Reorganization of multi-muscle and joint withdrawal reflex during arm movements in post-stroke hemiparetic patients Mariano Serrao a,b,⇑ , Alberto Ranavolo c , Ole Kaeseler Andersen d , Romildo Don c , Francesco Draicchio c , Carmela Conte b , Roberto Di Fabio a , Armando Perrotta e,f , Michelangelo Bartolo e,f , Luca Padua g , Valter Santilli h , Giorgio Sandrini f , Francesco Pierelli a,e a University of Rome ‘‘La Sapienza’’, Polo Pontino, Latina, Italy b Rehabilitation Centre, Policlinico Italia, Rome, Italy c Laboratory of Physiology, Ergonomics, Posture and Movement, National Institute of Occupational Safety and Prevention, Monte Porzio Catone, Rome, Italy d Center for Sensory–Motor Interaction, Department of Health Science and Technology, Aalborg University, Aalborg, Denmark e Rehabilitation Centre, IRCCS ‘‘Neuromed’’ Pozzilli, Isernia, Italy f IRCCS ‘‘C. Mondino Institute of Neurology’’ Foundation, University of Pavia, Italy g Fondazione Don Gnocchi, Piazzale Morandi 6, 20121 Milan, Italy h Department of Physical Medicine and Rehabilitation, La Sapienza University, Rome, Italy article info Article history: Accepted 11 July 2011 Available online 6 August 2011 Keywords: Withdrawal reflex Movement analysis Stroke Spinal cord highlights In a broader motor context these findings represent the spinal neuropathophysiological correlate of the movement abnormalities observed in hemiparetic patients. This study show a maladaptive behavior of the spinal cord during arm movements in stroke patients determined by new functional brain–spinal cord connections. This finding indicates a marked plastic rearrangement of the spinal cord which partially loses its capac- ity for functional modulation by descending motor pathways during a motor task. abstract Objectives: To investigate the behavior of the nociceptive withdrawal reflex (NWR) in the upper limb dur- ing reaching and grasping movements in post-stroke hemiparetic patients. Methods: Eight patients with chronic stroke and moderate motor deficits were included. An optoelec- tronic motion analysis system integrated with a surface EMG machine was used to record the kinematic and EMG data. The NWR was evoked through a painful electrical stimulation of the index finger during a movement which consisted of reaching out, picking up a cylinder, and returning it to the starting position. Results: We found that: (i) the NWR is extensively rearranged in hemiparetic patients, who were found to present different kinematic and EMG reflex patterns with respect to controls; (ii) patients partially lose the ability to modulate the reflex in the different movement phases; (iii) the impairment of the reflex modu- lation occurs at single-muscle, single-joint and multi-joint level. Conclusions: Patients with chronic and mild-moderate post-stroke motor deficits lose the ability to mod- ulate the NWR dynamically according to the movement variables at individual as well as at multi-muscle and joint levels. Significance: The central nervous system is unable to use the NWR substrate dynamically and flexibly in order to select the muscle synergies needed to govern the spatio–temporal interaction among joints. Ó 2011 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. 1. Introduction Reaching and grasping arm movements with the affected limb in hemiparetic patients are characterized by decreased movement speed, lack of smoothness and coordination, and abnormal pat- terns of muscle activation (Hammond et al., 1988; Gowland 1388-2457/$36.00 Ó 2011 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.clinph.2011.07.031 Abbreviations: NWR, nociceptive withdrawal reflex; PT, pain threshold; FM, forward movement; BM, backward movement; AF, acceleration forward; DF, deceleration forward; AB, acceleration backward; DB, deceleration backward; CMCs, coefficients of multiple correlations. ⇑ Corresponding author at: University of Rome ‘‘La Sapienza’’, Polo Pontino, Via Faggiana 34, 40100 Latina, Italy. Tel.: +39 0773 651473. E-mail address: [email protected] (M. Serrao). Clinical Neurophysiology 123 (2012) 527–540 Contents lists available at ScienceDirect Clinical Neurophysiology journal homepage: www.elsevier.com/locate/clinph

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Clinical Neurophysiology 123 (2012) 527–540
Contents lists available at ScienceDirect
Clinical Neurophysiology
journal homepage: www.elsevier .com/locate /c l inph
Reorganization of multi-muscle and joint withdrawal reflex during armmovements in post-stroke hemiparetic patients
Mariano Serrao a,b,⇑, Alberto Ranavolo c, Ole Kaeseler Andersen d, Romildo Don c, Francesco Draicchio c,Carmela Conte b, Roberto Di Fabio a, Armando Perrotta e,f, Michelangelo Bartolo e,f, Luca Padua g,Valter Santilli h, Giorgio Sandrini f, Francesco Pierelli a,e
a University of Rome ‘‘La Sapienza’’, Polo Pontino, Latina, Italyb Rehabilitation Centre, Policlinico Italia, Rome, Italyc Laboratory of Physiology, Ergonomics, Posture and Movement, National Institute of Occupational Safety and Prevention, Monte Porzio Catone, Rome, Italyd Center for Sensory–Motor Interaction, Department of Health Science and Technology, Aalborg University, Aalborg, Denmarke Rehabilitation Centre, IRCCS ‘‘Neuromed’’ Pozzilli, Isernia, Italyf IRCCS ‘‘C. Mondino Institute of Neurology’’ Foundation, University of Pavia, Italyg Fondazione Don Gnocchi, Piazzale Morandi 6, 20121 Milan, Italyh Department of Physical Medicine and Rehabilitation, La Sapienza University, Rome, Italy
a r t i c l e i n f o
Article history:Accepted 11 July 2011Available online 6 August 2011
Keywords:Withdrawal reflexMovement analysisStrokeSpinal cord
1388-2457/$36.00 � 2011 International Federation odoi:10.1016/j.clinph.2011.07.031
Abbreviations: NWR, nociceptive withdrawal reflforward movement; BM, backward movement; AFdeceleration forward; AB, acceleration backward;CMCs, coefficients of multiple correlations.⇑ Corresponding author at: University of Rome ‘‘La
Faggiana 34, 40100 Latina, Italy. Tel.: +39 0773 65147E-mail address: [email protected] (M. S
h i g h l i g h t s
� In a broader motor context these findings represent the spinal neuropathophysiological correlate of themovement abnormalities observed in hemiparetic patients.� This study show a maladaptive behavior of the spinal cord during arm movements in stroke patientsdetermined by new functional brain–spinal cord connections.� This finding indicates a marked plastic rearrangement of the spinal cord which partially loses its capac-ity for functional modulation by descending motor pathways during a motor task.
a b s t r a c t
Objectives: To investigate the behavior of the nociceptive withdrawal reflex (NWR) in the upper limb dur-ing reaching and grasping movements in post-stroke hemiparetic patients.Methods: Eight patients with chronic stroke and moderate motor deficits were included. An optoelec-tronic motion analysis system integrated with a surface EMG machine was used to record the kinematicand EMG data. The NWR was evoked through a painful electrical stimulation of the index finger during amovement which consisted of reaching out, picking up a cylinder, and returning it to the starting position.Results: We found that: (i) the NWR is extensively rearranged in hemiparetic patients, who were found topresent different kinematic and EMG reflex patterns with respect to controls; (ii) patients partially lose theability to modulate the reflex in the different movement phases; (iii) the impairment of the reflex modu-lation occurs at single-muscle, single-joint and multi-joint level.Conclusions: Patients with chronic and mild-moderate post-stroke motor deficits lose the ability to mod-ulate the NWR dynamically according to the movement variables at individual as well as at multi-muscleand joint levels.Significance: The central nervous system is unable to use the NWR substrate dynamically and flexibly inorder to select the muscle synergies needed to govern the spatio–temporal interaction among joints.� 2011 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights
reserved.
f Clinical Neurophysiology. Publish
ex; PT, pain threshold; FM,, acceleration forward; DF,DB, deceleration backward;
Sapienza’’, Polo Pontino, Via3.
errao).
1. Introduction
Reaching and grasping arm movements with the affected limbin hemiparetic patients are characterized by decreased movementspeed, lack of smoothness and coordination, and abnormal pat-terns of muscle activation (Hammond et al., 1988; Gowland
ed by Elsevier Ireland Ltd. All rights reserved.

