Response of Cucumber Plants to Low Doses of Different Synthetic Iron Chelates in Hydroponics I ! Journal of Plant Nutrition, 30: 795-809, 2007 Copyright © Taylor & Francis Group, LLC ISSN: 0190-4167 print /1532-4087 online DOl: 10.1080/01904160701290071 Q Taylor & Francis Taylor&.FrancisGroup I Juan J. Lucena 1 and Rufus L. Chaney2 1Dpto Qufmica Agricola, Universidad Aut6noma, Madrid, Spain 2USDA-ARS, Beltsville, Maryland, USA ABSTRACT The factors that control the use of iron (Fe) provided by iron chelates in strategy I plants are not well known. In this paper, the effectiveness of low concentrations of a series of pure Fe chelates to supply Fe to cucumber plants in hydroponics was stud- ied. The Fe Chelate Reductase (FCR) of the roots was measured using Fe-ethylene diamine tetraacetic acid (EDTA) as substrate. Despite the differences found in SPAD and biometric indexes among the treatments, FCR and Fe in xylem sap were only signif- icantly larger for the Fe- Ethylene diarnine di-(o-hydroxy-p-methylphenyl) acetic acid (EDDHMA) treatment. The trend in nutritional indexes was the opposite to the trend in the stability of the chelates, except for Fe-EDTA that gave the poorest results. A mechanism describing the uptake process, considering the re-oxidation of the Fe (II) reduced by the FCR and the formation of the Fe (II) complex is proposed. Keywords: chelates, iron, iron chelate reductase EDDHA, EDTA, EDDHMA, EDDHSA, HEED, reduction assays INTRODUCTION Iron (Fe) chlorosis is a nutritional disorder caused by the low availability of iron in calcareous soils (Marschner and Romheld, 1995). Iron chelates are the most efficient but costly remedy of Fe chlorosis. In fact, in Europe alone more than $60 million are spent every year on Fe chelates to treat Fe chloro- sis (.Alvarez-Fernandez et aI., 2005). The more widely used and more effec- tive Fe chelating agents are the diarnino-diphenolic-dicarboxylic acids, mainly Received 7 March 2005; accepted 27 July 2006. Address correspondence to Juan J. Lucena, Dpto Qufmica Agricola., Universidad Aut6noma, 28049 Madrid, Spain. E-mail: [email protected] 795

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Response of Cucumber Plants to Low Doses ofDifferent Synthetic Iron Chelates in Hydroponics
I!
Journal ofPlant Nutrition, 30: 795-809, 2007
Copyright © Taylor & Francis Group, LLC
ISSN: 0190-4167 print /1532-4087 online
DOl: 10.1080/01904160701290071
Q Taylor & Francis~ Taylor&.FrancisGroup
I
Juan J. Lucena1 and Rufus L. Chaney2
1Dpto Qufmica Agricola, Universidad Aut6noma, Madrid, Spain2USDA-ARS, Beltsville, Maryland, USA
ABSTRACT
The factors that control the use of iron (Fe) provided by iron chelates in strategy Iplants are not well known. In this paper, the effectiveness of low concentrations of aseries of pure Fe chelates to supply Fe to cucumber plants in hydroponics was studied. The Fe Chelate Reductase (FCR) of the roots was measured using Fe-ethylenediamine tetraacetic acid (EDTA) as substrate. Despite the differences found in SPADand biometric indexes among the treatments, FCR and Fe in xylem sap were only significantly larger for the Fe- Ethylene diarnine di-(o-hydroxy-p-methylphenyl) acetic acid(EDDHMA) treatment. The trend in nutritional indexes was the opposite to the trendin the stability of the chelates, except for Fe-EDTA that gave the poorest results. Amechanism describing the uptake process, considering the re-oxidation of the Fe (II)reduced by the FCR and the formation of the Fe (II) complex is proposed.
Keywords: chelates, iron, iron chelate reductase EDDHA, EDTA, EDDHMA,EDDHSA, HEED, reduction assays
INTRODUCTION
Iron (Fe) chlorosis is a nutritional disorder caused by the low availability ofiron in calcareous soils (Marschner and Romheld, 1995). Iron chelates arethe most efficient but costly remedy of Fe chlorosis. In fact, in Europe alonemore than $60 million are spent every year on Fe chelates to treat Fe chlorosis (.Alvarez-Fernandez et aI., 2005). The more widely used and more effective Fe chelating agents are the diarnino-diphenolic-dicarboxylic acids, mainly
Received 7 March 2005; accepted 27 July 2006.Address correspondence to Juan J. Lucena, Dpto Qufmica Agricola., Universidad
Aut6noma, 28049 Madrid, Spain. E-mail: [email protected]
795

796 J. J. Lucena and R. L. Chaney
ethylene diamine di-(o-hydroxy phenyl) acetic acid (EDDHA) and ethylene diamine di-(o-hydroxy-p-methyl phenyl) acetic acid (EDDHMA). However, dueto their high prices only cash crops may be treated with such chelates, whichleave some areas and crops with low productivity. Improving the efficacy of ironchelates should reduce the application rates and hence the cost of the treatments.
It has been hypothesized (Lindsay, 1995; Lucena, 2003) that Fe chelatesserve as carriers of native Fe in soil to the plant roots. This mechanism may bedescribed in four steps. First, the Fe(III) bonded to the chelating agent must bereleased. The uptake of the iron depends on the type of plants and the chelateused. Strategy I plants reduce the iron chelate by a reductase (Chaney et aI. 1972;Bienfait, 1985) and the Fe(II) formed is transported into the roots by a plasmamembrane transporter (Fox et aI., 1996). The rate of reduction and the uptake bythe plant depends on the chelate (Lucena and Chaney, 2006). In the second step,the free chelating agent moves from the root surface into the soil solution by adiffusion concentration gradient and then can react with the Fe present in thesoil. The chelating agent may also react with other metals, with protons or evenmay be retained by the soil surfaces. Only the most Fe specific chelating agentsas the diamino-diphenolic-dicarboxylic acids are able to complex iron in theappropriate soil conditions (Lindsay, 1979; Norvell, 1991; Lucena et aI., 1998).In the third step, the chelating agent binds native iron dissolved from the solidphases. This is a kinetically controlled process that depends on the chelatingagent (Stone, 1997), the nature of the solid phase (Schwertmann, 1991; PerezSanz and Lucena, 1995; Nowack and Sigg, 1997) and the soil conditions. Therate of dissolution may be the limiting factor for the whole process. It has beenhypothesized that kinetics, rather than equilibrium, controls the speciation oflow stability complexes (such as the phytosiderophores) (Stone, 1997). Finally,the iron chelate must return to the root surface, by diffusion or convection. Onits way to the plant the chelate may be retained by the soil surfaces, retardingits movement to the plant (Hermindez-Apaolaza and Lucena, 2001). In summary, the agronomic efficacy of a Fe chelate depends on the use that the plantcan make of the Fe provided by the chelates as Fe substrates, the competitionwith other cations, the solubilization of Fe hydroxides and sorption on soil surfaces. The first process, the plant-chelate interaction, is the focus of the presentpaper.
