ORIGINAL ARTICLE Responses of Benthic Bacteria to Experimental Drying in Sediments from Mediterranean Temporary Rivers Stefano Amalfitano & Stefano Fazi & Annamaria Zoppini & Anna Barra Caracciolo & Paola Grenni & Alberto Puddu Received: 13 September 2006 / Accepted: 24 January 2007 / Published online: 30 June 2007 # Springer Science + Business Media, LLC 2007 Abstract In the semiarid Mediterranean regions, water scarcity represents a common physiological stress for microbial communities residing in river sediments. How- ever, the effect of drying has not yet adequately been evaluated when analyzing riverine microbiological process- es. The bacterial community structure (abundance, biomass, composition) and functioning (carbon production, live cell percentage) were assessed during experimental desiccation in microcosms with sediments from different Mediterranean temporary rivers (Tagliamento, Krathis, Mulargia, Pardiela). Our results showed that the overall responses to drying of the bacterial community were independent from sediment origin and strictly related to water content. During desiccation, a prompt decline (up to 100%) of the initial bacterial carbon production was followed by a slower decrease in abundance and biomass, with an overall reduction of 74% and 78%, respectively. By the end of the experiment, live cells were still abundant but depressed in their main metabolic functions, thus resulting in a drastic increase in the community turnover time. Only 14% of the initial live cell biomass was available in dry sediments to immediately start the reactivation of the aquatic microbial food web after the arrival of new water. Community composition analysis showed a relative increase in alpha- and beta-Proteobacteria, when passing from wet to dry conditions. Our results suggest that the occurrence of drought events could affect carbon cycling through the freshwater microbial compartment, by temporarily limiting microbial mineralization and altering bacterial community structure. Introduction In southern European regions, arid and semiarid ecosystems are under high risk of desertification. The average temper- atures and the duration and frequency of dry periods are expected to rise as a consequence of climate change [29, 35]. Within this scenario the high hydrological variability that already characterizes Mediterranean river catchments [45] is likely to increase. Owing to the heterogeneity of riverine environments (geomorphology, soil properties and composition, rainfall patterns and vegetation coverage, land use and anthropogenic impact), river carbon fluxes are highly variable. Globally, the reported values range around 0.4 Pg C yr −1 for both inorganic and organic carbon (evenly divided between the particulate and dissolved phases) [52]. Although the total amount of C cycling through rivers corresponds to a small fraction of the total C stored in the terrestrial system (∼2000 Pg C), oceans (∼40,000 Pg C) and atmosphere (∼750 Pg C) [28], it is significant if compared to the net oceanic CO 2 uptake (∼2 Pg C yr –1 [9]). Hence, rivers represent a vital link in the biospheric C fluxes contributing to the net exchange of CO 2 between atmo- sphere, land, and water systems [51]. Ultimately, rivers regulate the amount of organic matter that, after escaping decomposition in terrestrial environments, is mineralized and/or exported to recipient water bodies [10]. In the semiarid Mediterranean regions, the seasonal and episodic nature of water flow has significant consequences for river C cycling. During summer, extensive evapotrans- piration, water abstraction, high irrigation demand, and overexploitation of groundwater drastically affect river flow, with extended reaches becoming completely dry [12]. Temporary river sediments are therefore extremely patchy substrates in terms either of space (“hot spots” with high process rates) or of time (“hot moments” with irregular Microb Ecol (2008) 55:270–279 DOI 10.1007/s00248-007-9274-6 S. Amalfitano (*) : S. Fazi : A. Zoppini : A. Barra Caracciolo : P. Grenni : A. Puddu Water Research Institute (IRSA-CNR), Via Reno 1, 00198 Rome, Italy e-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Responses of Benthic Bacteria to Experimental Dryingin Sediments from Mediterranean Temporary Rivers
Stefano Amalfitano & Stefano Fazi &Annamaria Zoppini & Anna Barra Caracciolo &
Paola Grenni & Alberto Puddu
Received: 13 September 2006 /Accepted: 24 January 2007 / Published online: 30 June 2007# Springer Science + Business Media, LLC 2007
Abstract In the semiarid Mediterranean regions, waterscarcity represents a common physiological stress formicrobial communities residing in river sediments. How-ever, the effect of drying has not yet adequately beenevaluated when analyzing riverine microbiological process-es. The bacterial community structure (abundance, biomass,composition) and functioning (carbon production, live cellpercentage) were assessed during experimental desiccationin microcosms with sediments from different Mediterraneantemporary rivers (Tagliamento, Krathis, Mulargia, Pardiela).Our results showed that the overall responses to drying of thebacterial community were independent from sediment originand strictly related to water content. During desiccation, aprompt decline (up to 100%) of the initial bacterial carbonproduction was followed by a slower decrease in abundanceand biomass, with an overall reduction of 74% and 78%,respectively. By the end of the experiment, live cells werestill abundant but depressed in their main metabolicfunctions, thus resulting in a drastic increase in thecommunity turnover time. Only 14% of the initial live cellbiomass was available in dry sediments to immediately startthe reactivation of the aquatic microbial food web after thearrival of new water. Community composition analysisshowed a relative increase in alpha- and beta-Proteobacteria,when passing from wet to dry conditions. Our results suggestthat the occurrence of drought events could affect carboncycling through the freshwater microbial compartment, bytemporarily limiting microbial mineralization and alteringbacterial community structure.
Introduction
In southern European regions, arid and semiarid ecosystemsare under high risk of desertification. The average temper-atures and the duration and frequency of dry periods areexpected to rise as a consequence of climate change [29,35]. Within this scenario the high hydrological variabilitythat already characterizes Mediterranean river catchments[45] is likely to increase. Owing to the heterogeneity ofriverine environments (geomorphology, soil properties andcomposition, rainfall patterns and vegetation coverage, landuse and anthropogenic impact), river carbon fluxes arehighly variable. Globally, the reported values range around0.4 Pg C yr−1 for both inorganic and organic carbon (evenlydivided between the particulate and dissolved phases) [52].Although the total amount of C cycling through riverscorresponds to a small fraction of the total C stored in theterrestrial system (∼2000 Pg C), oceans (∼40,000 Pg C) andatmosphere (∼750 Pg C) [28], it is significant if comparedto the net oceanic CO2 uptake (∼2 Pg C yr–1 [9]). Hence,rivers represent a vital link in the biospheric C fluxescontributing to the net exchange of CO2 between atmo-sphere, land, and water systems [51]. Ultimately, riversregulate the amount of organic matter that, after escapingdecomposition in terrestrial environments, is mineralizedand/or exported to recipient water bodies [10].
In the semiarid Mediterranean regions, the seasonal andepisodic nature of water flow has significant consequencesfor river C cycling. During summer, extensive evapotrans-piration, water abstraction, high irrigation demand, andoverexploitation of groundwater drastically affect riverflow, with extended reaches becoming completely dry[12]. Temporary river sediments are therefore extremelypatchy substrates in terms either of space (“hot spots” withhigh process rates) or of time (“hot moments” with irregular
Microb Ecol (2008) 55:270–279DOI 10.1007/s00248-007-9274-6
S. Amalfitano (*) : S. Fazi :A. Zoppini :A. Barra Caracciolo :P. Grenni :A. PudduWater Research Institute (IRSA-CNR),Via Reno 1,00198 Rome, Italye-mail: [email protected]

periods in which significant biogeochemical events mayoccur) [47].
