Annals of Applied Biology ISSN 0003-4746 RESEARCH ARTICLE Transcription factor profiling identifies an aleurone-preferred NAC family member involved in maize seed development N.C. Verza 1,2 , T.R.S. Figueira 1 , S.M. Sousa 1,3 & P. Arruda 1,4 1 Centro de Biologia Molecular e Engenharia Gen ´ etica, Universidade Estadual de Campinas (UNICAMP), Campinas, SP, Brazil 2 Present address: Amyris Pesquisa e Desenvolvimento de Biocombust´ ıveis, Rua James Clerk Maxwell, Campinas, SP, Brazil 3 Present address: Embrapa Milho e Sorgo, Sete Lagoas, MG, Brazil 4 Departamento de Gen ´ etica e Evoluc ¸˜ ao, Instituto de Biologia, Universidade Estadual de Campinas (UNICAMP), Campinas, SP, Brazil Keywords ABA- and GA-responsive gene; aleurone; maize endosperm; transcription factors; transcriptome. Correspondence P. Arruda, Departamento de Gen ´ etica e Evoluc ¸˜ ao, Instituto de Biologia, Universidade Estadual de Campinas, CP 6109, 13083-970, Campinas, SP, Brazil. Email: [email protected] Received: 24 July 2010; revised version accepted: 24 August 2010. doi:10.1111/j.1744-7348.2010.00447.x Abstract Transcription factors (TFs) preferentially expressed in developing endosperm were identified by searching a maize transcriptome database. One thousand and two hundred TFs were annotated, 414 of which were shown to express in the seeds. Of these, 113 presented preferential expression in developing endosperm. Among the endosperm-preferred TFs 13% were represented by members of the zinc-finger family, 17% by members of the bZIP family and 11% by members of the NAC family. Among the NAC family TFs, we identified APN-1, whose expression was detected 5 days after pollination (DAP) and peaked at 20 DAP. Analysis of the promoter sequence of gene encoding APN-1 revealed the presence of cis-elements related to endosperm-specificity, abscisic acid (ABA) and gibberellic acid (GA) signalling. Transient expression analysis of a construct containing the β -glucuronidase gene driven by the Apn-1 gene promoter showed that expression was restricted to the aleurone cell layer. Analysis of mRNA levels in developing maize endosperm treated with hormones revealed that the expression of the gene encoding APN-1 was stimulated by ABA and inhibited by GA. Introduction The processes underlying seed development and germi- nation require the concerted action of genes that are reg- ulated by transcription factors (TFs) (Schwechheimer & Bevan, 1998). Transcription factors are sequence-specific DNA-binding proteins that are capable of activating or repressing transcription of functionally related genes. They are responsible for the selective gene regulation, and are often expressed in a tissue-specific, developmental- stage-specific or stimulus-dependent manner (Pabo & Sauer, 1992; Riechmann & Ratcliffe, 2000; Gong et al., 2004). Transcription factors are generally grouped into families according to their DNA-binding domains (Pabo & Sauer, 1992; Bowman, 2000; Riechmann & Ratcliffe, 2000; Gong et al., 2004). The largest TF family found in the eukaryotes contains a DNA-binding motif known as zinc finger (Pabo & Sauer, 1992). On the basis of the structure and spatial arrangement of the zinc finger domain, this gene family can be further subdivided into several classes (Souer et al., 1996; Bowman, 2000; Riechmann et al., 2000). An important TF class in plants is the bZIP family that accounts for 5–6% of the TF content in Arabidopsis and rice. Another interesting TF group unique to plants is the NAC family (Riechmann et al., 2000). Members of this family are involved in developmental processes, including formation of the shoot apical meristem, floral organs and lateral shoots, as well as in stress responses and plant defence against diseases (Souer et al., 1996; Aida et al., 1997, Zimmermann & Werr, 2005). Two maize paralogues, ZmNAM1 and ZmNAM2, as well as the petunia PhNAM and Arabidopsis AtCUC2 contribute to shoot apical meristem establishment, whereas the maize ZmCUC3 is associated with leaf primordium development (Zimmermann & Werr, 2005). The expression of genes Ann Appl Biol 158 (2011) 115 – 129 © 2010 The Authors 115 Annals of Applied Biology © 2010 Association of Applied Biologists

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Annals of Applied Biology ISSN 0003-4746
R E S E A R C H A R T I C L E
Transcription factor profiling identifies an aleurone-preferredNAC family member involved in maize seed developmentN.C. Verza1,2, T.R.S. Figueira1, S.M. Sousa1,3 & P. Arruda1,4
1 Centro de Biologia Molecular e Engenharia Genetica, Universidade Estadual de Campinas (UNICAMP), Campinas, SP, Brazil
2 Present address: Amyris Pesquisa e Desenvolvimento de Biocombustıveis, Rua James Clerk Maxwell, Campinas, SP, Brazil
3 Present address: Embrapa Milho e Sorgo, Sete Lagoas, MG, Brazil
4 Departamento de Genetica e Evolucao, Instituto de Biologia, Universidade Estadual de Campinas (UNICAMP), Campinas, SP, Brazil
KeywordsABA- and GA-responsive gene; aleurone;
maize endosperm; transcription factors;
transcriptome.
CorrespondenceP. Arruda, Departamento de Genetica e
Evolucao, Instituto de Biologia, Universidade
Estadual de Campinas, CP 6109, 13083-970,
Campinas, SP, Brazil.
Email: [email protected]
Received: 24 July 2010; revised version
accepted: 24 August 2010.
doi:10.1111/j.1744-7348.2010.00447.x
Abstract
Transcription factors (TFs) preferentially expressed in developing endospermwere identified by searching a maize transcriptome database. One thousandand two hundred TFs were annotated, 414 of which were shown to expressin the seeds. Of these, 113 presented preferential expression in developingendosperm. Among the endosperm-preferred TFs 13% were represented bymembers of the zinc-finger family, 17% by members of the bZIP family and11% by members of the NAC family. Among the NAC family TFs, we identifiedAPN-1, whose expression was detected 5 days after pollination (DAP) andpeaked at 20 DAP. Analysis of the promoter sequence of gene encodingAPN-1 revealed the presence of cis-elements related to endosperm-specificity,abscisic acid (ABA) and gibberellic acid (GA) signalling. Transient expressionanalysis of a construct containing the β-glucuronidase gene driven by theApn-1 gene promoter showed that expression was restricted to the aleuronecell layer. Analysis of mRNA levels in developing maize endosperm treatedwith hormones revealed that the expression of the gene encoding APN-1 wasstimulated by ABA and inhibited by GA.
Introduction
The processes underlying seed development and germi-
nation require the concerted action of genes that are reg-
ulated by transcription factors (TFs) (Schwechheimer &
Bevan, 1998). Transcription factors are sequence-specific
DNA-binding proteins that are capable of activating or
repressing transcription of functionally related genes.
They are responsible for the selective gene regulation, and
are often expressed in a tissue-specific, developmental-
stage-specific or stimulus-dependent manner (Pabo &
Sauer, 1992; Riechmann & Ratcliffe, 2000; Gong
et al., 2004).
Transcription factors are generally grouped into
families according to their DNA-binding domains (Pabo
& Sauer, 1992; Bowman, 2000; Riechmann & Ratcliffe,
2000; Gong et al., 2004). The largest TF family found in the
eukaryotes contains a DNA-binding motif known as zinc
finger (Pabo & Sauer, 1992). On the basis of the structureand spatial arrangement of the zinc finger domain, thisgene family can be further subdivided into several classes(Souer et al., 1996; Bowman, 2000; Riechmann et al.,2000). An important TF class in plants is the bZIP familythat accounts for 5–6% of the TF content in Arabidopsisand rice. Another interesting TF group unique to plantsis the NAC family (Riechmann et al., 2000). Membersof this family are involved in developmental processes,including formation of the shoot apical meristem, floralorgans and lateral shoots, as well as in stress responsesand plant defence against diseases (Souer et al., 1996;Aida et al., 1997, Zimmermann & Werr, 2005). Twomaize paralogues, ZmNAM1 and ZmNAM2, as well as thepetunia PhNAM and Arabidopsis AtCUC2 contribute toshoot apical meristem establishment, whereas the maizeZmCUC3 is associated with leaf primordium development(Zimmermann & Werr, 2005). The expression of genes
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 115Annals of Applied Biology © 2010 Association of Applied Biologists

Transcription factor profiling in maize endosperm N.C. Verza et al.
encoding NACs have also been shown to respond toabscisic acid (ABA), whereas others act as negativeregulators of the response to gibberellic acid (GA) (Aidaet al., 1997; Fujita et al., 2004).
To date there are only few TFs associated with maizeseed ontogeny. One example is VP-1 that is required forABA induction of maturation and tolerance to desiccation(Hoecker et al., 1999; Suzuki et al., 2008). VP-1 inhibitsthe expression of germination-specific α-amylase genesin aleurone cells, preventing precocious hydrolysis ofstorage compounds in the endosperm (Hoecker et al.,1999). The gene encoding VP-1 is weakly expressed in thestarchy endosperm but is highly expressed in the aleuroneand embryo during seed maturation (Hoecker et al.,1999). VP-1 also regulates TRAB-1, which interacts withVP-1 to recognise ABA-responsive elements (ABREs) andmediate ABA signalling (Hobo et al., 1999). Other TFssuch as GAMYB, BPBF and SAD participate in seedmaturation and the post-germination phases (Gubleret al., 1995).
In this study we identified TFs that are preferentiallyexpressed in developing maize endosperm. One of theseendosperm-preferred TFs belongs to the NAC family andwas called APN-1. The gene encoding APN-1 was shownto be preferentially expressed in the aleurone cell layerand its expression was induced by ABA and repressedby GA. The results are discussed in the context of theregulatory process encompassing seed maturation andgermination.
