High genetic variability and lack of population structure in Prochilodus lineatus in the upper Pilcomayo and Bermejo River systems in South-east Bolivia Ruud van den Heuvel _________________________________________________________________________________________________________ Abstract: Prochilodus lineatus is a highly migratory fish species inhabiting the Paraná-Paraguay basin. In the Pilcomayo River system decline of P. lineatus due to anthropogenic activities and isolation of the Pilcomayo River from Paraguay River have caused interest in creating efficient management and conservation strategies. Currently, information to create these strategies is lacking. In particular, migratory patterns for P. lineatus in the upper Paraná are poorly known and only few population genetic studies are available. To asses genetic population structure we genotyped 201 individuals from four locations in the upper Pilcomayo and Bermejo Rivers (Villa Montes, El Pibe, Rio Pilaya and Bermejo). All individuals were genotyped for 5 microsatellite loci found to be informative to assess populations of P. lineatus. Our results show high genetic variability and lack of population structure within the Pilcomayo River system supporting our hypothesis that during the 1990’s a ‘reserve’ fish stock might had been present in the lower floodplains of the Grand Chaco area able to revitalize fish decline. Continued isolation of the Pilcomayo River with the Paraguay basin could pose a threat for this fish stock due to limited possibilities for new genetic diversity to be generated via gene flow. _________________________________________________________________________________________________________ Introduction In recent years, phylogeography and phylogeny have become powerful tools in understanding evolution of migratory fish populations, species and communities in distinct neotropic environments (Piorski, Sanches et al. 2008). By giving insight in how fish populations are influenced by genetic factors such as the impact of mutation, genetic drift and selection as well as historical factors (Piorski, Sanches et al. 2008) these tools provide a basis for assessment and development of efficient management and conservation strategies. Assessment and preservation of biodiversity of migratory fish populations is crucially important to minimize the loss of genetic variation, especially when these species are under threat of decline due to anthropogenic stressors. Decline in species genetic variation may reduce the ability of a species to adapt to environmental changes, ultimately decreasing long term survival (Frankham, Briscoe et al. 2002). So far, little is known about the genetic variation and population structure in migratory freshwater fish populations inhabiting large complex Neotropical River systems. Understanding how their seasonal migratory patterns influence their genetic makeup could have implications for fisheries, aquaculture, management and conservation. Prochilodus lineatus, (Characiformes, Prochilodontidae), commonly known as “sábalo” (in Argentina, Bolivia, Paraguay, Uruguay) or “curimbatá” (in Brazil) is a native freshwater fish with a geographical distribution in the Paraná-Uruguay and the Paraíba do Sul River basins located in South America (Reis, Kullander et al. 2003). Within these basins, P. lineatus is considered the most abundant fish species in terms of absolute biomass (Sverlij, Ros et al. 1993). The fish species is illiophagous, feeding on algae, detritus and associated microorganisms (Bayol and de Yuan Cordiviola 1996, Fugi, Hahn et al. 1996). Therefore Prochilodus is considered a key species for the role it plays in the cycle of organic matter and its position in the river food-chain (Pimm , Bowen 1983, Flecker 1996, Taylor, Flecker et al. 2006). Furthermore, sábalo is of great economic importance to commercial and artisanal fisheries in the Paraná River basin (Sivasundar, Bermingham et al. 2001, Smolders, Guerrero Hiza et al. 2002, Baigun, Minotti et al. 2013).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

High genetic variability and lack of population structure in Prochilodus lineatus
in the upper Pilcomayo and Bermejo River systems in South-east Bolivia
Ruud van den Heuvel
_________________________________________________________________________________________________________
Abstract: Prochilodus lineatus is a highly migratory fish species inhabiting the Paraná-Paraguay basin. In the Pilcomayo River system decline of
P. lineatus due to anthropogenic activities and isolation of the Pilcomayo River from Paraguay River have caused interest in creating efficient
management and conservation strategies. Currently, information to create these strategies is lacking. In particular, migratory patterns for P.
lineatus in the upper Paraná are poorly known and only few population genetic studies are available. To asses genetic population structure we
genotyped 201 individuals from four locations in the upper Pilcomayo and Bermejo Rivers (Villa Montes, El Pibe, Rio Pilaya and Bermejo). All
individuals were genotyped for 5 microsatellite loci found to be informative to assess populations of P. lineatus. Our results show high genetic
variability and lack of population structure within the Pilcomayo River system supporting our hypothesis that during the 1990’s a ‘reserve’ fish
stock might had been present in the lower floodplains of the Grand Chaco area able to revitalize fish decline. Continued isolation of the Pilcomayo
River with the Paraguay basin could pose a threat for this fish stock due to limited possibilities for new genetic diversity to be generated via gene
flow.
_________________________________________________________________________________________________________
Introduction
In recent years, phylogeography and phylogeny have
become powerful tools in understanding evolution of
migratory fish populations, species and communities in
distinct neotropic environments (Piorski, Sanches et al.
2008). By giving insight in how fish populations are
influenced by genetic factors such as the impact of mutation,
genetic drift and selection as well as historical factors
(Piorski, Sanches et al. 2008) these tools provide a basis for
assessment and development of efficient management and
conservation strategies. Assessment and preservation of
biodiversity of migratory fish populations is crucially
important to minimize the loss of genetic variation,
especially when these species are under threat of decline
due to anthropogenic stressors. Decline in species genetic
variation may reduce the ability of a species to adapt to
environmental changes, ultimately decreasing long term
survival (Frankham, Briscoe et al. 2002). So far, little is known
about the genetic variation and population structure in
migratory freshwater fish populations inhabiting large
complex Neotropical River systems. Understanding how
their seasonal migratory patterns influence their genetic
makeup could have implications for fisheries, aquaculture,
management and conservation.
Prochilodus lineatus,
(Characiformes, Prochilodontidae), commonly known as
“sábalo” (in Argentina, Bolivia, Paraguay, Uruguay) or
“curimbatá” (in Brazil) is a native freshwater fish with a
geographical distribution in the Paraná-Uruguay and the
Paraíba do Sul River basins located in South America (Reis,
Kullander et al. 2003). Within these basins, P. lineatus is
considered the most abundant fish species in terms of
absolute biomass (Sverlij, Ros et al. 1993). The fish species is
illiophagous, feeding on algae, detritus and associated
microorganisms (Bayol and de Yuan Cordiviola 1996, Fugi,
Hahn et al. 1996). Therefore Prochilodus is considered a key
species for the role it plays in the cycle of organic matter and
its position in the river food-chain (Pimm , Bowen 1983,
Flecker 1996, Taylor, Flecker et al. 2006). Furthermore,
sábalo is of great economic importance to commercial and
artisanal fisheries in the Paraná River basin (Sivasundar,
Bermingham et al. 2001, Smolders, Guerrero Hiza et al. 2002,
Baigun, Minotti et al. 2013).