In such systems, the microbial communities continuous-ly react to microclimate changes with rapid and short-termphysiological adjustments [44]. The benthic heterotrophicbacteria that generally dominate the microbial communitywithin river sediments [19, 33] mainly drive the mineral-ization processes. Water scarcity could affect the rate ofbacterial activity and periods of moisture limitation couldprovoke a strong selective pressure on community struc-ture, modifying resource competition among bacterialtaxonomic subgroups and consequently affecting the uppertrophic levels [24, 60].
Several studies have recently attempted to analyzemicrobial responses to drying or drying–rewetting cyclesin terrestrial soil, by measuring biomass or gross microbialprocesses with chemical determinants or other conventionalmethods (e.g., fumigation–extraction method, fatty acidanalysis) [56, 61]. The widespread opinion is that waterscarcity in soil directly affects cell physiological status andthe drying process causes a decrease in microbial biomassand degradation rates [1]. Moisture limitation imposesphysiological constraints that some bacteria can tolerateby adopting various survival strategies [4, 5]. However,only a few studies have analyzed bacterial physiology,viability, and diversity at the single-cell level under varyingwater regimes [18, 30, 49] and the microbial responses towater stress have not yet adequately been evaluated intemporary river sediments. Consequently, the effect ofdrying on benthic bacterial communities was not taken intoconsideration when analyzing river ecological processes ormodeling water quality in semiarid regions. A deeperunderstanding of microbiological processes is of criticalsignificance in predicting freshwater ecosystem functioningand the C budget in a warmer and drier climate [13].
This study, part of the TempQsim EU research project ontemporary rivers that typify the Mediterranean Region(www.tempqsim.net), was designed to describe benthicbacterial responses to the drying process. In experimentalmicrocosms, we simulated desiccation of sediments col-lected from four European temporary rivers, to describe theeffect of the decreasing moisture on benthic bacterialmetabolism and community structure.
Methods
Sampling Site Description and Sediment Characterization
Across the southern European catchments adopted as studysites by the TempQsim project, we selected four temporaryrivers located in different Mediterranean regions. Theserivers are typically characterized by the intermittency of the
water flow and experience recurrent dry phases of varyingduration and spatial extent, usually between June andSeptember [57].
The Mulargia, a tributary stream of the FlumendosaRiver, is located in the south-eastern part of Sardinia (Italy;39°38′N, 09°11′E). The main reach has a length of 18 km,with a catchment extension of approximately 70 km2. Thestrong seasonality of rainfall events is reflected by theintermittent flow regime of the river, which can becomecompletely dry during summer months. Sediments werecollected in the outlet section upstream of the Mulargiareservoir, as a part of a wider one-year period monitoringactivity with bimonthly sampling.
The River Tagliamento (Italy; 46°01′N, 12°55′E) drainsan area of approximately 2600 km2 and flows for 172 km tothe northern Adriatic Sea. This braided gravel-bed river isconsidered the last morphologically intact river in the Alpsthat has escaped intensive river management. Its hydrologyis characterized by a flashy pluvio-nival flow regime withpeaks in spring and autumn. First- and second-order streamsegments are intermittent and dry at the surface during lowflow periods in winter and summer [58]. Sampling tookplace in an island along a braided reach located in themiddle of the river course.
The River Krathis (Greece; 38°09′N, 27°17′E) is situatedin northern Peloponnesus and flows into the Corinthiangulf. Its length is 30 km and the catchment covers an areaof approximately 150 km2. Urban areas and agriculturalplains characterize the lowland region. Sediment sampleswere collected in the northern part of the river that dries outin late summer–autumn [59].
The River Pardiela (Portugal; 38°26′N, 07°41′W), atemporary tributary of the long transfrontier River Guadi-ana, has a catchment area of approximately 510 km2 andbelongs to the Degebe basin. Like many small streams inthe region, during summer months, there is a transitionfrom lotic to lentic conditions [41] through fragmentationof the flow channel into a series of isolated pools wheresamples were collected.
At the end of May 2004, wet sediments were sampled inthe four rivers from the uppermost oxic layer (0.5–5 cmdepth) of various homogeneous patches and immediatelysieved (2-mm mesh). The <2-mm fraction was stored(∼3 dm3) in plastic buckets and kept refrigerated (4°C)until the beginning of the experiment, which was within10 days of sampling. Immediately after sampling, sedimentaliquots from Mulargia were incubated in situ to estimatebacterial carbon production and fixed to determine cellabundance as described below.
For each sediment sample, the electrical conductivity(EC) was determined in a sediment slurry (10 g of drysediment to 25 mL of filter-sterilized Milli-Q water) aftershaking and allowing the suspension to settle for 30 min.
Responses of Benthic Bacteria to Experimental Drying 271
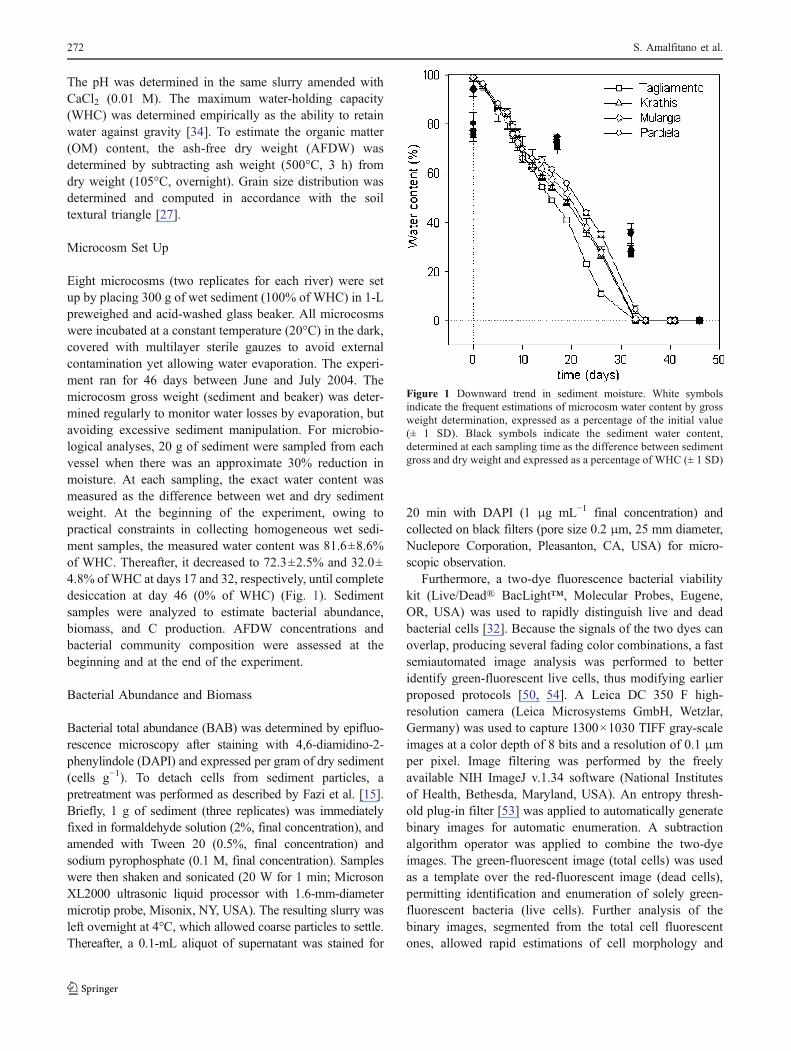
The pH was determined in the same slurry amended withCaCl2 (0.01 M). The maximum water-holding capacity(WHC) was determined empirically as the ability to retainwater against gravity [34]. To estimate the organic matter(OM) content, the ash-free dry weight (AFDW) wasdetermined by subtracting ash weight (500°C, 3 h) fromdry weight (105°C, overnight). Grain size distribution wasdetermined and computed in accordance with the soiltextural triangle [27].
