Reproductive Output of Uca vocator (Herbst, 1804) (Brachyura, Ocypodidae) from Three Subtropical Mangroves in Brazil Author(s): Karine Delevati Colpo and Maria Lucia Negreiros-Fransozo Source: Crustaceana, Vol. 76, No. 1 (Jan., 2003), pp. 1-11 Published by: BRILL Stable URL: http://www.jstor.org/stable/20105533 . Accessed: 07/02/2014 12:14 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . BRILL is collaborating with JSTOR to digitize, preserve and extend access to Crustaceana. http://www.jstor.org This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PM All use subject to JSTOR Terms and Conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Reproductive Output of Uca vocator (Herbst, 1804) (Brachyura, Ocypodidae) from ThreeSubtropical Mangroves in BrazilAuthor(s): Karine Delevati Colpo and Maria Lucia Negreiros-FransozoSource: Crustaceana, Vol. 76, No. 1 (Jan., 2003), pp. 1-11Published by: BRILLStable URL: http://www.jstor.org/stable/20105533 .
Accessed: 07/02/2014 12:14
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
BRILL is collaborating with JSTOR to digitize, preserve and extend access to Crustaceana.
http://www.jstor.org
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions

REPRODUCTIVE OUTPUT OF UCA VOCATOR (HERBST, 1804)
(BRACHYURA, OCYPODIDAE) FROM THREE SUBTROPICAL
MANGROVES IN BRAZIL
BY
KARINE DELEVATICOLPO1'2) and MARIA LUCIA NEGREIROS-FRANSOZO1'2) 1 ) NEBECC (Group of Studies on Crustacean Biology, Ecology and Culture)
2) Instituto de Bioci?ncias, Departamento de Zoologia, Campus de Botucatu,
Universidade Estadual Paulista (UNESP), C.P. 510, CEP-18618-000, Botucatu, SP, Brazil
ABSTRACT
Crab fecundity is widely known to vary proportionally to female size, but the female's nutritional
state also has an important effect on egg production. This study evaluates intraspecific variability of
reproductive output by monthly sampling Uca vocator populations from the Itapanha?, Indai?, and
Itamambuca mangroves on the southeastern coast of Brazil. The presence of ovigerous crabs, their
carapace width (CW) and their number of eggs were recorded. Additionally, the productivity of the
mangroves and the content of organic matter of the sediments were analysed in order to estimate food
availability in each locality. Size-specific fecundity relationships were obtained for each population
and compared among the three populations. Ovigerous females from Itamambuca are the largest
and their fecundities are also the highest among the populations studied. These results probably are
associated with the favourable environmental conditions in Itamambuca, as this is a young mangrove
with a high productivity level.
RESUMEN
La fecundidad de los cangrejos es muy variable respecto a la talla de la hembra, pero su
estado nutricional tambi?n puede tener importantes efectos en la producci?n de huevos. Este estudio
estudia la variabilidad intraespec?fica del potencial reproductor de Uca vocator mediante mu?streos
mensuales de poblaciones provenientes de los manglares de Itapanha?, Indai? e Itamambuca, en el
litoral sudeste de Brasil. Se registr? la presencia de hembras ov?geras, su anchura del caparaz?n
(CW) y n?mero de huevos. Asimismo, se analiz? la productividad del manglar y el contenido
en materia org?nica de los sedimentos con el fin de evaluar la disponibilidad del alimento en
cada localidad. La relaci?n talla-fecundidad se obtuvo para cada poblaci?n, compar?ndose las tres
poblaciones. Las hembras ov?geras de Itamambuca son las que tienen mayor talla y fecundidades
m?s elevadas de las tres poblaciones estudiadas. Estos resultados est?n probablemente asociados a
las buenas condiciones ambientales existentes en Itamambuca, que es un manglar joven de elevada
productividad.
? Koninklijke Brill NV, Leiden, 2003
Also available online: www.brill.nl
Crustaceana 76 (1): 1-11
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions

2 KARINE D. COLPO & MARIA L. NEGREIROS-FRANSOZO
INTRODUCTION
Mangroves are highly productive zones from which nutrients and organic matter are exported that supply consuming communities in adjacent estuarine
and coastal regions (Ottman et al., 1965; Macintosh, 1988; Robertson, 1991;
Lee, 1999; Schaeffer-Novelli, 1994). According to Schaeffer-Novelli (1994),
mangrove productivity can be evaluated by structure and maturation degree. The
deposit-feeding ocypodid crabs of the genus Uca are typical representatives of
the mangrove invertebrate fauna along the Brazilian coast. Their feeding and
burrowing activities contribute to the turnover of nutrients in those environments
(Aveline, 1980; Genoni, 1985; Macintosh, 1988).
