The FASEB Journal • FJ Express Full-Length Article Reoxygenation-specific activation of the antioxidant transcription factor Nrf2 mediates cytoprotective gene expression in ischemia-reperfusion injury Martin O. Leonard,* ,1 Niamh E. Kieran,* Katherine Howell,* Melissa J. Burne, ‡ Raghu Varadarajan, † Saravanakumar Dhakshinamoorthy, § Alan G. Porter, § Cliona O’Farrelly, † Hamid Rabb, ‡ and Cormac T. Taylor* *School of Medicine and Medical Science, UCD Conway Institute of Biomolecular and Biomedical Research and † Education and Research Center, St. Vincent’s University Hospital, University College Dublin, Ireland; ‡ Nephrology Division, Johns Hopkins University Hospital, Baltimore, Maryland, USA; and § Institute of Molecular and Cell Biology, Singapore, Republic of Singapore ABSTRACT Tissue reoxygenation following hypoxia is associated with ischemia-reperfusion injury (IRI) and may signal the development of ischemic preconditioning, an adaptive state that is protective against subsequent IRI. Here we used microarray RNA analysis of in vivo and in vitro models of IRI to delineate the underlying molecular mechanisms. Microarray analysis of renal tissue after ischemia-reperfusion revealed a number of highly up- regulated antioxidant genes including aldehyde dehydro- genases (ALDH1A1 and ALDH1A7), glutathione S-trans- ferases (GSTM5, GSTA2 and GSTP1), and NAD(P)H quinone oxidoreductase (NQO1). The transcription fac- tor NF-E2-related factor-2 (Nrf2), a master regulator of this antioxidant response, is also elevated in IRI. Further- more, microarray analysis of renal epithelial cells exposed to hypoxia/reoxygenation identified Nrf2 to be up-regu- lated on reoxygenation. We also reveal a reoxygenation- specific nuclear accumulation of Nrf2 protein and subse- quent activation of a NQO1 promoter reporter construct. Attenuating reactive oxygen species (ROS) in reoxygen- ation using the antioxidant N-acetyl cysteine results in inhibition of Nrf-2 activation. mRNA levels for Nrf2- dependent genes were detected in human liver biopsy 1 h after transplantation. These results indicate that reoxy- genation-dependent Nrf-2 activity facilitates ischemic pre- conditioning through the induction of antioxidant gene expression and that ROS may be critical in signaling this event.—Leonard, M. O., Kieran, N. E., Howell, K., Burne, M. J., Varadarajan, R., Dhakshinamoorthy, S., Porter, A. G., O’Farrelly, C., Rabb, H., Taylor, C. T. Reoxygen- ation-specific activation of the antioxidant transcription factor Nrf2 mediates cytoprotective gene expression in ischemia reperfusion injury. FASEB J. 20, E2166 –E2176 (2006) Key Words: antioxidant response element hypoxia ischemic preconditioning NQO1 Ischemic reperfusion injury, which occurs when blood flow is disrupted to a tissue or organ and subsequently reintroduced, is associated with a diverse array of disease states including myocardial infarction, stroke, acute renal failure, and post-transplantation injury (1– 4). In these conditions much of the damage occurs during the reperfusion period, when, for exam- ple, thrombolytic therapy has been administered or perfusion has been restored after transplantation. A primary factor in the initiation of the pathological response to reperfusion injury is the generation of high levels of ROS, which can covalently modify protein and lipid macromolecules leading to cell damage, DNA mutation, and initiation of apoptotic and necrotic cascades (5). Apart from ROS release from infiltrating cells such as neutrophils, oxygen deprivation (hypoxia) during ischemia and subsequent reoxygenation upon reperfusion are thought to be the major factors con- tributing to ROS production and the subsequent cellu- lar damage (5). It has been well characterized that exposure to ischemia and reperfusion leads to induc- tion of a protective state against subsequent ischemic- reperfusion events—namely, ischemic preconditioning (6). Mechanisms underlying the induction of an isch- emic preconditioned state remain unclear but likely involve the cells’ ability to maintain sufficient antioxi- dant and detoxification buffering capacity. An important mechanism by which cells adapt to oxidant stress is to transcriptionally up-regulate a dis- tinct array of cytoprotective genes responsible for buff- ering the cells’ antioxidant capacity. These genes act to maintain glutathione content and conjugational activ- ity; they are also responsible for detoxification of dam- aging electrophilic by-products of oxidant stress and include glutathione S-transferases, aldehyde dehygro- genases, and quinone oxidoreductases (7). A master regulator of this specific antioxidant phenotype is the 1 Correspondence: School of Medicine and Medical Sci- ences, The Conway Institute, University College Dublin, Belfield, Dublin 4, Ireland. E-mail: [email protected] doi: 10.1096/fj.05-5097fje E2166 0892-6638/06/0020-2166 © FASEB

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The FASEB Journal • FJ Express Full-Length Article
Reoxygenation-specific activation of the antioxidanttranscription factor Nrf2 mediates cytoprotectivegene expression in ischemia-reperfusion injury
Martin O. Leonard,*,1 Niamh E. Kieran,* Katherine Howell,* Melissa J. Burne,‡
Raghu Varadarajan,† Saravanakumar Dhakshinamoorthy,§ Alan G. Porter,§
Cliona O’Farrelly,† Hamid Rabb,‡ and Cormac T. Taylor**School of Medicine and Medical Science, UCD Conway Institute of Biomolecular and BiomedicalResearch and †Education and Research Center, St. Vincent’s University Hospital, University CollegeDublin, Ireland; ‡Nephrology Division, Johns Hopkins University Hospital, Baltimore, Maryland,USA; and §Institute of Molecular and Cell Biology, Singapore, Republic of Singapore
ABSTRACT Tissue reoxygenation following hypoxia isassociated with ischemia-reperfusion injury (IRI) and maysignal the development of ischemic preconditioning, anadaptive state that is protective against subsequent IRI.Here we used microarray RNA analysis of in vivo and invitro models of IRI to delineate the underlying molecularmechanisms. Microarray analysis of renal tissue afterischemia-reperfusion revealed a number of highly up-regulated antioxidant genes including aldehyde dehydro-genases (ALDH1A1 and ALDH1A7), glutathione S-trans-ferases (GSTM5, GSTA2 and GSTP1), and NAD(P)Hquinone oxidoreductase (NQO1). The transcription fac-tor NF-E2-related factor-2 (Nrf2), a master regulator ofthis antioxidant response, is also elevated in IRI. Further-more, microarray analysis of renal epithelial cells exposedto hypoxia/reoxygenation identified Nrf2 to be up-regu-lated on reoxygenation. We also reveal a reoxygenation-specific nuclear accumulation of Nrf2 protein and subse-quent activation of a NQO1 promoter reporter construct.Attenuating reactive oxygen species (ROS) in reoxygen-ation using the antioxidant N-acetyl cysteine results ininhibition of Nrf-2 activation. mRNA levels for Nrf2-dependent genes were detected in human liver biopsy 1 hafter transplantation. These results indicate that reoxy-genation-dependent Nrf-2 activity facilitates ischemic pre-conditioning through the induction of antioxidant geneexpression and that ROS may be critical in signaling thisevent.—Leonard, M. O., Kieran, N. E., Howell, K., Burne,M. J., Varadarajan, R., Dhakshinamoorthy, S., Porter,A. G., O’Farrelly, C., Rabb, H., Taylor, C. T. Reoxygen-ation-specific activation of the antioxidant transcriptionfactor Nrf2 mediates cytoprotective gene expression inischemia reperfusion injury. FASEB J. 20, E2166–E2176(2006)
Key Words: antioxidant response element � hypoxia � ischemicpreconditioning � NQO1
Ischemic reperfusion injury, which occurs whenblood flow is disrupted to a tissue or organ and
subsequently reintroduced, is associated with a diversearray of disease states including myocardial infarction,stroke, acute renal failure, and post-transplantationinjury (1–4). In these conditions much of the damageoccurs during the reperfusion period, when, for exam-ple, thrombolytic therapy has been administered orperfusion has been restored after transplantation. Aprimary factor in the initiation of the pathologicalresponse to reperfusion injury is the generation of highlevels of ROS, which can covalently modify protein andlipid macromolecules leading to cell damage, DNAmutation, and initiation of apoptotic and necroticcascades (5). Apart from ROS release from infiltratingcells such as neutrophils, oxygen deprivation (hypoxia)during ischemia and subsequent reoxygenation uponreperfusion are thought to be the major factors con-tributing to ROS production and the subsequent cellu-lar damage (5). It has been well characterized thatexposure to ischemia and reperfusion leads to induc-tion of a protective state against subsequent ischemic-reperfusion events—namely, ischemic preconditioning(6). Mechanisms underlying the induction of an isch-emic preconditioned state remain unclear but likelyinvolve the cells’ ability to maintain sufficient antioxi-dant and detoxification buffering capacity.
An important mechanism by which cells adapt tooxidant stress is to transcriptionally up-regulate a dis-tinct array of cytoprotective genes responsible for buff-ering the cells’ antioxidant capacity. These genes act tomaintain glutathione content and conjugational activ-ity; they are also responsible for detoxification of dam-aging electrophilic by-products of oxidant stress andinclude glutathione S-transferases, aldehyde dehygro-genases, and quinone oxidoreductases (7). A masterregulator of this specific antioxidant phenotype is the
1 Correspondence: School of Medicine and Medical Sci-ences, The Conway Institute, University College Dublin,Belfield, Dublin 4, Ireland. E-mail: [email protected]
doi: 10.1096/fj.05-5097fje
E2166 0892-6638/06/0020-2166 © FASEB

transcription factor Nrf2 (8). This transcription factoris held in the cytoplasm by a cytoskeletal-associatedspecific inhibitory protein KEAP1 under conditions ofnormal cellular redox state, where Nrf2 is continuouslytargeted to proteasomal degradation. Under conditionsof oxidative stress, cysteine residues within the hingeregion of KEAP1 become modified through mecha-nisms that involve thiol oxidation, resulting in a con-formational change in KEAP1 with the loss of Nrf2binding and proteasomal targeting. Nrf2 then accumu-lates and localizes to the nucleus, where it heterodimer-izes with specific cofactors, including members of themaf protein family, and coordinates up-regulation ofcytoprotective genes through the initiation of transac-tivation at antioxidant response elements (AREs)within the regulatory regions of these genes (9). Whilemuch of the work to characterize Nrf2 activation anddependent cytoprotective gene expression has beencarried out in response to chemical oxidants such assulforaphane and tBHQ, no studies have been per-formed to determine the role for Nrf2 in mediatingadaptive responses to reperfusion and reoxygenation-mediated cellular injury
In this study we demonstrate the induction of acohort of Nrf2-dependent antioxidant genes in a mu-rine model of renal IRI. We also demonstrate activationof Nrf2-dependent gene expression in liver biopsy aswell as a reoxygenation-specific induction of Nrf2 geneexpression and activation of Nrf2-dependent ARE tran-scriptional activity. These are the first data to demon-strate that reoxygenation-mediated activation of Nrf2 isa likely master regulator for the coordinated adaptivecytoprotective genomic response to oxidative stressduring reperfusion injury as a contributory mechanismfor the development of ischemic preconditioning.