In a previous work (Lucena and Chaney, 2006) the role of the Fe(III)chelates as substrates of the Fe-chelate reductase (FCR) in strategy I plants wasexamined. It was concluded that a high stability of the chelate decreases theFe reduction by the FCR in mildly chlorotic cucumber plants, but the relationbetween the reductase activity and the efficacy to provide Fe to plants was notdemonstrated. The main objective of this work is to study the effectivenessof pure Fe chelates in providing iron to cucumber plants in hydroponics. Thechelates used were low concentrations of Fe-ethylene diamine tetraacetic acid(EDTA), Fe-EDDHA, Fe-meso-EDDHA, Fe-racemic-EDDHA, Fe-EDDHMA,Fe-propylene diamine di-(o-hydroxyphenyl)acetic acid (PDDHA), Fe-ethylene

r\
~ Response of Cucumber Plants to Iron Chelates 797
diamine di-(2-hydroxy-5-sulphonylphenyl) acetic acid (EDDHSA), and FeN, N'-di(2-hydroxy benzyl)ethylene diamine-N, N'-diacetic acid (HBED).
MATERIALS AND METHODS
Plant Culture
Cucumber seeds (Cucumis sativus L. cv. Burpless, Meyer Seed Co., Lot 115947)were germinated in standard germination papers as in Lucena and Chaney, 2006,for 5 d. Uniform seedlings were selected. The stems of two individual plantswere wrapped together with polyurethane foam, and placed in a hole in a blackPlexiglas lid (three holes per lid). Three pair of seedlings per pot were grown in 2L vessels with pH 7.5 [0.1 M 4-(2-Hydroxyethyl)piperazine-l-ethanesulfonicacid (HEPES) + 2 g calcium carbonate (CaC03)] aerated nutrient solutioncontaining macros: 1 mM calcium nitrate [Ca(N03h], 0.9 mM potassiumnitrate (KN03), 0.3 mM magnesium sulfate (MgS04), 0.1 mM monopotassium phosphate (KH2P04); cationic micronutrients: 1.0 J,l,M manganese sulfate (MnS04), 0.5 J,l,M copper sulfate (CUS04), 0.5 J,l,M zinc sulfate (ZnS04),0.1 J,l,M cobalt sulfate (COS04), 0.1 J,l,M nickel sulfate (NiS04); anionic micronutrients: 35 J,l,M sodium chloride (NaCl), 10 J,l,M boric acid (H3B03), 0.05J,l,M sodium molybdate (Na2Mo04) and 5 J,l,M Fe added as the following treatments: Fe-EDTA, Fe-EDDHA, Fe-meso-EDDHA, Fe-racemic-EDDHA, FeEDDHMA, Fe-PDDHA, Fe-EDDHSA and Fe-HBED prepared as in Lucenaand Chaney (2006). Four replicate pots were prepared for each treatment.
Plants were grown in a growth chamber with cool-white fluorescent andincandescent lamps. The lights provided 200 J,l,E m-2 at plant height during16 h per day. The air temperature was 30125°C (day/night)
Water was added every two days. Eight d after transplanting, nutrient solutions were completely renewed. Two pairs of plants per pot were used on the17th day (13 d after transplanting) for the reduction assays (RA) and xylem sapcollection. The remaining pairs of plants per pot were allowed to grow in thesame nutrient solution (renewed again the day after the RA) for 11 more days(28 d after the beginning of the experiment) and then roots and shoots werewashed with deionized water, weighed and dried. Micronutrient content wasassessed after dry digestion procedure and Atomic Absorption determination.
During the experiment, readings with a chlorophyll-meter (SPAD)(Minolta, 1989), based on light transmission, were taken for all the leaves(average of three readings per leaf) at several times. SPAD readings were compared among treatments for each leaf stage (1st leaf and the oldest ones), butonly data from the leaves that were completing their development were considered. Young leaves may differ in area and degree of development so variabilityis large and SPAD results may not be representative. Readings for very smallleaves normally give relatively high (green) readings not related with the Festatus. Older leaves are also a poor material for Fe nutrition assessment.

798 J. J. Lucena and R. L. Chaney
Reduction Assays (RA) and Xylem Sap Collection
The root iron chelate reductase (FCR) activity of intact plants was estimated from the formation ofthe Fe(II)-bathophenantroline disulfonate complex(Fe(II)-BPDS3) from Fe(III)-EDTA and bathophenantroline disulfonic disodium salt (BPDS). One-hundred-twenty-five rnL disposable steriline beakersinserted in darkened boxes and with black lids were placed in the growth chamber. Each beaker contained 100 rnL RA solution consisting of macronutrient solution as in the growth period, 100 JLM Fe-EDTA chelate, 2 mM 2morpholinoethanesulfonic acid (MES) or 2 mM HEPES to buffer the pH at 6 or7.5 respectively, and 300 JLM BPDS as Fe2+ trapping and coloring reagent. Thelids had a hole for a plastic air dispersion tube, another for a pipette and a thirdhole to hold the pair of cucumber plants held by polyurethane foam. Each solution was continuously aerated and allowed to reach temperature equilibriumbefore transferring plants. Experiments started after 3 h into the daylight period.Initial 3 rnL samples were obtained for each beaker. The roots of plants werewashed three times in macronutrient solution containing 37.5 JLM Na2BPDS,and then transferred to the RA solutions. Three rnL samples were withdrawn at5, 10,20,60, and 120 m. Four replicates were prepared for each treatment andpH of the RA solution. Also 2 beakers without plants were included in order tocorrect reduction rates for slow photoreduction. '
When the RA finished, stems were excised 2 em over the root, these rootsplaced in RA solution without BPDS and the xylem sap pouring from theexcised plant collected during two hours following the procedure described inLucena and Chaney (2006). The fresh weight of blotted roots was obtained atthe end of the xylem sap collection assay. Also shoot dry weight and sap weightwere measured.
