of May 29, 2022. This information is current as Responses Subsets Impacts HIV Env-Induced Antibody BLyS-Mediated Modulation of Naive B Cell Cancro and Gunilla B. Karlsson Hedestam Karlsson, John R. Mascola, Richard T. Wyatt, Michael P. O'Dell, Emilie K. Grasset, Nadège Pelletier, Mikael C. I. Pia Dosenovic, Martina Soldemo, Jean L. Scholz, Sijy http://www.jimmunol.org/content/188/12/6018 doi: 10.4049/jimmunol.1200466 2012; 2012; 188:6018-6026; Prepublished online 4 May J Immunol Material Supplementary 6.DC1 http://www.jimmunol.org/content/suppl/2012/05/04/jimmunol.120046 References http://www.jimmunol.org/content/188/12/6018.full#ref-list-1 , 28 of which you can access for free at: cites 45 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. All rights reserved. 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on May 29, 2022 http://www.jimmunol.org/ Downloaded from by guest on May 29, 2022 http://www.jimmunol.org/ Downloaded from by guest on May 29, 2022 http://www.jimmunol.org/ Downloaded from by guest on May 29, 2022 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of May 29, 2022.This information is current as
ResponsesSubsets Impacts HIV Env-Induced Antibody BLyS-Mediated Modulation of Naive B Cell
Cancro and Gunilla B. Karlsson HedestamKarlsson, John R. Mascola, Richard T. Wyatt, Michael P.O'Dell, Emilie K. Grasset, Nadège Pelletier, Mikael C. I. Pia Dosenovic, Martina Soldemo, Jean L. Scholz, Sijy
http://www.jimmunol.org/content/188/12/6018doi: 10.4049/jimmunol.12004662012;
2012; 188:6018-6026; Prepublished online 4 MayJ Immunol
MaterialSupplementary
6.DC1http://www.jimmunol.org/content/suppl/2012/05/04/jimmunol.120046
Referenceshttp://www.jimmunol.org/content/188/12/6018.full#ref-list-1
, 28 of which you can access for free at: cites 45 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. All rights reserved.1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
BLyS-Mediated Modulation of Naive B Cell Subsets ImpactsHIV Env-Induced Antibody Responses
Pia Dosenovic,* Martina Soldemo,* Jean L. Scholz,† Sijy O’Dell,‡ Emilie K. Grasset,x
Nadege Pelletier,{ Mikael C. I. Karlsson,x John R. Mascola,‡ Richard T. Wyatt,{
Michael P. Cancro,† and Gunilla B. Karlsson Hedestam*
Neutralizing Abs provide the protective effect of the majority of existing human vaccines. For a prophylactic vaccine against
HIV-1, broadly neutralizing Abs targeting conserved epitopes of the viral envelope glycoproteins (Env) are likely required,
because the pool of circulating HIV-1 variants is extremely diverse. The failure to efficiently induce broadly neutralizing Abs
by vaccination may be due to the use of suboptimal immunogens or immunization regimens, or it may indicate that B cells specific
for broadly neutralizing Env determinants are selected against during peripheral checkpoints, either before or after Ag encounter.
To investigate whether perturbation of B cell subsets prior to immunization with recombinant Env protein affects the vaccine-
induced Ab response in mice, we used B lymphocyte stimulator (BLyS), a cytokine that regulates survival and selection of
peripheral B cells. We show that the transient BLyS treatment used in this study substantially affected naive B cell populations;
in particular, it resulted in more B cells surviving counter-selection at the transitional stages. We also observed more mature naive
B cells, especially marginal zone B cells, in BLyS-treated mice. Intriguingly, provision of excess BLyS prior to immunization led to
a consistent improvement in the frequency and potency of HIV-1 Env vaccine-induced neutralizing Ab responses, without
increasing the number of Env-specific Ab-secreting cells or the Ab-binding titers measured after boosting. The results presented
in this article suggest that an increased understanding of BLyS-regulated processes may help the design of vaccine regimens aimed
at eliciting improved neutralizing Ab responses against HIV-1. The Journal of Immunology, 2012, 188: 6018–6026.
Efforts to elicit broadly neutralizing Abs (bNAbs) againstHIV-1 through envelope glycoprotein (Env) vaccinationhave been unsuccessful, despite robust Ab titers to mul-
tiple epitopes on Env stimulated by current vaccine candidates.Even during chronic HIV-1 infection, bNAbs are elicited in onlya subset of infected individuals, usually only after years of activeviral replication (1). This suggests that effective B cell responsesagainst bNAb epitopes on Env are infrequent and subject to lim-itations imposed by extensive immune-selection pressure for re-sistant isolates during infection. The barriers to achievingappropriate Ab specificity and affinity maturation following vac-
cination are substantial and may be reflective of a variety offactors, including suboptimal presentation of bNAb epitopes oncandidate Env immunogens, insufficient affinity maturation ofcritical Ab specificities, as well as potential limitations in theB cell repertoire caused by events that occur either before or afterB cell exposure to Ag.Developing B cells undergo counter-selection at multiple
checkpoints during maturation, resulting in the loss of mostemerging BCR reactivities. At the transitional developmentalstages (2), about two thirds of newly formed B cells migratingfrom the bone marrow (BM) die before entering mature pre-immune pools. These losses reflect selection based on BCR signalstrength (3, 4) and mediate the elimination of autoreactive andpolyreactive specificities in both mice and humans (5, 6). Ac-cordingly, if clonotypes capable of broadly neutralizing activityagainst HIV-1 are prone to deletion at the transitional stage, theirfrequency in the preimmune repertoire may be low to nil. Indeed,some HIV-1 infection-elicited bNAbs share features with specif-icities prone to elimination during transitional differentiation, suchas long H chain CDR3 (7) or polyspecificity (8, 9). Alternatively,broadly neutralizing clonotypes or their precursors may survive topopulate the preimmune pools and respond to Ag exposure, yetfail to persist as the immune response evolves and peripheraltolerance mechanisms come into play (10). After Ag activationand costimulation, B cells enter the germinal center (GC) reactionwhere novel specificities are generated through somatic hyper-mutation (SHM). Among these newly arising specificities, thosethat most effectively compete for Ag and survival signals selec-tively persist and differentiate into memory and Ab-secretingplasma cells (11). Thus, if bNAb specificities are rarely gener-ated by SHM, or if these clones are poor competitors within theGC, their entrance into memory or Ab-forming pools may occur ata very low frequency.
*Department of Microbiology, Tumor and Cell Biology, Karolinska Institutet, S-17177 Stockholm, Sweden; †Department of Pathology and Laboratory Medicine, Uni-versity of Pennsylvania Perelman School of Medicine, Philadelphia, PA 19104;‡Vaccine Research Center, National Institute of Allergy and Infectious Diseases,National Institutes of Health, Bethesda, MD 20892; xDepartment of Medicine, Kar-olinska Institutet, S-171 77 Stockholm, Sweden; and {Department of Immunologyand Microbial Science, International AIDS Vaccine Initiative Neutralizing AntibodyCenter, The Scripps Research Institute, La Jolla, CA 92037
Received for publication February 7, 2012. Accepted for publication April 10, 2012.
This work was supported by the Swedish Research Council, the Swedish Interna-tional Development Cooperation Agency/Department for Research Cooperation, theKarolinska Institutet (to P.D. and G.B.K.H.), the International AIDS Vaccine Initia-tive (to G.B.K.H. and R.T.W.), and the National Institute of Allergy and InfectiousDiseases, National Institutes of Health for intramural funding to the Vaccine Re-search Center (to J.R.M.) and Grant AI073939S1 (to M.P.C.).
Address correspondence and reprint requests to Prof. Gunilla Karlsson Hedestam,Department of Microbiology, Tumor and Cell Biology, Karolinska Institutet, S-17177 Stockholm, Sweden. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: ALP, alkaline phosphatase; ASC, Ab-secretingcell; BLyS, B lymphocyte stimulator; BM, bone marrow; bNAb, broadly neutraliz-ing Ab; BR3, B lymphocyte stimulator receptor 3; CD4bs, CD4 binding site; Env,envelope glycoprotein; FO, follicular; GC, germinal center; LN, lymph node; MZ,marginal zone; ON, overnight; PC, phosphorylcholine; RT, room temperature; SHM,somatic hypermutation.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1200466
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

The B lineage-specific survival factor, B lymphocyte stimulator(BLyS; also termed BAFF), plays key roles in peripheral B celldevelopment, homeostasis, and selection. Although BLyS bindsthree different receptors, its most profound effects are mediatedby signaling through BLyS receptor 3 (BR3, also termed BAFF-R), which is expressed by transitional, mature naive, and GCB cells (12). There is ample evidence that the BLyS/BR3 axismodulates selection at the transitional stages, because mice andhumans deficient in either BLyS or BR3 show severely compro-mised transitional and mature naive B cell pools (13, 14), whereasBLyS overexpression yields B cell hyperplasia and signs of au-toimmunity (15, 16). Furthermore, in studies using transgenicmice, exogenous BLyS treatment rescues specificities normallylost at the transitional stage, allowing them to enter the maturepreimmune pools (5). There is evidence for a similar role of theBLyS/BR3 axis in GC evolution (17), although the mechanismshave not been determined. For example, GCs can be initiated, butare not sustained, in mice deficient in BLyS (18).In this study, we investigated whether potential counter-selection
of bNAb specificities, particularly at the transitional checkpoint,might be overcome by transient manipulation of BLyS levels. Wetreated mice with BLyS immediately prior to immunization andobserved a marked increase in transitional and mature naive B cellnumbers, indicating a significant increase in cells surviving se-lection at the transitional checkpoint. Upon termination of BLyStreatment, these subsets returned to pretreatment levels in less thana week. When BLyS- and control-treated mice were immunizedwith recombinant Env trimers (19), we found a consistent increasein the frequency of animals displaying HIV-1–neutralizing activityamong the BLyS-treated group. These results intriguingly suggestthat manipulation of selective processes in the peripheral B cellcompartment may be used to qualitatively improve vaccine-elicited neutralizing Ab responses against HIV-1.