According to Genoni (1985), food resources may constrain fiddler crab popu
lations. The food availability may regulate growth, reproductive output, and ulti
mately settlement rate and survivorship of both young and adult crabs (Christy,
1978). Their main food resource is organic matter, including the endofauna, spec
imens of which are sorted out from the substrate (Murai et al., 1982). Actual food
supply depends on the productivity of the ecosystem, microbial activity, substrate
texture, and tide action (Twilley et al., 1995; Moura et al., 1998).
Quantification of the egg production, from oog?nesis to actual fecundity, is often
used to assess reproductive potential in brachyuran crabs (e.g., Jones & Simons,
1983; Wenner et al., 1985; Hartnoll & Gould, 1988; Hines, 1989). Fecundity is
widely known to vary according to size, but the nutritional state of parental females
also plays an important role (Rabalais, 1991). The behavioural pattern and mating
system, which both vary considerably among species of fiddler crabs, may also
affect reproductive output in this group (Crane, 1975; Christy & Salmon, 1984).
The objective of this study is to assess intraspecific variability of reproductive
output, by analysing Uca vocator (Herbst, 1804) populations from three mangrove areas along the southeastern coast of Brazil, which have different productivity levels.
MATERIAL AND METHODS
Study sites and crab sampling Fiddler crabs were collected in three different mangroves on the coast of
S?o Paulo State, Brazil: Itapanha? (23?49'14"S 46?09/14//W), Indai? (23?24'51"S
45?03/14//W) and Itamambuca (23?24/43//S 45?01/03//W) (fig. 1).
Samples were taken monthly at each site from August 1999 to July 2000. During
fieldwork, crabs were removed from their burrows with the aid of diving knives.
Each month, a catch effort of 15 min. by two collectors was employed at each site.
The presence of ovigerous crabs was recorded and their proportion in the samples calculated.
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions
REPRODUCTIVE OUTPUT OF UCA VOCATOR 3
Fig. 1. Map of the northern coast of the State of S?o Paulo, indicating sampling sites at Bertioga and
Ubatuba.
Habitat analysis Data on the vegetation of each site were obtained according to C?ntr?n &
Schaeffer-Novelli (1984). Height, basal area, and density of trees per hectare
were measured to estimate the productivity of the mangroves. At each site, three
sediment samples were taken each month from which mean grain sizes were
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions

4 KARINE D. COLPO & MARIA L. NEGREIROS-FRANSOZO
calculated for each season. The organic matter content of sediment samples was
determined by ash-free dry weighing. Since there have been reports of crude oil
contamination over the study region, analyses of oil content and derived substances
were also carried out using N-hexane to separate oil from sediment.
Laboratory work
Maximum carapace width (CW) and the width of the abdomen at the anterior
margin of the 5th somite (AW) were measured with callipers (? 0.01 mm) for all
ovigerous crabs. The egg masses of all ovigerous females were removed and stored
in 70% ethanol until later analysis. Only females carrying early embryos were used
for fecundity estimates. Each clutch was immersed in a 1 : 100 freshwater solution
of 5-6% sodium hypochlorite (NaCIO) to detach the eggs from the pleopods. This
material was agitated for 5 minutes so as to completely dissociate the egg mass.
Subsamples were then obtained using a plankton splitter, from which egg counts
were obtained under a dissecting microscope.
Statistical analyses The characteristics of the mangrove vegetation were compared by the Kruskal
Wallis non-parametric procedure, complemented by Dunn's test. For each season,
the organic matter content in the sediments was compared among the three sites
using a one-way analysis of variance, complemented by the Tukey test (Sokal &
Rohlf, 1979). Median size (CW) of ovigerous crabs was compared among sites using the
Kruskal-Wallis non-parametric procedure, complemented by Dunn's test (Sokal
& Rohlf, 1979). The mean number of eggs (F) was calculated for each site and compared using
a one-way ANOVA, complemented by the Tukey test (Sokal & Rohlf, 1979).