Statistics
All the results were subject to statistical analysis using program SAS. Statisticalcomparison of means have been carried out to reveal the differences amongtreatments and pH of the RA solutions, using Duncan's Multiple Range Test(a = 0.05).
RESULTS
FCR Activity
Figure 1presents the FCR activity ofthe plants grown with low doses ofdifferentFe chelates. Statistical comparison of the means using the Duncan Test (a =0.05) revealed significant differences between pH (pH = 6: a, pH = 7.5: b)and among treatments (Fe-EDDHMA: a, Fe-EDTA: ab, Fe-EDDHA: b, Femeso-EDDHA: b, Fe-racemic-EDDHA: b, Fe-EDDHSA: b, Fe-PDDHA: b,
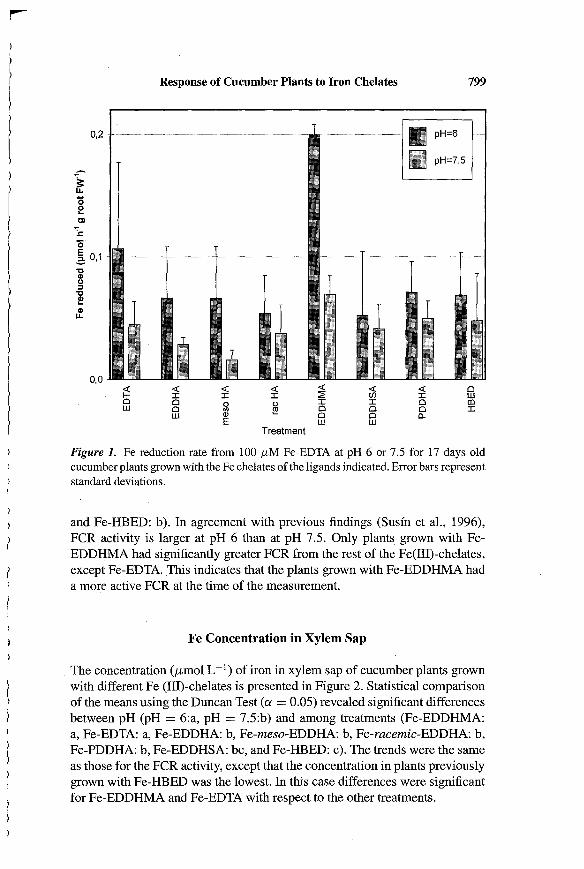
Response of Cucumber Plants to Iron Chelates 799
0,2 +.__ _ _ _ . ........._ 1. pH=6
pH=7.5
:c"0E2:: 0,1"0Gl(J:::l"0eGlu.
0,0
~ « « « « « « ClI I I :2: (/) I W
Cl Cl 0 u I I Cl IIIW Cl f/) ~ Cl Cl Cl I
W ()) Cl Cl 0-E w w
Treatment
Figure 1. Fe reduction rate from 100 p,M Fe-EDTA at pH 6 or 7.5 for 17 days oldcucumber plants grown with the Fe chelates of the ligands indicated. Error bars representstandard deviations.
and Fe-HBED: b). In agreement with previous findings (Susfn et aI., 1996),FCR activity is larger at pH 6 than at pH 7.5. Only plants grown with FeEDDHMA had significantly greater FCR from the rest of the Fe(III)-chelates,except Fe-EDTA.This indicates that the plants grown with Fe-EDDHMA hada more active FCR at the time of the measurement.
Fe Concentration in Xylem Sap
The concentration (p,mol L-1) of iron in xylem sap of cucumber plants grownwith different Fe (III)-chelates is presented in Figure 2. Statistical comparisonof the means using the Duncan Test (a = 0.05) revealed significant differencesbetween pH (pH = 6:a, pH = 7.5:b) and among treatments (Fe-EDDHMA:a, Fe-EDTA: a, Fe-EDDHA: b, Fe-meso-EDDHA: b, Fe-racemic-EDDHA: b,Fe-PDDHA: b, Fe-EDDHSA: be, and Fe-HBED: c). The trends were the sameas those for the FCR activity, except that the concentration in plants previouslygrown with Fe-HBED was the lowest. In this case differences were significantfor Fe-EDDHMA and Fe-EDTA with respect to the other treatments.
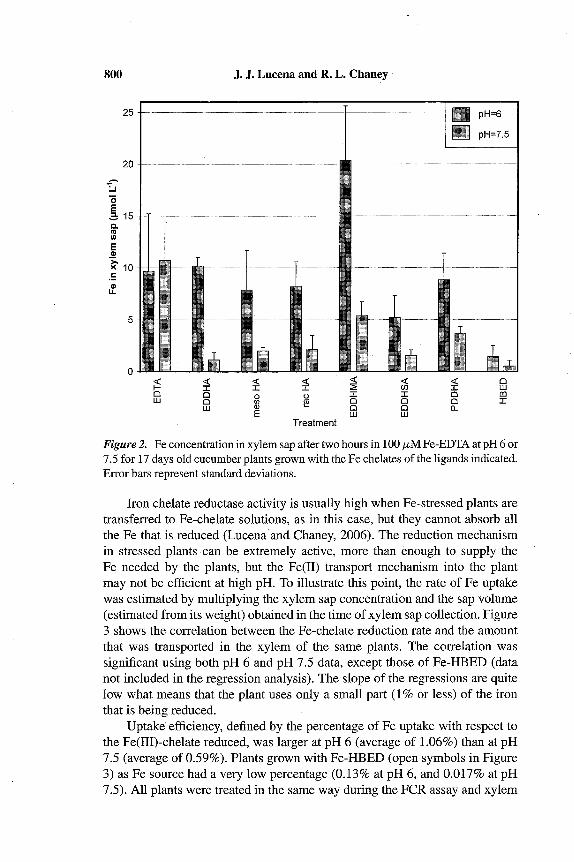
800 J. J. Lucena and R. L. Chaney
25 ----------------------------------- ---------------- • pH=6
pH=7.5
20 -------------------------------------
~(5E2: 15 +T--·················································· ---
Co
'"UI
Ea>
~ 10.5a>u.
