1966 American Journal of Botany 88(11): 1966–1976. 2001. REPRODUCTIVE BIOLOGY OF HENEQUE ´ N (AGAVE FOURCROYDES) AND ITS WILD ANCESTOR AGAVE ANGUSTIFOLIA (AGAVACEAE). I.GAMETOPHYTE DEVELOPMENT 1 NICKOLAI M. PIVEN, 2 FELIPE A. BARREDO-POOL, ILEANA C. BORGES-ARGA ´ EZ,MIGUEL A. HERRERA-ALAMILLO, ALBERTO MAYO-MOSQUEDA,JOSE ´ L. HERRERA-HERRERA, AND MANUEL L. ROBERT Centro de Investigacio ´n Cientı ´fica de Yucata ´n, Calle 43, No. 130, Col. Chuburna ´ de Hidalgo, CP 97200, Me ´rida, Yucata ´n, Me ´xico The pathways of micro- and megagametophyte development in Agave fourcroydes (heneque ´n) and A. angustifolia were studied. We used histology and light microscopy to observe anther ontogeny and ovary differentiation in relation to flower bud size. Both species have the same sexual reproductive strategies and gametophyte development that may be divided into three phases: (1) premeiotic, which includes the establishment of the megaspore mother cell and the pollen mother cell; (2) meiotic, the formation of mature microspores and functional megaspores; (3) postmeiotic, which encompasses the development of mature pollen grains and the formation of the embryo sac. A successive type microsporogenesis was found in both species with formation of T-shaped tetrads and binuclear pollen grains. In vitro germination tests revealed very low pollen fertility. The female gametophyte is formed from two micropylar megaspore cells after the first meiotic division (bisporic type). Male and female gametogenesis occur asynchronously with microspo- rogenesis finishing before macrosporogenesis. The results so far show that the formation of male and female gametophytes in heneque ´n is affected at different stages and that these alterations might be responsible for the low fertility shown by this species. Key words: Agavaceae; anther; embryo sac; heneque ´n; megasporogenesis; microsporogenesis; ovule; pollen. Agaves are monocotyledonous plants, many of which have their center of origin in Mexico, where some species have been domesticated and are of economic importance as sources of fiber, steroids, spirits, and other products (Gentry, 1982; Dahl- gren, Clifford, and Yeo, 1985; Robert et al., 1992; Colunga- Garcı ´a Marı ´n and May-Pat, 1993). The only cultivated species in the Yucatan Peninsula is heneque ´n (Agave fourcroydes Lem.) that descends from the wild ancestor Agave angustifolia Haw. (Colunga-Garcı ´a Marı ´n and May-Pat, 1997; Colunga- Garcı ´a Marı ´n et al., 1999). Both of these agaves are mono- carpic perennials that produce flowers only once toward the end of their life cycle of ;20 yr, after which they die. Throughout its life span, heneque ´n propagates mainly by means of its rhizomes whose apical meristems emerge at a distance from the parent plant, giving rise to new individuals. The flowers develop at the top of large inflorescences that can reach 3–8 m high and, after flowering, bulbils originate from buds beneath bracteoles on the inflorescence. However, in spite of their potential utility, bulbils and seeds are not usually used for commercial propagation and have not been used for breed- ing processes (Eastmond, Herrera, and Robert, 2000). The tra- ditional agricultural practice of cutting the heneque ´n inflores- cences soon after they begin to develop, in order to preserve the plant a little longer, has also limited the supply of seeds. 1 Manuscript received 7 November 2000; revision accepted 3 April 2001. The authors wish to thank two anonymous referees and Dr. Amarella East- mond for their thorough reviews of the manuscript and relevant comments, and Dr. Roger Orellana for allowing us to collect plant material from CICY’s Botanical Garden. This research was partially supported by funds provided by SISIERRA-CONACYT (Mexico), grant 980608. 2 Author for correspondence and reprint requests (e-mail: [email protected]). Fruits develop abnormally because carpels remain empty, probably due to insufficient pollination, and the few seeds that mature show low germination less than 10%. Both phenomena could be due to the pentaploid chromosome level of the spe- cies (5x 5 150; Castorena-Sa ´nchez, Escobedo, and Quiroz, 1991), which may be responsible for the low fertility of the species. Conversely, in the wild species A. angustifolia, which is a fertile hexaploid (6x 5 180; Castorena-Sa ´nchez, Escobe- do, and Quiroz, 1991), seed germination is as high as 73% (Colunga-Garcı ´a Marı ´n et al., 1999). This led us to include it in this study for comparative purposes. In spite of its economic importance, conventional breeding techniques have never been applied to heneque ´n, probably because its long life span and low fertility have made it almost impossible to carry out ge- netic improvement. No attempts have been made to cross hen- eque ´n with other agave species and no natural hybrids have been reported. The only successful program to hybridize hard fiber-producing agaves was started by Doughty in Tanzania in 1931 (Doughty, 1936) and culminated in the release of hybrid 11648 (A. angustifolia 3 A. amaniensis) 3 A. amaniensis (Lock, 1962). In order to develop alternative methods for the improvement of heneque ´n, Robert et al. (1987, 1992) used in vitro culture to micropropagate high-yielding individuals and to form a germplasm collection. The micropropagated plants that origi- nated in vitro by shoot multiplication are genetically stable and are performing satisfactorily in the field. However, the genetic homogeneity in the vegetatively propagated plantations is still a problem, and the generation of new genetic variability would be highly desirable in order to prevent susceptibility to micro- bial diseases and other threats to the plantations. Sexual re- production offers the best alternative to generate the variability needed for improvement programs. Nevertheless, until reliable

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1966
American Journal of Botany 88(11): 1966–1976. 2001.
REPRODUCTIVE BIOLOGY OF HENEQUEN
(AGAVE FOURCROYDES) AND ITS WILD ANCESTOR
AGAVE ANGUSTIFOLIA (AGAVACEAE).I. GAMETOPHYTE DEVELOPMENT1
NICKOLAI M. PIVEN,2 FELIPE A. BARREDO-POOL,ILEANA C. BORGES-ARGAEZ, MIGUEL A. HERRERA-ALAMILLO,ALBERTO MAYO-MOSQUEDA, JOSE L. HERRERA-HERRERA, AND
MANUEL L. ROBERT
Centro de Investigacion Cientıfica de Yucatan, Calle 43, No. 130, Col. Chuburna de Hidalgo, CP 97200, Merida, Yucatan, Mexico
The pathways of micro- and megagametophyte development in Agave fourcroydes (henequen) and A. angustifolia were studied. Weused histology and light microscopy to observe anther ontogeny and ovary differentiation in relation to flower bud size. Both specieshave the same sexual reproductive strategies and gametophyte development that may be divided into three phases: (1) premeiotic,which includes the establishment of the megaspore mother cell and the pollen mother cell; (2) meiotic, the formation of maturemicrospores and functional megaspores; (3) postmeiotic, which encompasses the development of mature pollen grains and the formationof the embryo sac. A successive type microsporogenesis was found in both species with formation of T-shaped tetrads and binuclearpollen grains. In vitro germination tests revealed very low pollen fertility. The female gametophyte is formed from two micropylarmegaspore cells after the first meiotic division (bisporic type). Male and female gametogenesis occur asynchronously with microspo-rogenesis finishing before macrosporogenesis. The results so far show that the formation of male and female gametophytes in henequenis affected at different stages and that these alterations might be responsible for the low fertility shown by this species.
