Restoration management of fly ash deposits crucially influence their conservation potential for terrestrial arthropods Robert Tropek a, b, * ,1 , Ilona Cerna a, b,1 , Jakub Straka c , Tomas Kadlec d , Pavel Pech e , Filip Tichanek b , Pavel Sebek a, b a Institute of Entomology, Biology Centre, Czech Academy of Sciences, Branisovska 31, CZ-37005 Ceske Budejovice, Czech Republic b Faculty of Science, University of South Bohemia, Branisovska 31, CZ-37005 Ceske Budejovice, Czech Republic c Faculty of Science, Charles University in Prague, Vinicna 7, CZ-12844 Prague, Czech Republic d Faculty of Environmental Sciences, Czech University of Life Sciences, Kamycka 129, CZ-165 21 Prague, Czech Republic e Faculty of Science, University of Hradec Kralove, Rokitanskeho 62, CZ-50003 Hradec Kralove, Czech Republic A R T I C L E I N F O Article history: Received 9 March 2014 Received in revised form 6 August 2014 Accepted 2 September 2014 Available online xxx Keywords: Biodiversity conservation Energy industry Restoration ecology Post-industrial biotopes Psammophilous species Human-modified habitats A B S T R A C T Fly ash deposits are a relatively common component of many industrial landscapes. Recently, these post- industrial sites were shown to host highly endangered communities of bees & wasps of vanishing continental drift sands. Nevertheless, nothing is known about this conservation potential for other arthropods, nor how it has been influenced by the practice of restoration. We sampled communities of bees wasps, moths, spiders, hoverflies, and ants colonizing several spontaneously developed and technically reclaimed plots in two fly ash deposits in northern Czech Republic, Central Europe. In addition to species richness, we also concentrated on the potential for colonization by species included in national red-lists and/or specialized to the highly threatened environments of continental drift sand dunes. In total, we recorded 585 species of the studied arthropods, including six considered as nationally extinct, 17 critically endangered, and 84 other red-listed species. From those numbers, 34 species were highly specialized for drift sands. bees & wasps and spiders comprised the overwhelming majority of the threatened species, and all the specialists. The overwhelming majority of the most threatened species colonized the non-reclaimed habitats and generally avoided afforested plots. Some of the endangered bees & wasps occurred also in the artificial grasslands, probably feeding on nectar. Summarizing, the studied fly ash deposits harbor arthropod communities of high conservation interest, importance of the spontaneously developed habitats differ among individual arthropod groups. However, this conservation potential is suppressed by artificial afforestation. Following the occurrence patterns of the most threatened species we suggest applying a mosaic-like combination of non-reclaimed habitats with reclaimed grasslands as a potentially effective solution combining both biodiversity conservation and environmental protection. The expected ensuing controversy of this approach should be one of the most essential challenges of current restoration ecology research and practice. ã 2014 Elsevier B.V. All rights reserved. 1. Introduction During the past few decades, effective restoration of various post-industrial sites, as typical symbols of man-made degradation of the European landscape, has been intensively studied. The majority of research efforts have targeted post-mining landscapes, which are usually the most common artificial habitats in Europe and some other regions (Walker, 1992; Tropek and Prach, 2012). As a consequence, a purely negative view of the post-mining barrens has been changed, as they often harbor endangered communities of many groups (e.g., insects and other arthropods (e.g., Benes et al., 2003; Krauss et al., 2009; Dolny and Harabis, 2012; Lenda et al., 2012; Heneberg et al., 2013), birds (e.g., Bejcek and Stastny, 1984; Salek, 2012), amphibians (Galan, 1997; Smolova et al., 2010), and plants (e.g., Wheater and Cullen, 1997; Tomlinson et al., 2008; Tropek et al., 2013b)). Recently, a simultaneous thorough research of restoration practices revealed spontaneous or directed * Corresponding author at: Institute of Entomology, Biology Centre, Czech Academy of Sciences, Branisovska 31, CZ-370 05 Ceske Budejovice, Czech Republic. Tel.: +420 387775030 fax: +420 389022263. E-mail addresses: [email protected] (R. Tropek), mufi[email protected] (I. Cerna), [email protected] (J. Straka), [email protected] (T. Kadlec), [email protected] (P. Pech), [email protected] (F. Tichanek), [email protected] (P. Sebek). 1 These authors contributed equally to this work. http://dx.doi.org/10.1016/j.ecoleng.2014.09.011 0925-8574/ ã 2014 Elsevier B.V. All rights reserved. Ecological Engineering 73 (2014) 45–52 Contents lists available at ScienceDirect Ecological Engineering journal homepage: www.else vie r.com/locat e/e coleng

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ecological Engineering 73 (2014) 45–52
Restoration management of fly ash deposits crucially influence theirconservation potential for terrestrial arthropods
Robert Tropek a,b,*,1, Ilona Cerna a,b,1, Jakub Straka c, Tomas Kadlec d, Pavel Pech e,Filip Tichanek b, Pavel Sebek a,b
a Institute of Entomology, Biology Centre, Czech Academy of Sciences, Branisovska 31, CZ-37005 Ceske Budejovice, Czech Republicb Faculty of Science, University of South Bohemia, Branisovska 31, CZ-37005 Ceske Budejovice, Czech Republicc Faculty of Science, Charles University in Prague, Vinicna 7, CZ-12844 Prague, Czech Republicd Faculty of Environmental Sciences, Czech University of Life Sciences, Kamycka 129, CZ-165 21 Prague, Czech Republice Faculty of Science, University of Hradec Kralove, Rokitanskeho 62, CZ-50003 Hradec Kralove, Czech Republic
A R T I C L E I N F O
Article history:Received 9 March 2014Received in revised form 6 August 2014Accepted 2 September 2014Available online xxx
Keywords:Biodiversity conservationEnergy industryRestoration ecologyPost-industrial biotopesPsammophilous speciesHuman-modified habitats
A B S T R A C T
Fly ash deposits are a relatively common component of many industrial landscapes. Recently, these post-industrial sites were shown to host highly endangered communities of bees & wasps of vanishingcontinental drift sands. Nevertheless, nothing is known about this conservation potential for otherarthropods, nor how it has been influenced by the practice of restoration. We sampled communities ofbees wasps, moths, spiders, hoverflies, and ants colonizing several spontaneously developed andtechnically reclaimed plots in two fly ash deposits in northern Czech Republic, Central Europe. In additionto species richness, we also concentrated on the potential for colonization by species included in nationalred-lists and/or specialized to the highly threatened environments of continental drift sand dunes. Intotal, we recorded 585 species of the studied arthropods, including six considered as nationally extinct,17 critically endangered, and 84 other red-listed species. From those numbers, 34 species were highlyspecialized for drift sands. bees & wasps and spiders comprised the overwhelming majority of thethreatened species, and all the specialists. The overwhelming majority of the most threatened speciescolonized the non-reclaimed habitats and generally avoided afforested plots. Some of the endangeredbees & wasps occurred also in the artificial grasslands, probably feeding on nectar. Summarizing, thestudied fly ash deposits harbor arthropod communities of high conservation interest, importance of thespontaneously developed habitats differ among individual arthropod groups. However, this conservationpotential is suppressed by artificial afforestation. Following the occurrence patterns of the mostthreatened species we suggest applying a mosaic-like combination of non-reclaimed habitats withreclaimed grasslands as a potentially effective solution combining both biodiversity conservation andenvironmental protection. The expected ensuing controversy of this approach should be one of the mostessential challenges of current restoration ecology research and practice.