5
0
~ « « « « « « 0:I: :I: :I: :2: (J) :I: W
0 0 0 <> :I: :I: 0 caw 0 I/l ~ 0 0 0 :I:
W (l) 0 0 0-E w w
Treatment
Figure 2. Fe concentration in xy!em sap after two hours in 100 fLM Fe-EDTA at pH 6 or7.5 for 17 days old cucumber plants grown with the Fe chelates of the ligands indicated.Error bars represent standard deviations.
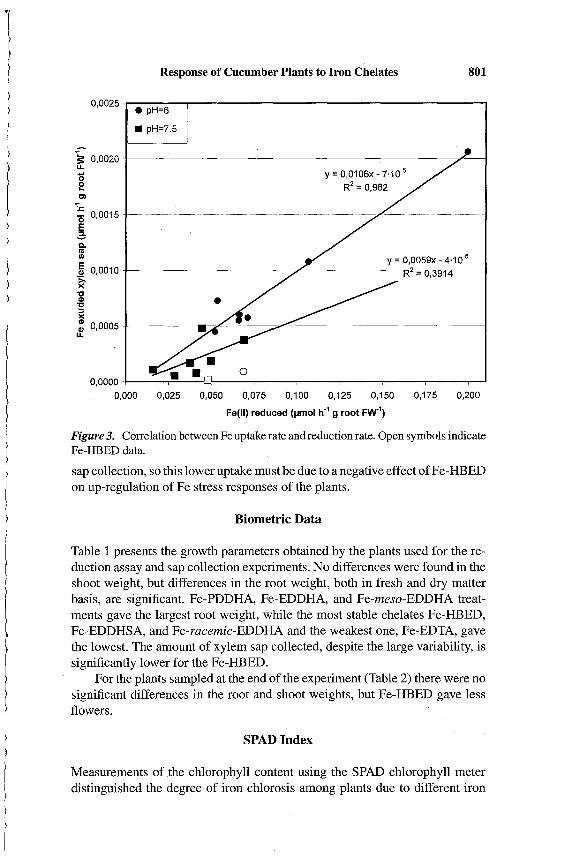
Iron chelate reductase activity is usually high when Fe-stressed plants aretransferred to Fe-chelate solutions, as in this case, but they cannot absorb allthe Fe that is reduced (Lucena and Chaney, 2006). The reduction mechanismin stressed plants -can be extremely active, more than enough to supply theFe needed by the plants, but the Fe(II) transport mechanism into the plantmay not be efficient at high pH. To illustrate this point, the rate of Fe uptakewas estimated by multiplying the xylem sap concentration and the sap volume(estimated from its weight) obtained in the time of xylem sap collection. Figure3 shows the correlation between the Fe-chelate reduction rate and the amountthat was transported in the xylem of the same plants. The correlation wassignificant using both pH 6 and pH 7.5 data, except those of Fe-HBED (datanot included in the regression analysis). The slope of the regressions are quitelow what means that the plant uses only a small part (1 % or less) of the ironthat is being reduced.
Uptake efficiency, defined by the percentage of Fe uptake with respect tothe Fe(III)-chelate reduced, was larger at pH 6 (average of 1.06%) than at pH7.5 (average of 0.59%). Plants grown with Fe-HBED (open symbols in Figure3) as Fe source had a very low percentage (0.13% at pH 6, and 0.017% at pH7.5). All plants were treated inthe same way during the FCR assay and xylem
Response of Cucumber Plants to Iron Chelates 801
0,0025 -r-:--:-:-:--...,-----------------------,• pH=6
• pH=7.5
y = 0,0106x _7'10.5
R2 = 0,962
o
•
~~ 0,0020 +-----------------------~~__Iu.'0eCl
:c'0 0,0015 + ,~................................................................ I
E.::Q.CIlIII
~ 0,0010 +--------------7-------
~"0G>
"0:::I
~G> 0,0005 +------.-zL........-'cu.
0,2000,1750,050 0,Q75 0,100 0,125 0,150
Fe(lI) reduced (llmol h-1 9 root FW1)
0,0000 +---..,.='----[-1---...,----,-----,-----,---...,...---.,-----'
0,000 0,025
Figure 3. Correlation between Fe uptake rate and reduction rate. Open symbols indicateFe-HEED data.
sap collection, so this lower uptake must be due to a negative effect ofFe-HBEDon up-regulation of Fe stress responses of the plants.
Biometric Data
Table 1 presents the growth parameters obtained by the plants used for the reduction assay and sap collection experiments. No differences were found in theshoot weight, but differences in the root weight, both in fresh and dry matterbasis, are significant. Fe-PDDHA, Fe-EDDHA, and Fe-meso-EDDHA treatments gave the largest root weight, while the most stable chelates Fe-HBED,Fe-EDDHSA, and Fe-racemic-EDDHA and the weakest one, Fe-EDTA, gavethe lowest. The amount of xylem sap collected, despite the large variability, issignificantly lower for the Fe-HBED.
For the plants sampled at the end of the experiment (Table 2) there were nosignificant differences in the root and shoot weights, but Fe-HBED gave lessflowers.
