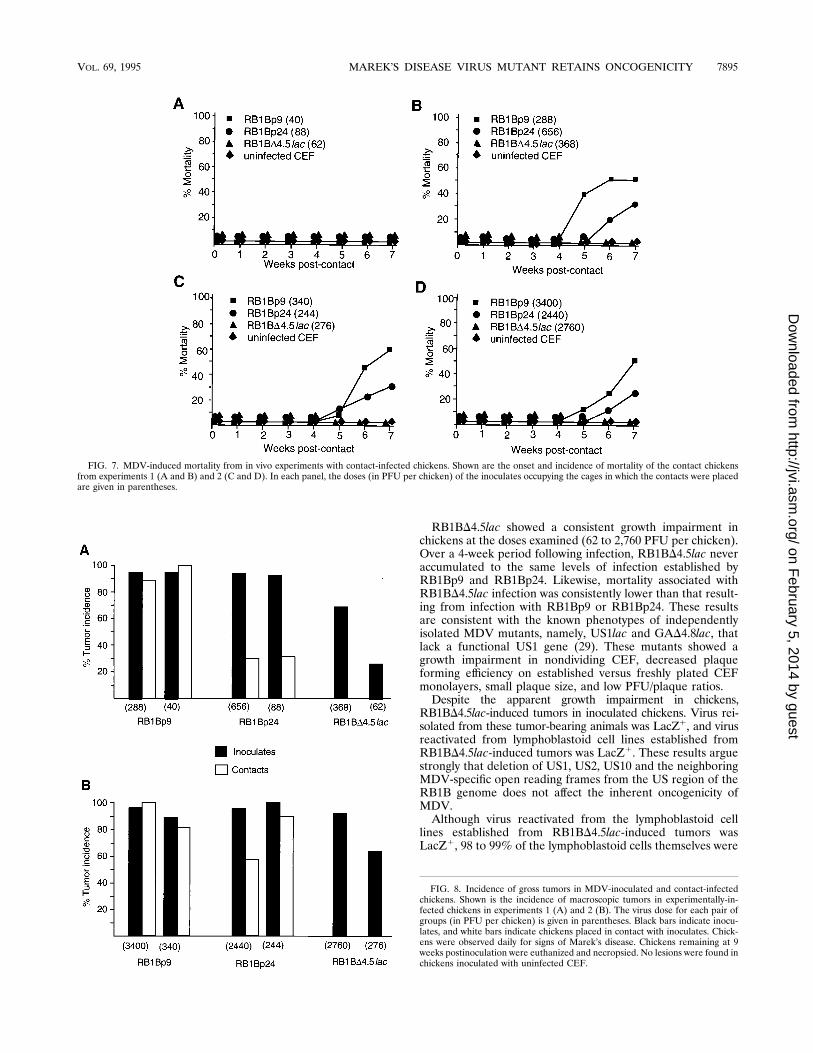
mortality among contact-infected chickens from these groupswas lower (Fig. 7). At low virus doses (,100 PFU per chicken),no mortality was observed for any of the virus-exposed groups(Fig. 7A) despite high mortality among the RB1Bp9 andRB1Bp24 inoculates (Fig. 6A). This may have been due to theshortened period of observation for the contact-exposed chick-ens (7 weeks), since many of these chickens had tumors (Fig.8A). For RB1Bp9-exposed contacts placed with chickens inoc-ulated with greater than 200 PFU per chicken, the mortalityreflected that of the inoculates, reaching approximately 50%by 7 weeks postexposure (Fig. 6B to D and 7B to D). RB1Bp24showed a consistently lower level of mortality in the contact-exposed chickens (Fig. 7B to D) compared with RB1Bp9, sug-gesting that RB1Bp24 had sustained some attenuation duringpassage in cell culture. In these experiments, RB1BD4.5lacinduced no mortality in contact-exposed chickens (Fig. 7).RB1BD4.5lac could not be recovered from contact-exposedchickens despite several virus reisolation attempts (data notshown).(iii) Tumor incidence. The incidence of MDV tumors is
characteristically high among chickens infected with RB1B(38), which has made it a challenge strain of choice for theevaluation of MDV vaccine efficacy. Like the observed inci-dence of mortality (Fig. 6), the incidence of gross lymphomas
was high (85 to 100%) among chickens inoculated withRB1Bp9 and RB1Bp24 (Fig. 8). The tumor incidence amongchickens placed in contact with RB1Bp9-infected birds wasalso high (80 to 100%), whereas the tumor incidence amongchickens in contact with RB1Bp24 was generally lower andmore variable (30 to 85%) (Fig. 8). The tumor incidence inRB1BD4.5lac inoculates was generally lower than that inRB1Bp9 and RB1Bp24 inoculates. In addition, the tumor in-cidence in RB1BD4.5lac inoculates was proportional to thedose of virus inoculated. In these experiments, no gross tumorswere seen in chickens placed in contact with RB1BD4.5lac-inoculated chickens. No tumors were observed in chickensexposed to uninfected CEF (data not shown).Reactivation of MDV from RB1BD4.5lac- and RB1Bp9-de-
rived lymphoblastoid cell lines. Cell lines were establishedfrom RB1BD4.5lac-induced (UD01, UD02, and UD03) andRB1Bp9-induced (UD14) tumors, and virus could be sponta-neously recovered from these cell lines by cocultivation withCEF monolayers (Fig. 9). MDVs reactivated from UD01,UD02, and UD03 expressed b-galactosidase but not the MDVUS1 protein (Fig. 9 and data not shown). MDV reactivatedfrom UD14, however, did not express b-galactosidase but hadhigh levels of the MDV US1 protein (Fig. 9). Southern blotanalysis indicated that DNAs purified from CEF infected with
FIG. 5. RB1BD4.5lac infection of chickens, experiment 2. Shown are virus reisolation results for chickens inoculated with different doses of uninfected or RB1Bp9-,RB1Bp24-, or RB1BD4.5lac-infected CEF. Virus doses in PFU per chicken are given in parentheses. Each point is the mean number of plaques obtained per 106 cellsplated. Plaques were counted by IFA.
VOL. 69, 1995 MAREK’S DISEASE VIRUS MUTANT RETAINS ONCOGENICITY 7893
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