ã 2014 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Ecological Engineering
journal homepage: www.else vie r .com/ locat e/e coleng
1. Introduction
During the past few decades, effective restoration of variouspost-industrial sites, as typical symbols of man-made degradation
* Corresponding author at: Institute of Entomology, Biology Centre, CzechAcademy of Sciences, Branisovska 31, CZ-370 05 Ceske Budejovice, Czech Republic.Tel.: +420 387775030 fax: +420 389022263.
E-mail addresses: [email protected] (R. Tropek), [email protected](I. Cerna), [email protected] (J. Straka), [email protected](T. Kadlec), [email protected] (P. Pech), [email protected] (F. Tichanek),[email protected] (P. Sebek).
1 These authors contributed equally to this work.
http://dx.doi.org/10.1016/j.ecoleng.2014.09.0110925-8574/ã 2014 Elsevier B.V. All rights reserved.
of the European landscape, has been intensively studied. Themajority of research efforts have targeted post-mining landscapes,which are usually the most common artificial habitats in Europeand some other regions (Walker, 1992; Tropek and Prach, 2012). Asa consequence, a purely negative view of the post-mining barrenshas been changed, as they often harbor endangered communitiesof many groups (e.g., insects and other arthropods (e.g., Benes et al.,2003; Krauss et al., 2009; Dolny and Harabis, 2012; Lenda et al.,2012; Heneberg et al., 2013), birds (e.g., Bejcek and Stastny, 1984;Salek, 2012), amphibians (Galan, 1997; Smolova et al., 2010), andplants (e.g., Wheater and Cullen, 1997; Tomlinson et al., 2008;Tropek et al., 2013b)). Recently, a simultaneous thorough researchof restoration practices revealed spontaneous or directed

46 R. Tropek et al. / Ecological Engineering 73 (2014) 45–52
succession as a sufficiently effective and cheap method encourag-ing the conservation potential of post-mining landscapes (e.g.,Tropek et al., 2010, 2012; Hodacova and Prach, 2003; Salek, 2012),excepting specific plots where other public concerns (e.g., erosionrisks, acid rock drainage, stream sedimentation, toxin leaks, andpublic safety issues) outweigh nature conservation (Prach andHobbs, 2008; Tropek et al., 2010). Noticeably less attention hasbeen paid to the biodiversity of other post-industrial sites.Nevertheless, some of them (e.g., road verges (Munguira andThomas, 1992; Hopwood, 2008; Ascensao et al., 2012), brownfields(Eversham et al., 1996; Eyre et al., 2003; Strauss and Biedermann,2005), or bases of electricity pylons (Tryjanowski et al., 2014)) arealso known to be colonized by rare species. Hence, similar patternsin still non-studied post-industrial sites could be expected,although wider surveys are still rather sporadic.
Deposits of coal combustion by-products (often called as fly ashdeposits because fly ash, i.e., very fine (average diameter <10 mm)glass-like particles of mineral residua, constitute more than 70% ofthe deposited wastes, Haynes, 2009), accompany practically everypower station, heating plant and many larger factories. Recently, asurvey of communities of aculeate hymenopterans revealed the flyash deposits in the Czech Republic as crucial refuges for criticallyendangered communities specialized for inland sand dunes(Tropek et al., 2013a), a highly endangered and rapidly vanishinghabitat throughout Europe (Tropek et al., 2013a; Fanta and Siepel,2010). Similarly interesting species were not found in a fewexisting published surveys (of plants (Ash et al., 1994; Vankova andKovar, 2004), lichens (Palice and Soldan, 2004), fungi (Kubatovaet al., 2002), and springtails (Shaw, 2003)), which was usuallyexplained by the hostile substrate chemistry. On the other hand,there is a wide lack of any published studies of other arthropods.Because various arthropods are often known to perceive thestructure of a microhabitat, including substrate, more sensitivelythan its chemical attributes (e.g., Shreeve et al., 2004; Dennis et al.,2006), we hypothesized the finding of patterns similar to the pilotsurvey of the aculeate hymenopterans (Tropek et al., 2013a).
Because of the strongly negative impacts of flying ash on humanhealth and the environment (e.g., high content of heavy metals andother toxins, extreme pH, lack of nitrogen and organic matter;Borm,1997; Adriano et al., 2002), almost all ecological research hasbeen motivated by fly ash deposits’ restoration. The most seriousproblems with fly ash are caused by its easy dispersibility by wind,and both main restoration approaches, technical and biotechno-logical (mainly phytoremediation and directed revegetation;Pandey et al., 2009, 2012; Pandey and Singh 2010), thus aim atrapid preventing of any aeolian erosion (Haynes, 2009; Pandey andSingh, 2012). Alternatively, the fly ash is used as a by-product incement and concrete production, agriculture, and soil stabilizationin other post-industrial sites’ reclamation (Sajwan et al., 2006;Gonzalez et al., 2009; Pandey et al., 2009; Ram and Masto, 2010).During technical reclamation (sensu Prach and Hobbs (2008) andTropek et al. (2010)), the ash surface is usually covered by fertiletopsoil and converted to productive forest, meadow or cropland(Haynes, 2009; Pandey and Singh, 2012). The biotechnologicalapproach is based on the adding of nutrients (often in the form ofother wastes) and acceleration of succession, usually by mycor-rhiza or microbe inoculation and/or other direct support ofselected successionally strong plants (Juwarkar and Jambhulkar,2008; Tandy et al., 2011). These interventions totally suppress theunique physical attributes of the bare ash, which seem to be crucialfor colonization by the specialized psammophilous insects (Tropekand Konvicka, 2011; Tropek et al., 2013a). Nevertheless, no researchconsidering the impact of different restoration methods on thebiodiversity of threatened arthropods exists. Because of this strongdiscrepancy between restoration methods protecting public healthand environment and those protecting biodiversity of critically
endangered species, restoration of fly ash deposits is one of themost current challenges of ecological engineering (see also Tropekand Konvicka, 2011; Pandey et al., 2009).
In this paper, we extend our pilot study of non-reclaimed fly ashdeposits habitats (Tropek et al., 2013a) for five groups ofarthropods differing in their natural history (bees & wasps, moths,spiders, hoverflies, ants) and compare it with communities oftechnically restored plots within the same localities. We concen-trated on three main questions: (1) What is the potential of fly ashdeposits to harbor endangered communities of different arthro-pods? (2) How does this potential differ among various habitatswithin the deposits? (3) How is this potential influenced bytechnical reclamation? So far, this is the first study evaluating thesuccess of restoration management of fly ash deposits for anyarthropods and the results can thus be crucial for any ecologicalengineering planning.