SPADlndex
Measurements of the chlorophyll content using the SPAD chlorophyll meterdistinguished the degree of iron chlorosis among plants due to different iron

802 J. J. Lucena and R. L. Chaney
Table IGrowth parameters of the 17 days old plants sampled for the reduction assay and xylemsap collection. Each value is the average f 8 data (4 pots/treatment x 2 bunch of plantsper pot). Different letter in the same column denotes significant differences betweenreatments (ex = 0.05, Duncan test)
Root fresh Root dry Shoot dryweight weight weight Sap weight
Treatment (gl2 plants) (g/2 plants) (gl2 plants) (gl2 plants/2h)
Fe-EDTA 8.73 bc 0.394 abc 1.42 ns 1.28 aFe-EDDHA 10.36 a 0.449 a 1.50 1.09 abFe-meso-EDDHA 10.07 a 00440 a 1.46 1.16 abFe-racemic-EDDHA 8.85 bc 0.389 bc 1.44 1.46 aFe-EDDHMA 9.20 ab 00400 ab 1.38 1.25 aFe-EDDHSA 7.79 c 0.343 c 1.36 1.02 abFe-PDDHA 10043 a 00449 a 1.49 1.23 aFe-HBED 8.60 bc 0.384 bc 1.411 0.763 b
treatments. Data for the SPAD index at different dates are shown in Table 3. Onlythe most indicative data corresponding to the leaves that were completing theirdevelopment are presented. At the end of the experiment values indicate thatplants presented different degrees ofFe chlorosis, confirmed by the visual aspectand as consequence of the low doses of chelates used during the experiment.
Trends are similar to those of the biometric parameters but more statisticaldifferences are observed. Fe-PDDHA, Fe-meso-EDDHA, and Fe-EDDHA gavehigher SPAD indexes, while the more stable chelates (Fe-racemic-EDDHA,
Table 2Growth parameters of the 28 days old plants sampled at the end of the experiment. Eachvalue is the average of 4 data (4 pots/treatment). Different letter in the same columndenotes significant differences between treatments (ex = 0.05, Duncan test)
Root fresh Root dry Shoot fresh Shoot dryweight weight weight weight # flowers
Treatment (gl2 plants) (g/2plants) (g/2 plants) (g/ 2 plants) (per 2 plants)
Fe-EDTA 12.18 ns 0.580 ns 15.99 ns 2.32 ns 9.5 abFe-EDDHA 14.60 0.619 17.81 2.62 10.0 abFe-meso-EDDHA 14.76 0.672 17.24 2.37 10.0 abFe-racemic-EDDHA 13.15 0.594 15.98 2.30 10.2 aFe-EDDHMA 14.33 0.600 17.26 2.56 13.2 aFe-EDDHSA 13.05 0.592 18.25 2044 10.2 aFe-PDDHA 13.59 0.617 16.87 2042 11.2 aFe-HBED 13.58 0.616 16.87 2042 7.8 b
Response of Cucumber Plants to Iron Chelates 803
Table 3SPAD index for the leaves (leaf 1 = oldest leaf) that were completing their developmentat different dates. Each value is the verage of 72 data (4 pots/treatment x 3 bunch ofplants/pot x 2 leaves/bunch x 3 readings per leaf) except for 28 days that are averagesof4 data (only 1 bunch per pot). Different letters in the same column denotes significantdifferences between treatments ()( = 0.05, Duncan test)
11 days 13 days 15 days 28 daysTreatment 1st leaf 2nd leaf 2nd leaf 3rd leaf
Fe-EDTA 36.1 c 32.2 b 36.3 b 25.5 bFe-EDDHA 39.4 b 36.0 a 40.8 a 30.5 aFe-meso-EDDHA 41.1 a 35.7 a 41.5 a 28.6 aFe-racemic-EDDHA 34.2d 33.0b 36.5 b 21.5 cFe-EDDHMA 39.5 b 34.9 a 40.0 a 30.2 aFe-EDDHSA 36.4 c 32.9 b 36.4 b 20.6 cdFe-PDDHA 39.6b 35.0 a 40.7 a 30.7 aFe-HEED 35.5 c 33.2 b 35.2 b 18.2 d
Fe-EDDHSA, and Fe-HBED) and the less stable Fe-EDTA gave lower SPADreadings.
Micronutrient Content in Leaves and Roots
Tables 4 and 5 present the micronutrient concentrations obtained in leaves androots at the end of the experiment. Fe concentrations in shoots are relativelylow to results of plants with adequate Fe nutrition confirming that were mildly
Table 4Micronutrient concentration in leaves of the 28 days old plants sampled at the end ofthe experiment. Each value is the average of 4 replicates (4 pots/treatment). Within acolumn treatment means followed by the same letter are significantly different (p <0.05, Duncan test)
mg·kg- I DM
Treatment Fe Mn Cu Zn Fe/Mn
Fe-EDTA 26.5 be 34.5 abc 8.16 a 19.3 ab 0.77 deFe-EDDHA 37.8 a 31.6 be 7.19 ab 22.4 a 1.19 aFe-meso- EDDHA 34.2 ab 30.7 c 6.99 ab 23.0 a 1.12 abFe-racemic- EDDHA 21.4 c 35.3 ab 7.67 a 19.2 ab 0.61 dFe-EDDHMA 27.6 be 31.0 c 7.26 ab 19.7 ab 0.89 beFe-EDDHSA 20.5 c 35.5 ab 6.79 ab 20.5 ab 0.58 dFe-PDDHA 29.6 abc 33.1 abc 6.12 b l8.0b 0.89 beFe-HBED 21.6 c 36.1 a 7.47 ab 19.2 ab 0.60d
804 J. J. Lucena and R. L. Chaney
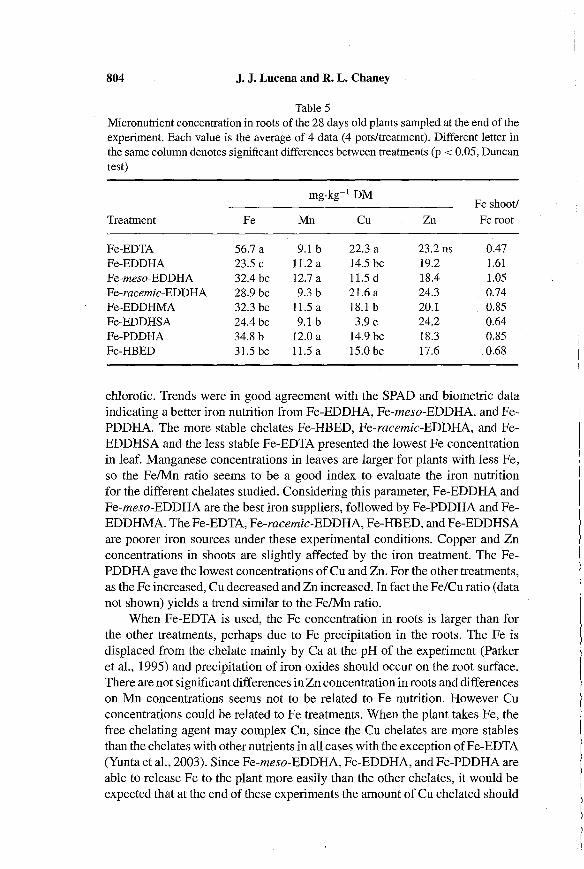
Table 5Micronutrient concentration in roots of the 28 days old plants sampled at the end of theexperiment. Each value is the average of 4 data (4 pots/treatment). Different letter inthe same column denotes significant differences between treatments (p < 0.05, Duncantest)
mg.kg-1 DMFe shoot/
Treatment Fe Mn Cu Zn Fe root
Fe-EDTA 56.7 a 9.1 b 22.3 a 23.2 ns 0.47Fe-EDDHA 23.5 c 11.2 a 14.5 bc 19.2 1.61Fe-meso-EDDHA 32.4 bc 12.7 a 11.5 d 18.4 1.05Fe-racemic-EDDHA 28.9 bc 9.3 b 21.6 a 24.3 0.74Fe-EDDHMA 32.3 bc U.S a 18.1 b 20.1 0.85Fe-EDDHSA 24.4 bc 9.1 b 3.9 e 24.2 0.64Fe-PDDHA 34.8 b 12.0 a 14.9 bc 18.3 0.85Fe-HEED 31.5 bc U.S a 15.0 bc 17.6 0.68
