International Journal of Food Microbiology 48 (1999) 37–50 Resistance of Escherichia coli and Salmonella against nisin and curvacin A 1 * ¨ Michael G. Ganzle , Christian Hertel, Walter P. Hammes ¨ ¨ Universitat Hohenheim, Institut f ur Lebensmitteltechnologie (ALT), Garbenstrasse 28, D-70599 Stuttgart, Germany Received 20 July 1998; received in revised form 15 January 1999; accepted 8 February 1999 Abstract We have determined the effects of the following factors on the resistance of Gram-negative bacteria against nisin and curvacin A: (i) chemotype of the lipopolysaccharide (LPS), (ii) addition of agents permeabilizing the outer membrane, (iii) the fatty acid supply of the growth medium, and (iv) the adaptation to acid and salt stress. Bacteriocin activity was determined against growing and resting cells as well as protoplasts. All smooth strains of Escherichia coli and Salmonella enterica serovar Typhimurium were highly resistant towards the bacteriocins, whereas mutants that possess the core of the LPS, but not the O antigen, as well as deep rough LPS mutants were sensitive. Antibiotics with outer membrane permeabilizing activity, polymyxin B and polymyxin B nonapeptide, increased the sensitivity of smooth E. coli towards 21 nisin, but not that of deep rough mutants. Incorporation of 1 g l of either oleic acid or linoleic acid to the growth media greatly increased the susceptibility of E. coli LTH1600 and LTH4346 towards bacteriocins. Both strains of E. coli were sensitive to nisin and curvacin A at a pH of less than 5.5 and more than 3% (w/v) NaCl. Adaptation to sublethal pH or higher NaCl concentrations (pH 4.54 and 5.35 or 4.5% (w / v) NaCl) provided only limited protection against the bacteriocidal activity of nisin and curvacin A. Adaptation to 4.5% (w/v) NaCl did not result in cross protection to bacteriocin activity at pH 4.4, but rendered the cells more sensitive towards bacteriocins. 1999 Elsevier Science B.V. All rights reserved. Keywords: Nisin; Curvacin A; Escherichia coli O157:H7; Salmonella; Resistance; LPS mutants 1. Introduction Escherichia coli and Salmonella enterica are of major concern for a wide variety foods that have not *Corresponding author. Tel.: 149-711-459-2305; fax: 149- undergone a germ reducing process. Virulent strains 711-459-4199. of E. coli have increasingly been recognized as E-mail address: [email protected] (W.P. Ham- foodborne pathogens. Among the six virotypes of E. mes) 1 coli, enterohemorrhagic E. coli (EHEC) are consid- ¨ ¨ ¨ Present address: Technische Universitat Munchen, Lehrstuhl fur Technische Mikrobiologie, D-85350 Freising-Weihenstephan. ered to be highly significant due to their low 0168-1605 / 99 / $ – see front matter 1999 Elsevier Science B.V. All rights reserved. PII: S0168-1605(99)00026-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

International Journal of Food Microbiology 48 (1999) 37–50
Resistance of Escherichia coli and Salmonella against nisin andcurvacin A
1 *¨Michael G. Ganzle , Christian Hertel, Walter P. Hammes
¨ ¨Universitat Hohenheim, Institut f ur Lebensmitteltechnologie (ALT), Garbenstrasse 28, D-70599 Stuttgart, Germany
Received 20 July 1998; received in revised form 15 January 1999; accepted 8 February 1999
Abstract
We have determined the effects of the following factors on the resistance of Gram-negative bacteria against nisin andcurvacin A: (i) chemotype of the lipopolysaccharide (LPS), (ii) addition of agents permeabilizing the outer membrane, (iii)the fatty acid supply of the growth medium, and (iv) the adaptation to acid and salt stress. Bacteriocin activity wasdetermined against growing and resting cells as well as protoplasts. All smooth strains of Escherichia coli and Salmonellaenterica serovar Typhimurium were highly resistant towards the bacteriocins, whereas mutants that possess the core of theLPS, but not the O antigen, as well as deep rough LPS mutants were sensitive. Antibiotics with outer membranepermeabilizing activity, polymyxin B and polymyxin B nonapeptide, increased the sensitivity of smooth E. coli towards
21nisin, but not that of deep rough mutants. Incorporation of 1 g l of either oleic acid or linoleic acid to the growth mediagreatly increased the susceptibility of E. coli LTH1600 and LTH4346 towards bacteriocins. Both strains of E. coli weresensitive to nisin and curvacin A at a pH of less than 5.5 and more than 3% (w/v) NaCl. Adaptation to sublethal pH orhigher NaCl concentrations (pH 4.54 and 5.35 or 4.5% (w/v) NaCl) provided only limited protection against thebacteriocidal activity of nisin and curvacin A. Adaptation to 4.5% (w/v) NaCl did not result in cross protection to bacteriocinactivity at pH 4.4, but rendered the cells more sensitive towards bacteriocins. 1999 Elsevier Science B.V. All rightsreserved.
Keywords: Nisin; Curvacin A; Escherichia coli O157:H7; Salmonella; Resistance; LPS mutants
1. Introduction
Escherichia coli and Salmonella enterica are ofmajor concern for a wide variety foods that have not
*Corresponding author. Tel.: 149-711-459-2305; fax: 149- undergone a germ reducing process. Virulent strains711-459-4199. of E. coli have increasingly been recognized as
E-mail address: [email protected] (W.P. Ham-foodborne pathogens. Among the six virotypes of E.mes)
1 coli, enterohemorrhagic E. coli (EHEC) are consid-¨ ¨ ¨Present address: Technische Universitat Munchen, Lehrstuhl furTechnische Mikrobiologie, D-85350 Freising-Weihenstephan. ered to be highly significant due to their low
0168-1605/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved.PI I : S0168-1605( 99 )00026-4

¨38 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
infectious dose and the severe consequences of brane, and therefore confers a high degree of resist-infection (Buchanan and Doyle, 1997). ance (Stevens et al., 1991; Schved et al., 1994).
