Reproductive Ecology of Black Mangrove (Avicennia germinans) Along the Louisiana Coast: Propagule Production Cycles, Dispersal Limitations, and Establishment Elevations Lauren K. Alleman & Mark W. Hester Received: 30 April 2010 /Revised: 29 October 2010 / Accepted: 11 April 2011 # Coastal and Estuarine Research Federation 2011 Abstract Black mangrove (Avicennia germinans) reprodu- ces by producing propagules through the process of cryptovivipary, which requires a significant energy invest- ment. We conducted a series of field and greenhouse studies to track propagule production in years of high and low disturbances (i.e., hurricanes), the effects of time and salinity on propagule dispersal potential, and the relationship between hydrology and propagule establishment elevations. Trees tended to produce greater numbers of propagules in years after hurricanes, and individual trees alternated the amount of energy they invested in reproduction in consec- utive years. In the greenhouse, propagule buoyancy was affected by salinity with propagules in 36 remaining buoyant for over 110 days and establishment success decreasing with dispersal period length. Finally, a field survey across five sites in southeastern Louisiana revealed that the 2009 cohort of propagules established at significantly lower elevations than mature trees occurred. These findings elucidate some salient features of black mangrove reproductive biology in the northern portion of its North American distribution. Keywords Avicennia germinans . Propagule dispersal . Louisiana . Elevation . Reproduction Introduction The range expansion of Avicennia germinans throughout the Gulf of Mexico and along the coasts of Central and South America is thought to be the result of long-distance dispersal events as evidenced by low genetic differentiation between West African and western Atlantic mangroves (Nettel and Dodd 2007). The propagules of A. germinans are cryptovi- viparous, buoyant, and salt-tolerant which allow for long- distance dispersal events. It is hypothesized that vivipary evolved as a reproductive strategy to facilitate the coloniza- tion of high-energy environments, whereby the embryo does not undergo a period of dormancy but rather germinates and develops while remaining attached to the parent tree, thus reducing the time to establishment (Farnsworth 2000). Supplied with maternal carbohydrates and nutrients during development, the cotyledons contain sufficient reserves for long dispersal periods (Farnsworth 2000). When A. germi- nans propagules reach maturity and are abscised from the parent tree, they undergo an “obligate dispersal period” (Rabinowitz 1978) during which they develop roots while floating on tidal currents until they arrive at suitable exposed substrate and establish. The process of vivipary is common among mangrove genera but not requisite (Tomlinson 1986; Saenger 2002). It has been suggested by Sousa et al. (2007) that “supply-side ecology” (Lewin 1986) may be as influential as abiotic factors or biotic interactions in determining the zonation, abundance, and composition of a mangrove community. As such, the arrival of large numbers of propagules at a site may initially outweigh other establish- ment factors, such as physiological tolerances or competi- tion (Sousa et al. 2007). However, once dispersed to a site, abiotic and biotic factors influence propagule stranding, successful establishment (Delgado et al. 2001), the “main- L. K. Alleman (*) Great Ecology & Environments, Inc., 2231 Broadway, New York, NY 10024, USA e-mail: [email protected] M. W. Hester Coastal Plant Ecology Laboratory, Department of Biology, University of Louisiana at Lafayette, Lafayette, LA 70503, USA Estuaries and Coasts DOI 10.1007/s12237-011-9404-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Reproductive Ecology of Black Mangrove (Avicenniagerminans) Along the Louisiana Coast: PropaguleProduction Cycles, Dispersal Limitations,and Establishment Elevations
Lauren K. Alleman & Mark W. Hester
Received: 30 April 2010 /Revised: 29 October 2010 /Accepted: 11 April 2011# Coastal and Estuarine Research Federation 2011
Abstract Black mangrove (Avicennia germinans) reprodu-ces by producing propagules through the process ofcryptovivipary, which requires a significant energy invest-ment. We conducted a series of field and greenhouse studiesto track propagule production in years of high and lowdisturbances (i.e., hurricanes), the effects of time and salinityon propagule dispersal potential, and the relationshipbetween hydrology and propagule establishment elevations.Trees tended to produce greater numbers of propagules inyears after hurricanes, and individual trees alternated theamount of energy they invested in reproduction in consec-utive years. In the greenhouse, propagule buoyancy wasaffected by salinity with propagules in 36 remaining buoyantfor over 110 days and establishment success decreasing withdispersal period length. Finally, a field survey across fivesites in southeastern Louisiana revealed that the 2009 cohortof propagules established at significantly lower elevationsthan mature trees occurred. These findings elucidate somesalient features of black mangrove reproductive biology inthe northern portion of its North American distribution.
Keywords Avicennia germinans . Propagule dispersal .
Louisiana . Elevation . Reproduction
Introduction
The range expansion of Avicennia germinans throughout theGulf of Mexico and along the coasts of Central and SouthAmerica is thought to be the result of long-distance dispersalevents as evidenced by low genetic differentiation betweenWest African and western Atlantic mangroves (Nettel andDodd 2007). The propagules of A. germinans are cryptovi-viparous, buoyant, and salt-tolerant which allow for long-distance dispersal events. It is hypothesized that viviparyevolved as a reproductive strategy to facilitate the coloniza-tion of high-energy environments, whereby the embryo doesnot undergo a period of dormancy but rather germinates anddevelops while remaining attached to the parent tree, thusreducing the time to establishment (Farnsworth 2000).Supplied with maternal carbohydrates and nutrients duringdevelopment, the cotyledons contain sufficient reserves forlong dispersal periods (Farnsworth 2000). When A. germi-nans propagules reach maturity and are abscised from theparent tree, they undergo an “obligate dispersal period”(Rabinowitz 1978) during which they develop roots whilefloating on tidal currents until they arrive at suitable exposedsubstrate and establish. The process of vivipary is commonamong mangrove genera but not requisite (Tomlinson 1986;Saenger 2002).
It has been suggested by Sousa et al. (2007) that“supply-side ecology” (Lewin 1986) may be as influentialas abiotic factors or biotic interactions in determining thezonation, abundance, and composition of a mangrovecommunity. As such, the arrival of large numbers ofpropagules at a site may initially outweigh other establish-ment factors, such as physiological tolerances or competi-tion (Sousa et al. 2007). However, once dispersed to a site,abiotic and biotic factors influence propagule stranding,successful establishment (Delgado et al. 2001), the “main-
L. K. Alleman (*)Great Ecology & Environments, Inc.,2231 Broadway,New York, NY 10024, USAe-mail: [email protected]
M. W. HesterCoastal Plant Ecology Laboratory, Department of Biology,University of Louisiana at Lafayette,Lafayette, LA 70503, USA
Estuaries and CoastsDOI 10.1007/s12237-011-9404-8

tenance” of long-term mangrove zonation (Ellison andFarnsworth 1993), and the long-term sustainability of aviable population (McKee 1995). The elevation at which A.germinans propagules strand and establish plays a crucialrole in whether they will survive to sexual maturity(Patterson et al. 1997). This establishment elevation andits relationship to local hydrology is a critical considerationin restoration (Kitaya et al. 2002; Lewis 2005). In fact, localhydrology has been invoked as the most fundamentalprinciple in mangrove restoration, and many projects havefailed because they did not adequately address thiscomponent (Lewis 2005; Sampson and Rollon 2008).