Microcosm Set Up
Eight microcosms (two replicates for each river) were setup by placing 300 g of wet sediment (100% of WHC) in 1-Lpreweighed and acid-washed glass beaker. All microcosmswere incubated at a constant temperature (20°C) in the dark,covered with multilayer sterile gauzes to avoid externalcontamination yet allowing water evaporation. The experi-ment ran for 46 days between June and July 2004. Themicrocosm gross weight (sediment and beaker) was deter-mined regularly to monitor water losses by evaporation, butavoiding excessive sediment manipulation. For microbio-logical analyses, 20 g of sediment were sampled from eachvessel when there was an approximate 30% reduction inmoisture. At each sampling, the exact water content wasmeasured as the difference between wet and dry sedimentweight. At the beginning of the experiment, owing topractical constraints in collecting homogeneous wet sedi-ment samples, the measured water content was 81.6±8.6%of WHC. Thereafter, it decreased to 72.3±2.5% and 32.0±4.8% of WHC at days 17 and 32, respectively, until completedesiccation at day 46 (0% of WHC) (Fig. 1). Sedimentsamples were analyzed to estimate bacterial abundance,biomass, and C production. AFDW concentrations andbacterial community composition were assessed at thebeginning and at the end of the experiment.
Bacterial Abundance and Biomass
Bacterial total abundance (BAB) was determined by epifluo-rescence microscopy after staining with 4,6-diamidino-2-phenylindole (DAPI) and expressed per gram of dry sediment(cells g−1). To detach cells from sediment particles, apretreatment was performed as described by Fazi et al. [15].Briefly, 1 g of sediment (three replicates) was immediatelyfixed in formaldehyde solution (2%, final concentration), andamended with Tween 20 (0.5%, final concentration) andsodium pyrophosphate (0.1 M, final concentration). Sampleswere then shaken and sonicated (20 W for 1 min; MicrosonXL2000 ultrasonic liquid processor with 1.6-mm-diametermicrotip probe, Misonix, NY, USA). The resulting slurry wasleft overnight at 4°C, which allowed coarse particles to settle.Thereafter, a 0.1-mL aliquot of supernatant was stained for
20 min with DAPI (1 μg mL−1 final concentration) andcollected on black filters (pore size 0.2 μm, 25 mm diameter,Nuclepore Corporation, Pleasanton, CA, USA) for micro-scopic observation.
Furthermore, a two-dye fluorescence bacterial viabilitykit (Live/Dead® BacLight™, Molecular Probes, Eugene,OR, USA) was used to rapidly distinguish live and deadbacterial cells [32]. Because the signals of the two dyes canoverlap, producing several fading color combinations, a fastsemiautomated image analysis was performed to betteridentify green-fluorescent live cells, thus modifying earlierproposed protocols [50, 54]. A Leica DC 350 F high-resolution camera (Leica Microsystems GmbH, Wetzlar,Germany) was used to capture 1300×1030 TIFF gray-scaleimages at a color depth of 8 bits and a resolution of 0.1 μmper pixel. Image filtering was performed by the freelyavailable NIH ImageJ v.1.34 software (National Institutesof Health, Bethesda, Maryland, USA). An entropy thresh-old plug-in filter [53] was applied to automatically generatebinary images for automatic enumeration. A subtractionalgorithm operator was applied to combine the two-dyeimages. The green-fluorescent image (total cells) was usedas a template over the red-fluorescent image (dead cells),permitting identification and enumeration of solely green-fluorescent bacteria (live cells). Further analysis of thebinary images, segmented from the total cell fluorescentones, allowed rapid estimations of cell morphology and
Figure 1 Downward trend in sediment moisture. White symbolsindicate the frequent estimations of microcosm water content by grossweight determination, expressed as a percentage of the initial value(± 1 SD). Black symbols indicate the sediment water content,determined at each sampling time as the difference between sedimentgross and dry weight and expressed as a percentage of WHC (± 1 SD)
272 S. Amalfitano et al.

biovolume. Eleven cell morphotypes were automaticallyidentified according to their distinctive shape by theCMEIAS v.1.27 software (Center for Microbial EcologyImage Analysis System [42]). Depending on the observedprevailing cell morphology, the mean cell biovolume (BVin μm3) was calculated from the measured mean cell area,by considering all cells to be spheres and hence applyingthe algorithm proposed by Massana el al. [46]. Thebacterial total biomass (BB in μg C g−1 dry sediment)was calculated by the allometric model (BB = (104.5×BV0.59×0.86) × BAB) proposed by Fischer and Pusch [21].The bacterial total abundance and biomass were multipliedby the percentages of live cells to estimate the live cellabundance and biomass.
Bacterial Carbon Production and Growth Rate
Bacterial carbon production (BCP) was estimated with[3H]leucine incorporation measurements following the meth-od proposed by Buesing and Gessner [6], in combinationwith the microcentrifugation technique proposed for watersamples by Smith and Azam [55] and applied to soil byBååth et al. [2]. Wet sediment (0.5 g, four replicates) wastransferred into 2-mL screw-cap and heat-resistant Sorensenmicrocentrifuge tubes (Sorenson Bioscience, Salt Lake City,UT, USA). Milli-Q water was added to create a final volumeof 1 mL in all tubes. An aqueous solution of radioactiveleucine (50 μM: unlabeled, 49.85 μM final concentration[L 8912, Sigma-Aldrich]; [3H]labeled, 0.15 μM final concen-tration [NEN Life Science Products, Boston, Massachusetts,USA]) was added to three replicates of each sedimentsample. A preliminary test showed that leucine saturationwas reached in all sediment samples at a final concentrationof 50 μM. Zero-time controls were run by killing sampleswith 100% trichloroacetic acid (TCA, 5% final concentra-tion), 15 min before leucine addition. Tubes were homoge-nized by vortexing and incubating (1 h) at 20°C in the dark.Incubations were stopped by adding 100% trichloroaceticacid (TCA). All tubes were centrifuged at 14,000 rcf for10 min at room temperature. The supernatant was discardedto separate macromolecules from the non-incorporated label.Four washing steps were then performed adding 1 mL of 5%TCA, unlabeled leucine solution (40 mM), 80% ethanol, andMilli-Q water, respectively. To extract protein, 1 mL ofNaOH (1 N) was added to the pellet and the tubes wereheated (1 h at 90°C), cooled down, and again centrifuged.The supernatant (0.1 mL) was transferred into a 2-mLEppendorf tube, 1 mL of liquid scintillation cocktail (UltimaGold, Packard Bioscience, Meriden, CT, USA) was addedand radioactivity was detected with the TRICARB 4430(Packard Bioscience) scintillation counter. The rates ofleucine incorporation were converted into units of C persediment dry weight (μg C h−1 g−1) by applying the
conversion factor of 1.44 kg C produced per mole ofincorporated leucine [8]. Considering the exponential growthmodel [39] and the live cell biomass (BBlive), bacterialgrowth rates (μ=ln((BBlive + BCP)/BBlive)/h) and turnovertimes (T2 = (ln2)/μ) were calculated. The per-cell specificproduction (BCPs in fg C h−1 cell−1) was computed bydividing C production by live cell abundance.