Materials and MethodsRecombinant BLyS and HIV-1 Env glycoproteins
Recombinant human BLyS was kindly provided by Human Genome Sciences(Rockville, MD). Recombinant soluble HIV-1 Env gp140 trimers based on theYU2 isolate of HIV-1, gp140-F (19), were produced by transient transfection,as previously described (20), and used for immunizations. In brief, cells weretransfected at a density of 1.1 3 106/ml in GIBCO FreeStyle 293 expressionmedia using 293Fectin, according to the manufacturer’s instructions (Invi-trogen). Supernatants were collected 4 d after transfection. Following col-lection, all supernatants were centrifuged at 3500 3 g to remove cells or celldebris, filtered through a 0.22-mm filter, and supplemented with complete,EDTA-free protease inhibitor mixture (Roche) and penicillin–streptomycin(Invitrogen). Proteins were captured via glycans by lentil–lectin affinitychromatography (GE Healthcare). After extensive washing with PBS, sup-plemented with 0.5M NaCl, the proteins were eluted with 1 M methyl-a-D-mannopyranoside and captured in the second step via the His-tag by nickel-chelation chromatography (GE Healthcare). Following a wash with 40 mMimidazole and 0.5 M NaCl in PBS, proteins were eluted with 300 mM im-idazole in PBS. Biotinylated Env probes used in the B cell ELISPOT assay(21) were produced in a similar fashion.
Treatment of mice and preparation of cells and tissue foranalysis
Adult BALB/c mice (Taconic A/S) were injected i.p. with 10 mgrecombinant BLyS or PBS once daily for 10 d. At the indicated time pointsafter treatment, mice were either sacrificed for analysis or immunized s.c.with 10 mg HIV-1 Env in 10 mg of the adjuvant AbISCO-100 (Isconova).Animals were boosted once or twice with the same regimen 14 d apart. Insome experiments (Fig. 5C, 5D), a third boost was given 60 d after thesecond boost. Animals were sacrificed at 4 or 21 d after the last immu-nization. Single-cell suspensions were prepared from spleen, BM (twofemur and two tibia/mouse), or inguinal lymph node (LN), as described(21). For immunostaining of spleen, tissue was immersed in OptimalCutting Medium (Histolab) and snap-frozen in 2-methyl butane that was
kept cold in liquid nitrogen. All animal experiments were approved bythe Committee for Animal Ethics (Stockholm, Sweden), and performedaccording to specified guidelines.
Flow cytometry
Single-cell suspensions from spleen and LNwere stained with the followingAbs: PerCP anti-B220 (RA3-62B), biotinylated anti-CD23 (B2B4), PE anti-IgM (R6-60.2), PE anti-CD95 (Jo2), allophycocyanin anti-CD8 (53-6.7) allfrom BD Biosciences, as well as allophycocyanin anti-CD93 (AA4.1),Alexa Fluor 488 anti-GL7, PE anti-CD1d (1B1), PE anti-CD3 PE (145-2C11) and FITC anti-CD4 (RM4-4), all from eBiosciences. BiotinylatedAb was visualized with Alexa Fluor 488-conjugated streptavidin (Invi-trogen). Stained cells were fixed in fixation buffer (BD Biosciences) andanalyzed on a FACSCalibur (BD Biosciences).
ELISA
The HIV-1 Env-specific ELISA was performed by coating 96-well ELISAplates (Nunc) with 100 ml soluble HIV-1 Env protein diluted to a concen-tration of 2 mg/ml in PBS. After overnight (ON) incubation at 4˚C, plateswere washed in PBS with 0.05% Tween (wash buffer). The plates were thenblocked for 1.5 h at room temperature (RT) in PBS with 2% dry milk(blocking buffer). Serum was added in fresh blocking buffer and incubated for1.5 h at RT. Plates were washed six times in wash buffer, and the secondaryAb, goat anti-mouse IgG-HRP, was added in wash buffer. Plates were incu-bated for 1.5 h at RT. The plates were washed six times in wash buffer, andthe assay was developed using the SigmaFAST OPD kit (Sigma-Aldrich).The reaction was stopped by the addition of 1 M H2SO4, and the OD wasread at 492 nm using an Asys Expert 96 ELISA reader (Biochrom).
For detection of anti-dsDNAAbs, ELISA plates were precoated with 50 mlmet-BSA (Sigma-Aldrich) diluted to a concentration of 5 mg/ml in PBS andincubated for 6 h at 4˚C. Plates were then washed four times in PBS with0.05% Tween (wash buffer) and coated with 50 ml DNA (Sigma-Aldrich)diluted to a concentration of 50 mg/ml in PBS. For detection of anti-phosphorylcholine (PC) Abs, ELISA plates were coated with 50 ml PC-BSA diluted to a concentration of 2.5 mg/ml in PBS at 4˚C ON. Afterwashing four times in wash buffer, both sets of plates were blocked with PBScontaining 1.5% BSA, 0.1% gelatin, and 3 mM EDTA (blocking buffer) for2 h at RT. Serum was then added in fresh blocking buffer and incubatedfor 2 h at RT prior to washing four times in wash buffer. Secondary Abs, anti-mouse IgM-alkaline phosphatase (ALP) (Mabtech) or anti-mouse IgM-HRP(Southern Biotech), were added in blocking buffer and incubated for 1 h atRT. Depending on the secondary Ab used, the ELISA was developed usingeither SIGMAFAST p-Nitrophenyl phosphate mix (Sigma-Aldrich), and thereaction was stopped by adding 3 N NaOH, or SureBlue TMB MicrowellPeroxidase Substrate (KPL), and the reaction was stopped by adding TMBStop Solution (KPL). The OD was measured at 405 or 450 nm, respectively,using an Asys Expert 96 ELISA reader (Biochrom).
The cardiolipin ELISAwas performed by coating 96-well ELISA plates(Nunc) with 50 ml cardiolipin (Sigma-Aldrich) diluted to a concentrationof 50 mg/ml in 99.5% EtOH. The plates were allowed to evaporate com-pletely at 4˚C ON. The plates were blocked in PBS with 10% FCS(blocking buffer) for 1 h at 37˚C. Serum was added in fresh blocking bufferand incubated for 1 h at 37˚C. The plates were then washed in PBS only.Secondary Ab anti-mouse IgM-HRP (Southern Biotech) was added inblocking buffer, and the plates were incubated for 1 h at 37˚C, washed inPBS only, and developed as described above.
To measure total IgM levels in serum, 96-well ELISA plates (Nunc) werecoated with 100 ml polyclonal goat anti-mouse IgM (Southern Biotech)diluted to a concentration of 2 mg/ml in 0.05 M carbonate buffer. Plateswere incubated ON at 4˚C. Next, ELISA plates were washed six times inPBS with 0.05% Tween (wash buffer) and blocked for 1.5 h in PBS with5% FCS (blocking buffer) at RT. Serum was added to fresh blocking bufferand incubated for 2 h at RT. The plates were washed six times in washbuffer and secondary Ab, anti-mouse IgM-ALP (Mabtech) or anti-mouseIgM-HRP, was added diluted in blocking buffer. After 1.5 h of incubationat RT the plates were washed six times in wash buffer. The assay wasdeveloped as described above.