528 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
et al., 1992; Trombly, 1992). This motor impairment is due mainlyto disruption of the planning and execution of reaching and preci-sion grasping due to primary motor cortex or corticospinal tractdamage, as has been extensively observed in primates and humans(Hepp-Reymond and Wiesendanger, 1972; Maier et al., 1993;Aruin, 2005; Gölge et al., 2004; Grichting et al., 2000; Muir andLemon, 1983; Nowak et al., 2003; Beer et al., 2000; Takahashiand Reinkensmeyer, 2003; Quaney et al., 2005; Wenzelburgeret al., 2005; Raghavan et al., 2006).
However, the spinal contribution to abnormal patterns ofmovement following stroke has not been well investigated. Spas-ticity and deficits in voluntary control of movement are both, insome way, related to disorders in the organization of segmentalreflex activity (e.g. Corcos et al., 1986; Powers et al., 1988; Levinand Feldman, 1994). During reaching tasks, a relationship hasbeen observed between an impaired voluntary range of move-ment and limitations in the range of stretch reflex regulationat individual muscle level (Levin and Feldman, 1994; Levinet al., 2000). Although a number of studies have focused onchanges occurring in monosynaptic reflexes in a single-joint fol-lowing stroke (Thilmann and Fellows, 1991; Wilson et al., 1999;Condliffe et al., 2005), there is a lack of research addressing therole of polysynaptic multi-joint reflexes. Thus, it is not knownhow these reflexes in the upper extremities are modulatedduring movement in stroke survivors compared to healthysubjects.
In a general motor context, control systems exploit reflexes inmovement and force or torque production, which is consistentwith the fact that reflexes are broadly adjustable rather than rigidstimulus–response constructs (Balasubramaniam and Feldman,2004). Spinal reflex pathways acting at multi-joint levels may beactivated by descending commands to generate and coordinatevoluntary movements (Lundberg, 1979).
The few studies that have investigated upper limb polysynapticreflexes in stroke patients were performed only at rest (Dewaldet al., 1999), during either passive (Black et al., 2007) or active,but machinery constrained, arm movements (Sangani et al.,2007). It would, however, be useful to investigate simultaneouslythe spinal reflexes in several muscles during unconstrained naturalarm movements in order to obtain a global picture of spinal func-tion and its modulation.
Of the various polysynaptic spinal reflexes, the withdrawal re-flex has been proved to be a useful tool for investigating changesin spinal cord function during lower limb movements in humans(Crenna and Frigo, 1984; Duysens et al., 1990,1992; Rossi and Dec-chi, 1994; Andersen et al., 1995; Andersen, 2007; Spaich et al.,2004, 2006; Sandrini et al., 2005; Emborg et al., 2009), given thatthese changes are due to converging inputs from the peripheralafferents and descending motor commands to the spinal cord neu-rons (Baldissera et al., 1981).
Although the flexion synergy evoked by painful stimuli serves aprimarily protective function, various studies have shown that the
Table 1Patients’ clinical characteristics. All patients were able to perform the required motor task
Pts Age Sex Time since stroke (years) Side Brain localization
1 58 M 3 R Precental gyrus2 63 M 5 R Internal capsule, corona ra3 69 M 2 L Precentral gyrus, capsule i4 65 M 2 R Internal capsule5 58 M 6 L Cortical and subcortical he6 53 M 4 R Internal capsule7 62 M 4 R Frontal cortical reigon and8 58 M 3 R Internal capsule, corona ra
nociceptive withdrawal reflex (NWR) also fulfills a more complexmotor function (for a review see Sandrini et al., 2005).
A few years ago (Serrao et al., 2006), we investigated the modu-lation of the NWR in human upper limbs during reaching and grasp-ing movements performed freely in three-dimensional space. Wefound that the NWR undergoes state- and phase-dependent modu-lation according to the mechanical function exerted by each musclein the course of the motor sequence. In particular, we observed thatthe amount and direction of the motion occurring at each joint inthe course of the motor task were the main factors determiningthe presence of either reflex reversal (presence of the reflex in eitheragonist or antagonist muscle in a phase reversal manner) or co-pres-ence (presence of the reflex in both antagonist muscles withoutphase reversal) patterns (Serrao et al., 2006). These findings suggestthat transmission along the excitatory reflex pathways is highlyadaptable to the motor context and related to the joint motion,and that supraspinal centers possibly exploit the same interneuro-nal spinal network in order to regulate voluntary movements andreflex responses in a complex, multisegmental way.
Application of this method in hemiparetic stroke survivors mayhelp to further understanding of the role of the descending path-ways in dynamically recruited spinal neurons (both motorneuronsand interneurons) involved in mediating reflexes, and also shedlight on the reorganization processes and adaptive changes occur-ring at multi-segmental level in the spinal cord.
We set out to investigate the behavior of the NWR in terms ofkinematic and EMG responses following electrical painful stimula-tions during reaching and grasping movements in the upper limbin a sample of post-stroke hemiparetic patients.
2. Methods
2.1. Patients
Eight patients (8 M, 56–64 years) with mild-moderate chronicpost-stroke hemiparesis were enroled in the study.
The inclusion criteria were: (i) the presence of a chronic(>9 months) ischemic brain lesion of the dominant hemisphereresulting in contralateral hemiparesis; and (ii) the ability to per-form the required motor task (reaching, grasping and transportingback an object).
The exclusion criteria were: (i) previous ischemic or hemor-rhagic stroke; (ii) chronic cerebrovascular encephalopathy; (iii)concurrent sensory disturbances; (iv) neglect; (v) apraxia (vi) cog-nitive impairment (MMSE <24); (vii) muscle or joint pain; and (viii)the presence of orthopedic disorders further impairing the perfor-mance of the affected arm.
In this study, motor function of the impaired upper limb inhemiparetic subjects was tested using the Fugl-Meyer scale(Fugl-Meyer et al., 1975). Muscle tone was evaluated using thesix-point (0–5) Ashworth scale (Ashworth, 1964).
.
Fugl-Meyer upper limb score Ashworth score
Wrist Elbow Shoulder
48diata 50 1 1 1nternal 46 2 1 2
52 2 2 1misphere 58 2 2 2
60 1 3 2corona radiata 52 1 2 1
diata 48 2 2 2

Fig. 1. Representation of the motor task. Forward (reaching to grasp the cylinder) and backward (grasping and transporting the cylinder back) movements.
M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540 529
Stroke subjects were evaluated by a physiotherapist to deter-mine their eligibility to participate. The patients’ clinical character-istics are reported in Table 1.
For comparison, eight healthy men, ranging in age from 46 to60 years, were also included in this study.
All the participants gave their written informed consent to thestudy, which had local ethics committee approval. The study com-plied with the terms of the Helsinki Declaration relating to humanexperimentation.
2.2. Instrumental evaluation
We used the SMART-E motion analysis system (BTS, Milan,Italy) to record the kinematic variables. The system consists ofeight infrared cameras (120 Hz sampling rate) which detect themotion of passive reflective markers. The markers were positionedon the right and left anterior superior iliac spines, on the spinousprocess of the seventh cervical vertebra, on the sacrum, on the leftand right acromions, on the right olecranon, on the right styloid ul-nar and radial processes, on the head of the third metacarpal bone,and on the dorsal aspects of the distal phalanx of thumb and indexfingers.
Surface EMG signals were recorded using a multi-channel Wi-FiPocketEMG system (BTS, Milan) operating at a sampling rate of1000 Hz, and bandpass-filtered at 10–400 Hz. The EMG activitywas recorded through pairs of Ag–AgCl surface electrodes pre-coated with electroconductive gel (diameter 1 cm, distance be-tween electrodes 2 cm), placed over eight muscles including theanterior and posterior deltoid (AD and PD), pectoralis major(PM), latissimus dorsi (LD), brachioradialis (BR), triceps brachii(TB), flexor carpii radialis (FCR), and extensor carpii radialis(ECR), according to standard anatomical landmarks (Hermenset al., 2000).
2.3. Painful stimulation technique
The stimulation technique was similar to that described previ-ously (Serrao et al., 2006; Don et al., 2008). Briefly, the right digitalnerve of the index finger was stimulated percutaneously through apair of ring electrodes. The stimulus consisted of a 20-ms train offive rectangular pulses (duration: 1 ms; frequency: 200 Hz) deliv-ered through a constant-current stimulator (Grass S-88; GrassMedical Instruments, Quincy, MA, USA). The stimulator triggeredthe motion analysis system.