Microbial Community Composition
At the beginning and at the end of the experiment,community composition was analyzed by Fluorescence InSitu Hybridization (FISH), in accordance with the protocolof Pernthaler et al. [48]. Before FISH, an additional cellpurification step was performed with the density gradientmedium Nycodenz (Nycomed, Oslo, Norway) to reducehigh-background fluorescence and nonspecific binding ofprobes to nonbacterial particles, as described in Fazi et al.[15]. Purified subsamples (0.5–1 ml) in duplicate for eachmicrocosm were filtered on 0.2 mm polycarbonate mem-branes (47 mm diameter, Nuclepore) by gentle vacuum(<0.2 bar) and washed with 10–20 mL of sterile ultrapurewater. Filter sections were hybridized by the followingoligonucleotide probes: ARCH915 specific for Archea;EUB338, EUB338-II and EUB338-III, targeted to mostBacteria; ALF1b, BET42a, GAM42a specific for the alpha-,beta-, and gamma- Proteobacteria subclasses, respectively;CF319a for Cytophaga-Flavobacterium; PLA46a forPlanctomycetales; HGC69a for Actinobacteria (high GCGram-positive Bacteria); and LGC354abc for Firmicutes(Gram-positive bacteria with low GC content). Furtherdetails on the above-mentioned probes are available atprobeBase [43]. All probes, 5′-labeled with Cy3 dye, werecommercially synthesized (Biomers.net, Ulm, Germany).
Results
Sediment Characterization
The <2-mm sediment fractions were mainly composed ofsand (>80%). Clay was below 2% in all sediments. Silt was12.2% and 7.6% in Tagliamento and Krathis, respectively,whereas in Mulargia and Pardiela it was only 3.0% and1.2%. Sediments from Tagliamento and Krathis wereclassified as loamy sand, Mulargia and Pardiela as coarsesand. The physicochemical and microbiological parametersmeasured in sediments at the beginning of the experimentare summarized in Table 1. Sediment from Pardiela differedfrom the others by having the lowest pH and conductivity,whereas sediment from Mulargia was characterized by thehighest conductivity and ash-free dry weight content. At theend of the experiment, the AFDW concentrations were not
Responses of Benthic Bacteria to Experimental Drying 273

significantly different from the initial values (P>0.05). Themaximum water-holding capacity showed similar values forall sediments. Microbiological characterization showed nomajor differences among the sediments from the four riversin term of bacterial total abundance, biomass, and Cproduction (Student–Newman–Keuls test, P>0.05). In situbacterial abundance, determined in Mulargia sediments atthe same sampling site and date when samples werecollected for microcosm setup, showed values (6.4±0.1×108 cells g−1) not significantly different from those found atthe beginning of the experiment (Student t-test, P<0.05).Moreover, in situ C production (1.04±0.03 μg C h−1 g−1)was statistically different (Student t-test, P>0.05), but stillcomparable to the microcosm initial rate (1.47±0.01 μgC h−1 g−1), when taking into account the wider range ofBCP registered in Mulargia sediments during the annualmonitoring activity (0.6–3.5 μg C h−1 g−1).
During desiccation, water content in all the microcosmsdecreased linearly (white symbols in Fig. 1) without anysignificant difference among the four rivers (analysis ofcovariance (ANOVA), P>0.05). After 35 days of incuba-tion, the amount of water lost by evaporation was no longerdetectable. The direct measurements of the water content bysediment dry weight determination (black symbols inFig. 1) revealed a slower decrease until day 46, when 0%of WHC was reached.
Effect of Drying on Bacterial Abundance and Biomass
Bacterial total abundance was significantly related to thesediment water content, following an exponential pattern(Tagliamento r2=0.79, Krathis r2=0.71, Pardiela r2=0.79,P<0.05), with the exception of Mulargia, where the bestinterpolation was linear (r2=0.85, P<0.05). Taking intoaccount the whole set of data, a significant exponentialrelation was found (y=2.0×108×e0.01x, r2=0.69, P<0.05)(Fig. 2a). On average, total abundance decreased by 74%,passing from 7.7±1.0×108 cells g−1 in the wet sedimentsdown to 2.0±0.7×108 cells g−1 in dry sediments. The
percentage of live cells also decreased from 48.3±3.7% to31.1±6.6% of the total abundance on passing from wet to
Table 1 Physicochemical and microbiological parameters of the sediments from the four rivers at the beginning of the experiment
Tagliamento Krathis Mulargia Pardiela
pH 8.3 8.2 8.1 7.3EC (μS cm−1) 88.5 89.1 104.3 47.7WHC (g g−1 %) 19.5 26.0 21.4 21.1AFDW (mg g−1) 5.9 (0.6) 4.4 (0.7) 9.2 (1.2) 4.5 (0.2)BAB (108 cells g−1) 6.9 (0.30) 8.9 (0.68) 7.0 (0.15) 8.1 (0.42)BB (μg C g−1) 38.8 (0.9) 40.3 (3.6) 35.2 (2.8) 46.0 (3.8)BCP (μg C h−1 g−1) 1.00 (0.01) 1.07 (0.04) 1.47 (0.01) 1.30 (0.06)
The maximum water holding capacity (WHC) is expressed as percentage of the weight of retained water versus dry sediment weight. Ash-free dryweight (AFDW), bacterial total abundance (BAB), biomass (BB), and C production (BCP) are expressed per gram of dry sediment as the averageof three replicates with standard deviation in parentheses.