Rabinowitz (1978) published some of the first studies onA. germinans propagule biology and reported that Avicen-nia propagules had the longest buoyant period of fivePanamanian mangrove species (>90 days), although theviability of the propagules decreased with the length of timespent in the water. While the cotyledons of the propaguleshave sufficient reserves for long dispersal periods, theirrecruitment is limited by factors such as propagulepredation, insect herbivory, desiccation, or decay (Ellisonand Farnsworth 1993; McKee 1995; Patterson et al. 1997;Allen et al. 2003). Numerous studies have been conductedto understand the role of Avicennia propagules in mangroveintertidal zonation (Smith 1987; Ellison and Farnsworth1993; Delgado et al. 2001; Clarke et al. 2001; Sousa et al.2007). Yet, questions remain regarding the dispersal potentialof propagules along salinity gradients, the viability of apropagule after a long dispersal period, and how propaguledevelopment during dispersal relates to establishment.Delgado et al. (2001) documented the growth of long rootsand the retention of buoyancy of A. germinans propagulesduring a 25-day dispersal period. Root development may beaffected by water turbulence; McMillan (1971) found thatA. germinans propagule root development was inhibited withturbulence. Clarke and Allaway (1993) found that propa-gules of Avicennia marina, an Indo-Pacific congener, havean obligate dispersal period of several weeks and remainviable for several months if they do not strand on sediment.
Though studies on propagule biology have been undertakenthroughout the neotropics, few have been tailored to Louisiana(but, see Patterson et al. 1997 and Pickens and Hester 2010).In addition to improving the understanding of the dispersalpotential and supply-side ecology of A. germinans at thenorthern boundary of its North American distribution, thereproductive ecology of the species has applications for therestoration of Louisiana’s salt marshes and barrier islands(e.g., A. germinans provides critical nesting habitat for thebrown pelican; Visser et al. 2005). Natural populations ofA. germinans are often utilized as a source of propagulesand/or seedlings for restoration. Additionally, hurricanes area regular disturbance in Louisiana, and a better compre-hension of A. germinans energy allocation and recovery
following these events is warranted. The overarching goalof this paper is to elucidate the key features of thereproductive ecology of A. germinans along the Louisianacoast by investigating the following: (1) propagule produc-tion cycles of adult trees of various size classes across yearswith severe hurricane disturbance or little disturbance, (2)constraints on propagule buoyancy, root development,dispersal potential, and viability, and (3) propagule estab-lishment patterns and local hydrology.
Materials and Methods
Propagule Production Cycles
Our study site was a coastal salt marsh of the Caminada-Moreau erosional headland, part of the Lafourche deltacomplex, which was an ancestral Mississippi delta lobeabandoned approximately 400 years ago (Boyd andPenland 1988). Specifically, the population of A. germi-nans in this study is located in the salt marsh that bordersBay Champagne, at approximately 29°6′22″N, 90°10′59″W. The Caminada-Moreau headland is experiencing someof the highest rates of shoreline erosion in the state,estimated at approximately 33 m of lateral erosionannually (McBride and Byrnes 1997). The region washeavily impacted by storm-surge inundation and overwashfrom Hurricane Katrina (landfall August 29, 2005) andHurricane Gustav (landfall September 1, 2008).
Vegetation of this erosional headland formerly includeddistinct dune, swale, and back-barrier marsh zones. Dominantbeach and dune vegetation included: beach tea (Crotonpunctatus), beach morning glory (Ipomoea spp.), marshhaycordgrass (Spartina patens), and common reed (Phragmitesaustralis; Hester and Mendelssohn 1991; Hester et al. 2005).However, there was extensive shoreline retreat, overwash,and re-deposition of dune sands into swale and marshhabitats in the 2005, and especially the 2008, hurricaneseasons. As a result, the high salt marsh zone rapidlyintergrades to swale and is vegetated with a mixed plantcommunity of marshhay cordgrass (S. patens), halophytes,such as glasswort (Salicornia spp.), as well smooth cordgrass(Spartina alterniflora) and black mangrove (A. germinans).
The first field study was initiated October 19, 2005,post-Hurricane Katrina at which time 50 randomly selectedtrees of representative size classes were tagged withindividual identification numbers. As the intent of thisstudy was to monitor reproductive trends, immature treeswere excluded from the selection process and only sexuallymature trees (or those becoming so) were chosen based onheight, branching patterns, and/or the presence of prop-agules. The following measurements were taken on each ofthe 50 trees: tree height, canopy diameter in two directions
Estuaries and Coasts

(first diameter being the widest region of the canopy, withthe second diameter at 90° to the first), elevation, and totalnumber of propagules produced. A subset of 10% of the totalnumber of propagules was collected and measured for length,width, and dry mass. Soil samples were taken at each tree andanalyzed for pH, salinity, and percent organic matter (loss onignition). These methods were repeated in 2006 and 2007within ±3 calendar days of October 19, around the time ofpeak propagule maturity prior to abscission (Hester, pers. obs.).However, on September 1, 2008, Hurricane Gustav madelandfall at Cocodrie, Louisiana (approximately 30 miles westof the study site) and deposited roughly 1.3 m of sand from thedunes onto the original 50 trees, causing massive mortality.Consequently, 50 new representative trees were selectedalong the edge of Bay Champagne slightly to the east of theoriginal site and the annual census was repeated in 2008 and2009 on those trees. A repeated-measures analysis of variance(ANOVA) was conducted to test for significant differencesamong years in height, area, and propagule production.
Propagule Buoyancy, Root Development, and Viability
The purpose of the propagule buoyancy study was toinvestigate the effects of salinity on propagule buoyancy,longevity, and viability. Additionally, we wished to evaluatethe probability of long-distance dispersal events given thesefindings. Three salinity levels were selected to span a rangeof riverine (0), estuarine (18), and full-strength sea water(36). The propagules, which were collected from theCaminada-Moreau headland population on November 15,2007, were transported from the coast in buckets ofartificial sea water at 24 (the salinity of the water atCaminada-Moreau). The majority of pericarps were shed bythe time the propagules reached the Center for Ecology andEnvironmental Technology in Lafayette, Louisiana, about24 h later, and those pericarps that were not shed werephysically removed (pericarps are shed in the field shortlyafter propagule abscission). Individual propagules wereplaced inside cylindrical plastic mesh enclosures that allowedpropagules to move vertically within the water column whileproviding a mechanism to keep track of individual propagulesfor the duration of the study. Larger reservoirs contained theenclosures within each salinity level (two reservoirs persalinity). Water and salinity levels were checked regularlyand fresh water added as necessary. Aerator stones werehaphazardly placed in reservoirs at the initiation of the studyand were moved sporadically throughout the 110 daysapproximately every 14 days, to avoid any potential unseeneffect of propagule proximity to aerator stone on buoyancy.
At the beginning of this study, all propagules werebuoyant and floating on the surface of the water. Thepropagules were monitored every 2 to 3 days for changes inposition (i.e., loss of buoyancy). The number of days a
propagule required to lose buoyancy and sink to the bottom ofthe enclosure was recorded, and any subsequent floating andsinking movements were also noted. After 110 days, the studywas terminated. All propagules were removed from theirenclosure and their roots scanned and analyzed for total rootlength, root surface area, and number of root tips usingWinRhizo software (Regent Instruments, Inc. 2005).