Within the Paraná basin the
Pilcomayo and Bermejo Rivers are considered important
migration routes for sábalo as they serve as “fish highways”
towards the spawning grounds located upstream in the
foothills of the Andean mountains. The Pilcomayo and
Bermejo Rivers are tributaries to the large La Plata system
and arise in the Bolivian Andes (Cordillera Oriental, at c 5000
m and 2000 meter above sea level respectively) after which
they cut down through the Andes through an inaccessible
terrain with rapids and narrow canyons ultimately meeting
with the Rio Paraguay in the lower Chaco plains (Figure 1).
Each year during the late dry season, when water levels and
water flow velocity are lowest and water temperature is cool
(<25°C) large shoals of adult sábalo start undertaking
upstream migrations towards the upper parts of the
Bermejo and Pilcomayo Rivers (Smolders, Guerrero Hiza et
al. 2002, Lucas and Baras 2008). During this migration
Prochilodus disperses from feeding grounds located in the
lower parts of the Gran Chaco area and the Paraguay-
Uruguay River system, often covering large distances. After
a migration covering a timespan of several months large
shoals of migrants reach the upstream spawning grounds
located in the Andean foothills. They gather in the open
water of the main channel or tributaries and spawn when
river discharges start increasing at the onset of the rainy
season (Bayley 1973). The peak of reproduction falls in
November-January (Mochek and Pavlov 1998). An
advantage of this reproduction strategy is that Individuals
who survive the migration will move, together with the
spawned eggs and larvae, downstream where they are
distributed over the newly inundated feeding grounds by the
increase in river discharge (Bayley 1973, de Yuan Cordiviola
1992, Araujo‐Lima and Oliveira 1998, Welcomme, Bene et al.
2006, Stassen, van de Ven et al. 2010).
In the Pilcomayo river a
considerable decline of sábalo catches was an important
concern during the 1990’s (Smolders, Guerrero Hiza et al.
2002). Since then, various explanations have been put
forward to explain the decline of P. lineatus in the 1990’s.
Mining operations located near the city of Potosi have
caused heavy metal pollution in the upper parts as well as in
the lower reaches of the river (Hudson-Edwards, Macklin et
al. 2001, Smolders, Lock et al. 2003, Strosnider, Lopez et al.
2014). Sábalo, being a detritivorous species which filters
Figure 1. Map of the Pilcomayo and Bermejo River systems part of the Erosive Andean sub-basin. The dashed lines are (ancient) river beds that remain dry for large parts of the year. The shaded area marks the actual floodplain of the river in 2002. Sample locations are highlighted as red dots on the map. (Edited from Smolders et al. 2002)

large amounts of sediment, could be expected to be very
sensitive to contamination of the sediment. This is
supported by studies showing the effect of heavy metal
contamination through the consumption of sábalo on
communities living along the upper and lower banks of the
river (Miller, Hudson-Edwards et al. 2004, Stassen, Preeker
et al. 2012). Intensive fishing, whose magnitude in the
Pilcomayo River is unknown, and yearly fish kills due to
suspended sediment (Swinkels, Van de Ven et al. 2014) are
also considered to be factors contributing to the decline of
sábalo. It is assumed that the main reason of the observed
sábalo decline during the 1990’s is the loss of the surface of
breeding and feeding areas (marshes in the lower basin) and
a decrease of interconnectivity between these areas and the
main river channel due to river discharge and sedimentation
processes (Smolders, Guerrero Hiza et al. 2002, Stassen, van
de Ven et al. 2010). The Pilcomayo River is characterized by
strong inter-annual variations in river discharge and
sediment load. A trend of declining rainfall in the upper basin
during the 1990’s related to the large-scale climatic “El Niño”
Southern Oscillation ( ENSO) system probably was the main
reason for sábalo decline (Smolders et al., 2000).
Since the 1990’s, ongoing
fluctuations in river discharge and sedimentation processes
have caused the Pilcomayo River to frequently change its
course. The mean sediment load of the Pilcomayo River is
among the highest in the world (Guyot, Calle et al. 1990,
Iriondo 1993, Smolders, Guerrero Hiza et al. 2002) with the
mean annual amount of sediment transported by the river
sufficient to cover as much as 5000 ha of land with one meter
of sediment. Huge amounts of sediment are transported
towards the lower reaches of the river in the Chaco plain
where its deposition provokes silting up as well as diversion
of the river bed, creating an alluvial fan with a total surface
area of 21,000 km2 (Iriondo 1993). During the last decades
the river has deposited its sediment into a tectonic
depression called Estero Patiño (Cordini 1947), located 250
km upstream from the Paraguay River. After the depression
filled up with sediment in the 1940’s (Pool, Usai et al. 1993)
the river started to fill up its own river channel, resulting in a
gradual retreat away from the Paraguay River (figure 2). The
gradual retreat of the Pilcomayo River in the lower Chaco
could have important consequences for sábalo and it is
hypothesized that this retreat has caused the sábalo
population of the Pilcomayo to become more or less isolated
from the populations in the La Plata basin (Stassen, van de
Ven et al. 2010). A recent study has shown the presence of a
single panmictic group with high levels of gene flow within
the La Plata basin (Sivasundar, Bermingham et al. 2001), with
specimens captured more than 1000 km away from the
tagging locality. This is in accordance with the assumption
made by (Castro 1990) that each river basin is thought to
contain a single panmictic population. However,
differentiation of a single panmictic sábalo population into
two or more (sub)populations due to siltation and retraction
of the Pilcomayo River could in the long run be detrimental
for the sábalo population(s) living there, as it results in a loss
of access to the La Plata basins gene pool while at the same
time being negatively affected by anthropogenic and
environmental stressors (Smolders, Guerrero Hiza et al.
2002, Smolders, Lock et al. 2003, Stassen, van de Ven et al.
2010, Swinkels, Van de Ven et al. 2014).
Migration patterns for P.
lineatus have been described for most of the Paraná and Rio
de La Plata basin on the basis of micro-data recording tags,
mark-recapture experiments, biotelemetry and elemental
analyses of hard tissues (Bayley 1973, Godinho and Kynard
2006, Lucas and Baras 2008, Peixer and Petrere Júnior 2009).
However, an evaluation of migration patterns and
population structure has not been described for P. lineatus
for the upper Pilcomayo and Bermejo River basins. To
Figure 1. Retreat of the Pilcomayo river and dynamic creation of new flood plains due to self-blockage (silting up) of the river channel. (from Stassen, van de Ven et al. 2010)
contribute to this lack of knowledge the aim of this study is
to provide insight in the population structure of P. lineatus
in the upper Pilcomayo basin and whether populations
inhabiting the Pilcomayo are affected by the retraction of
the Pilcomayo River from the Paraguay main River channel.