Size-specific fecundity relationships were obtained for each population using the
number of eggs as the dependent variable, and the power function Y = aZb,
in which carapace width (CW) was the reference dimension (Somers, 1991).
The obtained equations were linearized using log10. The equations from each
population were compared using a covariance analysis complemented by the Tukey test. A significance level of 5% was used for rejection of the null hypothesis in all
statistical tests.
RESULTS
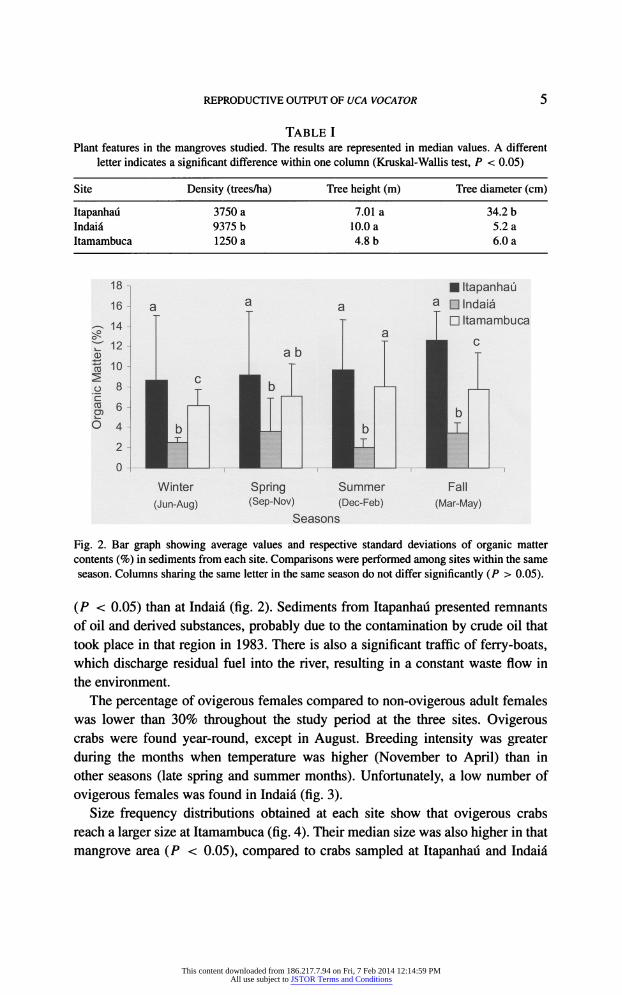
The mangrove characterization of each site is shown in table I. The amount of
organic matter found in the sediments from Itapanhau and Itamambuca was higher
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions
REPRODUCTIVE OUTPUT OF UCA VOCATOR 5
TABLE I Plant features in the mangroves studied. The results are represented in median values. A different
letter indicates a significant difference within one column (Kruskal-Wallis test, P < 0.05)
Site Density (trees/ha) Tree height (m) Tree diameter (cm)
Itapanhau Indai? Itamambuca
3750 a
9375 b 1250 a
7.01a
10.0 a
4.8 b
34.2 b 5.2 a
6.0 a
Itapanhau a B Indai?
Itamambuca
Winter Spring Summer Fall
(Jun-Aug) (Sep-Nov) (Dec-Feb) (Mar-May)
Seasons
Fig. 2. Bar graph showing average values and respective standard deviations of organic matter
contents (%) in sediments from each site. Comparisons were performed among sites within the same
season. Columns sharing the same letter in the same season do not differ significantly (P > 0.05).
(P < 0.05) than at Indai? (fig. 2). Sediments from Itapanhau presented remnants
of oil and derived substances, probably due to the contamination by crude oil that
took place in that region in 1983. There is also a significant traffic of ferry-boats, which discharge residual fuel into the river, resulting in a constant waste flow in
the environment.