A second study was designed to test the effects ofdispersal period length on viability. Propagules werecollected and transported from the Caminada-Moreauheadland population mentioned above and floated in 11.4-l buckets with an aerator stone in one of three salinity levels(0, 18, or 36). There were five buckets per salinity level,with ten propagules per bucket. Salinity and water levelswere checked regularly and adjusted as necessary. Prop-agules were monitored every 2 to 3 days for changes inposition, and upon sinking the propagule was removedfrom the reservoir and stranded on sand that was main-tained at the same salinity level to which the propagule hadbeen assigned. The time to establishment (or lack thereof)was monitored and recorded to observe the effect ofdispersal period length on establishment success.
Propagule Establishment Elevations
A second field study was conducted in June 2009 to: (1)survey the elevations at which black mangrove propagulesestablished in Louisiana salt marshes during the 2008–2009dispersal period and (2) relate this to local hydrology. Inearly June of 2009, five populations of A. germinans were
Fig. 1 Five A. germinans populations surveyed in June 2009. Awater-level gauge was located at Site 4
Estuaries and Coasts
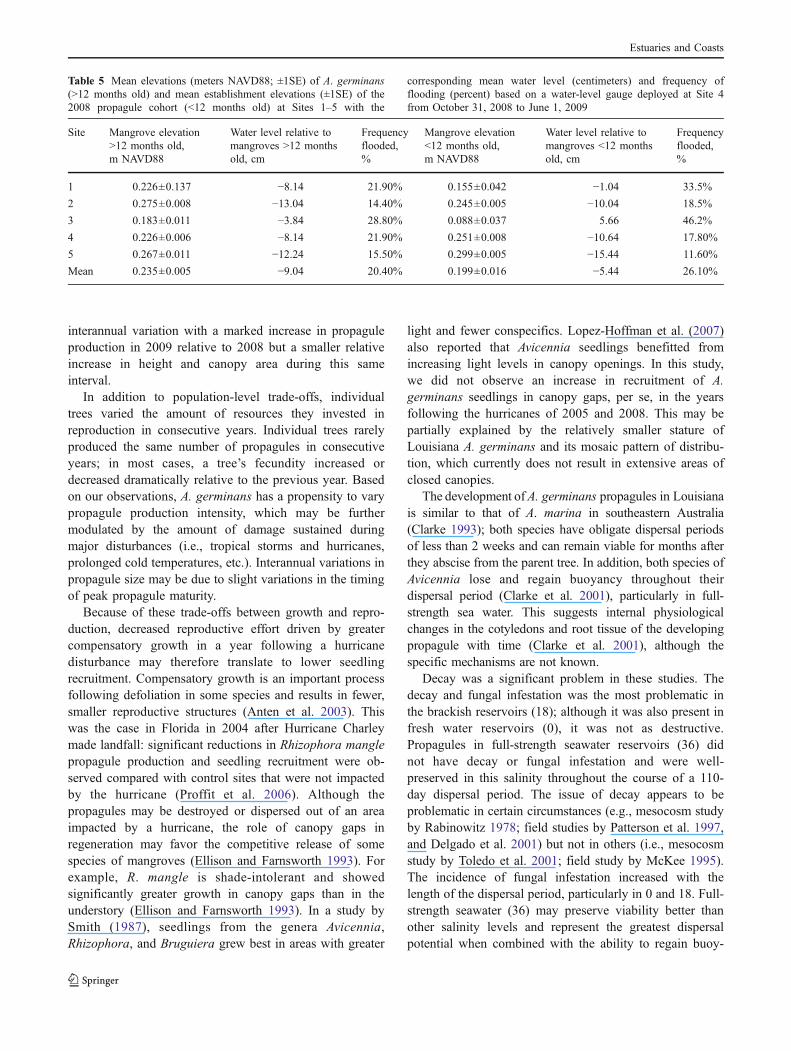
selected along a north-south gradient from the Caminada-Moreau headland (Fig. 1). The height and elevation of atleast 50 trees were surveyed at each site, including as manynewly established propagules as could be located. “Newlyestablished propagules” were classified as small seedlingsthat were shorter than 30 cm and had cotyledons (embry-onic structures) still attached to the stem. According toRabinowitz (1978), cotyledons of A. germinans remainattached to the seedling for months after establishment.Thus, we are confident that the seedlings with cotyledonsstill attached were members of the 2008 propagule cohortand had established between the fall of 2008 and the springof 2009 (out of the 272 trees that were sampled, 54 wereconsidered to be part of the 2008 cohort). Our research labhad previously deployed a water-level gauge located in theinterior marsh of the greater Lafourche delta headland, atapproximately 29°11′29″N, 90°14′55″W. This water-levelgauge was operational during the dispersal period of thepropagules, from approximately October 2008 to June2009, and was used to calculate the mean water levels ateach population. The benchmarks and the water-level gaugewere tied into the North American Vertical Datum of 1988(NAVD88) using the GULFNet GPS Real-Time Networkthat utilizes continuously operating reference stations (LSUCenter for Geomatics 2009). Differences in elevationbetween age classes were analyzed with ANOVA.
Results
Propagule Production Cycles
Hurricanes Katrina (2005) and Gustav (2008) impacted theCaminada-Moreau headland with high winds and stormsurges that resulted in the complete inundation of theheadland and marsh. The response of the trees affected byHurricane Katrina are captured in 2005 through 2007 data,after which all of the original 50 trees tagged in 2005 weredestroyed by Hurricane Gustav (2008), necessitating theselection of 50 new trees that were sampled in 2008 and2009. Interesting trends in propagule production andvegetative growth were observed for both sets of sampled
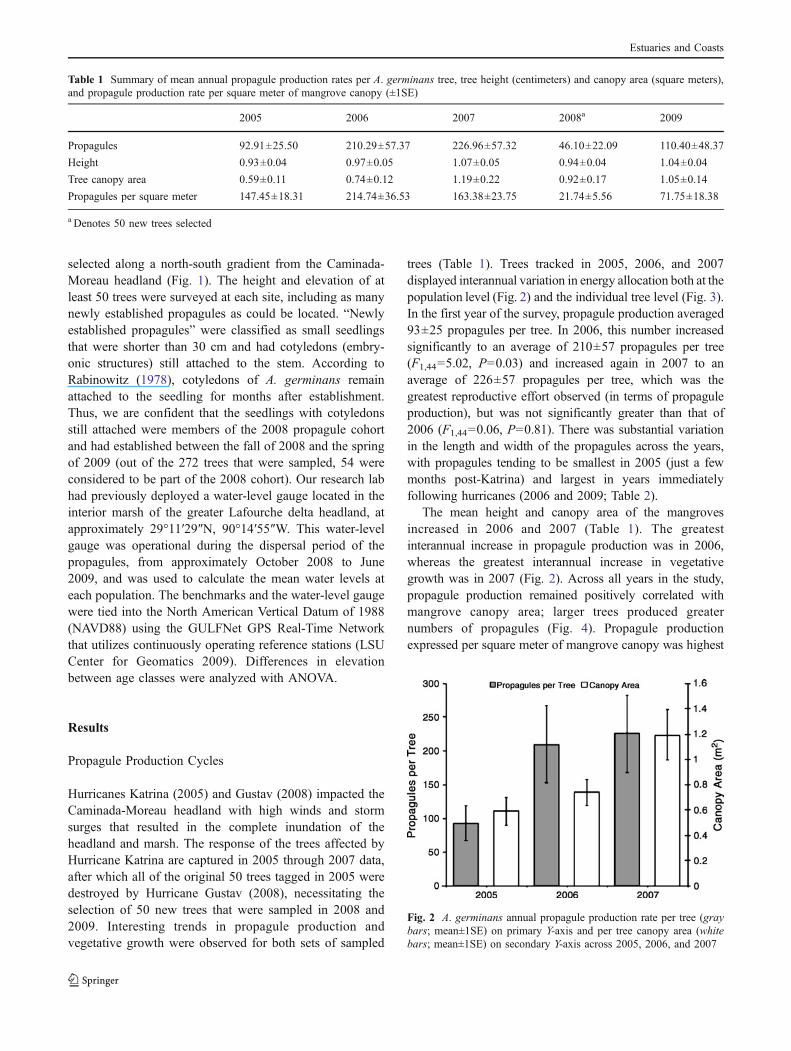
trees (Table 1). Trees tracked in 2005, 2006, and 2007displayed interannual variation in energy allocation both at thepopulation level (Fig. 2) and the individual tree level (Fig. 3).In the first year of the survey, propagule production averaged93±25 propagules per tree. In 2006, this number increasedsignificantly to an average of 210±57 propagules per tree(F1,44=5.02, P=0.03) and increased again in 2007 to anaverage of 226±57 propagules per tree, which was thegreatest reproductive effort observed (in terms of propaguleproduction), but was not significantly greater than that of2006 (F1,44=0.06, P=0.81). There was substantial variationin the length and width of the propagules across the years,with propagules tending to be smallest in 2005 (just a fewmonths post-Katrina) and largest in years immediatelyfollowing hurricanes (2006 and 2009; Table 2).