To conduct these measurements powerful molecular
markers and statistical approaches are required (Rueda,
Carriquiriborde et al. 2013). Molecular markers play an
important role as indispensable tools for determining the
genetic variation and biodiversity with high levels of
accuracy and reproducibility (Arif, Khan et al. 2011). Among
these molecular markers microsatellites are potentially the
most informative to questions concerning degree of
relatedness of individuals or groups due to their high
heterozygosity and high mutation rate and the advantage of
an relatively easy and low-cost detection by PCR (Hoshino,
Bravo et al. 2012). Combining microsatellite markers with
the Bayesian clustering approach (Pritchard, Stephens et al.
2000, Falush, Stephens et al. 2003) will allow for an
estimation of distinct populations and allocation of
individuals to discrete migratory stocks, potentially
providing insight into the population structure of P. lineatus.
Despite its ecological value to the ecosystem and its
economic importance to the communities living along the
banks of the Pilcomayo and Bermejo River, so far there has
been insufficient information about the genetic variability
and population structure in Prochilodus lineatus in the upper
Pilcomayo and Bermejo River systems. Such information
might help to formulate appropriate guidelines for proper
management and conservation. In particular, there is
uncertainty about the scope of migratory movements in
these basins and whether the isolation of the Pilcomayo
basin from the greater Parana river system has an effect on
genetic variability and population structure. By comparing
specimens from the Pilcomayo River, a system disconnected
from the La Plata basin, with specimens from the still
connected Bermejo River system we hope to contribute to a
better understanding of the genetic variability and
population structure of P. lineatus in the upper Pilcomayo
and Bermejo basins.
Material & Methods
Sampling and molecular methods
A total of 201 specimens were sampled from the Pilcomayo
and Bermejo River systems in the Tarija Department in
South-eastern Bolivia. The sample locations in the Pilcomayo
River were located near the city of Villa Montes ( 1 km
downstream of Villa Montes, 21°16'50.66"S; 63°27'6.44"W
and 5 km upstream of Villa Montes at a location locally
named ‘El Pibe’, 21°15'30.02"S; 63°32'43.44"W). Other
sample locations were located at a tributary 13 km from the
main channel of the Rio Pilaya (20°56'27.42"S;
Microsatellite Primer sequence (5'-3') Size range (bp) Ta (⁰C) Temp Time Cycle
PL-3 F: 5'-TCTGAGCTGTGAGGAATGGA-3' 185-203 50 95 ⁰C 3 min 1
R: 5'-AGAGCGCTCAAGCACAAGAT-3' 95 ⁰C 30 sec
PL-14 F: 5'-TGCCCAACACTGAAACTGAG-3' 104–134 61 variable 30 sec 29
R: 5'-CTCATCAACCTGCCTGGAAT-3' 72 ⁰C 35 sec
PL-23 F: 5'-TTGGCTACTTCCCCAAACAC-3' 244-252 59 95 ⁰C 30 sec
R: 5'-GGGGAACTAGTTTGACGATGC -3' 53 ⁰C 30 sec 8
PL-119 F: 5'-GAAAAAGGCTAGGGGACTGG-3' 161-207 58 72 ⁰C 35 sec
R: 5'-GAGGAAAAT TGCCTT TTGTAGG-3' 72 ⁰C 2 min 1
PL-139 F: 5'-CAGTGGCATGATGAT TAGATGG-3' 169–235 53
R: 5'-CACCTTTTGTTTGGCTTTTAGG-3'
PL-i61 F: 5'-ACCAAGGTGGGATTCCTTGGTC-3' 210-230 69
R: 5'-TTCTCCATCTCTGGCTGCGCA-3'
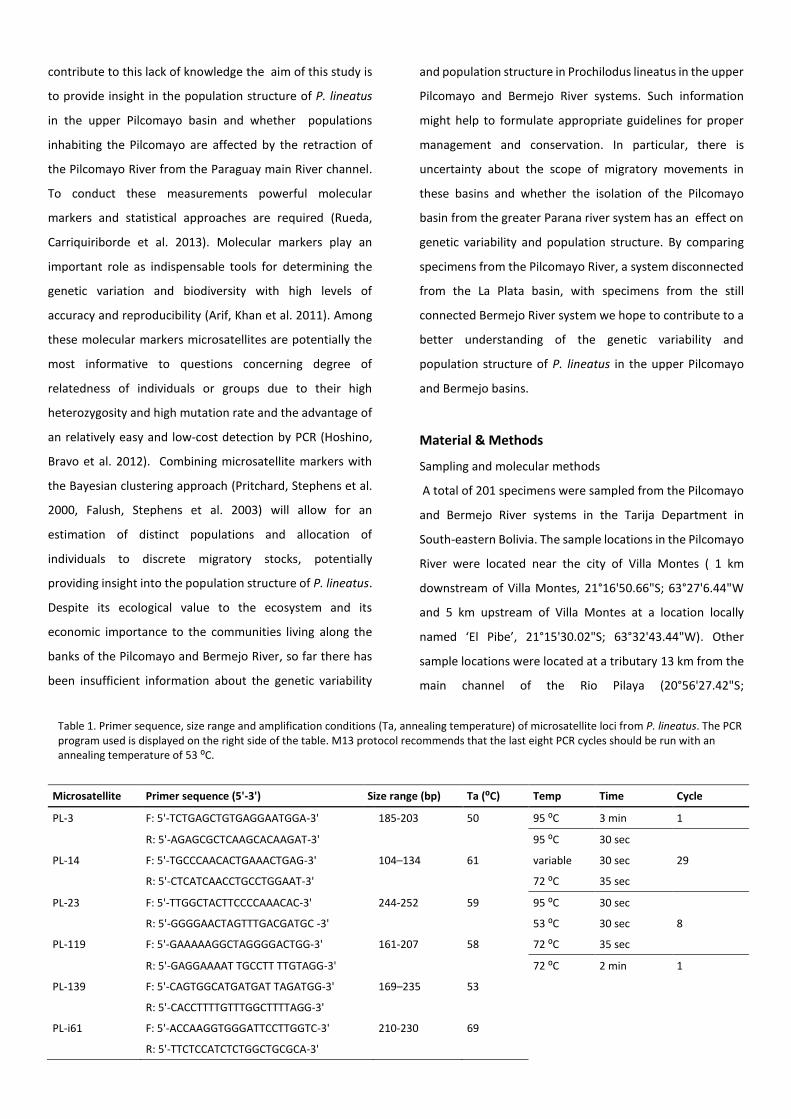
Table 1. Primer sequence, size range and amplification conditions (Ta, annealing temperature) of microsatellite loci from P. lineatus. The PCR program used is displayed on the right side of the table. M13 protocol recommends that the last eight PCR cycles should be run with an annealing temperature of 53 ⁰C.