Materials and methods
Plant material
Maize (Zea mays L.) plants from the F352 inbred line weregrown under standard greenhouse conditions. All plantswere individually hand pollinated. Maize seeds wereharvested at 5, 10, 15 and 20 days after pollination (DAP),frozen in liquid nitrogen and stored at −80◦C. Embryoswere dissected manually. Roots, leaves and coleoptileswere harvested from 5-day-old seedlings germinatedunder controlled conditions. The 10 DAP seeds for thetransient assays were used immediately after harvest.
Transcription factor identification
Transcription factors were identified by searches in theMAIZEST database (Verza et al., 2005) using annotatedTF sequences from TRANSFAC (Matys et al., 2003),GenBank (Benson et al., 2005) and Pfam databases(Bateman et al., 2004). We combined automated searchand manual inspection to generate a collection ofendosperm-expressed TFs. First, the TF sequences ofTRANSFAC were BLASTx against the 29 206 assembled
sequences of the MAIZEST database (Verza et al., 2005).Only matches with an E value ≤ 1.e-15 were retrieved.The retrieved sequences of the first run were BLASTxagainst the GenBank non-redundant proteins database.Then the selected sequences were manually inspected,and false positive matches including proteases, splicingfactors, kinases, translation factors and other non-TFproteins were removed. Next, the 29 206 assembledsequences of MAIZEST were BLASTx against Pfam, usingthe default settings (Bateman et al., 2004). Matches withTF motifs not identified by the previous analyses wereretrieved and allocated in one of the TRANSFAC classesor included in the ’Other’ group.
Identification of endosperm-preferred TFs
The MAIZEST database was constructed by sequencing30 531 Expressed Sequence Tags (ESTs) from 10, 15and 20 DAP developing endosperm and by maize ESTsfrom MaizeGDB (http://www.maizegdb.org/est.php) andGenoplante (http://genoplante-info.infobiogen.fr) (Verza et al., 2005). Alto-gether, the database contained 227 431 ESTs, 64 537 ofwhich were from developing endosperm and the remain-ing from BMS cell suspension cultures, field-grownunpollinated ears, embryos, glume, leaf primordium, api-cal meristem, premeiotic anthers, mature pollen, rootsand stem, among other tissues. To select the ESTswith endosperm-preferred expression, bioinformatic toolswere used to retrieve from the database the assembledsequences composed of ESTs originated exclusively fromendosperm cDNA libraries.
Isolation of the Apn-1 promoter, vector constructionand transient expression assays in immature maizeendosperm
A 1.9 kb fragment of the Apn-1 promoter was amplifiedfrom the maize F352 genomic DNA using the prim-ers ZmNAMprofwd (5′-CCAGTCAACATAGCCCAACT-3′) and ZmNAMprorev (5′-GAGGTCAGTCCTCGAGTCAGAGA-3′). A single polymerase chain reaction (PCR)product was isolated from agarose gel and then subclonedinto the pGEM-T EASY vector (Promega, USA). Thevector was subsequently digested with HincII and XhoIand the released fragment cloned into the promoter-less pRT103GUS vector (Yunes et al., 1994). Maize earsharvested at 20 DAP were surface sterilised for 15 minwith 5% commercial bleach and rinsed four times indistilled water. Seeds were dissected from the cobs andthe caps were manually dissected by removing the entirepericarp and half of the aleurone cell layer. Nine capswere placed in the centre of 100 mm diameter Petri dishes
116 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists

N.C. Verza et al. Transcription factor profiling in maize endosperm
containing 20 mL of MS medium, with the cap surfacefacing upward. Five micrograms of column-purifiedDNAs of constructs pApn1::GUS and p35S::GUS (used ascontrol) were used to coat 3 mg of 1–3 μm diameter goldparticles (Yunes et al., 1994). The endosperm caps werebombarded twice using a high-pressure helium gene gun(Yunes et al., 1994). After bombardment, the endospermswere incubated for 24 h in the dark at 25◦C. Theendosperms were then stained for GUS activity (Jeffersonet al., 1987). Endogenous β-glucuronidase activity wasinhibited by adding methanol to the assay mixture to afinal concentration of 20% (Kosugi et al., 1990).
RNA isolation
Total RNA was isolated from frozen endosperm andembryo by lithium chloride method, with minormodifications (Manning, 1991). For roots, leaves andcoleoptiles total RNA was isolated using the Trizol reagent(Invitrogen, Carlsbad, CA). The purity and integrity of theRNA were verified by the absorbance at 260/280 nm andagarose gel electrophoresis.
Effect of ABA and GA on Apn-1 expression
RNA samples were treated with RQ1 RNAse-free DNase I(Promega, Madison, WI) to eliminate contaminatinggenomic DNA. Reverse trascription polymerase chainreaction (RT-PCR) was carried out with 500 ng ofDNAse-treated total RNA using the one-step AccessRT-PCR System (Promega). The products were sepa-rated by electrophoresis in agarose gels and visualisedby ultraviolet (UV) excitation of ethidium bromide-stained DNA. The primers used to amplify the Apn-1transcript were ZmESN1fwd (5′-CATGGCGGCGGACC-3′) and ZmESN1rev (5′-GATGGCGTGTGGAAGTACTGA-3′), and the control primers were Actinfwd (5′-CATGCTTCTCTCTGCAGGCTGG-3′) and Actinrev (5′-TCATTAGGCA GTCAGTCAGATCACG-3′).
For hormone treatments, maize kernels harvested at 10DAP were surface sterilised with 1% sodium hypochloritefor 15 min, cut into two halves and placed with the innerside facing down on a plate containing MS mediumsupplemented with ABA (25 and 75 μM) or GA (25 and75 μM) (Sigma Chemical Co, St. Louis, USA). The plateswere incubated for 6 h in the dark at 25◦C. Hormonetreatments were performed in triplicate.
RNA from the hormone-treated kernels was isolatedas described above. For qPCR analysis, the primersused were forward: 5′-GCGACTTCGAGATTGTCAGCTT-3′ and reverse: 5′-ATGGCGGTGCAGGTGAA-3′. Apn-1expression was normalised relative to the expressionof 18S rRNA that was analysed in separate reactions
using TaqMan Ribosomal RNA Control Reagents – VICProbe (PE Applied Biosystems). The relative geneexpression was performed using the 2-DDCt method(Livak & Schmittgen, 2001). Reactions for the first-strandwere prepared using the TaqMan Reverse TranscriptionReagent (Applied Biosystems). A SYBR green PCR MasterMix (AB Applied Biosystems, Roche) was used for thesecond step, together with 10 μM of each primer and25 ng of cDNA.
Results
Transcription factors expressed in developing maizeendosperm
An EST database rich in endosperm transcripts (Verzaet al., 2005) was inspected for TFs expressing in thedeveloping maize seeds. The database comprised 227 431ESTs from cDNA libraries prepared from endospermsampled at 10, 15 and 20 DAP along with ESTs fromBMS cell suspension cultures, field-grown unpollinatedears, embryos, glume, leaf primordium, apical meristem,premeiotic anthers, mature pollen, roots and stem,among others tissues (Verza et al., 2005). Clusteringthese ESTs resulted in 29 206 assembled sequencesputatively encoding individual maize genes (Verza et al.,2005). Comparison of these assembled sequences withthe TRANSFAC (Matys et al., 2003), Pfam and theGenBank databases resulted in the identification of 1233sequences putatively encoding TFs, 414 of which camefrom endosperm cDNA libraries (Table 1). To estimatethe level of redundancy among the assembled sequences,the 414 TF sequences were compared with each otherusing BLASTN. Two sequences were considered asoriginating from the same transcript when they had98% nucleotide identity over a minimum of 100 bp.Around 10% of the TF sequences presented some degreeof redundancy, indicating that we have identified at least369 individual endosperm-expressed TFs. By searchingfor ESTs originated exclusively from endosperm cDNAlibraries, we identified 113 endosperm-preferred TFs,distributed among 53 contigs and 60 singletons (Table 1).The distribution of the endosperm-preferred TFs amongthe main families is shown in Table 2.
Functional annotation of endosperm-preferred TFs
The endosperm-preferred TFs were classified into familiesaccording to the roles established in TRANSFAC (Matyset al., 2003), GenBank (Benson et al., 2005) and Pfam(Bateman et al., 2004). When classification was notpossible, the TFs were placed in the ’Other’ group. Themost represented endosperm-expressed TF was the zinc-finger domain family, with 50 TFs (12%), followed by
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 117Annals of Applied Biology © 2010 Association of Applied Biologists
Transcription factor profiling in maize endosperm N.C. Verza et al.
Table 1 Transcription factors (TFs) expressed in developing maize
endosperm
Number of sequences analyseda 227 431Number of sequences from endospermb 64 537Number of assembled sequences analysed 29 206Number of assembled sequences representing TFsc 1 233Number of TFs expressed in endosperm 414Endosperm-preferred TFsd 113
aTotal sequences from MAIZEST database (Verza et al., 2005).bTotal sequences derived from developing endosperm cDNA libraries
(Verza et al., 2005).cTranscription factor (TF) sequences were identified by comparing
the assembled sequences with TRANSFAC Professional 8.2 (Biobase),
GenBank and with a set of TF domains from the Pfam database.dTFs found exclusevely in developing endosperm.