virus reactivated from UD01, the initial RB1BD4.5lac stock, orRB1BD4.5lac reisolated from chickens were indistinguishable(data not shown).
DISCUSSION
Reports of MDV mutants are accumulating. MDV mutantsthat have lacZ insertions into homologs of the US2 (8), US1(14, 25, 29), US10 (14, 25, 29), US3 (36), and US6 (gD) (28)genes within the US region of herpes simplex virus type 1 havebeen described. In addition, an MDV mutant having a largedeletion that removes US1, US2, US10 and the neighboringMDV-specific open reading frames has been reported (29). Anumber of mutant viruses that have sustained retroviral inser-tions resulting in the stable integration of long terminal repeatsinto the MDV genome have been reported (16, 17, 19). Theseinsertions have been clustered at the US-TRS and US-IRSjunctions, at the UL-TRL and UL-IRL junctions, and withinthe US6 (gD) coding region. Herpesvirus of turkeys insertionmutants, including mutants with disruptions in the thymidinekinase gene (33) or, the US10 homolog gene (24, 26, 43) anda mutant with the gpt gene inserted in the L-S junction region(22), have been reported. In addition, serotype 2 MDV mu-tants with pUC18 inserted into IRL (42) or with the gpt geneinserted into IRL (22) have been reported. These previouslyreported mutants, with the exception of some of the retrovirusinsertion mutants, have been constructed by using attenuatedor nononcogenic MDV strains or herpesvirus of turkeys as theparent strain. Most of the studies have led to conclusions thatthe particular genes disrupted are not required for replicationof the virus in cell culture. In general, the usefulness of thesemutants has been limited with regard to studying virus-en-
coded determinants of oncogenicity, since the parent virusesused cannot cause tumors.We now report a deletion mutant, RB1BD4.5lac, con-
structed in the genetic background of the highly oncogenicstrain RB1B. RB1BD4.5lac contains a deletion identical to thatof GAD4.8lac, spanning genes US1, US10, and US2 as well asMDV-specific open reading frames 1, 2 and 3. Southern blotanalysis of DNA purified from CEF infected with RB1BD4.5lac,RB1BD4.5lac reisolated from chickens, and RB1BD4.5lac re-activated from a tumor-derived cell line (UD01) all indicatedthat the deletion mutant was structurally stable and that thevirus stock was free of the parent virus. We attribute our abilityto construct RB1BD4.5lac to three factors. First, we were ableto purify RB1Bp9 DNA that had a relatively high transfectionefficiency, namely, 200 to 250 plaques per 10 mg of DNA.Second, we found that infection of CEF with oncogenic MDVscan be readily detected by IFA despite the presence of few orno visible cytopathic effects. Third, we reasoned that sinceplaques induced by the transfected RB1B are small and sincehomologous recombination between the plasmid and the viralgenome is uncommon, additional passage in culture prior toscreening might amplify the number of recombinants.Deletion of 4.5 kbp from the US region of the RB1B genome