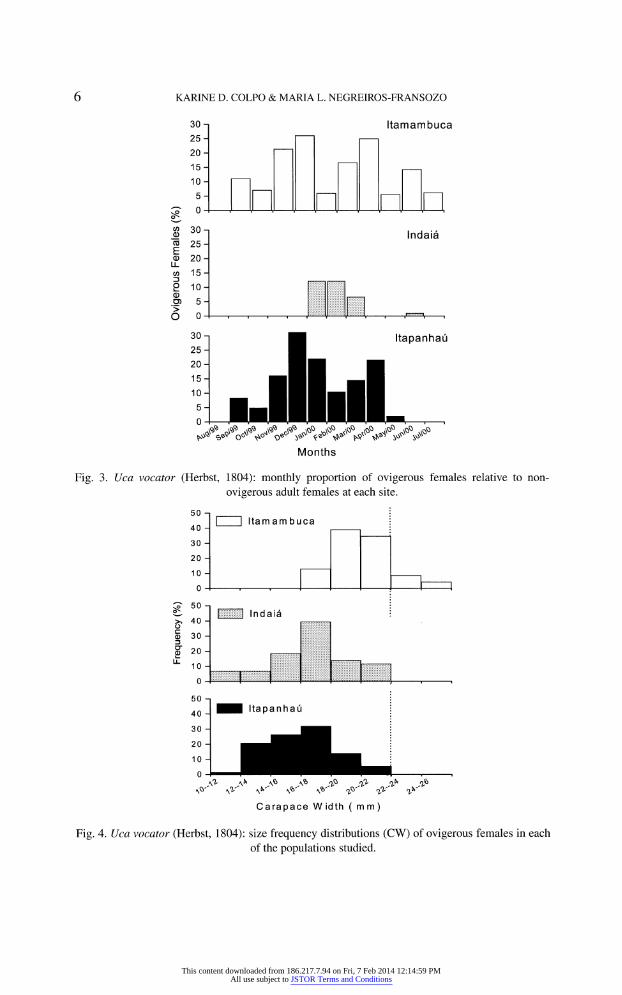
The percentage of ovigerous females compared to non-ovigerous adult females
was lower than 30% throughout the study period at the three sites. Ovigerous crabs were found year-round, except in August. Breeding intensity was greater
during the months when temperature was higher (November to April) than in
other seasons (late spring and summer months). Unfortunately, a low number of
ovigerous females was found in Indai? (fig. 3).
Size frequency distributions obtained at each site show that ovigerous crabs
reach a larger size at Itamambuca (fig. 4). Their median size was also higher in that
mangrove area (P < 0.05), compared to crabs sampled at Itapanhau and Indai?
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions
KARINE D. COLPO & MARIA L. NEGREIROS-FRANSOZO
30-1
25
20
15
10
5
o-L
Itamambuca
8 30
ro 25
<D 20 LL w 15
2 10 H ?) 5 -
? o
Indai?
Itapanhau
//////////// Months
Fig. 3. Uca vocator (Herbst, 1804): monthly proportion of ovigerous females relative to non
ovigerous adult females at each site.
\*' \V N?> ?jO rfl, rjfc
^ N?>' <$>' <$,'
Carapace Width ( mm)
Fig. 4. Uca vocator (Herbst, 1804): size frequency distributions (CW) of ovigerous females in each
of the populations studied.
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions
REPRODUCTIVE OUTPUT OF UCA VOCATOR 7
Itapanhau Indai? Itamambuca
Sites
I Min-Max
I I 25%-75%
o Median
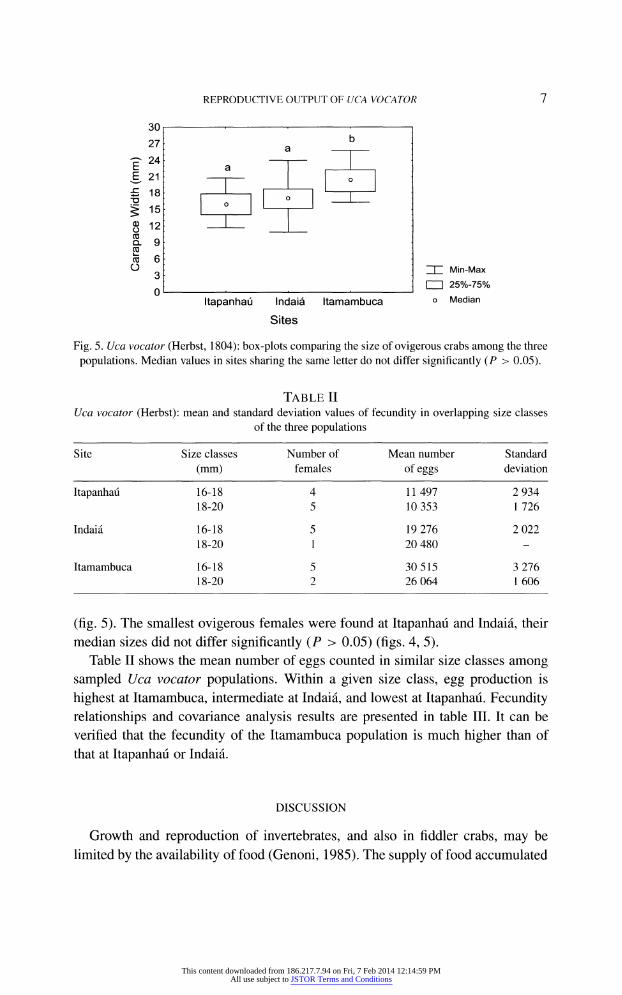
Fig. 5. Uca vocator (Herbst, 1804): box-plots comparing the size of ovigerous crabs among the three
populations. Median values in sites sharing the same letter do not differ significantly (P > 0.05).