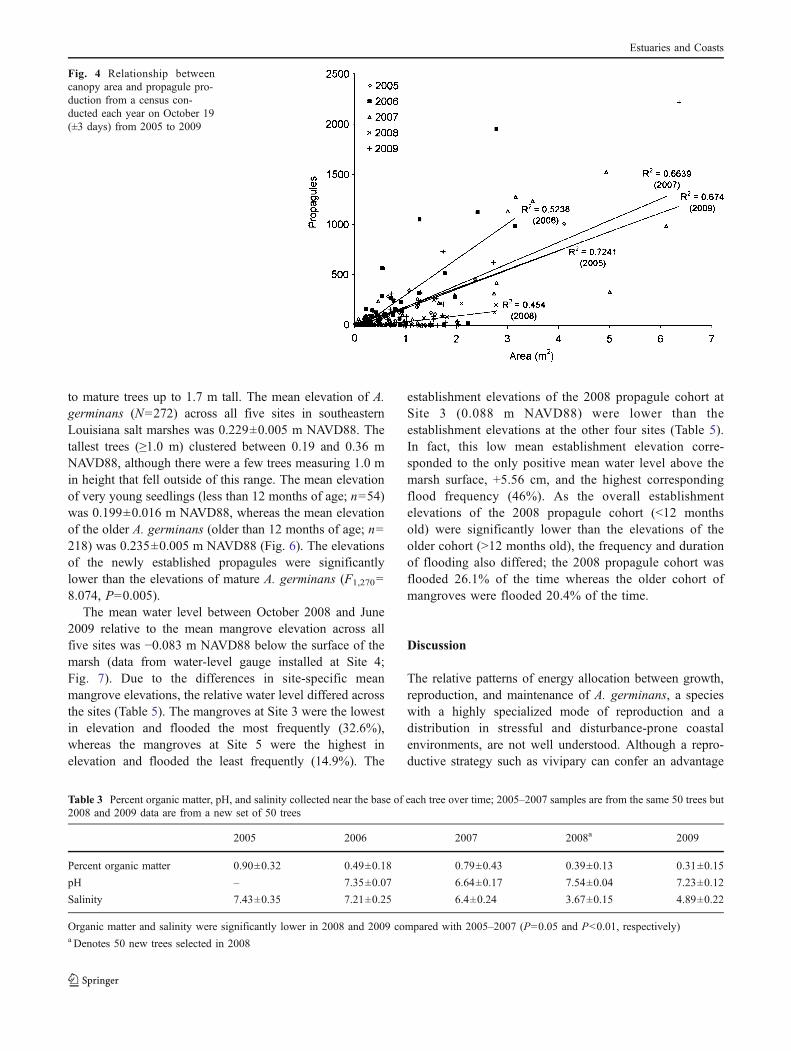
The mean height and canopy area of the mangrovesincreased in 2006 and 2007 (Table 1). The greatestinterannual increase in propagule production was in 2006,whereas the greatest interannual increase in vegetativegrowth was in 2007 (Fig. 2). Across all years in the study,propagule production remained positively correlated withmangrove canopy area; larger trees produced greaternumbers of propagules (Fig. 4). Propagule productionexpressed per square meter of mangrove canopy was highest
Table 1 Summary of mean annual propagule production rates per A. germinans tree, tree height (centimeters) and canopy area (square meters),and propagule production rate per square meter of mangrove canopy (±1SE)
2005 2006 2007 2008a 2009
Propagules 92.91±25.50 210.29±57.37 226.96±57.32 46.10±22.09 110.40±48.37
Height 0.93±0.04 0.97±0.05 1.07±0.05 0.94±0.04 1.04±0.04
Tree canopy area 0.59±0.11 0.74±0.12 1.19±0.22 0.92±0.17 1.05±0.14
Propagules per square meter 147.45±18.31 214.74±36.53 163.38±23.75 21.74±5.56 71.75±18.38
a Denotes 50 new trees selected
Fig. 2 A. germinans annual propagule production rate per tree (graybars; mean±1SE) on primary Y-axis and per tree canopy area (whitebars; mean±1SE) on secondary Y-axis across 2005, 2006, and 2007
Estuaries and Coasts
in 2006 with 214 propagules per square meter, as comparedwith 2005 with 147 propagules per square meter and 2007with 163 propagules per square meter. In 2008 and 2009,propagule production was again correlated with mangrovecanopy area, although the mean propagule production rateper square meter of mangrove canopy was considerablylower than in previous years (22 propagules per square meterin 2008 and 72 propagules per square meter in 2009).
Soil salinity and percent organic matter tended todecrease over time (Table 3). Soil salinity and percentorganic matter were both significantly lower in 2008 and2009 compared with 2005, 2006, and 2007 (F1,206=120.01,P<0.001 and F1,206=3.63, P=0.05, respectively). Thedecrease in soil organic matter coincides with the extensiveoverwash that occurred as a result of Hurricane Gustav in2008 and tropical storm Ida in 2009.