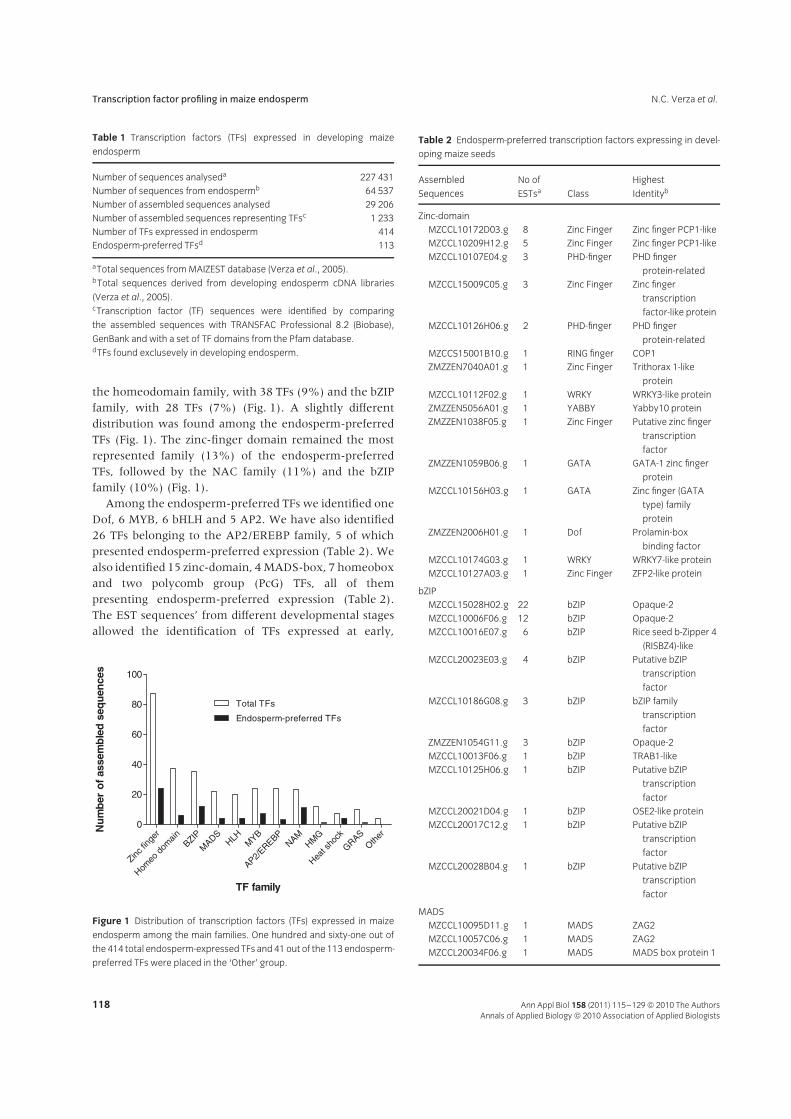
the homeodomain family, with 38 TFs (9%) and the bZIPfamily, with 28 TFs (7%) (Fig. 1). A slightly differentdistribution was found among the endosperm-preferredTFs (Fig. 1). The zinc-finger domain remained the mostrepresented family (13%) of the endosperm-preferredTFs, followed by the NAC family (11%) and the bZIPfamily (10%) (Fig. 1).
Among the endosperm-preferred TFs we identified oneDof, 6 MYB, 6 bHLH and 5 AP2. We have also identified26 TFs belonging to the AP2/EREBP family, 5 of whichpresented endosperm-preferred expression (Table 2). Wealso identified 15 zinc-domain, 4 MADS-box, 7 homeoboxand two polycomb group (PcG) TFs, all of thempresenting endosperm-preferred expression (Table 2).The EST sequences’ from different developmental stagesallowed the identification of TFs expressed at early,
Zinc fin
ger
Homeo
dom
ainBZIP
MADS
HLHM
YB
AP2/EREBP
NAMHM
G
Heat s
hock
GRASOth
er0
20
40
60
80
100
Total TFs
Endosperm-preferred TFs
TF family
Nu
mb
er o
f as
sem
ble
d s
equ
ence
s
Figure 1 Distribution of transcription factors (TFs) expressed in maize
endosperm among the main families. One hundred and sixty-one out of
the 414 total endosperm-expressed TFs and 41 out of the 113 endosperm-
preferred TFs were placed in the ‘Other’ group.
Table 2 Endosperm-preferred transcription factors expressing in devel-
oping maize seeds
Assembled
Sequences
No of
ESTsa Class
Highest
Identityb
Zinc-domainMZCCL10172D03.g 8 Zinc Finger Zinc finger PCP1-likeMZCCL10209H12.g 5 Zinc Finger Zinc finger PCP1-likeMZCCL10107E04.g 3 PHD-finger PHD finger
protein-relatedMZCCL15009C05.g 3 Zinc Finger Zinc finger
transcription
factor-like proteinMZCCL10126H06.g 2 PHD-finger PHD finger
protein-relatedMZCCS15001B10.g 1 RING finger COP1ZMZZEN7040A01.g 1 Zinc Finger Trithorax 1-like
proteinMZCCL10112F02.g 1 WRKY WRKY3-like proteinZMZZEN5056A01.g 1 YABBY Yabby10 proteinZMZZEN1038F05.g 1 Zinc Finger Putative zinc finger
transcription
factorZMZZEN1059B06.g 1 GATA GATA-1 zinc finger
proteinMZCCL10156H03.g 1 GATA Zinc finger (GATA
type) family
proteinZMZZEN2006H01.g 1 Dof Prolamin-box
binding factorMZCCL10174G03.g 1 WRKY WRKY7-like proteinMZCCL10127A03.g 1 Zinc Finger ZFP2-like protein
bZIPMZCCL15028H02.g 22 bZIP Opaque-2MZCCL10006F06.g 12 bZIP Opaque-2MZCCL10016E07.g 6 bZIP Rice seed b-Zipper 4
(RISBZ4)-likeMZCCL20023E03.g 4 bZIP Putative bZIP
transcription
factorMZCCL10186G08.g 3 bZIP bZIP family
transcription
factorZMZZEN1054G11.g 3 bZIP Opaque-2MZCCL10013F06.g 1 bZIP TRAB1-likeMZCCL10125H06.g 1 bZIP Putative bZIP
transcription
factorMZCCL20021D04.g 1 bZIP OSE2-like proteinMZCCL20017C12.g 1 bZIP Putative bZIP
transcription
factorMZCCL20028B04.g 1 bZIP Putative bZIP
transcription
factor
MADSMZCCL10095D11.g 1 MADS ZAG2MZCCL10057C06.g 1 MADS ZAG2MZCCL20034F06.g 1 MADS MADS box protein 1
118 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists
N.C. Verza et al. Transcription factor profiling in maize endosperm
Table 2 Continued
Assembled
Sequences
No of
ESTsa Class
Highest
Identityb
MZCCL10013G09.g 1 MADS MADS-box
transcription
factor-like
HomeodomainMZCCL10121F06.g 3 Homeobox Hox7-like proteinMZCCL10056F10.g 3 Homeo-Zip Putative Hox4 proteinMZCCL10216G12.g 1 Homeo-Zip OCL3 proteinZMZZEN6061C10.g 1 Homeo-Zip OCL5 proteinMZCCL15029D11.g 1 Homeobox Putative WUSCHEL
homeobox protein 2ZMZZEN6070H06.g 1 Homeobox Putative WUSCHEL
homeobox protein
11MZCCL10079H04.g 1 Homeobox Putative
homeodomain
protein
Helix-loop-helixMZCCS20044E10.g 11 bHLH bHLH protein familyZMZZEN5008D08.g 2 bHLH Putative bHLH
transcription factorZMZZEN7010B07.g 1 bHLH Putative transcription
factor PCF6MZCCL10202D08.g 1 bHLH bHLH protein familyMZCCL10075H11.g 1 bHLH bHLH protein familyMZCCS20019G07.g 1 bHLH Putative bHLH
transcription factor
NACZMZZEN3009C11.g 30 NAC NAM-related protein
1-likeMZCCL10018G09.g 23 NAC NAM-related protein
1-likeMZCCS15005C05.g 7 NAC NAM-related protein
1-likeMZCCL20006E06.g 4 NAC NAM-related protein
1-likeMZCCL10127E04.g 4 NAC OsNAC3 protein-likeZMZZEN6071D10.g 3 NAC NAM-related protein
1-likeMZCCL10058G04.g 2 NAC OsNAC2 protein-likeZMZZEN5053D04.g 2 NAC NAM-related protein
1-likeZMZZEN7014B11.g 2 NAC Putative NAM (no
apical meristem)
proteinMZCCL20010G12.g 1 NAC OsNAC1 protein-likeZMZZEN7015E08.g 1 NAC NAC2 protein-likeMZCCL10055H06.g 1 NAC NAM-like protein
Apetala2/EREBPMZCCL20025G09.g 13 ERF Putative transcription
factor EREBP1MZCCL10005A11.g 2 AP2 AP2
domain-containing
transcription factor
Table 2 Continued
Assembled
Sequences
No of
ESTsa Class
Highest
Identityb
MZCCL20042F02.g 1 AP2 AP2
domain-containing
transcription factorMZCCL10193B06.g 1 AP2 WRINKLED1-like
proteinMZCCL20022H07.g 1 ERF ERF1-like
transcription factor
HMGMZCCS15031G06.g 1 HMG Putative SSRP1
protein
MybMZCCS20011C07.g 4 MYB Putative typical
P-type R2R3 Myb
proteinMZCCL10097G05.g1 1 MYB Putative typical
P-type R2R3 Myb
proteinMZCCL10142D02.g 1 MYB Putative Myb-family
transcription factorMZCCL15026F02.g 1 MYB Putative Myb-family
transcription factorMZCCL10068G01.g 1 MYB Circadian clock
associated protein
LHY-likeZMZZEN6076B05.g 1 MYB Putative c-Myb-like
transcription factor
Heat shock factorMZCCL10023G08.g 3 HSF Heat shock factor
RHSF2-likeMZCCL10084F08.g 3 HSF Heat shock factor
RHSF4-likeMZCCL10160C11.g 1 HSF Heat shock factor
RHSF6-likeMZCCL20049C08.g 1 HSF Heat shock factor
RHSF4-like
GrasZMZZEN6040H08.g 1 GRAS Scarecrow
transcriptional
regulator-like
protein
OtherMZCCL10045B04.g 10 — Fertilisation-
independent
endosperm protein
1MZCCL10186E06.g 6 — Heat shock protein
HSP90MZCCL10038G04.g 6 — Putative VIP1
transcription factorMZCCL10026E03.g 5 — Heat shock protein
HSP82MZCCS20012A11.g 4 — Putative VIP2
transcription factor
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 119Annals of Applied Biology © 2010 Association of Applied Biologists
Transcription factor profiling in maize endosperm N.C. Verza et al.