Table II Uca vocator (Herbst): mean and standard deviation values of fecundity in overlapping size classes
of the three populations
Site Size classes
(mm)
Number of
females
Mean number
of eggs
Standard
deviation
Itapanhau
Indai?
Itamambuca
16-18
18-20
16-18
18-20
16-18
18-20
11497 10 353
19 276 20 480
30 515 26 064
2 934 1726
2 022
3 276 1606
(fig. 5). The smallest ovigerous females were found at Itapanhau and Indai?, their
median sizes did not differ significantly (P > 0.05) (figs. 4, 5).
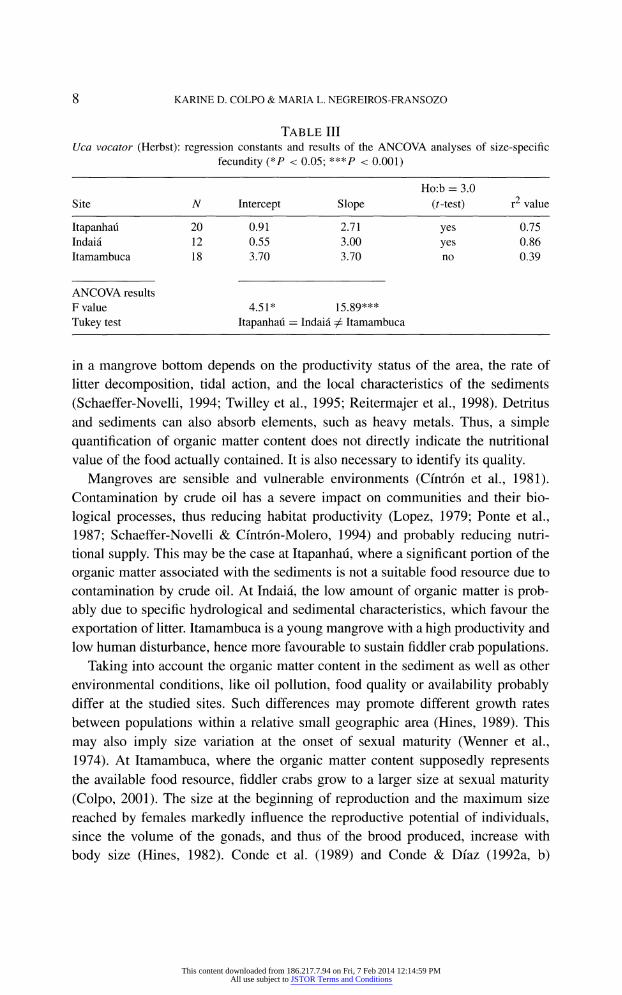
Table II shows the mean number of eggs counted in similar size classes among
sampled Uca vocator populations. Within a given size class, egg production is
highest at Itamambuca, intermediate at Indai?, and lowest at Itapanhau. Fecundity
relationships and covariance analysis results are presented in table III. It can be
verified that the fecundity of the Itamambuca population is much higher than of
that at Itapanhau or Indai?.