64°15'32.11"W) and near the city of Bermejo
(22°43'31.37"S; 64°21'32.60"W). See figure 1 for sampling
locations. Samples were obtained in four different months:
April of 2013 (El Pibe1, N=51 individuals), June of 2013 (El
Pibe2, N=30 individuals; Rio Pilaya, N=43 individuals), July of
2013 (Villa Montes, N=50 individuals; Bermejo, N=13
individuals), and August of 2013 (Bermejo, N=14 individuals).
The specimens were caught using cast nets. Specimens were
euthanized and muscle tissue samples and dorsal fin samples
where obtained and kept in 96% alcohol for further analysis.
Genomic DNA was extracted using an QIAGEN DNeasy Blood
& Tissue Kit. Each individual specimen was genotyped for 6
microsatellite loci and amplified by polymerase chain
reaction using the primers PL-3, PL-14, PL-23, PL-119, PL-139
(Rueda, Sommer et al. 2011) and PL-i61(Yazbeck and
Kalapothakis 2007) (table 1). The M13 (-21) tailed-primer
genotyping protocol (Schuelke 2000) was used to facilitate
fast and inexpensive analysis. PCR reactions were conducted
individually with each primer pair in 25 µl reaction volumes,
as follows: 17.5 µl H2O, 0.5 µl 50 mM MgCl2, 2.5 µl 10x NH4
reaction buffer, 0.5 µl 5 U BIOTAQ DNA polymerase, 0.5 µl
5mM dNTPs and 0.5 µl each primer (¾ M13 primer, ¼
forward primer). The PCR program and microsatellite
specific annealing temperatures are displayed in table 1. Five
of the six loci were shown to be selectively neutral and
unlinked for Prochilodus populations (Rueda, Sommer et al.
2011). After PCR microsatellite fragments were scored for
length with an Beckman Coulter CEQ 8800 analyzer
(GenomeLab™). Fragment lengths were assigned to allelic
classes with Genemarker V2.6.2®(SoftGenetics LLC).
Statistical analysis
To test for the occurrence of genotyping errors resulting
from low DNA quality ( large allele dropouts) or PCR artifacts
(null alleles) and for Hardy-Weinberg equilibrium (HWE) we
used MICRO-CHECKER (Van Oosterhout, Hutchinson et al.
2004). Next, basic descriptive parameters such as allele
number, gene diversity, inbreeding coefficient (Fis) and
observed and expected heterozygosity were calculated using
Arelequin v. 3.5.1.2 (Excoffier and Lischer 2010) and GenAlEx
v. 6.5 (Peakall and Smouse 2012). To measure genetic
differentiation between sample locations we used pairwise
Fst with routines implemented in Arlequin v. 3.5.1.2 and
finally, to assess population structure we used the Bayesian
clustering approach implemented in STRUCTURE (Pritchard,
Stephens et al. 2000, Falush, Stephens et al. 2003). Given the
number of clusters (K) and assuming Hardy-Weinberg and
linkage equilibrium within clusters, STRUCTURE estimates
posterior probabilities (allele frequencies in each cluster and
population memberships for every individual) for these K
clusters. Runs were performed using a burn-in of 100000
iterations followed by a Markov chain MonteCarlo of 150000
steps. POPFLAG in structure was disabled and the
“admixture model” and correlated allele frequencies
between groups were chosen following recommendations
by Falush et al. (2007). Analyses were run with a K ranging
from 1 to 10 recommended by Evanno et al. (2005). Analysis
of STRUCTURE results was performed with STRUCTURE
HARVESTER Web v.0.6.94 (Earl and Vonholdt 2012). K=3 was
chosen based on STRUCTURE HARVESTER results and
CLUMPP v.1.1.2b (Jakobsson and Rosenberg 2007) was used
to permute and match cluster output. Distruct v.1.1
(Rosenberg 2004) was used to graphically display results.
Finally, Bottleneck v.1.2.02 was run under three mutation
models (IAM, SSM & TPM), with proportion of SMM in the
TPM = 0.000 and variance of the geometric distribution for
TPM = 0.36, to detect recent effective population size
reduction (Piry, Luikart et al. 1999).
Table 2. Summary statistics of allelic variation of 6 microsatellite loci for P. lineatus
Locus N Allele number Gene diversity
PL-3 194 13 0.67
PL-14 196 20 0.94
PL-23 199 1 0
PL-119 192 64 0.97
PL-139 169 32 0.94
PL-i61 188 8 0.72
Results
All 201 individuals were genotyped for 6 microsatellite loci.
12 samples for which less than 5 loci could be determined
were excluded from analysis. During PCR 41 samples failed
to amplify and were excluded for further analysis. For all loci
and individuals combined, microchecker results suggest
that this population is possibly in Hardy Weinberg
equilibrium with loci PL139, PL-i61, PL-3, showing signs of a
null allele. The presence of null alleles indicates an excess
of homozygotes compared to the HWE. Null alleles are likely
to be encountered in populations with a large effective size
and their presence can result in the overestimation of Fst
values and genetic drift when these populations differ
significantly from one another (Chapuis and Estoup 2007).
No evidence of genotyping errors due to stuttering or allele
dropout were found. PL-23 was shown to be monomorphic
and also excluded from further analysis.
Table 3. Null allele frequency’s calculated with MICROCHECKER
using the methods described by Oosterhout et al. (2004),
Chakraborty et al. (1992) and Brookfield (1996)
Locus Oosterhout Chakraborty Brookfield 1 Brookfield 2
PL14 -0.04 -0.03 -0.03 0.00
PL139 0.30 0.44 0.29 0.42
PL-119 -0.01 -0.01 -0.01 0.00
PL-i61 0.26 0.38 0.23 0.23
PL-3 0.21 0.28 0.18 0.18
Genetic diversity at each locus ranged between 0.666-0.968,
number of alleles observed per locus between 8-64 (Table
2). Average observed heterozygosity values over all loci Ho=
0.61 (SD 0.24) and expected heterozygosity He= 0.8 (SD
0.16). Null allele frequencies are shown in table 3. These
results confirm that the microsatellite data is informative for
population structure analysis.