Key words: Agavaceae; anther; embryo sac; henequen; megasporogenesis; microsporogenesis; ovule; pollen.
Agaves are monocotyledonous plants, many of which havetheir center of origin in Mexico, where some species have beendomesticated and are of economic importance as sources offiber, steroids, spirits, and other products (Gentry, 1982; Dahl-gren, Clifford, and Yeo, 1985; Robert et al., 1992; Colunga-Garcıa Marın and May-Pat, 1993). The only cultivated speciesin the Yucatan Peninsula is henequen (Agave fourcroydesLem.) that descends from the wild ancestor Agave angustifoliaHaw. (Colunga-Garcıa Marın and May-Pat, 1997; Colunga-Garcıa Marın et al., 1999). Both of these agaves are mono-carpic perennials that produce flowers only once toward theend of their life cycle of ;20 yr, after which they die.Throughout its life span, henequen propagates mainly bymeans of its rhizomes whose apical meristems emerge at adistance from the parent plant, giving rise to new individuals.The flowers develop at the top of large inflorescences that canreach 3–8 m high and, after flowering, bulbils originate frombuds beneath bracteoles on the inflorescence. However, in spiteof their potential utility, bulbils and seeds are not usually usedfor commercial propagation and have not been used for breed-ing processes (Eastmond, Herrera, and Robert, 2000). The tra-ditional agricultural practice of cutting the henequen inflores-cences soon after they begin to develop, in order to preservethe plant a little longer, has also limited the supply of seeds.
1 Manuscript received 7 November 2000; revision accepted 3 April 2001.The authors wish to thank two anonymous referees and Dr. Amarella East-
mond for their thorough reviews of the manuscript and relevant comments,and Dr. Roger Orellana for allowing us to collect plant material from CICY’sBotanical Garden. This research was partially supported by funds providedby SISIERRA-CONACYT (Mexico), grant 980608.
2 Author for correspondence and reprint requests (e-mail: [email protected]).
Fruits develop abnormally because carpels remain empty,probably due to insufficient pollination, and the few seeds thatmature show low germination less than 10%. Both phenomenacould be due to the pentaploid chromosome level of the spe-cies (5x 5 150; Castorena-Sanchez, Escobedo, and Quiroz,1991), which may be responsible for the low fertility of thespecies. Conversely, in the wild species A. angustifolia, whichis a fertile hexaploid (6x 5 180; Castorena-Sanchez, Escobe-do, and Quiroz, 1991), seed germination is as high as 73%(Colunga-Garcıa Marın et al., 1999). This led us to include itin this study for comparative purposes. In spite of its economicimportance, conventional breeding techniques have never beenapplied to henequen, probably because its long life span andlow fertility have made it almost impossible to carry out ge-netic improvement. No attempts have been made to cross hen-equen with other agave species and no natural hybrids havebeen reported. The only successful program to hybridize hardfiber-producing agaves was started by Doughty in Tanzania in1931 (Doughty, 1936) and culminated in the release of hybrid11648 (A. angustifolia 3 A. amaniensis) 3 A. amaniensis(Lock, 1962).
In order to develop alternative methods for the improvementof henequen, Robert et al. (1987, 1992) used in vitro cultureto micropropagate high-yielding individuals and to form agermplasm collection. The micropropagated plants that origi-nated in vitro by shoot multiplication are genetically stable andare performing satisfactorily in the field. However, the genetichomogeneity in the vegetatively propagated plantations is stilla problem, and the generation of new genetic variability wouldbe highly desirable in order to prevent susceptibility to micro-bial diseases and other threats to the plantations. Sexual re-production offers the best alternative to generate the variabilityneeded for improvement programs. Nevertheless, until reliable

November 2001] 1967PIVEN ET AL.—REPRODUCTIVE BIOLOGY OF HENEQUEN
Figs. 1–4. Inflorescences in Agave. 1. Scape development of A. fourcroydes is arrested after the leaves are cut. 2. Scapes cut by growers in commercialplantations of henequen. 3. Flowering inflorescence of A. angustifolia. 4. Flowering inflorescence of A. fourcroydes.
knowledge of the sexual reproductive process of agaves ingeneral and of henequen in particular is available, this willremain a difficult task.
This is the first report of morphological and histologicalstudies of the sexual reproductive system of A. fourcroydes
and its wild relative A. angustifolia. We analyzed male ga-metophyte (pollen or microgametophyte) and female gameto-phyte (embryo sac or megagametophyte) development in bothspecies to understand how gametes are formed and what caus-es the differences in fertility.
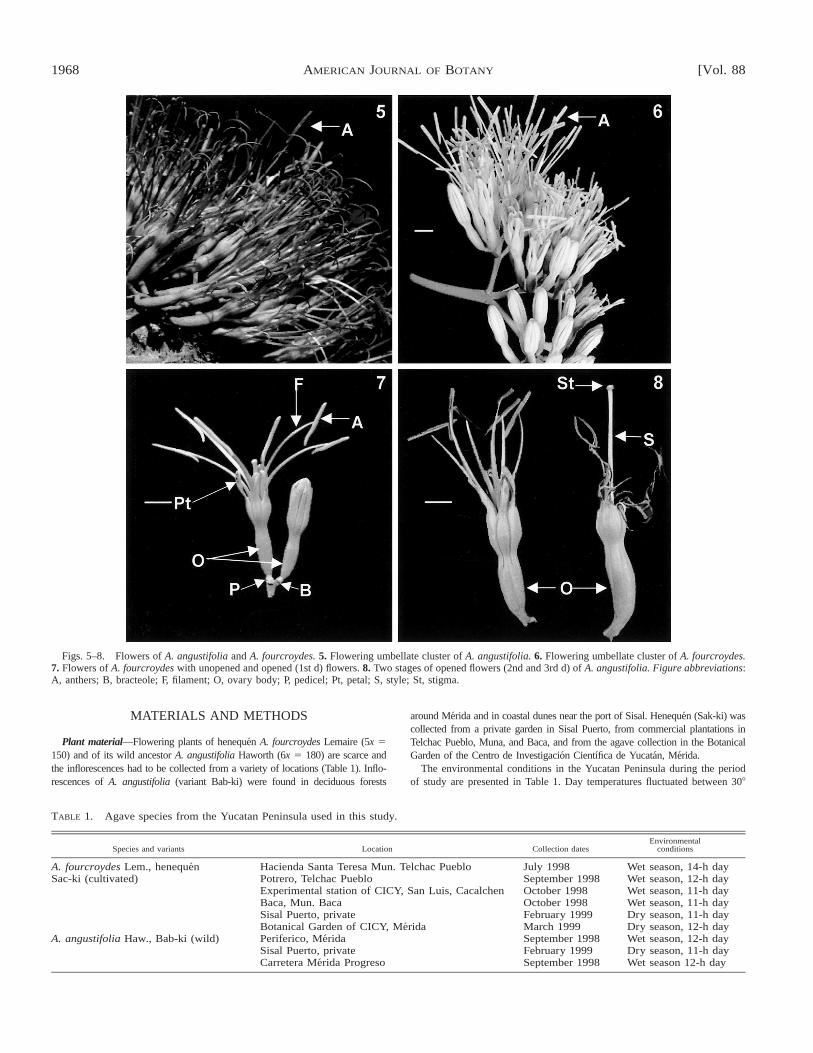
1968 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 5–8. Flowers of A. angustifolia and A. fourcroydes. 5. Flowering umbellate cluster of A. angustifolia. 6. Flowering umbellate cluster of A. fourcroydes.7. Flowers of A. fourcroydes with unopened and opened (1st d) flowers. 8. Two stages of opened flowers (2nd and 3rd d) of A. angustifolia. Figure abbreviations:A, anthers; B, bracteole; F, filament; O, ovary body; P, pedicel; Pt, petal; S, style; St, stigma.