Enteric bacteria are especially tolerant towards Chelating agents such as EDTA as well as theadverse environmental conditions such as low pH, application of sublethal stress such as heating orhigh salt concentrations (Small et al., 1994; Cheville freezing were shown to disrupt the permeabilityet al., 1996; Brown et al., 1997) and have been barrier of the LPS leading to an increased sensitivityshown to survive during storage in acid foods or of Salmonella enterica and E. coli towards nisinproducts with high concentrations of salt or organic (Stevens et al., 1991; Kalchayanand et al., 1992).acids (Presser et al., 1998; Glass et al. 1992; Leyer et We have recently shown that nisin and curvacin Aal., 1995; Reitsma and Henning, 1996). For example, in combination with low pH, . 5% NaCl, or pro-E. coli O157:H7 is most often associated with EHEC pylparabene also leads to a increased sensitivity of E.infections and remained viable on beef salads or raw coli and S. enterica towards nisin and curvacin A
¨salad vegetables for several days during refrigerated (Ganzle et al., 1999). These results suggested thatstorage (Abdul-Raouf et al., 1993a,b). Acid adapted bacteriocins may be active against E. coli at en-cells of E. coli O157:H7 exhibited an increased vironmental conditions near the growth limitingsurvival in acidic foods (Leyer et al., 1995). The acid factor levels even if a functional outer membrane istolerance of E. coli is inducible and involves the present. To investigate the role of the OM insynthesis of acid shock proteins, activation of meta- bacteriocin resistance in more detail, the effect of thebolic enzymes to maintain a high internal pH, and outer membrane composition on bacteriocin sen-the increased incorporation of cyclopropane fatty sitivity was assayed using sets of S. enterica and E.acids in the cytoplasmic membrane (Bearson et al., coli LPS mutants as well as agents that selectively1997; Rowbury, 1995; Brown et al., 1997). Several permeabilize the outer membrane. Furthermore, theof these systems require the alternative sigma factor effect of medium composition and pre-adaptation to
Ss , a regulator of late logarithmic and stationary sublethal acid and salt stress on the bacteriocingrowth, which furthermore provides cross protection sensitivity of strains of E. coli was investigated.towards heat and salt stress (Small et al., 1994;Cheville et al., 1996). Furthermore, acid tolerancewas induced by aerobiosis and pre-adaptation to 2. Materials and methodssublethal pH (Small et al., 1994; Benjamin andDatta, 1995; Brown et al., 1997). 2.1. Microorganism and media
Several reports suggest that bacteriocins of lacticacid bacteria may contribute to the inactivation of The susceptibility of Gram-negative bacteria to-Gram-negative microorganisms in food if these are wards nisin and curvacin A was assessed using twoapplied in combination with chelating agents (Shefet wild type strains of E. coli and sets of isogenic LPSet al., 1995; Scanell et al., 1997). The efficacy of mutants of E. coli and S. enterica. A compilation ofthese peptide antibiotics to inactivate Gram-positive the organisms used and their relevant characteristicspathogens and spoilage organisms such as Listeria are shown in Table 1 and a schematic representationmonocytogenes and Clostridia is well established of the S. enterica serovar Typhimurium LPS(Stiles, 1996; Hammes and Hertel, 1998). An im- chemotypes is shown in Fig. 1. The designations R1portant example is the lantibiotic nisin, produced by and K-12 for the E. coli strains ATCC 27325 andLactococcus lactis, which is approved as a food F470 refer to LPS chemotypes retaining the inneradditive in several countries. Furthermore, competi- core without the O-antigen (Nikaido, 1996; Brabetztive starter cultures are available producing bac- et al., 1997), and the E. coli deep rough (Re) mutantsteriocins in situ during food fermentations or upon have the same chemotype as the S. enterica Reapplication as protective cultures (Hammes and mutants (Nikaido, 1996; Brabetz et al., 1997). TheHertel, 1998). cultures of E. coli and S. enterica were incubated at
The architecture of the outer membrane (OM) of 378C and agitated at 200 rpm unless otherwiseGram-negative organisms prevents penetration of the indicated. The strains were grown in MRS (De Manbacteriocins to their target, the cytoplasmic mem- et al., 1960) without sodium acetate (mMRS, pH 6.5

¨M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50 39
Table 1Organisms tested for susceptibility towards nisin and curvacin A
Organism Strain designation Chemotype of LPS Reference
S. enterica serovar SL3770 Wild type Roantree et al., 1977Typhimurium LT2 (smooth)
SL3749 RaSL3750 Rb2
SL3748 Rb3
SL3769 Rb1
SL3789 Rb2
S. enterica serovar SA1355 Wild type Sanderson et al., 1974Typhimurium SA1377 ReE. coli F492 Wild type, O8:K27- Schmidt et al., 1970
F470 R1F515 Re
E. coli ATCC 27325 K-12WBB06 Re Brabetz et al., 1997
E. coli LTH1600 Smooth Our strain collectionE. coli LTH4346 Smooth, O157:H7
Fig. 1. Schematic representation of the LPS structure of S. Typhimurium LT2 and its LPS mutants according to Nikaido (1996). Ra, Rb1
etc. denote the LPS chemotype; smooth (wild type) LPS contains the O-chain. Abbreviations: GlcNAc: N-acetyl-D-glucosamine, Glc:D-glucose, Gal: D-galactose, Hep: L-glycero-D-manno-heptose, EtN: ethanolamine, Kdo: 3-deoxy-D-manno-oct-2-ulosonic acid.
21 21unless otherwise stated), or LB-broth (5 g l to LB-broth at a final concentration of 1 g l after21 21tryptone, 5 g l yeast extract, and 5 g l NaCl, pH autoclaving. For variation of the pH and the NaCl
7.2 unless otherwise stated). In experiments with concentration, 4 M NaOH, 4 M HCl, or NaCl werefatty acid supplementation, stearic acid (Merck, added to obtain the desired pH or NaCl concentrationDarmstadt), oleic acid (Sigma, Deisenhofen) or and the media were sterilized by filtration. Thelinoleic acid (Fluka, Buchs, Switzerland) were added concentrations of polymyxin B or polymyxin B

¨40 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
nonapeptide were adjusted by diluting sterile stock of 0.00160.0003 with indicator strains grown to the21solutions of these antibiotics (0.5 mg ml in 50% early logarithmic phase (OD 0.1–0.2) in600 nm
(v /v) ethanol) in mMRS to the desired concentration. mMRS. For the determination of the bacteriocinactivity at various pH and NaCl levels, mMRS with
2.2. Bacteriocin preparations the appropriate pH or NaCl content (pH 4.54, 4.80,5.35, 6.30, or 6.90; 0, 3, 5, 6, and 7% (w/v) NaCl)
Nisin (kindly provided by Aplin & Barret Ltd., was used. The plates were incubated at 378C withoutUK) was dissolved in 50% ethanol at a concentration agitation and the OD was measured in 60 min
21of 0.5 mg ml and stored at 2 208C. Curvacin A intervals during 48 h. The time when the indicator1was purified from culture supernatant of the curA strain in the control media (no addition of bac-
strain L. curvatus LTH1174 (Tichaczek et al., 1992) teriocin) had reached the logarithmic growth phasegrown to early stationary phase at 158C in MRS. The was determined. At this time, the OD-data of eachbacteriocin was precipitated by addition of 40% dilution series were used to calculate the bacteriocin(w/w) ammonium sulfate, the solids were collected activity as follows: (i) the data was fitted to theby centrifugation and redissolved in H O . NaCl following equation (Parente et al., 1995):2 demin
