Relationships between organohalogen contaminants and blood plasma clinical–chemical parameters in chicks of three raptor species from Northern Norway Christian Sonne a,n , Jan Ove Bustnes b , Dorte Herzke c , Veerle L.B. Jaspers d , Adrian Covaci d , Duncan J. Halley e , Truls Moum f , Igor Eulaers d , Marcel Eens d , Rolf A. Ims b , Sveinn A. Hanssen b , Kjell Einar Erikstad b , Trond Johnsen b , Lisbeth Schnug g , Frank F. Rig et a , Asger L. Jensen h a ˚ Arhus University, National Environmental Research Institute, Department of Arctic Environment, Frederiksborgvej 399, PO Box 358, DK-4000 Roskilde, Denmark b Norwegian Institute for Nature Research, Unit for Arctic Ecology, The Polar Environmental Centre, NO-9296 Tromsø, Norway c Norwegian Institute for Air Research, The Polar Environmental Centre, NO-9296 Tromsø, Norway d University of Antwerp, Ethology Research Group, Department of Biology and Toxicological Centre, B-2610 Antwerp, Belgium e Norwegian Institute for Nature Research, Department of Terrestrial Ecology, Tungasletta 2, NO-7485 Trondheim, Norway f Bodø University College, Faculty of Biosciences and Aquaculture, NO-8049 Bodø, Norway g Norwegian Institute for Agricultural and Environmental Research, Soil and Environment Division, Fr. A. Dahlsvei 20, NO-1432 ˚ As, Norway h Department of Basic Animal and Veterinary Sciences, Faculty of Life Sciences, University of Copenhagen, Frederiksberg, Denmark article info Article history: Received 3 April 2009 Received in revised form 25 August 2009 Accepted 29 August 2009 Available online 3 October 2009 Keywords: ALAT ALKP Cholesterol DDT GGT Golden eagle Goshawk PCBs Uric acid White-tailed eagle abstract Organohalogen contaminants (OHCs) may affect various physiological parameters in birds including blood chemistry. We therefore examined blood plasma clinical–chemical parameters and OHCs in golden eagle, white-tailed eagle and goshawk chicks from Northern Norway. Correlation analyses on pooled data showed that alkaline phosphatase (ALKP), glucose and creatinine were significantly negatively correlated to various OHCs (all: p o0.05; r: 0.43 to 0.55; n = 23), while alanine aminotransferase (ALAT), total protein, cholesterol, uric acid, total bilirubin, ratios protein:creatinine and uric acid:creatinine were significantly positively correlated to various OHCs (all: p o0.05; r: 0.43– 0.96). Based on these relationships, we suggest that the OHC concentrations found in certain raptor chicks of Northern Scandinavia may impact blood plasma biochemistry in a way that indicates impacts on liver, kidney, bone, endocrinology and metabolism. In order to elaborate further on these relationships and mechanisms, we recommend that a larger study should take place in the near future. & 2009 Elsevier Inc. All rights reserved. 1. Introduction Homeostasis is important for all vertebrate species in order to maintain the function of organ systems that are critical for survival and reproduction. Infectious diseases, genetic defects and environmental stressors are all factors that induce perturbations in blood biochemistry and in the homeostasis of vertebrate species in general (Braun, 2003; Harr, 2002; Richards and Proszkowiec-Weglarz, 2007; Schulz et al., 2000). Environmental stressors include extreme sound, temperatures, starvation, dehy- dration and environmental pollutants, such as organohalogen contaminants (OHCs) and heavy metals, that all have an impact on physiology and behaviour (AMAP, 1998, 2004; Chrousos and Gold, 1992; Johnson et al., 1992; Sharit and Salvendy, 1982). OHCs include polychlorinated biphenyls (PCBs), organochlorine pesticides (OCPs), brominated flame retardants (BFRs) and ARTICLE IN PRESS Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/ecoenv Ecotoxicology and Environmental Safety 0147-6513/$ - see front matter & 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.ecoenv.2009.08.017 n Corresponding author. Fax: +45 4630 1914. E-mail addresses: [email protected] (C. Sonne), [email protected] (J.O. Bustnes), [email protected] (D. Herzke), [email protected] (V.L. Jaspers). [email protected] (A. Covaci), [email protected] (D.J. Halley), [email protected] (T. Moum), [email protected] (I. Eulaers), [email protected] (M. Eens), [email protected] (R.A. Ims), [email protected] (S.A. Hanssen), [email protected] (K. Einar Erikstad), [email protected] (T. Johnsen). [email protected] (L. Schnug), [email protected] (F.F. Rig et), [email protected] (A.L. Jensen). URL: http://www.neri.dk (C. Sonne). Ecotoxicology and Environmental Safety 73 (2010) 7–17

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
Ecotoxicology and Environmental Safety 73 (2010) 7–17
Contents lists available at ScienceDirect
Ecotoxicology and Environmental Safety
0147-65
doi:10.1
n Corr
E-m
adrian.c
(M. Een
lisbeth.s
URL
journal homepage: www.elsevier.com/locate/ecoenv
Relationships between organohalogen contaminants and blood plasmaclinical–chemical parameters in chicks of three raptor species fromNorthern Norway
Christian Sonne a,n, Jan Ove Bustnes b, Dorte Herzke c, Veerle L.B. Jaspers d, Adrian Covaci d,Duncan J. Halley e, Truls Moum f, Igor Eulaers d, Marcel Eens d, Rolf A. Ims b, Sveinn A. Hanssen b,Kjell Einar Erikstad b, Trond Johnsen b, Lisbeth Schnug g, Frank F. Rig�et a, Asger L. Jensen h
a Arhus University, National Environmental Research Institute, Department of Arctic Environment, Frederiksborgvej 399, PO Box 358, DK-4000 Roskilde, Denmarkb Norwegian Institute for Nature Research, Unit for Arctic Ecology, The Polar Environmental Centre, NO-9296 Tromsø, Norwayc Norwegian Institute for Air Research, The Polar Environmental Centre, NO-9296 Tromsø, Norwayd University of Antwerp, Ethology Research Group, Department of Biology and Toxicological Centre, B-2610 Antwerp, Belgiume Norwegian Institute for Nature Research, Department of Terrestrial Ecology, Tungasletta 2, NO-7485 Trondheim, Norwayf Bodø University College, Faculty of Biosciences and Aquaculture, NO-8049 Bodø, Norwayg Norwegian Institute for Agricultural and Environmental Research, Soil and Environment Division, Fr. A. Dahlsvei 20, NO-1432 As, Norwayh Department of Basic Animal and Veterinary Sciences, Faculty of Life Sciences, University of Copenhagen, Frederiksberg, Denmark
a r t i c l e i n f o
Article history:
Received 3 April 2009
Received in revised form
25 August 2009
Accepted 29 August 2009Available online 3 October 2009
Keywords:
ALAT
ALKP
Cholesterol
DDT
GGT
Golden eagle
Goshawk
PCBs
Uric acid
White-tailed eagle
13/$ - see front matter & 2009 Elsevier Inc. A
016/j.ecoenv.2009.08.017
esponding author. Fax: +45 4630 1914.
ail addresses: [email protected] (C. Sonne), jan.o.b
[email protected] (A. Covaci), duncan.halley@ni
s), [email protected] (R.A. Ims), sveinn.a.hans
[email protected] (L. Schnug), [email protected] (
: http://www.neri.dk (C. Sonne).
a b s t r a c t
Organohalogen contaminants (OHCs) may affect various physiological parameters in birds including
blood chemistry. We therefore examined blood plasma clinical–chemical parameters and OHCs in
golden eagle, white-tailed eagle and goshawk chicks from Northern Norway. Correlation analyses on
pooled data showed that alkaline phosphatase (ALKP), glucose and creatinine were significantly
negatively correlated to various OHCs (all: po0.05; r: �0.43 to �0.55; n=23), while alanine
aminotransferase (ALAT), total protein, cholesterol, uric acid, total bilirubin, ratios protein:creatinine
and uric acid:creatinine were significantly positively correlated to various OHCs (all: po0.05; r: 0.43–
0.96). Based on these relationships, we suggest that the OHC concentrations found in certain raptor
chicks of Northern Scandinavia may impact blood plasma biochemistry in a way that indicates impacts
on liver, kidney, bone, endocrinology and metabolism. In order to elaborate further on these
relationships and mechanisms, we recommend that a larger study should take place in the near future.
& 2009 Elsevier Inc. All rights reserved.
1. Introduction
Homeostasis is important for all vertebrate species in order tomaintain the function of organ systems that are critical forsurvival and reproduction. Infectious diseases, genetic defects andenvironmental stressors are all factors that induce perturbationsin blood biochemistry and in the homeostasis of vertebratespecies in general (Braun, 2003; Harr, 2002; Richards and
ll rights reserved.
[email protected] (J.O. Bustnes), dor
na.no (D.J. Halley), Truls.Moum@h
[email protected] (S.A. Hanssen), kjell.e
F.F. Rig �et), [email protected] (A.L. Jens
Proszkowiec-Weglarz, 2007; Schulz et al., 2000). Environmentalstressors include extreme sound, temperatures, starvation, dehy-dration and environmental pollutants, such as organohalogencontaminants (OHCs) and heavy metals, that all have an impacton physiology and behaviour (AMAP, 1998, 2004; Chrousosand Gold, 1992; Johnson et al., 1992; Sharit and Salvendy, 1982).OHCs include polychlorinated biphenyls (PCBs), organochlorinepesticides (OCPs), brominated flame retardants (BFRs) and
[email protected] (D. Herzke), [email protected] (V.L. Jaspers).
ibo.no (T. Moum), [email protected] (I. Eulaers), [email protected]
[email protected] (K. Einar Erikstad), [email protected] (T. Johnsen).
en).

ARTICLE IN PRESS
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–178
perfluorinated compounds (PFCs). Due to transport mechanismsand chemical and biological properties, these compounds areretained within organisms and tend to biomagnify at the top ofmarine and terrestrial food webs (Muir et al., 1992; Norstrom andMuir, 1994; Rocca and Mantovani, 2006). OHCs are present in themarine environments of the Northern Hemisphere and areparticularly high in industrialised areas, such as the Baltic Sea.However, OHCs are also transported by atmospheric and seacurrent mechanisms from lower latitudes, such as Eurasia andNorth America, toward the north (AMAP, 1998, 2004; Colborn,2004; Macdonald et al., 2003, 2005) and consequently lipidsoluble organohalogen pollutants biomagnify in the marine foodchains of the Arctic. Top predators, such as polar bears (Ursus
maritimus) and raptors, accumulate high levels of OHC residuesthat may have adverse health impacts (AMAP, 1998, 2004).