TABLE 1. Agave species from the Yucatan Peninsula used in this study.
Species and variants Location Collection datesEnvironmental
conditions
A. fourcroydes Lem., henequen Hacienda Santa Teresa Mun. Telchac Pueblo July 1998 Wet season, 14-h daySac-ki (cultivated) Potrero, Telchac Pueblo September 1998 Wet season, 12-h day
Experimental station of CICY, San Luis, Cacalchen October 1998 Wet season, 11-h dayBaca, Mun. Baca October 1998 Wet season, 11-h daySisal Puerto, private February 1999 Dry season, 11-h dayBotanical Garden of CICY, Merida March 1999 Dry season, 12-h day
A. angustifolia Haw., Bab-ki (wild) Periferico, Merida September 1998 Wet season, 12-h daySisal Puerto, private February 1999 Dry season, 11-h dayCarretera Merida Progreso September 1998 Wet season 12-h day
MATERIALS AND METHODS
Plant material—Flowering plants of henequen A. fourcroydes Lemaire (5x 5150) and of its wild ancestor A. angustifolia Haworth (6x 5 180) are scarce andthe inflorescences had to be collected from a variety of locations (Table 1). Inflo-rescences of A. angustifolia (variant Bab-ki) were found in deciduous forests
around Merida and in coastal dunes near the port of Sisal. Henequen (Sak-ki) wascollected from a private garden in Sisal Puerto, from commercial plantations inTelchac Pueblo, Muna, and Baca, and from the agave collection in the BotanicalGarden of the Centro de Investigacion Cientıfica de Yucatan, Merida.
The environmental conditions in the Yucatan Peninsula during the periodof study are presented in Table 1. Day temperatures fluctuated between 308
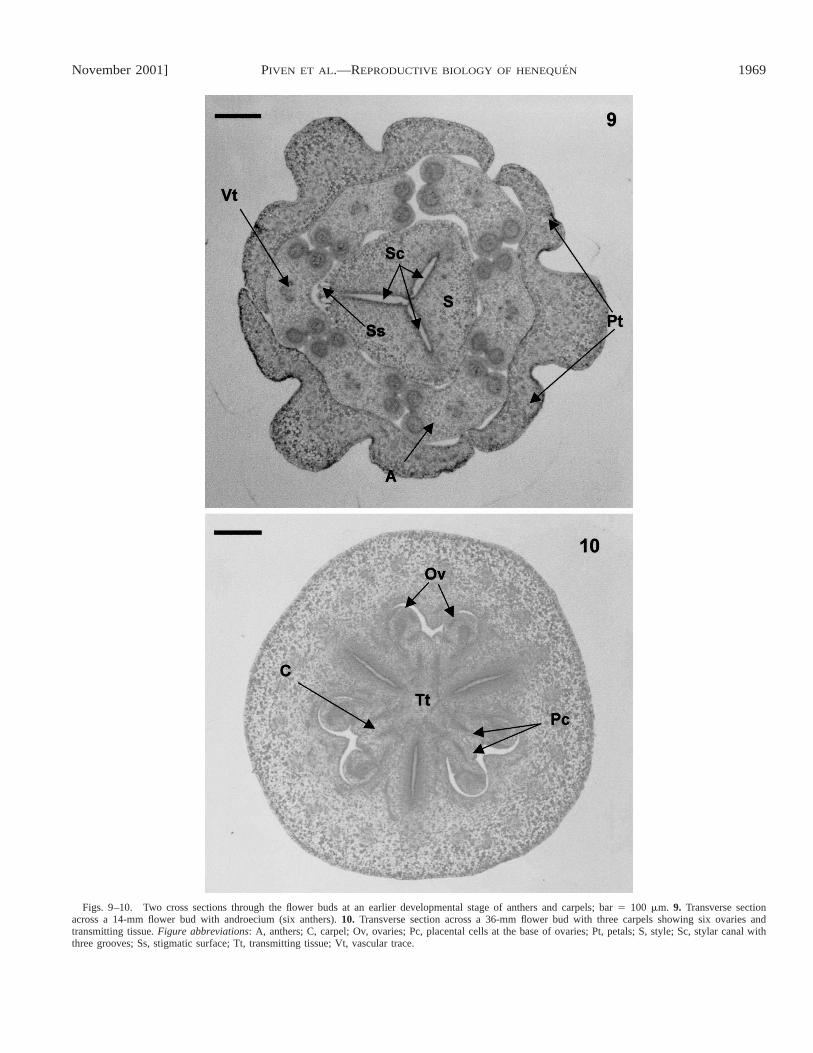
November 2001] 1969PIVEN ET AL.—REPRODUCTIVE BIOLOGY OF HENEQUEN
Figs. 9–10. Two cross sections through the flower buds at an earlier developmental stage of anthers and carpels; bar 5 100 mm. 9. Transverse sectionacross a 14-mm flower bud with androecium (six anthers). 10. Transverse section across a 36-mm flower bud with three carpels showing six ovaries andtransmitting tissue. Figure abbreviations: A, anthers; C, carpel; Ov, ovaries; Pc, placental cells at the base of ovaries; Pt, petals; S, style; Sc, stylar canal withthree grooves; Ss, stigmatic surface; Tt, transmitting tissue; Vt, vascular trace.