DISCUSSION
Growth and reproduction of invertebrates, and also in fiddler crabs, may be
limited by the availability of food (Genoni, 1985). The supply of food accumulated
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions
? KARINE D. COLPO & MARIA L. NEGREIROS-FRANSOZO
Table III Uca vocator (Herbst): regression constants and results of the ANCOVA analyses of size-specific
fecundity (*P < 0.05; ***P < 0.001)
Ho:b = 3.0
Site N Intercept Slope (i-test) r2 value
Itapanhau 20 0.91 2.71 yes 0.75
Indai? 12 0.55 3.00 yes 0.86 Itamambuca 18 3.70 3.70 no 0.39
ANCOVA results F value 4.51* 15.89***
Tukey test Itapanhau = Indai? ^ Itamambuca
in a mangrove bottom depends on the productivity status of the area, the rate of
litter decomposition, tidal action, and the local characteristics of the sediments
(Schaeffer-Novelli, 1994; Twilley et al., 1995; Reitermajer et al, 1998). Detritus
and sediments can also absorb elements, such as heavy metals. Thus, a simple
quantification of organic matter content does not directly indicate the nutritional
value of the food actually contained. It is also necessary to identify its quality.
Mangroves are sensible and vulnerable environments (C?ntr?n et al., 1981).
Contamination by crude oil has a severe impact on communities and their bio
logical processes, thus reducing habitat productivity (Lopez, 1979; Ponte et al.,
1987; Schaeffer-Novelli & C?ntr?n-Molero, 1994) and probably reducing nutri
tional supply. This may be the case at Itapanhau, where a significant portion of the
organic matter associated with the sediments is not a suitable food resource due to
contamination by crude oil. At Indai?, the low amount of organic matter is prob
ably due to specific hydrological and sedimental characteristics, which favour the
exportation of litter. Itamambuca is a young mangrove with a high productivity and
low human disturbance, hence more favourable to sustain fiddler crab populations.
Taking into account the organic matter content in the sediment as well as other
environmental conditions, like oil pollution, food quality or availability probably differ at the studied sites. Such differences may promote different growth rates
between populations within a relative small geographic area (Hines, 1989). This
may also imply size variation at the onset of sexual maturity (Wenner et al.,
1974). At Itamambuca, where the organic matter content supposedly represents
the available food resource, fiddler crabs grow to a larger size at sexual maturity
(Colpo, 2001). The size at the beginning of reproduction and the maximum size
reached by females markedly influence the reproductive potential of individuals,
since the volume of the gonads, and thus of the brood produced, increase with
body size (Hines, 1982). Conde et al. (1989) and Conde & D?az (1992a, b)
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions

REPRODUCTIVE OUTPUT OF UCA VOCATOR 9
reported this relationship between body size and reproductive effort in connection
with the productivity of mangrove forests in Aratus pisonii (H. Milne Edwards,
1837), a grapsid crab feeding on mangrove leaves. However, environmental factors
influence the absolute egg number (Jensen, 1958). During oog?nesis, there is a
great demand for proteins and lipids (Rabalais, 1991). Huner & Romaire (1979)
and Taylor (1983) reported that the freshwater crayfish, Procambarus clarkii
(Girard, 1852) and Procambarus spiculifer (Le Conte, 1856) reach higher size
and fecundity under favourable nutritional conditions as compared to those in an
unfavourable habitat. As mentioned by Momot (1986) and Huner & Lindqvist
(1991), environmental conditions such as food and water quality also potentially influence egg production in the freshwater crayfish, Orconectes virilis (Hagen,
1870). The fecundity of Uca vocator in Itamambuca was higher than in the other
two populations investigated. Such contrasting reproductive output was not only due to size differences. The brood size produced by females of similar size was
also higher at Itamambuca, which indicates a higher reproductive potential in
this population. Thus, it is suggested that favourable nutritional conditions are
promoting optimal reproductive output in that population.
ACKNOWLEDGEMENTS
Thanks are due to NEBECC co-workers for their kind help during fieldwork
and laboratory procedures. To F?lvio Freir? for drawing the map presented here.
To CNPq for a master science fellowship for the first author and to FAPESP for
financing part of the fieldtrips for the co-author (#94/4878-8; #98/03134-6).
LITERATURE CITED
Aveline, L. C, 1980. Fauna dos manguezais brasileiros. Rev. Brasileira Geogr., 42: 786-821.
Christy, J., 1978. Adaptative significance of reproductive cycles in the fiddler crab Uca pugllator. a hypothesis. Science, New York, 199: 453-455.