2. Methods
2.1. Studied groups and species categorization
We targeted five groups of arthropods with diverse naturalhistory: bees & wasps (Hymenoptera: Aculeata, excepting ants withtotally different habitat use), moths (Lepidoptera: Macrolepidop-tera), spiders (Araneae), hoverflies (Diptera: Syrphidae), and ants(Hymenoptera: Formicidae). All collected adult specimens fromthese groups were identified as to species level. Small part of thedataset, i.e., the bees & wasps collected in the three non-reclaimedplots (see below), were already included to Tropek et al. (2013a),here we use the complete dataset for answering differentquestions than in the cited work.
To show the importance of the different habitats for thearthropods’ conservation, we focused on the life-history patternsof the communities (Tropek et al., 2008). Hence, we focused on theconservation value of the communities, based on the status ofindividual species in the national red lists (with categories EX –
considered as extinct in the Czech Republic, CR – criticallyendangered, EN – endangered, VU – vulnerable, NT – nearly-threatened, and LI – low interest, not threatened), and on thepsammophilous specialisation of the communities, based on thelevel of individual species specialization for the sandy habitats(with categories DS – highly specialized psammophilous speciesfound almost exclusively in drift sands with a large proportion ofbare substrate, PS – psammophilous species found also on otherfinely-grained natural substrates such as alluvial sand, loess, ormarl, and NS – species not specialized to finely grained substrates).These categories were stated based on published data, the list ofreferences used for individual groups is included to Appendix 1.See Appendix 1 also for nomenclature references and a list of allrecorded species with their category memberships.
2.2. Study sites and sampling design
The Polabi region, northern Czech Republic, Central Europe, islowland with a relatively mildly warm temperate climate (sensuQuitt (1971); average temperature 8–9 �C, annual precipitation550–700 mm). It is a granary of the country, with extensiveagriculture, causing significant impoverishment of its naturalhabitats. As a consequence, continental drift sands, once relativelycommon in this region, have almost completely disappeared due toafforestation and/or succession during the last century (Tropeket al., 2013a).
In this region, we surveyed arthropods colonizing two relativelylarge fly ash deposits. The Chvaletice ash deposit (50�10N, 15�270E;250 m a.s.l.), where ash from the Chvaletice Power Station hassedimented since the late 1970s, and the Bukovina ash deposit

R. Tropek et al. / Ecological Engineering 73 (2014) 45–52 47
(50�70N, 15�500E; 250 m a.s.l.), where ash from the Opatovice nadLabem Power Station has been deposited since the 1960s. At thebeginning of the 2000s, the establishment of desulphurization inboth power stations changed the ash deposition technology,terminating the sedimentation of the wastes. Almost entiresedimentation lagoons were drained and the exposed fly ashwas left to spontaneous succession. At present, both localities arebeing technically reclaimed by covering with fertile topsoil andconversion to species-poor productive grasslands and/or forestplantations. During our research, the non-reclaimed ash surfacesstill covered ca 20 ha in the Chvaletice ash deposit and 10 ha in theBukovina ash deposit.
At both localities, insects were sampled in five habitats, threespontaneously developed (i–iii) and two established by differentrestoration methods (iv and v): (i) plots with barren-dry stronglydesiccative and thus with loose ash (herbaceous cover <10%); (ii)plots in the vicinity of water lagoons and/or in large depressions withbarren-wet and thus presumably more compact ash (herbaceouscover <10%); (iii) plots of ash overgrown by Calamagrostis epigejosand other grasses (herbaceous cover >80%); (iv) artificial forestsestablished bycovering with fertile topsoil and subsequent plantingof Pinus sylvestris; and (v) artificial meadows established by thesowing of species-poor grass and herb mixtures on plots covered byfertile topsoil. As the ash sedimentation terminated in both studieddeposits in the early 2000s, the majority of the studied plots werestill relatively young. All technically reclaimed plots were selectedto be also about 10 years old for better comparability with the non-reclaimedhabitats. During this decade, the meadows were regularlymown at least twice a year, while the other plots were unmanaged.All the three spontaneously developed habitat types and theirtransitions create the overwhelming majority of non-reclaimedparts of both studied deposits, while both reclaimed habitatsrepresent the two main technical reclamation methods prevailingin the restoration of various post-industrial sites in Europe (Prachand Hobbs, 2008; Tropek et al., 2012). Within each study site, wehaphazardly (in the sense of avoiding two neighboring plots of thesame habitat type) established three replicates of each, with at leastseveral tens of meters between any neighboring plots. In eachreplicate, we established one 3 m � 3 m study plot where 9 yellowpan traps (15 cm in diameter) filled with water and a small amountof detergent were exposed in a regular grid. Four exposure times (1–3 May, 13–15 June, 1–3 July, 30 July, 1 August, all 2009), all underoptimal weather (>20 �C in spring and >25 �C in summer, clear skies,not windy), covered the key emergence periods of the studiedgroups. In additional to pan-trapping, all ant workers were recordedduring detailed surveys of the study plots in August 2009. Mothswere collected by portable light traps (made by Hutur NGO, HradecKralove, Czech Republic, with two 3 W UV Cold Cathode Lights madeby Akasa Ltd. Middlesex, UK), which attract moths from maximallytens of meters and thus are ideal for microhabitat studies (Truxa andFiedler, 2012). One light trap was exposed in each plot for 6individual nights (8–9 May, 7–8 June, 22–23 June, 27–28 July, 9–10 August, 13–14 September, all 2010) throughout the seasoncovering flight periods of the greatest diversity of moths.
Because some of the studied groups could be affected by theactual plant food rewards more than the microhabitat structure,four nectar-related variables were recorded: actual nectar plants’abundance assessed within each study plot (NectAmountIn) and inits 5 m surroundings (NectAmountOut) on a ranked (0–3) scale,and the number of actually flowering plant species within thesame areas (NectRichIn and NectRichOut).
2.3. Statistical analyses
For all analyses, all specimens collected in each plot throughoutthe whole study and by all traps were pooled together in order to
avoid a pseudo-replicative design. As abundance of ants reflectsrather the distance from their nests than the microhabitatsuitability, only the presence of individual ant species in thestudied plots was analyzed.
The effects of particular habitats on species richness (i.e.,number of species per plot), conservation value (i.e., number of allindividuals per plot weighted by their red-list status: EX – 5; CR –
4; EN – 3; VU – 2; NT – 1; LI – 0), and psammophilousspecialisation (i.e., number of all individuals per plot weighted bytheir specialization level: DS – 2; PS – 1; NS – 0; see Tropek et al.,2010 for details) were tested separately for each of the five studiedgroups by generalized linear mixed-effect models (GLMM) withquasi-Poisson distribution. All analyses were performed in R 2.14.2software (R Development Core Team, 2011) using glmmPQLfunction in MASS package (Venables and Ripley, 2002). Thehabitat type represented a fixed-effect factor variable, particularash deposits acted as a random factor. Post-hoc comparisonsamong the habitat levels were done by setting different treatmentcontrasts. For bees & wasps, moths, and hoverflies, the fourvariables related to the nectar sources (see above) were averagedfor each plot, then tested by stepwise selection and the significantvariables were included into the models before testing the habitatvariable.