B cell ELISPOT
Env-specific and total IgG Ab-secreting cells (ASCs) were enumeratedin a B cell ELISPOT assay, as previously described (21). Briefly, 96-wellMultiScreen-IP filter plates (Millipore) were pretreated with 70% ethanoland washed three times in sterile PBS before being coated with 1 mg/well(10 mg/ml) a polyclonal goat anti-mouse IgG Ab (Mabtech). The plateswere incubated ON at 4˚C. Two hours before addition of the cells, theplates were washed five times in sterile PBS and blocked with complete
The Journal of Immunology 6019
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
RPMI 1640 medium at 37˚C for 2 h. Cells were added in duplicates to thewells in three-fold serial dilutions, starting at 106 cells/well. For in vitrostimulation of memory B cells, splenocytes were cultured or not with 2 mg/ml LPS for 6 d. Plates were then wrapped in plastic wrap and incubated for12 h at 37˚C. For detection of spots, the cells were removed by washing theplates six times in PBS with 0.05% Tween-20. Total IgG-secreting cellswere detected with 100 ng/well (1 mg/ml) of a biotinylated polyclonal goatanti-mouse IgG (Mabtech) in blocking buffer (PBS with 1% FCS and0.05% Tween 20). For the detection of Env-specific B cells, 200 ng/well(2 mg/ml) of biotinylated protein (variants of Env or b-galactosidaseas a control) was added diluted in blocking buffer. Biotinylated Ab andprotein were incubated in the plates for 2 h at RT. Plates were then washedsix times in PBS before the addition of 100 ml ALP-conjugated strepta-vidin (Mabtech), diluted 1:1000 in PBS. Plates were incubated for 45 minat RT and then washed six times in water. One hundred microliters ofBCIP/NBT-plus substrate (Mabtech) was then added and incubated for∼10 min at RT. Plates were washed extensively with water and air-dried.Spots were counted in an ImmunoSpot analyzer (Cellular Technology).Memory B cell-derived ASCs were calculated by subtracting the back-ground of non–LPS-stimulated cultures from LPS-stimulated cultures.
Immunohistology and confocal microscopy
Spleens were frozen in OCT medium and cryo-sectioned (6–8 mm) ontoSuperFrost Ultra Plus microscope slides (VWR). Staining was performedwith the following Abs: allophycocyanin anti-B220 (RA3-6B2; eBio-sciences), biotinylated MOMA-1 (Abcam), and Alexa Fluor 488 anti-TcRb (H57-597; BioLegend). Biotinylated Ab was visualized with Cy3-conjugated streptavidin from Jackson ImmunoResearch. Slides were scannedon a Leica TCS SP5X, using a 103/0.30 magnification objective: Leica506505 HCX FLUOTAR. Leica Application Suite Advanced Fluorescencesoftware was used to prepare images, and ImageJ was used for final ad-justment of brightness and contrast.
Neutralization assays
Neutralization assays were performed using a single round of infection ofHIV-1 Env pseudoviruses and TZM-bl target cells, as previously described(22, 23). In total, we had enough sera from individual mice to analyzeneutralization against four tier 1 viruses (MN, HXBc2, SF162, and BaL)and one tier 2 virus (6535). Results are reported as the serum neutralizationID50, the reciprocal of the serum dilution producing 50% virus neutrali-zation without subtracting prebleed values. For competition-neutralizationassays, test or control ligand was added to the serum 30 min prior to theaddition of virus. V3 sp. act. was mapped using a YU2-derived V3 peptideand a scrambled control peptide, as previously described (24). CD4binding site (CD4bs)-directed neutralizing activity was mapped using apair of probes referred to as TriMut and TriMut368/70 (25, 26).
Statistical analysis
Data that passed the D’Agostino and Pearson omnibus normality test wereanalyzed by unpaired, two-tailed Student t tests; data that did not pass wereanalyzed by the Mann–Whitney U test. GraphPad Prism software was usedfor all analyses: data were considered significant at *p# 0.05, **p# 0.01,and ***p # 0.001.
ResultsExogenous BLyS treatment increases primary B cell numbersbut does not alter splenic architecture
To characterize the effect of transient BLyS treatment on B cellcompartments, we injected mice with recombinant BLyS or PBS(controls) daily for 10 d prior to analysis. Consistent with priorreports (27), spleen size and weight increased in BLyS-treatedmice (Fig. 1A), reflecting expanded B cell numbers without sig-nificant effects on T cells (Fig. 1B, left panel). A similar effect onB cell populations was seen in LN (Fig. 1B, right panel). The
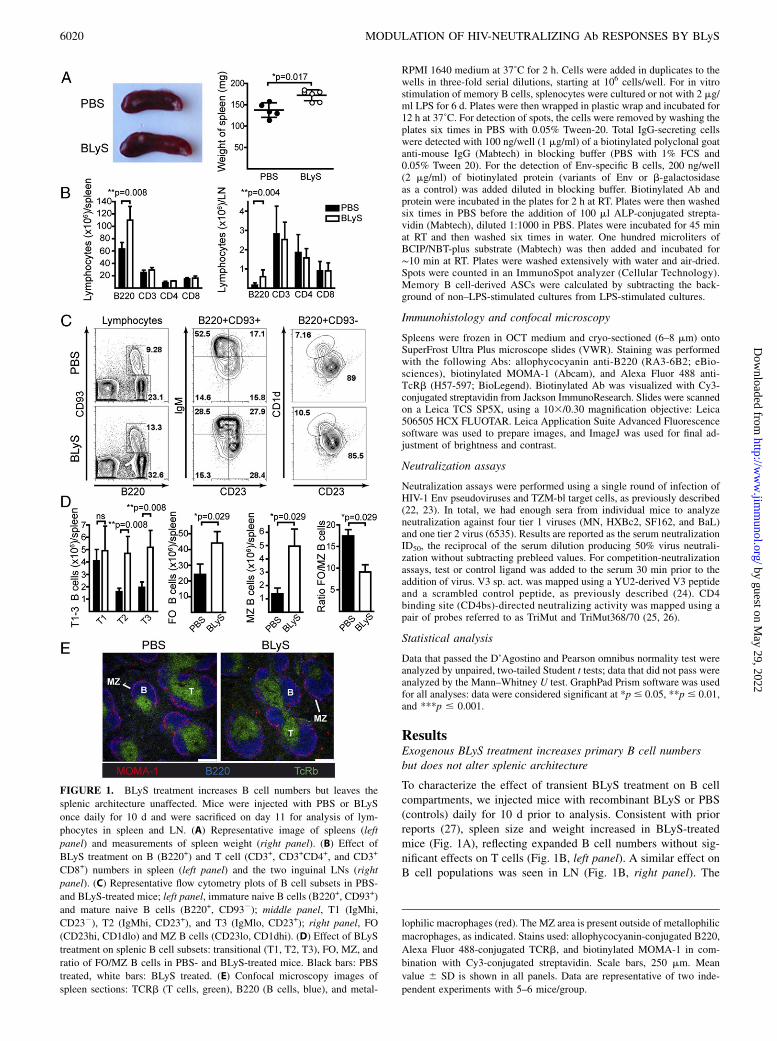
FIGURE 1. BLyS treatment increases B cell numbers but leaves the
splenic architecture unaffected. Mice were injected with PBS or BLyS
once daily for 10 d and were sacrificed on day 11 for analysis of lym-
phocytes in spleen and LN. (A) Representative image of spleens (left
panel) and measurements of spleen weight (right panel). (B) Effect of
BLyS treatment on B (B220+) and T cell (CD3+, CD3+CD4+, and CD3+
CD8+) numbers in spleen (left panel) and the two inguinal LNs (right
panel). (C) Representative flow cytometry plots of B cell subsets in PBS-
and BLyS-treated mice; left panel, immature naive B cells (B220+, CD93+)
and mature naive B cells (B220+, CD932); middle panel, T1 (IgMhi,
CD232), T2 (IgMhi, CD23+), and T3 (IgMlo, CD23+); right panel, FO
(CD23hi, CD1dlo) and MZ B cells (CD23lo, CD1dhi). (D) Effect of BLyS
treatment on splenic B cell subsets: transitional (T1, T2, T3), FO, MZ, and
ratio of FO/MZ B cells in PBS- and BLyS-treated mice. Black bars: PBS
treated, white bars: BLyS treated. (E) Confocal microscopy images of
spleen sections: TCRb (T cells, green), B220 (B cells, blue), and metal-
lophilic macrophages (red). The MZ area is present outside of metallophilic
macrophages, as indicated. Stains used: allophycocyanin-conjugated B220,
Alexa Fluor 488-conjugated TCRb, and biotinylated MOMA-1 in com-
bination with Cy3-conjugated streptavidin. Scale bars, 250 mm. Mean
value 6 SD is shown in all panels. Data are representative of two inde-
pendent experiments with 5–6 mice/group.