Population structure
Comparison of samples taken at different locations suggest
that the genetic signature varies greatly within the
Pilcomayo and Bermejo River basins and between different
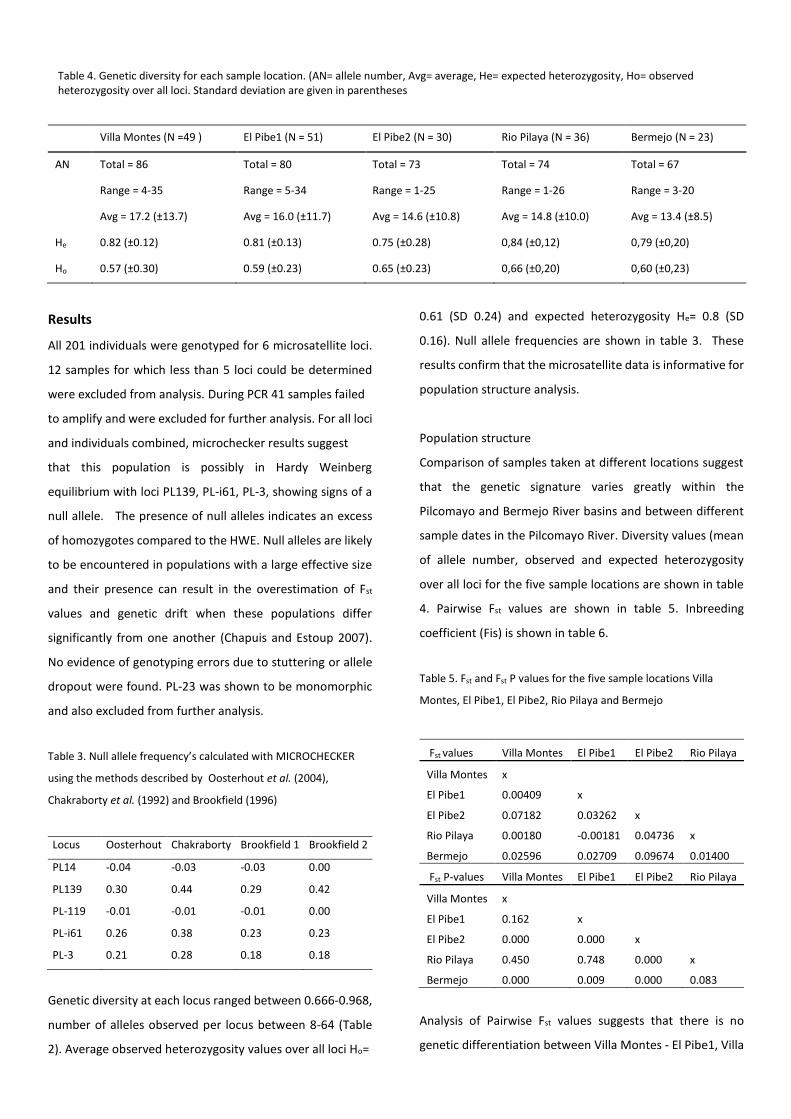
sample dates in the Pilcomayo River. Diversity values (mean
of allele number, observed and expected heterozygosity
over all loci for the five sample locations are shown in table
4. Pairwise Fst values are shown in table 5. Inbreeding
coefficient (Fis) is shown in table 6.
Table 5. Fst and Fst P values for the five sample locations Villa
Montes, El Pibe1, El Pibe2, Rio Pilaya and Bermejo
Fst values Villa Montes El Pibe1 El Pibe2 Rio Pilaya
Villa Montes x
El Pibe1 0.00409 x
El Pibe2 0.07182 0.03262 x
Rio Pilaya 0.00180 -0.00181 0.04736 x
Bermejo 0.02596 0.02709 0.09674 0.01400
Fst P-values Villa Montes El Pibe1 El Pibe2 Rio Pilaya
Villa Montes x
El Pibe1 0.162 x
El Pibe2 0.000 0.000 x
Rio Pilaya 0.450 0.748 0.000 x
Bermejo 0.000 0.009 0.000 0.083
Analysis of Pairwise Fst values suggests that there is no
genetic differentiation between Villa Montes - El Pibe1, Villa
Villa Montes (N =49 ) El Pibe1 (N = 51) El Pibe2 (N = 30) Rio Pilaya (N = 36) Bermejo (N = 23)
AN Total = 86 Total = 80 Total = 73 Total = 74 Total = 67
Range = 4-35 Range = 5-34 Range = 1-25 Range = 1-26 Range = 3-20
Avg = 17.2 (±13.7) Avg = 16.0 (±11.7) Avg = 14.6 (±10.8) Avg = 14.8 (±10.0) Avg = 13.4 (±8.5)
He 0.82 (±0.12) 0.81 (±0.13) 0.75 (±0.28) 0,84 (±0,12) 0,79 (±0,20)
Ho 0.57 (±0.30) 0.59 (±0.23) 0.65 (±0.23) 0,66 (±0,20) 0,60 (±0,23)
Table 4. Genetic diversity for each sample location. (AN= allele number, Avg= average, He= expected heterozygosity, Ho= observed heterozygosity over all loci. Standard deviation are given in parentheses
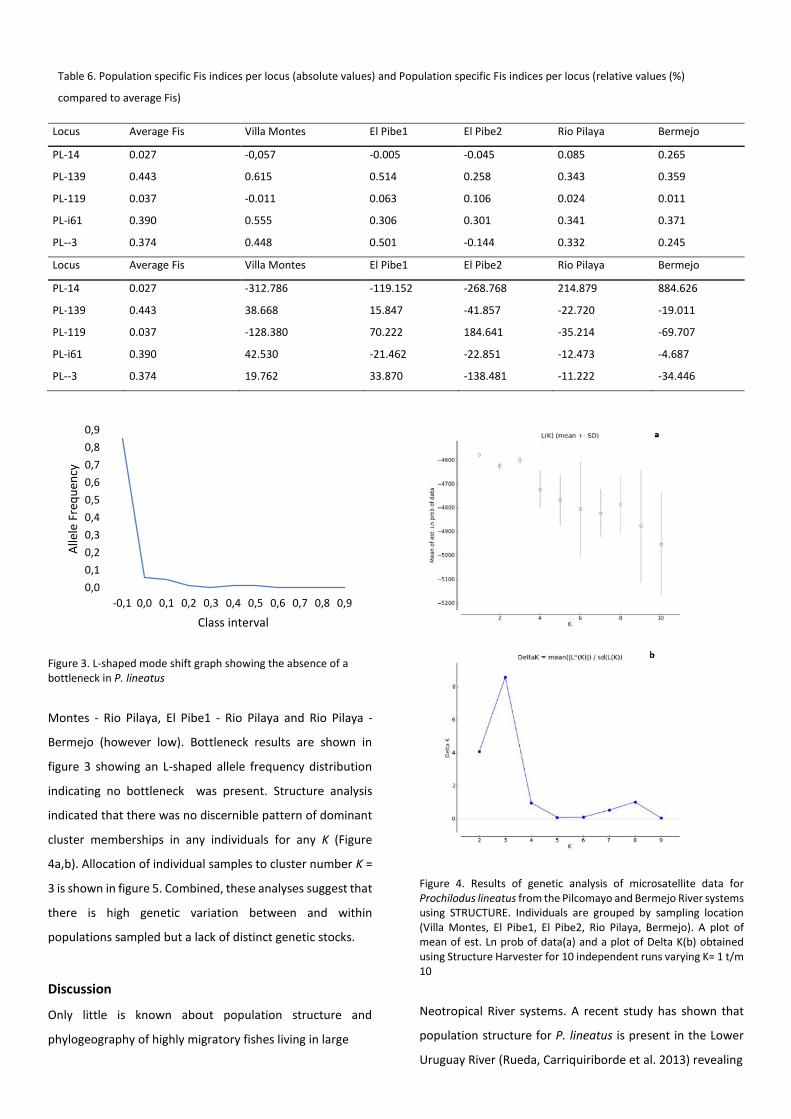
Figure 3. L-shaped mode shift graph showing the absence of a bottleneck in P. lineatus
Montes - Rio Pilaya, El Pibe1 - Rio Pilaya and Rio Pilaya -
Bermejo (however low). Bottleneck results are shown in
figure 3 showing an L-shaped allele frequency distribution
indicating no bottleneck was present. Structure analysis
indicated that there was no discernible pattern of dominant
cluster memberships in any individuals for any K (Figure
4a,b). Allocation of individual samples to cluster number K =
3 is shown in figure 5. Combined, these analyses suggest that
there is high genetic variation between and within
populations sampled but a lack of distinct genetic stocks.