We estimated the pain threshold (PT) using a staircase method(Willer, 1977). The intensity of the electrical shocks used for thesubsequent procedures was adjusted to three times of the PT(3xPT).
Subjects were asked to score their pain perception after eachstimulus on a 0–10 points visual analog scale (VAS).
2.4. Experimental procedure
Before starting formal measurements for the study, the subjectsunderwent an initial training session to familiarize them with theassessment procedures.
The general experimental methodology was similar to that de-scribed previously (Serrao et al., 2006; Don et al., 2008). Briefly, thesubjects were seated on a comfortable chair in front of a table in aquiet room in normal indoor lighting conditions. We evaluated amovement that consisted of reaching out from a starting position,picking up a cylinder (diameter: 3 cm; height: 6 cm; weight:300 g), and returning it to the starting position (Fig. 1). The startingposition and cylinder location were 40 cm apart.
The required speed was established during the initial practicesession. The participants were allowed periodic breaks to minimizefatigue. The trunk was restrained using a set of straps to minimizemotion.
We recorded 80 movement trials per healthy subject. During 60of these trials, randomly selected, a single painful stimulus wasdelivered (perturbed trials). The timing of stimulus delivery withineach perturbed trial was also randomly determined across thewhole motor sequence (from the beginning of movement to thecylinder placement). The remaining 20 trials, performed withoutstimulus delivery (unperturbed trials), were used for comparisons.
For the hemiparetic patients, fewer trials per person were col-lected (ranging from 45 to 55 trials) as 60 perturbed trials were,in all cases, considered excessive.
3. Data analysis
3.1. Kinematic data analysis
Data processing was performed using Analyzer software (BTS,Milan, Italy). We used a validated four-segment model of the trunkand upper limb to calculate angle and angular velocity of the wrist,elbow and shoulder joints (Rab et al., 2002). We analyzed adduc-tion-abduction and flexion–extension movements of the wrist,

Fig. 2. Algorithm used to compare the reflex window between unperturbed and perturbed trials (see Section 2 for detailed information).
530 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
flexion–extension movements of the elbow, and flexion–extensionand horizontal adduction-abduction of the shoulder.
The displacement and linear velocity of the radial marker inthe anterior-posterior direction were used to identify differentphases of the motor sequence (Serrao et al., 2006; Don et al.,2008). Forward movement (FM) was defined as the phase duringwhich the subject reached for the cylinder, and backward move-ment (BM) the one during which the subject transported the cyl-inder. The transition from forward to backward movementcorresponded to the point at which the direction of movementof the radial marker reversed. Then, each movement was furtherdivided into an accelerative and a decelerative phase, on the basisof the radial marker velocity peaks. The entire motor sequencewas thus divided into four subphases: acceleration forward (AF),deceleration forward (DF), acceleration backward (AB), and decel-eration backward (DB).
The mean duration of the entire motor sequence, as well as ofthe two phases and four subphases were calculated.
3.1.1. Kinematic reflex detectionTo evaluate the mechanical effect, the kinematic data in the
125–250 ms window after the painful electrical stimulation wereanalyzed.
For the shoulder, elbow and wrist, kinematic data for each sin-gle perturbed trial were compared to the mean of the subject’sunperturbed trials as reported in our previous study (Serraoet al., 2006) (Fig. 2). Measurements were performed as follows:(i) the instant at which the stimulus was delivered in each per-turbed trial was expressed as a percentage of the mean durationof the subject’s unperturbed trials; (ii) the joint angular velocitymean absolute value of each unperturbed trial was calculated ina time window of 125–250 ms (control window); (iii) the mean(and SD) angular velocity of all unperturbed trials was calculated;(iv) the mean angular velocity area was taken as the baseline valueand the mean value ±2SD was fixed as the mechanical responsethreshold; (v) the mechanical reflex responses were consideredpresent if the angular velocity values of the perturbed trial ex-ceeded this threshold; vi) reflex size (angular velocity changes)(dSDxp) was calculated using the following formula:
dSDxp ¼ ðxp�xuÞ=xSDu;
in which xp is the absolute value of joint angular velocity in theperturbed trial, xu is the corresponding baseline value, and xSDuis the standard deviation of the unperturbed trials.
Perturbed trials in which the reflex response overlapped twomovement phases were discarded.

M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540 531
For each perturbed trial, the sign of the reflex size (positive val-ues indicated acceleration and negative values indicated decelera-tion) was identified; moreover, it was noted whether themechanical perturbation induced by the electrical painful stimula-tion shifted the direction of the wrist marker trajectory (reversedirection with respect to the actual joint motion).
3.2. EMG data analysis
Data processing was performed using Matlab software (Matlab7.0.4, MathWorks, Natick, MA, USA).
Raw EMG signals recorded during the motor sequence werefull-wave rectified.
The reflexes were analyzed considering the EMG activity in thetime window from 60 to 200 ms after the stimulus (Serrao et al.,2006; Don et al., 2008). To analyze the presence and size of the re-flex in the perturbed trials, the shoulder, elbow and wrist muscleEMG data of each single perturbed trial were compared with themean of the subject’s unperturbed trials, assuming that until theinstant in which the stimulus was given, the execution of the mo-tor sequence was the same in the perturbed and unperturbed trials(Serrao et al., 2006; Don et al., 2008) (Fig. 2).
Measurements were performed as follows: (i) the instant atwhich the stimulus was delivered in each perturbed trial was ex-pressed as a percentage of the mean duration of the subject’sunperturbed trials; (ii) the EMG area of each unperturbed trialwas calculated in a time window of 60–200 ms; (iii) the mean(and SD) EMG area of all unperturbed trials was calculated; iv)the mean EMG area was taken as the baseline value and the meanvalue ± 2SD was fixed as the EMG response threshold; (v) the EMGreflex responses were considered present if the EMG area values ofthe perturbed trial exceeded this threshold; and (vi) the EMG NWRsize (dsdEMGp) was obtained using the following formula:
dsdEMGp ¼ ðEMGp� EMGuÞ=EMGSDu;
in which EMGp is the muscle EMG area in the single perturbed trial,EMGu is the baseline value, and EMGSDu is the SD of the baselinearea values. We used the SD of the baseline values to reduce thevariability of the EMG background activity of the unperturbed trialsduring movement thus avoiding under- or overestimation of theNWR size.
4. Statistical analysis
All analyses were performed using SPSS 17.0 software. Descrip-tive statistics included mean values and SDs. A p value <0.05 wasconsidered statistically significant.
4.1. Comparisons of kinematic variables (unperturbed trials)
The non-parametric Mann–Whitney test was used to comparemean durations of the motor sequence, movement phases and sub-phases and the angle values of the shoulder, elbow and wrist jointsbetween hemiparetic patients and controls.
We used coefficients of multiple correlations (CMCs) to com-pare radial marker displacement and velocity between hemipareticpatients and controls in the posterior–anterior (x), vertical (y) andlateral (z) directions.
The CMC is a measure of the overall waveform similarity of agroup of curves, and its magnitude is close to 1 if the waveformsare similar and close to 0 otherwise. A CMC value >0.7 indicatesgood waveform similarity (Kadaba et al., 1989).
4.2. Repeatability of kinematic variables
Repeatability of the radial marker displacement and velocitywere investigated using CMCs. Intrasubject repeatability was as-sessed by first computing the CMC for each subject over 20 unper-turbed trials and then averaging these values over all the subjectsin both of the groups. Intersubject repeatability was assessed bycomputing the CMCs for the unperturbed trials performed by allthe subjects in both of the groups (Kadaba et al., 1989).
4.3. Reflex comparisons and modulation
Kinematic and EMG ‘‘reflex probability’’ (expressed as a per-centage) was defined as the number of trials in which reflex re-sponses were detected in each movement subphase. As regardsthe EMG responses, the reflex probability in a single muscle wastermed ‘‘single-muscle probability’’ and the reflex probability inpaired antagonist muscles was termed ‘‘single-joint probability.’’
The Chi-square test was used to compare ‘‘single-muscle prob-ability’’ and ‘‘single-joint probability’’ between patients and con-trols for each subphase, and between subphases within patientsand controls.