6020 MODULATION OF HIV-NEUTRALIZING Ab RESPONSES BY BLyS
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

numbers of transitional (T2 and T3) B cells increased significantlyin BLyS-treated mice, and there was an accumulation of bothfollicular (FO) and marginal zone (MZ) B cells, especially MZB cells, as shown by the decreased FO/MZ B cell ratio (gatedpopulations are shown in Fig. 1C, calculated as absolute numbersin Fig. 1D). Immunofluorescence analysis of frozen spleen sec-tions from PBS- and BLyS-treated mice showed normal splenicarchitecture, suggesting that trafficking and homing of cells werelargely unaffected by BLyS treatment (Fig. 1E).Under conditions in which the BLyS level is permanently in-
creased and the homeostatic regulation of B cells is defective, suchas in BLyS transgenic mice, immature self-reactive B cells surviveperipheral selection checkpoints and progress to mature naive pools(5, 15, 16, 28, 29). To investigate whether BLyS treatment pro-moted survival of autoreactive B cells, we measured Ab titersagainst dsDNA, PC, and cardiolipin. Immediately after cessationof BLyS treatment, we found elevated levels of anti-dsDNA, anti-PC, and anti-cardiolipin Abs (Fig. 2A, left panels). However, thetotal Ab levels at this time point were also increased; when nor-malized to these, there was no difference between BLyS- andcontrol-treated animals (Fig. 2A, right panels). Furthermore, whenanti-dsDNA and anti-PC Ab responses were measured 19 d aftertermination of BLyS treatment, the levels were no longer in-creased in BLyS-treated mice (Fig. 2B). Within 5 d of the lastBLyS treatment, spleen weights (Fig. 3A), B lymphocyte numbers(Fig. 3B), and the number of IgM-secreting cells/spleen (Fig. 3C)also approached pretreatment levels. Accordingly, we chose toinitiate Env immunizations 1 d after the last day of BLyS treat-ment, when peripheral B cells were maximally affected.
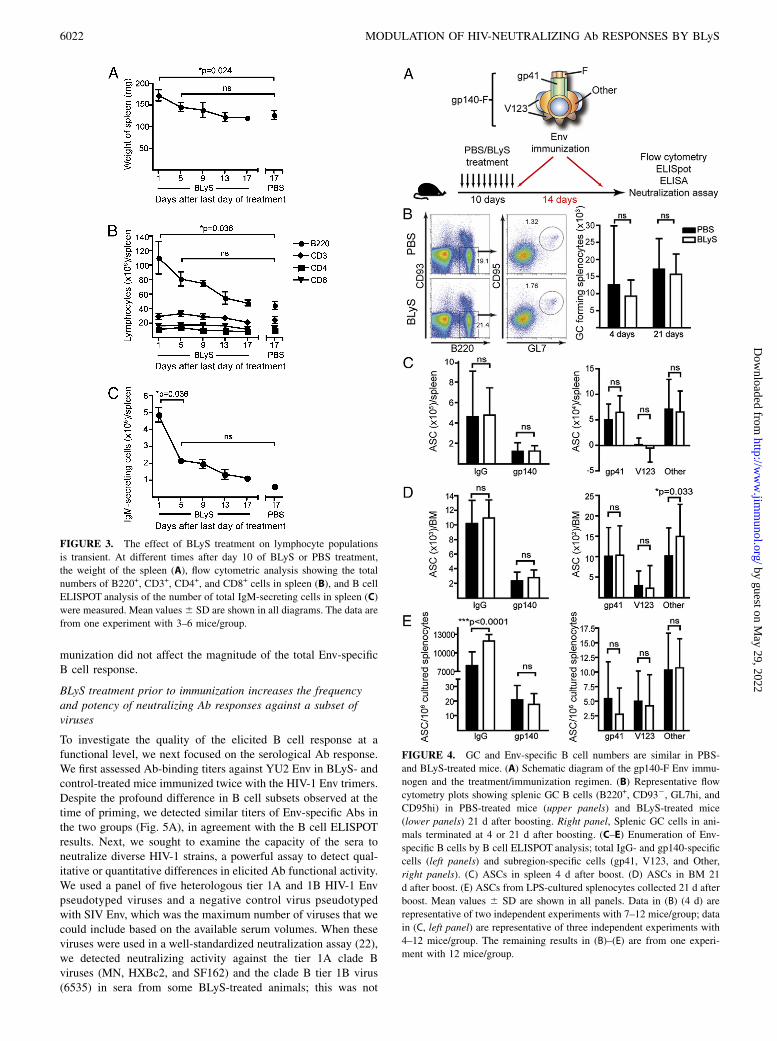
BLyS treatment prior to Env immunization does not affect themagnitude of the Env-specific B cell response
Codelivery of Ag fused to BLyS or the use of BLyS as an adjuvantcomponent administered at the same time as the Ag were reportedpreviously (30–32). In this study, we investigate the magnitude andquality of Env vaccine-elicited B cell responses after BLyS pre-treatment, using HIV-1 gp140-F trimers in adjuvant. The gp140-Ftrimers possess a heterologous trimerization motif (F) appended tothe Env ectodomain (gp140) and are derived from the primary HIV-1 isolate, YU2 (19) (Fig. 4A). Immunized mice were sacrificed foranalysis 4 or 21 d after the boost, and the presence of maturesplenic B cells with a GC phenotype was investigated. We detectedno significant difference in the number of cells with this phenotypebetween BLyS- and control-treated mice at either time point afterthe boost, suggesting that GC formation is not sensitive to BLySlevels (Fig. 4B), consistent with studies in BLyS-deficient mice(18). Furthermore, quantification of the number of ASCs by B cellELISPOT analyses of splenocytes collected 4 d after the boostshowed similar levels of total IgG- and Env-specific IgG-secretingcells in BLyS- and control-treated mice (Fig. 4C, left panel).We next asked whether the response directed against different
subregions of Env differed between BLyS-treated mice and con-trols. This analysis was performed using a differential B cellELISPOT assay that allows the enumeration of B cells specificfor distinct subdeterminants of gp140-F, including gp41, V region1, 2, and 3 (V123), and non-gp41, non-V123 reactivities not clearlydefined and referred to as “Other” (21). We found that a substantialfraction of the response after the boost was directed against gp41,consistent with previous results (21), with no difference betweenthe two groups of mice (Fig. 4C, right panel). On day 21 post-boost, a pool of Env-specific plasma cells is expected to haveaccumulated in the BM. When this population was analyzed, wedetected no marked differences between BLyS-treated mice andcontrols in the number of IgG-secreting cells against total Env or
Env subdeterminants, with the exception of an increase in the non-gp41, non-V123 fraction referred to as “Other” in BLyS-treatedanimals. However, this difference was not consistently observed(Fig. 4D). We also examined the Env-specific B cell memorycompartment after in vitro LPS stimulation of splenocytes col-lected 21 d after the second immunization. We observed an in-crease in the number of memory B cells in BLyS-treated animalscompared with controls but no difference in the number of Env-specific or Env-subspecific memory B cells (Fig. 4E). Taken to-gether, these data demonstrate that BLyS treatment prior to im-
FIGURE 2. Effect of BLyS treatment on anti-dsDNA, anti-PC, and anti-
cardiolipin Ab levels in serum. (A) IgM against dsDNA (upper left panel),
PC (middle left panel), and cardiolipin (lower left panel) in sera collected
1 d after the last day of PBS or BLyS treatment. Ratio of anti-dsDNA IgM
(at serum dilution 1:100), anti-PC IgM (at serum dilution 1:100), and anti-
cardiolipin (at serum dilution 1:50) against total IgM (at serum dilution
1:4050) in serum 1 d after the last day of PBS or BLyS treatment (right
panels). (B) IgM against dsDNA (left panel) or PC (right panel) in sera
collected 19 d after the last day of PBS or BLyS treatment. OD450 values
at a serum dilution of 1:25 are shown. In (A), pooled data from two indi-
vidual experiments are shown, with a total of 9–10 mice/group. In (B), data
from one experiment with 11–12 mice/group are shown. Diagrams show
mean 6 SD. *p # 0.05, **p # 0.01, ***p # 0.001.
The Journal of Immunology 6021
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
munization did not affect the magnitude of the total Env-specificB cell response.
BLyS treatment prior to immunization increases the frequencyand potency of neutralizing Ab responses against a subset ofviruses
To investigate the quality of the elicited B cell response at afunctional level, we next focused on the serological Ab response.We first assessed Ab-binding titers against YU2 Env in BLyS- andcontrol-treated mice immunized twice with the HIV-1 Env trimers.Despite the profound difference in B cell subsets observed at thetime of priming, we detected similar titers of Env-specific Abs inthe two groups (Fig. 5A), in agreement with the B cell ELISPOTresults. Next, we sought to examine the capacity of the sera toneutralize diverse HIV-1 strains, a powerful assay to detect qual-itative or quantitative differences in elicited Ab functional activity.We used a panel of five heterologous tier 1A and 1B HIV-1 Envpseudotyped viruses and a negative control virus pseudotypedwith SIV Env, which was the maximum number of viruses that wecould include based on the available serum volumes. When theseviruses were used in a well-standardized neutralization assay (22),we detected neutralizing activity against the tier 1A clade Bviruses (MN, HXBc2, and SF162) and the clade B tier 1B virus(6535) in sera from some BLyS-treated animals; this was not
FIGURE 3. The effect of BLyS treatment on lymphocyte populations
is transient. At different times after day 10 of BLyS or PBS treatment,
the weight of the spleen (A), flow cytometric analysis showing the total
numbers of B220+, CD3+, CD4+, and CD8+ cells in spleen (B), and B cell
ELISPOT analysis of the number of total IgM-secreting cells in spleen (C)
were measured. Mean values6 SD are shown in all diagrams. The data are
from one experiment with 3–6 mice/group.