chlorotic. Trends were in good agreement with the SPAD and biometric dataindicating a better iron nutrition from Fe-EDDHA, Fe-meso-EDDHA, and FePDDHA. The more stable chelates Fe-HBED, Fe-racemic-EDDHA, and FeEDDHSA and the less stable Fe-EDTA presented the lowest Fe concentrationin leaf. Manganese concentrations in leaves are larger for plants with less Fe,so the FelMn ratio seems to be a good index to evaluate the iron nutritionfor the different chelates studied. Considering this parameter, Fe-EDDHA andFe-meso-EDDHA are the best iron suppliers, followed by Fe-PDDHA and FeEDDHMA. The Fe-EDTA, Fe-racemic-EDDHA, Fe-HBED, and Fe-EDDHSAare poorer iron sources under these experimental conditions. Copper and Znconcentrations in shoots are slightly affected by the iron treatment. The FePDDHA gave the lowest concentrations of Cu and Zn. For the other treatments,as the Fe increased, Cu decreased and Zn increased. In fact the Fe/Cu ratio (datanot shown) yields a trend similar to the FelMn ratio.
When Fe-EDTA is used, the Fe concentration in roots is larger than forthe other treatments, perhaps due to Fe precipitation in the roots. The Fe isdisplaced from the chelate mainly by Ca at the pH of the experiment (Parkeret aI., 1995) and precipitation of iron oxides should occur on the root surface.There are not significant differences in Zn concentration in roots and differenceson Mn concentrations seems not to be related to Fe nutrition. However Cuconcentrations could be related to Fe treatments. When the plant takes Fe, thefree chelating agent may complex Cu, since the Cu chelates are more stablesthan the chelates with other nutrients in all cases with the exception ofFe-EDTA(Yunta et aI., 2003). Since Fe-meso-EDDHA, Fe-EDDHA, and Fe-PDDHA areable to release Fe to the plant more easily than the other chelates, it would beexpected that at the end of these experiments the amount of Cu chelated should

Response of Cucumber Plants to Iron Chelates 805
be considerably higher than in the others, reducing the Cu uptake by the plants,and diminishing the Cu concentration in roots. The Cu concentration in rootsis well related to the Cu concentration in shoots, except for Fe-EDDHSA thatgives very low tissue Cu levels. This result has to be taken with care, as the FeEDDHSA commercial samples employed in this study contain large amountsof impurities that could retain Cu and hence affect the Cu concentration inplant.
Among other factors, iron nutrition is related with the long distance transport ofFe. Thus, an accumulation ofFe in roots as that observed in the Fe-EDTAtreatment does not imply a better Fe nutrition. The ratio between Fe concentration in shoots and roots may be a good index of the long distance transport.Table 5 shows that this ratio is well correlated with all the other parameters thatare related to the quality of iron nutrition.
DISCUSSION
Several authors (Ellsworth et aI, 1997) have used the determination of the FCRas technique for screening Fe-chlorosis tolerant genotypes. Here FCR activitywas utilized as an index to evaluate the Fe nutrition of cucumber plants grownhydroponically with low concentrations of different Fe-chelates. However theresults obtained (Figure 1) did not present the same trend that the traditionaliron nutrition parameters, such as yield, chlorosis index or iron concentrationin shoots (Tables 1 to 5). In these experimental conditions plants are grownwith the same amount of Fe, so differences among treatments were expected tobe low and possibly hidden in the variability normally found in FCR activitymeasurements. Several causes may explain the variability, from the effect ofenvironmental factors to the effect of genetic differences on the expressionof an enzymatic activity. Also, FCR activity is expressed per unit root freshweight. Since the reduction is done only in a small part of the roots (near thetips) (Marschner and Romheld, 1995), reference to the whole root weight mayincrease the variability of the results. The fact that Fe-EDDHMA plants hada large FCR activity may be due to the bias associated with the experimentalmethodology.
Also Fe was determined in the sap collected after the FCR assay using thesame solutions, but without BPDS. It has to be considered that experimentswere done consecutively and not in parallel, so a decrease of the FCR can beexpected during the xylem sap collection with respect the FCR assay. However,this decrease can not explain the high differences observed in Fe reduced (Figure1) with respect to the Fe in sap (Figure 2). Without the trapping agent BPDS, theFe(II) produced in the xylem sap collection experiment and not absorbed by theplant, must be complexed by the EDTA or reoxidized. The ultimate electronacceptor for the reoxidation should be the oxygen, but the EDTA catalyzesreoxidation by favoring the Fe(III)-chelate, or even acting as electron carrier

806 J. J. Lucena and R. L. Chaney
(Kurimura et al., 1968). The role of the chelating agents in Fe(II) reoxidationin natural conditions of plant growth should receive more consideration since itmay affect the "shuttle effect" that has been proposed for the action of chelatesin soils (Lindsay, 1995).