Figure 2 Total and live cell abundance (a), biomass (b), and bacterialcarbon production (c) versus sediment water content, expressed as apercentage of WHC. Note the x-axis reverse scale: 100% identifies wetsediment at the beginning of the experiment, 0% corresponds to theending dry sediment. All data are normalized to gram of dry sediment.Error bars indicate standard deviations of three independent measure-ments. Regression curves are shown (see text)
274 S. Amalfitano et al.
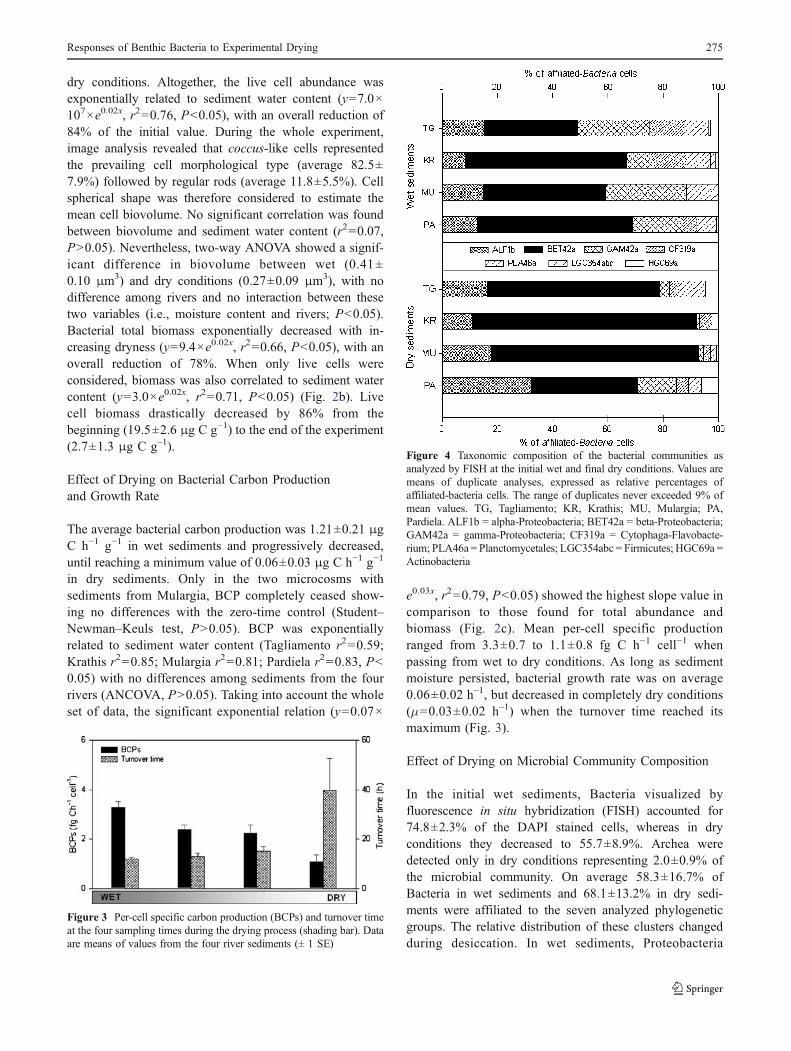
dry conditions. Altogether, the live cell abundance wasexponentially related to sediment water content (y=7.0×107×e0.02x, r2=0.76, P<0.05), with an overall reduction of84% of the initial value. During the whole experiment,image analysis revealed that coccus-like cells representedthe prevailing cell morphological type (average 82.5±7.9%) followed by regular rods (average 11.8±5.5%). Cellspherical shape was therefore considered to estimate themean cell biovolume. No significant correlation was foundbetween biovolume and sediment water content (r2=0.07,P>0.05). Nevertheless, two-way ANOVA showed a signif-icant difference in biovolume between wet (0.41±0.10 μm3) and dry conditions (0.27±0.09 μm3), with nodifference among rivers and no interaction between thesetwo variables (i.e., moisture content and rivers; P<0.05).Bacterial total biomass exponentially decreased with in-creasing dryness (y=9.4×e0.02x, r2=0.66, P<0.05), with anoverall reduction of 78%. When only live cells wereconsidered, biomass was also correlated to sediment watercontent (y=3.0×e0.02x, r2=0.71, P<0.05) (Fig. 2b). Livecell biomass drastically decreased by 86% from thebeginning (19.5±2.6 μg C g−1) to the end of the experiment(2.7±1.3 μg C g–1).
Effect of Drying on Bacterial Carbon Productionand Growth Rate
The average bacterial carbon production was 1.21±0.21 μgC h−1 g−1 in wet sediments and progressively decreased,until reaching a minimum value of 0.06±0.03 μg C h−1 g−1
in dry sediments. Only in the two microcosms withsediments from Mulargia, BCP completely ceased show-ing no differences with the zero-time control (Student–Newman–Keuls test, P>0.05). BCP was exponentiallyrelated to sediment water content (Tagliamento r2=0.59;Krathis r2=0.85; Mulargia r2=0.81; Pardiela r2=0.83, P<0.05) with no differences among sediments from the fourrivers (ANCOVA, P>0.05). Taking into account the wholeset of data, the significant exponential relation (y=0.07×
e0.03x, r2=0.79, P<0.05) showed the highest slope value incomparison to those found for total abundance andbiomass (Fig. 2c). Mean per-cell specific productionranged from 3.3±0.7 to 1.1±0.8 fg C h−1 cell−1 whenpassing from wet to dry conditions. As long as sedimentmoisture persisted, bacterial growth rate was on average0.06±0.02 h–1, but decreased in completely dry conditions(μ=0.03±0.02 h–1) when the turnover time reached itsmaximum (Fig. 3).
Effect of Drying on Microbial Community Composition
In the initial wet sediments, Bacteria visualized byfluorescence in situ hybridization (FISH) accounted for74.8±2.3% of the DAPI stained cells, whereas in dryconditions they decreased to 55.7±8.9%. Archea weredetected only in dry conditions representing 2.0±0.9% ofthe microbial community. On average 58.3±16.7% ofBacteria in wet sediments and 68.1±13.2% in dry sedi-ments were affiliated to the seven analyzed phylogeneticgroups. The relative distribution of these clusters changedduring desiccation. In wet sediments, Proteobacteria
Figure 3 Per-cell specific carbon production (BCPs) and turnover timeat the four sampling times during the drying process (shading bar). Dataare means of values from the four river sediments (± 1 SE)
Figure 4 Taxonomic composition of the bacterial communities asanalyzed by FISH at the initial wet and final dry conditions. Values aremeans of duplicate analyses, expressed as relative percentages ofaffiliated-bacteria cells. The range of duplicates never exceeded 9% ofmean values. TG, Tagliamento; KR, Krathis; MU, Mulargia; PA,Pardiela. ALF1b = alpha-Proteobacteria; BET42a = beta-Proteobacteria;GAM42a = gamma-Proteobacteria; CF319a = Cytophaga-Flavobacte-rium; PLA46a = Planctomycetales; LGC354abc = Firmicutes; HGC69a =Actinobacteria
Responses of Benthic Bacteria to Experimental Drying 275

accounted for 79.8±7.5% of the affiliated cells (alpha−=12.8±3.1%, beta−=48.3±11.3%, and gamma-=20.5±9.2%).Dry sediments from Mulargia, Krathis, and Tagliamentorivers were mostly dominated by beta-Proteobacteria (73.1±9.7%). Only in dry sediments from the River Pardiela,Proteobacteria remained more widely distributed, withalpha-Proteobacteria passing from 13% to 32%, and beta-Proteobacteria decreasing from 56% to 39% of the affiliatedcells (Fig. 4).
Discussion
Bacterial abundance and activity detected in Mulargiasediments were comparable to those found at the beginningof the experiment. Owing to logistic constraints, such directcomparison was not possible for the other three rivers.However, the similarity among sediments in terms ofphysicochemical and microbiological characteristics,allowed us to consider the stored samples from the otherrivers suitable to achieve the aim of this study. In addition,Fletcher et al. [25] did not observe major changes in benthiccommunity in sediments stored over several weeks.