MATERIALS AND METHODS
Ischemia reperfusion injury
Acute renal failure was induced using an established animalmodel of renal IRI (10). National Institutes of Health (NIH)Swiss mice (25 to 35 g) were anesthetized with 75 mg/kgintraperitoneal sodium pentobarbital. Incisions were made inthe abdomen and the renal pedicles were exposed throughblunt dissection. A microvascular clamp was placed on bothrenal pedicles for 30 min. During the procedure, animalswere kept hydrated with saline and body temperature wasmaintained at 36–38°C. After the procedure, clamps wereremoved and the wounds were sutured. Animals were allowedto recover for 24 h, then the kidneys were dissected and snapfrozen before processing for protein and RNA isolation.
Cell culture and experimental conditions
Human proximal tubular epithelial cells (HK-2; AmericanTissue Type Culture Collection; (11) were maintained inDulbecco/Vogt modified Eagle’s medium and Ham’s F-12medium (DMEM/F-12) containing 5 �g/ml insulin, 5 �g/mltransferrin, 5 ng/ml selenium, 36 ng/ml hydrocortisone, 4
pg/ml triiodo-l-thyronine, 10 ng/ml epidermal growth factor(EGF), 50U/ml penicillin, 50 �g/ml streptomycin, and 2 mMglutamine (Sigma-Aldrich, Dublin, Ireland). T84 intestinalepithelial cells were grown in DMEM/F-12 supplementedwith 2 mM l-glutamine, 100 U/ml penicillin, 100 �g/mlstreptomycin, and 10% fetal calf serum (Invitrogen, Paisley,UK). Cells were cultured to confluency in 35 mm diameterPetri dishes prior to experimental protocols. Cells weremaintained at 37°C in a humidified atmosphere under hyp-oxic [1% O2, 20 torr (1 torr�133Pa)] or normoxic/reoxy-genation [21% O2, 147 torr] conditions with a balance of95%N2/5%CO2. Hypoxic conditions were maintained usingan environmentally controlled hypoxia chamber (Coy Labs,Glass Lake, MI, USA). Cellular PO2 concentrations weremeasured by fluorescence quenching oxymetry (Oxylite-2000; Oxford Optronix, Oxford, UK) (12).
RNA preparation and cDNA synthesis
RNA was isolated from cell line and tissue samples usingTrireagent (Sigma-Aldrich). Briefly, after experimental pro-cedure, cells were scraped and lysed in 1 ml of reagent. Tissuesamples were homogenized in reagent using a Bertholdhand-held homogenizer. Samples were left to stand for 5 minat room temperature before the addition of 0.2 ml chloro-form. Samples were then shaken vigorously for 15 s andallowed to stand for 10 min at room temperature beforecentrifugation at 12,000 g for 15 min at 4°C. The upperaqueous phase was removed to a separate tube and RNA wasprecipitated using 0.5 ml isopropanol After centrifugationthe pellet was washed with 75% ethanol and resuspended in50 �l of TE buffer. RNA was quantified as absorbance at 260nM. RNA was reverse transcribed to single-stranded cDNAusing the Superscript Choice kit reagents (Invitrogen, Carls-bad, CA, USA). Pooled total RNA (5 �g) was denatured at65°C for 15 min and added to 1 nM of T7-(dT) primer(5�-(5�-GGCCAGTGAATTGTAATACGACTCACTATAGGGAGG-CGG-(dT)24-3�). The sample was incubated at 70°C for 10min, placed on ice, and centrifuged at 14,000 g for 30 s. Thesample was then added to 5 � first-strand cDNA buffer, 10mM DTT, and 500 �M dNTP mix, vortexed, and incubated at42°C for 2 min. After the addition of SuperScript II reversetranscriptase for 1 h at 42°C, samples were centrifuged brieflyand placed on ice. Second-strand cDNA synthesis was carriedout through the addition of 5 � second-strand reactionbuffer, 200 �M dNTP mix, DNA ligase (10U), DNA polymer-ase I (40U), and Rnase H (2U). After incubation at 16°C for2 h, 20U of T4 DNA polymerase was added and subsequentlyincubated at 16°C for 5 min. Double-stranded cDNA waspurified using the GeneChip sample cleanup module per themanufacturer’s instructions and eluted in TE buffer (Qiagen,Valencia, CA, USA).
Microarray analysis
Sample preparation and microarray analysis were carried outas described previously (13). Briefly, after total RNA extrac-tion and cDNA synthesis, complementary biotin-labeledcRNA was prepared using the Bioarray High Yield RNAtranscript labeling kit (ENZO Life Sciences Inc., Farming-dale, NY, USA). The labeled cRNA was washed using theGeneChip sample cleanup module per the manufacturer’sinstructions (Qiagen, Valencia, CA, USA). cRNA (20 �g) wasfragmented by addition of 5 � fragmentation buffer (200 mMTris acetate, pH 8.2, 500 mM KOAc, 150 mM MgOAc [madeup fresh in diethyl pyrocarbonate (DEPC) dH2O)] andincubated at 94°C for 35 min, placed on ice, and stored at
E2167Nrf2 ACTIVATION ON REOXYGENATION

–20°C until hybridization. After confirmation of cRNA integ-rity through hybridization to test microarrays, fragmentedcRNA was hybridized to human HG U95A or murine U77Amicroarrays (Affymetrix, Santa Clara, CA, USA) for 16 h,washed, and stained for 2 h with fluorescent detectionreagent streptavidin-phycoerythrin. Microarrays were thenscanned using an argon-ion laser (excitation 488 nm, detec-tion 570 nm) and results were presented as fluorescentintensities. Microarray Suite 5.0 software (Affymetrix) wasused to analyze the relative abundance of each gene from theaverage difference of fluorescent intensities.
Real-time and semiquantitative RT-polymerase chainreaction (RT-PCR) analysis
Target gene forward and reverse primers for real-time andsemiquantitative RT-PCR analysis were designed using thePrimer 3 software package (Table 1) and synthesized bySigma-Genosys (Haverhill, Suffolk, UK). Real-time polymer-ase chain reaction (PCR) was carried out using commerciallyavailable SYBR green-based detection reagents (Roche,Branchburg, NJ, USA). Each sample contained 50 ng ofcDNA, 0.4 mM each of the forward and reverse primers, and0.1 mM TaqMan™ probe (18S rRNA internal standard con-trol only). Temperature conditions consisted of a 5 min stepat 95°C, followed by 40 cycles of 60°C for 1 min and 95°C for15 s performed using a 7900HT sequence detector (AppliedBiosystems, Foster City, CA, USA). All measurements wereperformed in duplicate and water controls were negative inall runs. Data were collected during each extension phase ofthe PCR reaction and analyzed with the SDS software package(Applied Biosystems). Threshold cycles were determined foreach gene and quantification of templates was performedaccording to the standard curve method. The expression levelof each target gene was given as relative amount normalizedagainst 18S standard controls.
Semiquantitative RT-PCR analysis was carried out on cDNAusing a standard reaction mixture for amplification as describedpreviously (14). The amplification protocol included an initial 2min denaturation at 94°C, followed by 26 cycles of 94°C for 40 s,55°C for 40 s, 72°C for 1 min, with a final extension of 72°C for5 min. Amplification of 18S RNA (14 cycles) was carried out asa control for equal sample loading. PCR products were analyzedusing a 2% agarose gel in 0.5 � TAE buffer, stained withethidium bromide, and visualized under UV illumination(Uvipro GDS8000, UVItech, Cambridge, UK).
Immunofluorescent staining for Nrf2
Frozen kidney tissue embedded in OCT (optimal cuttingtemperature compound) was cut to 10 �m sections, dehy-drated using graded ethanol steps, rehydrated, and fixed withmethanol. After rinsing in PBS, tissue sections were perme-abilized in 0.2% Triton-X 100 in PBS, then blocked using 1%goat serum for 1 h at room temperature. Sections wereincubated with primary antibody (Ab) (polyclonal rabbitanti-Nrf2 (2 �g/ml), Abcam, Cambridge, UK) overnight at4°C. Sections were washed with PBS and incubated withsecondary Ab (Oregon green-linked goat anti-rabbit (1:500)Invitrogen, Paisley, UK) for 1 h at room temperature. DNAwas stained with propidium iodide (Sigma, Poole, Dorset,UK) for 5 min at room temperature. Specific staining wasassured using control slides where the primary Ab was omit-ted. Images were visualized using confocal microscopy with a�20 objective on a UV Zeiss 510 Meta System laser scanningmicroscope, analyzed using the LSM 5 browser imagingsoftware.
Whole-cell and nuclear protein extract preparation andWestern blot analysis
Whole-cell extracts were prepared in radio-immuno-precipi-tation assay (RIPA) lysis buffer (20 mmol/L Tris-HCl, pH 7.4,50 mmol/L NaCl, 5 mmol/L ethylene diaminetetraaceticacid, 1% Nonidet P-40, 0.1% SDS, 5 mmol/L NaF, 1 mmol/Lphenylmethylsulfonyl fluoride, 1 mmol/L Na3VO4, 1�mol/L leupeptin, 0.3 �mol/L aprotinin). Nuclear extractswere prepared from cells scraped into hypotonic buffer (10mM HEPES-NaOH buffer, pH7.9, containing 1.5 mM MgCl2,10 mM KCl, 0.5 mM DTT, and 0.5 mM PMSF.), incubated for10 min, and centrifuged for 10 min at 21,000 g at 4°C.Samples were then lysed for 10 min on ice in hypotonic buffercontaining 0.1% (v/v) Nonidet P-40, followed by centrifuga-tion at 21,000 g for 6 min. The resulting pellets wereresuspended in high-salt buffer [20 mM HEPES-NaOH bufferpH7.9 containing 420 mM NaCl, 1.5 mM MgCl2, 0.2 mMEDTA, 25% (w/v) glycerol, and 0.5 mM PMSF] for 15 min onice followed by centrifugation at 21,000 g for 6 min at 4°C.Supernatants were removed as the nuclear extracts. Proteincontent was quantified and normalized using the Bradfordmethod (Bio-Rad Laboratories, Hemel Hempstead, Hertford-shire, UK) and electrophoresed on 10% SDS-PAGE gels.Expression levels for Nrf1 and Nrf2 were measured usingspecific antibodies (Santa Cruz Biotechnology, Santa Cruz,CA, USA) by Western blot analysis as described previously
TABLE 1. Primer sequences for real-time PCR analysis
Forward Reverse
MurineAldh1a7 5�-TTTGTGAGGAGGAGTGTGG-3� 5�-TTGGCTCCTTCTTTCTTCC-3�NQO1 5�-ATCCTGGAAGGATGGAAG-3� 5�-CCTGCCTGGAAGTTTAGG-3�GSTM5 5�-GAACAGCTACCTGCACAGC-3� 5�-CCTTCAGGTTTGGGAACTC-3�GSTA2 5�-TGGAGATTGATGGGATGAAG-3� 5�-CAATCAGGGCTCTCTCCTTC-3�GBE1 5�-CTTCCGTTTTGATGGTGTC-3� 5�-CTGGGTACAACGTGTGAGC-3�ALDH1A1 5�-CCTTGGAAATCCTCTGACC-3� 5�-TCCTCCACATTCCAGTTTG-3�GSTP1 5�-GCCACCATACACCATTGTC-3� 5�-AGCCTTGCATCCAGGTATC-3�
HumanNRF2 5�-AAACCAGTGGATCTGCCAAC-3� 5�-GACCGGGAATATCAGGAACA-3�ALDH1A7 5�-TTTGTGAGGAGGAGTGTGG-3� 5�-TTGGCTCCTTCTTTCTTCC-3�GSTP1 5�-GCTCTATGGGAAGGACCAG-3� 5�-CTCAAAAGGCTTCAGTTGC-3�
E2168 Vol. 20 December 2006 LEONARD ET AL.The FASEB Journal
(15). Phosphorylation-specific (Ser-473) Akt Ab was obtainedfrom Abcam (Cambridge, UK).