was added to saturation and the solution was ex-tracted three times with 1/4 vol. isopropanol. The OD (1 2 a)16 hcombined organic phases were pooled, evaporated to ]] ]]]]5 a 1 ,b1c(ln d )OD 1 1 e0 hdryness, and loaded on a Superdex 30 gelfiltrationcolumn (Pharmacia, Uppsala, Sweden) using 50 mMtriethylamine (pH adjusted to 7.2 with CO ) in water where d is the dose (ml bacteriocin stock solution /2
and 25% (w/w) isopropanol as elution buffer. The ml total volume or mg nisin /ml), and a, b, and c areactive fractions were pooled, organic solvents re- regression coefficients. The SigmaPlot 1.0 softwaremoved under nitrogen, and the stock solution was package (SPSS Inc, Chicago, IL, USA) was used forstored in aliquots at 2 208C. Prior to use, the nisin all curve fit procedures. (ii) The regression coeffi-and curvacin A stock solutions were diluted 1:10 cients a, b and c were used to calculate the dilutionwith the assay media. level resulting in a 50% growth inhibition (d ). (iii)50
Bacteriocin activity was calculated as 1 /d and50212.3. Determination of growth rates and lag phase expressed as AU ml . If the concentration of the
bacteriocin at the various dilution levels was knownE. coli LTH1600 and E. coli LTH4346 were (nisin, polymyxin B and polymyxin B nonapeptide),
inoculated anaerobically in mMRS (pH 4.54, 4.80, the concentration resulting in 50% growth inhibition5.35, 6.30, or 6.90; 0, 3, 5, 6, and 7% (w/v) NaCl) (d ) was expressed as minimum inhibitory con-50
and the growth was recorded by measuring the centration (MIC). The MIC of polymyxin B againstoptical density at 600 nm. The media were inocu- E. coli F492 and F515 was determined accordinglylated to an OD of ca. 0.001 with the strains grown to in six independent experiments. For the determi-early logarithmic phase in mMRS, mMRS pH 5.3, or nation of bacteriocin activity during the purificationmMRS 4.5% (w/v) NaCl. The maximum specific of curvacin A and for the standardization of the stockgrowth rates m and the lag phases were de- solutions of nisin and curvacin A, L. curvatusmax
termined by fitting the OD vs. time data to the LTH1432 was used as indicator strain, mMRS (pHlogistic growth curve (Zwietering et al., 1990). 6.5) was used as assay medium and the optical
density of the cultures was measured after 16 h.2.4. Determination of bacteriocin activity againstgrowing cells
2.5. Determination of bacteriocin activity againstThe bacteriocin activity against growing cells was resting cells
measured by a critical dilution assay. Two-fold serialdilutions of the nisin or curvacin A solutions were Cells of strains E. coli LTH1600 and E. coliprepared with mMRS and inoculated to an OD LTH4346 were grown to stationary phase and added600 nm

¨M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50 41
to the assay buffer (50 mM citrate, pH 4.6) to a cell 3. Results7 21count of 460.4310 CFU l . Cell counts were
determined 0, 2, 4, and 8 h after addition of nisin or 3.1. Sensitivity of isogenic LPS mutant strainscurvacin A by plating appropriate dilutions in buf- against nisin and curvacin A
21 21fered peptone water (8 g l NaCl, 1 g l tryptone,21 21 21 To determine the constituents of the outer mem-1 g l CaCl , 1 g l Tween 80, 2.6 g l KH PO ,2 2 4
brane (OM) critical for the barrier effect againstpH 6.5) on LB-agar. Two experiments were per-bacteriocins, the activity of nisin and curvacin Aformed to determine the effect of the pH, the NaClagainst LPS mutants of E. coli and S. enterica wasconcentration, and fatty acid supplementation of theevaluated. The activity of nisin and curvacin Apreculture on the susceptibility towards bacteriocins.against the indicator strains was normalized to their(i) The cells were grown in mMRS at pH 4.5, 5.5,activity against one of the deep rough (Re) mutants,6.5, or 7.5, as well as mMRS pH 6.5 containingE. coli F515. As shown in Fig. 2, all deep rough (Re)4.5% (w/v) NaCl, and curvacin A was added to an
21 mutants were sensitive against both bacteriocins. Noactivity of 96 AU ml against L. curvatusactivity was detected against any of the wild typeLTH1432. (ii) Cells were grown in LB-broth orstrains of E. coli and S. enterica serovarLB-broth additionally containing stearic acid or oleicTyphimurium or the Ra mutant SL3749. The LPSacid and nisin was added to a concentration of 10
21 defective mutants derived from the smooth strainmg l . Control experiments were performed withoutSL3770 were bacteriocin sensitive when the LPSaddition of bacteriocin.was truncated beyond the galactose residue in thecarbohydrate mainchain (corresponding to the LPS
2.6. Determination of bacteriocin activity against chemotype Rb , Rc, Rd and Rd ). Strain SL37893 1 2
protoplasts (Rd2) was, however, more resistant towards nisinthan the Rd mutant, and inhibition by curvacin A1
Protoplasts were prepared from cells grown to was below the detection limit (relative activity 0.1).logarithmic phase according the method described by Remarkably, the curvacin A activity against strainsSambrook et al. (1989). In short, cells were grown to E. coli ATCC 27325 and F470 was comparable tomid logarithmic growth phase, washed twice in ice that of the corresponding deep rough (Re) mutants.cold 50 mM Tris3HCl (pH 8.0) containing 20% These strains possess the complete core regions of(w/v) sucrose and resuspended in buffer. Protoplast the LPS carbohydrate but are devoid of the O-formation was achieved by addition of 0.2 vol. antigen. The nisin activity against strains F470 and
21lysozyme solution (5 mg ml in 0.25 M Tris3 ATCC 27325 was 25 and 10%, respectively, com-HCL, pH 8.0) and 0.4 vol. EDTA (0.25 M, pH 8.0). pared to the corresponding Re mutants.The cell suspension was incubated at 378C until LPS mutants of E. coli are known to differ from.95% of the cells appeared spherical under the the wild type strains not only in the LPS chemotype,microscope (15–30 min). The protoplasts were col- but also in the composition of the cytoplasmiclected by centrifugation, and resuspended to an membrane. In wild type cells, components of theOD of 0.2 in 50 mM Tris buffer (pH 8.0) or 50 LPS are located exclusively in the outer leaflet of the595 nm
mM citrate buffer (pH 4.8), each buffer containing outer membrane (Magnuson et al., 1993). In deep10 mM CaCl and 20% (w/v) sucrose. Nisin was rough mutants, LPS components are incorporated in2
21added to a final concentration of 2.5 or 5 mg ml significant amounts in the cytoplasmic membrane, asand the lysis of protoplasts was followed by measur- it was shown by analysis of cell wall fractionsing the optical density at 595 nm. The percentage of prepared from heptose-less E. coli on 3-deoxy-D-intact protoplasts was calculated from the OD of the manno-oct-2-ulosonic acid (Kdo) and b-hydroxybacteriocin containing samples compared to the myristate (Koplow and Goldfine, 1974). To deter-control without addition of bacteriocin. It was ver- mine whether this altered composition of the cyto-ified during each trial that no change in optical plasmic membrane results in an increased resistancedensity occurred when solvent (0.005 ml ethanol /ml to bacteriocins, protoplasts were prepared of wildbuffer) was added in place of nisin. type strains and the corresponding LPS mutant

¨42 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
Fig. 2. Bacteriocin activity (nisin, and curvacin A, ) against LPS mutant strains of Salmonella enterica and E. coli. The activity wasexpressed relative to the inhibitory effect against E. coli F515. Error bars indicate the means6standard deviation of two independent
21experiments. The activity of the curvacin A and nisin stock solutions against E. coli F515 were 5764 and 420634 AU ml , respectively,and the detection limit of the assay was a relative activity of 0.1 and 0.01 for curvacin A and nisin, respectively.