Table 2 Continued
Assembled Sequences
No of
ESTsa Class Highest Identityb
ZMZZEN5005A11.g 4 — Putative transcription
factor X1MZCCL10039B11.g 3 — TATA box binding
protein-associated
factorMZCCL15012H09.g 3 — Transcription initiation
factor IIBMZCCM15002E07.g 3 — Putative co-repressor
proteinZMZZEN5055B11.g 3 — Transcription initiation
factor IIA small
subunitZMZZEN5026G12.g 3 — Putative
CCAAT-binding
transcription factorMZCCL20014D07.g 2 — Putative auxin
response factor 10
(ARF10)-likeMZCCL20034D01.g 2 — Heat shock protein
HSP82ZMZZEN6065G08.g 2 — Transcription initiation
factor IIHMZCCL20004F04.g 2 — Squamosa-promoter
binding-like proteinMZCCL10075G04.g 2 — TATA box binding
protein-associated
factorMZCCS10002E02.g 2 — TATA box binding
protein-associated
factorMZCCL10084A05.g 2 — Hd1-like proteinMZCCL10200C10.g 2 — Transcriptional
regulator FUSCA3ZMZZEN6050A03.g 2 — Putative
auxin-regulated
IAA22MZCCL10161E11.g 2 — Enhancer of zeste-like
protein 2ZMZZEN6049A09.g 2 — Putative
auxin-regulated
IAA8ZMZZEN6039F09.g 2 — Putative
CCAAT-binding
transcription factorZMZZEN6002G01.g 1 — Response regulator 10MZCCS20026C03.g 1 — Putative auxin
response factor 7
(ARF7)-likeMZCCS15007G06.g 1 — Putative auxin
response factor 7
(ARF7)-likeMZCCS15013B03.g 1 — Putative auxin
response factor 10
(ARF10)-likeZMZZEN5030B10.g 1 — Putative co-repressor
protein
Table 2 Continued
Assembled Sequences
No of
ESTsa Class Highest Identityb
ZMZZEN6061H10.g 1 — Putative auxin
response factor 1
(ARF1)-likeMZCCL20022G06.g 1 — Transcription initiation
factor IIBMZCCL15035F11.g 1 — Putative transcription
regulatory protein
SNF2MZCCL15009F05.g 1 — Putative
transcriptional
regulatory proteinMZCCL10077B06.g 1 — Putative
ethylene-insensitive
protein EIL3ZMZZEN6097G06.g 1 — Putative
transcriptional
co-repressor
LEUNIGMZCCL10178A09.g 1 — Putative HAP3
transcriptional-
activatorZMZZEN1009H11.g 1 — Putative transcription
factor IIIB 70 KD
subunitZMZZEN5062G11.g 1 — VIP1/ABI3-like
transcription factorMZCCL10212F09.g 1 — Putative
transcriptional
regulatory proteinMZCCL10210H11.g 1 — ABA response element
binding factorZMZZEN7004F11.g 1 — Putative trancription
factor
aNumber of ESTs in assembled sequences.bBest hit in the databases searches.
middle and late stages of seed development. The relative
expression of these TFs was calculated as the number
of ESTs from a given TF assembly divided by the total
number of ESTs of a particular stage (Table 3). This
allowed the identification of 36 TFs expressed only at
10 DAP, 12 TFs expressed only at 15 DAP and 18 TFs
expressed only at 20 DAP. Members of the NAC and
zinc finger families usually presented late expression
(15–20 DAP) while bZIP family members were found
expressed at all developmental stages. A total of 24
NAC family TFs was identified, 12 of which showed
endosperm-preferred expression (Table 2). The majority
of the endosperm-preferred NAC members presented late
expression (Table 3).
120 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists
N.C. Verza et al. Transcription factor profiling in maize endosperm
Table 3 Relative expression of endosperm-preferred transcription
factors in developing maize seeds
Relative Expressiona
Highest Identity Assembled Sequences 10 DAP 15 DAP 20 DAP
NAM-related
protein 1-like
MZCCL10018G09.g 0.6 1.6 17.4
Fertilisation-
independent
endosperm
protein 1
MZCCL10045B04.g 3.1 1.6 0
Zinc finger
PCP1-like
MZCCL10172D03.g 0.6 3.1 1.2
NAM-related
protein 1-like
MZCCL20006E06.g 0 1.6 3.7
NAM-related
protein 1-like
MZCCS15005C05.g 0 6.3 0
Transcription
initiation factor
IIB
MZCCL15012H09.g 0 4.7 0
Putative bZIP
transcription
factor
MZCCL20023E03.g 0 0 3.7
Zinc finger
transcription
factor-like
protein
MZCCL15009C05.g 0 3.1 0
Putative
transcription
factor EREBP1
MZCCL20025G09.g 0 0 2.5
Heat shock protein
HSP82
MZCCL20034D01.g 0 0 2.5
Opaque-2 MZCCL10006F06.g 4.3 1.6 3.7Putative typical
P-type R2R3
Myb protein
MZCCS20011C07.g 0 0 2.5
Putative VIP2
transcription
factor
MZCCS20012A11.g 0 0 2.5
Putative Hox4
protein
MZCCL10056F10.g 1.9 0 0
Heat shock factor
RHSF4-like
MZCCL10084F08.g 1.9 0 0
Putative VIP1
transcription
factor
MZCCL10038G04.g 1.2 1.6 0
Heat shock protein
HSP82
MZCCL10026E03.g 1.2 0 2.5
AP2 domain-
containing
transcription
factor
MZCCL10005A11.g 1.2 0 0
OsNAC2
protein-like
MZCCL10058G04.g 1.2 0 0
Hd1-like protein MZCCL10084A05.g 1.2 0 0OsNAC3
protein-like
MZCCL10127E04.g 1.2 0 0
Enhancer of
zeste-like
protein 2
MZCCL10161E11.g 1.2 0 0
Table 3 Continued
Relative Expressiona
Highest Identity Assembled Sequences 10 DAP 15 DAP 20 DAP
TATA box-binding
protein-
associated
factor
MZCCS10002E02.g 1.2 0 0
PHD finger
protein-related
MZCCL10107E04.g 0.6 1.6 1.2
Rice seed b-Zipper
4 (RISBZ4)-like
MZCCL10016E07.g 0.6 0 1.2
TATA box-binding
protein-
associated
factor
MZCCL10075G04.g 0.6 0 1.2
PHD finger
protein-related
MZCCL10126H06.g 0.6 0 1.2
bZIP family
transcription
factor
MZCCL10186G08.g 0.6 0 1.2
Zinc finger
PCP1-like
MZCCL10209H12.g 0.6 0 1.2
Opaque-2 MZCCL15028H02.g 0 1.6 1.2Putative auxin
response factor
10 (ARF10)-like
MZCCL20014D07.g 0 1.6 1.2
Putative
transcriptional
regulatory
protein
MZCCL15009F05.g 0 1.6 0
Putative
co-repressor
protein
MZCCM15002E07.g 0 1.6 0
Putative
Myb-family
transcription
factor
MZCCL15026F02.g 0 1.6 0
Putative WUSCHEL
homeobox
protein 2
MZCCL15029D11.g 0 1.6 0
Putative
transcription
regulatory
protein SNF2
MZCCL15035F11.g 0 1.6 0
COP1 MZCCS15001B10.g 0 1.6 0Putative auxin
response factor
7 (ARF7)-like
MZCCS15007G06.g 0 1.6 0
Putative auxin
response factor
10 (ARF10)-like
MZCCS15013B03.g 0 1.6 0
Putative SSRP1
protein
MZCCS15031G06.g 0 1.6 0
bHLH protein
family
MZCCS20044E10.g 0 0 1.2
Squamosa-
promoter
binding-like
protein
MZCCL20004F04.g 0 0 1.2
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 121Annals of Applied Biology © 2010 Association of Applied Biologists
Transcription factor profiling in maize endosperm N.C. Verza et al.
Table 3 Continued
Relative Expressiona
Highest Identity Assembled Sequences 10 DAP 15 DAP 20 DAP
OsNAC1 protein-like MZCCL20010G12.g 0 0 1.2Putative bZIP
transcription
factor
MZCCL20017C12.g 0 0 1.2
OSE2-like protein MZCCL20021D04.g 0 0 1.2Transcription
initiation factor
IIB
MZCCL20022G06.g 0 0 1.2
ERF1-like
transcription
factor
MZCCL20022H07.g 0 0 1.2
Putative bZIP
transcription
factor
MZCCL20028B04.g 0 0 1.2
MADS box protein 1 MZCCL20034F06.g 0 0 1.2AP2 domain-
containing
transcription
factor
MZCCL20042F02.g 0 0 1.2
Heat shock factor
RHSF4-like
MZCCL20049C08.g 0 0 1.2
Putative bHLH
transcription
factor
MZCCS20019G07.g 0 0 1.2
Putative auxin
response factor 7
(ARF7)-like
MZCCS20026C03.g 0 0 1.2
Hox7-like protein MZCCL10121F06.g 0.6 0 0TRAB1-like MZCCL10013F06.g 0.6 0 0MADS-box
transcription
factor-like
MZCCL10013G09.g 0.6 0 0
Heat shock factor
RHSF2-like
MZCCL10023G08.g 0.6 0 0
TATA box binding
protein-
associated
factor
MZCCL10039B11.g 0.6 0 0
NAM-like protein MZCCL10055H06.g 0.6 0 0ZAG2 MZCCL10057C06.g 0.6 0 0Circadian clock
associated
protein LHY-like
MZCCL10068G01.g 0.6 0 0
bHLH protein family MZCCL10075H11.g 0.6 0 0Putative ethylene-
insensitive
protein EIL3
MZCCL10077B06.g 0.6 0 0
Putative
homeodomain
protein
MZCCL10079H04.g 0.6 0 0
ZAG2 MZCCL10095D11.g 0.6 0 0Putative typical
P-type R2R3 Myb
protein
MZCCL10097G05.g1 0.6 0 0
WRKY3-like protein MZCCL10112F02.g 0.6 0 0
Table 3 Continued
Relative Expressiona
Highest Identity Assembled Sequences 10 DAP 15 DAP 20 DAP
Putative bZIP
transcription
factor
MZCCL10125H06.g 0.6 0 0
ZFP2-like protein MZCCL10127A03.g 0.6 0 0Putative Myb-family
transcription
factor
MZCCL10142D02.g 0.6 0 0
Zinc finger (GATA
type) family
protein
MZCCL10156H03.g 0.6 0 0
Heat shock factor
RHSF6-like
MZCCL10160C11.g 0.6 0 0
WRKY7-like protein MZCCL10174G03.g 0.6 0 0Putative HAP3
transcriptional-
activator
MZCCL10178A09.g 0.6 0 0
Heat shock protein
HSP90
MZCCL10186E06.g 0.6 0 0
WRINKLED1-like
protein
MZCCL10193B06.g 0.6 0 0
Transcriptional
regulator FUSCA3
MZCCL10200C10.g 0.6 0 0
bHLH protein family MZCCL10202D08.g 0.6 0 0ABA response
element binding
factor
MZCCL10210H11.g 0.6 0 0
Putative
transcriptional
regulatory protein
MZCCL10212F09.g 0.6 0 0
OCL3 protein MZCCL10216G12.g 0.6 0 0
aThe relative abundance of ESTs for each assembled sequence was
calculated as the number of ESTs present in a given library pool (10 DAP
pool, 15 DAP pool and 20 DAP pool) divided by the total number of ESTs in
that pool. The values were multiplied by 104 to aid better interpretation.