was accompanied by a loss of transcripts expressed from thisregion. The transcripts identified in RB1Bp24- andRB1BD4.5lac-infected CEF appeared identical in size and location to thoserecently reported for GAatt85 and GAD4.8lac, respectively(29). These findings, coupled with the similarity of restrictionenzyme recognition sites and the high level of sequence iden-tity for the US regions of strains RB1B and GA (5, 32, 34, 35),suggested that much of the US region of these strains is es-sentially identical.
FIG. 6. MDV-induced mortality from in vivo experiments with MDV-inoculated chickens. Shown are the onset and incidence of mortality of the inoculates fromexperiments 1 (A and B) and 2 (C and D). Virus doses in PFU per chicken are given (in parentheses) for each set of curves. In experiment 1, the total number ofinoculates was 30 chickens. In experiment 2, the number of chickens inoculated with uninfected CEF was 25 and the number of chickens inoculated with virus-infectedCEF was 35.
7894 PARCELLS ET AL. J. VIROL.
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
RB1BD4.5lac showed a consistent growth impairment inchickens at the doses examined (62 to 2,760 PFU per chicken).Over a 4-week period following infection, RB1BD4.5lac neveraccumulated to the same levels of infection established byRB1Bp9 and RB1Bp24. Likewise, mortality associated withRB1BD4.5lac infection was consistently lower than that result-ing from infection with RB1Bp9 or RB1Bp24. These resultsare consistent with the known phenotypes of independentlyisolated MDV mutants, namely, US1lac and GAD4.8lac, thatlack a functional US1 gene (29). These mutants showed agrowth impairment in nondividing CEF, decreased plaqueforming efficiency on established versus freshly plated CEFmonolayers, small plaque size, and low PFU/plaque ratios.Despite the apparent growth impairment in chickens,
RB1BD4.5lac-induced tumors in inoculated chickens. Virus rei-solated from these tumor-bearing animals was LacZ1, and virusreactivated from lymphoblastoid cell lines established fromRB1BD4.5lac-induced tumors was LacZ1. These results arguestrongly that deletion of US1, US2, US10 and the neighboringMDV-specific open reading frames from the US region of theRB1B genome does not affect the inherent oncogenicity ofMDV.Although virus reactivated from the lymphoblastoid cell
lines established from RB1BD4.5lac-induced tumors wasLacZ1, 98 to 99% of the lymphoblastoid cells themselves were
FIG. 7. MDV-induced mortality from in vivo experiments with contact-infected chickens. Shown are the onset and incidence of mortality of the contact chickensfrom experiments 1 (A and B) and 2 (C and D). In each panel, the doses (in PFU per chicken) of the inoculates occupying the cages in which the contacts were placedare given in parentheses.
FIG. 8. Incidence of gross tumors in MDV-inoculated and contact-infectedchickens. Shown is the incidence of macroscopic tumors in experimentally-in-fected chickens in experiments 1 (A) and 2 (B). The virus dose for each pair ofgroups (in PFU per chicken) is given in parentheses. Black bars indicate inocu-lates, and white bars indicate chickens placed in contact with inoculates. Chick-ens were observed daily for signs of Marek’s disease. Chickens remaining at 9weeks postinoculation were euthanized and necropsied. No lesions were found inchickens inoculated with uninfected CEF.