Discussion
Only little is known about population structure and
phylogeography of highly migratory fishes living in large
Figure 4. Results of genetic analysis of microsatellite data for Prochilodus lineatus from the Pilcomayo and Bermejo River systems using STRUCTURE. Individuals are grouped by sampling location (Villa Montes, El Pibe1, El Pibe2, Rio Pilaya, Bermejo). A plot of mean of est. Ln prob of data(a) and a plot of Delta K(b) obtained using Structure Harvester for 10 independent runs varying K= 1 t/m 10
Neotropical River systems. A recent study has shown that
population structure for P. lineatus is present in the Lower
Uruguay River (Rueda, Carriquiriborde et al. 2013) revealing
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
-0,1 0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9
Alle
le F
req
uen
cy
Class interval
Locus Average Fis Villa Montes El Pibe1 El Pibe2 Rio Pilaya Bermejo
PL-14 0.027 -0,057 -0.005 -0.045 0.085 0.265
PL-139 0.443 0.615 0.514 0.258 0.343 0.359
PL-119 0.037 -0.011 0.063 0.106 0.024 0.011
PL-i61 0.390 0.555 0.306 0.301 0.341 0.371
PL--3 0.374 0.448 0.501 -0.144 0.332 0.245
Locus Average Fis Villa Montes El Pibe1 El Pibe2 Rio Pilaya Bermejo
PL-14 0.027 -312.786 -119.152 -268.768 214.879 884.626
PL-139 0.443 38.668 15.847 -41.857 -22.720 -19.011
PL-119 0.037 -128.380 70.222 184.641 -35.214 -69.707
PL-i61 0.390 42.530 -21.462 -22.851 -12.473 -4.687
PL--3 0.374 19.762 33.870 -138.481 -11.222 -34.446
Table 6. Population specific Fis indices per locus (absolute values) and Population specific Fis indices per locus (relative values (%)
compared to average Fis)

seasonal variation between groups migrating upstream
towards the upper Paraná basin. This supports the idea by
Sanches et al. (2012) that different genetic populations of
migratory fishes can co-exist in a single hydrographic system
by segregating during the spawning season. In contrast,
studies done within the upper Paraná basin support the
presence of a single panmictic sábalo population
(Revaldaves, Renesto et al. 1997, Sivasundar, Bermingham et
al. 2001) These studies show high genetic variability and
gene flow but no genetic structure. Our results support these
findings. Within the Pilcomayo and between the Pilcomayo
and Bermejo River basins high levels of genetic variability
and gene flow can be found while population structure does
not seem to be present. The contrast between these studies
shows the that the existence of a single panmictic population
versus different genetic populations can vary greatly for P.
lineatus when sampling locations differ. Furthermore,
temporal effects have shown to be of significance when
sampling during spawning or non-reproductive season
(Sanches 2012). Within our results genetic differentiation is
observed between the El Pibe2 group and the other groups
sampled in the Pilcomayo River. No differentiation was
found between the Villa Montes, El Pibe1 and Rio Pilaya
group suggesting that the specimens collected at these sites
are more closely related than specimens from the El Pibe2
sample site. Specimens were collected in April (El Pibe1),
June (Rio Pilaya, El Pibe2) and July (Villa Montes). These
findings support findings by Sanches et al. (2012) and
highlight the importance of taking into account temporal
effects when doing population genetic research on highly
migratory species.
Observed heterozygosity
in our results is much lower than expected heterozygosity.
This could be ascribed to the presence of null alleles arising
from variation in the nucleotide sequences of flanking
regions that prevent the primer annealing to template DNA
during PCR amplification of the microsatellite locus [Dakin
and Avise, 2004, Pompanon et al., 2005]. The presence of
null alleles results in an excess of homozygous genotypes
within a population as compared to the expected proportion
under Hardy Weinberg Equilibrium (HWE) and Linkage
Equilibrium (LE) [Callen et al., 1993, Paetkau et al., 1995] and
an overestimation of Fst values and genetic drift (Chapuis and
Estoup 2007). Microchecker analysis suggests that loci
PL139, PL-i61, PL-3 show signs of a null allele. This results in
populations sampled having less genetic differentiation than
our analyses suggest. Nevertheless the following conclusions
can be drawn based on our analysis.
Results (table 4) suggest
that genetic variability is present in the Pilcomayo river
system despite the river being isolated from the La Plata
basin since the 1940’s and the massive decline of sábalo
numbers during the 1990’s. Since the last decade sábalo
population size seems to have been increasing. However, it
is well known that genetic variation does not rebound from
a decrease as quickly as population size does (Nei,
Maruyama et al. 1975). The presence of genetic variability
therefore suggests that despite the loss of an important
migration route and a decline in fish numbers the original
gene pool has more or less remained intact. This is
reinforced by Bottleneck results which show that no
bottleneck has occurred based on our allelic data. Our
hypothesis is that this genetic variability has been preserved
in the lower reaches of the Pilcomayo among the floodplains
of the Chaco plain. Each year different sábalo populations
from different regions in the lower floodplains start their
migration upstream. The populations able to start the
migration are chosen depending on the amount of river
Figure 5. Assignment probabilities for each individual (vertical bars) into one of three clusters. Each color represents the amount to which an individual belongs to a certain cluster.