For the kinematic reflex, the Chi-square test was used to com-pare the percentage positive/negative values of the reflex size signs(number of trials with either positive or negative signs) as well asthe direction shifts (number of trials showing reversal of the direc-tion of the radial marker) between patients and controls for eachsubphase.
The two-way ANOVA test was used to analyze the reflexsize depending on groups (patients and controls) and either move-ment subphases. Tukey post hoc test was used for pair-wisecomparisons.
The non-parametric Mann–Whitney test was used for compar-ing VAS values.
A Spearman rank correlation was performed to determinewhether the kinematic and reflex sizes were significantly corre-lated with the Ashworth scores in the hemiparetic patients.
5. Results
5.1. Kinematics
5.1.1. Comparisons of kinematic variables in the unperturbed trialsThe mean radial marker linear displacement and velocity in the
anterior-posterior direction and its division into phases and sub-phases in patients and controls are reported in Fig. 3.
Significant differences between patients and controls were ob-served for the total duration of the motor sequence (all compari-sons between patients vs controls; 1.52 ± 0.41 vs 1.06 ± 0.13 s,p < 0.05), the FM phase (0.86 ± 0.28 vs 0.52 ± 0.17 s, p < 0.05) andthe DF subphase (0.48 ± 0.21 vs 0.25 ± 0.12 s, p < 0.05).
No significant differences in the shoulder, elbow and wristrange of motion (ROM) were detected between subjects and pa-tients (ROMs: all, p > 0.05).
Mean between-groups CMCs for radial marker velocity in the x,y and z axes and displacement in the y and x axes were all >0.7,whereas CMC for marker displacement in the z axis was 0.609,
5.1.2. Repeatability of kinematic variablesThe mean intrasubject and intersubject CMCs of the curves
describing the radial marker displacement and velocity were all>0.79 and 0.75 in hemiparetic patients and >0.95 and >0.90 in con-trols, respectively.

Fig. 3. Displacement (m) and the linear velocity (m/s) of the right styloid radial marker in the anterior–posterior direction in the unperturbed trials in a patient (a) and control(b) (mean ± SD of 20 trials). The movement cycle is normalized with respect to the duration of the motor task (cycle %).
532 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
5.1.3. Type of joint motion in unperturbed and perturbed trialsThe types of joint motion observed in perturbed and unper-
turbed trials in both patients and controls are reported in Fig. 4.The motion of single joints did not differ between patients andcontrols in the unperturbed trials, whereas differences were ob-served in the perturbed ones. In the perturbed trials, the patientsshowed either shoulder extension (>70% of trials) or shoulder flex-ion (<30% of trials) in the AF and DF subphases, shoulder extensionin AB and DB and shoulder horizontal abduction, elbow flexion, aslight (<5�) and infrequent (<25% of trials) wrist abduction, andextension in all the movement subphases, while the controls pre-sented the same joint motion as seen in the unperturbed trials(Fig. 4). As regards the reflex size signs, no significant differenceswere observed in patients across movement subphases (Chi-square, all, p > 0.05), the signs were always positive (acceleration),whereas in the control subjects significant differences were ob-served between movement subphases. Indeed, the healthy subjectsexhibited decreased angular velocity (negative signs) in the shoul-der and elbow joint movements in the AF and AB subphases andincreased angular velocity (positive signs) in the DF and DB sub-phases (Chi-square, all, p < 0.01) (Fig. 4).
A significant difference in the occurrence of radial marker lineardirection shift after electrical stimulation was found between pa-tients and controls in both the AF and DF subphases (Chi-square,all, p < 0.01), this shift occurring with high probability (100%) inpatients and being nearly absent (<1%) in controls. Fig. 5 showsan example of direction shift of the radial marker trajectory in ahemiparetic patient compared to a control subject.
5.1.4. Reflex probability and size comparisonsKinematic responses (reflex probability and size) to painful
stimulation are reported in Table 2.
For the shoulder and elbow joints, no significant differences inreflex probability were found between patients and controls, con-sidering the different subphases (Chi-square, all, p > 0.05), the re-flex being present with high probability in both groups (Table 2).For the wrist joint, higher reflex probability values were found inhemiparetic patients than in controls in every subphase, as the re-flex probability in controls was very low (<1%) (Chi-square, all,p < 0.01) (Table 2).
Reflex size values for the shoulder joint in the sagittal planewere lower in patients than in controls in the AF, DF and AB sub-phases (Mann–Whitney, p < 0.05), while no differences were foundin the horizontal plane (Mann–Whitney, p > 0.05). On the contrary,higher reflex sizes in the elbow joint were found in patients than incontrols for every subphase (Mann–Whitney, p < 0.01) (Table 2).
5.1.5. Kinematic reflex probability and size modulationsNo significant differences in reflex probability or in reflex size
were found in either patients or controls when considering themovement subphases for each joint (Chi-square and ANOVA tests,all, p > 0.05).
5.2. EMG findings
5.2.1. Reflex comparisonsFigs. 6 and 7 show the individual muscle reflex probability and
size values recorded in the patients and controls. Several significantdifferences in reflex probability and size between patients and con-trols were detected when considering the different movement vari-ables in each investigated muscle. Briefly, the following significantresults were observed: (i) patients showed higher reflex probabilityvalues in almost all muscles (the PD was the exception) in the dif-ferent movement subphases, however, lower values were observed

Fig. 4. Shoulder, elbow and wrist joint motion across the motor task in both healthy subjects and patients. The type of joint motion and the accelerative and decelerative jointbehaviors are reported for both unperturbed (white cells) and perturbed trials (gray cells). Bold characters indicate joint motion differences between patients and controlsand between unperturbed and perturbed trials. FM = forward movement; BM = backward movement; AF = acceleration forward; DF = deceleration forward; AB = accelerationbackward; DB = deceleration backward; Nm = no motion; Flex = flexion; Ext = extension; Acc = acceleration; Dec = deceleration; SP = sagittal plane; HP = horizontal plane.
M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540 533
in the LD and PM muscles only in AF subphase and in DF and ABsubphases, respectively; (ii) patients showed higher reflex size val-ues than controls in all the muscles and movement subphases.
5.2.2. Reflex modulationFigs. 6 and 7 show the reflex probability modulation in the dif-
ferent movement subphases within patients and controls.

Fig. 5. A representative example of direction shift of the radial marker trajectory in a hemiparetic patient and a control.
Table 2Kinematic responses (reflex probability and reflex size) to electrical painful stimulation during movement. Trials considered for measurements: 160 unperturbed and 400perturbed trials in hemiparetic patients; 160 unperturbed and 480 perturbed trials in controls. Bold type indicates significant differences (p < 0.05) between healthy subjects andhemiparetic patients in the different movement variables. Empty cells indicate either non-measurable or non-comparable values. Cells with two values indicate either flexion orextension, respectively.
Motor sequence
Shoulder Elbow Wrist
Flexion–extension H. adduction–abduction Flexion–extension Flexion–extension Adduction–abduction
Hemiparetic patientsPhase Reflex
probabilityReflexsize
Reflexprobability
Reflexsize
Reflexprobability
Reflexsize
Reflexprobability
Reflexsize
Reflexprobability
Reflexsize
AF⁄ 100% +1.9 (0.4)+2.1 (0.4)
99.8% +2.7 (0.8) 100% +5.4 (2.0) 64.8% +2.2 (0.9) 22.1% +1.5 (0.6)
DF⁄ 100% +2.2 (0.6)+2.5 (0.5)
100% +2.5 (0.7) 100% +5.0 (2.1) 66.5% +2.4 (1.8) 23.9% +1.6 (0.5)
AB 99.3% +2.8 (0.7) 99.2% +2.9 (0.9) 100% +5.5 (1.8) 67.4% +1.5 (1.2) 23.3% +1.5 (0.6)DB 98.6% +2.9 (1.0) 98.6% +3.2 (0.9) 100% +5.2 (1.9) 69.6% +2.7 (1.3) 24.5% +2.7 (1.9)
ControlsAF 95.2% �3.2 (0.6) 92.0% �3.0 (0.5) 89.2% �3.3 (0.5) 0% – 0% –DF 94.8% +2.7 (0.6) 92.8% +2.6 (0.9) 90.5% +3.3 (0.6) 0% – 0% –AB 92.8% �3.8 (0.4) 96.8% �3.2 (0.7) 93.6% �3.4 (0.7) 0% – 0% –DB 95.8% +3.0 (0.6) 94.0% +3.5 (0.5) 90.4% +3.2 (0.7) 0% – 0% –
AF = acceleration forward; DF = deceleration forward; AB = acceleration forward; DB = deceleration backward.