Propagule Buoyancy, Root Development, and Viability
Fungus was observed on the cotyledons of the propagulesaround 60 days into the study. The incidence of fungus was
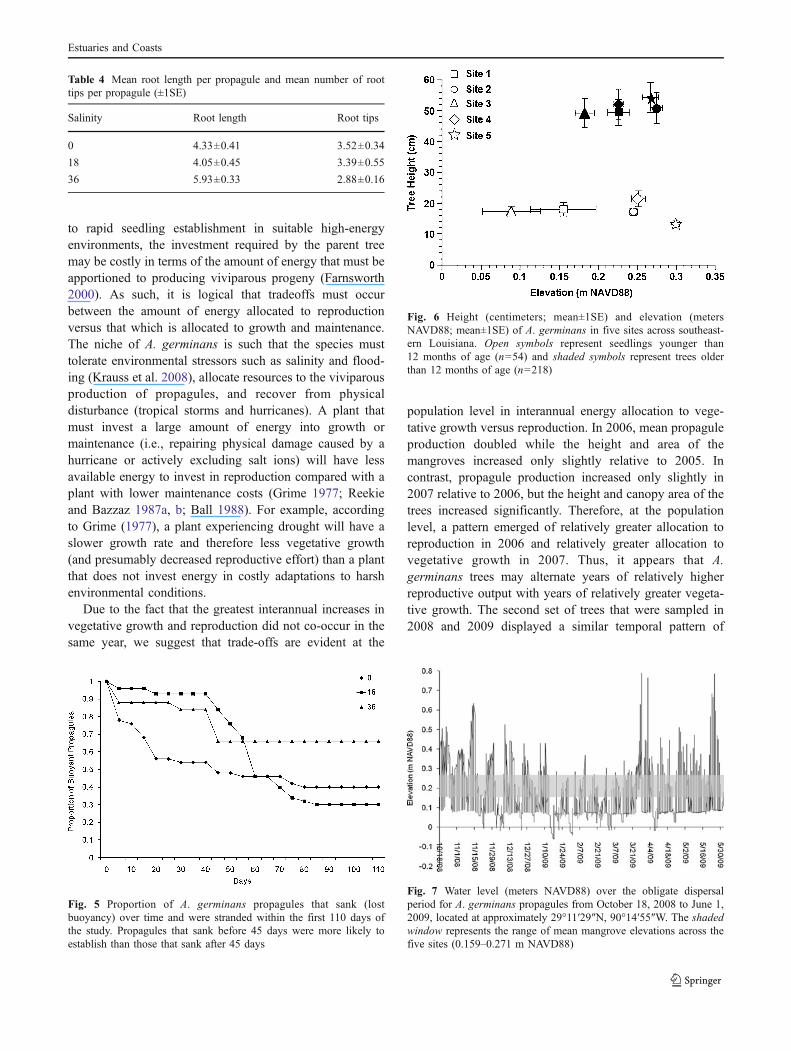
problematic in both of the propagule buoyancy studies, butonly in the reservoirs containing water at 0 or 18.Propagules in reservoirs at 36 did not have contaminationissues. The initial fungal pathogen likely belongs to thegenus Colletotrichum, and the secondary fungal pathogensmay be members of the genera Trichoderma or Aspergillus(pers. comm., Mary Aime, Louisiana State UniversityAgricultural Center). Due to the incidence of fungus andthe potential for this to affect results, data are not analyzed,but trends are presented (Table 4; Fig. 5).
Salinity affected certain aspects of propagule rootmorphology and the ability to lose and regain buoyancy.Propagules in 36 had the greatest number of root tips andshorter individual root lengths (Table 4). However, totalroot length and surface area were not affected by salinity.Ninety-seven percent of the propagules in 36 transitioned atleast once (i.e., lost buoyancy, then regained it), ascompared with 67% in 0 and 53% in 18. Propagules in 36remained buoyant for the longest period of time.
In the second study, 34% of the propagules lost buoyancy inthe first 45 days of the study, and an additional 21% sank in thetime between day 45 and day 110 (Fig. 5). Propagules thatwere stranded within the first 45 days had a greater chance ofsuccessfully establishing than those that sank after 45 days.The incidence of fungal infestation and rotting increased withthe amount of time spent floating in low-salinity water (0 and18 salinity levels), but not at 36. Only 25% of the propagulesthat sank within 110 days successfully established andsurvived to produce true leaves. The mean time fromstranding to establishment was 31 days (±3 days).
Propagule Establishment Elevations
A total of 272 A. germinans were measured, ranging fromnewly established propagules as small as 6.2 cm in height
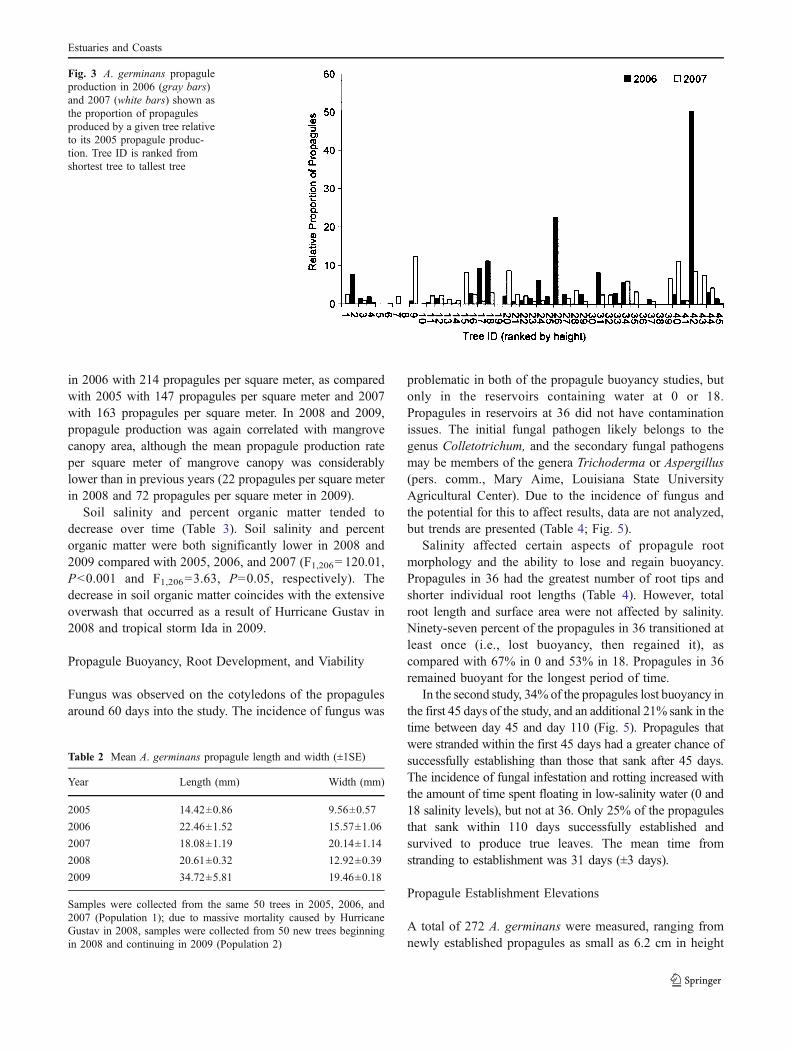
Fig. 3 A. germinans propaguleproduction in 2006 (gray bars)and 2007 (white bars) shown asthe proportion of propagulesproduced by a given tree relativeto its 2005 propagule produc-tion. Tree ID is ranked fromshortest tree to tallest tree
Table 2 Mean A. germinans propagule length and width (±1SE)
Year Length (mm) Width (mm)
2005 14.42±0.86 9.56±0.57
2006 22.46±1.52 15.57±1.06
2007 18.08±1.19 20.14±1.14
2008 20.61±0.32 12.92±0.39
2009 34.72±5.81 19.46±0.18
Samples were collected from the same 50 trees in 2005, 2006, and2007 (Population 1); due to massive mortality caused by HurricaneGustav in 2008, samples were collected from 50 new trees beginningin 2008 and continuing in 2009 (Population 2)
Estuaries and Coasts
to mature trees up to 1.7 m tall. The mean elevation of A.germinans (N=272) across all five sites in southeasternLouisiana salt marshes was 0.229±0.005 m NAVD88. Thetallest trees (≥1.0 m) clustered between 0.19 and 0.36 mNAVD88, although there were a few trees measuring 1.0 min height that fell outside of this range. The mean elevationof very young seedlings (less than 12 months of age; n=54)was 0.199±0.016 m NAVD88, whereas the mean elevationof the older A. germinans (older than 12 months of age; n=218) was 0.235±0.005 m NAVD88 (Fig. 6). The elevationsof the newly established propagules were significantlylower than the elevations of mature A. germinans (F1,270=8.074, P=0.005).