strains, and their susceptibility against nisin was where no activity could be detected against wild typedetermined. Protoplasts of E. coli are lysed by nisin strains.in a dose dependent manner (Schved et al., 1994).We verified the dose dependent lytic action of nisinagainst protoplasts using E. coli LTH1600 (Fig. 3A). 3.2. Effect of polymyxin B and polymyxin BCurvacin A failed to lyse protoplasts of E. coli nonapeptide (PMBN) on nisin activityLTH1600 if applied at ten times the growth inhib-itory concentration in the presence of EDTA (data Polymyxin B increases the permeability of the OMnot shown). to hydrophobic antibiotics (Rosenthal and Storm,
In Fig. 3B, lysis of E. coli strains ATCC 27325 1977). The lethal effect of polymyxin B is the result(K-12), and WBB06 (Re) as well as strains F492 of interaction with negatively charged head groups of(smooth), F470 (R1) and F515 (Re) is shown. the membrane lipids, and subsequent permeabiliza-Protoplasts prepared from wild type strains lysed tion of the cytoplasmic membrane (Teuber and
¨much more readily than those derived from strains Bader, 1976; Schroder et al., 1992). PMBN is awith truncated LPS. Remarkably, this finding is in polymyxin B derivative that lacks the fatty acidcontrast to the effect of nisin against whole cells, residue and one diaminobutyric acid moiety. PMBN
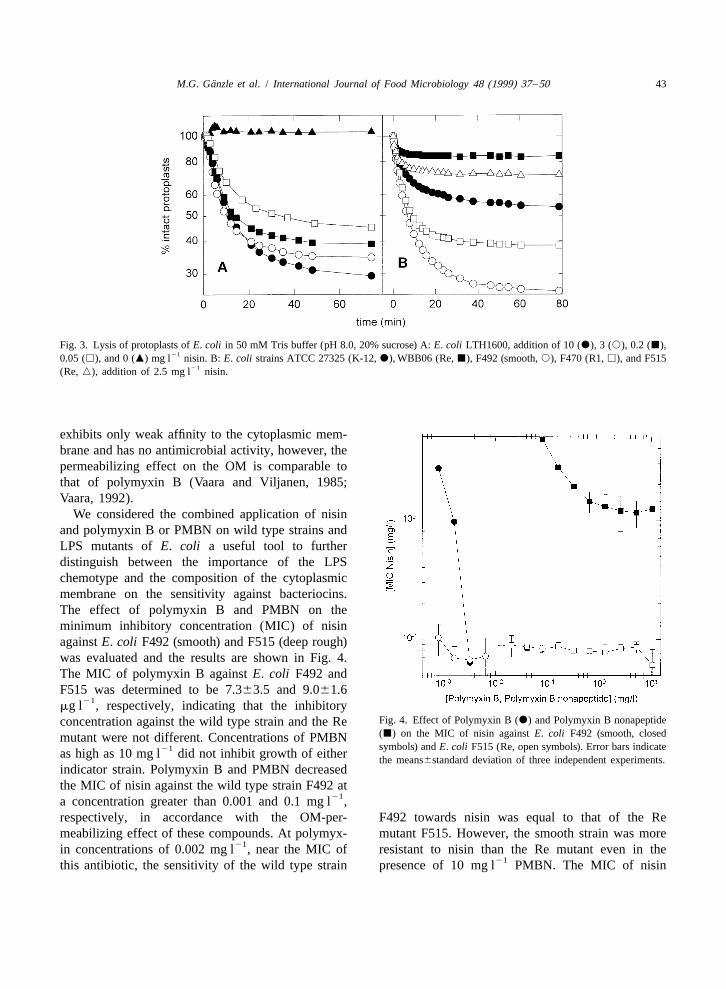
¨M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50 43
Fig. 3. Lysis of protoplasts of E. coli in 50 mM Tris buffer (pH 8.0, 20% sucrose) A: E. coli LTH1600, addition of 10 (d), 3 (s), 0.2 (j),210.05 (h), and 0 (m) mg l nisin. B: E. coli strains ATCC 27325 (K-12, d), WBB06 (Re, j), F492 (smooth, s), F470 (R1, h), and F515
21(Re, n), addition of 2.5 mg l nisin.
exhibits only weak affinity to the cytoplasmic mem-brane and has no antimicrobial activity, however, thepermeabilizing effect on the OM is comparable tothat of polymyxin B (Vaara and Viljanen, 1985;Vaara, 1992).
We considered the combined application of nisinand polymyxin B or PMBN on wild type strains andLPS mutants of E. coli a useful tool to furtherdistinguish between the importance of the LPSchemotype and the composition of the cytoplasmicmembrane on the sensitivity against bacteriocins.The effect of polymyxin B and PMBN on theminimum inhibitory concentration (MIC) of nisinagainst E. coli F492 (smooth) and F515 (deep rough)was evaluated and the results are shown in Fig. 4.The MIC of polymyxin B against E. coli F492 andF515 was determined to be 7.363.5 and 9.061.6
21mg l , respectively, indicating that the inhibitory
Fig. 4. Effect of Polymyxin B (d) and Polymyxin B nonapeptideconcentration against the wild type strain and the Re(j) on the MIC of nisin against E. coli F492 (smooth, closedmutant were not different. Concentrations of PMBNsymbols) and E. coli F515 (Re, open symbols). Error bars indicate21as high as 10 mg l did not inhibit growth of either the means6standard deviation of three independent experiments.
indicator strain. Polymyxin B and PMBN decreasedthe MIC of nisin against the wild type strain F492 at
21a concentration greater than 0.001 and 0.1 mg l ,respectively, in accordance with the OM-per- F492 towards nisin was equal to that of the Remeabilizing effect of these compounds. At polymyx- mutant F515. However, the smooth strain was more
21in concentrations of 0.002 mg l , near the MIC of resistant to nisin than the Re mutant even in the21this antibiotic, the sensitivity of the wild type strain presence of 10 mg l PMBN. The MIC of nisin

¨44 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
against the Re-mutant E. coli F515 remained unaf- effect of Tween 80 and an indirect effect exerted byfected by either polymyxin B or PMBN. the oleate moiety, the strain E. coli LTH1600 was
grown in LB-broth supplemented with oleate orstearate. The killing of E. coli by nisin was moni-
3.3. Effect of the fatty acid supply in the growth tored in citrate buffer (pH 4.75) containing neithermedia on nisin activity against E. coli Tween 80 nor any added fatty acids. As shown in
Fig. 5A, cells pre-grown in LB (oleate) were killedThe nisin activity against E. coli LTH1600 was more readily by nisin than those grown in LB
assayed in LB-broth and mMRS broth. Nisin as well (stearate) or LB without supplement. Thus, theas curvacin A activity against this strain in mMRS availability of oleic acid apparently results inbroth was much greater than in LB-broth (data not changes in the cell envelope of E. coli that rendersshown). To determine the medium components re- the organism more sensitive towards nisin.sponsible for this effect, LB-broth was supplied with To detect changes in the cytoplasmic membraneglucose and Tween 80 at the concentrations of these brought about by the addition of fatty acids, proto-compounds in mMRS, and the bacteriocin activity plasts were prepared from cells grown in LB-brothwas measured. The nisin activity determined in LB- without supplementation or with added oleic, stearic,broth was equal to that observed in mMRS broth if or linoleic acid. The lysis of the protoplasts by nisinTween 80 was supplied to the media, whereas was followed by measuring the optical density in
21 21addition of glucose, Mg or Mn had no effect Tris buffer (pH 8.0) (Fig. 5B). It is striking that the(data not shown). This effect was observed when order of sensitivities was reverted in comparison toTween 80 was added before or after the dilution of the bacteriocin sensitivity of whole cells. Whole cellsnisin on the micro-titerplates was carried out. An were more sensitive when oleic acid was present inincreased nisin activity as result of a reduced nisin the growth medium, but protoplasts prepared fromadsorption to plastic surfaces in the presence of cells grown in the presence of unsaturated fatty acidsTween 80 (Joosten and Nunez, 1995) can thus be were less sensitive to the lytic action of nisin thanexcluded. protoplasts from cells grown in LB-broth without
To differentiate between a possible detergent supplementation.
21 21Fig. 5. A: Killing of E. coli LTH1600 pre-grown in LB-broth (d), LB-broth with 1 g l stearate (j), and LB-broth with 1g l oleate (m)21in citrate buffer at pH 4.75. Closed symbols: 10 mg l nisin added, open symbols: control. B: Lysis of protoplasts of E. coli LTH1600 in
21Tris buffer (pH 8.0, 20% sucrose) containing 5 mg l nisin. The protoplasts were prepared from cells grown to late logarithmic phase in21 21 21LB-broth containing the following additions: none (d), 1 g l stearate (j), 1 g l oleate (m), or 1 g l linoic acid (.).