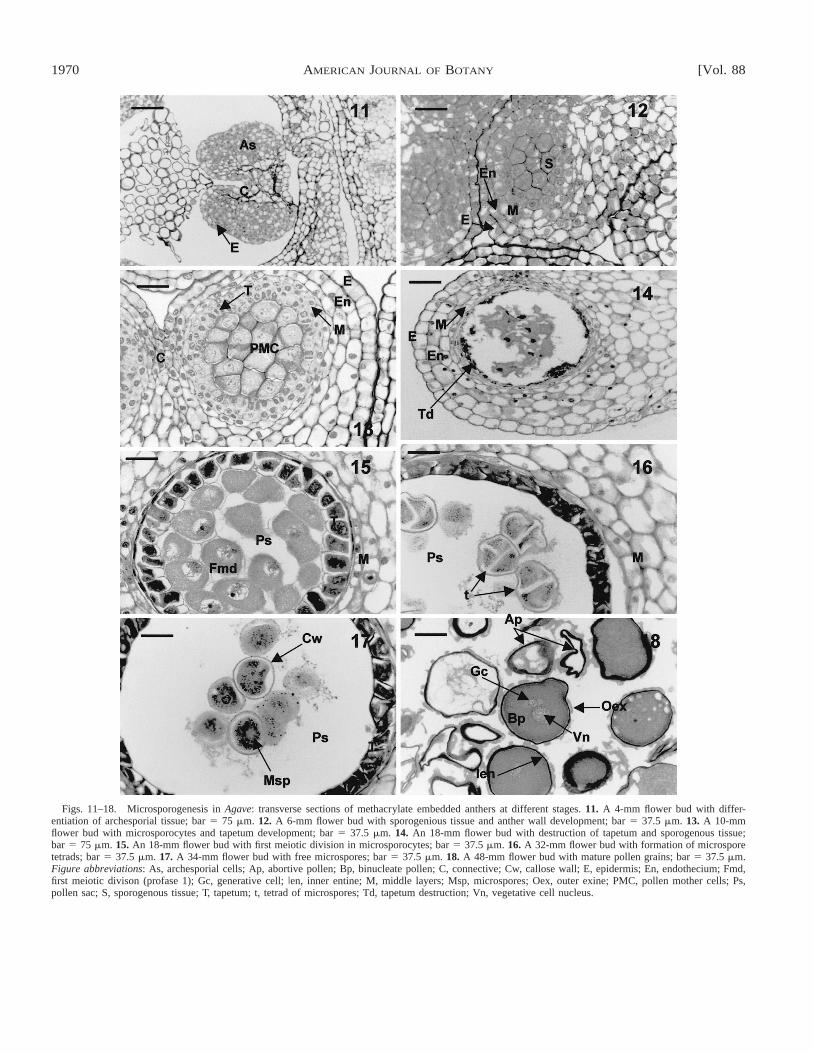
1970 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 11–18. Microsporogenesis in Agave: transverse sections of methacrylate embedded anthers at different stages. 11. A 4-mm flower bud with differ-entiation of archesporial tissue; bar 5 75 mm. 12. A 6-mm flower bud with sporogenious tissue and anther wall development; bar 5 37.5 mm. 13. A 10-mmflower bud with microsporocytes and tapetum development; bar 5 37.5 mm. 14. An 18-mm flower bud with destruction of tapetum and sporogenous tissue;bar 5 75 mm. 15. An 18-mm flower bud with first meiotic division in microsporocytes; bar 5 37.5 mm. 16. A 32-mm flower bud with formation of microsporetetrads; bar 5 37.5 mm. 17. A 34-mm flower bud with free microspores; bar 5 37.5 mm. 18. A 48-mm flower bud with mature pollen grains; bar 5 37.5 mm.Figure abbreviations: As, archesporial cells; Ap, abortive pollen; Bp, binucleate pollen; C, connective; Cw, callose wall; E, epidermis; En, endothecium; Fmd,first meiotic divison (profase 1); Gc, generative cell; Ien, inner entine; M, middle layers; Msp, microspores; Oex, outer exine; PMC, pollen mother cells; Ps,pollen sac; S, sporogenous tissue; T, tapetum; t, tetrad of microspores; Td, tapetum destruction; Vn, vegetative cell nucleus.
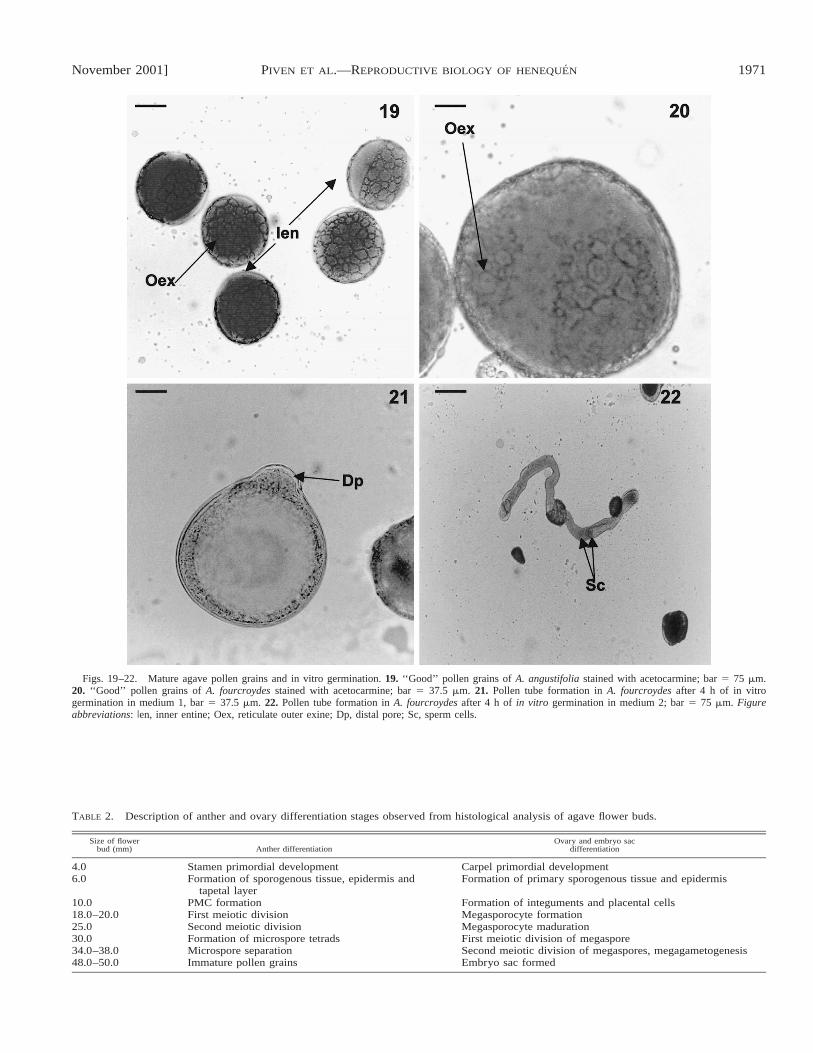
November 2001] 1971PIVEN ET AL.—REPRODUCTIVE BIOLOGY OF HENEQUEN
Figs. 19–22. Mature agave pollen grains and in vitro germination. 19. ‘‘Good’’ pollen grains of A. angustifolia stained with acetocarmine; bar 5 75 mm.20. ‘‘Good’’ pollen grains of A. fourcroydes stained with acetocarmine; bar 5 37.5 mm. 21. Pollen tube formation in A. fourcroydes after 4 h of in vitrogermination in medium 1, bar 5 37.5 mm. 22. Pollen tube formation in A. fourcroydes after 4 h of in vitro germination in medium 2; bar 5 75 mm. Figureabbreviations: Ien, inner entine; Oex, reticulate outer exine; Dp, distal pore; Sc, sperm cells.