Overall, bacterial total abundance was about one order ofmagnitude lower in comparison with other freshwateraerobic sediments [19, 23], and bacterial C production fellinto the lowest reported range, recalculated on a dry-weightbasis (0.3–30 μg C h−1 g−1 [38], ∼3 μg C h−1 g−1 [23],∼5 μg C h−1 g−1 [7]). Bacterial abundance and activity areassumed to be, at least partially, a function of the sedimentOM content and composition [20, 22]. Hence, the relativelylow values of these parameters could be mainly caused bythe low organic matter in our samples. In this context, thesmall differences in the sediment OM content among thefour rivers were reflected by similar initial bacterialbiomass and activity (Table 1). Moreover, the ratio betweenbacterial C production and total abundance was in line withthose reported for river sediment layers that provide “hotspots” of bacterial activity (0.4–4.4 fg C h−1 cell−1 [22]). Byconsidering the relatively high values of cell biovolume [8],bacterial total biomass was comparable to previous reports(∼50 μg C g−1 dry sediment [21]). Hence, the ratiosbetween C production and total biomass (specific growthrate) also fell within the reported range (0.004–0.3 h−1 [22],0.1 h−1 [7]), indicating that the organic matter wasefficiently utilized in water saturation conditions. Consid-ering a bacterial growth efficiency of 10–40% [38], weestimated that at the beginning of the experiment bacteriawere able to process 70–290 μg C per gram of dry sedimentper day, representing 1–3% of the organic matter of oursamples.
The results clearly showed that bacterial growth andmetabolic rates were dependent on sediment moisture.
During desiccation, a concomitant decreasing trend in theavailability of organic matter and nutrients may occur [3].However, the organic substrate should not represent alimiting factor in our experimental microcosms because itlargely exceeded the estimated bacterial carbon demand.This is also supported by the similar values of AFDWpercentages registered at the beginning and at the end of theexperiment.
When water was below the sediment saturation level, thefirst 30% decrease in water content induced a substantialreduction of bacterial C production (54%), abundance(31%), and consequently biomass (46%). Thereafter, Cproduction exponentially decreased, nearly ceasing in dryconditions, followed by a slower decrease in bacterialabundance and biomass. It cannot be excluded that longerincubation times of the leucine incorporation assay couldlead to a higher estimate of BCP, even in dry sediments.However, the linear relation between leucine incorporationand incubation time is not always guaranteed after 90 min[8], and prolonged incubations could provoke a shift ofbacterial metabolism [6].
Bacterial communities responded to dryness through celldeath and a break in carbon incorporation in the remaininglive cell biomass. Indeed, the removal of water through airdrying is known to be lethal to bacterial cells by damagingmembranes, proteins, and nucleic acids [5]. Because theper-cell production and turnover time did not show majorvariations while sediment moisture persisted, the reductionin community C production was mainly attributable to thedecrease in cell numbers. Conversely, in dry sediments atthe end of the experiment, a conspicuous number of livecells were still abundant, but mostly depressed in their mainmetabolic activity. Therefore, the significant decrease inper-cell production resulted in a drastic increase in thecommunity turnover time (Fig. 3). Our results indicated thatcomplete desiccation of sediments led to a delay inmineralization processes and synthesis of new biomass.The temporarily limited microbial degradation could affectthe accumulation of organic matter. New inundation mightrelease organic matter to downstream reaches without priordecomposition, thus allowing loss of higher quality organicmatter than would occur under constant flow conditions.Nevertheless, bacterial communities continue to be meta-bolically active even in sediment with reduced watercontent. During no-flow periods, when hyporheic waterssustain sediment moisture, microbial degradation may stilloccur and influence the sediment organic matter contentand composition, although with lower rates in comparisonwith those found in water saturation conditions. Theconsiderable loss of microbial organic carbon and thereduction of size of resistant cells during desiccation couldalso have a direct repercussion on the upper trophic levels.As soon as the sediment is rehydrated, only a minor part of
276 S. Amalfitano et al.

the initial live cell biomass (14%) is available to immedi-ately start the reactivation of the aquatic microbial foodweb.
It should be noted that the experiment was carried outunder controlled laboratory conditions and thus not takinginto consideration the combined effect of the dry-heatcondition, as well as the duration and the rate of drying.The combination of these factors could significantlyinfluence bacterial physiology [36]. Moreover, protistanpredation, a major factor in limiting bacterial abundanceand biomass [31], could differentially affect bacterialcommunities in wet and dry sediments because theincreasing sediment aggregate structure during desiccationcould restrict the movement of protists and their grazingefficiency [14].
To the best of our knowledge, the reported FISH resultsare the first attempt to describe bacterial communitycomposition in sediment from temporary rivers, suggestingan inherent resistance to drying of a part of the benthicbacterial communities. Water availability seemed to influ-ence community composition at least at division level, evenif the use of generic probes may hide additional variabilityat finer taxonomic levels.
Both in wet and dry sediments, the percentage of Bacteriacells visualized by EUB338 probes was higher than datareported for other freshwater sediments [15, 26]. However,the reduced percentages of hybridized cells in dry sedimentswere probably caused by the observed decrease in cellactivity. In this respect, recent evidence highlights that thelow ribosome content of slowly growing cells may affect thedetection efficiency of rRNA-targeted probes [16, 63].
Passing from wet to dry conditions, there was an increaseof about 10% in the total Bacteria affiliated to the sevenanalyzed groups, probably indicating a lower occurrence ofother groups in dry sediments. Although all bacterialtaxonomic groups decreased in terms of abundance by morethan 70%, an increase of the relative percentage of Bacteriaaffiliated to beta-Proteobacteria division was observed,except for Pardiela sediments where alpha-Proteobacteriaincreased. The alpha- and beta-Proteobacteria divisionscontain several cosmopolitan freshwater clusters and oftenrepresent the highest proportion of the domain Bacteria infreshwater systems [11, 26, 62, 64]. During the annualmonitoring activity of Mulargia sediments, these twogroups together represented about 50% of the wholecommunity at the end of winter period (March 2004),constantly increasing up to 70% at the end of summer(September 2004). In accordance to our findings, theseclusters appear to dominate the microbial community oftemporary river sediments and they could be considered asprimary targets for finer phylogenetic analyses.
Considering the lag between sediment sampling and thesetting up of the microcosms, our experimental design
could have limitations for community composition analysis,such as the possible growth of microbes quite differentfrom the native assemblage. However, the distribution oftaxa within the source community and the prevailingenvironmental conditions are considered the most relevantfactors in regulating the community composition dynamic[17, 37, 40]. Therefore, the comparison between wet anddry conditions still allowed us to estimate how dryingaffects the bacterial community structure.
Similar bacterial metabolism rates among different riversediments could indicate that drying stress conceivably ledto comparable physiological responses not completelyrelated to the initial community structure. Drying imposesa consistent bottleneck on function and relative composi-tion despite potential differences in the original community.Differently composed communities are able to cope with awide range of environmental conditions, owing to thepresence of generalist species, maintaining similar aggre-gated functions, such as biomass, production, and respira-tion [40]. Alternatively, drying stress severely affectedbacterial metabolism to such an extent as to overcomedifferences in community composition. In conclusion, ourfindings clearly showed a substantial influence of dryingstress on the bacterial community, affecting organic mattermineralization by temporarily limiting metabolic activityand reducing microbial biomass.