VOL. 69, 1995 MAREK’S DISEASE VIRUS MUTANT RETAINS ONCOGENICITY 7895
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
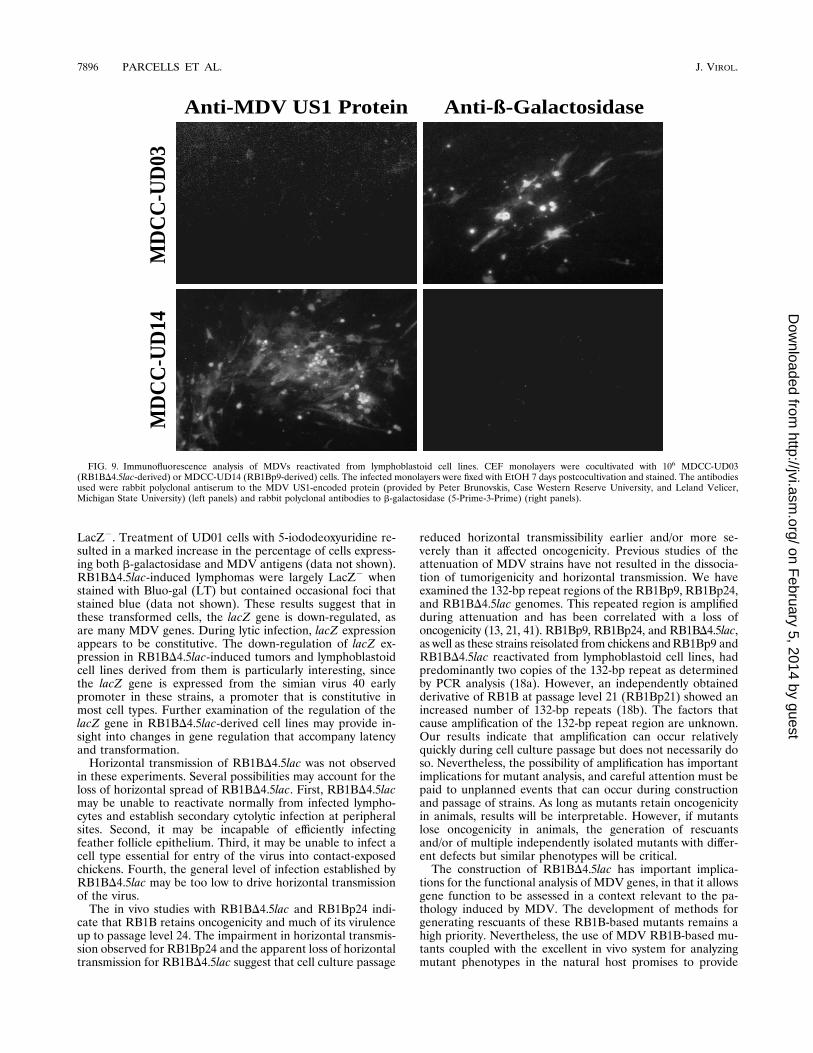
LacZ2. Treatment of UD01 cells with 5-iododeoxyuridine re-sulted in a marked increase in the percentage of cells express-ing both b-galactosidase and MDV antigens (data not shown).RB1BD4.5lac-induced lymphomas were largely LacZ2 whenstained with Bluo-gal (LT) but contained occasional foci thatstained blue (data not shown). These results suggest that inthese transformed cells, the lacZ gene is down-regulated, asare many MDV genes. During lytic infection, lacZ expressionappears to be constitutive. The down-regulation of lacZ ex-pression in RB1BD4.5lac-induced tumors and lymphoblastoidcell lines derived from them is particularly interesting, sincethe lacZ gene is expressed from the simian virus 40 earlypromoter in these strains, a promoter that is constitutive inmost cell types. Further examination of the regulation of thelacZ gene in RB1BD4.5lac-derived cell lines may provide in-sight into changes in gene regulation that accompany latencyand transformation.Horizontal transmission of RB1BD4.5lac was not observed
in these experiments. Several possibilities may account for theloss of horizontal spread of RB1BD4.5lac. First, RB1BD4.5lacmay be unable to reactivate normally from infected lympho-cytes and establish secondary cytolytic infection at peripheralsites. Second, it may be incapable of efficiently infectingfeather follicle epithelium. Third, it may be unable to infect acell type essential for entry of the virus into contact-exposedchickens. Fourth, the general level of infection established byRB1BD4.5lac may be too low to drive horizontal transmissionof the virus.The in vivo studies with RB1BD4.5lac and RB1Bp24 indi-
cate that RB1B retains oncogenicity and much of its virulenceup to passage level 24. The impairment in horizontal transmis-sion observed for RB1Bp24 and the apparent loss of horizontaltransmission for RB1BD4.5lac suggest that cell culture passage
reduced horizontal transmissibility earlier and/or more se-verely than it affected oncogenicity. Previous studies of theattenuation of MDV strains have not resulted in the dissocia-tion of tumorigenicity and horizontal transmission. We haveexamined the 132-bp repeat regions of the RB1Bp9, RB1Bp24,and RB1BD4.5lac genomes. This repeated region is amplifiedduring attenuation and has been correlated with a loss ofoncogenicity (13, 21, 41). RB1Bp9, RB1Bp24, and RB1BD4.5lac,as well as these strains reisolated from chickens andRB1Bp9 andRB1BD4.5lac reactivated from lymphoblastoid cell lines, hadpredominantly two copies of the 132-bp repeat as determinedby PCR analysis (18a). However, an independently obtainedderivative of RB1B at passage level 21 (RB1Bp21) showed