534 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
Patients did not show significant differences in reflex probabil-ity between subphases for any of the muscles, except for the ADand LD (Fig. 6). Conversely, for the healthy controls, significant dif-ferences in reflex probability between subphases were observedfor all the muscles, except the PD and AD (Fig. 6).
In hemiparetic patients, significant differences in reflex sizewere observed between the PD and PM muscles, while no differ-ence was found in any of the other muscles (Fig. 7). In controls sig-nificant differences in reflex size between subphases were found inall the muscles, except for the AD and PD (Fig. 7).
Fig. 6 show also the single-joint reflex probability modulation inboth patients and controls.
Patients did not show any significant reflex probability differ-ences between antagonistic muscles, except between the AD andPD muscles in the AF and DF subphases (Fig. 6). Controls showedsignificant differences between all antagonist muscles in all sub-phases, except between FCR and ECR in all subphases (Fig. 6).
5.2.3. VAS findingsDespite a trend towards lower VAS scores, the pain intensity re-
ported by the patients was not found to differ from controls during
the different movement subphases (patients vs controls, AF:6.3 ± 1.2 vs 6.5 ± 0.8, DF: 6.0 ± 1.5 vs 6.6 ± 1.3, AB: 6.3 ± 1.1 vs6.5 ± 0.8, DB: 6.2 ± 1.2 vs 6.5 ± 1.1).
Furthermore, there were no significant differences in painintensity between the phases and subphases within patients orwithin control subjects (all, p > 0.05).
5.2.4. Ashworth scale correlationsNo correlation was found between reflex sizes (kinematic and
EMG) and Ashworth scale scores (all, p > 0.05).
6. Discussion
In the present study the modulation of the upper limb NWR inpost-stroke hemiparetic patients in terms of kinematic and EMGreflex responses during reaching and grasping movements wasinvestigated. The study was conducted in a sample of patients withchronic stroke (>9 months) and moderate motor deficits (Fugl-Mayer >45) and spasticity (Ashworth <3 in all joints), and the find-ings were compared to those obtained in a group of age-matched

Fig. 6. Single-muscle and single-joint EMG reflex probability comparison and modulation between movement phases in patients and controls. AD = anterior deltoid;PD = posterior deltoid; PM = pectoralis major; LD = latissimus dorsi; BR = brachioradialis; TB = triceps brachi; FCR = flexor carpi radialis; ECR = extensor radialis carpi;FM = forward movement; BM = backward movement; AF = acceleration forward; DF = deceleration forward; AB = acceleration backward; DB = deceleration backward§ = statistical differences between patients and controls; � = statistical differences between subphases.
M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540 535
healthy controls. Since we did not evaluate patients with severemotor impairment and/or severe spasticity, the results cannot begeneralized to all stroke patients and thus it limits the potentialimpact of the observations. The fact that these patients were ableto perform the required motor task freely in three-dimensionalspace allowed us to obtain information on the transmissionchanges that occur in the neuronal substrate mediating the NWRas a result of the impairment of the dynamic descending controls.
The results of the study can be summarized as follows: (i) theNWR is extensively rearranged in hemiparetic patients, who pre-sented different kinematic and EMG reflex patterns with respectto controls; (ii) patients partially lose the ability to modulate thereflex in the different movement phases and subphases; and (iii)the impairment of the reflex modulation occurs at both single-muscle, single-joint and multi-joint levels.
6.1. Kinematics
6.1.1. Quality and repeatability of arm movementIn our study, even though the hemiparetic patients’ radial mar-
ker trajectories and velocities were mildly segmented and irregu-lar, their arm movements were not qualitatively different fromthose of healthy subjects, as shown by the CMCs, which were al-ways higher than 0.7. Indeed, for all subjects (post-stroke partici-pants and controls) the motor task consisted of a forwardfollowed by a backward movement, each movement being charac-terized by an acceleration followed by a deceleration subphase(Fig. 3). The main difference in reflex modulation concerned theduration of the DF subphase. As shown in Fig. 3, the radial markertended to decelerate drastically, leading to a flattening of the veloc-ity curve in the DF subphase. This behavior has also been reportedby other authors (van Vliet and Sheridan, 2007) and it probably re-
flects an extra attentive effort on the part of patients as they ap-proach the object prior to picking it up.
Although a slight difference in the intrasubject CMCs emergedbetween patients and controls, for both groups the CMCs wereabove 0.7 indicating high repeatability of the kinematic data inour series of subjects, patients as well as controls.
6.1.2. Kinematic reflex responsesHemiparetic patients showed a different reflex pattern across
the motor sequence compared to controls in terms of type ofmechanical response (joint motion), movement direction shift, re-flex probability and reflex size.
As regards the type of kinematic response, patients presented atshoulder joint level an extension and a horizontal abduction re-sponse. Some patients showed a flexion response which was pres-ent only in the first two movement subphases (Table 2 and Fig. 4).At elbow level, patients showed the same stereotyped reflex re-sponse characterized by a flexion in all subphases. At wrist level,patients showed a slight (<5�) and infrequent (<25% of trials)abduction and extension response which did not change acrossmovement subphases. In all patients and all joints, the mechanicalresponse consisted of an accelerative motion, regardless of themovement subphases (Table 2 and Fig. 4). This upper limb reflexpattern was quite different from that observed in the controls. In-deed, shoulder and elbow kinematic responses in the controls con-sisted of either an increase or a decrease of acceleration anddeceleration according to the movement variables, while no jointmotion was detected at wrist level (Fig. 4).
A reversal of the radial marker trajectory and velocity withrespect to the actual movement in the forward direction was foundin the patients indicating a movement direction shift. This ismainly attributed to an elbow flexion reflex response
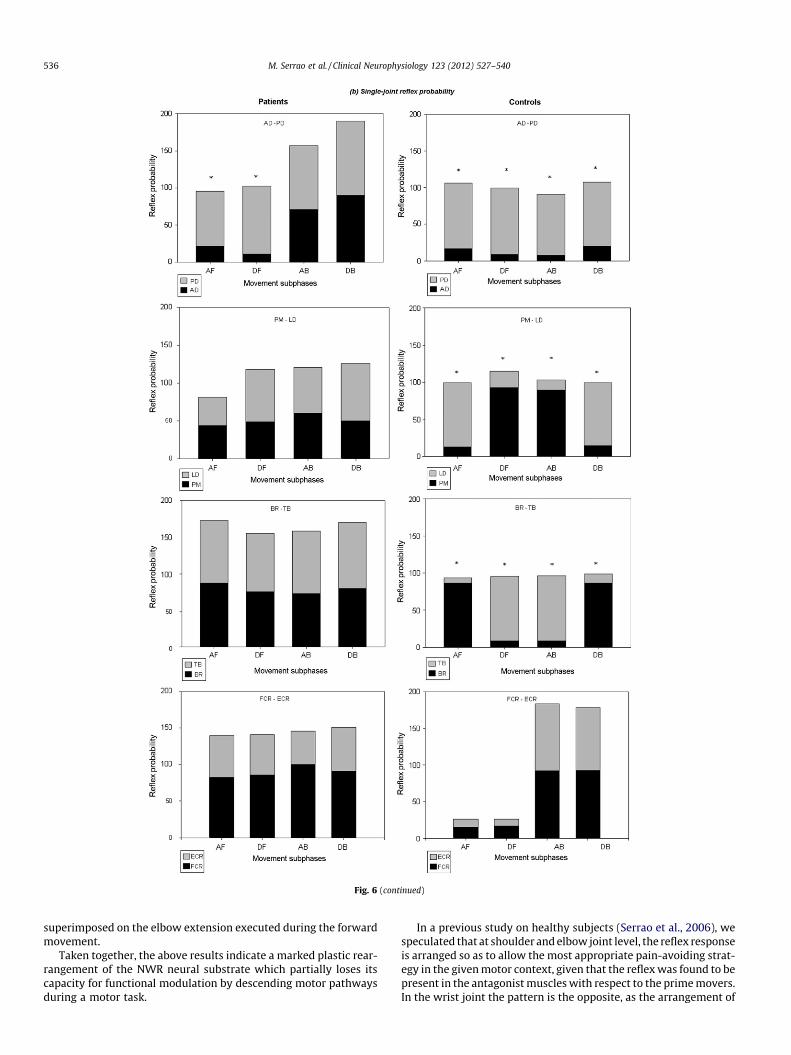
Fig. 6 (continued)
536 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
superimposed on the elbow extension executed during the forwardmovement.