Transient transfection
Cells were transfected with 2 �g of an ARE promoter-luciferase reporter construct [NQO1 wild-type (WT)]kindly provided by Prof. Alan Porter (Institute of Molecu-lar and Cell Biology, Proteos, Singapore; ref. 16). Trans-fection was carried out using Fugene 6 transfection reagent(Roche Applied Science, Lewes, East Sussex, UK) accord-ing to the manufacturer’s guidelines. After transfection for24 h and subsequent experimental procedures, cells werewashed with 2 ml of 1 � PBS (ice-cold) and lysed in 200 �l1 � luciferase reporter whole-cell lysis buffer (Promega,Southampton, UK). Luciferase activity was quantified usinga luciferin substrate (Promega) and luminometry (JuniorLB 9509, Berthold Technologies, Redbourn, Hertford-shire, UK). All readings were normalized to protein usingthe Bradford method (Bio-Rad).
Liver biopsy extraction and preparation
Liver wedge biopsy was obtained from donor liver at thebeginning of multiorgan donor retrieval before cold perfu-sion and stored at –80°C (retrieval biopsy) as describedpreviously (17). After a period of cold ischemia and reimplan-tation, a second wedge biopsy was performed 1 h after portalreperfusion and stored at –80°C (reperfusion biopsy). RNAwas isolated as described above.
Statistical analysis
All data are presented as mean � sem for n independentexperiments. Statistical significance was evaluated usingANOVA carried out using software package InStat (Graph-Pad, San Diego, CA, USA).
RESULTS
Ischemia-reperfusion injury induced Nrf2-regulatedantioxidant gene expression
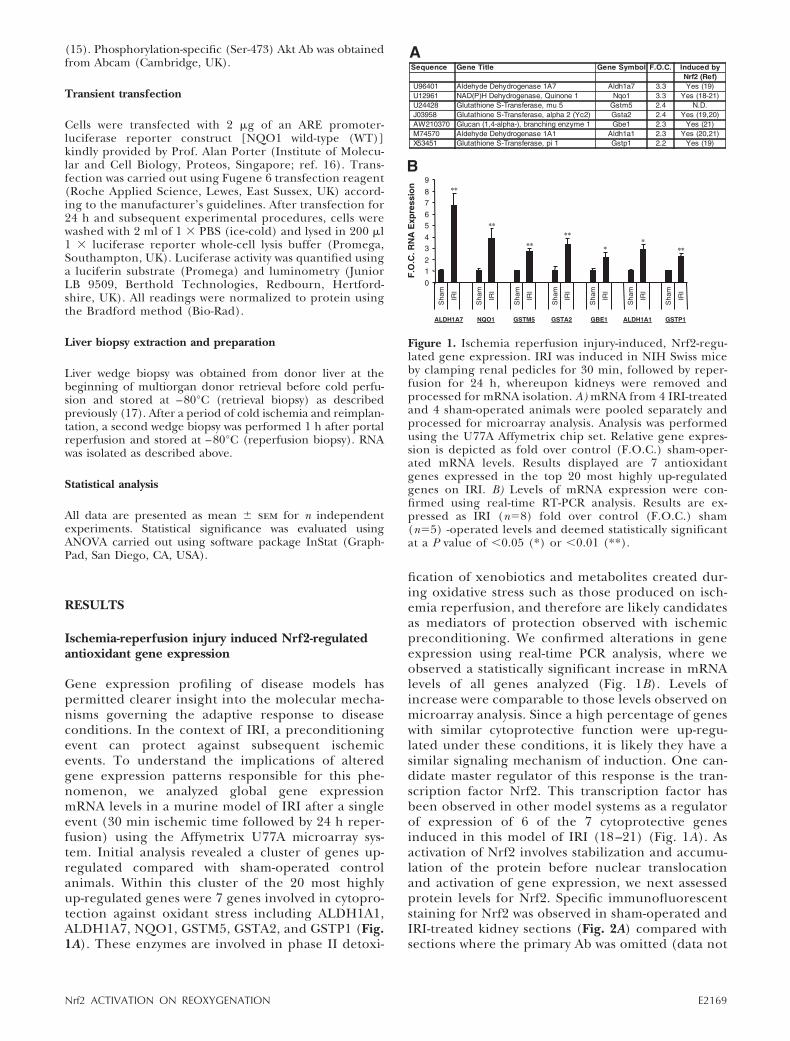
Gene expression profiling of disease models haspermitted clearer insight into the molecular mecha-nisms governing the adaptive response to diseaseconditions. In the context of IRI, a preconditioningevent can protect against subsequent ischemicevents. To understand the implications of alteredgene expression patterns responsible for this phe-nomenon, we analyzed global gene expressionmRNA levels in a murine model of IRI after a singleevent (30 min ischemic time followed by 24 h reper-fusion) using the Affymetrix U77A microarray sys-tem. Initial analysis revealed a cluster of genes up-regulated compared with sham-operated controlanimals. Within this cluster of the 20 most highlyup-regulated genes were 7 genes involved in cytopro-tection against oxidant stress including ALDH1A1,ALDH1A7, NQO1, GSTM5, GSTA2, and GSTP1 (Fig.1A). These enzymes are involved in phase II detoxi-
fication of xenobiotics and metabolites created dur-ing oxidative stress such as those produced on isch-emia reperfusion, and therefore are likely candidatesas mediators of protection observed with ischemicpreconditioning. We confirmed alterations in geneexpression using real-time PCR analysis, where weobserved a statistically significant increase in mRNAlevels of all genes analyzed (Fig. 1B). Levels ofincrease were comparable to those levels observed onmicroarray analysis. Since a high percentage of geneswith similar cytoprotective function were up-regu-lated under these conditions, it is likely they have asimilar signaling mechanism of induction. One can-didate master regulator of this response is the tran-scription factor Nrf2. This transcription factor hasbeen observed in other model systems as a regulatorof expression of 6 of the 7 cytoprotective genesinduced in this model of IRI (18 –21) (Fig. 1A). Asactivation of Nrf2 involves stabilization and accumu-lation of the protein before nuclear translocationand activation of gene expression, we next assessedprotein levels for Nrf2. Specific immunofluorescentstaining for Nrf2 was observed in sham-operated andIRI-treated kidney sections (Fig. 2A) compared withsections where the primary Ab was omitted (data not
Figure 1. Ischemia reperfusion injury-induced, Nrf2-regu-lated gene expression. IRI was induced in NIH Swiss miceby clamping renal pedicles for 30 min, followed by reper-fusion for 24 h, whereupon kidneys were removed andprocessed for mRNA isolation. A) mRNA from 4 IRI-treatedand 4 sham-operated animals were pooled separately andprocessed for microarray analysis. Analysis was performedusing the U77A Affymetrix chip set. Relative gene expres-sion is depicted as fold over control (F.O.C.) sham-oper-ated mRNA levels. Results displayed are 7 antioxidantgenes expressed in the top 20 most highly up-regulatedgenes on IRI. B) Levels of mRNA expression were con-firmed using real-time RT-PCR analysis. Results are ex-pressed as IRI (n�8) fold over control (F.O.C.) sham(n�5) -operated levels and deemed statistically significantat a P value of �0.05 (*) or �0.01 (**).
E2169Nrf2 ACTIVATION ON REOXYGENATION
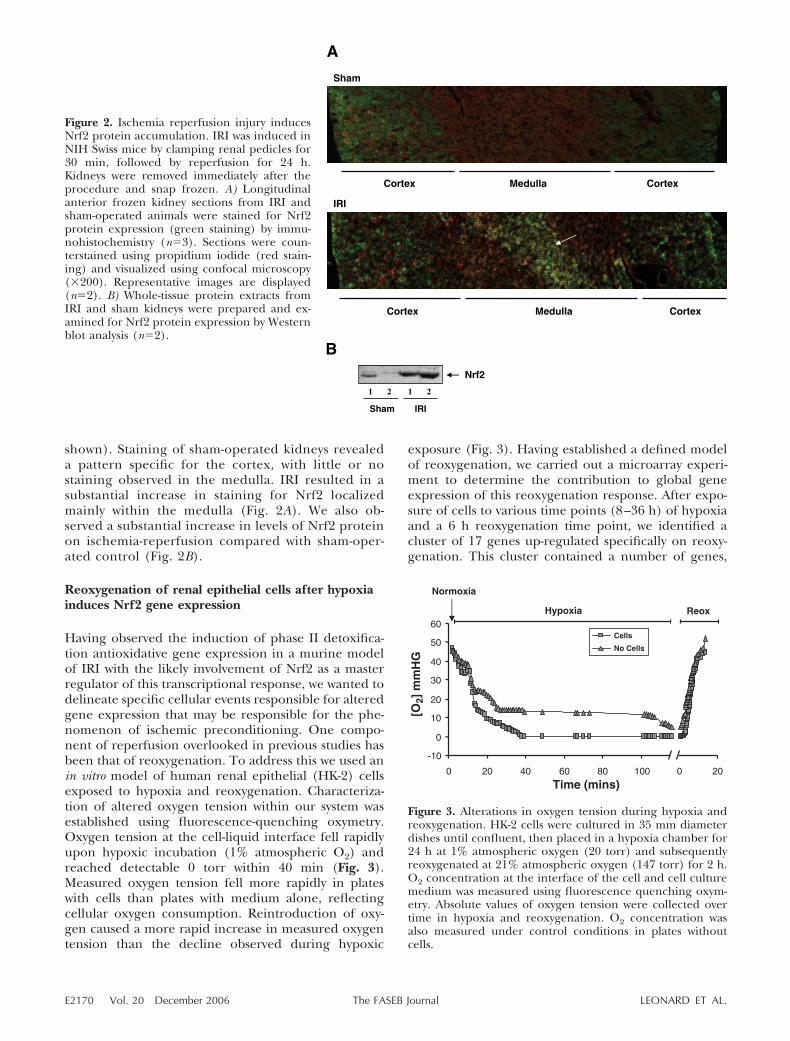
shown). Staining of sham-operated kidneys revealeda pattern specific for the cortex, with little or nostaining observed in the medulla. IRI resulted in asubstantial increase in staining for Nrf2 localizedmainly within the medulla (Fig. 2A). We also ob-served a substantial increase in levels of Nrf2 proteinon ischemia-reperfusion compared with sham-oper-ated control (Fig. 2B).