Christy, J. & M. Salmon, 1984. Ecology and evolution of mating systems of fiddler crabs (genus
Uca). Biol. Rev., 59: 483-509.
C?ntr?n, G., A. E. Lugo, R. Martinez, B. B. C?ntr?n & L. Encarnaci?n, 1981. Impact
of oil in the tropical marine environment: 1-29. (Technical Publication, Division of Marine
Resources. Department of natural Resources of Puerto Rico).
C?ntr?n, G. & Y. Schaeffer-Novelli, 1984. Methods for studying mangrove structure. In:
S. C. Snedaker & J. G. Snedaker (eds.), The mangrove ecosystem: research methods, 1:
91-113. (Unesco, Rome).
Colpo, K. D., 2001. Biologia populacional comparativa de Uca vocator (Herbst, 1804) (Brachyura,
Ocypodidae) em tres localidades do litoral norte Paulista: 1-104. (M.Sc. Thesis, Instituto de
Bioci?ncias Universidade Estadual Paulista, Botucatu).
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions

10 KARINE D. COLPO & MARIA L. NEGREIROS-FRANSOZO
Conde, J. E. & H. D?AZ, 1992a. Variations in intraspecific relative size at the onset of maturity
(RSOM) in Aratus pisonli (H. Milne Edwards, 1837) (Decapoda, Brachyura, Grapsidae).
Crustaceana, 62: 214-216.
-&-,1992b. Extension of the stunting range in ovigerous female of the mangrove crab
Aratus plsonii (H. Milne Edwards, 1837) (Decapoda, Brachyura, Grapsidae). Crustaceana, 62:
319-322.
Conde, J. E., H. D?az & G. Rodriguez, 1989. Crescimiento reducido en el cangrejo de mangle
Aratus plsonii (H. Milne Edwards) (Brachyura: Grapsidae). Acta cient. Venezolana, 40: 159
160.
Crane, J., 1975. Fiddler crabs of the world, Ocypodidae: genus Uca: 1-736. (Princeton University
Press, New Jersey).
Genoni, G. P., 1985. Food limitation in salt marsh fiddler crabs Uca rapax (Smith) (Decapoda:
Ocypodidae). Journ. exp. mar. Biol. Ecol., 87: 97-110.
HARTNOLL, R. G. & P. GOULD, 1988. Brachyuran life history strategies and the optimization of
egg production. Symp. zool. Soc, London, 59: 1-9.
Hines, A. H., 1982. Allometric constraints and variables of reproductive effort in brachyuran crabs.
Mar. Ecol., 69: 309-320.
-, 1989. Geographic variation in size at maturity in brachyuran crabs. Bull. mar. Sei., 45 (2):
356-368.
Huner, J. V. & O. V. Lindqvist, 1991. Special problems in freshwater crayfish egg production. In: A. Wenner & A. Kuris (eds.), Crustacean egg production. Crust. Issues, 1: 235-246.
(A. A. Balkema, Rotterdam).
Huner, J. V & R. P. Romaire, 1979. Size at maturity as a means of comparing populations
of Procambarus clarkii (Girard) (Crustacea, Decapoda) from different habitats. Freshwater
Crayfish, 4: 53-64.
Jensen, J. P., 1958. The relation between body size and number of eggs in marine Malacostraca.
Meddel. Danmarks Fisk- og Havunders0g., (n.s.) 2: 1-25.
Jones, M. B. & M. J. Simons, 1983. Latitudinal variation in reproductive characteristics of a mud
crab, Heiice crassa (Grapsidae). Bull. mar. Sei., 33 (3): 656-670.
Lee, S. Y, 1999. The effect of mangrove leaf litter enrichment on macrobenthic colonization of
defaulted sandy substrates. Estuar. coast. Shelf Sei., 49: 703-712.
Lopez, J. M., 1979. Studies on mangroves of Guayanilla Bay. Proceedings of the Symposium of
Energy Industry and the Marine Environment of Guayanilla Bay: 73-82.
Macintosh, D. J., 1988. The ecology and physiology of decapods of mangrove swamps. Symp.
zool. Soc, London, 59: 315-341.
Moura, D. E., C. C. Lamparelli, F. O. Rodrigues & R. C. Vincent, 1998. Decomposi?ao
de folhas em manguezais na regi?o de Bertioga, S?o Paulo, Brasil. In: Proceedings of the IV
Simposio de Ecossistemas Brasileiros, 1: 130-148.