The mean water level between October 2008 and June2009 relative to the mean mangrove elevation across allfive sites was −0.083 m NAVD88 below the surface of themarsh (data from water-level gauge installed at Site 4;Fig. 7). Due to the differences in site-specific meanmangrove elevations, the relative water level differed acrossthe sites (Table 5). The mangroves at Site 3 were the lowestin elevation and flooded the most frequently (32.6%),whereas the mangroves at Site 5 were the highest inelevation and flooded the least frequently (14.9%). The
establishment elevations of the 2008 propagule cohort atSite 3 (0.088 m NAVD88) were lower than theestablishment elevations at the other four sites (Table 5).In fact, this low mean establishment elevation corre-sponded to the only positive mean water level above themarsh surface, +5.56 cm, and the highest correspondingflood frequency (46%). As the overall establishmentelevations of the 2008 propagule cohort (<12 monthsold) were significantly lower than the elevations of theolder cohort (>12 months old), the frequency and durationof flooding also differed; the 2008 propagule cohort wasflooded 26.1% of the time whereas the older cohort ofmangroves were flooded 20.4% of the time.
Discussion
The relative patterns of energy allocation between growth,reproduction, and maintenance of A. germinans, a specieswith a highly specialized mode of reproduction and adistribution in stressful and disturbance-prone coastalenvironments, are not well understood. Although a repro-ductive strategy such as vivipary can confer an advantage
Fig. 4 Relationship betweencanopy area and propagule pro-duction from a census con-ducted each year on October 19(±3 days) from 2005 to 2009
Table 3 Percent organic matter, pH, and salinity collected near the base of each tree over time; 2005–2007 samples are from the same 50 trees but2008 and 2009 data are from a new set of 50 trees
2005 2006 2007 2008a 2009
Percent organic matter 0.90±0.32 0.49±0.18 0.79±0.43 0.39±0.13 0.31±0.15
pH – 7.35±0.07 6.64±0.17 7.54±0.04 7.23±0.12
Salinity 7.43±0.35 7.21±0.25 6.4±0.24 3.67±0.15 4.89±0.22
Organic matter and salinity were significantly lower in 2008 and 2009 compared with 2005–2007 (P=0.05 and P<0.01, respectively)a Denotes 50 new trees selected in 2008
Estuaries and Coasts
to rapid seedling establishment in suitable high-energyenvironments, the investment required by the parent treemay be costly in terms of the amount of energy that must beapportioned to producing viviparous progeny (Farnsworth2000). As such, it is logical that tradeoffs must occurbetween the amount of energy allocated to reproductionversus that which is allocated to growth and maintenance.The niche of A. germinans is such that the species musttolerate environmental stressors such as salinity and flood-ing (Krauss et al. 2008), allocate resources to the viviparousproduction of propagules, and recover from physicaldisturbance (tropical storms and hurricanes). A plant thatmust invest a large amount of energy into growth ormaintenance (i.e., repairing physical damage caused by ahurricane or actively excluding salt ions) will have lessavailable energy to invest in reproduction compared with aplant with lower maintenance costs (Grime 1977; Reekieand Bazzaz 1987a, b; Ball 1988). For example, accordingto Grime (1977), a plant experiencing drought will have aslower growth rate and therefore less vegetative growth(and presumably decreased reproductive effort) than a plantthat does not invest energy in costly adaptations to harshenvironmental conditions.
Due to the fact that the greatest interannual increases invegetative growth and reproduction did not co-occur in thesame year, we suggest that trade-offs are evident at the
population level in interannual energy allocation to vege-tative growth versus reproduction. In 2006, mean propaguleproduction doubled while the height and area of themangroves increased only slightly relative to 2005. Incontrast, propagule production increased only slightly in2007 relative to 2006, but the height and canopy area of thetrees increased significantly. Therefore, at the populationlevel, a pattern emerged of relatively greater allocation toreproduction in 2006 and relatively greater allocation tovegetative growth in 2007. Thus, it appears that A.germinans trees may alternate years of relatively higherreproductive output with years of relatively greater vegeta-tive growth. The second set of trees that were sampled in2008 and 2009 displayed a similar temporal pattern of
Table 4 Mean root length per propagule and mean number of roottips per propagule (±1SE)
Salinity Root length Root tips
0 4.33±0.41 3.52±0.34
18 4.05±0.45 3.39±0.55
36 5.93±0.33 2.88±0.16
Fig. 5 Proportion of A. germinans propagules that sank (lostbuoyancy) over time and were stranded within the first 110 days ofthe study. Propagules that sank before 45 days were more likely toestablish than those that sank after 45 days
Fig. 6 Height (centimeters; mean±1SE) and elevation (metersNAVD88; mean±1SE) of A. germinans in five sites across southeast-ern Louisiana. Open symbols represent seedlings younger than12 months of age (n=54) and shaded symbols represent trees olderthan 12 months of age (n=218)
Fig. 7 Water level (meters NAVD88) over the obligate dispersalperiod for A. germinans propagules from October 18, 2008 to June 1,2009, located at approximately 29°11′29″N, 90°14′55″W. The shadedwindow represents the range of mean mangrove elevations across thefive sites (0.159–0.271 m NAVD88)
Estuaries and Coasts
interannual variation with a marked increase in propaguleproduction in 2009 relative to 2008 but a smaller relativeincrease in height and canopy area during this sameinterval.
In addition to population-level trade-offs, individualtrees varied the amount of resources they invested inreproduction in consecutive years. Individual trees rarelyproduced the same number of propagules in consecutiveyears; in most cases, a tree’s fecundity increased ordecreased dramatically relative to the previous year. Basedon our observations, A. germinans has a propensity to varypropagule production intensity, which may be furthermodulated by the amount of damage sustained duringmajor disturbances (i.e., tropical storms and hurricanes,prolonged cold temperatures, etc.). Interannual variations inpropagule size may be due to slight variations in the timingof peak propagule maturity.
Because of these trade-offs between growth and repro-duction, decreased reproductive effort driven by greatercompensatory growth in a year following a hurricanedisturbance may therefore translate to lower seedlingrecruitment. Compensatory growth is an important processfollowing defoliation in some species and results in fewer,smaller reproductive structures (Anten et al. 2003). Thiswas the case in Florida in 2004 after Hurricane Charleymade landfall: significant reductions in Rhizophora manglepropagule production and seedling recruitment were ob-served compared with control sites that were not impactedby the hurricane (Proffit et al. 2006). Although thepropagules may be destroyed or dispersed out of an areaimpacted by a hurricane, the role of canopy gaps inregeneration may favor the competitive release of somespecies of mangroves (Ellison and Farnsworth 1993). Forexample, R. mangle is shade-intolerant and showedsignificantly greater growth in canopy gaps than in theunderstory (Ellison and Farnsworth 1993). In a study bySmith (1987), seedlings from the genera Avicennia,Rhizophora, and Bruguiera grew best in areas with greater
light and fewer conspecifics. Lopez-Hoffman et al. (2007)also reported that Avicennia seedlings benefitted fromincreasing light levels in canopy openings. In this study,we did not observe an increase in recruitment of A.germinans seedlings in canopy gaps, per se, in the yearsfollowing the hurricanes of 2005 and 2008. This may bepartially explained by the relatively smaller stature ofLouisiana A. germinans and its mosaic pattern of distribu-tion, which currently does not result in extensive areas ofclosed canopies.