The substrate for the iron chelate reductase determination was always FeEDTA, so the ratio between Fe reduction and Fe uptake should be constant foreach pH value. However, plants treated with Fe-HEED had a lower uptake ratethan the expected from the reduction rate (Figure 3). Additionally, the SPADindex (Table 3) and other measurements appear to support the conclusion thatplants supplied with Fe-HBED had more chlorosis than with the other Fechelates. This raises a question about the effect of the chelate, not only onthe reduction mechanism, but also on the transporter across the root plasmamembrane.
With respect to the comparison of the iron nutrition from the differentFe-chelates it is useful to consider the difference between responses to FeEDTA and to the other chelates. The Fe complexed as Fe-EDTA is readilydisplaced in the nutrient solution by Mn, Zn, or Cu at pH 6, and also by Caat pH 7.5, precipitating in an inorganic form (Lindsay, 1995) and retained onthe root surface (Kosegarten and Koyro, 2001). Occasionally the plants usethe Fe from this inorganic form, or some Fe-EDTA (due to the kinetic effects)may reach the root surface. As expected, Fe nutrition from Fe-EDTA was lesseffective than from other chelates (Table 4). All the other chelating agentscontain two o-hydroxyphenyl, two carboxylic, and two amino groups that givea very highly stable Fe (III) complex. In fact, when there is enough Fe present,only the Fe chelate is formed with the chelating agent. However, when theiron concentration is low, a second cation, e.g., Cu, can be complexed by thechelating agent (Yunta et al., 2003). When present in adequate levels, all ofthe Fe(III)-chelates are good Fe sources for the strategy I plants. When addedhydroponically at low concentrations, the differences in iron nutrition shouldbe attributed to differences between different Fe-chelates as substrates of theFCR and their effect on the Fe(II) uptake mechanism. The ability to give ironto plants follows the order: Fe-meso-EDDHA = Fe-EDDHA = Fe-PDDHA >Fe-EDDHMA > Fe-racemic-EDDHA > Fe-HEED = Fe-EDDHSA. This isthe same order that was obtained when these chelates were used as substrates ofthe FCR activity measured on mildly Fe-stressed cucumber plants (Lucena andChaney, 2006). This sequence is the opposite of the Fe-chelate stability (Yuntaet al., 2003; Lucena and Chaney, 2006).
Two different associated mechanisms to explain the differences in ironnutrition from the different Fe(III)-chelates are proposed. The first one is relatedwith Fe reduction. Before the Fe is reduced the Fe-chelate has to bind to theFCR. Then, an electron is transferred to the Fe(III)-chelate and Fe(III) is reduced(Robinson et al., 1999). As the Fe(II)-chelate is weaker than the Fe(III)-chelate,Fe (II) is separated from the chelating agent. Since the whole process depends

Response of Cucumber Plants to Iron Chelates 807
on the chelate (or Fe) binding to the FCR enzyme and the stability of bothFe(II) and Fe(III) chelates (Marschner and Romheld, 1995) there may be norelationship between the stability constant for a Fe(III)-chelate and the ability ofthe FCR to reduce Fe(III) in it. However, in this case all the chelates (except forFe-EDTA) had the same donor groups and the stability of the Fe(III)-chelateswas inversely correlated with the FCR activity (Lucena and Chaney, 2006).This is also in good agreement with the sorption characteristics of meso andracemic isomers of Fe-EDDHA and Fe-EDDHMA onto Fe oxides (HernandezApaolaza and Lucena, 2001). The second mechanism is related to the fate ofthe Fe(II) once it is reduced and dissociated from the chelating agent. ThisFe(II) may bind to the fer,rous permease of the root epidermal cells and then beabsorbed by the plant (Fox et aI., 1996). However, since little of the Fe reducedis taken up (Figure 3), most ofthis Fe(II) must be reoxidized, as in equation 1,where Y is the chelating agent, which catalyzes the auto-oxidation of Fe(II) bydissolved O2 (X) (Kurimura et al., 1968).
Fe(II) - Y + X ---+ Fe(III) - Y + X(e-) (1)
This reaction should occur more rapidly for the more stable Fe (III)-chelatesimpeding Fe (II) absorption. Both mechanisms are consistent with experimentaldata, so it is believed that both may occur simultaneously. The conclusion isthe same: the less stable chelates give more Fe to the stressed cucumber plants.
In addition ofbeing the most stable chelate, the negative effect ofFe-HBEDon Fe uptake should also be considered, since this gave the lowest ratio betweenFe uptake and Fe reduction.
In order to predict the behavior of an iron chelate applied to a soil, not onlyplant uptake characteristics must be considered, but also the soil chemistry ofthe Fe (III)-chelate. Low stable Fe (III)-complexes (e.g., those formed withweak organic acids, fulvates, and EDTA-like chelating agents), are retained onthe soil surfaces or the iron may be displaced by competing metals (Norvell,1991). While the chelators that form weak Fe (III)-chelates react in the soil, thechelators that form stronger Fe (III)-chelates retard iron reduction and uptake.Thus the best results will be obtained with Fe (III)-chelates of intermediatestrength.
CONCLUSIONS
When plants are grown hydroponically with low concentration of Fe-chelates,the less stable Fe (III)-chelates are the more effective, exception made of thosein which the displacement of the Fe by other metals takes place, as in the caseof Fe-EDTA. The use of Fe (llI)-EDTA as iron supplier for plants is clearlydisadvantageous compared to the rest of the compounds tested in the presentwork.

808 J. J. Lucena and R. L. Chaney
ACKNOWLEDGMENTS
The authors gratefully acknowledge the technical assistance of Carrie E. Greenand Francisca BIas for their help in conducting the experiments and to Prof. M.G6mez-Gallego for her helpful review of the manuscript. This work was doneduring a research visit of J.J. Lucena in the USDA supported by a MEC-CICyTgrant. Part of the work has been supported by project AGL2004-07849 fromthe Spanish Ministry of Education and Science.