TABLE 2. Description of anther and ovary differentiation stages observed from histological analysis of agave flower buds.
Size of flowerbud (mm) Anther differentiation
Ovary and embryo sacdifferentiation
4.0 Stamen primordial development Carpel primordial development6.0 Formation of sporogenous tissue, epidermis and
tapetal layerFormation of primary sporogenous tissue and epidermis
10.0 PMC formation Formation of integuments and placental cells18.0–20.0 First meiotic division Megasporocyte formation25.0 Second meiotic division Megasporocyte maduration30.0 Formation of microspore tetrads First meiotic division of megaspore34.0–38.0 Microspore separation Second meiotic division of megaspores, megagametogenesis48.0–50.0 Immature pollen grains Embryo sac formed
1972 [Vol. 88AMERICAN JOURNAL OF BOTANY
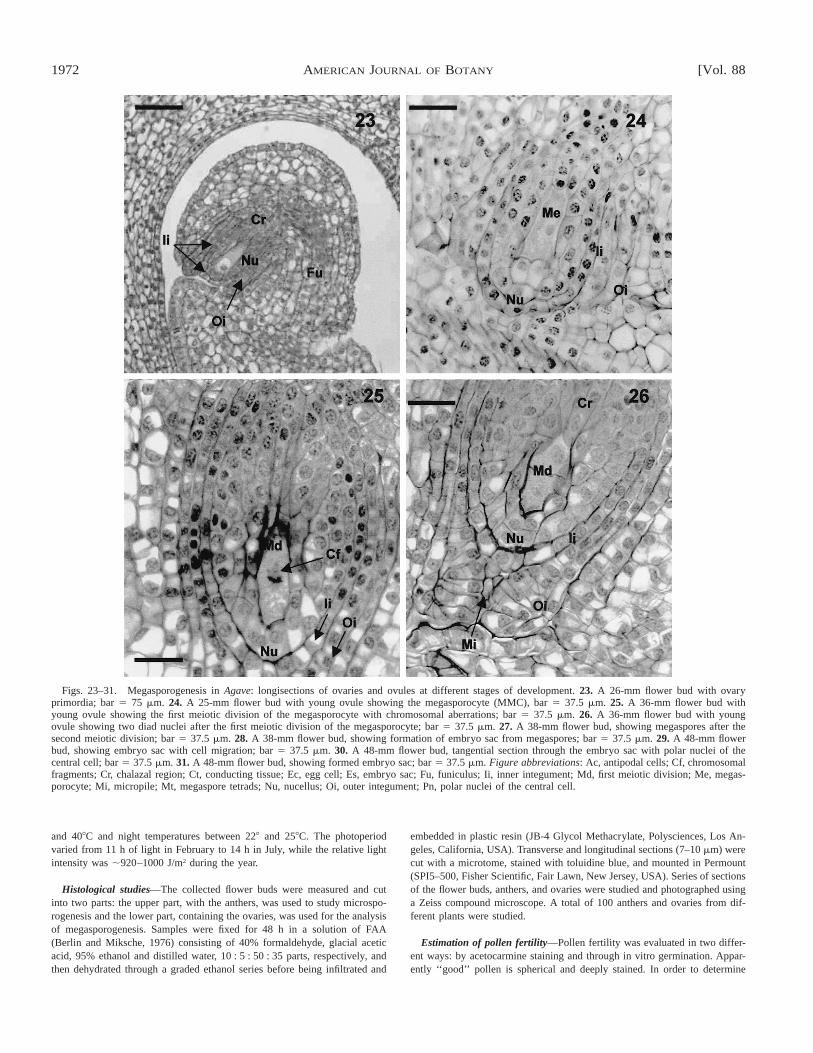
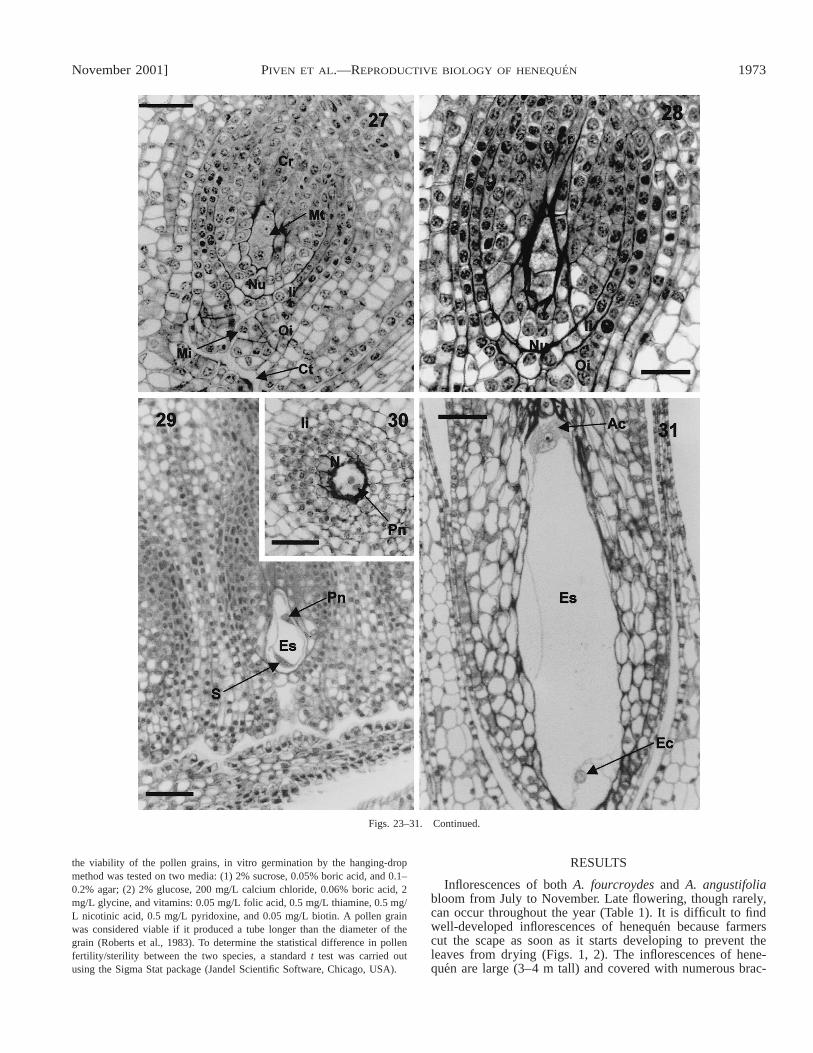
Figs. 23–31. Megasporogenesis in Agave: longisections of ovaries and ovules at different stages of development. 23. A 26-mm flower bud with ovaryprimordia; bar 5 75 mm. 24. A 25-mm flower bud with young ovule showing the megasporocyte (MMC), bar 5 37.5 mm. 25. A 36-mm flower bud withyoung ovule showing the first meiotic division of the megasporocyte with chromosomal aberrations; bar 5 37.5 mm. 26. A 36-mm flower bud with youngovule showing two diad nuclei after the first meiotic division of the megasporocyte; bar 5 37.5 mm. 27. A 38-mm flower bud, showing megaspores after thesecond meiotic division; bar 5 37.5 mm. 28. A 38-mm flower bud, showing formation of embryo sac from megaspores; bar 5 37.5 mm. 29. A 48-mm flowerbud, showing embryo sac with cell migration; bar 5 37.5 mm. 30. A 48-mm flower bud, tangential section through the embryo sac with polar nuclei of thecentral cell; bar 5 37.5 mm. 31. A 48-mm flower bud, showing formed embryo sac; bar 5 37.5 mm. Figure abbreviations: Ac, antipodal cells; Cf, chromosomalfragments; Cr, chalazal region; Ct, conducting tissue; Ec, egg cell; Es, embryo sac; Fu, funiculus; Ii, inner integument; Md, first meiotic division; Me, megas-porocyte; Mi, micropile; Mt, megaspore tetrads; Nu, nucellus; Oi, outer integument; Pn, polar nuclei of the central cell.