anincreased number of 132-bp repeats (18b). The factors thatcause amplification of the 132-bp repeat region are unknown.Our results indicate that amplification can occur relativelyquickly during cell culture passage but does not necessarily doso. Nevertheless, the possibility of amplification has importantimplications for mutant analysis, and careful attention must bepaid to unplanned events that can occur during constructionand passage of strains. As long as mutants retain oncogenicityin animals, results will be interpretable. However, if mutantslose oncogenicity in animals, the generation of rescuantsand/or of multiple independently isolated mutants with differ-ent defects but similar phenotypes will be critical.The construction of RB1BD4.5lac has important implica-
tions for the functional analysis of MDV genes, in that it allowsgene function to be assessed in a context relevant to the pa-thology induced by MDV. The development of methods forgenerating rescuants of these RB1B-based mutants remains ahigh priority. Nevertheless, the use of MDV RB1B-based mu-tants coupled with the excellent in vivo system for analyzingmutant phenotypes in the natural host promises to provide
Anti-MDV US1 Protein Anti-ß-Galactosidase
MD
CC
-UD
03M
DC
C-U
D14
FIG. 9. Immunofluorescence analysis of MDVs reactivated from lymphoblastoid cell lines. CEF monolayers were cocultivated with 106 MDCC-UD03(RB1BD4.5lac-derived) or MDCC-UD14 (RB1Bp9-derived) cells. The infected monolayers were fixed with EtOH 7 days postcocultivation and stained. The antibodiesused were rabbit polyclonal antiserum to the MDV US1-encoded protein (provided by Peter Brunovskis, Case Western Reserve University, and Leland Velicer,Michigan State University) (left panels) and rabbit polyclonal antibodies to b-galactosidase (5-Prime-3-Prime) (right panels).
7896 PARCELLS ET AL. J. VIROL.
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

insight into herpesvirus latency and oncology in general andinto the particulars of how herpesviruses interact with cells oflymphoid lineage.
ACKNOWLEDGMENTS
We thank Peter Brunovskis and Leland Velicer for sharing antiseraand for helpful conversations and Marie Meyer for technical assistancewith the in vivo experiments.This work was supported by the Delaware Research Partnership;
Intervet International, Boxmeer, The Netherlands; and AmericanCancer Society grant VM-44.
REFERENCES
1. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A.Smith, and K. Struhl (ed.). 1987. Current protocols in molecular biology.John Wiley & Sons, New York.
2. Becker, Y., Y. Asher, E. Tabor, I. Davidson, and M. Malkinson. 1994. Openreading frames in a 4556 nucleotide sequence within MDV-1 BamHI-DDNA fragment: evidence for splicing of mRNA from a new viral glycopro-tein gene. Virus Genes 8:55–69.