Reoxygenation of renal epithelial cells after hypoxiainduces Nrf2 gene expression
Having observed the induction of phase II detoxifica-tion antioxidative gene expression in a murine modelof IRI with the likely involvement of Nrf2 as a masterregulator of this transcriptional response, we wanted todelineate specific cellular events responsible for alteredgene expression that may be responsible for the phe-nomenon of ischemic preconditioning. One compo-nent of reperfusion overlooked in previous studies hasbeen that of reoxygenation. To address this we used anin vitro model of human renal epithelial (HK-2) cellsexposed to hypoxia and reoxygenation. Characteriza-tion of altered oxygen tension within our system wasestablished using fluorescence-quenching oxymetry.Oxygen tension at the cell-liquid interface fell rapidlyupon hypoxic incubation (1% atmospheric O2) andreached detectable 0 torr within 40 min (Fig. 3).Measured oxygen tension fell more rapidly in plateswith cells than plates with medium alone, reflectingcellular oxygen consumption. Reintroduction of oxy-gen caused a more rapid increase in measured oxygentension than the decline observed during hypoxic
exposure (Fig. 3). Having established a defined modelof reoxygenation, we carried out a microarray experi-ment to determine the contribution to global geneexpression of this reoxygenation response. After expo-sure of cells to various time points (8–36 h) of hypoxiaand a 6 h reoxygenation time point, we identified acluster of 17 genes up-regulated specifically on reoxy-genation. This cluster contained a number of genes,
Figure 2. Ischemia reperfusion injury inducesNrf2 protein accumulation. IRI was induced inNIH Swiss mice by clamping renal pedicles for30 min, followed by reperfusion for 24 h.Kidneys were removed immediately after theprocedure and snap frozen. A) Longitudinalanterior frozen kidney sections from IRI andsham-operated animals were stained for Nrf2protein expression (green staining) by immu-nohistochemistry (n�3). Sections were coun-terstained using propidium iodide (red stain-ing) and visualized using confocal microscopy(�200). Representative images are displayed(n�2). B) Whole-tissue protein extracts fromIRI and sham kidneys were prepared and ex-amined for Nrf2 protein expression by Westernblot analysis (n�2).
Figure 3. Alterations in oxygen tension during hypoxia andreoxygenation. HK-2 cells were cultured in 35 mm diameterdishes until confluent, then placed in a hypoxia chamber for24 h at 1% atmospheric oxygen (20 torr) and subsequentlyreoxygenated at 21% atmospheric oxygen (147 torr) for 2 h.O2 concentration at the interface of the cell and cell culturemedium was measured using fluorescence quenching oxym-etry. Absolute values of oxygen tension were collected overtime in hypoxia and reoxygenation. O2 concentration wasalso measured under control conditions in plates withoutcells.
E2170 Vol. 20 December 2006 LEONARD ET AL.The FASEB Journal
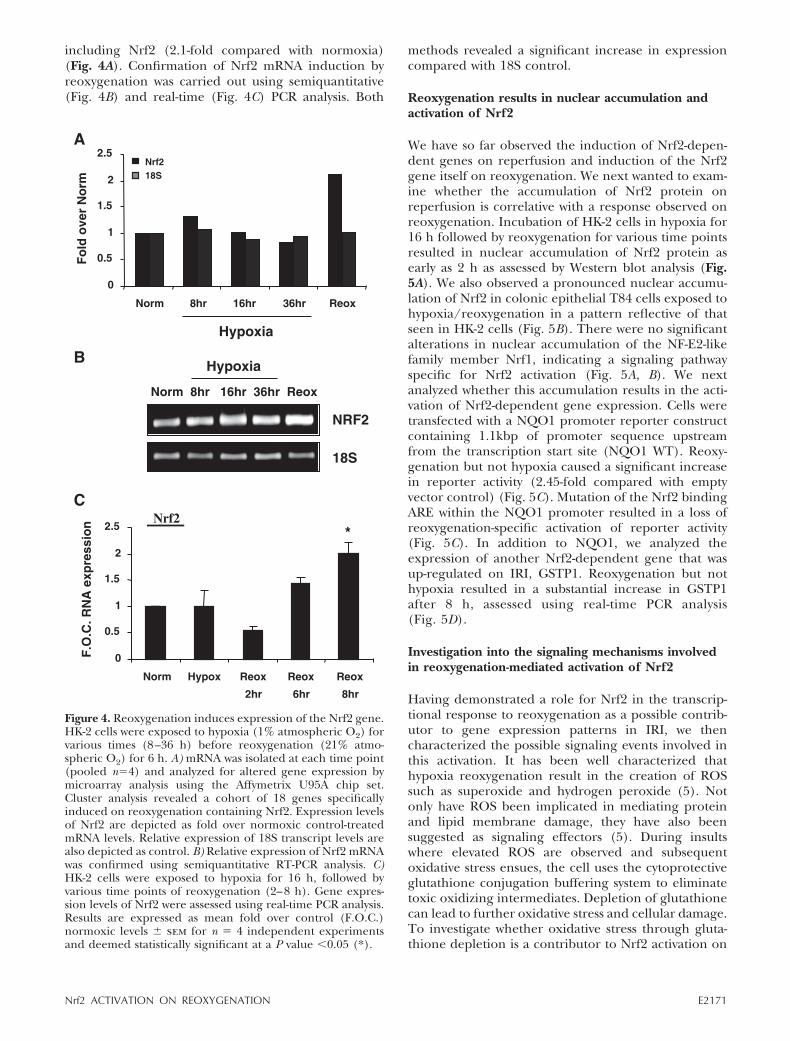
including Nrf2 (2.1-fold compared with normoxia)(Fig. 4A). Confirmation of Nrf2 mRNA induction byreoxygenation was carried out using semiquantitative(Fig. 4B) and real-time (Fig. 4C) PCR analysis. Both
methods revealed a significant increase in expressioncompared with 18S control.
Reoxygenation results in nuclear accumulation andactivation of Nrf2
We have so far observed the induction of Nrf2-depen-dent genes on reperfusion and induction of the Nrf2gene itself on reoxygenation. We next wanted to exam-ine whether the accumulation of Nrf2 protein onreperfusion is correlative with a response observed onreoxygenation. Incubation of HK-2 cells in hypoxia for16 h followed by reoxygenation for various time pointsresulted in nuclear accumulation of Nrf2 protein asearly as 2 h as assessed by Western blot analysis (Fig.5A). We also observed a pronounced nuclear accumu-lation of Nrf2 in colonic epithelial T84 cells exposed tohypoxia/reoxygenation in a pattern reflective of thatseen in HK-2 cells (Fig. 5B). There were no significantalterations in nuclear accumulation of the NF-E2-likefamily member Nrf1, indicating a signaling pathwayspecific for Nrf2 activation (Fig. 5A, B). We nextanalyzed whether this accumulation results in the acti-vation of Nrf2-dependent gene expression. Cells weretransfected with a NQO1 promoter reporter constructcontaining 1.1kbp of promoter sequence upstreamfrom the transcription start site (NQO1 WT). Reoxy-genation but not hypoxia caused a significant increasein reporter activity (2.45-fold compared with emptyvector control) (Fig. 5C). Mutation of the Nrf2 bindingARE within the NQO1 promoter resulted in a loss ofreoxygenation-specific activation of reporter activity(Fig. 5C). In addition to NQO1, we analyzed theexpression of another Nrf2-dependent gene that wasup-regulated on IRI, GSTP1. Reoxygenation but nothypoxia resulted in a substantial increase in GSTP1after 8 h, assessed using real-time PCR analysis(Fig. 5D).
Investigation into the signaling mechanisms involvedin reoxygenation-mediated activation of Nrf2
Having demonstrated a role for Nrf2 in the transcrip-tional response to reoxygenation as a possible contrib-utor to gene expression patterns in IRI, we thencharacterized the possible signaling events involved inthis activation. It has been well characterized thathypoxia reoxygenation result in the creation of ROSsuch as superoxide and hydrogen peroxide (5). Notonly have ROS been implicated in mediating proteinand lipid membrane damage, they have also beensuggested as signaling effectors (5). During insultswhere elevated ROS are observed and subsequentoxidative stress ensues, the cell uses the cytoprotectiveglutathione conjugation buffering system to eliminatetoxic oxidizing intermediates. Depletion of glutathionecan lead to further oxidative stress and cellular damage.To investigate whether oxidative stress through gluta-thione depletion is a contributor to Nrf2 activation on
Figure 4. Reoxygenation induces expression of the Nrf2 gene.HK-2 cells were exposed to hypoxia (1% atmospheric O2) forvarious times (8–36 h) before reoxygenation (21% atmo-spheric O2) for 6 h. A) mRNA was isolated at each time point(pooled n�4) and analyzed for altered gene expression bymicroarray analysis using the Affymetrix U95A chip set.Cluster analysis revealed a cohort of 18 genes specificallyinduced on reoxygenation containing Nrf2. Expression levelsof Nrf2 are depicted as fold over normoxic control-treatedmRNA levels. Relative expression of 18S transcript levels arealso depicted as control. B) Relative expression of Nrf2 mRNAwas confirmed using semiquantitative RT-PCR analysis. C)HK-2 cells were exposed to hypoxia for 16 h, followed byvarious time points of reoxygenation (2–8 h). Gene expres-sion levels of Nrf2 were assessed using real-time PCR analysis.Results are expressed as mean fold over control (F.O.C.)normoxic levels � sem for n � 4 independent experimentsand deemed statistically significant at a P value �0.05 (*).
E2171Nrf2 ACTIVATION ON REOXYGENATION
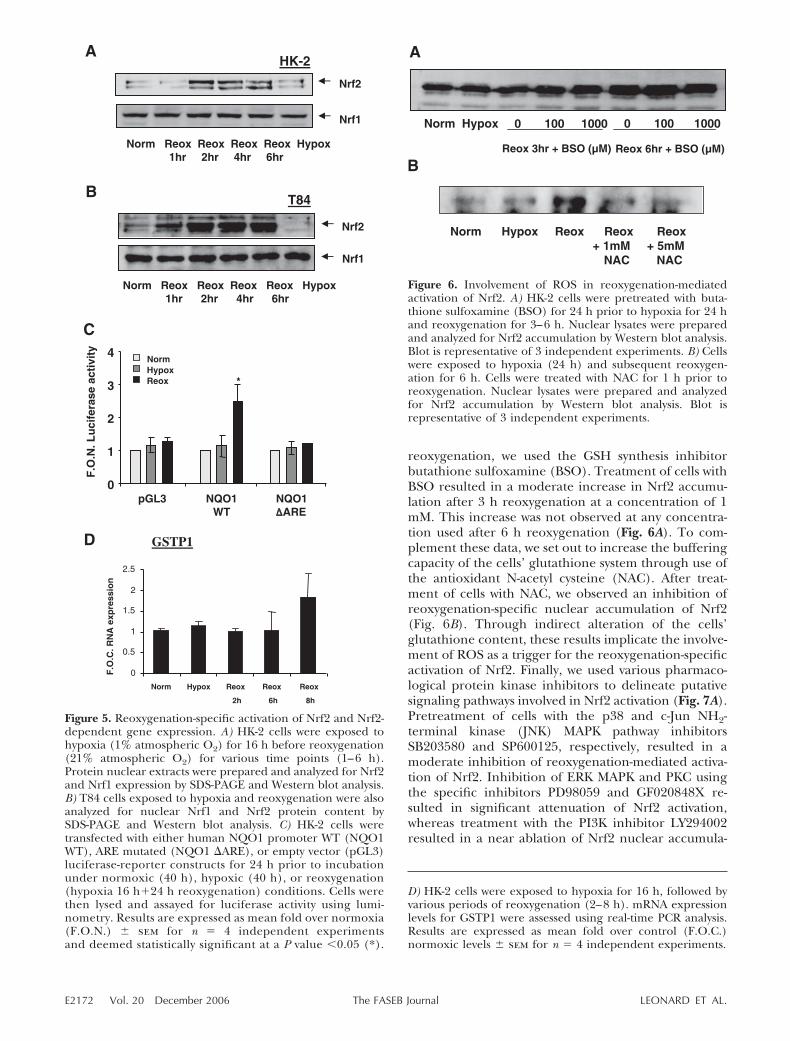
reoxygenation, we used the GSH synthesis inhibitorbutathione sulfoxamine (BSO). Treatment of cells withBSO resulted in a moderate increase in Nrf2 accumu-lation after 3 h reoxygenation at a concentration of 1mM. This increase was not observed at any concentra-tion used after 6 h reoxygenation (Fig. 6A). To com-plement these data, we set out to increase the bufferingcapacity of the cells’ glutathione system through use ofthe antioxidant N-acetyl cysteine (NAC). After treat-ment of cells with NAC, we observed an inhibition ofreoxygenation-specific nuclear accumulation of Nrf2(Fig. 6B). Through indirect alteration of the cells’glutathione content, these results implicate the involve-ment of ROS as a trigger for the reoxygenation-specificactivation of Nrf2. Finally, we used various pharmaco-logical protein kinase inhibitors to delineate putativesignaling pathways involved in Nrf2 activation (Fig. 7A).Pretreatment of cells with the p38 and c-Jun NH2-terminal kinase (JNK) MAPK pathway inhibitorsSB203580 and SP600125, respectively, resulted in amoderate inhibition of reoxygenation-mediated activa-tion of Nrf2. Inhibition of ERK MAPK and PKC usingthe specific inhibitors PD98059 and GF020848X re-sulted in significant attenuation of Nrf2 activation,whereas treatment with the PI3K inhibitor LY294002resulted in a near ablation of Nrf2 nuclear accumula-
Figure 5. Reoxygenation-specific activation of Nrf2 and Nrf2-dependent gene expression. A) HK-2 cells were exposed tohypoxia (1% atmospheric O2) for 16 h before reoxygenation(21% atmospheric O2) for various time points (1–6 h).Protein nuclear extracts were prepared and analyzed for Nrf2and Nrf1 expression by SDS-PAGE and Western blot analysis.B) T84 cells exposed to hypoxia and reoxygenation were alsoanalyzed for nuclear Nrf1 and Nrf2 protein content bySDS-PAGE and Western blot analysis. C) HK-2 cells weretransfected with either human NQO1 promoter WT (NQO1WT), ARE mutated (NQO1 �ARE), or empty vector (pGL3)luciferase-reporter constructs for 24 h prior to incubationunder normoxic (40 h), hypoxic (40 h), or reoxygenation(hypoxia 16 h�24 h reoxygenation) conditions. Cells werethen lysed and assayed for luciferase activity using lumi-nometry. Results are expressed as mean fold over normoxia(F.O.N.) � sem for n � 4 independent experimentsand deemed statistically significant at a P value �0.05 (*).