Raptors including white-tailed eagles (Haliaeetus albicilla),golden eagles (Aquila chrysaetos) and northern goshawks (Accipiter
gentilis) are top predators of either marine or terrestrial foodwebs. As a result, significant OHC concentrations are found inspecific birds of prey (Colborn, 1991; Donaldson et al., 1999;Herzke et al., 2002, 2005). For example, white-tailed eagles relymainly on marine fish-eating seabird species, such as greatcormorant (Phalacrocorax carbo) and several gull species (Laridae)and also feed on a variety of marine fish species of which themajority is picked up from gulls or fishing vessels (Willgohs,1961). Studies have shown that several marine bird species andfish from Northern Norway are heavily polluted with OHCs(Bustnes et al., 2008; Helberg et al., 2005; Julshamn et al.,2004), which subsequently accumulate in eagles and hawks(Gjershaug et al., 2008). Coastal goshawks and golden eagles canalso prey on seabirds, but usually rely on terrestrial food sources.The primary main prey species in Northern Norway includeamongst others redwing (Turdus iliacus), fieldfare (Turdus pilaris),carrion crow (Corvus corone cornix), grouse species (Lagopus spp.;Tetrao spp.) and small mammals such as mountain hare (Lepus
timidus) and red squirrels (Sciurus vulgaris) (Grønnesby andNygard, 2000). The golden eagle’s diet is dominated by grouse,with mountain hares and reindeer (Rangifer tarandus) as im-portant secondary sources, the latter often found as carrion(Johnsen et al., 2007; Halley et al., 2007). Terrestrial prey speciesare all relatively low in OHCs compared with food from themarine environment (AMAP, 1998, 2004). Therefore, raptors thathave specialised on the terrestrial food web are likely to be lowerin OHCs compared with the marine predator species (Gjershauget al., 2008; Herzke et al., 2002, 2005).
The mode of toxic action of OHCs on vertebrate organisms isvariable and includes responses of neuro-endocrine pathwayssuch as the hypothalamic–hypophysis–target organ axis and up-regulation of liver cytochrome (CYP) metabolism (Bandiera et al.,1997; Grasman et al., 2000; Henriksen et al., 2000; Kennedy et al.,2003; Letcher et al., 1996; Østby et al., 2005). Briefly, thesechanges lead to disruptions of specific endogenous peptide andsteroid hormones like thyroid hormones T3/T4, estrogens andtestosterone with fatal consequences for development, reproduc-tive success and immune competence (Colborn, 2004; Darnerudet al., 2001; Erikstad et al., 2009; Guvenius et al., 2002; Hamerset al., 2006; Harju et al., 2007; Letcher et al., 2000; Verreault et al.,2004, 2006, 2007). A fatal impact on raptor species is a thinning ofegg-shells that subsequently results in low reproductive rates dueto fragile/porous eggs although the exposure that caused this toxicresponse seems to have decreased over the last 10 years (Elliottand Martin, 1994; Falk et al., 2006; Helander et al., 2002;Scharenberg and Looft, 2004). Moreover, studies of birds andmammals have shown that OHCs have a toxic impact on liver andkidney function and the general homeostasis measured by bloodclinical–chemical parameters (Dieter et al., 1977; Fischbein, 1985;
Hayes et al., 1984; Kutlu et al., 2007). Due to the low endocrineplasticity and low homeostatic buffering capacity at the foetal andneonatal stages, these effects are probably most severe at thesedevelopmental stages (Damstra et al., 2002; Grandjean andLandrigan, 2006).
To further investigate the potential OHC impacts on the healthof raptors, we conducted a study on blood plasma clinical–chemical parameters in golden eagle, white-tailed eagle andgoshawk chicks in the Troms and Finnmark counties of NorthernNorway (Fig. 1). The study species and nest locations were chosenin order to have breeding pairs and chicks that were supposedlyhigh (white-tailed eagles breeding in the coastal environment)and low (terrestrial goshawks and golden eagles) in their OHCbody burdens due to their reliance on the marine versusterrestrial food webs in this area (Cramp and Simmons, 1980).The present study reports on the relationship between bloodplasma OHCs and biochemical parameters and thereby thepotential health impacts on the chicks of these three raptorspecies.
2. Materials and methods
The study was conducted on white-tailed eagles and northern goshawks from
Troms County, and golden eagles from Finnmark County, Northern Norway, from
April to the end of June 2008. The study area ranged from 691 to 711N and from 181
to 261E (Fig. 1). Nests of the three species were checked for breeding activity from
late March to the middle of May using binoculars and telescopes, whilst keeping a
distance to avoid disturbing the breeding pairs. The presence of at least one bird
lying on the nest was used as a confirmation of breeding activity.
2.1. Study design and sampling
In winter 2008 (January–April), a large number of nests of all three species
were visited and birds with territorial behaviour were recorded. In late April and
early May it was determined if the birds had laid eggs. In May–June nestlings in
successful nests were inspected for the first time, and a second time at beginning of
July shortly before fledging, approximately 2 weeks later. Due to variation in
hatching date, the first visit took place at a nestling age of between 1 and 3 weeks.
The nestlings (2 golden eagles, 16 goshawks and 5 white-tailed eagles) were
lowered from the nest in a nylon bag and blood (0.1–4.0 mL) was sampled from the
brachial vein during the second visit after which the chicks were returned to their
nests. Blood was centrifuged at 8000 rpm for 10 min and 1 mL supernatant plasma
was transferred to a sterile 1.5 mL Eppendorfs tube and frozen at �20 1C the same
day prior to analyses. The study was approved by the National Animal Research
Authority of Norway.
2.2. Analyses of blood plasma clinical–chemical parameters
The analyses were conducted at the Central Clinical Laboratory at the
Department of Small Animal Clinical Sciences, Faculty of Life Sciences, University
of Copenhagen and included the following 19 components: albumin (Alb; g/L),
glucose (Glu; mmol/L), total protein (TP; g/L), alkaline phosphatase (ALKP; U/L),
alanine aminotransferase (ALAT; U/L), total bilirubin (TB; mmol/L), fructosamine
(Fructo; mmol/L), cholesterol (Cho; mmol/L), creatinine (Cre; mmol/L), inorganic
phosphate (Iph; mmol/L), bile acids (BA; mmol/L), amylase (Amy; U/L), urea (Urea;
mmol/L), gamma glutamyl transferase (GGF; U/L), calcium (Ca; mmol/L),
magnesium (Mg; mmol/L), uric acid (UA; U/L), sodium (Na; mmol/L) and
potassium (K; mmol/L). The analyses were routinely conducted at the laboratory
using an automated spectrophotometrical analyser also containing ion-selective
electrodes (ADVIA 1800, Siemens). All assays were subjected to daily internal and
quarterly external quality control. Only results from accepted analytical runs are
reported here. Information on methods can be found at the Department of Small
Animal Clinical Sciences (http://www.life.ku.dk).
2.3. OHC analyses
Details regarding the analyses can be found in Herzke et al. (2005) and Gotsch
et al. (2004). Briefly, plasma samples from nestlings were analyzed for a set of OCs
(organochlorines: PCBs [polychlorinated biphenyls: CB-18, CB-28, CB-99, CB-101,
CB-105, CB-118, CB-138, CB-153, CB-180, CB-183, CB-187, CB-194], p,p0-DDE
[dichlorodiphenyldichloroethylene], b-HCH [hexachlorohexane], HCB [hexachlor-
obenzene], heptachlor epoxide, CHLs [chlordanes: t-chlordane, c-chlordane,

ARTICLE IN PRESS
Fig. 1. Map identifying Troms and Finnmark provinces in Northern Norway from which we obtained blood plasma samples of golden eagles (n=2), goshawks (n=16) and
white-tailed eagles (n=5).
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–17 9
oxy-chlordane, t-Nonachlor, c-Nonachlor] and Mirex), PBDEs (polybrominated
diphenylethers: BDE-28, BDE-47, BDE-99, BDE-100, BDE-153, BDE-154), TBA
(tribromoanisole) and perfluorinated compounds (PFCs: PFHpS, PFOS, PFDcS,
PFPA, PFOA, PFNA, PFDcA, PFUnA, PFDoA, PFTriA). The analyses of environmental
contaminants in blood plasma were carried out at the Norwegian Institute of Air
Research (Norsk Institutt for Luftforskning, NILU) in Tromsø and followed standard
procedures. Briefly, in order to extract OHCs from the plasma, an aliquot of each
sample was denatured with saturated ammonium sulphate solution and ethanol
after the addition of an internal standard solution (13C-labelled compounds). The
solution was extracted twice with n-hexane and the combined extract was further
cleaned up by Florisil column chromatography. To all samples, octachloronaphta-
lene was added as recovery standard prior to analyses. Samples were quantita-
tively analyzed by gas chromatography coupled with a mass selective detector
(GC/MS). For quantification of all compounds, crystalline reference standards was
obtained from Cambridge Isotope Laboratories (Woburn, MA, USA). Solvents of
pesticide grade were employed (E. Merck, Darmstadt, Germany). 13C-labeled
compounds were used as internal standards representing each group of analytes,
containing between 77 and 500 pg/mL of 13C-p,p0-DDE, 13C-p,p0-DDT, 13C-labeled
CB-28, -52, -101, -118, -153 and -180, 13C-labeled trans-nonachlor, HCB and b-HCH.
The results were not corrected for recovery. PBDEs and TBA were analysed by
applying negative chemical ionisation (NCI) in the GC/MS. The masses m/z 79
and 81 were analysed for brominated compounds in addition to comparison
of retention times of standard compounds for each individual PBDE congener. 13C-
labeled CB-118 was used for quantification.
Prior to PFC analysis, ca. 0.2 g of plasma was fortified with internal standard13C-labeled PFOS and PFOA (Wellington, US) and extracted with 1 mL acetonitrile
for three times 10 min in an ultrasonic bath. Vortex was applied in between the
application of the ultrasound. After centrifugation, 1 mL of the supernatant
solution was added to 25 mg ENVI-Carb and 50 mL glacial acetic acid and vortexed
thoroughly. After additional centrifugation at 10,000 rpm for 10 min, 0.5 mL of the
solution was transferred in an autoinjector vial. Prior to analysis, 3,7-dimethyl-
perfluorooctanoic acid (3,7-di-methyl-PFOA, 97%, ABCR; Germany) and 0.5 mL of a
22 mM ammonium acetate solution was added to each sample. We analysed the
ionic PFCs: perfluorocarboxyl acids (PFCA) with a carbon chain containing 4–13
carbons, as well as perfluorosulphonates (PFS) as perfluorobutane-, perfluorohex-
ane-, perfluorooctane-, and perfluordecane-sulphonate in addition to perfluor-
ooctane sulfonamide (PFOSA). All standards were purchased as crystalline
substances from Aldrich, Fluka and ABCR with purities greater than 95%. Aliquots

ARTICLE IN PRESS
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–1710
(25 mL) were injected automatically on a HPLC (Agilent 1100; Agilent Technologies,
Palo Alto, CA) coupled to ESI time-of-flight-high-resolution MS in the negative ion
mode (HPLC–ToF–HRMS ESI) (LCT, Micromass, Manchester, England). The values
for the pollutant concentrations are all given in pg/g wet weight (ww).
2.4. Statistical analyses
SAS statistical software package (SAS 9.1 and enterprise guide V4.0, SAS
Institute Inc., Cary, NC, USA) was used for the initial statistical analyses. All OHC
and blood clinical–chemical parameters (BCCP) data were log-transformed to meet
the requirement of equal variance and homogeneity of the data distribution
between groups (Zar, 1984), and the level of significance was set to a=0.05, while
0.05opo0.1 was considered a trend. First, a non-parametric Kruskal–Wallis test
was applied to test for differences in mean concentrations of OHCs (P
PCB, p,p0-
DDE, b-HCH, HCB, heptachlor epoxide, chlordanes [P
CHL], Mirex,P
PBDE, TBA
andP
PFC) and BCCPs between species, which is applicable despite the low
samples size of 2, 5, 16 individuals and the relative uncertainty of the w2
approximation according to Siegel (1988). Shapiro–Wilk tests showed that pooled
data (n=23) were normally distributed and therefore Pearson’s correlation analysis
was applied to test for the relationships between OHCs and BCCPs. Only in cases
with species differences in OHC and BCCP concentrations the correlation analyses
were split into each species; and subsequently the non-parametric Spearman rank
test was applied to white-tailed eagles. The statistics were not Bonferroni
corrected in order not to introduce type II errors (i.e. rejecting true null-
hypothesis) (Perneger, 1998) and as each specific OHCs vs. BCCPs was of clinical/
biological interest rather than just the pattern.