and 408C and night temperatures between 228 and 258C. The photoperiodvaried from 11 h of light in February to 14 h in July, while the relative lightintensity was ;920–1000 J/m2 during the year.
Histological studies—The collected flower buds were measured and cutinto two parts: the upper part, with the anthers, was used to study microspo-rogenesis and the lower part, containing the ovaries, was used for the analysisof megasporogenesis. Samples were fixed for 48 h in a solution of FAA(Berlin and Miksche, 1976) consisting of 40% formaldehyde, glacial aceticacid, 95% ethanol and distilled water, 10 : 5 : 50 : 35 parts, respectively, andthen dehydrated through a graded ethanol series before being infiltrated and
embedded in plastic resin (JB-4 Glycol Methacrylate, Polysciences, Los An-geles, California, USA). Transverse and longitudinal sections (7–10 mm) werecut with a microtome, stained with toluidine blue, and mounted in Permount(SPI5–500, Fisher Scientific, Fair Lawn, New Jersey, USA). Series of sectionsof the flower buds, anthers, and ovaries were studied and photographed usinga Zeiss compound microscope. A total of 100 anthers and ovaries from dif-ferent plants were studied.
Estimation of pollen fertility—Pollen fertility was evaluated in two differ-ent ways: by acetocarmine staining and through in vitro germination. Appar-ently ‘‘good’’ pollen is spherical and deeply stained. In order to determine
November 2001] 1973PIVEN ET AL.—REPRODUCTIVE BIOLOGY OF HENEQUEN
Figs. 23–31. Continued.
the viability of the pollen grains, in vitro germination by the hanging-dropmethod was tested on two media: (1) 2% sucrose, 0.05% boric acid, and 0.1–0.2% agar; (2) 2% glucose, 200 mg/L calcium chloride, 0.06% boric acid, 2mg/L glycine, and vitamins: 0.05 mg/L folic acid, 0.5 mg/L thiamine, 0.5 mg/L nicotinic acid, 0.5 mg/L pyridoxine, and 0.05 mg/L biotin. A pollen grainwas considered viable if it produced a tube longer than the diameter of thegrain (Roberts et al., 1983). To determine the statistical difference in pollenfertility/sterility between the two species, a standard t test was carried outusing the Sigma Stat package (Jandel Scientific Software, Chicago, USA).
RESULTS
Inflorescences of both A. fourcroydes and A. angustifoliabloom from July to November. Late flowering, though rarely,can occur throughout the year (Table 1). It is difficult to findwell-developed inflorescences of henequen because farmerscut the scape as soon as it starts developing to prevent theleaves from drying (Figs. 1, 2). The inflorescences of hene-quen are large (3–4 m tall) and covered with numerous brac-

1974 [Vol. 88AMERICAN JOURNAL OF BOTANY
teal leaves. Flowers from in umbellate clusters (i.e., lateralracemes), starting from the lowest branches. Flowering con-tinues for several weeks from the base upwards, so that afterthe first few days the inflorescence presents flowers in all stag-es of development (Figs. 3, 4). Flowers are bisexual (her-maphrodite) with both pistil and stamens in the same flower.The morphology of the flowers is very similar in both species,but some clones of A. angustifolia have darker anthers than A.fourcroydes (Figs. 5, 6). The flowers are epigyneous, with pet-als, stamens, and anthers located above the ovary (Figs. 7, 8).The six narrow stamens are united at the base of the tube ofthe corolla. Their basal parts are fused with the pistil wall andform part of the ovary, a characteristic that is common to allagave species. The flowers are protandrous; the anthers (theandroecium) are supported by long filaments and protrude 3–4 cm from the corolla (Figs. 7, 8). Flower development of thisspecies is initiated by periclinal cell divisions in the protoderm(a meristem covered by a bract) where flower primordia (spe-cific organ meristem layers) are formed. Six primordial sta-mens are initiated in the third whorl after the sepal and petalprimordia, but prior to the initiation of carpel formation. Atthis stage the development of the gametophyte is initiated.
Developmental stages of anthers and male gametophyticcells—Androecia of A. angustifolia and A. fourcroydes consistof six anthers, and are arcuate, tetrasporangiate, and dithecalwith laterally positioned microsporangia that open longitudi-nally (Fig. 9). The stages of microsporangial development areshown in Table 2. The male gametophytic cycle begins withthe formation of the microsporangium in the stamen and con-sists of archesporial cells (Fig. 11). This tissue forms a primaryparietal and sporogenous layer of cells as well as secondaryparietal layers (Fig. 12), with specific middle wall layers: thetapetum and the endothecium. The tapetum, whose cells formfrom the endothecium, is glandular (secretory). The microspo-rocytes or pollen mother cells (PMC) form via mitotic celldivision of sporogenous tissue and can be observed, compactlyarranged and connected through plasmodesmata, at the centerof the microsporangium (Fig. 13). During the early meioticprophase, the original plasmodesmata disintegrate and thePMC become spherical (Fig. 15). The tapetum in both agavesis best developed during microspore formation (Figs. 15–17),but, as can be seen in Fig. 14, in some anthers of A. four-croydes the tapetum and the generative tissue are completelydestroyed before microsporogenesis is initiated and thereforemicrospores and pollen grains do not develop.
Microsporogenesis in these agaves takes place in two suc-cessive meiotic divisions forming T-shaped microspore tetrads(Figs. 15, 16). The uninucleate microspores grow after theyare released from the tetrads (Fig. 17) and divide asymmetri-cally resulting in two types of cells (vegetative and genera-tive). After meiosis, the tapetum begins to break down and thepollen grains are released. The mature pollen wall is composedof two layers: an outer one of exine and an inner one of intine(Fig. 18).