All multivariate ordinations were computed in Canoco forWindows 4.5 (ter Braak and Smilauer, 2002). Differences in speciescomposition of the communities sampled in particular habitatswere analyzed by detrended canonical correspondence analyses(DCCA). We used logarithmic transformation of the species dataand downweighting of the rare species option. The permutationdesign reflected the spatial arrangements of the samples (theMonte Carlo permutation tests, 999 runs under the full permuta-tion model, study plots as split plots and study sites as whole plots,both levels freely permuted). Possible effects of the nectar relatedfactors (see above) were tested by the CANOCO forward selectionprocedure for bees & wasps, moths, and hoverflies; as none of thefactors showed a significant effect (p > 0.05) for any group theywere not included into the final models.
3. Results
In total, we recorded 4731 individuals of 277 bee & wasp species(from these numbers, 3479 individuals of 227 species sampled innon-reclaimed habitats were already included to Tropek et al.,2013a), 1547 individuals of 202 moth species, 504 individuals of65 spider species, 363 individuals of 30 hoverfly species, and 11 antspecies. From these numbers, 6 species were considered asnationally extinct (only bees & wasps), 17 as critically endangered(15 bees & wasps, and 2 spiders), 26 as endangered (24 bees &wasps, 1 spider, and 1 hoverfly), 41 as vulnerable (35 bees & wasps,1 moth, 4 spiders, and 1 hoverfly), and 17 as nearly-threatened(1 moth, and 16 spiders; this category was not used in the bees &wasps national red lists at all). In addition, 34 specialists of driftsands (33 bees & wasps, and 1 spider) were found. In total, 20% ofbee & wasp species, 1% of moth species, 34% of spider species, 7% ofhoverfly species, and 0% of ant species recorded during this studywere included in the national red lists.
The species richness significantly depended on the habitat typein all studied groups (see Fig. 1 for results of the post-hoccomparisons). The species richness (with significant effect ofNectAmountIn: F1,23 = 21.65, P = 0.0001) of bees & wasps wassignificantly lower in forest than all other habitats, and higher inbarren-wet than in both reclaimed habitats (F4,23 = 19.44, P< 0.0001). The lowest species richness of moths (with significanteffects of NectRichOut: F1,22 = 6.47, P = 0.019; and NectRichIn:F1,22 = 7.33, P = 0.013) were revealed in barren-wet and the highestin barren-dry habitats (F4,22 = 3.82, P = 0.016). For spiders, meadow
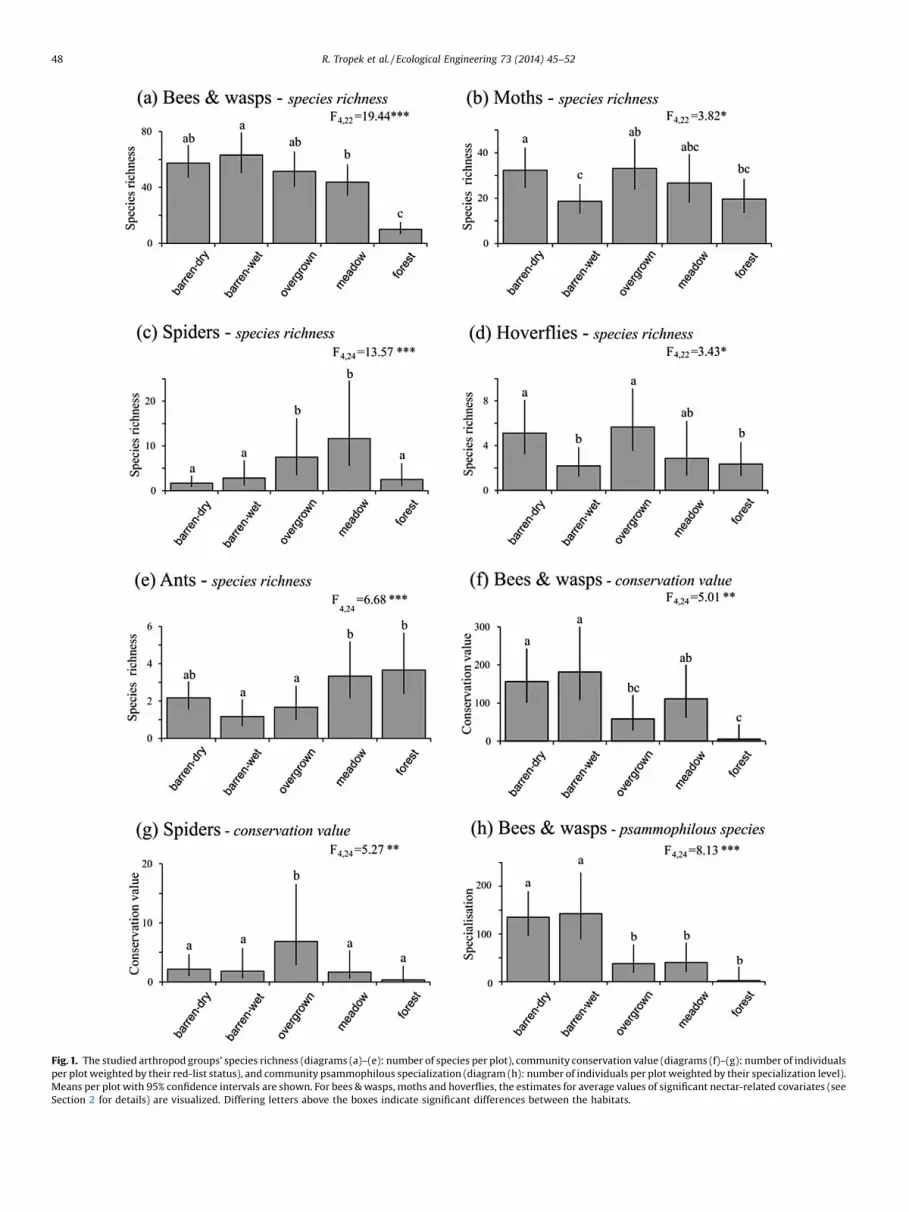
Fig. 1. The studied arthropod groups’ species richness (diagrams (a)–(e): number of species per plot), community conservation value (diagrams (f)–(g): number of individualsper plot weighted by their red-list status), and community psammophilous specialization (diagram (h): number of individuals per plot weighted by their specialization level).Means per plot with 95% confidence intervals are shown. For bees & wasps, moths and hoverflies, the estimates for average values of significant nectar-related covariates (seeSection 2 for details) are visualized. Differing letters above the boxes indicate significant differences between the habitats.
48 R. Tropek et al. / Ecological Engineering 73 (2014) 45–52

Table 1Results of the detrended canonical correspondence analy-ses (DCCA) of influence of the studied habitats on thecommunity composition of studied taxa.