¨M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50 45
3.4. Effect of pH and NaCl on bacteriocin growth of both organisms was below 4.5. E. coliresistance LTH1600 was able to grow at 7% NaCl, whereas
growth of E. coli LTH4346 (O157:H7) is inhibited3.4.1. Growth inhibition of E. coli by addition of 6% NaCl (data not shown). Adapta-
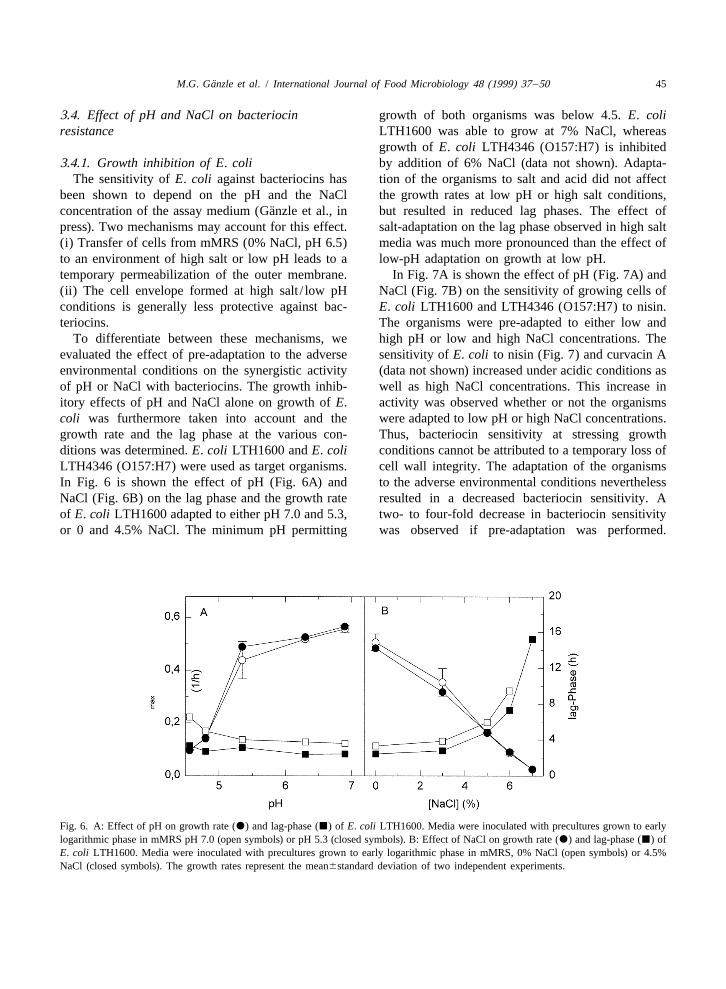
The sensitivity of E. coli against bacteriocins has tion of the organisms to salt and acid did not affectbeen shown to depend on the pH and the NaCl the growth rates at low pH or high salt conditions,
¨concentration of the assay medium (Ganzle et al., in but resulted in reduced lag phases. The effect ofpress). Two mechanisms may account for this effect. salt-adaptation on the lag phase observed in high salt(i) Transfer of cells from mMRS (0% NaCl, pH 6.5) media was much more pronounced than the effect ofto an environment of high salt or low pH leads to a low-pH adaptation on growth at low pH.temporary permeabilization of the outer membrane. In Fig. 7A is shown the effect of pH (Fig. 7A) and(ii) The cell envelope formed at high salt / low pH NaCl (Fig. 7B) on the sensitivity of growing cells ofconditions is generally less protective against bac- E. coli LTH1600 and LTH4346 (O157:H7) to nisin.teriocins. The organisms were pre-adapted to either low and
To differentiate between these mechanisms, we high pH or low and high NaCl concentrations. Theevaluated the effect of pre-adaptation to the adverse sensitivity of E. coli to nisin (Fig. 7) and curvacin Aenvironmental conditions on the synergistic activity (data not shown) increased under acidic conditions asof pH or NaCl with bacteriocins. The growth inhib- well as high NaCl concentrations. This increase initory effects of pH and NaCl alone on growth of E. activity was observed whether or not the organismscoli was furthermore taken into account and the were adapted to low pH or high NaCl concentrations.growth rate and the lag phase at the various con- Thus, bacteriocin sensitivity at stressing growthditions was determined. E. coli LTH1600 and E. coli conditions cannot be attributed to a temporary loss ofLTH4346 (O157:H7) were used as target organisms. cell wall integrity. The adaptation of the organismsIn Fig. 6 is shown the effect of pH (Fig. 6A) and to the adverse environmental conditions neverthelessNaCl (Fig. 6B) on the lag phase and the growth rate resulted in a decreased bacteriocin sensitivity. Aof E. coli LTH1600 adapted to either pH 7.0 and 5.3, two- to four-fold decrease in bacteriocin sensitivityor 0 and 4.5% NaCl. The minimum pH permitting was observed if pre-adaptation was performed.
Fig. 6. A: Effect of pH on growth rate (d) and lag-phase (j) of E. coli LTH1600. Media were inoculated with precultures grown to earlylogarithmic phase in mMRS pH 7.0 (open symbols) or pH 5.3 (closed symbols). B: Effect of NaCl on growth rate (d) and lag-phase (j) ofE. coli LTH1600. Media were inoculated with precultures grown to early logarithmic phase in mMRS, 0% NaCl (open symbols) or 4.5%NaCl (closed symbols). The growth rates represent the mean6standard deviation of two independent experiments.

¨46 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
Fig. 7. A: Effect of pH on nisin activity against E. coli LTH1600 (j) and E. coli O157:H7 LTH4346 (.). Media were inoculated withprecultures grown to early logarithmic phase in mMRS pH 7.0 (open symbols) or pH 5.3 (closed symbols). B: Effect of NaCl on nisinactivity against E. coli LTH1600 (j) and E. coli O157:H7 LTH4346 (.). Media were inoculated with precultures grown to earlylogarithmic phase in mMRS, 0% NaCl (open symbols) or 4.5% NaCl (closed symbols). Lines dropping below the x-axes indicate bacteriocin
21activity below the detection level (2 AU ml ).
Adaptation of E. coli O157:H7 to pH 5.3 did notresult in a increased nisin resistance at low pHvalues. It is remarkable that at all levels of pH andNaCl taken into consideration, E. coli O1547:H7 wasabout ten times as sensitive towards both bac-teriocins than E. coli LTH1600.
3.4.2. Effect of pre-adaptation on bacteriocinresistance of resting cells and protoplasts of E.coli
To confirm that the adaptation to a low pHenvironment increases bacteriocin resistance at lowpH, the killing of resting cells of E. coli LTH1600and LTH4346 (O157:H7) by curvacin A in citratebuffer (pH 4.4) was investigated. Additionally, thesestrains were pre-grown at 4.5% NaCl to assesswhether adaptation to salt stress provides crossprotection against bacteriocin activity at acidic con-ditions. In Fig. 8A is shown the survival of E. coliLTH1600 pre-grown at pH 4.54, 5.35, 6.45, and7.28, or pH 6.45 and 4.5% NaCl. With a decreasingpH of the preculture, E. coli became increasinglyresistant to curvacin A compared to those cellsgrown at a pH near neutral. However, independent of Fig. 8. Killing of E. coli LTH1600 (A) and E. coli O157:H7
LTH4346 (B) in citrate buffer (pH 4.4) with 96 (closed symbols)the pH of the preculture, curvacin A exhibited a21or 0 (open symbols) AU ml curvacin A added. The preculturesbactericidal activity against E. coli. Cells adapted to
were grown to early logarithmic phase in mMRS pH 4.54 (d), pHa high NaCl concentration were less resistant to 5.35 (j), pH 6.45 (m), pH 7.28 (.) or mMRS, pH 6.45, 4.5%curvacin A than cells pre-grown without NaCl. Thus, NaCl (♦). Lines dropping below the x-axes indicate cell counts
24adaptation of E. coli to pH 5.3 decreased the below the detection limit (2310 % surviving cells).