3. Bradley, G., M. Hayashi, G. Lancz, A. Tanaka, and M. Nonoyama. 1989.Structure of the Marek’s disease virus BamHI-H gene family: genes ofputative importance for tumor induction. J. Virol. 63:2534–2542.
4. Bradley, G., G. Lancz, A. Tanaka, and M. Nonoyama. 1989. Loss of Marek’sdisease virus tumorigenicity is associated with truncation of RNAs tran-scribed within BamHI-H. J. Virol. 63:4129–4135.
5. Brunovskis, P., X. Chen, and L. F. Velicer. 1995. The Marek’s disease virusunique short region: alphaherpesvirus-homologous, fowlpox virus-homolo-gous, and MDV-specific genes. Virology 206:324–338.
6. Buckmaster, A. E., S. D. Scott, M. J. Sanderson, M. E. G. Boursnell, N. L. J.Ross, and M. M. Binns. 1988. Gene sequence and mapping data fromMarek’s disease virus and herpesvirus of turkeys: implications for herpesvi-rus classification. J. Gen. Virol. 69:2033–2042.
7. Calnek, B. W., and R. L. Witter. 1991. Marek’s disease, p. 342–385. In B. W.Calnek (ed.), Diseases of poultry. Iowa State University Press, Ames.
8. Cantello, J. L., A. S. Anderson, A. Francesconi, and R. W. Morgan. 1991.Isolation of a Marek’s disease virus (MDV) recombinant containing the lacZgene of Escherichia coli stably inserted within the MDV US2 gene. J. Virol.65:1584–1588.
9. Chen, X., P. J. A. Sondermeijer, and L. F. Velicer. 1992. Identification of aunique Marek’s disease virus gene which encodes a 38-kilodalton phospho-protein and is expressed in both lytically infected cells and latently infectedlymphoblastoid tumor cells. J. Virol. 66:85–94.
10. Churchill, A. E., R. C. Chubb, and W. Baxendale. 1969. The attenuation withloss of antigenicity of the herpes-type virus of Marek’s disease, strain HPRS-16, on passage in cell culture. J. Gen. Virol. 4:557–564.
11. Delecluse, H.-J., and W. Hammerschmidt. 1993. Status of Marek’s diseasevirus in established lymphoma cell lines: herpesvirus integration is common.J. Virol. 67:82–92.
12. Fukuchi, K., M. Sudo, Y.-S. Lee, A. Tanaka, and M. Nonoyama. 1984.Structure of Marek’s disease virus DNA: detailed restriction enzyme map. J.Virol. 51:102–109.
13. Fukuchi, K., M. Sudo, Y.-S. Lee, A. Tanaka, and M. Nonoyama. 1985. Thestructure of Marek disease virus DNA: the presence of unique expansion innonpathogenic viral DNA. Proc. Natl. Acad. Sci. USA 82:751–754.
14. Hirai, K., M. Sakaguchi, H. Maeda, Y. Kino, H. Nakamura, G.-S. Zhu, andM. Yamamoto. 1992. Construction of recombinant Marek’s disease virustype 1 expressing the lacZ gene of Escherichia coli, p. 150–155. InG. de Boerand S. H. M. Jeurissen (ed.), Proceedings of the Fourth International Sym-posium on Marek’s Disease. Ponsen & Looijen, Wageningen, The Nether-lands.
15. Hong, Y., and P. M. Coussens. 1994. Identification of an immediate-earlygene in the Marek’s disease virus long internal repeat region which encodesa unique 14-kilodalton polypeptide. J. Virol. 68:3593–3603.
16. Isfort, R., D. Jones, R. Kost, R. Witter, and H.-J. Kung. 1992. Retrovirusinsertion into herpesvirus in vitro and in vivo. Proc. Natl. Acad. Sci. USA89:991–995.
17. Jones, D., R. Isfort, R. Witter, R. Kost, and H.-J. Kung. 1993. Retroviralinsertions into a herpesvirus are clustered at the junctions of the short repeatand short unique sequences. Proc. Natl. Acad. Sci. USA 90:3855–3859.
18. Jones, D., L. Lee, J.-L. Liu, H.-J. Kung, and J. K. Tillotson. 1992. Marek’sdisease virus encodes a basic-leucine zipper gene resembling the fos/junoncogene that is highly expressed in lymphoblastoid tumors. Proc. Natl.Acad. Sci. USA 89:4042–4046.