The development ofA. germinans propagules in Louisianais similar to that of A. marina in southeastern Australia(Clarke 1993); both species have obligate dispersal periodsof less than 2 weeks and can remain viable for months afterthey abscise from the parent tree. In addition, both species ofAvicennia lose and regain buoyancy throughout theirdispersal period (Clarke et al. 2001), particularly in full-strength sea water. This suggests internal physiologicalchanges in the cotyledons and root tissue of the developingpropagule with time (Clarke et al. 2001), although thespecific mechanisms are not known.
Decay was a significant problem in these studies. Thedecay and fungal infestation was the most problematic inthe brackish reservoirs (18); although it was also present infresh water reservoirs (0), it was not as destructive.Propagules in full-strength seawater reservoirs (36) didnot have decay or fungal infestation and were well-preserved in this salinity throughout the course of a 110-day dispersal period. The issue of decay appears to beproblematic in certain circumstances (e.g., mesocosm studyby Rabinowitz 1978; field studies by Patterson et al. 1997,and Delgado et al. 2001) but not in others (i.e., mesocosmstudy by Toledo et al. 2001; field study by McKee 1995).The incidence of fungal infestation increased with thelength of the dispersal period, particularly in 0 and 18. Full-strength seawater (36) may preserve viability better thanother salinity levels and represent the greatest dispersalpotential when combined with the ability to regain buoy-
Table 5 Mean elevations (meters NAVD88; ±1SE) of A. germinans(>12 months old) and mean establishment elevations (±1SE) of the2008 propagule cohort (<12 months old) at Sites 1–5 with the
corresponding mean water level (centimeters) and frequency offlooding (percent) based on a water-level gauge deployed at Site 4from October 31, 2008 to June 1, 2009
Site Mangrove elevation>12 months old,m NAVD88
Water level relative tomangroves >12 monthsold, cm
Frequencyflooded,%
Mangrove elevation<12 months old,m NAVD88
Water level relative tomangroves <12 monthsold, cm
Frequencyflooded,%
1 0.226±0.137 −8.14 21.90% 0.155±0.042 −1.04 33.5%
2 0.275±0.008 −13.04 14.40% 0.245±0.005 −10.04 18.5%
3 0.183±0.011 −3.84 28.80% 0.088±0.037 5.66 46.2%
4 0.226±0.006 −8.14 21.90% 0.251±0.008 −10.64 17.80%
5 0.267±0.011 −12.24 15.50% 0.299±0.005 −15.44 11.60%
Mean 0.235±0.005 −9.04 20.40% 0.199±0.016 −5.44 26.10%
Estuaries and Coasts

ancy (Sousa et al. 2007). Our study differs from previousstudies (namely Rabinowitz 1978) because we confirm theviability of A. germinans propagules after a long dispersalperiod of 110 days.
McKee (1995) and Sousa et al. (2007) observed thatpropagules seldom maximize their full dispersal potentialby rarely dispersing more than several meters or in veryrare cases, greater than 50 km, from their parent trees(Clarke 1993). The proximity of a propagule to an intertidalcreek or bay may account for greater dispersal distance,especially in conjunction with flood tides and/or high-energy storms (Clarke 1993). Therefore, long-distancedispersal is indeed physiologically possible but may berelatively rare due to decreasing viability with increasingdispersal period length, physical barriers, and the direction-al flow of tides and currents.
Interestingly, in the field, propagules not only established atsimilar elevations as adult trees but also at lower positions inthe intertidal where physical energy and frequency of floodingare two factors that are known to potentially limit recruitment(Sousa et al. 2007). Strong north winds typical of Louisianawinters can cause periods of low water of up to 79 cm belowmean sea level (Hester and Mendelssohn 1989) and may haveexisted several times throughout the 2008–2009 obligatedispersal period and acted as low water “establishmentwindows.” For example, during an 18-day period fromJanuary 13, 2009 to February 1, 2009, mean water levelwas 0.048 m NAVD88, which is approximately −0.181 mbelow the surface of the marsh (0.229 m NAVD88). Meanwater level during this period was lower than mean waterlevel throughout the rest of the season, thereby exposingsubstrate previously unavailable for establishment that year.Concurrent with low mean water level, the absence ofincoming tidal energy may have permitted propagules toanchor roots in low-elevation areas during this window.This data supports estimates that have shown that A.germinans propagules can establish if the substrate is notflooded 70% of the time (McMillan 1971; Lewis andEstevez 1988; Patterson et al. 1997). Rabinowitz (1978)reported that A. germinans propagules can firmly anchorthemselves into the substrate within weeks; similarly, wereport establishment times of less than 3 weeks.
There is evidence that once a propagule establishes, theseedling may be able to persist at this elevation even if it islower than usual because the physical energy and not thephysico-chemical environment of low-elevation sites iswhat limits Avicennia’s recruitment (Patterson et al. 1997;Sousa et al. 2007). However, the higher mean elevation ofthe older trees (0.239 m NAVD88) may suggest thatpropagules below this elevation have established outsidetheir “optimal” range and may not survive to sexualmaturity, or may become physiologically stressed oretiolated over time (Ye et al. 2004; Krauss et al. 2006).
In summary, hydrology plays a key role in the reproductivebiology, establishment patterns, and zonation of A. germinansin Louisiana. Winter climatic events such as strong northerlywinds may create “establishment windows” by loweringwater levels in the marsh, thereby exposing substrate atlower elevations that become temporally available forstranding and establishment. However, due to the overalllow rate of propagule establishment in the mesocosmstudies, we hypothesize that A. germinans produces manymore propagules than actually establish and survive toadulthood. Furthermore, the success of truly long-distancedispersal events seems low given the severely decreasedviability of propagules after dispersal periods exceeding45 days. In light of the dynamic interactions of climatechange, hurricane disturbance, and regional subsidencealong the coastline of Louisiana, the information presentedherein on A. germinans is anticipated to be valuable tocoastal managers in the restoration of salt marshes andbarrier islands of northern coast of the Gulf of Mexico.
Acknowledgments We would like to extend our gratitude to supportprovided by NOAA/CREST (National Oceanic and AtmosphericAdministration/Coastal Restoration and Enhancement Through Sci-ence and Technology) and CSAP (Coastal Science AssistantshipProgram) provided by LA OCPR (Louisiana Office of CoastalProtection and Restoration) and Louisiana Sea Grant. We would alsolike to thank the members of the Coastal Plant Ecology Lab whohelped with the collection of data in the field: Christine Pickens,Michael Dupuis, and Jonathan Willis. Additional thanks to TherynHenkel, Joe Baustian, and Sean Graham for providing data andsupport for this project.
References
Allen, J.A., K.W. Krauss, and R.D. Hauff. 2003. Factors limiting theintertidal distribution of the mangrove species Xylocarpusgranatum. Oecologia 135: 110–121.
Anten, N.P.R., M. Martinez-Ramos, and D.D. Ackerly. 2003. Defoliationand growth in an understory palm: quantifying the contributions ofcompensatory responses. Ecology 84(11): 2905–2918.
Ball, M.C. 1988. Ecophysiology of mangroves. Trees 2: 129–142.Boyd, R., and S. Penland. 1988. Geomorphologic model for
Mississippi-Delta evaluation AAPG Bulletin. American Associa-tion of Petroleum Geologists 72(9): 1110.