FIGURE 4. GC and Env-specific B cell numbers are similar in PBS-
and BLyS-treated mice. (A) Schematic diagram of the gp140-F Env immu-
nogen and the treatment/immunization regimen. (B) Representative flow
cytometry plots showing splenic GC B cells (B220+, CD932, GL7hi, and
CD95hi) in PBS-treated mice (upper panels) and BLyS-treated mice
(lower panels) 21 d after boosting. Right panel, Splenic GC cells in ani-
mals terminated at 4 or 21 d after boosting. (C–E) Enumeration of Env-
specific B cells by B cell ELISPOT analysis; total IgG- and gp140-specific
cells (left panels) and subregion-specific cells (gp41, V123, and Other,
right panels). (C) ASCs in spleen 4 d after boost. (D) ASCs in BM 21
d after boost. (E) ASCs from LPS-cultured splenocytes collected 21 d after
boost. Mean values 6 SD are shown in all panels. Data in (B) (4 d) are
representative of two independent experiments with 7–12 mice/group; data
in (C, left panel) are representative of three independent experiments with
4–12 mice/group. The remaining results in (B)–(E) are from one experi-
ment with 12 mice/group.
6022 MODULATION OF HIV-NEUTRALIZING Ab RESPONSES BY BLyS
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

observed in control-treated mice (Fig. 5B). To confirm this effect,we performed two independent immunization experiments withadditional boosts (3 3 Env and 4 3 Env), but otherwise replicatedthe same conditions that we used in the first experiment (23 Env).Overall, the frequency of mice displaying detectable neutral-ization titers was increased in these repeat experiments comparedwith the first experiment, likely reflecting higher Env-bindingtiters elicited by the additional boosts (Fig. 5C). Consistent withthe first experiment, we saw no difference in total Env-bindingAb titers between BLyS- and control-treated animals, yet theneutralizing activity against MN, HXBc2, SF162, and 6335 wasmore frequent and more potent in BLyS-treated animals; almostall mice displayed neutralizing activity, some at relatively hightiters (Fig. 5D).
When the number of mice responding with positive neutraliz-ing activity against MN, HXBc2, SF162, and 6335 in the control-and BLyS-treated groups was compared, we observed a statisti-cally significant difference (Fig. 5E). We also observed statisti-cally significant differences between the PBS- and BLyS-treatedmice when the average ID50 neutralization titers against MN andSF162, but not against HXBc2 and 6535, were compared (Fig.5F). There was no correlation between YU2 Env-binding titersand virus-neutralizing activity (Supplemental Fig. 1A), suggestingthat the improved neutralizing activity of the serum Abs was notsimply due to an increase in overall Ab titers against the Ag usedfor immunization. Because YU2 Env is heterologous to the virusesused in the neutralization assay, we also measured serum-bindingtiters against recombinant MN and SF162 Env proteins. These
FIGURE 5. Sera from mice immunized twice (23) with Env in adjuvant collected 21 d after the second immunization were analyzed for total Env-
binding Abs. OD50 values are shown for total Env-specific Ab titers. Each point in the ELISA analysis represents data from one mouse and the bar indicates
the mean of all values 6 SD. (B) The sera in (A) were also analyzed for neutralizing activity against a panel of heterologous HIV-1 isolates. Neutralization
data are shown as the reciprocal dilution giving 50% neutralization (ID50 titer). (C) Sera from animals immunized three (33) or four times (43) with Env in
adjuvant collected 4 d after the last immunization were analyzed for Env-binding Abs as in (A). (D) The sera from 3 3 and 4 3 Env-immunized animals
described in (C) were analyzed for neutralization activity in the same way as described in (B). (E) The number of mice with detectable neutralization activity
against MN, HXBc2, SF162, or 6535, combining all experiments (two sequential injections [2 3 Env], three sequential injections [3 3 Env], and four
sequential injections [43 Env] of Env), was plotted and analyzed by a nonparametric Mann–Whitney test. To have an equal number of mice in each group,
one mouse was removed from the 4 3 Env BLyS group. This mouse (number 46) displayed the highest ID50 neutralization titer against several of the
viruses. Thus, by removing this mouse from the comparison, we underestimate rather than overestimate the effect caused by the BLyS treatment. (F) The
potency of neutralization was also analyzed by calculating the average ID50 neutralization titers against MN, HXBc2, SF162, and 6535 among PBS- and
BLyS-treated mice, combining the three experiments (2 3 Env, 3 3 Env, and 4 3 Env). Statistical significance was analyzed using the nonparametric
Mann–Whitney test. The ELISA data from 23 are from one experiment with 12 mice per group; the results from 33 are representative data from two
independent experiments with 5–7 mice per group, and the data from 43 are from one experiment with 4–5 mice per group. The neutralization data are
from one experiment each for the 23 (12 mice per group), 33 (7 mice per group) and 43 (4–5 mice per group) groups.
The Journal of Immunology 6023
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
analyses showed increased binding titers against MN, providinga possible explanation for the improved MN-neutralizing activityin BLyS-treated animals. However, a similar relationship betweenbinding and neutralization was not observed for SF162 (Supple-mental Fig. 1B).The specificity of the HIV-1–neutralizing activity was confirmed
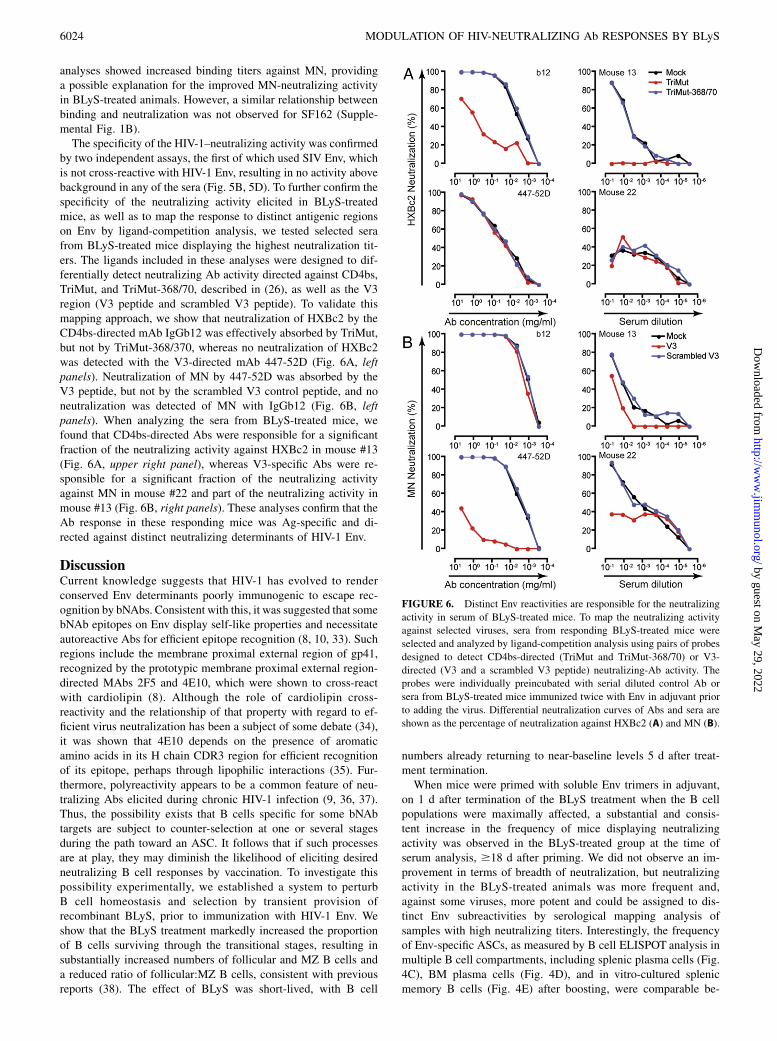
by two independent assays, the first of which used SIV Env, whichis not cross-reactive with HIV-1 Env, resulting in no activity abovebackground in any of the sera (Fig. 5B, 5D). To further confirm thespecificity of the neutralizing activity elicited in BLyS-treatedmice, as well as to map the response to distinct antigenic regionson Env by ligand-competition analysis, we tested selected serafrom BLyS-treated mice displaying the highest neutralization tit-ers. The ligands included in these analyses were designed to dif-ferentially detect neutralizing Ab activity directed against CD4bs,TriMut, and TriMut-368/70, described in (26), as well as the V3region (V3 peptide and scrambled V3 peptide). To validate thismapping approach, we show that neutralization of HXBc2 by theCD4bs-directed mAb IgGb12 was effectively absorbed by TriMut,but not by TriMut-368/370, whereas no neutralization of HXBc2was detected with the V3-directed mAb 447-52D (Fig. 6A, leftpanels). Neutralization of MN by 447-52D was absorbed by theV3 peptide, but not by the scrambled V3 control peptide, and noneutralization was detected of MN with IgGb12 (Fig. 6B, leftpanels). When analyzing the sera from BLyS-treated mice, wefound that CD4bs-directed Abs were responsible for a significantfraction of the neutralizing activity against HXBc2 in mouse #13(Fig. 6A, upper right panel), whereas V3-specific Abs were re-sponsible for a significant fraction of the neutralizing activityagainst MN in mouse #22 and part of the neutralizing activity inmouse #13 (Fig. 6B, right panels). These analyses confirm that theAb response in these responding mice was Ag-specific and di-rected against distinct neutralizing determinants of HIV-1 Env.