A principal component analysis (PCA) was applied to investigate the patterns
of BCCPs in the three raptorial bird species. The PCA was performed on the
correlation matrix and differences in mean scores of the PCA components between
bird species were tested by analysis of variance (ANOVA) followed by Tukey’s
multiple comparisons of means.
The multiple relationships between contaminants (OHCs) and BCCPs were
analysed by redundancy analysis (RDA). The RDA expresses how much of the
variance in one data set (BCCPs) can be explained by the other data set (OHCs). The
RDA is a PCA of the predicted values of a multivariate regression with BCCPs as
dependent variables and OHCs as independent variables. For a detailed description
of RDA, see Ter Braak and Prentice (1988). Because of the relative low number of
samples, the variables included in the analysis were limited to the following: ALKP,
ALAT, amylase, glucose, albumin, total protein, cholesterol, bile acid, uric acid, total
bilirubin, magnesium, potassium, creatinine, SPCB, p,p0-DDE, b-HCH, HCB,
heptachlor epoxide, SCHL, Mirex, TBA, SPBDE and SPFC. The PCAs and RDAs
were performed using the free software R version 2.8.1.
3. Results
3.1. Blood plasma clinical–chemical parameters
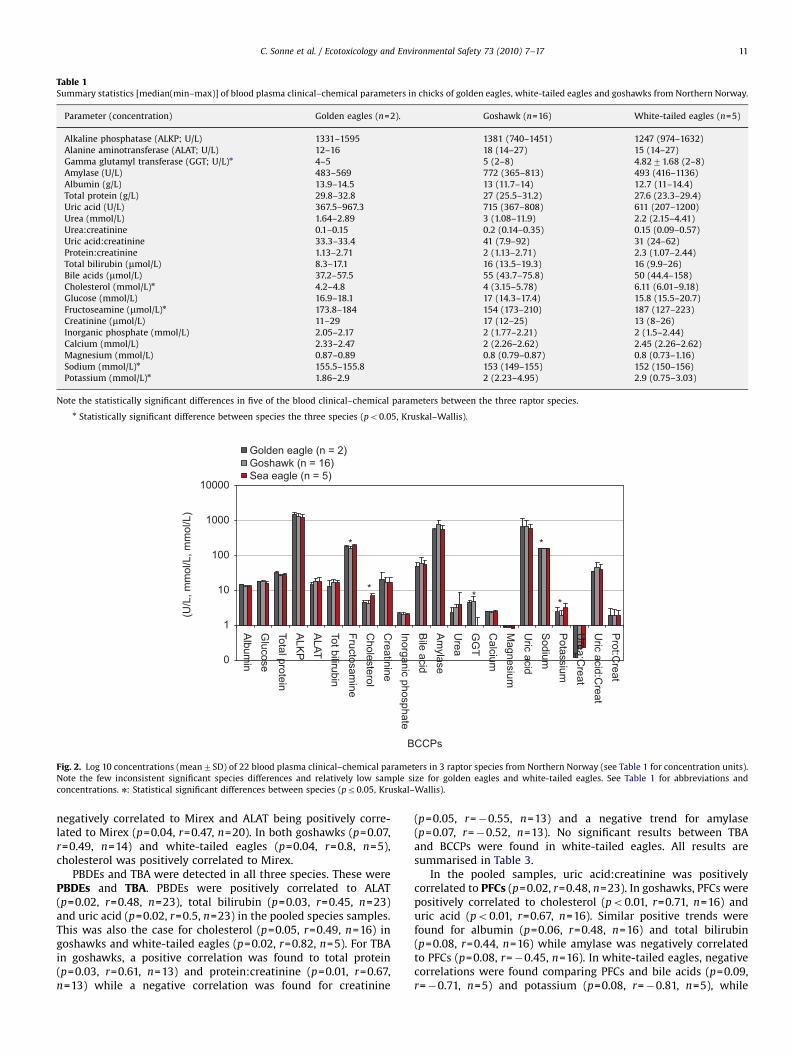
Twenty-two blood plasma clinical–chemical parameters(BCCPs) were measured in all 23 raptor specimens (Table 1). Insummary, these were composed of 3 liver enzymes, 1 digestiveenzyme, 2 protein groups, erythrocyte metabolism wasteproducts, cholesterol, 2 carbohydrates, creatinine (muscleprotein metabolism) and 5 electrolytes/minerals. In addition,protein:creatinine, urea:creatinine and uric acid:creatinine wereintroduced as supportive parameters for renal functioning. Ingeneral, the species differences were few and inconsistent,although fructosamine, cholesterol, gamma glutamyl transferase,sodium and potassium differed significantly between species(Kruskal–Wallis: all po0.05 and w246.6) (Table 1, Fig. 2).Therefore, the statistical analyses of OHCs versus BCCPs wereconducted separately for these five BCCPs within goshawks(Pearson’s correlation analysis) and white-tailed eagles(Spearman rank test). For the remaining 17 BCCPs, data werepooled across species in order to increase the statistical power ofthe analyses with an ntotal=23.
3.2. OHC concentrations
Ten distinct OHC classes/types were measured in blood plasmaof all three raptor species (Table 2). These were
PPCB, p,p0-DDE,
b-HCH, HCB, heptachlor epoxide, chlordanes (P
CHL), Mirex,
PPBDE, TBA and
PPFC. The mean blood plasma OHC
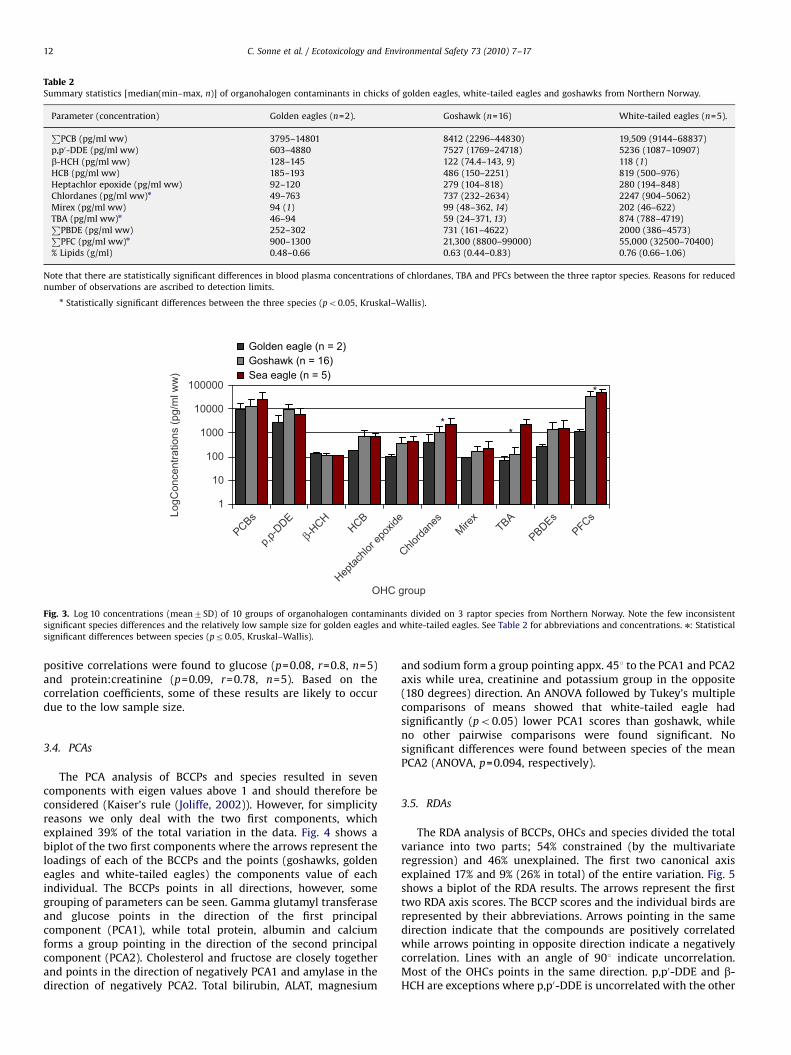
concentrations (pg/mL ww) are shown for each species in Fig. 3.It is seen that the most dominating OHC groups were PCBs, PFCsand p,p0-DDE and the concentrations generally increased white-tailed eagles ‘‘goshawks’’ golden eagles except for p,p0-DDE andb-HCH. In all three species, the most dominating PCB congenerswere the higher chlorinated CB-138, �153 and �180 whilefor PBDEs it was BDE-47, �99 and �100. Oxy-chlordane andt-nonachlor were the chlordanes found in highest concentrationsin all three species. PFOS was the most dominating fluorinatedcompound and, together with PFNA, the only PFC found in allspecimens. PFPA was only found in a single goshawk, while PFDcSwas found in three goshawks and one white-tailed eagle. The PFCsincreased in the following order: PFDcS�PFHpS)PFDoA�PFTriA�PFPA)PFDcA�PFOA)PFUnA)PFNA)PFOS. From Table 2 itcan be seen that the OHC difference between white-tailed eaglesversus goshawks is ranging up to a factor ca. 1.5–15 (only p,p0-DDEwas lowest in the white-tailed eagles), whilst golden eagleconcentrations for some compounds were comparable to thoseof goshawks (e.g. Mirex) and a factor 20–60 lower than bothgoshawks and white-tailed eagles (e.g. PFCs). There were,however, only significant differences in concentrations ofchlordanes, TBA and PFCs between species with white-tailedeagles carrying the highest concentrations followed by goshawksand golden eagles, respectively (Kruskal–Wallis: all po0.05 andw246.2) (Table 2, Fig. 3). Therefore, the statistical analyses ofOHCs vs. BCCPs were conducted separately for chlordanes, PFCsand TBA within each species. For the remaining 7 OHCs, data werepooled across species in order to increase the statistical power ofthe analyses with an ntotal=23.
3.3. BCCPs versus contaminant concentrations
Due to species-specific differences, the relationships betweenchlordanes, PFCs, TBA and BCCPs, respectively, were evaluatedseparately for goshawks and white-tailed eagles. This was also thecase for OHCs and fructose, cholesterol, gamma glutamyltransferase, sodium and potassium, respectively. In case of goldeneagles the sample size was too small (n=2) to conduct anyseparate statistical analyses. All results are summarised in Table 3in which only significant differences and trends are given. Thepooled species samples showed a highly negative correlation ofglucose to PCBs (po0.01, r=�0.53, n=23). Specifically ingoshawks, cholesterol (p=0.002, r=0.72, n=16) was highlypositively correlated to PCBs and that was also the case forwhite-tailed eagles (p=0.03, r=0.85, n=5).