D) HK-2 cells were exposed to hypoxia for 16 h, followed byvarious periods of reoxygenation (2–8 h). mRNA expressionlevels for GSTP1 were assessed using real-time PCR analysis.Results are expressed as mean fold over control (F.O.C.)normoxic levels � sem for n � 4 independent experiments.
Figure 6. Involvement of ROS in reoxygenation-mediatedactivation of Nrf2. A) HK-2 cells were pretreated with buta-thione sulfoxamine (BSO) for 24 h prior to hypoxia for 24 hand reoxygenation for 3–6 h. Nuclear lysates were preparedand analyzed for Nrf2 accumulation by Western blot analysis.Blot is representative of 3 independent experiments. B) Cellswere exposed to hypoxia (24 h) and subsequent reoxygen-ation for 6 h. Cells were treated with NAC for 1 h prior toreoxygenation. Nuclear lysates were prepared and analyzedfor Nrf2 accumulation by Western blot analysis. Blot isrepresentative of 3 independent experiments.
E2172 Vol. 20 December 2006 LEONARD ET AL.The FASEB Journal
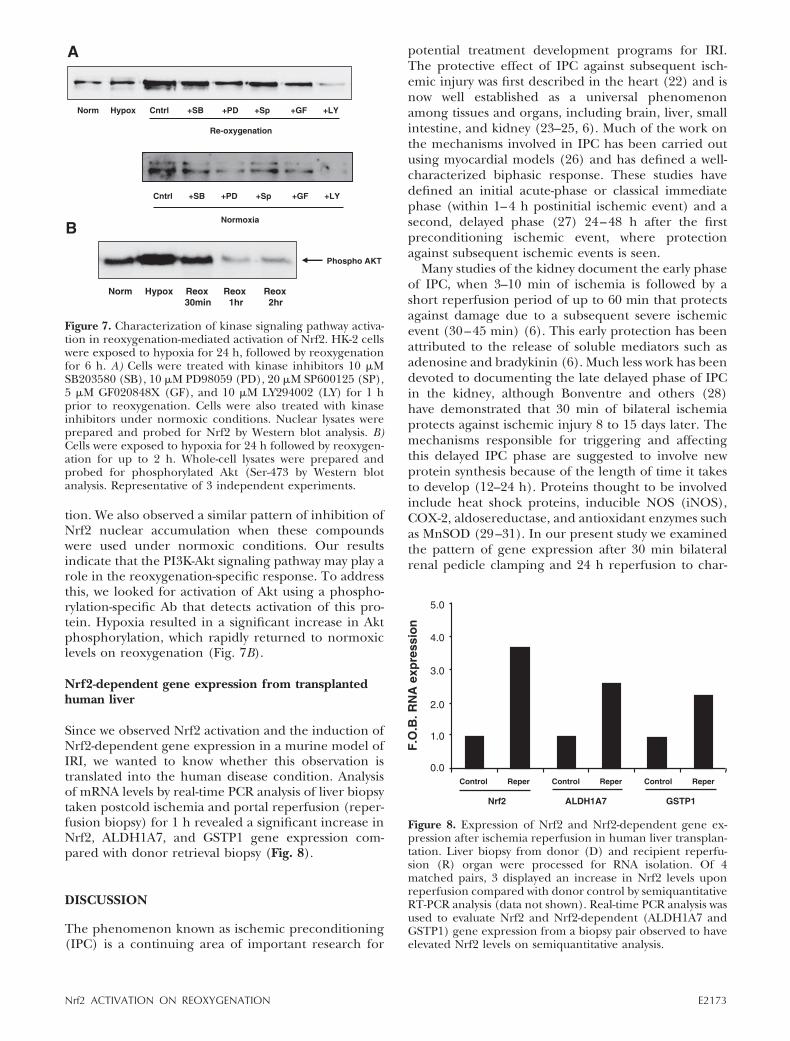
tion. We also observed a similar pattern of inhibition ofNrf2 nuclear accumulation when these compoundswere used under normoxic conditions. Our resultsindicate that the PI3K-Akt signaling pathway may play arole in the reoxygenation-specific response. To addressthis, we looked for activation of Akt using a phospho-rylation-specific Ab that detects activation of this pro-tein. Hypoxia resulted in a significant increase in Aktphosphorylation, which rapidly returned to normoxiclevels on reoxygenation (Fig. 7B).
Nrf2-dependent gene expression from transplantedhuman liver
Since we observed Nrf2 activation and the induction ofNrf2-dependent gene expression in a murine model ofIRI, we wanted to know whether this observation istranslated into the human disease condition. Analysisof mRNA levels by real-time PCR analysis of liver biopsytaken postcold ischemia and portal reperfusion (reper-fusion biopsy) for 1 h revealed a significant increase inNrf2, ALDH1A7, and GSTP1 gene expression com-pared with donor retrieval biopsy (Fig. 8).
DISCUSSION
The phenomenon known as ischemic preconditioning(IPC) is a continuing area of important research for
potential treatment development programs for IRI.The protective effect of IPC against subsequent isch-emic injury was first described in the heart (22) and isnow well established as a universal phenomenonamong tissues and organs, including brain, liver, smallintestine, and kidney (23–25, 6). Much of the work onthe mechanisms involved in IPC has been carried outusing myocardial models (26) and has defined a well-characterized biphasic response. These studies havedefined an initial acute-phase or classical immediatephase (within 1–4 h postinitial ischemic event) and asecond, delayed phase (27) 24–48 h after the firstpreconditioning ischemic event, where protectionagainst subsequent ischemic events is seen.
Many studies of the kidney document the early phaseof IPC, when 3–10 min of ischemia is followed by ashort reperfusion period of up to 60 min that protectsagainst damage due to a subsequent severe ischemicevent (30–45 min) (6). This early protection has beenattributed to the release of soluble mediators such asadenosine and bradykinin (6). Much less work has beendevoted to documenting the late delayed phase of IPCin the kidney, although Bonventre and others (28)have demonstrated that 30 min of bilateral ischemiaprotects against ischemic injury 8 to 15 days later. Themechanisms responsible for triggering and affectingthis delayed IPC phase are suggested to involve newprotein synthesis because of the length of time it takesto develop (12–24 h). Proteins thought to be involvedinclude heat shock proteins, inducible NOS (iNOS),COX-2, aldosereductase, and antioxidant enzymes suchas MnSOD (29–31). In our present study we examinedthe pattern of gene expression after 30 min bilateralrenal pedicle clamping and 24 h reperfusion to char-
Figure 8. Expression of Nrf2 and Nrf2-dependent gene ex-pression after ischemia reperfusion in human liver transplan-tation. Liver biopsy from donor (D) and recipient reperfu-sion (R) organ were processed for RNA isolation. Of 4matched pairs, 3 displayed an increase in Nrf2 levels uponreperfusion compared with donor control by semiquantitativeRT-PCR analysis (data not shown). Real-time PCR analysis wasused to evaluate Nrf2 and Nrf2-dependent (ALDH1A7 andGSTP1) gene expression from a biopsy pair observed to haveelevated Nrf2 levels on semiquantitative analysis.
Figure 7. Characterization of kinase signaling pathway activa-tion in reoxygenation-mediated activation of Nrf2. HK-2 cellswere exposed to hypoxia for 24 h, followed by reoxygenationfor 6 h. A) Cells were treated with kinase inhibitors 10 �MSB203580 (SB), 10 �M PD98059 (PD), 20 �M SP600125 (SP),5 �M GF020848X (GF), and 10 �M LY294002 (LY) for 1 hprior to reoxygenation. Cells were also treated with kinaseinhibitors under normoxic conditions. Nuclear lysates wereprepared and probed for Nrf2 by Western blot analysis. B)Cells were exposed to hypoxia for 24 h followed by reoxygen-ation for up to 2 h. Whole-cell lysates were prepared andprobed for phosphorylated Akt (Ser-473 by Western blotanalysis. Representative of 3 independent experiments.
E2173Nrf2 ACTIVATION ON REOXYGENATION

acterize putative effectors of this preconditioning effectobserved at later time points.
Analysis of microarray data revealed the induction ofa group of 7 phase II detoxification and antioxidantgenes all within the top 20 most highly up-regulated,including aldehyde dehydrogenases, glutathione Stransferases, and NQO1. The high incidence of antiox-idant gene expression suggests a highly specific andimportant coordinated response to protect the cellagainst ongoing or future oxidative stress. This coordi-nated response likely involves Nrf2, as this transcriptionfactor has been identified as an activator of theseantioxidant genes in other model systems. Evidence tosupport this was demonstrated through the accumula-tion of Nrf2 protein in ischemia-reperfused kidneycompared with sham-operated controls. We also ob-served specific medullary staining for Nrf2 within thekidney on IRI. Further analysis of the specific cell typeor types within the medulla responsible for this stainingwill reveal greater insight into the mechanisms of Nrf2activation and its importance in protecting againstoxidant stress within the kidney.