¨M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50 47
bacteriocin sensitivity at pH 4.4, whereas adaptation foods. In our studies, we have taken into accountof the pre-culture to 4.5% NaCl provided no cross additional aspects to elucidate the sensitivity ofprotection to bacteriocin activity at low pH. These enteric bacteria to bacteriocins. The role of the cellresults furthermore confirm that curvacin A is bac- envelope of the organisms was considered by assess-tericidal to E. coli without causing lysis of proto- ment of the bacteriocin sensitivity of LPS mutantplasts. strains and evaluation of the effect of OM per-
In Fig. 8B, the survival of E. coli LTH4346 meabilizing agents. Additionally, effects of environ-(O157:H7) pre-grown at pH 4.50, 6.45, or 6.45 and mental conditions (fatty acid supply during growth,
14.5% NaCl is shown. The results indicate that the H and NaCl concentration) and the physiologicalbacteriocin resistance of E. coli LTH4346 response of E. coli to adverse environmental con-(O157:H7) was comparable to that of strain ditions on bacteriocin sensitivity was examined. TheLTH1600 with respect to the effect of the various nisin concentrations found to be active against E.conditions of the pre-culture. Adaptation to pH 5.3 coli are well within the range of commercial nisin
21decreased bacteriocin sensitivity, and adaptation to application in the food industry, i.e. 1–50 mg l4.5% NaCl increased bacteriocin sensitivity. As (De Vuyst and Vandamme, 1994). The effective doseshown for the bacteriocin activity against growing of curvacin A did not exceed the bacteriocin activitycells, E. coli LTH4346 (O157:H7) was more sensi- of culture supernatant of bacteriocin producing lac-tive to curvacin A than E. coli LTH1600. tobacilli. We considered synergistic effects of bac-
In order to determine the role of the outer mem- teriocins with low pH or high NaCl concentrations asbrane, the nisin mediated lysis of protoplasts pre- these factors are among the most commonly appliedpared from E. coli LTH1600 grown at pH 5.1, 5.5, preservative principles for foods, including those6.5, and 7.5 was determined. The protoplasts were foods such as sausages or meat based salads thatincubated in Tris HCl buffer at pH 8.0 or in citrate allow the application of bacteriocinogenic cultures.buffer at pH 4.8. Protoplasts obtained from cells To establish the relevance of the results to wellgrown at the different pH values were not affected in recognized foodborne pathogens, E. coli O157:H7their lytic behavior in the presence of nisin (data not was used as target organism in addition to non-shown). No appreciable differences were observed pathogenic wild type strains. Thus, the results maybetween lysis of protoplasts prepared from E. coli provide a rationale to allow the development ofLTH1600 or E. coli LTH O157:H7 (data not shown). feasible processes employing bacteriocins to improveThus, the differences in sensitivity to nisin and the hygienic status of food.curvacin A between these strains is likely to be due The outer membrane composition of LPS mutantto the different outer membrane composition. In strains and the role of mutant LPS in the suscep-direct contrast to nisin activity against whole cells, tibility against therapeutic antibiotics is well char-the lytic activity of nisin against protoplasts was acterized (Vaara, 1993; Nikaido, 1996). Deep roughmore pronounced at pH 8.0 than at 4.8. mutants are characterized by a decreased protein
content of the outer membrane and the occurrence ofphospholipid patches on the outer leaflet of the OM
4. Discussion (Koplow and Goldfine, 1974; Smit et al., 1975;Gmeiner and Schlecht, 1980; Nikaido, 1996). The
The inhibitory spectra of bacteriocins of lactic acid biophysical structure of mutant LPS differs from thatbacteria generally do not include Gram-negative or wild type LPS as it has a higher tendency to formbacteria. Thus, processes that render these micro- non-lammellar structures, and the hydrophobicity oforganisms sensitive to bacteriocins can greatly en- the molecules is decreased (Seydel et al., 1993). Thehance the applications of bacteriocins. Combination susceptibility of deep rough mutants to otherwiseof bacteriocins with chelators or physical disruption inactive antibiotics has been attributed to theseof the cells have been successfully applied by Shefet phospholipids on the cell surface (Vaara et al., 1990;et al. (1995) and Kalchayanand et al. (1992) to kill Nikaido, 1996; Vaara, 1993). We found all LPSE. coli. However, these processes may not be mutants with Re chemotype highly susceptible tofeasible for a wide range of minimally processed nisin and curvacin A, indicating that the truncated

¨48 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
LPS with a more disordered structure and phos- synergism was observed with the Re mutant, indicat-pholipids facing outside greatly facilitates access of ing that the outer membrane of the latter organismbacteriocins to the cytoplasmic membrane. The provides no appreciable protection against nisin.acquisition of sensitivity by Re mutants is compar- PMBN failed to decrease the MIC of nisin of the
21able to that of EDTA-treated wild type cells (Vaara, smooth strain (1 mg l ) to the corresponding level211992; Nikaido, 1996), and our results thus confirm of the Re mutant (0.1 mg l ), indicating that the
the synergistic activity of EDTA and nisin on E. coli LPS of the smooth E. coli strain provides protection¨(Shefet et al., 1995; Schved et al., 1994; Ganzle et against nisin even in the presence of PMBN. In
al., 1999). contrast, the MIC of several antibiotics of smooth S.In LPS mutants with Rc and Rb chemotype, enterica serovar Typhimurium in the presence of
phospholipids are not found on the outer leaflet of PMBN were the same as those of the correspondingthe OM or are not accessible from outside, e.g. by Re mutant (Vaara and Vaara, 1983a).phospholipases (Smit et al., 1975; Van Alphen et al., Supplementation of growth media with unsatu-1977). The protein and phospholipid content of their rated fatty acids increased the bacteriocin sensitivityOM is comparable to wild type strains rather than of growing and resting cells of E. coli. Based on thedeep rough mutants (Smit et al., 1975). However, we observation that the OM composition is crucial forfound the susceptibility to nisin or curvacin A of S. bacteriocin activity, it can be assumed that the fattyenterica with Rb chemotype, E. coli K-12 and R1 acid composition of the LPS was altered to facilitate3
to be similar to that of the corresponding deep rough the diffusion of bacteriocins through the outer mem-mutants. These findings indicate that the occurrence brane. Marked differences were observed for theof phospholipids on the cell surface and the bio- lytic behavior of protoplasts prepared from LB, LBphysical changes occurring in truncated LPS are not (stearate) or LB (oleate) which point to an altereda prerequisite for bacteriocin susceptibility of enteric composition of the cytoplasmic membrane as well.bacteria. This finding may be of importance for the elimina-
The lytic activity of nisin towards propoplasts tion of E. coli in food as most plant fat used for foodprepared from LPS mutant strains was directly production contains a high amount of unsaturatedopposite to the sensitivity of growing cells against and polyunsaturated fatty acids.the bacteriocin. The growth inhibitory activity of As pointed out in the introduction of this paper, E.bacteriocins, brought about by pore formation in the coli, including strains of the pathogenic E. colicytoplasmic membrane, does not necessarily corre- O157:H7, has an especially high tolerance to adverselate with their lytic activity. Both nisin and curvacin environmental conditions such as low pH, high saltA were bactericidal to E. coli, but only nisin was concentration, and prolonged storage at refrigerationfound to lyse protoplasts. However, our data are temperatures. Remarkably, the E. coli O157:H7 wasconsistent with the assumption that the cytoplasmic more susceptible to both nisin and curvacin than E.membrane of LPS mutants is more resistant towards coli LTH1600. We therefore expect that bacteriocinsnisin due to the incorporation of LPS into the of lactic acid bacteria may contribute to the killing ofcytoplasmic membrane. Whole cells of LPS mutants this organism in foods.are nevertheless more sensitive towards nisin than Adaptation to acid and NaCl decreased the bac-the corresponding wild type cells which possess an teriocin sensitivity of E. coli at low pH and high saltintact OM permeability barrier. concentrations, respectively. However, the bac-
Polymyxin B and PMBN act synergistically with teriocin sensitivities of acid and salt adapted cellsother antibiotics due to interactions with the outer were nevertheless increased up to 100-fold by in-
1membrane of Gram-negative cells, causing the loss creasing H and NaCl concentrations. A change ofof a functional permeability barrier without release the composition of the cytoplasmic membrane isof outer and inner membrane components (Rosenthal unlikely to account for the decreased sensitivity ofand Storm, 1977; Vaara and Vaara, 1983a,b; Dixon acid adapted cells. No differences could be detectedand Chopra, 1986). We found synergistic effects of in the lytic activity of nisin against protoplastspolymyxin B of PMBN with nisin only if the smooth prepared from cells grown at different pH values.E. coli strain was used as target organism. No Adaptation of E. coli to acid and NaCl results in

¨M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50 49
S al and growth of Escherichia coli O157:H7 in ground, roastedsimilar changes in the fatty acid composition, i.e. sbeef as affected by pH, acidulants, and temperature. Appl.mediated increase of cyclopropane fatty acidsEnviron. Microbiol. 59, 2364–2368.