For p,p0-DDE, a positive correlation was found to the pooledsamples of alanine aminotransferase (ALAT) (p=0.01, r=0.52,n=23), total bilirubin (p=0.03, r=0.46, n=23) and protein:creati-nine (p=0.04, r=0.43, n=23). HCB was positively correlated to uricacid (p=0.005, r=0.57, n=23) and uric acid:creatinine (p=0.003,r=0.58, n=23). Specifically in goshawk, HCB was positivelycorrelated to cholesterol (po0.01, r=0.74, n=23). Heptachlorepoxide showed a positive correlation with uric acid:creatinine(p=0.04, r=0.43, n=23). Specifically in goshawks, heptachlorepoxide was positively correlated to cholesterol (po0.01,r=0.63, n=16) and a similar positive trend was found for totalprotein (p=0.06, r=0.48, n=16).
In goshawks, cholesterol was highly positively correlated tochlordanes (po0.01, r=0.65, n=16) and a similar trend was foundfor total protein (p=0.07, r=0.47, n=16). In white-tailed eagles,chlordanes were positively correlated to cholesterol (p=0.04,r=0.82, n=5). Regarding Mirex, three significant results werefound in the pooled samples. These were glucose (p=0.05,r=�0.43, n=20) and ALKP (p=0.04, r=�0.45, n=20) being
ARTICLE IN PRESS
Table 1Summary statistics [median(min–max)] of blood plasma clinical–chemical parameters in chicks of golden eagles, white-tailed eagles and goshawks from Northern Norway.
Parameter (concentration) Golden eagles (n=2). Goshawk (n=16) White-tailed eagles (n=5)
Alkaline phosphatase (ALKP; U/L) 1331–1595 1381 (740–1451) 1247 (974–1632)
Alanine aminotransferase (ALAT; U/L) 12–16 18 (14–27) 15 (14–27)
Gamma glutamyl transferase (GGT; U/L)n 4–5 5 (2–8) 4.8271.68 (2–8)
Amylase (U/L) 483–569 772 (365–813) 493 (416–1136)
Albumin (g/L) 13.9–14.5 13 (11.7–14) 12.7 (11–14.4)
Total protein (g/L) 29.8–32.8 27 (25.5–31.2) 27.6 (23.3–29.4)
Uric acid (U/L) 367.5–967.3 715 (367–808) 611 (207–1200)
Urea (mmol/L) 1.64–2.89 3 (1.08–11.9) 2.2 (2.15–4.41)
Urea:creatinine 0.1–0.15 0.2 (0.14–0.35) 0.15 (0.09–0.57)
Uric acid:creatinine 33.3–33.4 41 (7.9–92) 31 (24–62)
Protein:creatinine 1.13–2.71 2 (1.13–2.71) 2.3 (1.07–2.44)
Total bilirubin (mmol/L) 8.3–17.1 16 (13.5–19.3) 16 (9.9–26)
Bile acids (mmol/L) 37.2–57.5 55 (43.7–75.8) 50 (44.4–158)
Cholesterol (mmol/L)n 4.2–4.8 4 (3.15–5.78) 6.11 (6.01–9.18)
Glucose (mmol/L) 16.9–18.1 17 (14.3–17.4) 15.8 (15.5–20.7)
Fructoseamine (mmol/L)n 173.8–184 154 (173–210) 187 (127–223)
Creatinine (mmol/L) 11–29 17 (12–25) 13 (8–26)
Inorganic phosphate (mmol/L) 2.05–2.17 2 (1.77–2.21) 2 (1.5–2.44)
Calcium (mmol/L) 2.33–2.47 2 (2.26–2.62) 2.45 (2.26–2.62)
Magnesium (mmol/L) 0.87–0.89 0.8 (0.79–0.87) 0.8 (0.73–1.16)
Sodium (mmol/L)n 155.5–155.8 153 (149–155) 152 (150–156)
Potassium (mmol/L)n 1.86–2.9 2 (2.23–4.95) 2.9 (0.75–3.03)
Note the statistically significant differences in five of the blood clinical–chemical parameters between the three raptor species.
n Statistically significant difference between species the three species (po0.05, Kruskal–Wallis).
0
1
10
100
1000
10000
Album
inG
lucoseTotal proteinA
LKP
ALAT
Tot bilirubinFructosam
ineC
holesterolC
reatinineInorganic phosphateB
ile acidA
mylase
Urea
GG
TC
alciumM
agnesiumU
ric acidS
odiumP
otassiumU
rea:Creat
Uric acid:C
reatP
rot:Creat
BCCPs
(U/L
, mm
ol/L
, mm
ol/L
)
Golden eagle (n = 2)Goshawk (n = 16)Sea eagle (n = 5)
*
**
**
Fig. 2. Log 10 concentrations (mean7SD) of 22 blood plasma clinical–chemical parameters in 3 raptor species from Northern Norway (see Table 1 for concentration units).
Note the few inconsistent significant species differences and relatively low sample size for golden eagles and white-tailed eagles. See Table 1 for abbreviations and
concentrations. n: Statistical significant differences between species (pr0.05, Kruskal–Wallis).
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–17 11
negatively correlated to Mirex and ALAT being positively corre-lated to Mirex (p=0.04, r=0.47, n=20). In both goshawks (p=0.07,r=0.49, n=14) and white-tailed eagles (p=0.04, r=0.8, n=5),cholesterol was positively correlated to Mirex.
PBDEs and TBA were detected in all three species. These werePBDEs and TBA. PBDEs were positively correlated to ALAT(p=0.02, r=0.48, n=23), total bilirubin (p=0.03, r=0.45, n=23)and uric acid (p=0.02, r=0.5, n=23) in the pooled species samples.This was also the case for cholesterol (p=0.05, r=0.49, n=16) ingoshawks and white-tailed eagles (p=0.02, r=0.82, n=5). For TBAin goshawks, a positive correlation was found to total protein(p=0.03, r=0.61, n=13) and protein:creatinine (p=0.01, r=0.67,n=13) while a negative correlation was found for creatinine
(p=0.05, r=�0.55, n=13) and a negative trend for amylase(p=0.07, r=�0.52, n=13). No significant results between TBAand BCCPs were found in white-tailed eagles. All results aresummarised in Table 3.
In the pooled samples, uric acid:creatinine was positivelycorrelated to PFCs (p=0.02, r=0.48, n=23). In goshawks, PFCs werepositively correlated to cholesterol (po0.01, r=0.71, n=16) anduric acid (po0.01, r=0.67, n=16). Similar positive trends werefound for albumin (p=0.06, r=0.48, n=16) and total bilirubin(p=0.08, r=0.44, n=16) while amylase was negatively correlatedto PFCs (p=0.08, r=�0.45, n=16). In white-tailed eagles, negativecorrelations were found comparing PFCs and bile acids (p=0.09,r=�0.71, n=5) and potassium (p=0.08, r=�0.81, n=5), while
ARTICLE IN PRESS
Table 2Summary statistics [median(min–max, n)] of organohalogen contaminants in chicks of golden eagles, white-tailed eagles and goshawks from Northern Norway.
Parameter (concentration) Golden eagles (n=2). Goshawk (n=16) White-tailed eagles (n=5).
PPCB (pg/ml ww) 3795–14801 8412 (2296–44830) 19,509 (9144–68837)
p,p0-DDE (pg/ml ww) 603–4880 7527 (1769–24718) 5236 (1087–10907)
b-HCH (pg/ml ww) 128–145 122 (74.4–143, 9) 118 (1)
HCB (pg/ml ww) 185–193 486 (150–2251) 819 (500–976)
Heptachlor epoxide (pg/ml ww) 92–120 279 (104–818) 280 (194–848)
Chlordanes (pg/ml ww)n 49–763 737 (232–2634) 2247 (904–5062)
Mirex (pg/ml ww) 94 (1) 99 (48–362, 14) 202 (46–622)
TBA (pg/ml ww)n 46–94 59 (24–371, 13) 874 (788–4719)P
PBDE (pg/ml ww) 252–302 731 (161–4622) 2000 (386–4573)P
PFC (pg/ml ww)n 900–1300 21,300 (8800–99000) 55,000 (32500–70400)
% Lipids (g/ml) 0.48–0.66 0.63 (0.44–0.83) 0.76 (0.66–1.06)
Note that there are statistically significant differences in blood plasma concentrations of chlordanes, TBA and PFCs between the three raptor species. Reasons for reduced
number of observations are ascribed to detection limits.
n Statistically significant differences between the three species (po0.05, Kruskal–Wallis).
1
10
100
1000
10000
100000
PCBs
p,p-D
DE
β-HCH
HCB
Heptac
hlor e
poxid
e
Chlorda
nes
Mirex
TBA
PBDEsPFCs
OHC group
LogC
once
ntra
tions
(pg/
ml w
w)
Golden eagle (n = 2)Goshawk (n = 16)Sea eagle (n = 5)
*
*
*
Fig. 3. Log 10 concentrations (mean7SD) of 10 groups of organohalogen contaminants divided on 3 raptor species from Northern Norway. Note the few inconsistent
significant species differences and the relatively low sample size for golden eagles and white-tailed eagles. See Table 2 for abbreviations and concentrations. n: Statistical
significant differences between species (pr0.05, Kruskal–Wallis).
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–1712
positive correlations were found to glucose (p=0.08, r=0.8, n=5)and protein:creatinine (p=0.09, r=0.78, n=5). Based on thecorrelation coefficients, some of these results are likely to occurdue to the low sample size.
3.4. PCAs
The PCA analysis of BCCPs and species resulted in sevencomponents with eigen values above 1 and should therefore beconsidered (Kaiser’s rule (Joliffe, 2002)). However, for simplicityreasons we only deal with the two first components, whichexplained 39% of the total variation in the data. Fig. 4 shows abiplot of the two first components where the arrows represent theloadings of each of the BCCPs and the points (goshawks, goldeneagles and white-tailed eagles) the components value of eachindividual. The BCCPs points in all directions, however, somegrouping of parameters can be seen. Gamma glutamyl transferaseand glucose points in the direction of the first principalcomponent (PCA1), while total protein, albumin and calciumforms a group pointing in the direction of the second principalcomponent (PCA2). Cholesterol and fructose are closely togetherand points in the direction of negatively PCA1 and amylase in thedirection of negatively PCA2. Total bilirubin, ALAT, magnesium
and sodium form a group pointing appx. 451 to the PCA1 and PCA2axis while urea, creatinine and potassium group in the opposite(180 degrees) direction. An ANOVA followed by Tukey’s multiplecomparisons of means showed that white-tailed eagle hadsignificantly (po0.05) lower PCA1 scores than goshawk, whileno other pairwise comparisons were found significant. Nosignificant differences were found between species of the meanPCA2 (ANOVA, p=0.094, respectively).