1st axis F,Pa, andexplained variationb
All axes F, Pa, andexplained variationb
Bees &wasps
3.1**; 11.0% 2.2**; 26.4%
Moths 2.1**; 10.1% 1.6**; 26.4%Spiders 1.8*; 7.6% 1.8**; 24.2%Hoverflies 3.04**; 10.9% 2.83**; 71.1%Ants 4.2n.s.; n.a. 2.1n.s.; n.a.
a P-values: n.s. P � 0.05; * P < 0.05; ** P < 0.001.b The variation in species data explained by the first or all
ordination axes.
R. Tropek et al. / Ecological Engineering 73 (2014) 45–52 49
and overgrown were species-richer than all other habitats(F4,24 = 13.57, P < 0.0001). For hoverflies, the species richness (withsignificant effects of NectRichOut: F1,22 = 8.97, P = 0.007; andNectRichIn: F1,22 = 8.67, P = 0.008) was highest in barren-dry andovergrown and lowest in barren-wet and forest (F4,22 = 3.43,P = 0.025). The highest species richness of ants were revealed inboth reclaimed habitats (F4,24 = 6.68, P = 0.0009).
Fig. 2. DCCA diagrams of relationships of the arthropod groups to habitats in the studied fl
status ((a), (b), (c) and (f): red diamonds – regionally extinct or critically endangered;
crosses – not threatened species) or psammophilous specialization ((d) and (e): red diamspecies able to occur also in other finely grained substrates; black crosses – non-psamminformation for individual species’ categorization. (For interpretation of the references to
Because only few or even no specimens of threatened moths,hoverflies and ants were found and the data thus did not showenough response variation, the differences in conservation valueamong particular habitats were tested only for bees & wasps andspiders (Fig. 1). For bees & wasps, the significantly most valuedcommunities (with no significant effect of the nectar-relatedvariables) were revealed in barren-dry and barren-wet, while thesignificantly least valued were in forest (F4,24 = 5.01, P = 0.0045).Overgrown hosted significantly more valued communities ofspiders than all other habitats (F4,24 = 5.27, P = 0.0034). Similarly,the response variation of psammophilous specialisation wassufficient only in bees & wasps (Fig. 1), where barren-dry andbarren-wet were revealed to host the significantly highestspecialized communities (F4,24 = 8.14, P = 0.0003). SeeAppendix 2 for detailed results of all the final models.
Species composition of all studied groups except antssignificantly differed among the habitats (Table 1). For bettercomprehensibility we chose visualization of threatened andspecialized species of each significant group (except moths andhoverflies, where no specialized species were recorded) separately(Fig. 2). A gradient between the spontaneously developed (i.e.,barren-dry, barren-wet and overgrown) and the technicallyreclaimed (i.e., forest and meadow) plots along the main ordinationaxis was obvious in all significant analyses. While these differenceswere more evident for moths and spiders, communities of bees &wasps, and hoverflies recorded in meadow were closer to
y ash deposits. The symbols distinguish individual species according their red-listedgreen triangles – endangered; blue stars – vulnerable or nearly-threatened; blackonds – species highly specialized to drift sands; green triangles – psammophilousophilous species). See Table 1 for associated statistics and Table S1 in Supporting
color in this figure legend, the reader is referred to the web version of this article.)

50 R. Tropek et al. / Ecological Engineering 73 (2014) 45–52
spontaneously developed plots than forest along the main axis.Among the groups with significant responses, both threatened andsand-specialized species generally preferred spontaneous succes-sion to technical reclamation, with very rare exceptions.
4. Discussion
4.1. Conservation potential of fly ash deposits for different arthropods
Extending the dataset with more groups of arthropods, togetherwith covering formerly neglected technically reclaimed habitats,confirmed the noticeable conservation potential of fly ash depositsrevealed by the pilot study of bees & wasps (Tropek et al., 2013a)for threatened spiders but not for the other groups. More than athird of the recorded spider species (including two criticallyendangered ones: Arctosa perita and Clubiona juvenis) are includedin the national red list. The finding of two more nationally extinct(Nomada minuscula and Nomada stigma) and two more criticallyendangered (Arachnospila wesmaeli and Chrysura simplex) speciesof bees & wasps, which were not sampled in spontaneouslydeveloped plots (Tropek et al., 2013a), even strengthened theimportance of these post-industrial barrens for conservation ofthis group. Both these groups are generally threatened in CentralEurope, with many extinct or critically endangered speciesoverwhelmingly related to the strong decrease in their habitats(Buchar and Ruzicka, 2002; New, 2012). Various vanishing speciesfrom both groups were also repeatedly revealed to colonize post-industrial or other anthropogenic secondary habitats (Tropek andKonvicka, 2008; Heneberg et al., 2013; Krauss et al., 2009). On theother hand, it was already shown that arthropods at fly ashdeposits could contain higher concentrations of some toxins(Brieger et al., 1992). Considering relatively short life cycle of thespecies recorded in our study and high abundances of many ofthem, including some of the most critically endangered ones, wehypothesized that this contamination is less serious than strongdecrease of their natural habitats, but more research on this isurgently needed.
Especially in bees & wasps, a significant number of recordedspecies, including the most threatened ones, are drift sandspecialists as was already published using part of the data inTropek et al. (2013a). Hence, it seems that the fine structure of flyash physically mimics natural drift sand for at least part of thisgroup and that these secondary habitats crucially contribute to itsconservation (Tropek et al., 2013a). In spiders, the criticallyendangered A. perita is specialized for drift sand dunes and thenearly-threatened Steatoda albomaculata for other sandy habitats,such as riverbeds. The absence of other drift sand specializedspiders known from the region (Buchar and Ruzicka, 2002;Dolansky, 2003) could imply either that the fine structure of flyash is a sufficient factor for only some of the psammophilousspiders or that these specialists are less effective in colonizing thenewly established habitats than the specialized bees & wasps. Onthe other hand, the pan-trapping method used is not as effective incapturing spiders as it is for bees & wasps (Samways et al., 2009)and other present specialized species could be thus overlooked.Comparing this study to the only other detailed survey of spiders(Rezac, 2004) from a deposit of anthropogenic finely-grainedsubstrate, a dried sludge from pyrite mining, where no more driftsand specialists were found by quantitative pitfall-trapping, itseems to be less probable. Some other recorded threatened bees &wasps and a majority of the recorded threatened spiders arespecialized for sunny rocks and well-preserved xeric grasslands orforest steppes and warm forest edges, which is congruent withalready known communities in various post-industrial habitats.Nevertheless, records of the critically endangered spider C. juvenis
specialized for non-destroyed wetlands implies that even wetparts of the deposits could have some conservation potential.