DiscussionCurrent knowledge suggests that HIV-1 has evolved to renderconserved Env determinants poorly immunogenic to escape rec-ognition by bNAbs. Consistent with this, it was suggested that somebNAb epitopes on Env display self-like properties and necessitateautoreactive Abs for efficient epitope recognition (8, 10, 33). Suchregions include the membrane proximal external region of gp41,recognized by the prototypic membrane proximal external region-directed MAbs 2F5 and 4E10, which were shown to cross-reactwith cardiolipin (8). Although the role of cardiolipin cross-reactivity and the relationship of that property with regard to ef-ficient virus neutralization has been a subject of some debate (34),it was shown that 4E10 depends on the presence of aromaticamino acids in its H chain CDR3 region for efficient recognitionof its epitope, perhaps through lipophilic interactions (35). Fur-thermore, polyreactivity appears to be a common feature of neu-tralizing Abs elicited during chronic HIV-1 infection (9, 36, 37).Thus, the possibility exists that B cells specific for some bNAbtargets are subject to counter-selection at one or several stagesduring the path toward an ASC. It follows that if such processesare at play, they may diminish the likelihood of eliciting desiredneutralizing B cell responses by vaccination. To investigate thispossibility experimentally, we established a system to perturbB cell homeostasis and selection by transient provision ofrecombinant BLyS, prior to immunization with HIV-1 Env. Weshow that the BLyS treatment markedly increased the proportionof B cells surviving through the transitional stages, resulting insubstantially increased numbers of follicular and MZ B cells anda reduced ratio of follicular:MZ B cells, consistent with previousreports (38). The effect of BLyS was short-lived, with B cell
numbers already returning to near-baseline levels 5 d after treat-ment termination.When mice were primed with soluble Env trimers in adjuvant,
on 1 d after termination of the BLyS treatment when the B cellpopulations were maximally affected, a substantial and consis-tent increase in the frequency of mice displaying neutralizingactivity was observed in the BLyS-treated group at the time ofserum analysis, $18 d after priming. We did not observe an im-provement in terms of breadth of neutralization, but neutralizingactivity in the BLyS-treated animals was more frequent and,against some viruses, more potent and could be assigned to dis-tinct Env subreactivities by serological mapping analysis ofsamples with high neutralizing titers. Interestingly, the frequencyof Env-specific ASCs, as measured by B cell ELISPOT analysis inmultiple B cell compartments, including splenic plasma cells (Fig.4C), BM plasma cells (Fig. 4D), and in vitro-cultured splenicmemory B cells (Fig. 4E) after boosting, were comparable be-
FIGURE 6. Distinct Env reactivities are responsible for the neutralizing
activity in serum of BLyS-treated mice. To map the neutralizing activity
against selected viruses, sera from responding BLyS-treated mice were
selected and analyzed by ligand-competition analysis using pairs of probes
designed to detect CD4bs-directed (TriMut and TriMut-368/70) or V3-
directed (V3 and a scrambled V3 peptide) neutralizing-Ab activity. The
probes were individually preincubated with serial diluted control Ab or
sera from BLyS-treated mice immunized twice with Env in adjuvant prior
to adding the virus. Differential neutralization curves of Abs and sera are
shown as the percentage of neutralization against HXBc2 (A) and MN (B).
6024 MODULATION OF HIV-NEUTRALIZING Ab RESPONSES BY BLyS
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

tween control and BLyS-treated mice. We also did not observea difference between the groups in terms of the total binding Abtiters to the YU2 Env immunogen, suggesting that, despite a largernaive B cell pool at the time of priming, the magnitude of thevaccine-induced response was not augmented. This may be con-sistent with effective feedback mechanisms, such as FCRgII sig-naling, regulating the magnitude of the elicited B cell response.When the relationship between YU2 Env binding and neutralizingactivity was analyzed for individual mice, we observed no cor-relation for any of the viruses against which neutralizing activitywas detected (r2 = 0.20 for MN, r2 = 0.21 for HXBc2, r2 = 0.040for SF162, and r2 = 0.085 for 6435) (Supplemental Fig. 1A). In-stead, several mice displaying high Env-binding titers did notpossess detectable neutralizing activity; conversely, neutralizingactivity was occasionally detected in serum displaying modestEnv-binding titers. When binding titers were measured against thematched Env glycoprotein, improved neutralization of MN couldbe explained by an increased binding titer to the homologous(MN) Env in some mice. However, a similar effect was not ob-served for SF162-directed neutralization, because there was nodifference between BLyS-treated and control mice in terms ofbinding Abs against SF162 Env, and the individual mice in theBLyS group displaying neutralizing activity against SF162 (#19and #22) were not the same as those with the highest binding titersto SF162 Env (Supplemental Fig. 1B). Collectively, this analysissuggests that BLyS pretreatment had a qualitative effect on theEnv-specific B cell response.There are several possible explanations for the observed effects,
because BLyS can affect B cell survival at multiple stages. Thebest-established activity of BLyS is at the transitional checkpoint,where, under normal circumstances, immature B cells compete foravailable BLyS before surviving clones mature and populate theB cell follicles (14, 39). Recent evidence suggests that BLyS alsomodulates GC evolution (reviewed in Ref. 40). In the studiespresented in this article, we believe that it is most likely that BLySimproved survival of B cells prior to BCR-specific activation,thereby affording increased representation of Ag-responsive neu-tralizing clonotypes in BLyS-treated mice. We consider it lesslikely that the effect arose from BLyS modulating the GC reaction,because we only administered BLyS prior to the first Env immu-nization, and the effects of the BLyS treatment that we measuredwere very short-lived. In fact, we limited the time frame duringwhich exogenous BLyS was given to minimize the possibility thatthe exogenous BLyS would affect the fate of B cells after they hadencountered Ag. Thus, the experimental conditions used in thisstudy were chosen to modulate the B cell pools prior to immu-nization rather than to affect processes at later time points. This isa distinctly different approach from that used in a recent study byMelchers et al. (31), in which BLyS (BAFF) was coadministeredwith an HIV-1 Env Ag through plasmid DNA vaccination, thus theeffect of BLyS in that system is expected to be different from theexperimental set-up described in this article.We have not shown directly that naive B cells capable of pro-
ducing neutralizing Abs were rescued by the exogenous BLyStreatment and responded to the vaccination. However, our cellularanalyses strongly suggest that excess BLyS levels substantiallyaffected the composition of transitional and mature naive B cellpools, consistent with an altered naive repertoire. This is re-flected by the disproportionate enlargement of the MZ pool, as wellas the shift in the transitional B cells to a greater proportion andnumber of later transitional subsets (T2 and T3 cells; CD23+
CD93+) in BLyS-treated mice compared with PBS-treated mice.Although we have not analyzed the B cell repertoire at the mo-lecular level, the MZ is well established as a pool that contains
a different repertoire of B cells (41, 42). Furthermore, an en-largement of the late transitional pool indicates a relaxation ofnegative selection that normally occurs at this checkpoint (5, 28,43, 44). Thus, our cellular analysis strongly supports the statementthat we have altered the conditions under which the Env-specificB cell response is elicited, which is reflected in a higher frequencyof mice responding with detectable neutralizing activity and, inseveral cases, higher neutralizing titers (Fig. 4). A molecularcharacterization of the preimmune B cell repertoire in BLyS-treated mice will help to elucidate this question and is the focusof our future efforts. Similarly, an analysis of the Env-specificB cell repertoire in control and BLyS-treated mice is needed tofully dissect the effects observed in this study.It is interesting to note that increased levels of transitional
B cells were reported in individuals chronically infected with HIV-1(45). Along with CD4+ Th cell destruction caused by the infec-tion, HIV-1–infected individuals often present multiple alterationsin peripheral B cell subsets, including decreased stringency atperipheral B cell selection checkpoints, which may be a commonconsequence of chronic immune activation (46). The observationthat a subset of chronically infected individuals develop potentand broad serum-neutralizing activity after several years of in-fection (47–49) suggests that the human immune system is ca-pable of generating such Ab responses. Consistent with this,bNAbs elicited by natural infection display extensive SHM and,when back-mutated to their germ-line sequence, lose broadlyneutralizing activity (49). Thus, activation of rare B cell clonesand successive rounds of GC selection to enhance otherwisepoorly competitive specificities may be required before broadlyneutralizing activity is achieved.In summary, we established a system that can be used to improve
our understanding of HIV-1 Env vaccine-induced B cell responses.We show that manipulation of naive B cell subsets by exogenousBLyS treatment prior to vaccination with recombinant Env trimersin adjuvant resulted in an improvement in the elicited HIV-1–specific neutralizing Ab response. These studies provide a foun-dation for mechanistic investigations of BLyS-regulated peripheralB cell selection processes and their roles for vaccine-inducedneutralizing Ab responses.