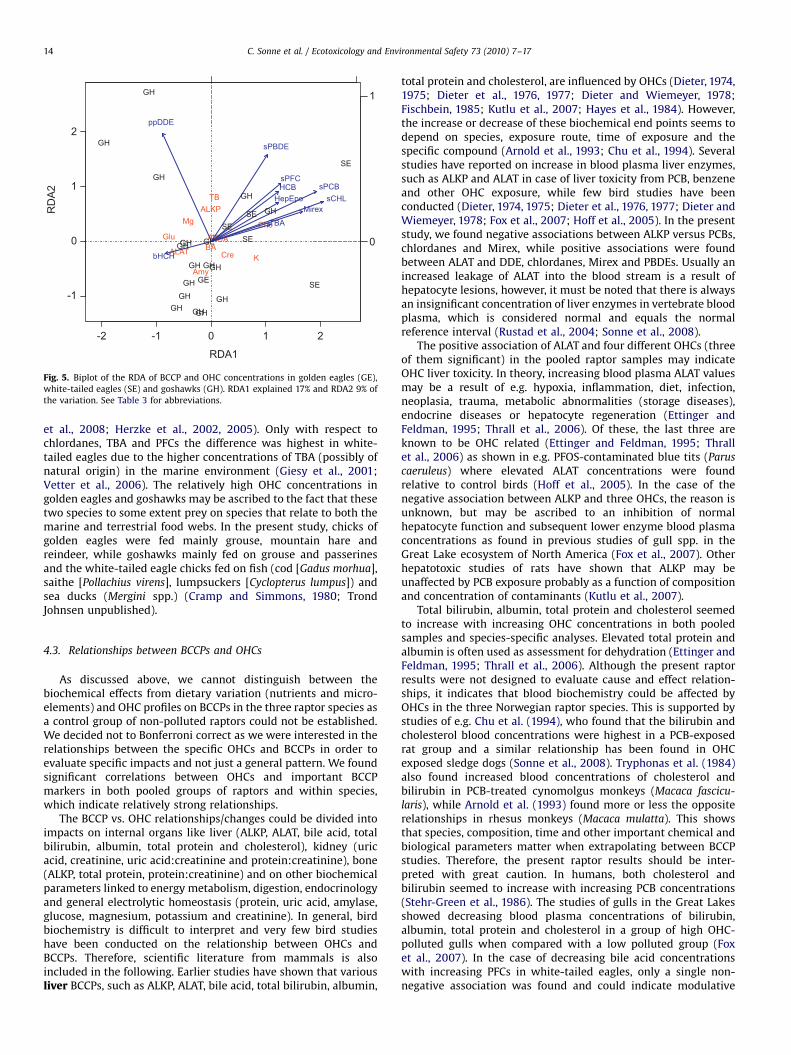
3.5. RDAs
The RDA analysis of BCCPs, OHCs and species divided the totalvariance into two parts; 54% constrained (by the multivariateregression) and 46% unexplained. The first two canonical axisexplained 17% and 9% (26% in total) of the entire variation. Fig. 5shows a biplot of the RDA results. The arrows represent the firsttwo RDA axis scores. The BCCP scores and the individual birds arerepresented by their abbreviations. Arrows pointing in the samedirection indicate that the compounds are positively correlatedwhile arrows pointing in opposite direction indicate a negativelycorrelation. Lines with an angle of 901 indicate uncorrelation.Most of the OHCs points in the same direction. p,p0-DDE and b-HCH are exceptions where p,p0-DDE is uncorrelated with the other
ARTICLE IN PRESS
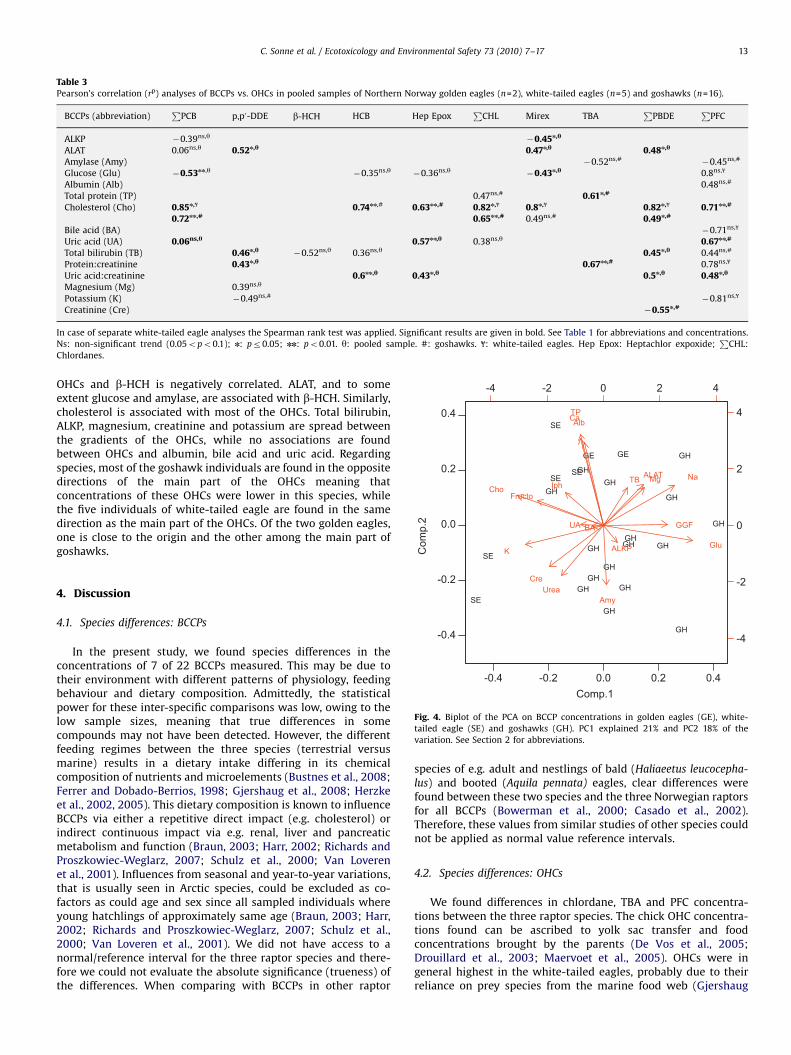
Table 3Pearson’s correlation (rp) analyses of BCCPs vs. OHCs in pooled samples of Northern Norway golden eagles (n=2), white-tailed eagles (n=5) and goshawks (n=16).
BCCPs (abbreviation)P
PCB p,p0-DDE b-HCH HCB Hep EpoxP
CHL Mirex TBAP
PBDEP
PFC
ALKP �0.39ns,y�0.45n,h
ALAT 0.06ns,y 0.52n,h 0.47n,h 0.48n,h
Amylase (Amy) �0.52ns,#�0.45ns,#
Glucose (Glu) �0.53nn,y�0.35ns,y
�0.36ns,y�0.43n,h 0.8ns,f
Albumin (Alb) 0.48ns,#
Total protein (TP) 0.47ns,# 0.61n,#
Cholesterol (Cho) 0.85n,f 0.74nn,# 0.63nn,# 0.82n,f 0.8n,f 0.82n,f 0.71nn,#
0.72nn,# 0.65nn,# 0.49ns,# 0.49n,#
Bile acid (BA) �0.71ns,f
Uric acid (UA) 0.06ns,h 0.57nn,h 0.38ns,y 0.67nn,#
Total bilirubin (TB) 0.46n,h�0.52ns,y 0.36ns,y 0.45n,h 0.44ns,#
Protein:creatinine 0.43n,h 0.67nn,# 0.78ns,f
Uric acid:creatinine 0.6nn,h 0.43n,h 0.5n,h 0.48n,h
Magnesium (Mg) 0.39ns,y
Potassium (K) �0.49ns,#�0.81ns,f
Creatinine (Cre) �0.55n,#
In case of separate white-tailed eagle analyses the Spearman rank test was applied. Significant results are given in bold. See Table 1 for abbreviations and concentrations.
Ns: non-significant trend (0.05opo0.1); n: pr0.05; nn: po0.01. y: pooled sample. #: goshawks. f: white-tailed eagles. Hep Epox: Heptachlor expoxide;P
CHL:
Chlordanes.
Com
p.2
GEGE
GH
GH
GH
GH
GHGH
GH
GH
GHGH
SE
SE
SE
SE
-4
Alb
Glu
TP
ALAT
ALKP
TB
FructoCho Iph
BA GGF
Ca
Mg
UA
Na
K
0.0
0.2
0.4
0
2
4
-2 0 2 4
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–17 13
OHCs and b-HCH is negatively correlated. ALAT, and to someextent glucose and amylase, are associated with b-HCH. Similarly,cholesterol is associated with most of the OHCs. Total bilirubin,ALKP, magnesium, creatinine and potassium are spread betweenthe gradients of the OHCs, while no associations are foundbetween OHCs and albumin, bile acid and uric acid. Regardingspecies, most of the goshawk individuals are found in the oppositedirections of the main part of the OHCs meaning thatconcentrations of these OHCs were lower in this species, whilethe five individuals of white-tailed eagle are found in the samedirection as the main part of the OHCs. Of the two golden eagles,one is close to the origin and the other among the main part ofgoshawks.
-0.4Comp.1
GH
GH
GHGH
GHGHSE
Cre
AmyUrea
-0.4
-0.2
-0.2 0.0 0.2 0.4
-4
-2
Fig. 4. Biplot of the PCA on BCCP concentrations in golden eagles (GE), white-
tailed eagle (SE) and goshawks (GH). PC1 explained 21% and PC2 18% of the
variation. See Section 2 for abbreviations.
4. Discussion
4.1. Species differences: BCCPs
In the present study, we found species differences in theconcentrations of 7 of 22 BCCPs measured. This may be due totheir environment with different patterns of physiology, feedingbehaviour and dietary composition. Admittedly, the statisticalpower for these inter-specific comparisons was low, owing to thelow sample sizes, meaning that true differences in somecompounds may not have been detected. However, the differentfeeding regimes between the three species (terrestrial versusmarine) results in a dietary intake differing in its chemicalcomposition of nutrients and microelements (Bustnes et al., 2008;Ferrer and Dobado-Berrios, 1998; Gjershaug et al., 2008; Herzkeet al., 2002, 2005). This dietary composition is known to influenceBCCPs via either a repetitive direct impact (e.g. cholesterol) orindirect continuous impact via e.g. renal, liver and pancreaticmetabolism and function (Braun, 2003; Harr, 2002; Richards andProszkowiec-Weglarz, 2007; Schulz et al., 2000; Van Loverenet al., 2001). Influences from seasonal and year-to-year variations,that is usually seen in Arctic species, could be excluded as co-factors as could age and sex since all sampled individuals whereyoung hatchlings of approximately same age (Braun, 2003; Harr,2002; Richards and Proszkowiec-Weglarz, 2007; Schulz et al.,2000; Van Loveren et al., 2001). We did not have access to anormal/reference interval for the three raptor species and there-fore we could not evaluate the absolute significance (trueness) ofthe differences. When comparing with BCCPs in other raptor
species of e.g. adult and nestlings of bald (Haliaeetus leucocepha-
lus) and booted (Aquila pennata) eagles, clear differences werefound between these two species and the three Norwegian raptorsfor all BCCPs (Bowerman et al., 2000; Casado et al., 2002).Therefore, these values from similar studies of other species couldnot be applied as normal value reference intervals.
4.2. Species differences: OHCs
We found differences in chlordane, TBA and PFC concentra-tions between the three raptor species. The chick OHC concentra-tions found can be ascribed to yolk sac transfer and foodconcentrations brought by the parents (De Vos et al., 2005;Drouillard et al., 2003; Maervoet et al., 2005). OHCs were ingeneral highest in the white-tailed eagles, probably due to theirreliance on prey species from the marine food web (Gjershaug
ARTICLE IN PRESS
-2
RDA1
RD
A2
AlbGlu TP
ALKP
ALAT
TB
Cho
CreBA
Amy
Mg
UA
K
sPCB
ppDDE
HepEpo
bHCH
HCBsCHL
Mirex
TBA
sPBDE
sPFC
GE
GE
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
GH
SE
SE
SE
SE
SE
-1
0
1
2
-1 0 1 2
1
0
Fig. 5. Biplot of the RDA of BCCP and OHC concentrations in golden eagles (GE),
white-tailed eagles (SE) and goshawks (GH). RDA1 explained 17% and RDA2 9% of
the variation. See Table 3 for abbreviations.