For the other studied arthropods, the conservation potential ofthe studied sites is significantly lower, if there is any at all, aspractically all recorded species are relatively common in theadjoining cultural landscape. Especially in moths and hoverflies, alow abundance of many of the recorded species, despite theoptimality of the used methods for recording of these groups (Fryand Waring, 2001; Samways et al., 2009), is evident (cf.Appendix 1). It even indicates that the majority of the commonspecies do not reproduce at the studied deposits and the recordedspecimens were just passing through. Considering that manythreatened lepidopteran species have been recorded in variouspost-mining habitats (e.g., Benes et al., 2003; Lenda et al., 2012;Tropek et al., 2010, 2013b), it seems that fly ash itself is hostile forthis group. To the best of our knowledge, no comprehensive studyof hoverflies colonizing post-industrial sites exists for comparison.Since both groups are presumably herbivorous, which differ-entiates them from the other studied arthropods, we hypothesizethat the dearth of any suitable food sources, caused by accumula-tion of heavy metals (Jambhulkar and Juwarkar, 2009) which couldbe toxic for the larvae, could cause this pattern. Higherconcentrations of some heavy metals, such as arsenic, cadmiumand selenium, have been proved in vertebrates living in coal ashdeposits (Bryan et al., 2012; Metts et al., 2013), but no study ofinvertebrates exists. The plant communities of fly ash deposits arealso not known to be formed of psammophytic species (Ash et al.,1994; Vankova and Kovar, 2004), which could also be a constraintfor the most specialized herbivorous arthropods.
Ants are the only group where no species of conservation valuewere found. It is known, however, that in the forming of antcommunities, inter-species interactions are often more importantthan available niches, and that the presence of the top-competitorscan cause low richness of ant species (e.g., Rosengren, 1986;Puntilla et al., 1996). Both studied deposits were colonized byterritorial and aggressive Tetramorium sp. and Lasius niger, whichprobably prevent a colonization of Lasius psammophilus, aregionally present sand specialist, and other relatively smallerspecies.
4.2. Habitat preferences and restoration practice
From the biodiversity conservation point of view, the technicalreclamation of the studied ash deposits is significantly lesseffective than spontaneous succession, as the overwhelmingmajority of threatened and specialized arthropods preferrednon-reclaimed plots and almost generally avoided the afforestedplots. This is fully congruent with studies of post-mining sitesrevealing traditional technical reclamation as squandering conser-vation potential and funds (e.g., Tropek et al., 2010, 2012; Hodacovaand Prach, 2003; Salek, 2012).
The two study groups containing high numbers of red-listedspecies, i.e., bees & wasps and spiders, differed in preferences of thenon-reclaimed habitats. While the spiders’ conservation value wassignificantly higher in overgrown plots than all other studiedhabitats, both types of barren habitats were more important thanthe overgrown plots and the artificial meadows for bees & wasps(and especially for the drift sand specialists). This could beexplained by needs of the available resources differing among thegroups. The majority of the most endangered recorded species ofbees & wasps require barren, finely-grained substrate for their nestbuilding (New, 2012), which is generally suppressed by substrateovergrowth. On the other hand, almost all the endangered speciesof spiders are specialists of xeric grasslands or forest steppes, forwhich the bare fly ash has too extreme a microclimate (Fanta andSiepel, 2010), which is probably partly suppressed by spontaneous

R. Tropek et al. / Ecological Engineering 73 (2014) 45–52 51
overgrowth. As a confirmation, both of the recorded sandspecialists preferred barren habitats.
Both studied technical reclamation approaches diminishmicrotopographic heterogeneity and increase nutrients (Tropeket al., 2010) and thus create rather ruderal habitats dominated bycompetitively strong common generalists, which suppress thecolonization potential of endangered and/or specialized bees &wasps and spiders. This effect of technical reclamation onarthropods has been shown in other post-industrial sites (Tropeket al., 2010, 2012, 2013b; Hendrychova et al., 2012). On the otherhand, the sown meadows offer sufficient nectar and pollen sourcesand they are thus visited by psammophilous bees & wasps nestingin non-reclaimed parts of the deposits with limited nectar sources.These patterns are often contradictory to the species richnessanalyses, which showed the overgrown and/or technicallyreclaimed plots fully comparable with the barren habitats. Thiswas, however, caused by the presence of the generalist species,which are not the priority of the modern conservation approaches,as they are relatively common in the surrounding landscape(Tropek et al., 2008; Samways et al., 2009). Patterns within theother studied groups are not consistent; these communities,however, generally consist of common species with very lowconservation value and, as we concluded that the studied depositshave inconsiderable conservation potential for those groups, therestoration practice would not affect them.
To summarize, artificial afforestation is the worst reclamationmethod for the arthropod biodiversity of the fly ash deposits inCentral Europe, and probably also in other temperate areas. Acombination of spontaneous development with carefully planneddisturbances, maintaining a mosaic of plots in different succes-sional stages (including the earliest bare ones) seems to be mosteffective for conservation of the most threatened species (cf. Prachand Hobbs, 2008; Tropek et al., 2013b). As shown for bees & wasps,artificially established non-intensive grasslands could provide animportant nectar source. Examples of restoration of differentthreatened habitats (such as mesotrophic sparse grasslands: Tandyet al., 2011) on the finely-grained anthropogenic substrates are alsoknown, but they must not be applied on large areas with apotential for conservation of more valued communities (Tropekand Konvicka, 2011) and the biodiversity-focused restorationshould be more mosaic-like. This approach could be in congruencewith preventing aerial erosion of the substrate by the establish-ment of windbreaks protecting the bare plots, but other appliedresearch is necessary.
4.3. Future of fly ash deposits
At least for the next few decades, coal combustion will remainan important component of energetic industry in many parts of theworld, and fly ash will thus remain an unavoidable waste.Nevertheless, the technology of its separation and depositinghas rapidly changed with power plant modernization throughoutthe world (e.g., Sajwan et al., 2006). The practice of sedimentationof all the coal combustion wastes in artificial water lagoons withfluctuating water levels has been widely replaced by collecting drywastes directly from the smoke and depositing them as a drymixture with calcium sulfate from flue-gas desulphurization(Sajwan et al., 2006). A subsequent hydration of the mixtureleads to its chemical stabilization, preventing wind and watererosion. No study of the biodiversity of these mixtures depositsexists, but the microhabitat structure is totally different, especiallybecause of the lack of any finely-grained substrate, which seems tobe the crucial factor for the drift sand specialists.
Simultaneously, there is strong public pressure for rapidreclamation of the remaining drained fly ash sedimentationlagoons (Borm, 1997; Pandey and Singh, 2012). This pattern is
similar throughout Central Europe and probably elsewhere (Borm,1997; Haynes, 2009), and it is demonstrable even in the studieddeposits, as many of the studied plots have already been reclaimedsince the time of our survey and the rest are currently underreclamation. As a consequence, we have the last chance forstudying these post-industrial barrens colonizations and trying toconserve their biodiversity by responsible restoration.
In conclusion, the conservation potential of the fly ash depositsfor colonizing by vanishing drift sand specialists of at least somearthropod groups is tremendous. Nevertheless, further research ofthe biodiversity of other crucial bioindicator groups with aconsiderable amount of the psammophilous specialists from otherareas is greatly needed. On the other hand, we are far fromadvocating coal combustion and the establishment of any newdeposits, as their enormous impacts on the environment and risksfor human health are indisputable. Wisely restored alreadyexisting fly ash deposits combining the needs of both environ-mental risks prevention and biodiversity conservation couldharbor the rapidly vanishing communities of continental sanddunes. Being optimistic, we believe there will be landscape-scaledrestoration of the afforested and/or overgrown natural drift sandssooner or later. The ecologically restored fly ash deposits couldthen offer a crucial regional species pool for the rehabilitation ofnatural communities.