Though much work has established the basic patho-logical characteristics of IRI and delayed ischemicpreconditioning, the precise signaling mechanisms re-sponsible remain unclear. It was recently demonstratedthat ROS are generated during IRI (5). During isch-emia-reperfusion, increased intracellular calcium levelsare considered to contribute to cellular injury and celldeath through an alteration in mitochondrial respira-tory function causing an increase in damaging ROS,including superoxide anion radical (O2), the hy-droxyl radical (OH�), hydrogen peroxide (H2O2), andperoxynitrate (ONOO). These ROS have also beenpostulated as cytoprotective signaling intermediates forpreconditioning, although downstream targets havenot been extensively investigated to date. The rise inlevels of ROS on ischemia/reperfusion appears to beessential for the development of delayed IPC (32–34).An important overlooked source of ROS on ischemiareperfusion involves those produced upon reoxygen-ation. Hypoxia causes accumulation of hypoxanthine,which on reoxygenation forms O2– through the actionof xanthine oxidase. The spontaneous production ofhydroxyl radical forms from H2O2 can also occurthrough the action of free Fe2� (Fenton reaction)(35–36). The generation of ROS on reoxygenation hasbeen demonstrated in various model systems, includingHUVECs through mitochondrial sources (37).
It has been widely demonstrated that Nrf2 activationoccurs due to an alteration in the redox state of the celldue to the presence of increased amounts of electro-philes or ROS. In the context of our current work, weidentified Nrf2 in a model of hypoxia/reoxygenation ofrenal epithelial cells within a cluster genes up-regulatedspecifically on reoxygenation through microarray anal-ysis. We postulate that this reoxygenation-specific up-regulation is a mechanism through which cells activatean antioxidant response to protect themselves from
future oxidant damage. It has been demonstrated thatNrf2 activation induces the expression of the Nrf2 geneitself (38). We also demonstrated a rapid nuclearaccumulation of Nrf2 on reoxygenation and the induc-tion of promoter reporter activity of one of the identi-fied antioxidant genes, NQO1, which was abolished ondeletion of the ARE sequence. We also observed areoxygenation-specific up-regulation of the Nrf2-de-pendent gene GSTP1. All these data suggest that reoxy-genation-specific activation of Nrf2 results in accumu-lation and transactivation of Nrf2-dependent Phase IIdetoxification and antioxidant gene expression, includ-ing Nrf2 itself, and this is a vital mechanism throughwhich these genes are up-regulated in IRI to protect thecell from further damage. We also suggest this as acontributory mechanism through which the develop-ment of ischemic preconditioning occurs. Activation ofNrf2 has been observed in response to different ROS,and in our current study we present evidence to sup-port that reoxygenation-specific activation of Nrf2 ismediated through ROS generation. This was evidencedthrough the use of the antioxidant NAC, which resultedin an inhibition of reoxygenation-specific activation ofNrf2. Additional evidence was generated through mod-ification of the cells redox status using the glutathionesynthesis inhibitor BSO, resulting in further activationof Nrf2 on reoxygenation.
To investigate other possible signaling events in-volved in the activation of Nrf2 on reoxygenation, weused an array of protein kinase inhibitors to the MAPKsp38, JNK, ERK, PI3kinase, and PKC. We observedsubstantial inhibition of reoxygenation-stimulated Nrf2accumulation using the PKC and ERK MAPK pathwayinhibitors and an abolition of activation using thePI3kinase pathway inhibitor LY294002. We observed amodest inhibition using p38 and JNK MAPK inhibitors.These results are interesting in the context that all ofthese pathways are activated in various models of IRIand hypoxia/reoxygenation. Indeed, the PI3K-Akt andERK MAPK pathways are recruited in the setting of IPC(39–40). Inhibition of either PI3K or ERK kinasecascades abolished the protective effect of precondi-tioning to a subsequent ischemic event in these models.In the heart, inhibition of PKC prevented the protec-tion seen with ischemic preconditioning (41). Theinvolvement of PKC may not be all that clear; forexample, PKC activation during ischemia is reported tobe harmful and the specific actions of PKC-epsilon arereported to be inhibited by potent inhibitors of Ca2�-dependent PKC inhibitors such as GF109203X. There-fore, inhibition of Nrf2 accumulation observed in ourmodel system using GF109203X could be due to mul-tiple PKC isoforms, not just PKC. There is compellingevidence for the involvement of these signaling cas-cades as specific triggers or mediators for reoxygen-ation and reperfusion-stimulated activation of Nrf2.However, treatment of cells with these kinase inhibitorsunder normoxic conditions resulted in a nearly identi-cal pattern of inhibition of Nrf2 expression, as was
E2174 Vol. 20 December 2006 LEONARD ET AL.The FASEB Journal

observed under reoxygenation. This result indicatesthat these signaling pathways are likely necessary forbasal expression of Nrf2 but are not the reoxygenation-specific trigger. This is further emphasized by the lackof reoxygenation-specific activation of Akt in thesecells. It is therefore likely that some as yet undefinedsignaling cascade specifically activated on reoxygen-ation likely involving ROS is responsible for Nrf2 acti-vation.
It has been put forward as to whether IPC actuallyoccurs in humans. Some studies have been carried out,for example, using a human forearm model, wherepreconditioning 3 � 5 min ischemia before subsequent20 min ischemia protects against endothelial dysfunc-tion and neutrophil activation (42). There has alsobeen evidence in the heart for the existence of IPC. Ithas been observed that patients with preinfarctionangina have smaller infarct sizes, better functionalrecovery, and better prognosis, which the authors havesuggested as a functional readout of ischemic precon-ditioning. We have demonstrated an up-regulation ofNrf2 and Nrf2-dependent antioxidant gene expressionin ischemic reperfused human liver biopsies pre- andpost-transplant, which we suggest as a mechanism ofprotection against current and subsequent oxidantdamage as would occur with future ischemic events.
In summary we have demonstrated for the first timethe induction of Nrf2 activation and Nrf2-dependentantioxidant gene expression in an in vivo model of IRI.We also demonstrate for the first time a reoxygenation-specific activation of Nrf2 and Nrf2-dependent antiox-idant gene expression in a renal epithelial cell in vitromodel. We therefore postulate that reoxygenation-specific activation of the Nrf2 antioxidant pathway as acontributory mechanism to the adaptive cytoprotectiveresponse to ongoing and subsequent oxidant damagein IRI. We also present evidence for the involvement ofROS in the reoxygenation-specific activation of Nrf2.Further investigation into the signaling pathways in-volved in this response may help our understanding ofischemic preconditioning toward future therapeuticpotential.
We acknowledge technical assistance from Annemarie Grif-fin. This work was supported by grants from the HealthResearch Board of Ireland, the Wellcome Trust (to C.T.T.),the Science Foundation of Ireland (to C.T.T.), and from theNational Institutes of Health (M.J.B. and H.R.: N100K R01and NH/LBI Lung Injury SCCOR).
REFERENCES
1. Moens, A. L., Claeys, M. J., Timmermans, J. P., and Vrints, C. J.(2005) Myocardial ischemia/reperfusion-injury, a clinical viewon a complex pathophysiological process. Int. J. Cardiol. 100,179–190
2. Zheng, Z., Lee, J. E., and Yenari, M. A. (2003) Stroke: molecularmechanisms and potential targets for treatment. Curr. Mol. Med.3, 361–372
3. Rabb, H., O’Meara, Y. M., Maderna, P., Coleman, P., and Brady,H. R. (1997) Leukocytes, cell adhesion molecules and ischemicacute renal failure. Kidney Int. 51, 1463–1468
4. Husted, T. L., and Lentsch, A. B. (2005) Anti-inflammatoryapproaches to the prevention of ischemia/reperfusion injuryin solid organ transplantation. Curr. Opin. Invest. Drugs. 6,508 –512
5. Li, C., and Jackson, R. M. (2002) Reactive species mechanisms ofcellular hypoxia-reoxygenation injury. Am. J. Physiol. 282, C227–C241
6. Bonventre, J. V. (2002) Kidney ischemic preconditioning. Curr.Opin. Nephrol. Hypertens. 11, 43–48
7. Mathers, J., Fraser, J. A., McMahon, M., Saunders, R. D., Hayes,J. D., and McLellan, L. I. (2004) Antioxidant and cytoprotectiveresponses to redox stress. Biochem. Soc Symp. 71, 157–176
8. Jaiswal, A. K. (2004) Nrf2 signaling in coordinated activation ofantioxidant gene expression. Free Radic. Biol. Med. 36, 1199–1207
9. Kobayashi, M., and Yamamoto, M. (2005) Molecular mecha-nisms activating the Nrf2-Keap1 pathway of antioxidant generegulation. Antioxid. Redox Signal. 7, 385–394
10. Leonard, M. O., Hannan, K., Burne, M. J., Lappin, D. W.,Doran, P., Coleman, P., Stenson, C., Taylor, C. T., Daniels, F.,Godson, C., et al. (2002) 15-Epi-16-(para-fluorophenoxy)-li-poxin A(4)-methyl ester, a synthetic analogue of 15-epi-lipoxinA(4), is protective in experimental ischemic acute renal failure.J. Am. Soc. Nephrol. 13, 1657–1662
11. Ryan, M. J., Johnson, G., Kirk, J., Fuerstenberg, S. M., Zager,R. A., and Torok-Storb, B. (1994) HK-2: an immortalizedproximal tubule epithelial cell line from normal adult humankidney. Kidney Int. 45, 48–57
12. Griffiths, J. R., and Robinson, S. P. (1999) The OxyLite: afibre-optic oxygen sensor. Br. J. Radiol. 72, 627–630
13. Leonard, M. O., O’Reilly, S., Comerford, K M, and Taylor, C. T.(2004) Identification of cyclic AMP response element-bindingprotein-dependent transcriptional responses in hypoxia by mi-croarray analysis. Methods Enzymol. 381, 511–524