Pollen morphology and fertility—The morphology of thepollen of A. fourcroydes is very similar to that of A. angus-tifolia. The pollen grains are binuclear with vegetative andgenerative nuclei, monosulcate with two distal pores, and havea reticulate exine (Figs. 19, 20). A few differences betweenthe two species can be observed in the pollen ornamentationand apertures (Ludlow-Wiechers and Ojeda, 1983; Ojeda,
Ludlow, and Orellana, 1984). The proportion of spherical, ma-ture pollen grains (‘‘good pollen’’) differs between the twospecies: ;48.9 6 1.5% for A. angustifolia and ;33.6 60.08% for A. fourcroydes (Figs. 19, 20). This difference isstatistically significant (t 5 8.98 with 4.00 degrees of freedomat P # 0.005, 95% confidence interval). The sterility of thepollen grains was also different: a very high frequency (66.46 0.8%) of abnormal pollen grains was observed in henequen(Fig. 18) and 51.1 6 1.5% for A. angustifolia. The differencebetween means is statistically significant (t 5 6.65 with 2.00degrees of freedom at P # 0.022, 95% confidence interval).The percentage of germinated pollens that produced only shortpollen tubes was very low (0.5 6 0.08%). Occasionally, itwas observed that in media 2, adjusted with vitamins, therewas a slight increase in germination (1%) and in the length ofthe pollen tubes (Figs. 21, 22). On the other hand, pollen vi-ability in A. angustifolia was ;3 6 0.8% but the differencewas not statistically significant. During germination the gen-erative cells formed two sperm cells after another mitotic di-vision (Fig. 22).
Ovule and embryo sac formation—The gynoecium of A.fourcroydes and A. angustifolia consists of three united carpelsand a trilocular ovary with axillary placentation (Fig. 10).Each locule has many anatropous ovules. The stages of theovary differentiation process that are inferred from histologicalanalyses are presented in Table 2. The ovules are specializedstructures, derived from the placenta of the ovary, that producethe megasporocyte (Fig. 23). The latter is known as the mega-spore mother cell (MMC) and is the site of the embryo sacformation, fertilization, and embryogenesis. The placenta isconfined to the central axis of the pistil. The ovules are con-nected to the ovary wall that consists of three basic structures:a nucellus or megasporangium, two integuments (inner andouter), and a funiculus (Figs. 23, 24). The MMC, which formsfrom the archesporial tissue, is located below the apex of thenucellus (Fig. 24) while the integuments form at its base dur-ing megasporogenesis (Figs. 23, 24). The inner integument islocated at the nucellus, whereas the outer integument is locatedabove the inner integument and consists of two cell layers(Figs. 25, 26). The micropyle pore is found at the top of theouter integument (Fig. 26). The funiculus is fused with oneside of the ovule and is visible as a rib (chalazal region) andlocated at the opposite end of the micropyle (Fig. 26).
During megasporogenesis the MMC undergoes a meioticdivision parallel to the micropylar-chalasal axis of the nucel-lus. At this stage, we observed some structures that could in-dicate that chromosomal fragmentations occur (Fig. 25). Wallformation is perpendicular to the micropylar-chalasal axis,forming a first dyad of megaspores (Fig. 26). Only one mega-spore undergoes a second meiotic division creating two func-tional megaspores (bisporic type of megasporogenesis, Fig.27). Thus the female gametophyte in henequen develops fromthe two viable micropile megaspores directly into an embryosac (Fig. 28). However, their formation is not regular for anormal eight-nucleate megagametophyte. Positioned at thecenter of the embryo sac, the central cell contains two polarnuclei (Figs. 29, 30) that partially fuse before fertilization,generating the primary endosperm. The egg cell is located inthe micropylar part of the embryo sac, while the synergids,which could be located on either side of the egg cell region,are not present in the mature embryo sac (Figs. 29, 31). Theantipodal cells are placed at the chalazal region (Fig. 31).

November 2001] 1975PIVEN ET AL.—REPRODUCTIVE BIOLOGY OF HENEQUEN
DISCUSSION
Henequen (Agave fourcroydes Lem.) is an economically im-portant fiber crop for the state of Yucatan, Mexico (Cruz, Or-ellana, and Robert, 1985; Colunga-Garcıa Marın and May-Pat,1993). It is, however, a relatively infertile pentaploid specieswith a very long life span, which is vegetatively propagatedand has never been genetically improved. We studied the re-productive biology of this species in order to understand thebasis of its sexual reproductive patterns and to predict the dif-ficulties involved in a crossbreeding program. We decided toinclude its putative wild ancestor A. angustifolia in this studybecause, being phylogenetically related to the former, it couldprovide clues to its reproductive characteristics. A limitationof the present study was posed by the difficulty of findingenough flowering plants, particularly of henequen, and someof the variations observed could be due to the different envi-ronmental conditions under which the plants were growing.
Both species have the same vegetative and sexual repro-ductive strategies and are remarkably similar at the morpho-logical and biochemical levels (Colunga-Garcıa Marın andMay-Pat, 1997; Colunga-Garcıa Marın et al., 1999). However,A. angustifolia is a hexaploid species that produces a highpercentage of fertile seeds. It is interesting to note that thereis very little information regarding the development of thereproductive system of the Agavaceae (Wunderlich, 1950;Gentry, 1982), and these studies are limited to some aspectsof pollination (Arizaga et al., 2000; Slauson, 2000), nectarproduction, and seed viability (Schaffer and Schaffer, 1977,1979; Freeman and Reid, 1985), while little is known aboutother aspects such as gametogenesis. In this paper we discussthe stages of anther and ovary development in henequen re-lated to flower growth, compare them to those of A. angusti-folia, and discuss the possible causes of the low fertility of theformer.
Principal stages of gametophyte development—Flower de-velopment and maturation begin at the lowermost buds in theinflorescence and proceed upwards. The principal stages ofgametophyte development are described in Table 2 and showthe correlation of anther ontogeny and ovary differentiationwith flower bud length. Both processes may be divided intothree phases. Phase I (premeiosis) encompasses the early pro-liferative stages, the differentiation of the locules as well asthe establishment of the PMC, and the formation of MMC.Phase II (meiotic stage) extends from the onset of the PMCand MMC meiosis to the appearance of the microspores andfunctional megaspores. Phase III (postmeiosis) begins at mi-crospore mitosis and extends to the maturation of pollen grainsof the male gametophyte and the formation of the embryo sac.
The results in Table 2 show that some important events ofmale and female gametogenesis are asynchronous. Microspo-rogenesis starts before megasporogenesis and, as a result, pol-len maturity and anther dehiscence occur before the stigmabecomes receptive. This pattern (protandria) is similar for bothA. fourcroydes and A. angustifolia and has been described forother species such as A. sisalana (Lock, 1962) and A. lechug-illa (Freeman and Reid, 1985). This asynchrony of gameto-phyte development might be a common pattern for the genusAgave. Self-pollination is therefore unlikely within the sameflower, but mature pollen could fall on receptive flowers de-veloping on lower parts of the scape.
Male gametophyte development—Microsporogenesis in A.angustifolia and A. fourcroydes with the ‘‘successive type’’ ofmeiosis was recognized because cell walls formed after thefirst cell division (Fig. 16). Although most stages of the pro-cess of male gamete formation in both agaves are apparentlynormal, pollen grains showed a very low in vitro germinationrate: 3% for A. angustifolia and 0.5% for A. fourcroydes. Thiscould reflect some meiotic and mitotic aberrations during mi-crosporogenesis, due to their polyploid nature. Alternatively,it could be due to the influence of tapetal malfunction (Fig.14), since it is widely assumed that the tapetum plays an im-portant role during microsporogenesis and the maturation ofthe pollen grains (Chapman, 1987).