Clarke, P.J. 1993. Dispersal of gray mangrove (Avicennia marina)propagules in southeastern Australia. Aquatic Botany 45(2–3):195–204.
Clarke, P.J., and W.G. Allaway. 1993. The regeneration niche of thegray mangrove (Avicennia marina)—effects of salinity, light, andsediment factors on establishment, growth, and survival in thefield. Oecologia 93(4): 548–556.
Clarke, P.J., R.A. Kerrigan, and C.J. Westphal. 2001. Dispersalpotential and early growth in 14 tropical mangroves: do earlylife history traits correlate with patterns of adult distribution?Journal of Ecology 89(4): 648–659.
Delgado, P., P.F. Hensel, J.A. Jimenez, and J.W. Day. 2001. Theimportance of propagule establishment and physical factors inmangrove distributional patterns in a Costa Rican estuary.Aquatic Botany 71: 157–178.
Estuaries and Coasts

Ellison, A.M., and E.J. Farnsworth. 1993. Seedling survivorship,growth, and response to disturbance in Belizean mangal.American Journal of Botany 80(10): 1137–1145.
Farnsworth, E.J. 2000. The ecology and physiology of viviparous andrecalcitrant seeds. Annual Review of Ecology, Evolution, andSystematics 31: 107–138.
Grime, J.P. 1977. Evidence for the existence of three primarystrategies in plants and its relevance to ecological andevolutionary theory. The American Naturalist 111(982): 1169–1194.
Hester, M.W., and I.A. Mendelssohn. 1989. Water relations andgrowth responses of Uniola paniculata (sea oats) to soil moistureand water-table depth. Oecologia 78: 289–296.
Hester, M.W., and I.A. Mendelssohn. 1991. Expansion patterns andsoil physicochemical characterization of three Louisiana popula-tions of Uniola paniculata (Sea oats). Journal of CoastalResearch 7(2): 387–401.
Hester, M.W., E.A. Spalding, and C.D. Franze. 2005. Biologicalresources of the Louisiana coast: part 1. An overview of coastalplant communities of the Louisiana gulf shoreline. Journal ofCoastal Research 44: 134–145.
Kitaya, Y., V. Jintana, S. Piriyayotha, D. Jaijing, K. Yabunki, S.Izutani, A. Nishimiya, and M. Iwasaki. 2002. Early growth ofseven mangrove species planted at different elevations in a Thaiestuary. Trees 16: 150–154.
Krauss, K.W., T.W. Doyle, R.R. Twilley, V.H. Rivera-Monroy, and J.K. Sullivan. 2006. Evaluating the relative contributions ofhydroperiod and soil salinity on growth of south Floridamangroves. Hydrobiologia 569: 311–324.
Krauss, K.W., C.E. Lovelock, K.L. McKee, L. Lopez-Hoffman, S.M.L. Ewe, and W.P. Sousa. 2008. Environmental drivers inmangrove establishment and early development: a review.Aquatic Botany 89: 105–127.
Lewin, R. 1986. Supply-side ecology. Science 234: 25–27.Lewis III, R.R. 2005. Ecological engineering for successful management
and restoration of mangrove forests. Ecological Engineering 24:403–418.
Lewis, R.R. III and E.D. Estevez. 1988. The ecology of TampaBay, Florida: An estuarine profile. National Wetlands ResearchCenter, US Fish and Wildlife Service. Biological Report No85. 132 pp.
Lopez-Hoffman, L., D.D. Ackerly, N.P.R. Anten, J.L. Denoyer, andM. Martinez-Ramos. 2007. Gap-dependence in mangrove life-history strategies: a consideration of the entire life cycle andpatch dynamics. Journal of Ecology 95: 1222–1233.
McBride, R.A., and M.R. Byrnes. 1997. Regional variations in shoreresponse along barrier island systems of the Mississippi Riverdelta plain: historical change and future prediction. Journal ofCoastal Research 13(3): 628–655.
McKee, K.L. 1995. Seedling recruitment patterns in a Belizeanmangrove forest: effects of establishment ability and physico-chemical factors. Oecologia 101: 448–460.
McMillan, C. 1971. Environmental factors affecting seedling estab-lishment of the black mangrove on the central Texas coast.Ecology 52(5): 927–930.
Nettel, A., and R.S. Dodd. 2007. Drifting propagules and recedingswamps: genetic footprints of mangrove recolonization anddispersal along tropical coasts. Evolution 61(4): 958–971.
Patterson, C.S., K.L. McKee, and I.A. Mendelssohn. 1997. Effects oftidal inundation and predation on Avicennia germinans seedlingestablishment and survival in a sub-tropical mangal/salt marshcommunity. Mangroves and Salt Marshes 1: 103–111.
Pickens, C.N., and M.W. Hester. 2010. Temperature tolerance of earlylife history stages of black mangrove Avicennia germinans:implications for range expansion. Estuaries and Coasts.doi:10.1007/sl12237-010-9358-2.
Proffit, C.E., E.C. Milbrandt, and S.E. Travis. 2006. Red mangrove(Rhizophora mangle) reproduction and seedling colonizationafter Hurricane Charley: comparisons of Charlotte Harbor andTampa Bay. Estuaries and Coasts 29(6): 972–978.
Rabinowitz, D. 1978. Dispersal properties of mangrove propagules.Biotropica 10(1): 47–57.
Reekie, E.G., and F.A. Bazzaz. 1987a. Reproductive effort in plants.1. Carbon allocation to reproduction. The American Naturalist129(6): 876–896.
Reekie, E.G., and F.A. Bazzaz. 1987b. Reproductive effort in plants.2. Does carbon reflect the allocation of other resources? TheAmerican Naturalist 129(6): 897–906.
Saenger, P. 2002. Mangrove ecology, silviculture, and conservation.Dordrecht: Kluwer.
Sampson, M.S., and R.N. Rollon. 2008. Growth performance ofplanted mangroves in the Philippines: revisiting forest manage-ment strategies. Royal Swedish Academy of Science 37(4): 234–240.
Smith III, T.J. 1987. Effects of light and intertidal position on seedlingsurvival and growth in tropical tidal forests. Journal ofExperimental Marine Biology and Ecology 110: 133–146.
Sousa, W.P., P.G. Kennedy, B.J. Mitchell, and B.M. Ordonez. 2007.Supply-side ecology in mangroves: do propagule dispersal andseedling establishment explain forest structure? EcologicalMonographs 77(1): 53–76.
Toledo, G., A. Rojas, and Y. Bashan. 2001. Monitoring of blackmangrove restoration with nursery-reared seedlings on an aridcoastal lagoon. Hydrobiologia 444: 101–109.
Tomlinson, P.B. 1986. The Botany of Mangroves. Cambridge: Cam-bridge University Press.
Visser, J.M., W.G. Vermilion, D.E. Evers, R.G. Linscombe, and C.E.Sasser. 2005. Nesting habitat requirements for brown pelican andtheir management implications. Journal of Coastal Research 21(2): e27–e35.
Ye, Y., N.F.Y. Tam, Y.S. Wong, and C.Y. Lu. 2004. Does sea level riseinfluence propagule establishment, early growth and physiologyof Kandelia candel and Bruguiera gymnorrhiza? Journal ofExperimental Marine Biology and Ecology 306: 197–215.
Estuaries and Coasts
Related Documents