Identification of an aleurone-preferred NAC TF
The 12 endosperm-preferred NAC TFs correspond to10 non-redundant family members. As most of theseTFs presented late expression (Table 3), we examinedwhether they may play a role in the response to droughtstress that is associated with seed desiccation. We choseAPN-1 (Table 4) for further characterisation. AnotherNAC with higher number of ESTs has already beencharacterised as NRP-1(Guo et al., 2003).
The assembled consensus cDNA sequence of Apn-1 isavailable at GenBank under accession number C0440312.This consensus sequence was used to design primers toamplify, clone and resequence the complete Apn-1 cDNAsequence. The amino acid sequence deduced from thecDNA of Apn-1 was 73% identical to the petunia NAM(accession number X92205) and 84% identical to the
122 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists

N.C. Verza et al. Transcription factor profiling in maize endosperm
Table 4 Endosperm-preferred NAC-family transcription factors
Assembled
Sequence
Number of
Sequencesa Highest Identityb
ZMZZEN3009C11.g 30 NAM-related protein 1-likeMZCCL10018G09.gc 23 NAM-related protein 1-likeMZCCS15005C05.g 7 NAM-related protein 1-likeMZCCL20006E06.g 4 NAM-related protein 1-likeMZCCL10127E04.g 4 OsNAC3 protein-likeZMZZEN6071D10.g 3 NAM-related protein 1-likeMZCCL10058G04.g 2 OsNAC2 protein-likeZMZZEN5053D04.g 2 NAM-related protein 1-likeZMZZEN7014B11.g 2 Putative NAM proteinMZCCL20010G12.g 1 OsNAC1 protein-likeZMZZEN7015E08.g 1 NAC2 protein-likeMZCCL10055H06.g 1 NAM-like protein
aNumber of ESTs in the assembled sequence.bBest GenBank hit.cAssembled sequence corresponding to the Apn-1 gene.
maize NRP-1 (accession number AY325313). NRP-1 isan endosperm-specific NAC member regulated by gene-specific imprinting (Guo et al., 2003). The alignment ofthe amino acid sequence of APN-1 with the amino acidsequences of related NAC-family members shows thatthe NAC domains are highly conserved among all of thesequences (Fig. 2A). The C-terminal region of APN-1 wasless conserved when compared with the correspondingsequences of the other NAC members as exemplified byNRP-1 that shared only 65% identity with APN-1 in thisregion (Fig. 2A).
We retrieved the genomic sequence of Apn-1 fromthe TIGR database (http://www.tigr.org) to determine itsgene structure. Comparison of the Apn-1 full-length cDNAsequence with the Apn-1 genomic sequence showedthat the gene consists of three exons and two introns(Fig. 2B). This gene structure differed from that of thepetunia phNam gene and the Arabidopsis AraCuc-1 gene,both of which have four exons and three introns. Incontrast, the Apn-1 gene structure was very similar tothat of maize Nrp-1, which also has three exons andtwo introns. The first exon of Nrp-1 was less than halfthe size of that of Apn-1 whereas the first intron ofNrp-1 was several-fold larger than that of Apn-1. Thesegenomic comparisons indicated that Apn-1 is distinct fromthe other genes encoding NAC family members alreadycharacterised in maize and other plants (Souer et al., 1996;Guo et al., 2003).
The Apn-1 gene is preferentially expressedin the developing endosperm
To confirm the expression pattern of the Apn-1gene, we examined its mRNA levels by RT-PCR
in developing endosperm, root, leaf, coleoptiles andembryonic tissues. The Apn-1 gene was preferentiallyexpressed in developing endosperm, with a pattern ofexpression that started 5 DAP and reached a maximumaround 20 DAP (Fig. 3). The Apn-1 mRNA was alsodetected in coleoptiles and embryo, but the level ofexpression was several-fold lower than that observedfor the developing endosperm (Fig. 3). Very faint bandswere also observed in leaves and roots, indicating lowexpression of Apn-1 in these tissues.
The Apn-1 gene promoter has conservedendosperm-specific, ABA and GA cis-acting elements
The promoter sequence of Apn-1 was retrievedfrom the maize contig assembly available at TIGR(http://www.tigr.org). A 1.9 kb Apn-1 promoter fragmentlocated upstream from the translation start codon wasamplified and cloned. The PlantCare (Lescot et al., 2002)and Place (Higo et al., 1999) tools were used to identifyputative conserved cis-regulatory elements in the Apn-1promoter (data not shown). We identified two TATAboxes, one located at position 113 and another at position352 from the initial ATG. The Apn-1 promoter containedputative elements related to endosperm-specificity suchas the prolamin-box that is conserved in maize seed stor-age proteins, a GCN-4-like motif that plays a central rolein controlling endosperm-specific expression (De Freitaset al., 1994; Yunes et al., 1998), and an RY/Sph motif thatregulates the expression of several seed-specific genes(Bobb et al., 1997). The RY/Sph motif may also regu-late the response to ABA signalling through the bindingof maize VP1. A number of hormone-related cis-actingelements were also found in the Apn-1 promoter, includ-ing an ACGT-containing ABA response elements (ABREs)(Ezcurra et al., 2000), an amylase box (also known as AmyBox and Box I) conserved in the promoter of α-amylase
genes, a pyrimidine box that occurs as an accessory motifin the promoter of barley α-amylase genes (Amy2/32b)(Gomez-Cadenas et al., 2001), and two TATC-boxes thatare related to GA responsiveness.
The Apn-1 gene promoter drives aleurone-specificexpression
Transient expression in developing endosperm was usedto examine the pattern of expression driven by theApn-1 gene promoter (Fig. 4). The 1.9 kb Apn-1 promoterfragment was cloned into the promoter-less pRT103GUSvector (Yunes et al., 1994) to drive the expression ofthe GUS gene. The pApn-1-GUS construct along withthe control construct pRT103GUS containing the GUSgene driven by the constitutive CaMV35S promoter
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 123Annals of Applied Biology © 2010 Association of Applied Biologists

Transcription factor profiling in maize endosperm N.C. Verza et al.