Taken together, the above results indicate a marked plastic rear-rangement of the NWR neural substrate which partially loses itscapacity for functional modulation by descending motor pathwaysduring a motor task.
In a previous study on healthy subjects (Serrao et al., 2006), wespeculated that at shoulder and elbow joint level, the reflex responseis arranged so as to allow the most appropriate pain-avoiding strat-egy in the given motor context, given that the reflex was found to bepresent in the antagonist muscles with respect to the prime movers.In the wrist joint the pattern is the opposite, as the arrangement of

Fig. 7. EMG reflex sizes modulation between movement variables in patients and controls.
M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540 537
the reflex is determined by the stabilizing role that the CNS conferson this joint during reaching and grasping movements (Cruse et al.,1993; Dean and Brüwer, 1994; Haggard et al., 1995).
Unlike healthy subjects, hemiparetic patients lack, mainly at el-bow and wrist joint levels, the capacity for functional adaptation of
the nocifensive response to the motor context, given that the reflexresponse in these joints was inflexibly invariant across the entiremovement. The main mechanical response consisted of a retrac-tion of the arm from the painful source by an elbow flexion, regard-less of the movement subphases.

538 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
The very high reflex probability and sizes primarily observed atthe elbow and wrist joints (Table 2) suggest an abnormal spreadingof cutaneous afferent inputs into the multi-segmental spinal cordwith enhanced kinematic responses to electrical stimulation acrossthe movement in these joints.
6.2. EMG reflex responses
Hemiparetic patients exhibit a partial lack of reflex probabilityand size modulation in the different movement subphases. In par-ticular, the patients did not show changes in reflex probability dis-tribution across the movement phases in the FRC and ERC muscles,as observed in controls, whereas they did show changes in the ADmuscle (Fig. 6). Furthermore, in almost all muscles patients did notshow changes in reflex probability distribution across subphases,even though changes in the AD and LD were recorded.
In healthy subjects (Figs. 6 and 7) it is clear that in the shoulderand elbow joints, modulation of the NWR was determined by twoprinciples, i.e., reflex reversal and the mechanical relevance ofmuscle activity in the course of the movement, as also observedin our previous study (Serrao et al., 2006). For example, consideringthe acceleration-deceleration of the elbow joint, the reflex proba-bility was low in the triceps and high in the brachioradialis musclein the AF subphase when the triceps was the prime mover, while itwas high in the triceps and low in the brachioradialis in the DFsubphase when the biceps was the prime mover (Fig. 6). This pat-tern was reversed during the backward movement (Fig. 6). Thisbehavior suggests that transmission in the excitatory reflex path-ways is modulated in order to allow a wide range of withdrawalstrategies. In contrast, patients did not show this reflex modulationin most of the muscles individually and, when it was present, as inAD and LD (Fig. 6), this modulation was not linked to any mechan-ical or physiological principle.
The impaired reflex modulation was even more pronounced bya differential distribution of single-joint reflex probability betweenantagonist muscles (Fig. 6). In the healthy subjects the reflex wasalternately present in antagonistic muscles around the shoulderand elbow joints with a reflex reversal behavior characterizingthe transition from one subphase to another. In patients, thisbehavior was lost; partly with the exception of the AD and PD mus-cles. Indeed, a ‘‘coupling reflex pattern’’ (co-presence of the reflexin the antagonistic muscles) in the elbow and shoulder joint mus-cles without any reflex reversal was seen in the patients. In con-trast, in wrist muscles a ‘‘reciprocal reflex pattern’’ instead of‘‘coupling reflex pattern’’ without any phase modulation was ob-served (Fig. 6).
6.3. Functional considerations
In a broader motor context these findings may represent theneuropathophysiological correlate of the movement abnormalitiesobserved in hemiparetic patients, as reported in several previousstudies (Levin, 1996; Roby-Brami et al., 1997, 2003; Cirstea and Le-vin, 2000). It has been suggested that alterations in the trajectoriesof goal-directed movements is due to a deficit of smooth temporalinterjoint coordination between elbow and shoulder joints (Levin,1996), as well as to a deficit in the regulation of interactive jointtorques (Beer et al., 2000). On this basis, our data indicate that alack of interjoint coordination and interaction torques may be sec-ondary to the inability of the descending commands to controltemporally and spatially the spinal reflexes in a multi-joint coordi-nated manner and to adapt them to a functional motor context.
Disruption of the motor cortex and cortico-spinal tract afterstroke is probably the main reason for such spinal rearrangement.However, it is important to consider that descending pathways arenot a mere modulatory system but an integral part of the substrate
mediating the NWR. Indeed, it has been demonstrated thatdescending modulation determines the modular organization andthe adaptability of the NWR circuit during the development ofthe CNS (Vecchierini-Blineau and Guihneuc, 1982; Levinssonet al., 1999; Waldenstrom et al., 2003; Hart and Giszter, 2004;Grillner, 2004). Moreover, spinal cord injury leads to expansionof the reflex receptive fields in humans (Andersen et al., 2004) indi-cating a pivotal role of the descending activity in maintaining func-tionally optimal spinal reflex pathways. In this light, the spinalreorganization in chronic hemiplegic patients indicates that theplastic rearrangement of the substrate mediating the NWR isdetermined by new functional brain-spinal cord connections.
The reflexes in the shoulder joint seem to be different from theelbow and wrist joints and muscles. Indeed, the patients seemed tomaintain a certain differential distribution of the reflex probabilityin the AD and PD muscles (Fig. 6). In addition, the shoulder kine-matic response was either an abduction or an adduction duringmovement. Thus, in our patient series the shoulder seemed to con-serve, in some respects, the EMG and kinematic reflex behavior ob-served in healthy subjects. This indicates that the plastic changesin dynamic brain-spinal cord functional connections in post-strokepatients are expressed in a different way according to the specificrole of each single muscle and joint. It is known that the CNS as-signs an important role to the shoulder joint in executing grasp-ing-reaching movements. Hence, a pre-programmed control ofthe movement is exerted mainly by the shoulder muscles deter-mining a ‘‘shoulder-centred pattern’’ during arm reaching move-ments (Soechting and Flanders, 1989; Sainburg and Kalakanis,2000). It has been demonstrated that hemiparetic patients may de-velop alternative grasping strategies, using less elbow extensionand shoulder flexion for reaching (Roby-Brami et al., 1997) andincreasing the redundancy of the motor system, particularly as aresult of recruitment of excessive trunk movement (Roby-Bramiet al., 1997; Cirstea and Levin, 2000).
Given the motor task performed in the present study, in whichthe trunk was restrained but the patients were able to perform thefull range of shoulder movements, it can be speculated that hemi-paretic patients use the shoulder as the leading joint (Dounskaia,2005) and that particular attention should be paid to rehabilitationand recovery of the shoulder (Michaelsen et al., 2001; Roby-Bramiet al., 2003).
6.4. Comparisons with previous studies
To our knowledge, no previous studies have been performed onthe upper limb NWR in hemiparetic and spastic patients duringmovement. Thus, our data can be compared only with investiga-tions on the human upper limb at rest (Dewald et al., 1999) oron the human lower limb at rest (Dimitrijevic and Nathan, 1971;Meinck et al., 1985; Shahani and Young, 1971, 1980; Bhagwanet al., 1971; Bussel et al., 1989; Hornby et al., 2004) and duringwalking (Spaich et al., 2006).
Dewald et al. (1999) recorded EMG, reflex forces, and joint tor-ques from the arms of eight brain-injured hemiparetic patientswith different degrees of spasticity at rest. The authors found laterreflex onset both of EMG activity and of the associated shoulderand elbow torques relative to the unimpaired limb. A systematicchange in the spatial distribution of muscle activation in the im-paired limb and changes in the mechanical response patterns werealso observed (Dewald et al., 1999). This finding fits well with thepresent observations concerning the lack of differential spatial andtemporal distribution of the reflex probability in the different mus-cles and joints in patients compared to controls.