Although in vitro germination cannot reproduce exact invivo conditions (Roberts et al., 1983), we believe it can be auseful tool to relate pollen quality (viability) to normal malegametophyte development, as previously discussed by Janssenand Hermsen (1976).
Female gametophyte: ovule and embryo sac development—According to Dahlgren, Clifford, and Yeo (1985), the femalegametophyte of the Agavoideae is generated from the onlyfunctional megaspore via megagametogenesis (Polygonumtype). However, in A. fourcroydes and A. angustifolia, wefound a bisporic type of megasporogenesis (similar to Allium)but not the Polygonum type. The female gametophyte in hen-equen develops directly into an embryo sac from the two vi-able megaspores. These megaspores divide mitotically twiceresulting in the migration of the nuclei and in the maturationof the embryo sac. Failures in meiosis might have resulted inthe formation of abortive megaspores with a distorted chro-mosome balance that lead to the degradation of the nuclei andthe occurrence of numerous empty embryo sacs (Fig. 25). Itis apparent from the anatomy of the carpels and ovaries thatstructural maturation of the embryo sacs is not regular in someparts of the locules, and this could be the reason for the lowfertility of the female gametophyte.
The results so far show that the formation of male and fe-male gametophyte in henequen is abnormal at the meiotic andpostmeiotic stages and that these alterations might be respon-sible for the low fertility shown by this species. These resultsshould be useful for future henequen breeding research usingpollination, fertilization, and zygotic embryo rescue.
LITERATURE CITED
ARIZAGA, S., E. EZCURRA, E. PETERS, F. RAMıREZ DE ARELLANO, AND E.VEGA. 2000. Pollination biology of Agave macrocantha (Agavaceae) ina Mexican tropical desert. I. Floral biology and pollination mechanisms.American Journal of Botany 87: 1004–1010.
BERLIN, G. P., AND J. P. MIKSCHE. 1976. Botanical microtechnique and cy-tochemistry. 3rd ed. Iowa State University Press, Ames, Iowa, USA.
CASTORENA-SANCHEZ, I., R. M. ESCOBEDO, AND A. QUIROZ. 1991. Newcytotaxonomical determinants recognized in six taxa of Agave in thesections Rigidae and Sisalanae. Canadian Journal of Botany 69: 1257–1264.
CHAPMAN, G. P. 1987. The tapetum. In K. L. Giles and J. Pracash [eds.],Pollen: cytology and development. International Review of Cytology 107:111–125.
COLUNGA-GARCIA MARIN, P., J. COELLO-COELLO, L. E. EGUIARTE, AND D.PINERO. 1999. Isozymatic variation and phylogenetic relationships be-tween henequen (Agave fourcroydes) and its wild ancestor A. angustifolia(Agavaceae). American Journal of Botany 86: 115–123.
COLUNGA-GARCIA MARIN, P., AND F. MAY-PAT. 1993. Agave studies in Yu-catan, Mexico. I. Past and present germplasm diversity and uses. Eco-nomic Botany 47: 312–327.

1976 [Vol. 88AMERICAN JOURNAL OF BOTANY
COLUNGA-GARCIA MARIN, P., AND F. MAY-PAT. 1997. Morphological vari-ation of henequen (Agave fourcroydes, Agavaceae) germplasm and itswild ancestor (A. angustifolia) under uniform growth conditions: diver-sity and domestication. American Journal of Botany 84: 1449–1465.
CRUZ, C., R. ORELLANA, AND M. L. ROBERT. 1985. Agave research progressin Yucatan. Desert Plants 7: 71–73, 80, 89–92.
DAHLGREN, R. M. T., H. T. CLIFFORD, AND P. F. YEO. 1985. The families ofthe monocotyledons: structure, evolution and taxonomy. Springer-Verlag,Berlin, Germany.
DOUGHTY, L. R. 1936. Chromosome behavior in relation to genetics of Aga-ve. 1. Seven species of fibre Agaves. Journal of Genetics 33: 197–205.
EASTMOND, A., J. L. HERRERA, AND M. L. ROBERT. 2000. La biotecnologıaaplicada al henequen: alternativas para el futuro. CICY (Centro de In-vestigacion Cientıfica de Yucatan, A.C.), Merida, Yucatan, Mexico.
FREEMAN, C. E., AND W. H. REID. 1985. Aspects of the reproductive biologyof Agave lechuguilla Torr. Desert Plants 7: 75–80.
GENTRY, H. S. 1982. Agaves of continental North America. University ofArizona Press, Tucson, Arizona, USA.
JANSSEN, A. W. B., AND J. G. TH. HERMSEN. 1976. Estimating pollen fertilityin Solanum species and haploids. Euphytica 25: 577–586.
LOCK, G. W. 1962. Sisal. Longmans, Green, London, UK.LUDLOW-WIECHERS, B., AND L. OJEDA. 1983. El pollen del genero Agave
de la Penınsula de Yucatan. Boletın de la Sociedad Botanica de Mexico44: 29–42.
OJEDA, L., B. LUDLOW-WIECHERS, AND R. ORELLANA. 1984. Palinologıa dela familia Agavaceae para la Penınsula de Yucatan. Biotica 9: 379–398.
ROBERT, M. L., J. L. HERRERA, J. L. CHAN, AND F. CONTRERAS. 1992. Mi-cropropagation of Agave spp. In Y. P. S. Bajaj [ed.], Biotechnology inagriculture and forestry, vol. 19, High-tech and micropropagation III,306–309. Springer-Verlag, Berlin, Germany.
ROBERT, M. L., J. L. HERRERA, F. CONTRERAS, AND K. N. SCORER. 1987. Invitro propagation of Agave fourcroides Lem. (Henequen). Plant Cell,Tissue and Organ Culture 8: 37–48.
ROBERTS, I. N., T. C. GAUDE, G. HARROD, AND H. G. DICKINSON. 1983.Pollen-stigma interections in Brassica oleracea; a new pollen germina-tion medium and its use in elucidating the mechanism of self incompat-ibility. Theoretical and Applied Genetics 65: 231–238.
SCHAFFER, W. M., AND V. SCHAFFER. 1977. The reproductive biology ofAgavaceae: pollen and nectar production in four Arizona agaves. South-western Naturalist 22: 157–168.
SCHAFFER, W. M., AND V. SCHAFFER. 1979. The adaptive significance ofvariations in reproductive habit in the Agavaceae II: pollinator foragingbehavior and selection for increased reproductive expenditure. Ecology60: 1051–1069.
SLAUSON, L. A. 2000. Pollination biology of two chripterophilous agaves inArizona. American Journal of Botany 87: 825–836.
WUNDERLICH, R. 1950. Die Agavaceae Hutchinsons im Lichte ihrer Em-bryologie, ihres Gynozeuim-Staubblatt-Blattbaues. Osterreichische Bo-tanische Zeitschrift 97: 437–502.
Related Documents