A
ZmASN-1 1 MAADQQPQLQEEMNDDAVGGGLRLPPGFRFHPSDFEIVSFYLTNKVLNTR-FTCTAITEAZmNRP-1 1 MADQQQPQQQPQEMDVDRTGGLELPPGFRFHPSDFEIINDYLTKKVHDRD-YSCIAIADAOsNAC1 1 ---------------------MDLPPGFRFHPTDEELITHYLLRKAADPAGFAARAVGEAPhNAM 1 ----------MENYQHFDCSDSNLPPGFRFHPTDEELITYYLLKKVLDSN-FTGRAIAEVANAC021/22 1 -----METEEEMKESSISMVEAKLPPGFRFHPKDDELVCDYLMRRSLHNNHRPPLVLIQV
ZmASN-1 60 DLNKIEPWDLPSKAKMGEKEWYFFYQKDRKYPTGLRANRATEAGYWKATGKDKEVYN--A ZmNRP-1 60 DLNKTEPWDLPKVAKMGEKEWCFFYQKDRKYPTGLRANRATEAGYWKATGKDKEVYNPFA OsNAC1 40 DLNKCEPWDLPSRATMGEKEWYFFCVKDRKYPTGLRTNRATESGYWKATGKDREIFR--- PhNAM 50 DLNKCEPWELPEKAKMGEKEWYFFSLRDRKYPTGLRTNRATEAGYWKATGKDREIYS--- ANAC021/22 56 DLNKCEPWDIPKMACVGGKDWYFYSQRDRKYATGLRTNRATATGYWKATGKDRTILR---
ZmASN-1 118 AEGVAVLVGMKKTLVFYRGRAPRGDKTNWVMHEYRLEGSGRLPAGLASATGSAAANAAAAZmNRP-1 120 AEG-LLLVGMKKTLVFYKGRAPRGDKTNWVMHEYRLEGSGRLPASPASASGSATNIAAAMOsNAC1 97 -GK--ALVGMKKTLVFYTGRAPRGGKTGWVMHEYRIHG----------------KHAAANPhNAM 107 -SKTSALVGMKKTLVFYRGRAPKGEKSNWVMHEYRLDG----------------KFAYHY ANAC021/22 113 ---KGKLVGMRKTLVFYQGRAPRGRKTDWVMHEFRLQG---------------SHHPPNH
ZmASN-1 178 LKASA--YKDEWVVCRVFHKTTGIKKTTTAAPAYQVAMAGAEMDQNQNNIP--------- ZmNRP-1 179 MKASASACKDEWVVCRVFNKTTGIKKT--AAPAYQVAMAGPEMDQNQNNIPAIPIPMPLQ OsNAC1 138 SKQD-----QEWVLCRVFKKSLELAPAAAAAVGRRGAGAGTDVGPSSMPMADDVV----- PhNAM 150 ISRSS---KDEWVISRVFQKS-------CSTVGTTSNGGKKRLNSSFNNMYQEVS----- ANAC021/22 155 SLSSP---KEDWVLCRVFHKN------------TEGVICRDNMGSCFDETAS--------
ZmASN-1 227 ----FPMPMQFSMLPDFSLDP----VPPYYPYPNAGAGMPMLPMAAGIGGGAGG------ ZmNRP-1 237 LPLPVPMQMQFPILPDFAMDP----VAPYYPNPNAGAGM-MPPMALAGMGGAGG------ OsNAC1 188 ----GLAPCALPPLMDVSGGGGGAGTTSLSATAGAAAAPPAAHVTCFSNALEGQFLDTPY PhNAM 195 ----SPSSVSLPPLLESS---------PYNNTATSAAASKKEHVSCFSTIST-------- ANAC021/22 192 --------ASLPPLMDPYINFD---QEPSSYLSDDHHYIINEHVPCFSNLSQNQ------
ZmASN-1 273 --LHLNGAALFGNPMAAPQPMSFYH-QMGTGTAC--------------------AGGFDV ZmNRP-1 286 --LQING-ALFGNPVPAPLPMNFYHHQMGMGAAAGQVDMGAAAGQMDMGAAGAGAGGFDV OsNAC1 244 LLPAADPADHLAMSSASPFLEALQMQYVQDAAAAGGAGMVHE---------LLMGGGWYC PhNAM 234 --PSFDPSSVFDISSNSNTLHSLPAPSFSAILDPSSTFSRNS---------VFPS---LR ANAC021/22 235 ----TLNSNLTNSVSELKIPCKNPNPLFTGGSASATLTGLDS----------------FC
ZmASN-1 310 SAPESRPSSMVSQKD----DQANGAEISSMMSVAGPGPATTTT--IEMDGVWKY-- ZmNRP-1 343 AAPESRPSSMVSQKD----EQANAAEISSMMSVTGPG-SATTT--IEMDGIWKYKY OsNAC1 295 NKGERERLSGASQDTGLTSSEVNPGEISSSSRHNAWIITTRPSGPIEIFIHHH--- PhNAM 280 SLQENLHLPLFSGGT----SAMHGG--FSSPLANWPVPETQKVDHSELDCMWSY-- ANAC021/22 275 SSDQMVLRALLSQLT-----KIDGSLGPKESQSYGEGSSESLLTDIGIPSTVWNC-
B
PhNam
AraCuc-1
ZmNrp-1
ZmApn-1
I II III IV
B
B
C
D
E
A
Figure 2 (A) Alignment of the amino acid sequences of the five NAC-family transcription factors (TFs); maize APN-1 and NRP-1 (AAP86221); rice OsNAC1
(BAC53810); petunia PhNAM (CAA63101) and Arabidopsis CUC-1 (AB049069). Lines above the sequences indicate the conserved NAC subdomains A
to E. Amino acid identification: white on black, identical residues and white on light grey, conserved residues. (B) Schematic representation of the gene
structure encoding NAC-family proteins. Boxes represent exons and lines introns. Lengths were scaled up. Petunia NAM (CAA63101), Arabidopsis CUC-1
(AB049069), maize NRP-1 (AAP86221) and maize APN-1. Roman numbers indicate exons.
124 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists
N.C. Verza et al. Transcription factor profiling in maize endosperm
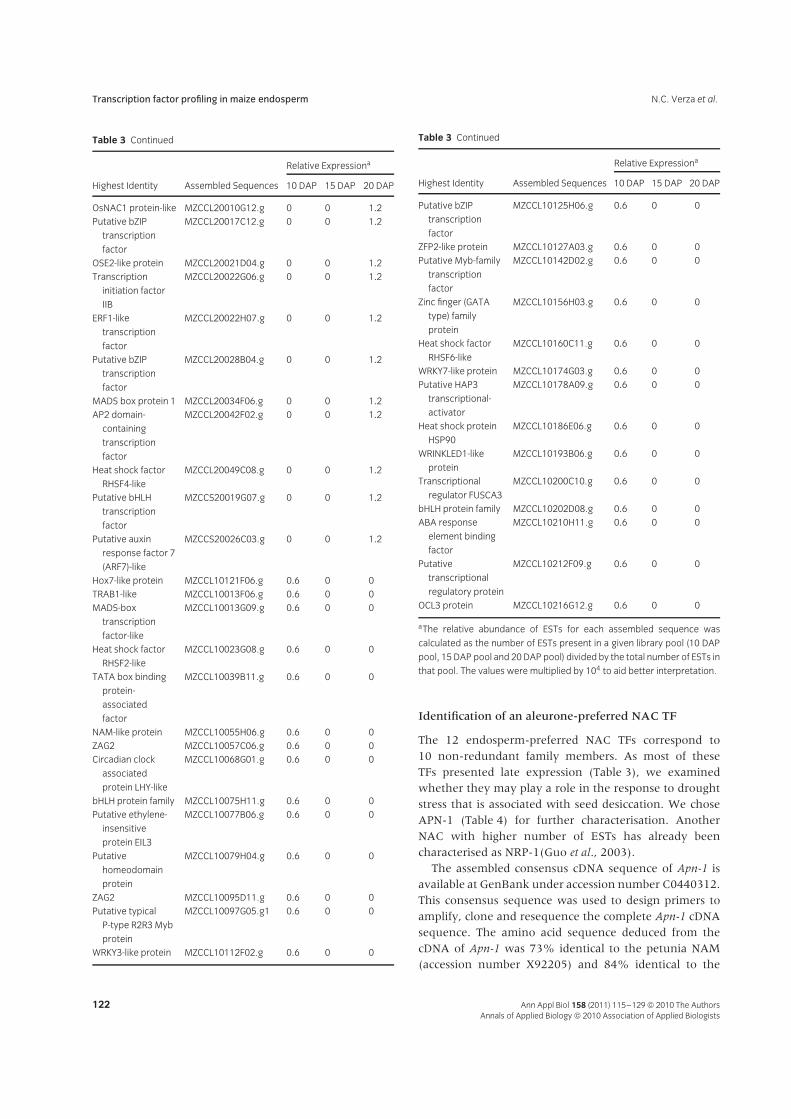
Figure 3 Reverse trascription polymerase chain reaction (RT-PCR)
analysis of Apn-1expression during seed development and among
different tissues. L, leaf; R, root; Co, coleoptile; Emb, embryo. Actin
was used as an endogenous control for RT-PCR analysis.
was used in transient assays (Fig. 4A). Immature maizeseeds were sectioned transversally and the caps wereprepared by peeling back the pericarp and removingthe aleurone layer from half of the cap area whileleaving the other half of the cap with an intactaleurone layer. The caps were bombarded with goldmicroprojectiles coated with DNA constructs and theGUS activity was assayed histologically (Fig. 4B). GUSexpression driven by the CaMV35S promoter occurredin aleurone and sub-aleurone endosperm cells, whereasthe Apn-1 promoter directed GUS expression exclusivelyin the aleurone layer. This result clearly shows that theApn-1 gene is preferentially expressed in the aleurone celllayer.
The expression of Apn-1 is induced by ABA andinhibited by GA
To examine the possible involvement of hormones inthe regulation of Apn-1 expression, 10 DAP endospermwas treated with ABA or GA and Apn-1 mRNA levelswere estimated by Q-PCR. The Apn-1 transcript level wasreduced by 3- and 12-fold when developing endospermswere treated with 25 and 75 μM GA, respectively (Fig. 5).In contrast, the transcript levels increased by 40%when developing seeds were treated with 75 μM ABA(Fig. 5). These findings indicated that Apn-1 expression isregulated by ABA and GA, two plant hormones involvedin seed maturation and seed germination, respectively.
Discussion
The maize endosperm development comprises definedphases associated with cell differentiation, development
Figure 4 Tissue distribution of GUS activity driven by the Apn-1 promoter
in 20 days after pollination (DAP) maize endosperm caps. Half of the
aleurone layers were removed from the caps just before bombardment. (A)
Schematic representation of the pApn1::GUS and p35S::GUS constructs.
(B) p35S::GUS bombarded caps showing expression in the aleurone and
sub-aleurone cell layers (left) and pApn-1::GUS bombarded caps showing
expression exclusively in the aleurone cell layer (right).
0
20
40
60
80
100
120
Control 25 75 25 75
GA (<M) ABA (<M)
APN
-1 m
RNA
rela
tive
exp
ress
ion
(%)
Figure 5 Apn-1 mRNA relative expression in 10 days after pollination
(DAP) maize kernels incubated in MS medium supplemented with
gibberellic acid (GA) or abscisic acid (ABA). The kernels were treated
with 25 μM and 75 μM of each hormone for 6 h in the dark at 25◦C. Three
biological replicates were performed for each treatment. Quantitative
reverse trascription polymerase chain reaction (qRT-PCR) was normalised
by 18S ribosomal (Taqman).
and metabolism. During the first 4 DAP, the endospermnuclei divide synchronously without cell wall formation.After this phase the tissue changes from a multinucleatesingle cell to a uninucleate multicellular morphologywith most cells being produced by 12 DAP. Around12 DAP, the endosperm begins to accumulate starch
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 125Annals of Applied Biology © 2010 Association of Applied Biologists

Transcription factor profiling in maize endosperm N.C. Verza et al.
and storage proteins and by 16 DAP, the maturationprogramme is initiated, preparing the seeds for desiccationand dormancy. At 25–30 DAP, the water content of theendosperm begins to decrease, and the seed desiccationsignals the arrest of germinative development (Lopes &Larkins, 1993; Olsen, 2001). One important challenge isto dissect the regulatory process orchestrated by TF sets,along each phase of endosperm development. Both tissue-preferred and non-preferred TFs regulate endospermdevelopment and metabolism. There are TFs related tospecific functions such as storage of protein and starchsynthesis and others that regulate genes that play a rolein endosperm development and metabolism as well as inother tissues.