discharge that year and the resulting connectivity of the
main river channel with the floodplains. This interannual and
seasonal variation of interconnectivity between the
floodplains selects the regions from which sábalo is able to
migrate and spawn upstream keeping genetic variation
intact despite anthropogenic and environmental stressors.
For nearly all Pilcomayo
groups differentiation with the Bermejo group can be found
indicating that sábalo inhabiting the Bermejo River are
genetically more distant from the groups in the Pilcomayo
than they are among themselves. These findings support the
idea made by Stassen et al. 2010, that the populations living
in the Pilcomayo River system are more or less isolated from
the bigger sábalo population inhabiting the La Plata basin.
While genetic variation within the Pilcomayo seems to be
currently present a lasting isolation of the river with the
Paraguay channel might have a negative effect on the
sábalo population in the long run. Due to limited possibilities
for new genetic diversity to be generated via gene flow, it
might be expected that sábalo populations living in the
Pilcomayo River will become more vulnerable to extinction
following a population crash. While a ‘reserve’ fish stock
exists in the floodplains able to revitalize sábalo through
interannual and seasonal fluctuations in river discharge,
continued environmental and anthropogenic effects and
isolation of the Pilcomayo River might deplete this reserve
fish stock resulting in a loss of genetic variation for
Prochilodus populations in the Pilcomayo River basin.
Results gained from this
study could be useful for future management and
conservation strategies regarding P. lineatus within the
Pilcomayo River system. Such as the introduction of
Prochilodus from other parts of the Parana River system to
increase genetic diversity or monitoring genetic diversity
within the area of Villa Montes for the detection of possible
future trends.
References
Araujo‐Lima, C. and E. Oliveira (1998). "Transport of larval fish in the Amazon." Journal of fish Biology 53(sA): 297-306.
Arif, I. A., H. A. Khan, A. H. Bahkali, A. A. Al Homaidan, A. H. Al Farhan, M. Al Sadoon and M. Shobrak (2011). "DNA marker technology for wildlife conservation." Saudi Journal of Biological Sciences 18(3): 219-225.
Baigun, C., P. Minotti and N. Oldani (2013). "Assessment of sábalo (Prochilodus lineatus) fisheries in the lower Paraná River basin (Argentina) based on hydrological, biological, and fishery indicators." Neotropical Ichthyology 11: 199-210.
Bayley, P. B. (1973). "Studies on the migratory characin, Prochilodus platensis Holmberg 1889, (Pisces, Characoidei) in the River Pilcomayo, South America." Journal of Fish Biology 5(1): 25-40.
Bayol, V. and E. de Yuan Cordiviola (1996). "Food assimilation of a neotropical riverine detritivorous fish, Prochilodus lineatus, studied by fatty acid composition (Pisces, Curimatidae)." Hydrobiologia 330(2): 81-88.
Bowen, S. (1983). "Detritivory in neotropical fish communities." Environmental Biology of Fishes 9(2): 137-144.
Brookfield, J. (1996). "A simple new method for estimating null allele frequency from heterozygote deficiency." Molecular Ecology 5(3): 453-455.
Castro, R. (1990). "Revisão taxonômica da família Prochilodontidae (Ostriophysi, Characiformes)." Unpublished Ph. D. dissertation. São Paulo: Universidade de São Paulo. 293pp.
Chakraborty, R., M. d. Andrade, S. Daiger and B. Budowle (1992). "Apparent heterozygote deficiencies observed in DNA typing data and their implications in forensic applications." Annals of human genetics 56(1): 45-57.
Chapuis, M.-P. and A. Estoup (2007). "Microsatellite Null Alleles and Estimation of Population Differentiation." Molecular Biology and Evolution 24(3): 621-631.

Cordini, I. R. (1947). Los ríos Pilcomayo en la región del Patiño. Buenos Aires.
de Yuan Cordiviola, E. (1992). "Fish populations of lentic environments of the Paraná River." Hydrobiologia 237(3): 159-173.
Earl, D. A. and B. M. Vonholdt (2012). "STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method." Conservation Genetics Resources 4(2): 359-361.
Evanno, G., S. Regnaut and J. Goudet (2005). "Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study." Molecular Ecology 14(8): 2611-2620.
Excoffier, L. and H. E. L. Lischer (2010). "Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows." Molecular Ecology Resources 10(3): 564-567.
Falush, D., M. Stephens and J. K. Pritchard (2003). "Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies." Genetics 164(4): 1567-1587.
Falush, D., M. Stephens and J. K. Pritchard (2007). "Inference of population structure using multilocus genotype data: dominant markers and null alleles." Molecular ecology notes 7(4): 574-578.
Flecker, A. S. (1996). "Ecosystem Engineering by a Dominant Detritivore in a Diverse Tropical Stream." Ecology 77(6): 1845-1854.
Frankham, R., D. A. Briscoe and J. D. Ballou (2002). Introduction to conservation genetics, Cambridge University Press.
Fugi, R., N. Hahn and A. Agostinho (1996). "Feeding styles of five species of bottom-feeding fishes of the high Paraná river." Environmental Biology of Fishes 46(3): 297-307.
Godinho, A. L. and B. Kynard (2006). "Migration and spawning of radio-tagged zulega Prochilodus argenteus in a dammed Brazilian River." Transactions of the American Fisheries Society 135(3): 811-824.
Guyot, J. L., H. Calle, J. Cortes and M. Pereira (1990). "Transport of Suspended Sediment and Dissolved Material from the Andes to the Rio-De-La-Plata by the Bolivian Tributaries of the Rio Paraguay (Rios Pilcomayo and Bermejo)." Hydrological Sciences Journal-Journal Des Sciences Hydrologiques 35(6): 653-665.
Hoshino, A. A., J. P. Bravo, P. M. Nobile and K. A. Morelli (2012). "Microsatellites as tools for genetic diversity analysis." Genetic Diversity in Microorganisms: 149-170.
Hudson-Edwards, K. A., M. G. Macklin, J. R. Miller and P. J. Lechler (2001). "Sources, distribution and storage of heavy metals in the Rio Pilcomayo, Bolivia." Journal of Geochemical Exploration 72(3): 229-250.
Iriondo, M. (1993). "Geomorphology and Late Quaternary of the Chaco (South-America)." Geomorphology 7(4): 289-303.
Jakobsson, M. and N. A. Rosenberg (2007). "CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure." Bioinformatics 23(14): 1801-1806.
Lucas, M. and E. Baras (2008). Migration of Freshwater Fishes, Wiley.
Miller, J. R., K. A. Hudson-Edwards, P. J. Lechler, D. Preston and M. G. Macklin (2004). "Heavy metal contamination of water, soil and produce within riverine communities of the Rio Pilcomayo basin, Bolivia." Science of the Total Environment 320(2-3): 189-209.