Acknowledgments This research was supported by the EuropeanUnion TempQsim Project (EVK1-CT2002-00112). We thank theproject coordinator and partners, in particular Hydrocontrol (Italy),EAWAG (Switzerland), TUC (Greece), and IMAR (Portugal), for theassistance in sampling. Our acknowledgements to Nanna Buesing forwhat we learned about bacterial production determination in sedi-ments, Ilaria Pizzetti for helping in FISH analyses, and FrancescaFalconi for laboratory assistance. We also thank two anonymousreviewers for the valuable suggestions.
References
1. Austin AT, Yahdjian L, Stark JM, Belnap J, Porporato A, NortonU, Ravetta DA, Schaeffer SM (2004) Water pulses and biogeo-chemical cycles in arid and semiarid ecosystems. Oecologia 141:221–235
2. Bååth E, Pettersson M, Söderberg K (2001) Adaptation of a rapidan economical microcentrifugation method to measure thymidineand leucine incorporation by soil bacteria. Soil Biol Biochem 33:1571–1574
3. Baldwin DS, Mitchell AM (2000) The effects of drying and re-flooding on the sediment and soil nutrient dynamics of lowlandriver-floodplain systems: a synthesis. Regulated Rivers: Researchand Management 16: 457–467
4. Bär M, von Hardenberg J, Meron E, Provenzale A (2002)Modeling the survival of bacteria in drylands: the advantage ofbeing dormant. Proc R Soc Lond B 269: 937–942
Responses of Benthic Bacteria to Experimental Drying 277

5. Billi D, Potts M (2002) Life and death of dried prokaryotes. ResMicrobiol 153: 7–12
6. Buesing N, Gessner MO (2003) Incorporation of radiolabeledleucine into protein to estimate bacterial production in plant littersediment epiphytic biofilms and water samples. Microb Ecol 45:291–301
7. Buesing N, Gessner MO (2006) Benthic bacterial and fungalproductivity and carbon turnover in a freshwater marsh. ApplEnviron Microbiol 72: 596–605
8. Buesing N, Marxsen J (2005) Theoretical and empirical conver-sion factors for determining bacterial production in freshwatersediments via leucine incorporation. Limnol Oceanogr 3: 101–107
9. Caldeira K, Akai M (2005) Ocean storage. In: Coninck HC, LoosMA, Metz B, Davidson O, Meyer LA (Eds.) IntergovernmentalPanel on Climate Change Special Report on CarbonDioxide Captureand Storage. Cambridge University Press, Cambridge, pp 277–317
10. Cole JJ, Caraco NF (2001) Carbon in catchments: connectingterrestrial carbon losses with aquatic metabolism. Mar Freshw Res52: 101–110
11. Crump BC, Kling GW, Bahr M, Hobbie JE (2003) Bacterioplanktoncommunity shifts in an arctic lake correlate with seasonal changes inorganic matter source. Appl Environ Microbiol 69: 2253–2268
12. Dahm CN, Baker MA, Moore DI, Thibault JR (2003) Coupledbiogeochemical and hydrological responses of streams and riversto drought. Freshw Biol 48: 1219–1231
13. Darrel Jenerette G, Lal R (2005) Hydrologic sources of carboncycling uncertainty throughout the terrestrial-aquatic continuum.Glob Chang Biol 11: 1873–1882
14. Ekelund F, Frederiksen HB, Rønn R (2002) Population dynamicsof active and total ciliate populations in arable soil amended withwheat. Appl Environ Microbiol 68: 1096–1101
15. Fazi S, Amalfitano S, Pernthaler J, Puddu A (2005) Bacterialcommunities associated with benthic organic matter in headwaterstream microhabitats. Environ Microbiol 7: 1633–1640
16. Fazi S, Amalfitano S, Pizzetti I, Pernthaler J (2007) Efficiency offluorescence in situ hybridization for bacterial cell identification intemporary river sediments with contrasting water content. SystAppl Microbiol (in press). DOI 10.1016/j.syapm.2007.03.003
17. Fenchel T, Finlay BJ (2004) The ubiquity of small species:patterns of local and global diversity. BioScience 54: 777–784
18. Fierer N, Schimel JP, Holden PA (2003) Influence of drying-rewetting frequency on soil bacterial community structure. MicrobEcol 45: 63–71
19. Findlay S, Tank J, Dye S, Valett HM,Mulholland PJ,McDowellWH,Johnson SL, Hamilton SK, Edmonds J, Dodds WK, Bowden WB(2002) A cross-system comparison of bacterial and fungal biomass indetritus pools of headwater streams. Microb Ecol 43: 55–66
20. Findlay SEG, Sinsabaugh RL, Sobczak WV, Hoostal M (2003)Metabolic and structural response of hyporheic microbial com-munities to variations in supply of dissolved organic matter.Limnol Oceanogr 48: 1608–1617
21. Fischer H, Pusch M (2001) Comparison of bacterial production insediments, epiphyton and the pelagic zone of a lowland river.Freshw Biol 46: 1335–1348
22. Fischer H, Sachse A, Steinberg CEW, Pusch M (2002) Differen-tial retention and utilization of dissolved organic carbon bybacteria in river sediments. Limnol Oceanogr 47: 1702–1711
23. Fischer H, Wanner SC, Pusch M (2002) Bacterial abundance andproduction in river sediments as related to the biochemical compositionof particulate organic matter (POM). Biogeochemistry 61: 37–55
24. Flanagan KM, McCauley E, Wrona F (2006) Freshwater foodwebs control carbon dioxide saturation through sedimentation.Glob Chang Biol 12: 644–651
25. Fletcher R., Reynoldson TB, Taylor WD (2001) Maintenance ofintact sediment box cores as laboratory mesocosms. EnvironPollut 115: 183–189
26. Gao X, Olapade AO, Leff LG (2005) Comparison of benthicbacterial community composition in nine streams. Aquat MicrobEcol 40: 51–60
27. Gerakis A, Baer B (1999) A computer program for soil texturalclassification. http://nowlin.css.msu.edu/software/triangle_form.html/. Soil Sci Soc Am J 63: 807–808
28. Grace J (2004) Understanding and managing the global carboncycle. J Ecol 92: 189–202
29. Gregory JM, Mitchell JFB, Brady AJ (1997) Summer drought innorthern mid-latitudes in a time-dependent CO2 climate experi-ment. J Climate 10: 662–686
30. Griffiths RI, Whiteley AS, O'Donnell AG, Bailey MJ (2003)Physiological and community responses of established grasslandbacterial populations to water stress. Appl Environ Microbiol 69:6961–6968
31. Gücker B, Fischer H (2003) Flagellates and ciliates in sedimentsof a lowland river: taxonomy, spatial distribution, and potentialrole in the microbial food web. Aquat Microb Ecol 31: 67–76
32. Haglund AL, Lantz P, Törnblom E, Tranvik L (2003) Depthdistribution of active bacteria and bacterial activity in lakesediment. FEMS Microbiol Ecol 46: 31–38
33. Hall RO, Meyer JL (1998) The trophic significance of bacteria ina detritus-based food web. Ecology 79: 1995–2012