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–1714
et al., 2008; Herzke et al., 2002, 2005). Only with respect tochlordanes, TBA and PFCs the difference was highest in white-tailed eagles due to the higher concentrations of TBA (possibly ofnatural origin) in the marine environment (Giesy et al., 2001;Vetter et al., 2006). The relatively high OHC concentrations ingolden eagles and goshawks may be ascribed to the fact that thesetwo species to some extent prey on species that relate to both themarine and terrestrial food webs. In the present study, chicks ofgolden eagles were fed mainly grouse, mountain hare andreindeer, while goshawks mainly fed on grouse and passerinesand the white-tailed eagle chicks fed on fish (cod [Gadus morhua],saithe [Pollachius virens], lumpsuckers [Cyclopterus lumpus]) andsea ducks (Mergini spp.) (Cramp and Simmons, 1980; TrondJohnsen unpublished).
4.3. Relationships between BCCPs and OHCs
As discussed above, we cannot distinguish between thebiochemical effects from dietary variation (nutrients and micro-elements) and OHC profiles on BCCPs in the three raptor species asa control group of non-polluted raptors could not be established.We decided not to Bonferroni correct as we were interested in therelationships between the specific OHCs and BCCPs in order toevaluate specific impacts and not just a general pattern. We foundsignificant correlations between OHCs and important BCCPmarkers in both pooled groups of raptors and within species,which indicate relatively strong relationships.
The BCCP vs. OHC relationships/changes could be divided intoimpacts on internal organs like liver (ALKP, ALAT, bile acid, totalbilirubin, albumin, total protein and cholesterol), kidney (uricacid, creatinine, uric acid:creatinine and protein:creatinine), bone(ALKP, total protein, protein:creatinine) and on other biochemicalparameters linked to energy metabolism, digestion, endocrinologyand general electrolytic homeostasis (protein, uric acid, amylase,glucose, magnesium, potassium and creatinine). In general, birdbiochemistry is difficult to interpret and very few bird studieshave been conducted on the relationship between OHCs andBCCPs. Therefore, scientific literature from mammals is alsoincluded in the following. Earlier studies have shown that variousliver BCCPs, such as ALKP, ALAT, bile acid, total bilirubin, albumin,
total protein and cholesterol, are influenced by OHCs (Dieter, 1974,1975; Dieter et al., 1976, 1977; Dieter and Wiemeyer, 1978;Fischbein, 1985; Kutlu et al., 2007; Hayes et al., 1984). However,the increase or decrease of these biochemical end points seems todepend on species, exposure route, time of exposure and thespecific compound (Arnold et al., 1993; Chu et al., 1994). Severalstudies have reported on increase in blood plasma liver enzymes,such as ALKP and ALAT in case of liver toxicity from PCB, benzeneand other OHC exposure, while few bird studies have beenconducted (Dieter, 1974, 1975; Dieter et al., 1976, 1977; Dieter andWiemeyer, 1978; Fox et al., 2007; Hoff et al., 2005). In the presentstudy, we found negative associations between ALKP versus PCBs,chlordanes and Mirex, while positive associations were foundbetween ALAT and DDE, chlordanes, Mirex and PBDEs. Usually anincreased leakage of ALAT into the blood stream is a result ofhepatocyte lesions, however, it must be noted that there is alwaysan insignificant concentration of liver enzymes in vertebrate bloodplasma, which is considered normal and equals the normalreference interval (Rustad et al., 2004; Sonne et al., 2008).
The positive association of ALAT and four different OHCs (threeof them significant) in the pooled raptor samples may indicateOHC liver toxicity. In theory, increasing blood plasma ALAT valuesmay be a result of e.g. hypoxia, inflammation, diet, infection,neoplasia, trauma, metabolic abnormalities (storage diseases),endocrine diseases or hepatocyte regeneration (Ettinger andFeldman, 1995; Thrall et al., 2006). Of these, the last three areknown to be OHC related (Ettinger and Feldman, 1995; Thrallet al., 2006) as shown in e.g. PFOS-contaminated blue tits (Parus
caeruleus) where elevated ALAT concentrations were foundrelative to control birds (Hoff et al., 2005). In the case of thenegative association between ALKP and three OHCs, the reason isunknown, but may be ascribed to an inhibition of normalhepatocyte function and subsequent lower enzyme blood plasmaconcentrations as found in previous studies of gull spp. in theGreat Lake ecosystem of North America (Fox et al., 2007). Otherhepatotoxic studies of rats have shown that ALKP may beunaffected by PCB exposure probably as a function of compositionand concentration of contaminants (Kutlu et al., 2007).
Total bilirubin, albumin, total protein and cholesterol seemedto increase with increasing OHC concentrations in both pooledsamples and species-specific analyses. Elevated total protein andalbumin is often used as assessment for dehydration (Ettinger andFeldman, 1995; Thrall et al., 2006). Although the present raptorresults were not designed to evaluate cause and effect relation-ships, it indicates that blood biochemistry could be affected byOHCs in the three Norwegian raptor species. This is supported bystudies of e.g. Chu et al. (1994), who found that the bilirubin andcholesterol blood concentrations were highest in a PCB-exposedrat group and a similar relationship has been found in OHCexposed sledge dogs (Sonne et al., 2008). Tryphonas et al. (1984)also found increased blood concentrations of cholesterol andbilirubin in PCB-treated cynomolgus monkeys (Macaca fascicu-
laris), while Arnold et al. (1993) found more or less the oppositerelationships in rhesus monkeys (Macaca mulatta). This showsthat species, composition, time and other important chemical andbiological parameters matter when extrapolating between BCCPstudies. Therefore, the present raptor results should be inter-preted with great caution. In humans, both cholesterol andbilirubin seemed to increase with increasing PCB concentrations(Stehr-Green et al., 1986). The studies of gulls in the Great Lakesshowed decreasing blood plasma concentrations of bilirubin,albumin, total protein and cholesterol in a group of high OHC-polluted gulls when compared with a low polluted group (Foxet al., 2007). In the case of decreasing bile acid concentrationswith increasing PFCs in white-tailed eagles, only a single non-negative association was found and could indicate modulative

ARTICLE IN PRESS
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–17 15
effects on hepatocyte bile synthesis/secretion and hepatocyte andbile duct lesions although that is highly speculative (Fox et al.,2007).
BCCP renal parameters also showed significant relationships toOHC concentrations. These were total protein (increase), uric acid(increase), creatinine (decrease), uric acid:creatinine (increase)and protein:creatinine (increase). Furthermore, the statisticalrelationships indicated elevated levels of protein and uric acidcompared with creatinine with increasing OHC concentrations.The increased blood protein and uric acid concentrations mayindicate metabolic shifts, high protein diet – which is not unlikelyfor the raptor species – or dehydration as discussed above(Ettinger and Feldman, 1995; Thrall et al., 2006). Furthermore,the elevated levels of protein and uric acid might be related tochick muscle metabolism and growth (Waikar and Bonventre,2008). Chu et al. (1994) found a similar urea increase in theirstudy of PCB-exposed female rats. In addition, previous OHCstudies of Arctic species have shown renal histopathologicalchanges that would allow such interpretations (Sonne et al., 2006,2007, 2008). However, histopathological end points were un-fortunately not possible to investigate in the present Norwegianraptor chicks.
Similarly for bones, decreasing ALKP concentrations withincreasing OHC concentrations could indicate changes in boneturn-over and metabolism (Andrews, 1989). Such impacts aresupported by OHC-driven eggshell thinning theories and observa-tions (Lundholm, 1997; Elliott and Martin, 1994; Falk et al., 2006;Helander et al., 2002; Hernandez et al., 2008; Scharenberg andLooft, 2004) and may even be a result of trans-generationaltoxicity (Steinberg et al., 2008). Unfortunately, bone end pointmarkers were also not possible to investigate in the present raptorchicks.
Changes in the more overall and broader biochemical endpoint OHC markers were found (amylase, glucose, magnesium andpotassium). The decreasing blood glucose levels as a function ofvarious OHCs could indicate endocrine changes as serumfructosamine was not affected and as birds in general are knownto maintain high blood glucose concentrations (Braun andSweazea, 2008). The endocrine pathways for this could be dueto an inverse relationship between maximum corticosterone-driven stress response and PCB concentrations as seen in fishspecies (Jørgensen et al., 2006; Lacroix and Hontela, 2003). Formagnesium and potassium the relationships may be dietary and/or OHC explained while a linkage to renal disorders cannot beexcluded (Musso, in press).
4.4. Interpretations
The present study may suggest a relationship between OHCsand specific BCCPs in the three raptor species. However, thepresent results should be interpreted with great caution as BCCPsare influenced by multiple factors such as age, diseases, diet,hydration and other environmental factors as well as species-specific differences driven by physiology, diet and OHC loadsespecially in case of goshawks and white-tailed eagles. Therefore,the present pilot study points towards a larger study whereanalyses can be conducted within each species and on a largerdata set likely including histopathology and bone biomarkers ifpossible. It is not easy to conclude on the health impacts from theOHC relationships to liver, kidney and general homeostasis in thethree raptor species. It is either not possible to assess whetherthere are indications of a decline in the raptor populations ofNorthern Norway although climate-driven year-to-year variationsare seen (Cramp and Simmons, 1980). Especially the high OHCcontaminated goshawk and white-tailed eagle populations may
suffer from increased risks of infections and lower reproductionrates resulting in impaired fitness, survival and adaptations toecological changes like global warming.
5. Conclusions
Three OHC groups and five BCCPs showed species-specificdifferences. Furthermore, the relationships between OHCs andvarious BCCPs suggested an impact from dietary composition and/or persistent organic contaminants on liver/kidney function,endocrinology and metabolism in raptor chicks of NorthernNorway in a direction that may indicate health impacts.Ultimately, the chemical stress on blood plasma biochemistrycould have consequences on goshawk and white-tailed eaglepopulations in particular as they carry the highest body burdens.The pathways for this are due to increased infection risks,decreased reproductive performance and thereby impaired fit-ness, survival and adaptations to ecological changes such as globalwarming. We therefore recommend that a larger study shouldtake place in the near future in order to monitor potentialpopulation impacts.
Acknowledgments
The Norwegian Research Council and the Funds of ScientificResearch Flanders (personally to Dr. VLB Jaspers and Dr. A Covaci);are acknowledged for funding. Laboratory technicians at theDepartment of Small Animal Clinical Sciences, University ofCopenhagen and at the Norwegian Institute for Air Research areacknowledged for conducting the BCCP and OHC analyses,respectively.
References
AMAP, 1998. Arctic pollution 1998; Arctic Monitoring and Assessment Programme,Oslo, Norway.
AMAP 2004. Persistent Organic Pollutants in the Arctic. Arctic Pollution 2002,Arctic Monitoring and Assessment Programme, Oslo, Norway.