18a.Kent, J. Personal communication.18b.Kent, J., and A. Anderson. Personal communication.19. Kost, R., D. Jones, R. Isfort, R. Witter, and H.-J. Kung. 1993. Retrovirus
insertion into herpesvirus: characterization of a Marek’s disease virus har-
boring a solo LTR. Virology 192:161–169.20. Makimura, K., F.-Y. Peng, M. Tsuji, S. Hasegawa, Y. Kamai, M. Nonoyama,
and A. Tanaka. 1994. Mapping of Marek’s disease virus genome: identifica-tion of junction sequences between unique long and inverted repeat regions.Virus Genes 8:15–24.
21. Maotani, K., A. Kanamori, K. Ikuta, S. Ueda, S. Kato, and K. Hirai. 1986.Amplification of a tandem repeat within inverted repeats of Marek’s diseasevirus DNA during serial in vitro passage. J. Virol. 58:657–660.
22. Marshall, D. R., J. D. Reilly, X. Liu, and R. F. Silva. 1993. Selection ofMarek’s disease virus recombinants expressing the Escherichia coli gpt gene.Virology 195:638–648.
23. Morgan, R. W., J. L. Cantello, and C. H. McDermott. 1990. Transfection ofchicken embryo fibroblasts with Marek’s disease virus DNA. Avian Dis.34:345–351.
24. Morgan, R. W., J. Gelb, Jr., C. S. Schreurs, D. Lutticken, J. K. Rosenberger,and P. J. A. Sondermeijer. 1992. Protection of chickens from Newcastle andMarek’s diseases with a recombinant herpesvirus of turkeys vaccine express-ing the Newcastle disease virus fusion protein. Avian Dis. 36:858–870.
25. Morgan, R. W., and M. S. Parcells. 1992. The isolation and characterizationof mutants mapping to the unique short region of the Marek’s disease virusgenome, p. 156–159. InG. de Boer and S. H. M. Jeurissen (ed.), Proceedingsof the Fourth International Symposium on Marek’s Disease. Ponsen &Looijen, Wageningen, The Netherlands.
26. Nakamura, H., M. Sakaguchi, Y. Hirayama, N. Miki, M. Yamamoto, and K.Hirai. 1992. Protection against Newcastle disease by recombinant Marek’sdisease virus serotype-1 expressing the fusion protein of Newcastle diseasevirus, p. 332–335. In G. de Boer and S. H. M. Jeurissen (ed.), Proceedings ofthe Fourth International Symposium onMarek’s Disease. Ponsen & Looijen,Wageningen, The Netherlands.
27. Ohashi, K., P. H. O’Connell, and K. A. Schat. 1994. Characterization ofMarek’s disease virus BamHI-A-specific cDNA clones obtained from aMarek’s disease lymphoblastoid cell line. Virology 199:275–283.
28. Parcells, M. S., A. S. Anderson, and R. W. Morgan. 1994. Characterizationof a Marek’s disease virus mutant containing a lacZ insertion in the US6(gD) homologue gene. Virus Genes 9:5–13.
29. Parcells, M. S., A. S. Anderson, and R. W. Morgan. 1994. Characterizationof Marek’s disease virus insertion and deletion mutants that lack US1 (ICP22homolog), US10, and/or US2 and neighboring short-component open read-ing frames. J. Virol. 68:8239–8253.
30. Peng, F., G. Bradley, A. Tanaka, G. Lancz, and M. Nonoyama. 1992. Isola-tion and characterization of cDNAs from BamHI-H gene family RNAsassociated with the tumorigenicity of Marek’s disease virus. J. Virol. 66:7389–7396.
31. Peng, F., S. Specter, A. Tanaka, and M. Nonoyama. 1994. A 7 kDa proteinencoded by the BamHI-H gene family of Marek’s disease virus is producedin lytically and latently infected cells. Int. J. Oncol. 4:799–802.
32. Ross, L. J. N., M. M. Binns, and J. Pastorek. 1991. DNA sequence andorganization of genes in a 5.5 kbp EcoRI fragment mapping in the shortunique segment of Marek’s disease (strain RB1B). J. Gen. Virol. 72:949–954.