Acknowledgements
The authors would like to thank the fly ash deposits operatorsfor access to the study sites, M. Konvicka, O. Cizek, D. Mach, andJ. Rehounek for valuable discussions, L. Mazanek for somehoverflies revision, and M. Sweney for English corrections. Thestudy was funded by the Czech Science Foundation (P504/12/2525), the University of South Bohemia (GAJU 160/2010/P, 144/2010/P, and 168/2013/P) and the Institutional grantRVO:60077344. The authors declare no conflicts of interest.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.eco-leng.2014.09.011.
References
Adriano, D.C., Weber, J., Bolan, N.S., Paramasivam, S., Koo, B.J., Sajwan, K.S., 2002.Effects of high rates of coal fly ash on soil, turfgrass, and groundwater quality.Water Air Soil Pollut. 139, 365–385.
Ascensao, F., Clevenger, A.P., Grilo, C., Filipe, J., Santos-Reis, M., 2012. Highway vergesas habitat providers for small mammals in agrosilvopastoral environments.Biodivers. Conserv. 21, 3681–3697.
Ash, H.J., Gemmell, R.P., Bradshaw, A.D., 1994. The introduction of native plantspecies on industrial waste heaps: a test of immigration and other factorsaffecting primary succession. J. Appl. Ecol. 31, 74–84.
Bejcek, V., Stastny, K., 1984. The succession of bird communities on spoil banks aftersurface brown coal mining. Ekol. Polska 32, 245–259.
Benes, J., Kepka, P., Konvicka, M., 2003. Limestone quarries as refuges for Europeanxerophilous butterflies. Conserv. Biol. 17, 1058–1069.
Borm, P.J.A., 1997. Toxicity and occupational health hazards of coal fly ash (CFA). Areview of data and comparison to coal mine dust. Ann. Occup. Hyg. 6, 659–676.
Bryan Jr., A.L., Hopkins, W.A., Parikh, J.H., Jackson, B.P., Unrine, J.M., 2012. Coal fly ashbasins as an attractive nuisance to birds: parental provisioning exposesnestlings to harmful trace elements. Environ. Pollut. 161, 170–177.
Buchar, J., Ruzicka, V., 2002. Catalogue of Spiders of the Czech Republic. Peres,Praque.
Dennis, R.L.H., Shreeve, T.G., Van Dyck, H., 2006. Habitats and resources: the needfor a resource-based definition to conserve butterflies. Biodivers. Conserv. 15,1943–1966.
Dolansky, J., 2003. Spiders of East Bohemian sandy habitats and sliding chalk slopes.Vychodocesky sbornik prirodovedny. Prace studie 10, 285–310.
Dolny, A., Harabis, F., 2012. Underground mining can contribute to freshwaterbiodiversity conservation: allogenic succession forms suitable habitats fordragonflies. Biol. Conserv. 145, 109–117.

52 R. Tropek et al. / Ecological Engineering 73 (2014) 45–52
Eversham, B.C., Roy, D.B., Telfer, M.G., 1996. Urban, industrial and other manmadesites as analogues of natural habitats for Carabidae. Ann. Zool. Fenn. 33,149–156.
Eyre, M.D., Luff, M.L., Woodward, J.C., 2003. Beetles (Coleoptera) on brownfield sitesin England: an important conservation resource? J. Insect Conserv. 7, 223–231.
Fanta, J., Siepel, H., 2010. Inland Drift Sand Landscapes. KNNV Publishing, Zeist.Fry, R., Waring, P., 2001. A guide to moth traps and their use. The Amateur
Entomologists’ Society. Kent.Galan, P., 1997. Colonization of spoil benches of an opencast lignite mine in
Northwest Spain by amphibians and reptiles. Biol. Conserv. 79, 187–195.Gonzalez, A., Navia, R., Moreno, N., 2009. Fly ashes from coal and petroleum coke
combustion: current and innovative potential applications. Waste Manage. Res.27, 976–987.
Haynes, R.J., 2009. Reclamation and revegetation of fly ash disposal sites –challenges and research needs. J. Environ. Manage. 90, 43–53.
Hendrychova, M., Salek, M., Tajovsky, K., Rehor, M., 2012. Soil properties and speciesrichness of invertebrates on afforested sites after brown coal mining. Restor.Ecol. 20, 561–567.
Heneberg, P., Bogusch, P., Rehounek, J., 2013. Sandpits provide critical refuge forbees and wasps (Hymenoptera: Apocrita). J. Insect Conserv. 17, 473–490.
Hodacova, D., Prach, K., 2003. Spoil heaps from brown coal mining: technicalreclamation versus spontaneous revegetation. Restor. Ecol. 11, 385–391.
Hopwood, J.L., 2008. The contribution of roadside grassland restorations to nativebee conservation. Biol. Conserv. 141, 2632–2640.
Jambhulkar, H.P., Juwarkar, A.A., 2009. Assessment of bioaccumulation of heavymetals by different plant species grown on fly ash dump. Ecotoxicol. Environ.Saf. 72, 1122–1128.
Juwarkar, A.A., Jambhulkar, H.P., 2008. Restoration of fly ash dump throughbiological interventions. Environ. Monit. Asses. 139, 355–365.
Krauss, J., Alfert, T., Steffan-Dewenter, I., 2009. Habitat area but not habitat agedetermines wild bee richness in limestone quarries. J. Appl. Ecol. 46, 194–202.
Kubatova, A., Prasil, K., Vanova, M., 2002. Diversity of soil microscopic fungi onabandoned industrial deposits. Cryptogam. Mycol. 23, 205–219.
Lenda, M., Skorka, P., Moron, D., Rosin, Z.M., Tryjanowski, P., 2012. The importance ofthe gravel excavation industry for the conservation of grassland butterflies. Biol.Conserv. 148, 180–190.
Metts, B.S., Buhlmann, K.A., Tuberville, T.D., Scott, D.E., Hopkins, W.A., 2013.Maternal transfer of contaminants and reduced reproductive success ofSouthern Toads (Bufo terrestris) exposed to coal combustion waste. Environ.Sci. Technol. 47, 2846–2853.
Munguira, M.L., Thomas, J.A., 1992. Use of road verges by butterfly and burnetpopulations, and the effect of roads on adult dispersal and mortality. J. Appl.Ecol. 29, 316–329.
New, T.R., 2012. Hymenoptera and Conservation. Willey-Blackwell, Hoboken.Palice, Z., Soldan, Z., 2004. Lichen and bryophyte species diversity on toxic
substrates in the abandoned sedimentation basins of Chvaletice and Bukovina.In: Kovar, P. (Ed.), Natural Recovery of Human-made Deposits in Landscape.Academia, Prague, pp. 200–221.
Pandey, V.C., Singh, B., 2010. Impact of fly ash incorporation in soil systems. Agric.Ecosyst. Environ. 136, 16–27.
Pandey, V.C., Singh, B., 2012. Rehabilitation of coal fly ash basins: current need to useecological engineering. Ecol. Eng. 49, 190–192.