(McGarrity and Armstrong, 1975; Brown et al., Bearson, S., Bearson, B., Foster, J.W., 1997. Acid stress responses1997; Magnuson et al., 1993), but NaCl adaptation in enterobacteria. FEMS Microbiol. Lett. 147, 173–180.
Benjamin, M.M., Datta, A.R., 1995. Acid tolerance of en-provides no cross protection towards bacteriocin atterohemorrhagic Escherichia coli. Appl. Environ. Microbiol.acidic pH. We found the lytic activity of nisin higher61, 1669–1672.
at pH 8.0 than at pH 4.8. This is in agreement with ¨Brabetz, W., Muller-Loennies, S., Holst, O., Brade, H., 1997.data of Winkowski et al. (1996) who reported that Deletion of the heptosyltransferase genes rfaC and rfaF inleakage of carboxyfluorescin from Listeria mono- Escherichia coli K-12 results in an Re-type lipopolysaccharide
with a high degree of 2-aminoethanol phosphate substitution.cytogenes derived liposomes was strongly decreasedEur. J. Biochem. 247, 716–724.at low pH although nisin activity against whole cells
Brown, J.L., Ross, T., McMeekin, T.A., Nichols, P.D., 1997. Acidwas increased. habituation of Escherichia coli and the potential role of
A possible explanation for the increased bac- cyclopropane fatty acids in low pH tolerance. Int. J. Food.1teriocin activity towards E. coli at elevated H and Microbiol. 37, 163–173.
1 Buchanan, R.L., Doyle, M.P., 1997. Foodborne disease signifi-Na concentrations is the replacement of divalentcance of Escherichia coli O157:H7 and other enterohemor-cations from their LPS binding sites, rendering cellsrhagic E. coli. Food Technol. 51 (10), 69–76.
more susceptible (Vaara, 1992). However, this mech- Cheville, A.M., Arnold, K.W., Buchrieser, C., Cheng, C.M.,anism does not account for the effect of pre-adapta- Kaspar, C.W., 1996. rpoS regulation of acid, heat, and salttion on bacteriocin activity, so that alterations of the tolerance in Escherichia coli O157:H7. Appl. Environ. Mi-
crobiol. 62, 1822–1824.LPS chemotype upon exposure to acid and salt stressDe Man, J.C., Rogosa, M., Sharpe, M.E., 1960. A medium for thecan be expected. It must be further taken into
cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130–135.account that, as the outer membrane does not com- De Vuyst, L., Vandamme, E.J., 1994. Nisin, a lantibiotic producedpletely prevent influx of inhibitors, the resistance of by Lactococcus lactis supsp. lactis: properties, biosynthesis,E. coli depends on energy dependent efflux systems fermentation and applications. In: De Vuyst, J., Vandamme, E.J.
(Eds.), Bacteriocins of Lactic Acid Bacteria. Microbiology,which use the pH component of the proton motiveGenetics, and Applications, Chapman and Hall, London.force (pmf) to transport a wide range of unrelated
Dixon, R., Chopra, I., 1986. Leakage of periplasmic proteins fromamphiphilic substrates (Fralick and Burns-Keliher, Escherichia coli mediated by polymyxin B nonapeptide. Anti-1994; Nikaido, 1994; Thanassi et al., 1997). The microb. Agents Chemother. 29, 781–788.application of salt and acid stress is likely to Fralick, J.A., L Burns-Keliher, L., 1994. Additive effect of tolC
and rfa mutations on the hydrophobic barrier of the outeradversely affect the efficacy of these pmf dependentmembrane of Escherichia coli K-12. J. Bacteriol. 176, 6404–transport systems, resulting in increased bacteriocin6406.
sensitivity of the cells. ¨Ganzle, M.G., Weber, S., Hammes, W.P., 1999. Effect of ecologi-cal factors on the inhibitory spectrum and activity of bac-teriocins. Intern. J. Food Microbiol. 46, 207–217.
Glass, K.A., Loeffelholz, J.M., Ford, J.P., Doyle, M.P., 1992. FateAcknowledgementsof Escherichia coli O157:H7 as affected by pH or sodiumchloride and in fermented, dry sausage. Appl. Environ. Mi-
We wish to thank Dr. Werner Brabetz, crobiol. 58, 2513–2516.Gmeiner, J., Schlecht, S., 1980. Molecular composition of the¨Forschungszentrum Borstel, Institut fur Experimen-
outer membrane of Escherichia coli and the importance oftelle Biologie und Medizin, Borstel, for providingprotein-lipopolysaccharide interactions. Arch. Microbiol. 127,bacterial strains, and for the helpful discussions81–86.
during the work. Hammes, W.P., Hertel, C., 1998. New developments in meatstarter cultures. Meat Sci. 149, S125–S138.
Joosten, H.M.L.J., Nunez, M., 1995. Adsorption of nisin andenterocin 4 to polypropylene and glass surfaces and its
References prevention by Tween 80. Lett. Appl. Microbiol. 21, 389–392.Kalchayanand, N., Hanlin, M.B., Ray, B., 1992. Sublethal injury
Abdul-Raouf, U.M., Beuchat, L.R., Ammar, M.S., 1993a. Survival makes Gram-negative and resistant Gram-positive bacteriaand growth of Escherichia coli O157:H7 on salad vegetables. sensitive to the bacteriocins, pediocin AcH and nisin. Lett.Appl. Environ. Microbiol. 59, 1999–2006. Appl. Microbiol. 15, 239–243.