AcknowledgmentsWe thank T.-S. Migone and Human Genome Sciences for recombinant
BLyS, M. Forsell, K. Sandgren, and R. Goenka for scientific discussions
and assistance with microscopy, and the personnel at the Department of Mi-
crobiology, Tumor and Cell Biology animal facility at Karolinska Institutet
for excellent technical assistance.
DisclosuresThe authors have no financial conflicts of interest.
References1. Stamatatos, L., L. Morris, D. R. Burton, and J. R. Mascola. 2009. Neutralizing
antibodies generated during natural HIV-1 infection: good news for an HIV-1vaccine? Nat. Med. 15: 866–870.
2. Allman, D., R. C. Lindsley, W. DeMuth, K. Rudd, S. A. Shinton, andR. R. Hardy. 2001. Resolution of three nonproliferative immature splenic B cellsubsets reveals multiple selection points during peripheral B cell maturation. J.Immunol. 167: 6834–6840.
3. Gu, H., D. Tarlinton, W. Muller, K. Rajewsky, and I. Forster. 1991. Most pe-ripheral B cells in mice are ligand selected. J. Exp. Med. 173: 1357–1371.
4. Levine, M. H., A. M. Haberman, D. B. Sant’Angelo, L. G. Hannum,M. P. Cancro, C. A. Janeway, Jr., and M. J. Shlomchik. 2000. A B-cell receptor-specific selection step governs immature to mature B cell differentiation. Proc.Natl. Acad. Sci. USA 97: 2743–2748.
5. Thien, M., T. G. Phan, S. Gardam, M. Amesbury, A. Basten, F. Mackay, andR. Brink. 2004. Excess BAFF rescues self-reactive B cells from peripheral de-letion and allows them to enter forbidden follicular and marginal zone niches.Immunity 20: 785–798.
The Journal of Immunology 6025
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

6. Wardemann, H., S. Yurasov, A. Schaefer, J. W. Young, E. Meffre, andM. C. Nussenzweig. 2003. Predominant autoantibody production by earlyhuman B cell precursors. Science 301: 1374–1377.
7. Saphire, E. O., P. W. Parren, R. Pantophlet, M. B. Zwick, G. M. Morris,P. M. Rudd, R. A. Dwek, R. L. Stanfield, D. R. Burton, and I. A. Wilson. 2001.Crystal structure of a neutralizing human IGG against HIV-1: a template forvaccine design. Science 293: 1155–1159.
8. Haynes, B. F., J. Fleming, E. W. St Clair, H. Katinger, G. Stiegler, R. Kunert,J. Robinson, R. M. Scearce, K. Plonk, H. F. Staats, et al. 2005. Cardiolipinpolyspecific autoreactivity in two broadly neutralizing HIV-1 antibodies. Science308: 1906–1908.
9. Mouquet, H., J. F. Scheid, M. J. Zoller, M. Krogsgaard, R. G. Ott, S. Shukair,M. N. Artyomov, J. Pietzsch, M. Connors, F. Pereyra, et al. 2010. Polyreactivityincreases the apparent affinity of anti-HIV antibodies by heteroligation. Nature467: 591–595.
10. Verkoczy, L., M. Diaz, T. M. Holl, Y. B. Ouyang, H. Bouton-Verville,S. M. Alam, H. X. Liao, G. Kelsoe, and B. F. Haynes. 2010. Autoreactivity in anHIV-1 broadly reactive neutralizing antibody variable region heavy chaininduces immunologic tolerance. Proc. Natl. Acad. Sci. USA 107: 181–186.
11. Gatto, D., and R. Brink. 2010. The germinal center reaction. J. Allergy Clin.Immunol. 126: 898–907; quiz 908–909.
12. Stadanlick, J. E., M. Kaileh, F. G. Karnell, J. L. Scholz, J. P. Miller, W. J. Quinn,III, R. J. Brezski, L. S. Treml, K. A. Jordan, J. G. Monroe, et al. 2008. TonicB cell antigen receptor signals supply an NF-kappaB substrate for prosurvivalBLyS signaling. Nat. Immunol. 9: 1379–1387.
13. Gross, J. A., S. R. Dillon, S. Mudri, J. Johnston, A. Littau, R. Roque, M. Rixon,O. Schou, K. P. Foley, H. Haugen, et al. 2001. TACI-Ig neutralizes moleculescritical for B cell development and autoimmune disease. impaired B cell mat-uration in mice lacking BLyS. Immunity 15: 289–302.
14. Schiemann, B., J. L. Gommerman, K. Vora, T. G. Cachero, S. Shulga-Morskaya,M. Dobles, E. Frew, and M. L. Scott. 2001. An essential role for BAFF in thenormal development of B cells through a BCMA-independent pathway. Science293: 2111–2114.
15. Gross, J. A., J. Johnston, S. Mudri, R. Enselman, S. R. Dillon, K. Madden,W. Xu, J. Parrish-Novak, D. Foster, C. Lofton-Day, et al. 2000. TACI and BCMAare receptors for a TNF homologue implicated in B-cell autoimmune disease.Nature 404: 995–999.
16. Mackay, F., S. A. Woodcock, P. Lawton, C. Ambrose, M. Baetscher,P. Schneider, J. Tschopp, and J. L. Browning. 1999. Mice transgenic for BAFFdevelop lymphocytic disorders along with autoimmune manifestations. J. Exp.Med. 190: 1697–1710.
17. Rahman, Z. S., S. P. Rao, S. L. Kalled, and T. Manser. 2003. Normal inductionbut attenuated progression of germinal center responses in BAFF and BAFF-Rsignaling-deficient mice. J. Exp. Med. 198: 1157–1169.
18. Vora, K. A., L. C. Wang, S. P. Rao, Z. Y. Liu, G. R. Majeau, A. H. Cutler,P. S. Hochman, M. L. Scott, and S. L. Kalled. 2003. Cutting edge: germinalcenters formed in the absence of B cell-activating factor belonging to theTNF family exhibit impaired maturation and function. J. Immunol. 171:547–551.
19. Yang, X., J. Lee, E. M. Mahony, P. D. Kwong, R. Wyatt, and J. Sodroski. 2002.Highly stable trimers formed by human immunodeficiency virus type 1 envelopeglycoproteins fused with the trimeric motif of T4 bacteriophage fibritin. J. Virol.76: 4634–4642.
20. Forsell, M. N., B. Dey, A. Morner, K. Svehla, S. O’dell, C. M. Hogerkorp,G. Voss, R. Thorstensson, G. M. Shaw, J. R. Mascola, et al. 2008. B cell rec-ognition of the conserved HIV-1 co-receptor binding site is altered by endoge-nous primate CD4. PLoS Pathog. 4: e1000171.
21. Dosenovic, P., B. Chakrabarti, M. Soldemo, I. Douagi, M. N. Forsell, Y. Li,A. Phogat, S. Paulie, J. Hoxie, R. T. Wyatt, and G. B. Karlsson Hedestam. 2009.Selective expansion of HIV-1 envelope glycoprotein-specific B cell subsetsrecognizing distinct structural elements following immunization. J. Immunol.183: 3373–3382.
22. Li, M., F. Gao, J. R. Mascola, L. Stamatatos, V. R. Polonis, M. Koutsoukos,G. Voss, P. Goepfert, P. Gilbert, K. M. Greene, et al. 2005. Human immuno-deficiency virus type 1 env clones from acute and early subtype B infections forstandardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 79:10108–10125.
23. Shu, Y., S. Winfrey, Z. Y. Yang, L. Xu, S. S. Rao, I. Srivastava, S. W. Barnett,G. J. Nabel, and J. R. Mascola. 2007. Efficient protein boosting after plasmidDNA or recombinant adenovirus immunization with HIV-1 vaccine constructs.Vaccine 25: 1398–1408.
24. Morner, A., I. Douagi, M. N. Forsell, C. Sundling, P. Dosenovic, S. O’Dell,B. Dey, P. D. Kwong, G. Voss, R. Thorstensson, et al. 2009. Human immuno-deficiency virus type 1 env trimer immunization of macaques and impact ofpriming with viral vector or stabilized core protein. J. Virol. 83: 540–551.