33. Ross, L. J. N., M. M. Binns, P. Tyers, J. Pastorek, V. Zelnik, and S. Scott.1992. Construction and properties of a herpesvirus of turkeys recombinantexpressing the Marek’s disease virus homologue of glycoprotein B (gB) ofherpes simplex virus, p. 144–149. In G. de Boer and S. H. M. Jeurissen (ed.),Proceedings of the Fourth International Symposium on Marek’s Disease.Ponsen & Looijen, Wageningen, The Netherlands.
34. Sakaguchi, M., T. Urakawa, Y. Hirayama, N. Miki, M. Yamamoto, and K.Hirai. 1992. Sequence determination and genetic content of an 8.9-kb re-striction fragment in the short unique region and the internal inverted repeatof Marek’s disease virus type I DNA. Virus Genes 6:365–378.
35. Sakaguchi, M., T. Urakawa, Y. Hirayama, N. Miki, M. Yamamoto, and K.Hirai. 1993. Sequence determination and genetic content of an 8.9-kb re-striction fragment in the short unique region and the internal inverted repeatof Marek’s disease virus type 1 DNA. Virus Genes 7:211. (Erratum.)
36. Sakaguchi, M., T. Urakawa, Y. Hirayama, N. Miki, M. Yamamoto, G.-S.Zhu, and K. Hirai. 1993. Marek’s disease virus protein kinase gene identifiedwithin the short unique region of the viral genome is not essential for viralreplication in cell culture and vaccine-induced immunity in chickens. Virol-ogy 195:140–148.
37. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
38. Schat, K. A., B. W. Calnek, and J. Fabricant. 1982. Characterization of twohighly oncogenic strains of Marek’s disease virus. Avian Pathol. 11:593–605.
39. Schat, K. A., B. W. Calnek, J. Fabricant, and D. L. Graham. 1985. Patho-genesis of infection with attenuated Marek’s disease virus strains. AvianPathol. 14:127–146.
40. Schat, K. A., C.-L. Chen, W. R. Shek, and B. W. Calnek. 1982. Surfaceantigens on Marek’s disease lymphoblastoid cell lines. JNCI 69:715–720.
41. Silva, R. F., and R. L. Witter. 1985. Genomic expansion of Marek’s diseasevirus DNA is associated with serial in vitro passage. J. Virol. 54:690–696.
42. Silva, R. F., and R. L. Witter. 1992. A non-defective recombinant Marek’sdisease virus with two copies of inserted foreign DNA, p. 140–143. In G. de
VOL. 69, 1995 MAREK’S DISEASE VIRUS MUTANT RETAINS ONCOGENICITY 7897
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from

Boer and S. H. M. Jeurissen (ed.), Proceedings of the Fourth InternationalSymposium on Marek’s Disease. Ponsen & Looijen, Wageningen, The Neth-erlands.
43. Sondermeijer, P. J. A., J. A. J. Claessens, P. E. Jenniskens, A. P. A. Mockett,R. A. J. Thijssen, M. J. Willemse, and R. W. Morgan. 1993. Avian herpes-virus as a live viral vector for the expression of heterologous antigens.Vaccine 11:349–358.
44. Wilson, M. R., and P. M. Coussens. 1991. Purification and characterizationof infectious Marek’s disease virus genomes using pulse field electrophoresis.
Virology 185:673–680.45. Zelnik, V., R. Darteil, J. C. Audonnet, G. D. Smith, M. Riviere, J. Pastorek,
and L. J. N. Ross. 1993. The complete sequence and gene organization of theshort unique region of herpesvirus of turkeys. J. Gen. Virol. 74:2151–2162.
46. Zhu, G.-S., A. Iwata, M. Gong, S. Ueda, and K. Hirai. 1994. Marek’s diseasevirus type 1-specific phosphorylated proteins pp38 and pp24 with commonamino acid termini are encoded from the opposite junction regions betweenthe long unique and inverted repeat sequences of viral genome. Virology200:816–820.
7898 PARCELLS ET AL. J. VIROL.
on February 5, 2014 by guest
http://jvi.asm.org/
Dow
nloaded from
Related Documents