Abdul-Raouf, U.M., Beuchat, L.R., Ammar, M.S., 1993b. Surviv- Koplow, J., Goldfine, H., 1974. Alterations in the outer membrane

¨50 M.G. Ganzle et al. / International Journal of Food Microbiology 48 (1999) 37 –50
of the cell envelope of heptose-deficient mutants of Es- Seydel, U., Koch, M.H.J., Brandenburg, K., 1993. Structuralcherichia coli. J. Bacteriol. 117, 527–543. polymorphisms of rough mutant lipopolysaccharides Rd to Ra
Leyer, G.J., Wang, L.L., Johnson, E.A., 1995. Acid adaptation of from Salmonella minnesota. J. Structural Biol. 110, 232–243.Escherichia coli O157:H7 increases survival in acidic foods. Shefet, S.M., Sheldon, B.W., Klaenhammer, T.R., 1995. EfficacyAppl. Environ. Microbiol. 61, 3752–3755. of optimized nisin-based treatments to inhibit Salmonella
Magnuson, K., Jackowski, S., Rock, C.O., Cronan, Jr. J.E., 1993. typhimurium and extend shelf life of broiler carcasses. J. Food.Regulation of fatty acid biosynthesis in Escherichia coli. Prot. 58, 1077–1082.Microbiol. Rev. 57, 522–542. Small, P., Blankenhorn, D., Welty, D., Zinser, E., Slonczewski,
McGarrity, J.T., Armstrong, J.B., 1975. The effect of salt on J.L., 1994. Acid and base resistance in Escherichia coli andphospholipid fatty acid composition in Escherichia coli K12. Shigella flexneri: role of rpoS and growth pH. J. Bacteriol. 176,Biochim. Biophys. Acta. 398, 258–264. 1729–1737.
Nikaido, H., 1994. Prevention of drug access to bacterial targets: Smit, J., Kamio, Y., Nikaido, H., 1975. Outer membrane ofpermeability barriers and active efflux. Science 264, 382–387. Salmonella typhimurium: chemical analysis and freeze-fracture
Nikaido, H., 1996. Outer membrane. In: Neidhardt, F.C. (Ed.), studies with lipopolysaccharide mutants. J. Bacteriol. 124,Escherichia and Salmonella: Cellular and Molecular Biology, 942–958.2nd ed, ASM Press, Washington DC, Chapter 5. Stevens, K.A., Sheldon, B.W., Klapes, N.A., Klaenhammer, T.R.,
Parente, E., Brienza, C., Moles, M., Ricciardi, A., 1995. A 1991. Nisin treatment for inactivation of Salmonella speciescomparison of methods for the measurement of bacteriocin and other Gram-negative bacteria. Appl. Environ. Microbiol.activity. J. Microbiol. Methods 22, 95–108. 57, 3612–3615.
Presser, K.A., Ross, T., Ratkowsky, D.A., 1998. Modelling the Stiles, M.E., 1996. Biopreservation by lactic acid bacteria. An-growth limits (growth/no growth interface) of Escherichia coli tonie van Leeuwenhoek 70, 331–345.as a function of temperature, pH, lactic acid concentration, and Teuber, M., Bader, J., 1976. Action of polymyxin B on bacterialwater activity. Appl. Environ. Microbiol. 64, 1773–1779. membranes. Arch. Microbiol. 109, 51–58.
Reitsma, C.J., Henning, D.R., 1996. Survival of enterohemor- Thanassi, D.G., Cheng, L.W., Nikaido, H., 1997. Active efflux ofrhagic Escherichia coli O157:H7 during the manufacture and bile salts by Escherichia coli. J. Bacteriol. 179, 2512–2518.curing of cheddar cheese. J. Food. Prot. 59, 460–464. Tichaczek, P.S., Nissen-Meyer, J., Nes, I.F., Vogel, R.F., Hammes,
Roantree, R.J., Kuo, T.T., MacPhee, D.G., 1977. The effect of W.P., 1992. Characterization of the bacteriocins curvacin Adefined lipopolysaccharide core defects upon antibiotic re- from Lactobacillus curvatus LTH1174 and sakacin P from L.sistances of Salmonella Typhimurium. J. Gen. Microbiol. 103, sake LTH673. Sys. Appl. Microbiol. 15, 460–468.223–234. Vaara, M., 1992. Agents that increase the permeability of the outer
Rosenthal, K.S., Storm, D.R., 1977. Disruption of the Escherichia membrane. Microbiol. Rev. 56, 395–411.coli outer membrane permeability barrier by immobilized Vaara, M., 1993. Antibiotic supersusceptible mutants of Es-polymyxin B. J. Antibiotics 30, 1087–1092. cherichia coli and Salmonella typhimurium. Antimicrob.
Rowbury, R.J., 1995. An assessment of environmental factors Agents Chemother. 37, 2255–2260.influencing acid tolerance and sensitivity in Escherichia coli, Vaara, M., Viljanen, P., 1985. Binding of polymyxin B nonapep-Salmonella spp. and other enterobacteria. Lett. Appl. Mi- tide to Gram-negative bacteria. Antimicrob. Agents Chemother.crobiol. 20, 333–337. 27, 548–554.
Sanderson, K.E., Van Wyngaarden, J., Luderitz, O., Stocker, B.A., Vaara, M., Vaara, T., 1983a. Polycations sensitize enteric bacteria1974. Rough mutants of Salmonella typhimurium with defects to antibiotics. Antimicrob. Agents Chemother. 24, 107–113.in the heptose region of the lipopolysaccharide core. Can. J. Vaara, M., Vaara, T., 1983b. Polycations as outer membraneMicrobiol. 20, 1127–1134. disorganizing agents. Antimicrob. Agents Chemother. 24, 114–
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Clon- 122.ing. A Laboratory Manual, 2nd ed, Cold Spring Harbor Vaara, M., Plachy, W.Z., Nikaido, H., 1990. Partitioning ofLaboratory Press. hydrophobic probes into lipopolysaccharide bilayers. Biochim.
Scanell, A.G.M., Hill, C., Buckley, D.J., Arendt, E.K., 1997. Biophys. Acta 1024, 152–158.Determination of the influence of organic acids and nisin on Van Alphen, L., Lugtenberg, B., van Boxtel, R.,Verhoef, K., 1977.shelf-life and microbiological safety aspects of fresh pork. J. Architecture of the outer membrane of Escherichia coli K12: I.Appl. Microbiol. 83, 407–412. Action of phospholipases A2 and C on wild type strains and
Schmidt, G., Jann, B., Jann, K., 1970. Immunochemistry of R outer membrane mutants. Biochim. Biophys. Acta 466, 257–lipopolysaccharides of Escherichia coli. Eur. J. Biochem. 16, 268.382–392. Winkowski, K., Ludescher, R.D., Montville, T.J., 1996. Physico-
¨Schroder, G., Brandenburg, K., Seydel, U., 1992. Polymyxin B chemical characterization of the nisin-membrane interactioninduces transient permeability fluctuations in asymmetric pla- with liposomes derived from Listeria monocytogenes. Appl.nar lipopolysaccharide /phospholipid bilayers. Biochemistry 31, Environ. Microbiol. 62, 323–327.631–638. Zwietering, M.H., Jongenburger, I., Rombouts, F.M., van’t Riet,
Schved, F., Henis, Y., Juven, B.J., 1994. Response of spheroplasts K., 1990. Modeling of the bacterial growth curve. Appl.and chelator-permeabilized cells of Gram-negative bacteria to Environ. Microbiol. 56, 1875–1881.the action of the bacteriocins pediocin SJ-1 and nisin. Int. J.Food. Microbiol. 21, 305–314.
Related Documents