With regard to the lower limbs a reduction of the early compo-nent of the NWR has been observed in numerous studies (Dimitrij-evic and Nathan, 1971; Fisher, 1979; Bhagwan et al., 1971; Shahani

M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540 539
and Young, 1980) while the late component is preserved anddesynchronized (Meinck et al., 1985). Meinck et al. (1985) investi-gated the NWR in detail in the upper motor neuron (UMN)syndrome revealing a net increase in reflex activity, de-syncroniza-tion, an abnormal sensitivity to reflex facilitation and irradiation tomuscles not normally involved in the reflex, irrespective of the siteof the UMN, spinal cord, brainstem or cerebrum. On the otherhand, other authors found differences in NWR following spinalcord transection and cerebral hemisphere lesions (Shahani andYoung, 1980). In our study, we did not differentiate between theearly and late components of the NWR. Several researchersreported the appearance of the early and late components in thelower limbs (mainly in the biceps femoris or tibialis anteriormuscles) with latencies in ranges of 56–65 ms for the early andof 85–120, 90–130, 90–180 ms for the late (for review see Sandriniet al., 2005).
In the upper limbs (biceps, triceps, flexor and extensor carpiradialis muscles) withdrawal responses with similar latencies wereobserved (Floeter et al., 1998). Considering that we used a timewindow of 60–200 ms to detect the reflex activity, the reflexes ob-served in the present study should be interpreted as related to thelate component of the NWR. From this perspective, the higher re-flex probabilities and reflex sizes and the spreading of the reflexactivity among antagonist muscles are findings similar to those ob-served by Meinck et al. on the UNM in lower limbs.
In a more recent study, executed on hemiparetic stroke patientsduring gait, Spaich et al. (2006) revealed a mixed behavior of bothEMG and kinematic reflex responses related to the role assigned tothe given muscles and joints during gait. Indeed, they observed alarger reflex in the tibialis anterior, as well as increased ankle dor-si-flexion at heel-off and during mid-swing, enlarged soleus reflexreceptive fields, smaller responses in the vastus lateralis and bicepsfemoris muscles, and in contralateral muscles and joints.
Although it is not easy to compare upper and lower limbs, theseauthors’ results, with regard to the differential distribution of theabnormalities among joints, are in accordance with the presentdata which show a main involvement of the elbow and wrist jointswith respect to the shoulder.
7. Conclusions
Patients with chronic and mild-moderate post-stroke motordeficits lose the ability to modulate the NWR dynamicallyaccording to the movement variables at individual as well as atmulti- and coordinated muscle and joint levels. These findingsmay indicate an inability of the central nervous system to usethe NWR substrate dynamically and flexibly in order to select themuscle synergies needed to govern the spatio-temporal interactionamong joints. They may also explain some of the typical movementabnormalities observed in hemiparetic patients during voluntarymotor activities.
References
Andersen OK, Jensen LM, Brennum J, Arendt-Nielsen L. Modulation of the humannociceptive reflex by cyclic movements. Eur J Appl Physiol Occup Physiol1995;70:311–21.
Andersen OK, Finnerup NB, Spaich EG, Jensen TS, Arendt-Nielsen L. Expansion ofnociceptive withdrawal reflex receptive fields in spinal cord injured humans.Clin Neurophysiol 2004;115:2798–810.
Andersen OK. Studies of the organization of the human nociceptive withdrawalreflex. Focus on sensory convergence and stimulation site dependency. ActaPhysiol (Oxf) 2007;654:1–35.
Aruin AS. Support-specific modulation of grip force in individuals with hemiparesis.Arch Phys Med Rehabil 2005;86:768–75.
Ashworth B. Preliminary trial of carisoprodol in multiple sclerosis. Practitioner1964;192:540–2.
Balasubramaniam R, Feldman AG. Guiding movements without redundancyproblems. In: Jirsa VK, Kelso JAS, editors. Coordination dynamics : issues andtrends. New York–Berlin–Heidelberg: Springer; 2004. p. 531–6.
Baldissera F, Hultborn H, Illert M. Integration in the spinal cord. In: Brookhart JM,Mountcastle VB, Brooks VB, Geiger SR, editors. Handbook of physiology, section1, vol II. Bethesda: American Physiological Society; 1981. p. 509–95.
Beer RF, Dewald JP, Rymer Z. Deficits in the coordination of multijoint armmovements in patients with hemiparesis: evidence for disturbed control oflimb dynamics. Exp Brain Res 2000;131:305–19.
Bhagwan T, Shahani BT, Young RR. Human flexor reflexes. J Neurol NeurosurgPsychiatry 1971;34:616–27.
Black I, Nichols D, Pelliccio M, Hidler J. Quantification of reflex activity in strokesurvivors during an imposed multi-joint leg extension movement. Exp Brain Res2007;183:271–81.
Bussel B, Roby-Brami A, Yakovleff A, Bennis N. Late flexion reflex in paraplegicpatients. Evidence for a spinal stepping generator. Brain Res Bull 1989;22:53–6.
Crenna P, Frigo C. Evidence of phase-dependent nociceptive reflexes duringlocomotion in man. Exp Neurol 1984;85:336–45.
Cirstea MC, Levin MF. Compensatory strategies for reaching in stroke. Brain2000;123:940–53.
Condliffe EG, Clark DJ, Patten C. Reliability of elbow stretch reflex assessment inchronic post-stroke hemiparesis. Clin Neurophysiol 2005;116:1870–8.
Corcos DM, Gottlieb GL, Penn RD, Myklebust B, Agarwal GC. Movement deficitscaused by hyperexcitable stretch reflexes in spastic humans. Brain1986;109:1043–58.
Cruse H, Brüwer M, Dean J. Control of three- and four-joint arm movement:strategies for a manipulator with redundant degrees of freedom. J Mot Behav1993;25:131–9.
Dean J, Brüwer M. Control of human arm movements in two dimensions: paths andjoint control in avoiding simple linear obstacles. Exp Brain Res1994;97:497–514.
Dewald JP, Beer RF, Given JD, McGuire JR, Rymer WZ. Reorganization of flexionreflexes in the upper extremity of hemiparetic subjects. Muscle Nerve1999;22:1209–21.
Dimitrijevic MR, Nathan PW. Studies of spasticity in man. Dishabituation of theflexion reflex in spinal man. Brain 1971;94:77–90.
Don R, Pierelli F, Ranavolo A, Serrao M, Mangone M, Paoloni M, et al. Modulation ofspinal inhibitory reflex responses to cutaneous nociceptive stimuli duringupper limb movement. Eur J Neurosci 2008;28:559–68.
Dounskaia N. The internal model and the leading joint hypothesis: implications forcontrol of multi-joint movements. Exp Brain Res 2005;166:1–16.
Duysens J, Tax AA, Trippel M, Dietz V. Ipsilateral reflex reversal during humanlocomotion suggests that the biceps femoris reflex is not a simple flexion reflex.In: Brandt T, Paulus W, Bles W, Dieterich M, Krafczyk S, Straube A, editors.Disorders of posture and gait. Stuttgart: Georg Thieme Verlag; 1990. p. 124–7.
Duysens J, Tax AA, Trippel M, Dietz V. Phase-dependent reversal of reflexly inducedmovements during human gait. Exp Brain Res 1992;90:404–14.
Emborg J, Spaich EG, Andersen OK. Withdrawal reflexes examined during humangait by ground reaction forces: site and gait phase dependency. Med Biol EngComput 2009;47:29–39.
Fisher MJ. Electrophysiologic analysis of the motor system after stroke: the flexorreflex. Arch Phys Med Rehabil 1979;60:7–11.
Floeter MK, Gerloff C, Kouri J, Hallett M. Cutaneous withdrawal reflexes of the upperextremity. Muscle Nerve 1998;21:591–8.
Fugl-Meyer AR, Jääskö L, Leyman I, Olsson S, Steglind S. The post-stroke hemiplegicpatient: I. A method for evaluation of physical performance. Scand J RehabilMed 1975;7:13–31.
Gölge M, Müller M, Dreesmann M, Hoppe B, Wenzelburger R, Kuhtz-Buschbeck JP.Recovery of the precision grip in children after traumatic brain injury. Arch PhysMed Rehabil 2004;85:1435–44.