Inspections of large collection of maize endospermESTs and their comparison with ESTs from a set ofvegetative maize tissues allowed us to identify TFs thatare preferentially expressed in developing endosperm.In addition, because endosperm ESTs were sequencedfrom libraries prepared with mRNA isolated from 10, 15and 20 DAP it was possible to identify the early andlate endosperm-preferred TFs. The Arabidopsis containsaround 1533 TFs, which account for approximately 5.9%of its total number of genes (Riechmann et al., 2000).If maize contains TFs in a proportion similar to that ofArabidopsis, one could estimate that the maize genomecodes for approximately 2300 TFs. Thus the 1233 TFswe identified in all maize tissues represents around54% of the maize TFs set. The 414 TFs expressed indeveloping endosperm accounts for 33% of the totalTFs while the endosperm-preferred TFs accounts for9.2% of the total TFs. This set of endosperm-preferredTFs may contain important candidates for regulation ofendosperm specification and development. One exampleis the members of the AP2/EREBP (Table 2). In ArabidopsisApetala2, a member of the AP2/EREBP family controlsseed mass through regulation of gene expression inmaternal sporophyte and endosperm (Jofuku et al., 2005;Ohto et al., 2005). Other examples of potential candidatesfor regulating nitrogen and carbohydrate metabolismare the endosperm-preferred Dof, MYB, bHLH and AP2(Table 2) as members of these groups of TFs have beenshown to regulate storage compounds deposition inseeds (Giroux et al., 1994; Price et al., 2004; Yanagisawaet al., 2004).
Endosperm-preferred TFs belonging to the zinc-domain, MADS-box, homeobox and polycomb groupwere also identified in this work (Table 2). Transcrip-tion factors of these groups have been shown to beinvolved in the regulation of various developmental pro-cesses (Kohler & Grossniklaus, 2002; Sung et al., 2003).Examples are FIE (fertilisation-independent endosperm)and MEA (Medea). Fertilisation-independent endosperm
regulates endosperm and embryo development andrepresses flowering during embryo and seedlingdevelopment whereas MEA regulates endosperm andembryo development (Kiyosue et al., 1999; Kohleret al., 2003).
One of the most interesting groups of endosperm-preferred TFs identified in this work belongs to theNAC family. The NAC family comprises a group ofTFs specific to plants and is found to play roles indiverse processes, including developmental programmesand response to biotic and abiotic stresses. Examples arethe AtNAM that is up-regulated in Arabidopsis developingseeds (Duval et al., 2002); NRP-1 that was shown to beexpressed in an endosperm-preferred manner in maize(Guo et al., 2003) and ZmNAM1, ZmNAM2, PhNAM,AtCUC2, AmCUP and ZmCUC3 that contribute to shootapical meristem establishment and formation of leafprimordium (Zimmermann & Werr, 2005). Twenty-four NAC members were identified in this work, 12 ofwhich show endosperm-preferred expression (Table 2).The majority of the endosperm-preferred NAC memberspresented late expression. After 20 DAP, when the NACtranscripts seem to accumulate in endosperm, the relativewater content of the endosperm begins to decrease andseeds initiate desiccation. Therefore, the endosperm-preferred NAC TFs identified in this study are goodcandidates to play a role in this physiological process.
To investigate the role of endosperm-preferred NACmembers in endosperm development, we chose APN-1 forfurther characterisation. The Apn-1 gene presented higherexpression at 20 DAP suggesting that it may play a rolein regulating the late phases of endosperm development,although the low expression seen in the early stagesmay also indicate a regulatory function right from thestart of development. The demonstration that the Apn-1expression in seeds was confined to the aleurone layeris of major interest. Aleurone is a tissue with molecularprogrammes related to the regulation of the transitionbetween seed development and germination. Both lateseed development and germination are controlled bylevels of the plant hormones ABA and GA. Inspectionof the promoter sequence of the Apn-1 gene revealed anumber of target sequences such as the pyrimidine andTATC boxes that are part of a GA response complex thatprovides a high level of GA-regulated expression. Thedemonstration that the expression of Apn-1 is inhibitedby GA and stimulated by ABA suggests that this TFmay have a regulatory role during the transition fromseed maturation to germination. In maize, ABA playsa central role in repressing precocious germination ofdeveloping seeds by regulating the expression of diversegenes involved in seed maturation. In developing seeds,ABA is synthesised in the embryo and is also transferred
126 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists

N.C. Verza et al. Transcription factor profiling in maize endosperm
from maternal tissues to the seed during desiccation (Ober& Setter, 1992). Maize kernels deficient in ABA synthesisare viviparous, germinating on the ear midway to kernelmaturity (Neill et al., 1986). While the ABA levelsare important to prevent precocious seed germination,GAs play a crucial role in promoting germination. Inwheat and barley, GAs promote the expression of genesencoding hydrolytic enzymes, leading to the mobilisationof endosperm reserves for the embryo nutrition (Gubleret al., 1995). In maize, GA and ABA play antagonisticroles and GA levels in maize embryos decline prior tothe peak of ABA (White et al., 2000). Several NAC geneshave been shown to be induced by ABA (Greve et al.,2003), while others act as negative regulators of GAresponse in barley aleurone, inhibiting GA up-regulationof alpha amylase (Robertson, 2004). Accordingly, wespeculate that the maize APN-1 may be involved in theinhibition of precocious seed germination, for example, bytranscriptional repression of genes encoding α-amylases,proteinases and glucanases. During germination, Apn-1expression in the aleurone cell layer is inhibited byGA produced by the embryo, thereby allowing theup-regulation of genes encoding these hydrolases and,consequently, seed germination. Apn-1 also showedrelatively weak expression in the embryo. It may alsoparticipate in the embryo’s desiccation tolerance. Furtherstudies are required to decipher the regulatory actions ofABA and GA on Apn-1 expression, and to clarify whetherthere is any interaction between VP1 and APN-1 duringseed development.
Acknowledgements
N. C. V. was supported by a postgraduate fellowshipfrom Coordenacao de Aperfeicoamento de Pessoal deNıvel Superior (CAPES), S. M. S was supported by apostgraduate fellowship from Conselho Nacional deDesenvolvimento Cientıfico e Tecnologico (CNPq) andT. R. S. F was supported by a postgraduate fellowshipfrom Fundacao de Amparo a Pesquisa do Estado de SaoPaulo (FAPESP).
Database submissions
The nucleotide sequence of Apn-1 gene has beendeposited in the NR division of GenBank under theaccession numbers 1148883 and 1148889.
References
Aida M., Ishida T., Fukaki H., Fujisawa H., Tasaka M.(1997) Genes involved in organ separation in Arabidopsis:an analysis of the cup-shaped cotyledon mutant. Plant
Cell, 9, 841–857.
Bateman A., Coin L., Durbin R., Finn R.D., Hollich V.,
Griffiths-Jones S., Khanna A., Marshall M., Moxon S.,
Sonnhammer E.L.L., Studholme D.J., Yeats C., Eddy S.R.
(2004) The Pfam protein families database. Nucleic Acids
Research, 32, D138–D141.
Benson D.A., Karsch-Mizrachi I., Lipman D.J., Ostell J.,
Wheeler D.L. (2005) GenBank. Nucleic Acids Research, 33,
D34–D38.
Bobb A.J., Chern M.S., Bustos M.M. (1997) Conserved
RY-repeats mediate transactivation of seed-specific
promoters by the developmental regulator PvALF. Nucleic
Acids Research, 25, 641–647.
Bowman J.L. (2000) The YABBY gene family and abaxial
cell fate. Current Opinion in Plant Biology, 3, 17–22.
De Freitas F.A., Yunes J.A., Da Silva M.J., Arruda P.,
Leite A. (1994) Structural characterization and promoter
activity analysis of the gamma-kafirin gene from
sorghum. Molecular & General Genetics, 245, 177–186.
Duval M., Hsieh T.F., Kim S.Y., Thomas T.L. (2002)
Molecular characterization of AtNAM: a member of the
Arabidopsis NAC domain superfamily. Plant Molecular
Biology, 50, 237–248.
Ezcurra I., Wycliffe P., Nehlin L., Ellerstrom M., Rask L.
(2000) Transactivation of the Brassica napus napin
promoter by ABI3 requires interaction of the conserved
B2 and B3 domains of ABI3 with different cis-elements:
B2 mediates activation through an ABRE, whereas B3
interacts with an RY/G-box. The Plant Journal, 24, 57–66.
Fujita M., Fujita Y., Maruyama K., Seki M., Hiratsu K.,
Ohme-Takagi M., Tran L.-S.P., Yamaguchi-Shinozaki K.,
Shinozaki K. (2004) A dehydration-induced NAC protein,
RD26, is involved in a novel ABA-dependent
stress-signaling pathway. The Plant Journal, 39, 863–876.
Giroux M.J., Boyer C., Feix G., Hannah L.C. (1994)
Coordinated transcriptional regulation of storage product
genes in the maize endosperm. Plant Physiology, 106,
713–722.
Gomez-Cadenas A., Zentella R., Sutliff T.D., Ho T.D.H.
(2001) Involvement of multiple cis-elements in the
regulation of GA responsive promoters: definition of a
new cis-element in the Amy32b gene promoter of barley
(Hordeum vulgare). Physiologia Plantarum, 112, 211–216.