Andrews, J.E., 1989. Polychlorinated biphenyl (Aroclor 1254) induced changesin femur morphology calcium metabolism and nephrotoxicity. Toxicology 57,83–96.
Arnold, D.L., Bryce, F., Karpinski, K., Mes, J., Fernie, S., Tryphonas, H., Truelove, J.,McGuire, P.F., Burns, D., Tanner, J.R., Stapley, R., Zawidzka, Z.Z., Basford, D., 1993.Toxicological consequences of Aroclor 1254 ingestion by female rhesus(Macaca mulatta) monkeys. Part 1B. Prebreeding phase: clinical and analyticallaboratory findings. Food Chem. Toxicol. 11, 811–824.
Bandiera, S.M., Torok, S.M., Letcher, R.J., Nostrom, R.J., 1997. Immunoquantitation ofcytochromes P450 1A and P450 2B and comparison with chlorinatedhydrocarbon levels in archived polar bear liver samples. Chemosphere 34,1469–1479.
Bowerman, W.W., Stickle, J.E., Sikarskie, J.G., Giesy, J.P., 2000. Hematology andserum chemistries of nestling bald eagles (Haliaeetus leucocephalus) in thelower peninsula of MI, USA. Chemosphere 41, 1575–1579.
Braun, E.J., 2003. Regulation of renal and lower gastrointestinal function: role influid and electrolyte balance. Comp. Biochem. Physiol. A Mol. Integr. Physiol.136, 499–505.
Braun, Sweazea, K.L., 2008. Glucose regulation in birds. Comp. Biochem. Physiol. BBiochem. Mol. Biol. 151, 1–9.
Bustnes, J.O., Tveraa, T., Fauchald, P., Helberg, M., Skaare, J.U., 2008. The potentialimpact of environmental variation on the concentrations and ecological effectsof pollutants in a marine avian top predator. Environ. Int. 34, 193–201.
Casado, E., Balbontin, J., Ferrer, M., 2002. Plasma chemistry in booted eagle(Hieraaetus pennatus) during breeding season. Comp. Biochem. Physiol. A 131,233–241.
Chrousos, G.P., Gold, P.W., 1992. The concepts of stress and stress system disorders:overview of physical and behavioral homeostasis. JAMA 267, 1244–1252.
Chu, I., Villeneuve, D.C., Yagminas, A., Lecavalier, P., Poon, R., Feeley, M., Kennedy,S.W., Seegal, R.F., Hakansson, H., Ahlborg, U.G., Valli, V.E., 1994.Subchronic toxicity of 3,30 ,4,40 ,5-pentachlorobiphenyl in the rat. 1. Clinical,biochemical, hamatological, and histopathological changes. Fund. Appl.Toxicol. 22, 457–468.
Colborn, T., 1991. Epidemiology of Great Lakes bald eagles. J. Toxicol. Environ.Health 33, 395–453.

ARTICLE IN PRESS
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–1716
Colborn, T., 2004. Neurodevelopment and endocrine disruption. Environ. HealthPerspect. 112, 944–949.
Cramp, S., Simmons, K.E.L., 1980. In: Cramp, S., Simmonds, K.E.L. (Eds.), The Birds ofthe Western Palearctic, vol. III. Oxford University Press, UK 695pp.
Damstra, T., Barlow, S., Bergman, A., Kavlock, R., Kraak, G.V.D., 2002. In: GlobalAssessment of the State-of-the-Science of Endocrine Disruptors. WHO, Geneva,Switzerland.
Darnerud, P.O., Eriksen, G.S., Johannesson, T., Laresen, P.B., Viluksela, M., 2001.Polybrominated diphenyl ethers: occurrence, dietary exposure, and toxicology.Environ. Health Perspect. 109, 49–68.
De Vos, S., Verschueren, D., De Schrijver, R., 2005. Digestibility, retention andincorporation of low-level dietary PCB contents in laying hens. Chemosphere58, 1553–1562.
Dieter, M.P., 1974. Plasma enzyme activities in Coturnix quail fed graded doses ofDDE, polychlorinated biphenyl, malathion and mercuric chloride. Toxicol. Appl.Pharmacol. 27, 86–98.
Dieter, M.P., 1975. Further studies of the use of enzyme profiles to monitor residueaccumulation in wildlife: plasma enzymes in starlings fed graded concentra-tions of morsodren, DDE, Aroclor 1254, and malathion. Arch. Environ. Contam.Toxicol. 3, 142–150.
Dieter, M.P., Perry, M.C., Mulhern, B.M., 1976. Lead and PCB’s in canvasback ducks:relationship between enzyme levels and residues in blood. Arch. Environ.Contam. Toxicol. 5, 1–13.
Dieter, M.P., Perry, M.C., Mulhern, B.M., 1977. Lead and PCBs in canvasback ducks:relationship between enzyme levels and residues in blood. Arch. Environ.Contam. Toxicol. 5 (1), 1–13.
Dieter, M.P., Wiemeyer, S.N., 1978. Six different plasma enzymes in bald eagles(Haliaeetus leucocephalis) and their usefulness in pathological diagnosis. Comp.Biochem. Physiol. 61C, 153–155.
Donaldson, G.M., Shutt, J.L., Hunter, P., 1999. Organochlorine contamination in baldeagle eggs and nestlings from the Canadian Great Lakes. Arch. Environ.Contam. Toxicol. 36, 70–80.
Drouillard, K.G., Norstrom, R.J., Fox, G.A., Gilman, A., Peakall, D.B., 2003.Development and validation of a herring gull embryo toxicokinetic model forPCBs. Ecotoxicology 12, 55–68.
Elliott, J.E., Martin, P.A., 1994. Chlorinated hydrocarbons and shell thinning in eggsof (Accipiter) hawks in Ontario, 1986–1989. Environ. Pollut. 86, 189–200.
Ettinger, S.J., Feldman, E.C., 1995. In: Ettinger, S.J., Feldman, E.C. (Eds.), Textbook ofVeterinary Internal Medicine. W.B. Saunders, Philadelphia, USA.
Erikstad, K.E., Bustnes, J.O., Lorentsen, S.H., Reiertsen, T.K., 2009. Sex ratio in lesserblack-backed gull in relation to environmental pollutants. Behav. Ecol.Sociobiol. 57, 357–365.
Falk, K., Møller, S., Mattox, W.G., 2006. A long-term increase in eggshell thicknessof Greenlandic Peregrine Falcons Falco peregrinus tundrius. Sci. Total Environ.355, 127–134.
Ferrer, Miguel, Dobado-Berrios, Pablo, 1998. Factors affecting plasma chemistryvalues of the Spanish Imperial Eagle, Aquila adalberti. Comp. Biochem. Phys. A120, 209–217.
Fischbein, A., 1985. Liver function tests in workers with occupational exposure topolychlorinated biphenyls (PCBs): comparison with Yusho and Yu-cheng.Environ. Health Perspect. 60, 145–150.
Fox, G.A., Jeffrey, D.A., Williams, K.S., Kennedy, S.W., Grasman, K.A., 2007. Health ofherring gulls (Larus argentatus) in relation to breeding location in the Early1990s. I. Biochem. Meas. J Toxicol. Environ. Health A 70, 1443–1470.
Giesy, J.P., Kannan, K., Jones, P.D., 2001. Global Biomonitoring of PerfluorinatedOrganics. Sci. World 1, 627–629.
Gjershaug, J.O., Kalas, J.A., Nygard, T., Herzke, D., Folkestad, A.O., 2008. Monitoringof raptors and their contamination levels in Norway. Ambio 37, 420–424.
Grandjean, P., Landrigan, P.J., 2006. Review: developmental neurotoxicity ofindustrial chemicals. Lancet 16, 2167–2178.
Grasman, K.A., Scanlon, P.F., Fox, G.A., 2000. Geographic variation in hematologicalvariables in adult and prefledgling herring gulls (Larus argentatus) and possibleassociations with organochlorine exposure. Arch. Environ. Contam. Toxicol. 38,244–253.
Grønnesby, S., Nygard, T., 2000. Using time-lapse video monitoring to study foodselection by breeding Goshawks Accipiter gentilis in Central Norway. OrnisFenn. 77, 117–129.
Guvenius, D.M., Hassanzadeh, P., Bergman, A, Nor�en, K., 2002. Metabolites ofpolychlorinated biphenyls in human liver and adipose tissue. Environ. Toxicol.Chem. 21, 362–372.
Gotsch, A., Mariussen, E., von der Recke, R., Herzke, D., Berger, U., Vetter, W., 2004.Analytical strategies for successful enantioselective separation of atropiso-meric polybrominated biphenyls 132 and 149 in environmental samples.J. Chromatogr. A 1063, 193–199.
Halley, D.J., Nygard, T., Minagawa, M., Systad, G.H., Jacobsen, K.O., Johnsen, T.V.,2007. Rein som næring hos kongørn I hekketida I et omrade I Finnmarkundersøkt ved hjelp av stabilisotopteknikk. Prosjektrapport 2004–2006.
Hamers, T., Kamstra, J.H., Sonneveld, E., Murk, A.J., Kester, M.H.A., Andersson, P.L.,Legler, J., Brouwer, A., 2006. In vitro profiling of the endocrine-disruptingpotency of brominated flame retardants. Toxicol. Sci. 92, 157–173.
Harju, M., Hamers, T., Kamstra, J.H., Sonneveld, E., Boon, J.P., Tysklind, M.,Andersson, P.L., 2007. Quantitative structure–activity relationship modelingon in vitro endocrine effects and metabolic stability involving 26 selectedbrominated flame retardants. Environ. Toxicol. Chem. 26, 816–826.
Harr, K.E., 2002. Clinical chemistry of companion avian species: a review. Vet. Clin.Pathol. 31, 140–151.
Hayes, M.A., Roberts, E., Roomi, M.W., Safe, S.H., Farber, E., Cameron, R.G., 1984.Comparative influences of different PB-type and 3-MC-type polychlorinatedbiphenyl-induced phenotypes on cytocidal hepatotoxicity of bromobenzeneand acetaminophen. Toxicol. Appl. Pharmacol. 76 (1), 118–127.
Helberg, M., Bustnes, J.O., Erikstad, K.E., Kristiansen, K.O., Skaare, J.U., 2005.Relationships between reproductive performance and organochlorinepollutants in great-black backed gulls (Larus marinus). Environ. Pollut. 134,475–483.
Helander, B., Olsson, A., Bignert, A., Asplund, L., Litz�en, K., 2002. The role of DDE,PCB, coplanar PCB and eggshell parameters for reproduction in the white-tailed sea eagle (Haliaeetus albicilla) in Sweden. Ambio 31, 386–403.
Henriksen, E.O., Gabrielsen, G.W., Trudeau, S., Wolkers, J., Sagerup, K., Skaare, J.U.,2000. Organochlorines and possible biochemical effects in glaucous gulls(Larus hyperboreus) from Bjørnøya, the Barents Sea. Arch. Environ. Contam.Toxicol. 38, 234–243.
Hernandez, M., Gonz�alez, L.M., Oria, J., S�anchez, R., Arroyo, B., 2008. Influence ofcontamination by organochlorine pesticides and polychlorinated biphenyls onthe breeding of the Spanish imperial eagle (Aquila adalberti). Environ. Toxicol.Chem. 27, 433–441.