14. Leonard, M. O., Cottell, D. C., Godson, C., Brady, H. R., andTaylor, C. T. (2003) The role of HIF-1 alpha in transcriptionalregulation of the proximal tubular epithelial cell response tohypoxia. J. Biol. Chem. 278, 40296–40304
15. Leonard, M. O., Godson, C., Brady, H. R., and Taylor, C. T.(2005) Potentiation of glucocorticoid activity in hypoxiathrough induction of the glucocorticoid receptor. J. Immunol.174, 2250–2257
16. Dhakshinamoorthy, S., and Porter, A. G. (2004) Nitric oxide-induced transcriptional up-regulation of protective genes byNrf2 via the antioxidant response element counteracts apoptosisof neuroblastoma cells. J. Biol. Chem. 279, 20096–20107
17. Varadarajan, R., Golden-Mason, L., Young, L., McLoughlin, P.,Nolan, N., McEntee, G., Traynor, O., Geoghegan, J., Hegarty,J. E., and O’Farrelly, C. (2004) Nitric oxide in early ischaemiareperfusion injury during human orthotopic liver transplanta-tion. Transplantation 78, 250–256
18. Lee, J. M., and Johnson, J. A. (2004) An important role ofNrf2-ARE pathway in the cellular defense mechanism. J. Bio-chem. Mol. Biol. 37, 139–143
19. Thimmulappa, R. K., Mai, K. H., Srisuma, S., Kensler, T. W.,Yamamoto, M., and Biswal, S. (2002) Identification of Nrf2-regulated genes induced by the chemopreventive agent sul-foraphane by oligonucleotide microarray. Cancer Res. 62,5196 –5203
20. Kwak, M. K., Wakabayashi, N., Itoh, K., Motohashi, H.,Yamamoto, M., and Kensler, T. W. (2003) Modulation of geneexpression by cancer chemopreventive dithiolethiones throughthe Keap1-Nrf2 pathway. Identification of novel gene clustersfor cell survival. J. Biol. Chem. 278, 8135–8145
21. Lee, J. M., Calkins, M. J., Chan, K, Kan, Y. W., Johnson, J. A.(2003) Identification of the NF-E2-related factor-2-dependentgenes conferring protection against oxidative stress in primarycortical astrocytes using oligonucleotide microarray analysis.J. Biol. Chem. 278, 12029–12038
22. Murry, C. E., Jennings, R. B., and Reimer, K. A. (1986) Precon-ditioning with ischemia: a delay of lethal cell injury in ischemicmyocardium. Circulation 74, 1124–1136
E2175Nrf2 ACTIVATION ON REOXYGENATION

23. Glazier, S. S., O’Rourke, D. M., Graham, D. I., and Welsh, F. A.(1994) Induction of ischemic tolerance following brief focalischemia in rat brain. J. Cereb. Blood Flow Metab. 14, 545–553
24. Peralta, C., Closa, D., Hotter, G., Gelpi, E., Prats, N., andRosello-Catafau, J. (1996) Liver ischemic preconditioning ismediated by the inhibitory action of nitric oxide on endothelin.Biochem. Biophys. Res. Commun. 229, 264–270
25. Hotter, G., Closa, D., Prados, M., Fernandez-Cruz, L., Prats, N.,Gelpi, E., and Rosello-Catafau, J. (1996) Intestinal precondition-ing is mediated by a transient increase in nitric oxide. Biochem.Biophys. Res. Commun. 222, 27–32
26. Eisen, A., Fisman, E. Z., Rubenfire, M., Freimark, D., McKech-nie, R., Tenenbaum, A., Motro, M., and Adler, Y. (2004)Ischemic preconditioning: nearly two decades of research. Acomprehensive review. Atherosclerosis 172, 201–210
27. Yellon, D. M., and Baxter, G. F. (1995) A “second window ofprotection” or delayed preconditioning phenomenon: futurehorizons for myocardial protection? J. Mol. Cell Cardiol. 27,1023–1034
28. Park, K. M., Byun, J. Y., Kramers, C., Kim, J. I., Huang, P. L., andBonventre, J. V. (2003) Inducible nitric-oxide synthase is animportant contributor to prolonged protective effects of isch-emic preconditioning in the mouse kidney. J. Biol. Chem. 278,27256–27266
29. Nayeem, M. A., Hess, M. L., Qian, Y. Z., Loesser, K. E., andKukreja, R. C. (1997) Delayed preconditioning of cultured adultrat cardiac myocytes: role of 70- and 90-kDa heat stress proteins.Am. J. Physiol. 273, H861–H868
30. Cutrn, J. C., Perrelli, M. G., Cavalieri, B., Peralta, C., RosellCatafau, J, and Poli, G. (2002) Microvascular dysfunction in-duced by reperfusion injury and protective effect of ischemicpreconditioning. Free Radic. Biol. Med. 33, 1200–1208
31. Bolli, R. (2000) The late phase of preconditioning. Circ. Res. 87,972
32. Kaeffer, N., Richard, V., and Thuillez, C. (1997) Delayedcoronary endothelial protection 24 hours after preconditioning:role of free radicals. Circulation 96, 2311–2316
33. Tang, X. L., Takano, H., Rizvi, A., Turrens, J. F., Qiu, Y., Wu,W. J., Zhang, Q., and Bolli, R. (2002) Oxidant species trigger
late preconditioning against myocardial stunning in consciousrabbits. Am. J. Physiol. 282, H281–H291
34. Sun, J. Z., Tang, X. L., Park, S. W., Qiu, Y., Turrens, J. F., andBolli, R. (1996) Evidence for an essential role of reactive oxygenspecies in the genesis of late preconditioning against myocardialstunning in conscious pigs. J. Clin. Invest. 97, 562–576
35. Weinberg, J. M. (1991) The cell biology of ischemic renal injury.Kidney Int. 39, 476–500
36. Fleury, C., Mignotte, B., and Vayssiere, J. L. (2002) Mitochon-drial reactive oxygen species in cell death signaling. Biochimie(Paris) 84, 131–141
37. Therade-Matharan, S., Laemmel, E., Duranteau, J., and Vicaut,E. (2004) Reoxygenation after hypoxia and glucose depletioncauses reactive oxygen species production by mitochondria inHUVEC. Am. J. Physiol. 287, R1037–R1043
38. Lee, J. M., Li, J., Johnson, D. A., Stein, T. D., Kraft, A. D.,Calkins, M. J., Jakel, R. J., and Johnson, J. A. (2005) Nrf2, amulti-organ protector? FASEB J. 19, 1061–1066
39. Tong, H., Chen, W., Steenbergen, C., and Murphy, E. (2000)Ischemic preconditioning activates phosphatidylinositol-3-ki-nase upstream of protein kinase C. Circ. Res. 87, 309–315
40. Mocanu, M. M., Bell, .R M., and Yellon, D. M. (2002) PI3 kinaseand not p42/p44 appears to be implicated in the protectionconferred by ischemic preconditioning. J. Mol. Cell Cardiol. 34,661–668
41. Node, K, Kitakaze, M., Sato, H., Minamino, T., Komamura, K.,Shinozaki, Y., Mori, H., and Hori, M. (1997) Role of intracellu-lar Ca2� in activation of protein kinase C during ischemicpreconditioning. Circulation 96, 1257–1265
42. Kharbanda, R. K., Peters, M., Walton, B., Kattenhorn, M.,Mullen, M., Klein, N., Vallance, P., Deanfield, J., and MacAl-lister, R. (2001) Ischemic preconditioning prevents endothe-lial injury and systemic neutrophil activation during isch-emia-reperfusion in humans in vivo. Circulation 103, 1624 –
1630
Received for publication November 29, 2005.Accepted for publication August 7, 2006.
E2176 Vol. 20 December 2006 LEONARD ET AL.The FASEB Journal
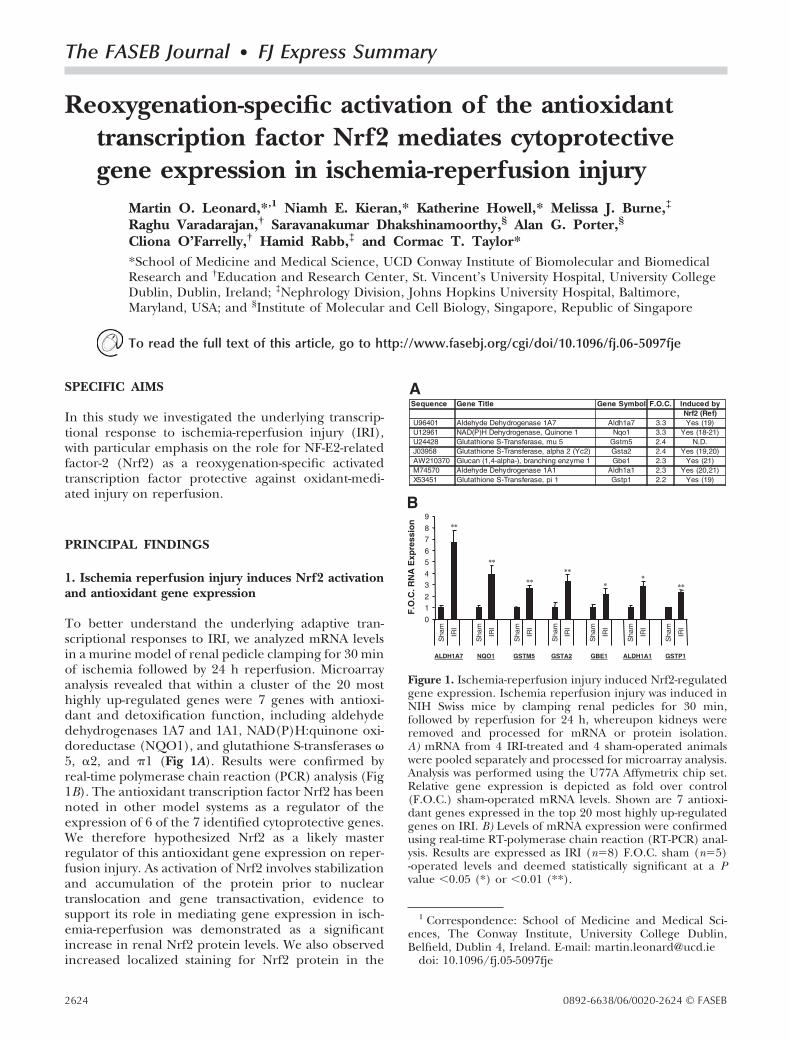
The FASEB Journal • FJ Express Summary
Reoxygenation-specific activation of the antioxidanttranscription factor Nrf2 mediates cytoprotectivegene expression in ischemia-reperfusion injury
Martin O. Leonard,*,1 Niamh E. Kieran,* Katherine Howell,* Melissa J. Burne,‡
Raghu Varadarajan,† Saravanakumar Dhakshinamoorthy,§ Alan G. Porter,§
Cliona O’Farrelly,† Hamid Rabb,‡ and Cormac T. Taylor**School of Medicine and Medical Science, UCD Conway Institute of Biomolecular and BiomedicalResearch and †Education and Research Center, St. Vincent’s University Hospital, University CollegeDublin, Dublin, Ireland; ‡Nephrology Division, Johns Hopkins University Hospital, Baltimore,Maryland, USA; and §Institute of Molecular and Cell Biology, Singapore, Republic of Singapore
To read the full text of this article, go to http://www.fasebj.org/cgi/doi/10.1096/fj.06-5097fje
SPECIFIC AIMS
In this study we investigated the underlying transcrip-tional response to ischemia-reperfusion injury (IRI),with particular emphasis on the role for NF-E2-relatedfactor-2 (Nrf2) as a reoxygenation-specific activatedtranscription factor protective against oxidant-medi-ated injury on reperfusion.
PRINCIPAL FINDINGS
1. Ischemia reperfusion injury induces Nrf2 activationand antioxidant gene expression
To better understand the underlying adaptive tran-scriptional responses to IRI, we analyzed mRNA levelsin a murine model of renal pedicle clamping for 30 minof ischemia followed by 24 h reperfusion. Microarrayanalysis revealed that within a cluster of the 20 mosthighly up-regulated genes were 7 genes with antioxi-dant and detoxification function, including aldehydedehydrogenases 1A7 and 1A1, NAD(P)H:quinone oxi-doreductase (NQO1), and glutathione S-transferases �5, �2, and �1 (Fig 1A). Results were confirmed byreal-time polymerase chain reaction (PCR) analysis (Fig1B). The antioxidant transcription factor Nrf2 has beennoted in other model systems as a regulator of theexpression of 6 of the 7 identified cytoprotective genes.We therefore hypothesized Nrf2 as a likely masterregulator of this antioxidant gene expression on reper-fusion injury. As activation of Nrf2 involves stabilizationand accumulation of the protein prior to nucleartranslocation and gene transactivation, evidence tosupport its role in mediating gene expression in isch-emia-reperfusion was demonstrated as a significantincrease in renal Nrf2 protein levels. We also observedincreased localized staining for Nrf2 protein in the
1 Correspondence: School of Medicine and Medical Sci-ences, The Conway Institute, University College Dublin,Belfield, Dublin 4, Ireland. E-mail: [email protected]
doi: 10.1096/fj.05-5097fje
Figure 1. Ischemia-reperfusion injury induced Nrf2-regulatedgene expression. Ischemia reperfusion injury was induced inNIH Swiss mice by clamping renal pedicles for 30 min,followed by reperfusion for 24 h, whereupon kidneys wereremoved and processed for mRNA or protein isolation.A) mRNA from 4 IRI-treated and 4 sham-operated animalswere pooled separately and processed for microarray analysis.Analysis was performed using the U77A Affymetrix chip set.Relative gene expression is depicted as fold over control(F.O.C.) sham-operated mRNA levels. Shown are 7 antioxi-dant genes expressed in the top 20 most highly up-regulatedgenes on IRI. B) Levels of mRNA expression were confirmedusing real-time RT-polymerase chain reaction (RT-PCR) anal-ysis. Results are expressed as IRI (n�8) F.O.C. sham (n�5)-operated levels and deemed statistically significant at a Pvalue �0.05 (*) or �0.01 (**).