MURAI, M., S. Goshima & Y. NAKASONE, 1982. Some behavioral characteristics related to food
supply and soil texture of burrowing habitats observed on Uca vocans and U. l?ctea perplexa.
Mar. Biol., Berlin, 66: 191-197.
Ottmann, F., T. Okuda, L. Cavalcanti, O. C. Silva, J. V. A. Ara?jo, P. A. Coelho,
M. N. Paranagu? & E. Eskinazi, 1965. Estudo da Barra das Jangadas-Parte, V Efeitos da
poluic?o sobre a ecologia do estuario. Trab. Inst. Ocean. Univ. fed. Pernambuco, 7/8: 7-16.
Ponte, A. C. E., I. A. Z. Fonseca & S. M. C. A. Claro, 1987. Impacto causado por petr?leo
no manguezal do Canal da Bertioga ? Estrutura da vegetac?o. In: Proceedings of the Simposio
sobre Ecosistemas da Costa Sul e Sudeste: s?ntese dos conhecimentos, 54: 138-147.
Rabal ais, N. N., 1991. Egg production in crabs with abbreviated development. In: A. Wenner
& A. Kuris (eds.), Crustacean egg production. Crust. Issues, 1: 217-234. (A. A. Balkema,
Rotterdam).
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions

REPRODUCTIVE OUTPUT OF UCA VOCATOR 1 1
Reitermajer, D., J. C. Viana, A. F. D. S. Queiroz, R. M. Barbosa, S. A. Rocha
& J. B. Souza, 1998. Caracteriza?ao da distribui?ao da materia org?nica em zonas de
manguezais do estuario do rio Sau?pe/ Entre Rios-BA. In: Proceedings of the IV Simposio
de Ecossistemas Brasileiros, 1: 195-201.
Robertson, A. L, 1991. Plant-animal interactions and the structure and function of mangrove
forest ecosystems. Australian Journ. Ecol., 16: 433-443.
Schaeffer-Novelli, Y, 1994. Manguezais. In: Diagn?stico ambiental oce?nico e costeiro das
regi?es sul e sudeste do Brasil: 128-196. (Fundac?o de Estudos em Pesquisas Aqu?ticas ?
FUNDESPA). Schaeffer-Novelli, Y. & G. C?ntr?n-Molero, 1994. Manguezais brasileiros: urna s?ntese
sobre os aspectos hist?ricos (s?culos XVI a XIX), zona?ao, estrutura e impactos ambient?is.
In: Proceedings of the III Simposio de Ecossistema da Costa Brasileira, 1: 333-341.
Sokal, R. R. & F. J. Rohlf, 1979. Biometr?a. Principios y m?todos estad?sticos en la investigaci?n
biol?gica: 1-832. (H. Blume Ediciones, Madrid).
Somers, K. M., 1991. Characterizing size-specific fecundity in crustaceans. In: A. Wenner
& A. Kuris (eds.), Crustacean egg production. Crust. Issues, 1: 357-378. (A. A. Balkema,
Rotterdam).
Taylor, R. C, 1983. Drought-induced changes in crayfish populations along a stream continuum.
American Midi. Natural., 110: 286-298.
Twilley, R. R., S. C. Snedaker, A. Y??ez-Arancibia& E. Medina, 1995. Mangroves sys
tems. In: V. H. He Y WOOD (ed.), Global biodiversity assessment, Biodiversity and ecosystem
function: ecosystem analyses, 1: 387-393. (Cambridge University Press, Cambridge, U.K.).
Wenner, A. M., C. Fusaro & A. Oaten, 1974. Size at onset of sexual maturity and growth rate
in crustacean populations. Canadian Journ. Zool., 52 (9): 1095-1106.
Wenner, A. M., H. M. Page & P. R. Siegel, 1985. Variation in size at onset of egg production. In: A. M. Wenner (ed.), Factors in adult growth. Crust. Issues, 1: 149-164. (A. A. Balkema,
Rotterdam).
First received 28 February 2002.
Final version accepted 24 October 2002.
This content downloaded from 186.217.7.94 on Fri, 7 Feb 2014 12:14:59 PMAll use subject to JSTOR Terms and Conditions
Related Documents