Herzke, D., Kallenborn, R., Nygard, T., 2002. Organochlorines in egg samples fromNorwegian birds of prey: congener-, isomer- and enantiomer specificconsiderations. Sci. Total Environ. 291, 59–71.
Herzke, D., Berger, U., Kallenborn, R., Nygard, T., Vetter, W., 2005. Brominated flameretardants and other organobromines in Norwegian predatory bird eggs.Chemosphere 61, 441–449.
Hoff, P.T., Van de Vijver, K., Dauwe, T., Covaci, A., Maervoet, J., Eens, M., Blust, R., DeCoen, W., 2005. Evaluation of biochemical effects related to perfluorooctanesulfonic acid exposure in organohalogen-contaminated great tit (Parus major)and blue tit (Parus caeruleus) nestlings. Chemosphere 61, 1558–1569.
Johnsen, T.V., Systad, G.H., Jacobsen, K.O., Nygard, T., Bustnes, J.O., 2007. Theoccurrence of reindeer calves in the diet of nesting golden eagles in Finnmark,northern Norway. Ornis Fenn. 84, 112–118.
Johnson, E.O., Kamilaris, T.C., Chrousos, G.P., Gold, P.W., 1992. Mechanisms ofstress: a dynamic overview of hormonal and behavioral homeostasis. Neurosci.Biobehav. R 16, 115–130.
Joliffe, I.T., 2002. In: Principal Component Analysis second ed. SpringerISBN:0387954422, 9780387954424, 487pp.
Julshamn, K., Lundebye, A.K., Heggstad, K., Berntssen, M.H., Boe, B., 2004.Norwegian monitoring programme on the inorganic and organic contaminantsin fish caught in the Barents Sea, Norwegian Sea and North Sea, 1994–2001.Food Addit. Contam. 21, 365–376.
Jørgensen, E.H., Vijayan, M.M., Killie, J.-E.A., Aluru, N., Aas-Hansen, Ø, Maule, A.,2006. Toxicokinetics and effects of PCBs in Arctic fish: a review of studies onArctic charr. J. Toxicol. Environ. Health A 69, 37–52.
Kennedy, S.W., Fox, G.A., Jones, S.P., Trudeau, S.F., 2003. Hepatic EROD activity isnot a useful biomarker of polychlorinated biphenyl exposure in the adultherring gull (Larus argentatus). Ecotoxicology 12, 153–161.
Kutlu, S., Colakoglu, N., Halifeoglu, I., Sandal, S., Seyran, A.D., Aydin, M., Yilmaz, B.,2007. Comparative evaluation of hepatotoxic and nephrotoxic effects ofaroclors 1221 and 1254 in female rats. Cell Biochem. Funct. 25, 167–172.
Lacroix, M., Hontela, A., 2003. The organochlorine o,p0-DDD disrupts the adrenalsteroidogenic signaling pathway in rainbow trout (Oncorhynchus mykiss).Toxicol. Appl. Pharmacol. 190, 197–205.
Letcher, R.J., Norstrom, R.J., Lin, S., Ramsay, M.A., Bandiera, S.M., 1996. Immuno-quantitation and microsomal monooxygenase activities of hepatic cyto-chromes P4501A and P4502B and chlorinated hydrocarbon contaminantlevels in polar bear (Ursus maritimus). Toxicol. Appl. Pharmacol. 137, 127–140.
Letcher, R.J., Norstrom, R.J., Muir, D.C.G., Sandau, C.D., Koczanski, K., Michaud, R.,De Guise, S., B�eland, P., 2000. Methylsulfone polychlorinated biphenyl and2,2-bis(chlorophenyl)-1,1-dichloroethylene metabolites in beluga whale(Delphinapterus leucas) from the St. Lawrence River estuary and westernHudson Bay, Canada. Environ. Toxicol. Chem. 19, 1378–1388.
Lundholm, C.D., 1997. DDE-induced eggshell thinning in birds: effects of p,p0-DDEon the calcium and prostaglandin metabolism of the eggshell gland. Comp.Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 118, 113–128.
Macdonald R.W., Harner T., Fyfe J., Loeng H.,Weingartner T. 2003. AMAPAssessment 2002. The Influence of Global Change on Contaminant Pathwaysto, within and from the Arctic. Arctic Monitoring and Assessment Programme,Oslo, Norway, xii+65pp.
Macdonald, R.W., Harner, T., Fyfe, J., 2005. Recent climate change in the Arctic andits impact on contaminant pathways and interpretation of temporal trenddata. Sci. Total Environ. 342, 5–86.
Maervoet, J., Beck, V., Roelens, S.A., Covaci, A., Voorspoels, S., Geuns, J.M., Darras,V.M., Schepens, P., 2005. Uptake and tissue-specific distribution of selectedpolychlorinated biphenyls in developing chicken embryos. Environ. Toxicol.Chem. 24, 597–602.
Muir, D.C., Wagemann, R., Hargrave, B.T., Thomas, D.J., Peakall, D.B., Norstrom, R.J.,1992. Arctic marine ecosystem contamination. Sci. Total Environ. 122, 75–134.
Musso C.G.In press. Magnesium metabolism in health and disease. Int. Urol.Nephrol.
Norstrom, R.J., Muir, D.C., 1994. Chlorinated hydrocarbon contaminants in arcticmarine mammals. Sci. Total Environ. 154, 107–128.
Perneger, T.V., 1998. What’s wrong with Bonferroni adjustments. BMJ 316,1236–1238.
Richards, M.P., Proszkowiec-Weglarz, M., 2007. Mechanisms regulating feed intake,energy expenditure, and body weight in poultry. Poult. Sci. 86, 1478–1490.

ARTICLE IN PRESS
C. Sonne et al. / Ecotoxicology and Environmental Safety 73 (2010) 7–17 17
Rocca, C.L., Mantovani, A., 2006. From environment to food: the case of PCB. Ann.Ist Super Sanit�a 42, 410–416.
Rustad, P., Felding, P., Lahti, A., Nordic Reference Interval Project 2000, 2004.Proposal for guidelines to establish common biological reference intervals inlarge geographical areas for biochemical quantities measured frequently inserum and plasma. Clin. Chem. Lab. Med. 42, 783–791.
Scharenberg, W., Looft, V., 2004. Reduction of organochlorine residues in Goshawkeggs (Accipiter gentilis) from Northern Germany (1971–2002) and increasingeggshell index. Ambio 33, 495–498.
Schulz, J.H., Bermudez, A.J., Tomlinson, J.L., Firman, J.D., He, Z., 2000. Blood plasmachemistries from wild mourning doves held in captivity. J. Wildlife Dis. 36,541–545.
Sharit, J., Salvendy, G., 1982. Occupational stress: review and reappraisal. Hum.Factor 24, 129–162.
Siegel S. 1988. Nonparametric Statistics for the Behavioral Sciences. McGraw-HillInternational Book Company.
Sonne, C., Dietz, R., Leifsson, P.S., Born, E.W., Kirkegaard, M., Letcher, R.J., Muir,D.C.G., Riget, F.F., Hyldstrup, L., 2006. Are organohalogen contaminants a co-factor in the development of renal lesions in East Greenland polar bears (Ursusmaritimus)?. Environ. Toxicol. Chem. 25 (69), 1551–1557.
Sonne, C., Leifsson, P.S., Dietz, R., Kirkegaard, M., Møller, P., Jensen, A.L., Letcher, R.J.,Shahmiri, S., 2007. Renal lesions in Greenland sledge dogs (Canis familiaris)exposed to a natural dietary cocktail of persistent organic pollutants. Toxicol.Environ. Chem. 89 (3), 563–576.
Sonne, C., Dietz, R., Kirkegaard, M., Letcher, R.J., Shahmiri, S., Andersen, S., Møller, P.,Olsen, A.K., Jensen, A.L., 2008. Effects of organohalogen pollutants onhaematological and urine clinical–chemical parameters in Greenland sledgedogs (Canis familiaris). Ecotoxicol. Environ. Saf. 69, 381–390.
Stehr-Green, P.A., Welty, E., Steele, G., Steinberg, K., 1986. Evaluation of potentialhealth effects associated with serum polychlorinated biphenyl levels. Environ.Health Perspect. 70, 255–259.
Steinberg, R.M., Walker, D.M., Juenger, T.E., Woller, M.J., Gore, A.C., 2008. Effects ofperinatal polychlorinated biphenyls on adult female rat reproduction:development, reproductive physiology, and second generational effects. Biol.Reprod. 78, 1091–1101.
Ter Braak, C.J.F., Prentice, I.C., 1988. A theory of gradient analysis. Adv. Ecol. Res. 18,271–317.
Thrall, M.A., Baker, D.C., Campbell, T.W., DeNicola, D.B., Fettman, M.J., Lassen, E.D.,Rebar, A., Weiser, A., 2006. In: Veterinary Hematology and Clinical Chemistry:Text and Clinical Case Presentations Set. Blackwell Publishing, Iowa, USA.
Tryphonas, L., Truelove, J., Zawidzka, Z., Wong, J., Mes, J., Charbonneau, S., Grant,D.L., Campbell, J.S., 1984. Polychlorinated biphenyl (PCB) toxicity inadult cynomolgus monkeys (M. fascicularis): a pilot study. Toxicol. Pathol. 12,10–25.
Van Loveren, H., van Amsterdam, J.G., Vandebriel, R.J., Kimman, T.G., Rumke, H.C.,Steerenberg, P.S., Vos, J.G., 2001. Vaccine-induced antibody responses asparameters of the influence of endogenous and environmental factors. Environ.Health Perspect. 109, 757–764.
Verreault, J., Skaare, J.U., Jenssen, B.M.D., Gabrielsen, G.W., 2004. Effects oforganochlorine contaminants on thyroid hormone levels in arctic breedingglaucous gulls, Larus hyperboreus. Environ. Health Perspect. 112, 532–537.
Verreault, J., Letcher, R.J., Ropstad, E., Dahl, El, Gabrielsen, G.W., 2006. Organohalo-gen contaminants and reproductive hormones in incubating glaucous gulls(Larus hypervoreus) from the Norwegian Arctic. Environ. Toxicol. Chem. 25,2990–2996.
Verreault, J., Bech, C., Letcher, R.J., Ropstad, E., Dahl, E., Gabrielsen, G.W., 2007.Organohalogen contamination in breeding glaucous gulls from the NorwegianArctic: associations with basal metabolism and circulating thyroid hormones.Environ. Pollut. 45, 138–145.
Vetter, W., Recke, R.V.D., Herzke, D., Nygard, T., 2006. Natural and man-madeorganobromine compounds in marine biota from Central Norway. Environ. Int.33, 17–26.
Waikar, S.S., Bonventre, J.V., 2008. Biomarkers for the diagnosis of acute kidneyinjury. Nephron. Clin. Pract. 109, c192.
Willgohs, J.F., 1961. The white-tailed eagle Haliaeetus albicilla in Norway. Univ.Bergen Arbok Mat. Nart. Ser. 12, 1–212.
Zar, J.H., 1984. Biostatistical analysis second ed. Prentice-Hall Inc., New Jersey, USA.Østby, L., Gabrielsen, G.W., Krøkje, A., 2005. Cytochrome P4501A induction and
DNA adduct formation in glaucous gulls (Larus hyperboreus), fed withenvironmentally contaminated gull eggs. Ecotoxicol. Environ. Saf. 62, 363–375.
Related Documents