REFERENCES
Alvarez-Fermmdez, A., S. Garcia-Marco, and J. J. Lucena. 2005. Evaluation of synthetic iron (III)-chelates (EDDHA/Fe3+, EDDHMA/Fe3+ andEDDHSAlFe3+) to correct iron chlorosis. European Journal ofAgronomy22: 119-130.
Bienfait, H. F. 1985. Regulated redox processes at the plasmalemma of plantroot cells and their function in iron uptake. Journal ofBioenergetics andBiomembranes 17:73-83.
Chaney, R. L., J. C. Brown, and L. O. Tiffin. 1972. Obligatory reduction offerric chelates in iron uptake by soybeans. Plant Physiology 50: 208-213.
Ellsworth, J. w., V. D. Jolley, D. S. Nuland, and A. D.BIaylock. 1997. Screeningfor resistance to iron deficiency chlorosis in dry bean using iron reductioncapacity, Journal ofPlant Nutrition 20: 1489-1502.
Fox, T. c., J. E. Shaff, M. A. Grusak, W. A. Norvell, Y. Chen, R. L. Chaney,and L.V. Kochian. 1996. Direct measurement of59Fe-labeled Fe2+ influx inroots of pea using a chelator buffer system to control free Fe2+ in solution.Plant Physiology 111: 93-100.
Hemandez-Apaolaza, L., and J. J. Lucena. 2001. Fe (III)-EDDHA and EDDHMA sorption on Ca-montmorillonite, ferrihydrite and peat. JournalofAgricultural Food Chemistry 49: 5258-5264.
Kosegarten, H., and H. W. Koyro. 2001. Apoplastic accumulation of iron in theepidermis of maize (Zea mays) roots grown in calcareous soil. PhysiologiaPlantarum 113: 515-522.
Kurimura, Y., R. Ochai, and N. Matsuura. 1968. Oxygen oxidation of ferrousions induced by chelation. Bulletin of the Chemical Society of Japan 41:2234-2239.
Lindsay, W. L. 1979. Chemical Equilibria in Soils. New York: John Wiley andsons.
Lindsay, W. L. 1995. Chemical reactions in soils that affect iron availability toplants. A quantitative approach. In Iron Nutrition in Soils and Plants, ed.J. Abadfa, 7-14. Dordrecht, The Netherlands: Kluwer Academic Pubs.
Lucena J. J. 2003. Fe chelates for remediation of Fe chlorosis in Strategy Iplants. Journal ofPlant Nutrition 26 (10&11): 1969-1984.

Response of Cucumber Plants to Iron Chelates 809
Lucena, l l, and R. L. Chaney. 2006. Root ferric chelate reductase and ironconcentration in xylem sap as indexes of the iron uptake by green stressedcucumber plants from synthetic iron chelates. Journal of Plant Nutrition29: 423-439.
Lucena, l l, A Garate, and O. Carpena. 1998. Theoretical and practical studieson chelate-Ca-pH system in solution. Journal ofPlant Nutrition 11: 10511061.
Marschner, H., and V. Rornheld. 1995. Strategies of plants for acquisition ofiron. In Iron Nutrition in Soils and Plants, ed. l Abadfa, 375-9378. Dordrecht, The Netherlands: Kluwer Academic Pubs.
Minolta. 1989. Chlorophyll meterSpad 502 operation manual, Mino1ta Camera.Japan.
Norvell, W. A. 1991. Reactions of metal chelates in soils and nutrient solutions.In Micronutrients in Agriculture, eds. l J. Morvedt, S. R. Cox, L. M.Shuman, and R. M. Welch, 187-223. Madison, Wisconsin: SSSA
Nowack, B., and L. Sigg. 1997 Dissolution of iron (III) (hydr) oxides by metalEDTA complexes. Geochimica Cosmochimica Acta 61: 951-963.
Parker, D. R., R. L. Chaney, and W. A Norvell. 1995. Chemical EquilibriaModels: Applications to Plant Nutrition Research. In Chemical Equilibrium and Reaction Models, eds. R. M. Loeppert, A P. Schwab, and S.Goldberg, 163-200. Madison, Wisconsin: SSSA
Perez-Sanz, A, and l J. Lucena. 1995. Synthetic ion oxides as sources of Fein a hydroponic culture of sunflower. In Iron Nutrition in Soils and Plants,ed. J. Abadfa, 241-246. Dordrecht, The Netherlands: Kluwer AcademicPubs.
Robinson, N.J., C. M. Procter, E. L. Conolly, and M. L. Guerinot. 1999. Aferric-chelatereductase for iron uptake from soils. Nature 397: 694-697.
Schwertmann, U. 1991. Solubility and dissolution of iron oxides. In Iron Nutrition and Interaction in Plants, eds. Y. Chen and Y. Hadar, 3-27. Dordrecht,The Netherlands: Kluwer Academic Pubs.
Stone, A T. 1997. Reactions of extracellular organic ligands with dissolvedmetal ions and mineral surfaces. In Reviews in Mineralogy (35), Geomicrobiology: Interactions Between Microbes and Minerals, eds. J. E. Banfieldand K. H. Nealson, 309-343. Washington, D.C.: Mineralogical Society ofAmerica.
Susin, S., A Abadfa, J. A Gonzalez-Reyes, J. J. Lucena, and J. Abadia. 1996.The pH requirement for in vivo activity of the iron-deficiency-induced"Turbo" ferric chelate reductase. Plant Physiology 110: 111-123.
Yunta, E, S. Garcia-Marco, J. J. Lucena, M. G6mez-Gallego, R. Alcazar, andM. A. Sierra. 2003. Chelating Agents Related to Ethylenediamine bis (2hydroxyphenyl) acetic acid (EDDHA): Synthesis, Characterization, andEquilibrium Studies of the Free Ligands and their Mg2+, Ca2+, Cu2+ andFe3+ Chelates. Inorganic Chemistry 42: 5412-5421.
Related Documents