Pandey, V.C., Singh, K., Singh, R.P., Singh, B., 2012. Naturally growing Saccharummunja L. on the fly ash lagoons: a potential ecological engineer for therevegetation and stabilization. Ecol. Eng. 40, 95–99.
Pandey, V.C., Abhilash, P.C., Singh, B., 2009. The Indian perspective of utilizing fly ashin phytoremediation, phytomanagement and biomass production. J. Environ.Manage. 90, 2943–2958.
Prach, K., Hobbs, R.J., 2008. Spontaneous succession versus technical reclamation inthe restoration of disturbed sites. Restor. Ecol. 16, 363–366.
Puntilla, P., Haila, Y., Tukia, H., 1996. Ant communities in taiga clearcuts: habitateffects and species interactions. Ecography 19, 16–28.
Quitt, E., 1971. Klimaticke Oblasti Ceskoslovenska. Academia, Prague.R Development Core Team, 2011. R: A language and environment for statistical
computing. R Foundation for Statistical Computing. Vienna, Austria.http://www.r-project.org/.
Ram, L.C., Masto, R.E., 2010. An appraisal of the potential use of fly ash for reclaimingcoal mine spoil. J. Environ. Manage. 91, 603–617.
Rezac, M., 2004. Spiders and harvestmen (Arachnida: Araneae, Opiliones) on anabandoned ore-washery sedimentation basin near Chvaletice. In: Kovar, P. (Ed.),Natural Recovery of Human-made Deposits in Landscape. Academia,Prague, pp. 311–323.
Rosengren, R., 1986. Competition and coexistence in an insular ant community – amanipulation experiment (Hymenoptera: Formicidae). Ann. Zool. Fennici 23,297–302.
Sajwan, K.S., Punshon, T., Seaman, J.C., 2006. Production of coal combustionproducts and their potential uses. In: Sajwan, K.S., Twardowska, I., Punshon, T.,Alva, A.K. (Eds.), Coal Combustion Byproducts and Environmental Issues.Springer, New York.
Salek, M., 2012. Spontaneous succession on opencast mining sites: implications forbird biodiversity. J. Appl. Ecol. 49, 1417–1425.
Samways, M., McGeoch, M.A., New, T.R., 2009. Insect Conservation: A Handbook ofApproaches and Methods. Oxford University Press, Oxford.
Shaw, P., 2003. Collembola of pulverised fuel ash sites in east London. Eur. J. Soil Biol.39, 1–8.
Shreeve, T.G., Dennis, R.L.H., Van Dyck, H., 2004. Resources, habitats andmetapopulations – whither reality? Oikos 106, 404–408.
Smolova, D., Dolezalova, J., Vojar, J., Solsky, M., Kopecky, O., 2010. Summary offaunistic records and evaluation of amphibian occurrence on spoil banks innorthern Bohemia. Sbornik Severoceskeho Muzea, Prirodni Vedy, Liberec 28,155–163.
Strauss, B., Biedermann, R., 2005. The use of habitat models in conservation of rareand endangered leafhopper species (Hemiptera, Auchenorrhyncha). J. InsectConserv. 9, 245–259.
Tandy, S., Wallace, H.L., Jones, D.L., Nason, M.A., Williamson, J.C., Healey, J.R., 2011.Can a mesotrophic grassland community be restored on a post-industrial sandysite with compost made from waste materials? Biol. Conserv. 144, 500–510.
ter Braak, C.J.F., Smilauer, P., 2002. CANOCO reference manual and CanoDraw forWinndows User’s Guide: Software for Canonical Community Ordination(Version 4.5). Microcomputer Power, Ithaca.
Tomlinson, S., Matthes, U., Richardson, P., Larson, D.W., 2008. The ecologicalequivalence of quarry floors to alvars. Appl. Veg. Sci. 11, 73–82.
Tropek, R., Cerna, I., Straka, J., Cizek, O., Konvicka, M., 2013a. Is coal combustion thelast chance for vanishing insects of inland drift sand dunes in Europe? Biol.Conserv. 162, 60–64.
Tropek, R., Hejda, M., Kadlec, T., Spitzer, L., 2013b. Local and landscape factorsaffecting communities of plants and diurnal Lepidoptera in black coal spoilheaps: implications for restoration management. Ecol. Eng. 57, 252–260.
Tropek, R., Kadlec, T., Hejda, M., Kocarek, P., Skuhrovec, J., Malenovsky, I., Vodka, S.,Spitzer, L., Banar, P., Konvicka, M., 2012. Technical reclamations are wasting theconservation potential of post-mining sites. A case study of black coal spoildumps. Ecol. Eng. 43, 13–18.
Tropek, R., Kadlec, T., Karesova, P., Spitzer, L., Kocarek, P., Malenovsky, P., Banar, P.,Tuf, I.H., Hejda, M., Konvicka, M., 2010. Spontaneous succession in limestonequarries as an effective restoration tool for endangered arthropods and plants. J.Appl. Ecol. 47, 139–147.
Tropek, R., Konvicka, M., 2008. Can quarries supplement rare xeric habitats in apiedmont region? Spiders of the Blansky les Mts. Czech Republic. Land Degrad.Dev. 19, 104–114.
Tropek, R., Konvicka, M., 2011. Should restoration damage rare biotopes? Biol.Conserv. 144, 1299.
Tropek, R., Prach, K., 2012. Mining and post-industrial sites. In: Jongepierova, I.,Prach, K., Jongepier, J.W., Pesout, K. (Eds.), Ecological Restoration in the CzechRepublic. AOPK CR, Prague, pp. 87–93.
Tropek, R., Spitzer, L., Konvicka, M., 2008. Two groups of epigeic arthropods differ incolonising of piedmont quarries: the necessity of multi-taxa and life-historytraits approaches in the monitoring studies. Commun. Ecol. 9, 177–184.
Truxa, C., Fiedler, K., 2012. Attraction to light – from how far do moths (Lepidoptera)return to weak artificial sources of light? Eur. J. Entomol. 109, 77–84.
Tryjanowski, P., Sparks, T.H., Jerzak, L., Rosin, Z.M., Skorka, P., 2014. A paradox forconservation: electricity pylons benefit avian diversity in intensively usedfarmland. Conserv. Lett. 7, 34–40.
Vankova, J., Kovar, P., 2004. Plant species diversity in the biotopes of unreclaimedindustrial deposits as artificial islands in the landscape. In: Kovar, P. (Ed.),Natural Recovery of Human-Made Deposits in Landscape. Academia, Prague, pp.30–45.
Venables, W.N., Ripley, B.D., 2002. Modern Applied Statistics with S. Fourth Edition.New York, Springer.
Ecosystems of disturbed ground. In: Walker, L.R. (Ed.), Ecosystems of the World 16.Elsevier, Amsterdam.
Wheater, C.P., Cullen, W.R., 1997. The flora and invertebrate fauna of abandonedlimestone quarries in Derbyshire. Restor. Ecol. 5, 77–84.
Related Documents