Mochek, A. and D. Pavlov (1998). "The ecology of Sábalo Prochilodus lineatus (Curimatidae, Characoidei) of the Pilcomayo River (South America)." Journal of ichthyology 38(1): 28-36.
Nei, M., T. Maruyama and R. Chakraborty (1975). "The bottleneck effect and genetic variability in populations." Evolution: 1-10.
Peakall, R. and P. E. Smouse (2012). "GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update." Bioinformatics 28(19): 2537-2539.

Peixer, J. and M. Petrere Júnior (2009). "Sport fishing in Cachoeira de Emas in Mogi-Guaçu River, State of São Paulo, Brazil." Brazilian Journal of Biology 69(4): 1081-1090.
Pimm, S. L. "Food webs too food webs: integration of patterns and dynamics." Trends in Ecology & Evolution 11(8): 349.
Piorski, N. M., A. Sanches, L. F. Carvalho-Costa, T. Hatanaka, M. Carrillo-Avila, P. D. Freitas and P. M. Galetti (2008). "Contribution of conservation genetics in assessing neotropical freshwater fish biodiversity." Brazilian Journal of Biology 68(4): 1039-1050.
Piry, S., G. Luikart and J.-M. Cornuet (1999). "BOTTLENECK: a program for detecting recent effective population size reductions from allele data frequencies." Montpellier, France.
Pool, M., E. Usai and J. Baas (1993). "Estudio del Rio Pilcomayo." Brussels, EURAGRI Temporary Association.[Links].
Pritchard, J. K., M. Stephens and P. Donnelly (2000). "Inference of population structure using multilocus genotype data." Genetics 155(2): 945-959.
Reis, R. E., S. O. Kullander, C. J. Ferraris and P. U. C. d. R. G. d. S. M. d. C. e. Tecnologia (2003). Check List of the Freshwater Fishes of South and Central America, EDIPUCRS.
Revaldaves, E., E. Renesto and M. F. Machado (1997). "Genetic variability of Prochilodus lineatus (Characiformes, Prochilodontidae) in the upper Paraná river." Brazilian Journal of Genetics 20(3).
Rosenberg, N. A. (2004). "DISTRUCT: a program for the graphical display of population structure." Molecular Ecology Notes 4(1): 137-138.
Rueda, E. C., P. Carriquiriborde, A. M. Monzon, G. M. Somoza and G. Orti (2013). "Seasonal variation in genetic population structure of sabalo (Prochilodus lineatus) in the Lower Uruguay River." Genetica 141(7-9): 401-407.
Rueda, E. C., J. Sommer, P. Scarabotti, R. Markariani and G. Orti (2011). "Isolation and characterization of polymorphic microsatellite loci in the migratory freshwater fish Prochilodus lineatus (Characiformes:
Prochilodontidae)." Conservation Genetics Resources 3(4): 681-684.
Sanches, A. and P. M. Galetti (2012). "Population genetic structure revealed by a school of the freshwater migratory fish, Brycon hilarii." Latin American Journal of Aquatic Research 40(2): 408-417.
Sanches, A. G. J., Pedro M Galzerani,Felipe Derazo,Janeth Cutilak-Bianchi,Beatriz Hatanaka,Terumi (2012). "Genetic population structure of two migratory freshwater fish species (Brycon orthotaenia and Prochilodus argenteus) from the São Francisco River in Brazil and its significance for conservation." Latin american journal of aquatic research.
Schuelke, M. (2000). "An economic method for the fluorescent labeling of PCR fragments." Nature Biotechnology 18(2): 233-234.
Sivasundar, A., E. Bermingham and G. Ortí (2001). "Population structure and biogeography of migratory freshwater fishes (Prochilodus: Characiformes) in major South American rivers." Molecular Ecology 10(2): 407-417.
Smolders, A. J. P., M. A. Guerrero Hiza, G. van der Velde and J. G. M. Roelofs (2002). "Dynamics of discharge, sediment transport, heavy metal pollution and Sábalo (Prochilodus lineatus) catches in the lower Pilcomayo river (Bolivia)." River Research and Applications 18(5): 415-427.
Smolders, A. J. P., R. A. C. Lock, G. Van der Velde, R. I. M. Hoyos and J. G. M. Roelofs (2003). "Effects of mining activities on heavy metal concentrations in water, sediment, and macroinvertebrates in different reaches of the Pilcomayo River, South America." Archives of Environmental Contamination and Toxicology 44(3): 314-323.
Stassen, M. J., M. W. van de Ven, T. van der Heide, M. A. G. Hiza, G. van der Velde and A. J. Smolders (2010). "Population dynamics of the migratory fish Prochilodus lineatus in a neotropical river: the relationships with river discharge, flood pulse, El Niño and fluvial megafan behaviour." Neotropical Ichthyology 8(1): 113-122.
Stassen, M. J. M., N. L. Preeker, A. M. J. Ragas, M. W. P. M. van de Ven, A. J. P. Smolders and N. Roeleveld (2012). "Metal exposure and reproductive disorders in

indigenous communities living along the Pilcomayo River, Bolivia." Science of the Total Environment 427: 26-34.
Strosnider, W. H. J., F. S. L. Lopez, J. A. LaBar, K. J. Palmer and R. W. Nairn (2014). "Unabated acid mine drainage from Cerro Rico de Potosi, Bolivia: uncommon constituents of concern impact the Rio Pilcomayo headwaters." Environmental Earth Sciences 71(7): 3223-3234.
Sverlij, S. B., A. E. Ros and G. Orti (1993). Sinopsis de los datos biológicos y pesqueros del sábalo, Prochilodus lineatus (Valenciennes, 1847), Food & Agriculture Org.
Swinkels, L. H., M. W. P. M. Van de Ven, M. J. M. Stassen, G. Van der Velde, H. J. R. Lenders and A. J. P. Smolders (2014). "Suspended sediment causes annual acute fish mortality in the Pilcomayo River (Bolivia)." Hydrological Processes 28(1): 8-15.
Taylor, B. W., A. S. Flecker and R. O. Hall (2006). "Loss of a Harvested Fish Species Disrupts Carbon Flow in a Diverse Tropical River." Science 313(5788): 833-836.
Van Oosterhout, C., W. F. Hutchinson, D. P. M. Wills and P. Shipley (2004). "MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data." Molecular Ecology Notes 4(3): 535-538.
Welcomme, R. L., C. Bene, C. A. Brown, A. Arthington, P. Dugan, J. M. King and V. Sugunan (2006). Predicting the water requirements of river fisheries. Wetlands and Natural Resource Management, Springer: 123-154.
Yazbeck, G. M. and E. Kalapothakis (2007). "Isolation and characterization of microsatellite DNA in the piracema fish Prochilodus lineatus (Characiformes)." Genetics and Molecular Research 6(4): 1026-1034.
Related Documents