25. Xiang, S. H., P. D. Kwong, R. Gupta, C. D. Rizzuto, D. J. Casper, R. Wyatt,L. Wang, W. A. Hendrickson, M. L. Doyle, and J. Sodroski. 2002. Mutagenicstabilization and/or disruption of a CD4-bound state reveals distinct con-formations of the human immunodeficiency virus type 1 gp120 envelope gly-coprotein. J. Virol. 76: 9888–9899.
26. Feng, Y., K. McKee, K. Tran, S. O’Dell, S. D. Schmidt, A. Phogat, M. N. Forsell,G. B. Karlsson Hedestam, J. R. Mascola, and R. T. Wyatt. 2012. Biochemicallydefined HIV-1 envelope glycoprotein variant immunogens display differential
binding and neutralizing specificities to the CD4-binding site. J. Biol. Chem.287: 5673–5686.
27. Moore, P. A., O. Belvedere, A. Orr, K. Pieri, D. W. LaFleur, P. Feng, D. Soppet,M. Charters, R. Gentz, D. Parmelee, et al. 1999. BLyS: member of the tumornecrosis factor family and B lymphocyte stimulator. Science 285: 260–263.
28. Ota, M., B. H. Duong, A. Torkamani, C. M. Doyle, A. L. Gavin, T. Ota, andD. Nemazee. 2010. Regulation of the B cell receptor repertoire and self-reactivity by BAFF. J. Immunol. 185: 4128–4136.
29. Khare, S. D., I. Sarosi, X. Z. Xia, S. McCabe, K. Miner, I. Solovyev, N. Hawkins,M. Kelley, D. Chang, G. Van, et al. 2000. Severe B cell hyperplasia and autoimmunedisease in TALL-1 transgenic mice. Proc. Natl. Acad. Sci. USA 97: 3370–3375.
30. Gor, D. O., X. Ding, Q. Li, D. Sultana, S. S. Mambula, R. J. Bram, andN. S. Greenspan. 2011. Enhanced immunogenicity of pneumococcal surfaceadhesin A (PsaA) in mice via fusion to recombinant human B lymphocytestimulator (BLyS). Biol. Direct 6: 9.
31. Melchers, M., I. Bontjer, T. Tong, N. P. Chung, P. J. Klasse, D. Eggink,D. C. Montefiori, M. Gentile, A. Cerutti, W. C. Olson, et al. 2012. TargetingHIV-1 envelope glycoprotein trimers to B cells by using APRIL improves an-tibody responses. J. Virol. 86: 2488–2500.
32. Kanagavelu, S. K., V. Snarsky, J. M. Termini, S. Gupta, S. Barzee, J. A. Wright,W. N. Khan, R. S. Kornbluth, and G. W. Stone. 2012. Soluble multi-trimericTNF superfamily ligand adjuvants enhance immune responses to a HIV-1 GagDNA vaccine. Vaccine 30: 691–702.
33. Verkoczy, L., Y. Chen, H. Bouton-Verville, J. Zhang, M. Diaz, J. Hutchinson,Y. B. Ouyang, S. M. Alam, T. M. Holl, K. K. Hwang, et al. 2011. Rescue of HIV-1 broad neutralizing antibody-expressing B cells in 2F5 VH x VL knockin micereveals multiple tolerance controls. J. Immunol. 187: 3785–3797.
34. Scherer, E. M., M. B. Zwick, L. Teyton, and D. R. Burton. 2007. Difficulties ineliciting broadly neutralizing anti-HIV antibodies are not explained by car-diolipin autoreactivity. AIDS 21: 2131–2139.
35. Scherer, E. M., D. P. Leaman, M. B. Zwick, A. J. McMichael, and D. R. Burton.2010. Aromatic residues at the edge of the antibody combining site facilitateviral glycoprotein recognition through membrane interactions. Proc. Natl. Acad.Sci. USA 107: 1529–1534.
36. Liao, H. X., X. Chen, S. Munshaw, R. Zhang, D. J. Marshall, N. Vandergrift,J. F. Whitesides, X. Lu, J. S. Yu, K. K. Hwang, et al. 2011. Initial antibodiesbinding to HIV-1 gp41 in acutely infected subjects are polyreactive and highlymutated. J. Exp. Med. 208: 2237–2249.
37. Morris, L., X. Chen, M. Alam, G. Tomaras, R. Zhang, D. J. Marshall, B. Chen,R. Parks, A. Foulger, F. Jaeger, et al. 2011. Isolation of a human anti-HIV gp41membrane proximal region neutralizing antibody by antigen-specific singleB cell sorting. PLoS ONE 6: e23532.
38. Hsu, B. L., S. M. Harless, R. C. Lindsley, D. M. Hilbert, and M. P. Cancro. 2002.Cutting edge: BLyS enables survival of transitional and mature B cells throughdistinct mediators. J. Immunol. 168: 5993–5996.
39. Batten, M., J. Groom, T. G. Cachero, F. Qian, P. Schneider, J. Tschopp,J. L. Browning, and F. Mackay. 2000. BAFF mediates survival of peripheralimmature B lymphocytes. J. Exp. Med. 192: 1453–1466.
40. Kalled, S. L. 2006. Impact of the BAFF/BR3 axis on B cell survival, germinalcenter maintenance and antibody production. Semin. Immunol. 18: 290–296.
41. Chen, X., F. Martin, K. A. Forbush, R. M. Perlmutter, and J. F. Kearney. 1997.Evidence for selection of a population of multi-reactive B cells into the splenicmarginal zone. Int. Immunol. 9: 27–41.
42. Carey, J. B., C. S. Moffatt-Blue, L. C. Watson, A. L. Gavin, and A. J. Feeney.2008. Repertoire-based selection into the marginal zone compartment duringB cell development. J. Exp. Med. 205: 2043–2052.
43. Lesley, R., Y. Xu, S. L. Kalled, D. M. Hess, S. R. Schwab, H. B. Shu, andJ. G. Cyster. 2004. Reduced competitiveness of autoantigen-engaged B cells dueto increased dependence on BAFF. Immunity 20: 441–453.
44. Thorn, M., R. H. Lewis, A. Mumbey-Wafula, S. Kantrowitz, and L. A. Spatz.2010. BAFF overexpression promotes anti-dsDNA B-cell maturation and anti-body secretion. Cell. Immunol. 261: 9–22.
45. Malaspina, A., S. Moir, J. Ho, W. Wang, M. L. Howell, M. A. O’Shea,G. A. Roby, C. A. Rehm, J. M. Mican, T. W. Chun, and A. S. Fauci. 2006.Appearance of immature/transitional B cells in HIV-infected individuals withadvanced disease: correlation with increased IL-7. Proc. Natl. Acad. Sci. USA103: 2262–2267.
46. Moir, S., A. Malaspina, O. K. Pickeral, E. T. Donoghue, J. Vasquez, N. J. Miller,S. R. Krishnan, M. A. Planta, J. F. Turney, J. S. Justement, et al. 2004. Decreasedsurvival of B cells of HIV-viremic patients mediated by altered expression ofreceptors of the TNF superfamily. J. Exp. Med. 200: 587–599.
47. Walker, L. M., M. Huber, K. J. Doores, E. Falkowska, R. Pejchal, J. P. Julien,S. K. Wang, A. Ramos, P. Y. Chan-Hui, M. Moyle, et al; Protocol G PrincipalInvestigators. 2011. Broad neutralization coverage of HIV by multiple highlypotent antibodies. Nature 477: 466–470.
48. Scheid, J. F., H. Mouquet, B. Ueberheide, R. Diskin, F. Klein, T. Y. Olivera, J.Pietzsch, D. Fenyo, A. Abadir, K. Velinzon, et al. 2011. Sequence and structuralconvergence of broad and potent HIV antibodies that mimic CD4 binding. Sci-ence 333: 1633–1637.
49. Wu, X., Z. Y. Yang, Y. Li, C. M. Hogerkorp, W. R. Schief, M. S. Seaman,T. Zhou, S. D. Schmidt, L. Wu, L. Xu, et al. 2010. Rational design of envelopeidentifies broadly neutralizing human monoclonal antibodies to HIV-1. Science329: 856–861.
6026 MODULATION OF HIV-NEUTRALIZING Ab RESPONSES BY BLyS
by guest on May 29, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from


Supplementary Figure 1. Correlation between total Env binding titers and neutralizing
activity. (A) Correlation between total YU2 Env binding titers and neutralizing activity for
indicated viruses for individual mice presented in Figure 5 A-D. (B) Binding titers against
recombinant MN (left panel) and SF162 (right panel) Env proteins in serum from mice shown
in Figure 5 A and B. Mice showing detectable neutralization activity are indicated with a red
circle. Each point represents data from one mouse and the bar shown in (B) indicates the
mean of all values ±SD.
Related Documents