34. Hillel D (1971) Soil and water: physical principles and processes.In: Kozlowski TT (Ed.) Physiological Ecology: A Series ofMonographs, Texts and Treatises, Academic Press, New York, pp288–302
35. IPCC (2001) Climate Change 2001: the scientific basis. Contri-bution of working group to the third assessment report of theintergovernmental panel on climate change. In: Houghton JT,Ding Y, Griggs DJ, Noguer M, van der Linden PJ, Dai X, Maskell K,Johnson CA (Eds.) Cambridge University Press, Cambridge andNew York, pp 1–881
36. Keryn P (2001) Temperature and moisture effects on decompo-sition. In: Proceedings of the workshop on net ecosystemexchange, 18–20 April, Canberra, Australia, pp 95–102
37. Kirchman DL, Dittel AI, Findlay SEG, Fischer D (2004) Changesin bacterial activity and community structure in response todissolved organic matter in the Hudson River, New York. AquatMicrob Ecol 35: 243–257
38. Kirschner AKT, Velimirov B (1999) Benthic bacterial secondaryproduction measured via simultaneous 3H-thymidine and 14C-leucine incorporation, and its implication for the carbon cycle of ashallowmacrophyte dominated backwater system. Limnol Oceanogr44: 1871–1881
39. Koch AL (1994). Growth measurement. In: Gerhardt P, MurrayRGE, Wood WA, Krieg NR (Eds.) Methods for General andMolecular Bacteriology, American Society for Microbiology,Washington, DC, pp 248–277
40. Langenheder S, Lindström ES, Tranvik LJ (2006) Structure andfunction of bacterial communities emerging from different sourcesunder identical conditions. Appl Environ Microbiol 72: 212–220
41. Lillebø AI, Morais M, Guilherme P, Fonseca R, Serafim A, Neves R(2006) Nitrogen and phosphorus dynamics in channel-bedprocesses of temporary streams: a case study in Pardielacatchment (Degebe River, Portugal). Aquatic Conserv: MarFreshw Ecosyst (in press)
42. Liu J, Dazzo FB, Glagoleva O, Yu B, Jain AK (2001) CMEIAS:A computer-aided system for the image analysis of bacterialmorphotypes in microbial communities. http://cme.msu.edu/cmeias/. Microb Ecol 41: 173–194
43. Loy A., HornM,WagnerM (2003) ProbeBase—an online resource forrRNA-targeted oligonucleotide probes. Nucleic Acids Res 31: 514–516
44. Mamilov AS, Dilly OM (2002) Soil microbial eco-physiology asaffected by short-term variations in environmental conditions. SoilBiol Biochem 34: 1283–1290
278 S. Amalfitano et al.

45. Mariotti A, Struglia MV, Zeng N, Lau KM (2002) The hydrologicalcycle in the Mediterranean region and implications for the waterbudget of the Mediterranean Sea. J Climate 15: 1674–1690
46. Massana R, Gasol JM, Bjørnsen PK, Blackburn NT, Hagström Å,Hietanen S, Hygum BH, Kuparinen J, Pedrós-Alió C (1997).Measurement of bacterial size via image analysis of epifluor-escence preparations: description of an inexpensive system andsolutions to some of the most common problems. Sci Mar 61:397–407
47. McClain NJ, Boyer E, Dent L, Gergel S, Grimm N, Groffman P,Hart S, Harvey J, Johnston C, Mayorga E, McDowell W, Pinay G(2003) Biogeochemical hot spots and hot moments at the interfaceof terrestrial and aquatic ecosystems. Ecosystems 6: 301–312
48. Pernthaler J, Glöckner FO, Schönhuber W, Amann R (2001)Fluorescence in situ hybridization (FISH) with rRNA-targetedoligonucleotide probes. Methods Microbiol 30: 207–226
49. Pesaro M, Nicollier G, Zeyer J, Widmer F (2004) Impact of soildrying-rewetting stress on microbial communities and activitiesand on degradation of two crop protection products. Appl EnvironMicrobiol 70: 2577–2587
50. Posch T, Pernthaler J, Alfreider A, Psenner R (1997) Cell-specificrespiratory activity of aquatic bacteria studied with the tetrazoliumreduction method, Cyto-Clear slides, and image analysis. ApplEnviron Microbiol 63: 867–873
51. Raymond PA (2005) Carbon cycle: the age of the Amazon’s breath.Nature 436: 469–470
52. Richey JE (2004) Pathways of atmospheric CO2 through fluvialsystems. In: Fields C (Ed.) Toward CO2 Stabilization Issues,Strategies, and Consequences. A SCOPE GCP Rapid AssessmentProject, Island Press, Washington, DC, pp 329–340
53. Sahoo PK, Soltani S, Wong KC, Chen YC (1988) A survey ofthresholding techniques. Comput Vis Graph Image Process 41: 233–260
54. Shopov A, Williams SC, Verity PG (2000) Improvements inimage analysis and fluorescence microscopy to discriminate and
enumerate bacteria and viruses in aquatic samples. Aquat MicrobEcol 22: 103–110
55. Smith DC, Azam F (1992) A simple, economical method for mea-suring bacterial protein synthesis rates in seawater using 3H-leucine.Mar Microb Food Webs 6: 107–114
56. Steenwerth KL, Jackson LE, Calderón FJ, Stromberg MR, ScowKM (2003) Soil microbial community composition and land usehistory in cultivated and grassland ecosystems of coastal Cal-ifornia. Soil Biol Biochem 35: 489–500
57. TempQsim-Consortium (2006) Critical issues in the water qualitydynamics of temporary waters. Evaluation and recommendationsfrom the TempQsim project. In: Froebrich J, Bauer M (Eds.)Enduser Summary, University of Hannover, Germany, pp 1–69
58. Tockner K, Ward JV, Arscott DB, Edwards PJ, Kollmann J, GurnellAM, Petts GE, Maiolini B (2003) The Tagliamento River: a modelecosystem of European importance. Aquat Sci 65: 239–253
59. Tzoraki O, Amaxidis Y, Skoulikidis N, Íikolaidis N (2004) Instream geochemical processes of temporary rivers—Krathis RiverCase Study. International Conference of Protection and Restora-tion VII of the Environment, June 28, Mykonos, Greece
60. Wardle DA, Verhoef HA, Clarholm M (1998) Trophic relation-ships in the soil microfood-web: predicting the responses to achanging global environment. Glob Chang Biol 4: 713–727
61. Wu J, Brookes PC (2005) The proportional mineralization ofmicrobial biomass and organic matter caused by air-drying andrewetting of a grassland soil. Soil Biol Biochem 37: 507–515
62. Zwart G, Crump BC, Kamst-van Agterveld MP, Hagen F, Han SK(2002) Typical freshwater bacteria: an analysis of available 16SrRNA gene sequences from plankton of lakes and rivers. AquatMicrob Ecol 28: 141–155
63. Zwirglmaier K (2005) Fluorescence in situ hybridisation (FISH)—the next generation. FEMS Microbiol Lett 246: 151–158
64. Zwisler W, Selje N, Simon M (2003) Seasonal patterns of thebacterioplankton community composition in a large mesotrophiclake. Aquat Microb Ecol 31: 211–225
Responses of Benthic Bacteria to Experimental Drying 279
Related Documents