Gowland C, deBruin H, Basmajian JV, Plews N, Burcea I. Agonist and antagonistactivity during voluntary upper-limb movement in patients with stroke. PhysTher 1992;72:624–33.
Grichting B, Hediger V, Kaluzny P, Wiesendanger M. Impaired proactive andreactive grip force control in chronic hemiparetic patients. Clin Neurophysiol2000;111:1661–71.
Grillner S. Muscle twitches during sleep shape the precise muscles of thewithdrawal reflex. Trends Neurosci 2004;27:169–71.
Haggard P, Hutchinson K, Stein J. Patterns of coordinated multi-joint movement.Exp Brain Res 1995;107:254–66.
Hammond MC, Kraft GH, Fitts SS. Recruitment and termination ofelectromyographic activity in the hemiparetic forearm. Arch Phys MedRehabil 1988;69:106–10.
Hart CB, Giszter SF. Modular premotor drives and unit bursts as primitives for frogmotor behaviors. J Neurosci 2004;24:5269–82.
Hepp-Reymond MC, Wiesendanger M. Unilateral pyramidotomy in monkeys: effecton force and speed of a conditioned precision grip. Brain Res 1972;36:117–31.
Hermens HJ, Freriks B, Disselhorst-Klug C, Rau G. Development of recommendationsfor SEMG sensors and sensor placement procedures. J Electromyogr Kinesiol2000;10:361–74.
Hornby TG, Tysseling-Mattiace VM, Benz EN, Schmit BD. Contribution of muscleafferents to prolonged flexion withdrawal reflexes in human spinal cord injury.J Neurophysiol 2004;92:3375–84.
Kadaba MP, Ramakrishnan HK, Wootten ME, Gainey J, Gorton G, Cochran GV.Repeatability of kinematic, kinetic, and electromyographic data in normal adultgait. J Orthop Res 1989;7:849–60.

540 M. Serrao et al. / Clinical Neurophysiology 123 (2012) 527–540
Levin MF, Feldman AG. The role of stretch reflex threshold regulation in normal andimpaired motor control. Brain Res 1994;657:23–30.
Levin MF, Selles RW, Verheul MH, Meijer OG. Deficits in the coordination of agonistand antagonist muscles in stroke patients: implications for normal motorcontrol. Brain Res 2000;853:352–69.
Levin MF. Interjoint coordination during pointing movements is disrupted in spastichemiparesis. Brain 1996;119:281–94.
Levinsson A, Garwicz M, Schouenborg J. Sensorimotor transformation in catnociceptive withdrawal reflex system. Eur J Neurosci 1999;11:4327–32.
Lundberg A. Multisensory control of spinal reflex pathways. In: Granit R, PomeianoO, editors. Reflex control of posture and movement, progress in brain research.Amsterdam: Elsevier; 1979. p. 11–28.
Maier MA, Bennett KM, Hepp-Reymond MC, Lemon RN. Contribution of the monkeycorticomotoneuronal system to the control of force in precision grip. JNeurophysiol 1993;69:772–85.
Meinck HM, Benecke R, Conrad B. Spasticity and flexor reflex. In: Delwaide PJ, YongRR, editors. Clinical neurophysiology in spasticity. Amsterdam: Elsevier SciencePublishers B.V. Biomedical Division; 1985. p. 41–54.
Michaelsen SM, Luta A, Roby-Brami A, Levin MF. Effect of trunk restraint on therecovery of reaching movements in hemiparetic patients. Stroke2001;32:1875–83.
Muir RB, Lemon RN. Corticospinal neurons with a special role in precision grip.Brain Res 1983;261:312–6.
Nowak DA, Hermsdörfer J, Topka H. Deficits of predictive grip force control duringobject manipulation in acute stroke. J Neurol 2003;250:850–60.
Powers RK, Marder-Meyer J, Rymer WZ. Quantitative relations between hypertoniaand stretch reflex threshold in spastic hemiparesis. Ann Neurol1988;23:115–24.
Quaney BM, Perera S, Maletsky R, Luchies CW, Nudo RJ. Impaired grip forcemodulation in the ipsilesional hand after unilateral middle cerebral arterystroke. Neurorehabil Neural Repair 2005;19:338–49.
Rab G, Petuskey K, Bagley A. A method for determination of upper extremitykinematics. Gait Posture 2002;15:113–9.
Roby-Brami A, Fuchs S, Mokhtari M, Bussel B. Reaching and grasping strategies inhemiparetic patients. Motor Control 1997;1:72–91.
Roby-Brami A, Feydy A, Combeaud M, Biryukova EV, Bussel B, Levin MF. Motorcompensation and recovery for reaching in stroke patients. Acta Neurol Scand2003;107:369–81.
Raghavan P, Krakauer JW, Gordon AM. Impaired anticipatory control of fingertipforces in patients with a pure motor or sensorimotor lacunar syndrome. Brain2006;129:1415–25.
Rossi A, Decchi B. Flexibility of lower limb reflex responses to painful cutaneousstimulation in standing humans: evidence of load-dependent modulation. JPhysiol 1994;481:521–32.
Sainburg RL, Kalakanis D. Differences in control of limb dynamics during dominantand nondominant arm reaching. J Neurophysiol 2000;83:2661–75.
Sangani SG, Starsky AJ, McGuire JR, Schmit BD. Multijoint reflexes of the stroke arm:neural coupling of the elbow and shoulder. Muscle Nerve 2007;36:694–703.
Sandrini G, Serrao M, Rossi P, Romaniello A, Cruccu G, Willer JC. The lower limbflexion reflex in humans. Progr Neurobiol 2005;77:353–95.
Shahani BT, Young RR. Human flexor reflexes. J Neurol Neurosurg Psychiatry1971;34:616–27.
Shahani BT, Young RR. The flexor reflex in spasticity. In: Feldman RG, Young RR,Koella WP, editors. Spasticity: disordered motor control. Chicago: MedicalPublishers; 1980. p. 287–95.
Serrao M, Pierelli F, Don R, Ranavolo A, Cacchio A, Currà A, et al. Kinematic andelectromyographic study of the nociceptive withdrawal reflex in the upperlimbs during rest and movement. J Neurosci 2006;26:3505–13.
Soechting JF, Flanders M. Sensorimotor representations for pointing to targets inthree-dimensional space. J Neurophysiol 1989;2:582–94.
Spaich EG, Arendt-Nielsen L, Andersen OK. Modulation of lower limb withdrawalreflexes during gait: a topographical study. J Neurophysiol 2004;91:258–66.
Spaich EG, Hinge HH, Arendt-Nielsen L, Andersen OK. Modulation of the withdrawalreflex during hemiplegic gait: effect of stimulation site and gait phase. ClinNeurophysiol 2006;117:2482–95.
Takahashi CD, Reinkensmeyer DJ. Hemiparetic stroke impairs anticipatory controlof arm movement. Exp Brain Res 2003;149:131–40.
Thilmann AF, Fellows SJ. The time-course of bilateral changes in the reflexexcitability of relaxed triceps surae muscle in human hemiparetic spasticity. JNeurol 1991;238:293–8.
Trombly CA. Deficits of reaching in subjects with left hemiparesis: a pilot study. AmJ Occup Ther 1992;46:887–97.
van Vliet PM, Sheridan MR. Coordination between reaching and grasping in patientswith hemiparesis and healthy subjects. Arch Phys Med Rehabil2007;88:1325–31.
Vecchierini-Blineau MF, Guihneuc P. Lower limb cutaneous polysynaptic reflexes inthe child, according to age and state of waking or sleeping. J Neurol NeurosurgPsychiatry 1982;45:531–8.
Waldenstrom A, Thelin J, Thimansson E, Levinsson A, Schouenborg J. Developmentallearning in a pain-related system: evidence for a crossmodality mechanism. JNeurosci 2003;23:7719–25.
Wenzelburger R, Kopper F, Frenzel A, Stolze H, Klebe S, Brossmann A, et al. Handcoordination following capsular stroke. Brain 2005;128:64–74.
Willer JC. Comparative study of perceived pain and nociceptive flexion reflex inman. Pain 1977;3:69–80.
Wilson LR, Gandevia SC, Inglis JT, Gracies J, Burke D. Muscle spindle activity in theaffected upper limb after a unilateral stroke. Brain 1999;122:2079–88.
Related Documents