2624 0892-6638/06/0020-2624 © FASEB
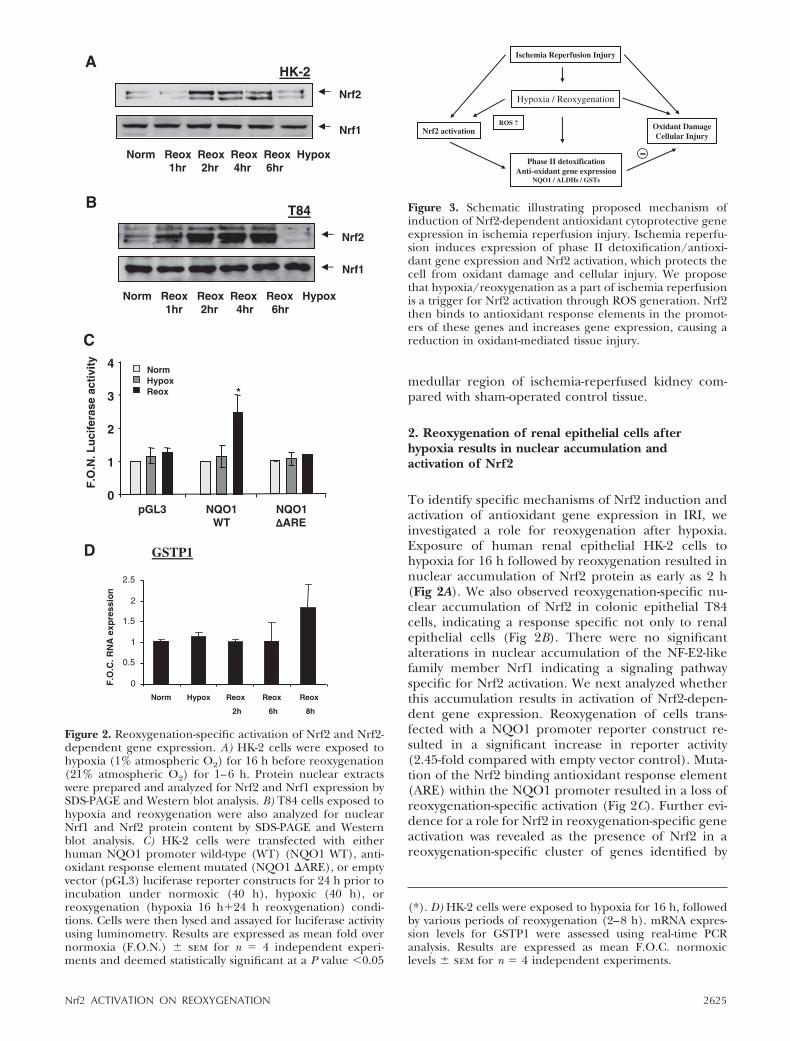
medullar region of ischemia-reperfused kidney com-pared with sham-operated control tissue.
2. Reoxygenation of renal epithelial cells afterhypoxia results in nuclear accumulation andactivation of Nrf2
To identify specific mechanisms of Nrf2 induction andactivation of antioxidant gene expression in IRI, weinvestigated a role for reoxygenation after hypoxia.Exposure of human renal epithelial HK-2 cells tohypoxia for 16 h followed by reoxygenation resulted innuclear accumulation of Nrf2 protein as early as 2 h(Fig 2A). We also observed reoxygenation-specific nu-clear accumulation of Nrf2 in colonic epithelial T84cells, indicating a response specific not only to renalepithelial cells (Fig 2B). There were no significantalterations in nuclear accumulation of the NF-E2-likefamily member Nrf1 indicating a signaling pathwayspecific for Nrf2 activation. We next analyzed whetherthis accumulation results in activation of Nrf2-depen-dent gene expression. Reoxygenation of cells trans-fected with a NQO1 promoter reporter construct re-sulted in a significant increase in reporter activity(2.45-fold compared with empty vector control). Muta-tion of the Nrf2 binding antioxidant response element(ARE) within the NQO1 promoter resulted in a loss ofreoxygenation-specific activation (Fig 2C). Further evi-dence for a role for Nrf2 in reoxygenation-specific geneactivation was revealed as the presence of Nrf2 in areoxygenation-specific cluster of genes identified by
Figure 2. Reoxygenation-specific activation of Nrf2 and Nrf2-dependent gene expression. A) HK-2 cells were exposed tohypoxia (1% atmospheric O2) for 16 h before reoxygenation(21% atmospheric O2) for 1–6 h. Protein nuclear extractswere prepared and analyzed for Nrf2 and Nrf1 expression bySDS-PAGE and Western blot analysis. B) T84 cells exposed tohypoxia and reoxygenation were also analyzed for nuclearNrf1 and Nrf2 protein content by SDS-PAGE and Westernblot analysis. C) HK-2 cells were transfected with eitherhuman NQO1 promoter wild-type (WT) (NQO1 WT), anti-oxidant response element mutated (NQO1 �ARE), or emptyvector (pGL3) luciferase reporter constructs for 24 h prior toincubation under normoxic (40 h), hypoxic (40 h), orreoxygenation (hypoxia 16 h�24 h reoxygenation) condi-tions. Cells were then lysed and assayed for luciferase activityusing luminometry. Results are expressed as mean fold overnormoxia (F.O.N.) � sem for n � 4 independent experi-ments and deemed statistically significant at a P value �0.05
(*). D) HK-2 cells were exposed to hypoxia for 16 h, followedby various periods of reoxygenation (2–8 h). mRNA expres-sion levels for GSTP1 were assessed using real-time PCRanalysis. Results are expressed as mean F.O.C. normoxiclevels � sem for n � 4 independent experiments.
Figure 3. Schematic illustrating proposed mechanism ofinduction of Nrf2-dependent antioxidant cytoprotective geneexpression in ischemia reperfusion injury. Ischemia reperfu-sion induces expression of phase II detoxification/antioxi-dant gene expression and Nrf2 activation, which protects thecell from oxidant damage and cellular injury. We proposethat hypoxia/reoxygenation as a part of ischemia reperfusionis a trigger for Nrf2 activation through ROS generation. Nrf2then binds to antioxidant response elements in the promot-ers of these genes and increases gene expression, causing areduction in oxidant-mediated tissue injury.
2625Nrf2 ACTIVATION ON REOXYGENATION

microarray analysis and through induction of the Nrf2-dependent gene GSTP1 (Fig 2D).
3. Investigation into intracellular signaling pathwaysinvolved in reoxygenation-mediated activation of Nrf2
Having demonstrated a role for Nrf2 in the transcrip-tional response to reoxygenation as a possible contrib-utor to gene expression patterns in IRI, we character-ized possible signaling events involved in its activation.Reactive oxygen species (ROS) generation after hyp-oxia and reoxygenation has been implicated as a triggerfor gene expression in reperfusion injury. To addresswhether such signals are responsible for Nrf2 activationon reoxygenation, we altered glutathione content toadjust the cells’ response to ROS and oxidative stress.Using the glutathione synthesis inhibitor butathionesulfoxamine (BSO), we observed a moderate increasein Nrf2 accumulation after 3 h reoxygenation. Treat-ment of cells with the antioxidant N-acetyl cysteine(NAC) resulted in inhibition of reoxygenation-specificinduction of Nrf2 nuclear accumulation.
We used pharmacological protein kinase inhibitorsto delineate signaling cascade involvement in Nrf2activation. Pretreatment of HK-2 cells with the p38 andc-Jun NH2-terminal kinase (JNK) MAPK pathway inhib-itors SB203580 and SP600125 resulted in a slight inhi-bition of reoxygenation-mediated activation of Nrf2.Inhibition of ERK MAPK and PKC using specific inhib-itors PD98059 and GF020848X resulted in significantattenuation of Nrf2 activation, and treatment with thePI3K inhibitor LY294002 resulted in near ablation ofNrf2 nuclear accumulation. A similar pattern of inhibi-tion of Nrf2 nuclear accumulation was observed withthese compounds under normoxic conditions.
4. Induction of Nrf2-dependent gene expression intransplanted ischemic reperfused human liver
We wanted to know whether Nrf2 activation and induc-tion of Nrf2-dependent gene expression in a murinemodel of IRI were translated into human disease.Analysis of mRNA levels by real-time PCR analysis ofliver biopsy taken after cold ischemia and portal reper-fusion (reperfusion biopsy) for 1 h revealed a signifi-cant increase in Nrf2, ALDH1A7, and GSTP1 geneexpression compared with donor retrieval biopsy.
CONCLUSIONS AND SIGNIFICANCE
In this study microarray analysis revealed induction of agroup of 7 phase II detoxification and antioxidantgenes all within the top 20 most highly up-regulated onrenal IRI. This coordinated response likely involvesNrf2-regulated transactivation of cis-acting AREs withinthe regulatory regions of these genes, as Nrf2 has beenidentified as a master regulator of these genes in othermodel systems. We demonstrate an increased stainingfor and accumulation of Nrf2 protein in ischemia-reperfused kidney, implicating activation of this tran-scription factor as a mechanism of induction of cyto-protective antioxidant gene expression. From our invitro studies, we demonstrate a reoxygenation-specificinduction of Nrf2 protein nuclear accumulation. Wealso demonstrate the reoxygenation-specific activationof NQO1 promoter reporter activity, which is inhibitedupon deletion of the ARE through which Nrf2 binds.RNA microarray analysis identified Nrf2 specificallywithin a cluster of genes induced on reoxygenationalone.
Investigation into the intracellular signaling mecha-nisms in reoxygenation-specific activation of Nrf2 re-vealed a role for ROS by use of the antioxidant NACand the glutathione synthesis inhibitor BSO. We noteda role for kinase signaling pathways in Nrf2 expressionthat likely contributes to basal but not reoxygenation-triggered expression of Nrf2. Evidence points to activa-tion of a pathway specific for cytoprotection againstoxidative damage in IRI induced by reoxygenation afterhypoxia/ischemia.We have demonstrated for the first time induction ofNrf2 activation and Nrf2-dependent antioxidant geneexpression in an in vivo model of IRI, as well asreoxygenation-specific activation of Nrf2 and Nrf2-dependent antioxidant gene expression in a renalepithelial cell in vitro model. We postulate that reoxy-genation-specific activation of the Nrf2 antioxidantpathway is a contributory mechanism to the adaptivecytoprotective response to ongoing and subsequentoxidant damage in ischemia-reperfusion. Further delin-eation of the signaling mechanisms involved in thisresponse—specifically the involvement of ROS signal-ing pathways—may enhance our understanding of isch-emic preconditioning in protecting against oxidativedamage.
2626 Vol. 20 December 2006 LEONARD ET AL.The FASEB Journal
Related Documents