Gong W., Shen Y.P., Ma L.G., Pan Y., Du Y.L., Wang D.H.,
Yang J.Y., Hu L.D., Liu X.F., Dong C.X., Ma L., Chen Y.H.,
Yang X.Y., Gao Y., Zhu D.M., Tan X.L., Mu J.Y.,
Zhang D.B., Liu Y.L., Dinesh-Kumar S.P., Li Y.,
Wang X.P., Gu H.Y., Qu L.J., Bai S.N., Lu Y.T., Li J.Y.,
Zhao J.D., Zuo J.R., Huang H., Deng X.W., Zhu Y.X.
(2004) Genome-wide ORFeome cloning and analysis of
Arabidopsis transcription factor genes. Plant Physiology,
135, 773–782.
Greve K., La Cour T., Jensen M.K., Poulsen F.M., Skriver K.
(2003) Interactions between plant RING-H2 and
plant-specific NAC (NAM/ATAF1/2/CUC2) proteins:
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 127Annals of Applied Biology © 2010 Association of Applied Biologists

Transcription factor profiling in maize endosperm N.C. Verza et al.
RING-H2 molecular specificity and cellular localization.
Biochemical Journal, 371, 97–108.
Gubler F., Kalla R., Roberts J.K., Jacobsen J.V. (1995)
Gibberellin-regulated expression of a myb gene in barley
aleurone cells – Evidence for Myb transactivation of a
high-pl alpha-amylase gene promoter. Plant Cell, 7,
1879–1891.
Guo M., Rupe M.A., Danilevskaya O.N., Yang X.F., Hut Z.H.
(2003) Genome-wide mRNA profiling reveals
heterochronic allelic variation and a new imprinted gene
in hybrid maize endosperm. The Plant Journal, 36, 30–44.
Higo K., Ugawa Y., Iwamoto M., Korenaga T. (1999) Plant
cis-acting regulatory DNA elements (PLACE) database.
Nucleic Acids Research, 27, 297–300.
Hobo T., Asada M., Kowyama Y., Hattori T. (1999)
ACGT-containing abscisic acid response element (ABRE)
and coupling element 3 (CE3) are functionally
equivalent. The Plant Journal, 19, 679–689.
Hoecker U., Vasil I.K., McCarty D.R. (1999) Signaling from
the embryo conditions Vp1-mediated repression of
alpha-amylase genes in the aleurone of developing maize
seeds. The Plant Journal, 19, 371–377.
Jefferson R.A., Kavanagh T.A., Bevan M.W. (1987) Gus
fusions – beta-glucuronidase as a sensitive and versatile
gene fusion marker in higher-plants. Embo Journal, 6,
3901–3907.
Jofuku K.D., Omidyar P.K., Gee Z., Okamuro J.K. (2005)
Control of seed mass and seed yield by the floral homeotic
gene APETALA2. Proceedings of the National Academy of
Sciences of the United States of America, 102, 3117–3122.
Kiyosue T., Ohad N., Yadegari R., Hannon M., Dinneny J.,
Wells D., Katz A., Margossian L., Harada J.J.,
Goldberg R.B., Fischer R.L. (1999) Control of
fertilization-independent endosperm development by the
MEDEA polycomb gene in Arabidopsis. Proceedings of the
National Academy of Sciences of the United States of America,
96, 4186–4191.
Kohler C., Grossniklaus U. (2002) Epigenetic inheritance of
expression states in plant development: the role of
Polycomb group proteins. Current Opinion in Cell Biology,
14, 773–779.
Kohler C., Hennig L., Bouveret R., Gheyselinck J.,
Grossniklaus U., Gruissem W. (2003) Arabidopsis MSI1 is
a component of the MEA/FIE Polycomb group complex
and required for seed development. Embo Journal, 22,
4804–4814.
Kosugi S., Ohashi Y., Nakajima K., Arai Y. (1990) An
improved assay for β-glucuronidase in transformed
cells – methanol almost completely suppresses a putative
endogenous β-glucuronidase activity. Plant Science, 70,
133–140.
Lescot M., Dehais P., Thijs G., Marchal K., Moreau Y., Van
de Peer Y., Rouze P., Rombauts S. (2002) PlantCARE, a
database of plant cis-acting regulatory elements and a
portal to tools for in silico analysis of promoter sequences.
Nucleic Acids Research, 30, 325–327.
Livak K.J., Schmittgen T.D. (2001) Analysis of relative gene
expression data using real-time quantitative PCR and the
2-DDCt method. Methods, 25, 402–408.
Lopes M.A., Larkins B.A. (1993) Endosperm origin,
development, and function. Plant Cell, 5, 1383–1399.
Manning K. (1991) Isolation of nucleic acids from plants by
differential solvent precipitation. Analytical Biochemistry,
195, 45–50.
Matys V., Fricke E., Geffers R., Gossling E., Haubrock M.,
Hehl R., Hornischer K., Karas D., Kel A.E.,
Kel-Margoulis O.V., Kloos D.U., Land S.,
Lewicki-Potapov B., Michael H., Munch R., Reuter I.,
Rotert S., Saxel H., Scheer M., Thiele S., Wingender E.
(2003) TRANSFAC (R): transcriptional regulation, from
patterns to profiles. Nucleic Acids Research, 31, 374–378.
Neill S.J., Horgan R., Parry A.D. (1986) The carotenoid and
abscisic acid content of viviparous kernels and seedlings of
Zea mays L. Planta, 169, 87–96.
Ober E.S., Setter T.L. (1992) Water deficit induces abscisic
acid accumulation in endosperm of maize viviparous
mutants. Plant Physiology, 98, 353–356.
Ohto M., Fischer R.L., Goldberg R.B., Nakamura K.,
Harada J.J. (2005) Control of seed mass by APETALA2.
Proceedings of the National Academy of Sciences of the United
States of America, 102, 3123–3128.
Olsen O.A. (2001) Endosperm development: cellularization
and cell fate specification. Annual Review of Plant Physiology
and Plant Molecular Biology, 52, 233–267.
Pabo C.O., Sauer R.T. (1992) Transcription factors –
structural families and principles of DNA recognition.
Annual Review of Biochemistry, 61, 1053–1095.
Price J., Laxmi A., St Martin S.K., Jang J.C. (2004) Global
transcription profiling reveals multiple sugar signal
transduction mechanisms in Arabidopsis. Plant Cell, 16,
2128–2150.
Riechmann J.L., Heard J., Martin G., Reuber L., Jiang C.-Z.,
Keddie J., Adam L., Pineda O., Ratcliffe O.J.,
Samaha R.R., Creelman R., Pilgrim M., Broun P.,
Zhang J.Z., Ghandehari D., Sherman B.K., Yu G.-L.
(2000) Arabidopsis transcription factors: genome-wide
comparative analysis among eukaryotes. Science, 290,
2105–2110.
Riechmann J.L., Ratcliffe O.J. (2000) A genomic perspective
on plant transcription factors. Current Opinion in Plant
Biology, 3, 423–434.
Robertson M. (2004) Two transcription factors are negative
regulators of gibberellin response in the HvSPY-signaling
pathway in barley aleurone. Plant Physiology, 136,
2747–2761.
Schwechheimer C., Bevan M. (1998) The regulation of
transcription factor activity in plants. Trends in Plant
Science, 3, 378–383.
128 Ann Appl Biol 158 (2011) 115–129 © 2010 The AuthorsAnnals of Applied Biology © 2010 Association of Applied Biologists

N.C. Verza et al. Transcription factor profiling in maize endosperm
Souer E., vanHouwelingen A., Kloos D., Mol J., Koes R.(1996) The no apical meristem gene of petunia is requiredfor pattern formation in embryos and flowers and isexpressed at meristem and primordia boundaries. Cell, 85,159–170.
Sung Z.R., Chen L.J., Moon Y.H., Lertpiriyapong K. (2003)Mechanisms of floral repression in Arabidopsis. Current
Opinion in Plant Biology, 6, 29–35.Suzuki M., Latshaw S., Sato Y., Settles A.M., Koch K.E.,
Hannah L.C., Kojima M., Sakakibara H., McCarty D.R.(2008) The maize Viviparous8 locus, encoding a putativeALTERED MERISTEM PROGRAM1-like peptidase,regulates abscisic acid accumulation and coordinatesembryo and endosperm development. Plant Physiology,146, 1193–1206.
Verza N.C., Silva T.R., Neto G.C., Nogueira F.T.S.,Fisch P.H., De Rosa V.E., Rebello M.M., Vettore A.L., daSilva F.R., Arruda P. (2005) Endosperm-preferredexpression of maize genes as revealed bytranscriptome-wide analysis of expressed sequence tags.Plant Molecular Biology, 59, 363–374.
White C.N., Proebsting W.M., Hedden P., Rivin C.J. (2000)Gibberellins and seed development in maize. I. Evidence
that gibberellin/abscisic acid balance governs germinationversus maturation pathways. Plant Physiology, 122,1081–1088.
Yanagisawa S., Akiyama A., Kisaka H., Uchimiya H.,Miwa T. (2004) Metabolic engineering with Dof1transcription factor in plants, improved nitrogenassimilation and growth under low-nitrogen conditions.Proceedings of the National Academy of Sciences of the United
States of America, 101, 7833–7838.Yunes J.A., Neto G.C., Da Silva M.J., Leite A., Ottoboni
L.M.M., Arruda P. (1994) The transcriptional activatorOpaque2 recognizes 2 different target sequencesin the 22-Kd-like alpha-prolamin genes. Plant Cell, 6,237–249.
Yunes J.A., Vettore A.L., da Silva M.J., Leite A., Arruda P.(1998) Cooperative DNA binding and sequencediscrimination by the Opaque2 bZIP factor. Plant Cell, 10,1941–1955.
Zimmermann R., Werr W. (2005) Pattern formation in themonocot embryo as revealed by NAM and CUC3orthologues from Zea mays L. Plant Molecular Biology, 58,669–685.
Ann Appl Biol 158 (2011) 115–129 © 2010 The Authors 129Annals of Applied Biology © 2010 Association of Applied Biologists
Related Documents











