AMER. ZOOL., 21:63-81 (1981) Relationships Between Invertebrate Phyla Based on Functional-Mechanical Analysis of the Hydrostatic Skeleton 1 WOLFGANG FRIEDRICH GUTMANN Forschungs-Institut Senckenberg, 6000 Frankfurt am Main, W. Germany SYNOPSIS. The phylogeny of the major groups of deuterostome coelomates—the chor- dates, hemichordates and echinoderms—is discussed based on a mechanical-functional analysis of the hydrostatic skeleton and associated structures. The basic approach is to first establish transformation series of individual features and of functional complexes of features and second to determine their "Lesrichtung" by showing the direction of in- creased economy (i.e., better adaptation) with respect to environmental factors. It is ar- gued that a metameric coelom is primitive with respect to an oligomeric one and that the ancestral form of the deuterostome coelomates is a metameric, coelomate worm-like an- imal with a complex set of circular, transverse and longitudinal body muscles. The coelom plus the complex body musculature formed the hydrostatic skeleton. The sequence of structural modifications leading to chordates is: (a) appearance of the notochord; (b) specialization of the dorsal longitudinal muscles with a reduction and disappearance of the transverse and circular muscles; (c) simultaneous appearance of the dorsal hollow nerve cord; (d) development of a postanal tail; and (e) appearance and specialization of the branchial basket with gill slits as a filter feeding apparatus. The primitive chordate would be most similar to the lancelet (Acrania). Tunicates are advanced chordates spe- cialized for sessile life and lost most chordate features in the adult, but retained them in the larvae as adaptations for active dispersal. Enteropneusts (acorn worms) are another advanced group specialized for burrowing in fine sediments and that evolved the anterior proboscis as a peristaltic burrowing organ. The notochord was lost as was the dorsal nerve cord and segmented condition of the coelom. A collar originated as a means to prevent discharged water from re-entering the mouth. Pterobranchs arose from enteropneust- like forms; their major structural changes are reduction of the branchial basket and mod- ification of the collar into tentacles which are associated with life in a closed tube. Finally, echinoderms arose from a pterobranch-like ancestor by specializing for sessile life and feeding with tentacles and by final loss of the branchial basket. Groups such as the tuni- cates, hemichordates and echinoderms could be eliminated as ancestral forms within the deuterostome coelomates because the evolution of acraniates and vertebrates from each of these groups would involve the appearance of gill slits before the notochord and/or the evolution of a metameric coelom from an oligomeric one, both of which are exceed- ingly improbable. Central to the methods used to establish the transformation series of features and their direction of evolutionary change (Lesrichtung) are functional (me- chanical) analysis and adaptive interpretation of features; hence, functional-adaptive anal- yses are an integral and essential part of the methodology of phylogenetic investigation. INTRODUCTION , , , , , , , . . . . . _ , the hydrostatic skeleton and on the evo- The phylogenetic relationship of the ludon o f t h e coelom including the ques . phyla and subphyla of animals has been d o n o f t h e pri mitiveness of the metameric the focus of active research ever since the arrangerri ent of coelomic cavities, I under- pubhcationofthe Origin of Species, but took a research project with the coopera- with almost no consensus of opinion de- d o n o f several coll eagues at the Sencken- yeloping. Moreover, no methods existed . Museum (K. Bonik, T. L. Franzen, M. for choosing in any convincing way be- Gra & sshoff) D . Mollenhauer, D. S. Peters; tween the various conflicting phylogenetic the resuks are presented in Schafer, 1973, hypotheses for the invertebrate phyla. Us- 1975 ig76 , g 7 8 ) w k h t h e major , s o f ing ideas on the functional properties of establishing major me tazoan groups and elucidating their phylogenetic relation- ships. The basic assumption underlying 'From the Symposium on Functional-AdapUveAnal- ^ . { h • evo l utionary ysts in Systematics presented at the Annual Meeting or r J • •• • / the American Society of Zoologists, 27-30 December changes are gradualists and adaptive as 1979, at Tampa, Florida. advocated by Simpson (1953) and Bock 63 Downloaded from https://academic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AMER. ZOOL., 21:63-81 (1981)
Relationships Between Invertebrate Phyla Based onFunctional-Mechanical Analysis of the
Hydrostatic Skeleton1
WOLFGANG FRIEDRICH GUTMANN
Forschungs-Institut Senckenberg, 6000 Frankfurt am Main, W. Germany
SYNOPSIS. The phylogeny of the major groups of deuterostome coelomates—the chor-dates, hemichordates and echinoderms—is discussed based on a mechanical-functionalanalysis of the hydrostatic skeleton and associated structures. The basic approach is tofirst establish transformation series of individual features and of functional complexes offeatures and second to determine their "Lesrichtung" by showing the direction of in-creased economy (i.e., better adaptation) with respect to environmental factors. It is ar-gued that a metameric coelom is primitive with respect to an oligomeric one and that theancestral form of the deuterostome coelomates is a metameric, coelomate worm-like an-imal with a complex set of circular, transverse and longitudinal body muscles. The coelomplus the complex body musculature formed the hydrostatic skeleton. The sequence ofstructural modifications leading to chordates is: (a) appearance of the notochord; (b)specialization of the dorsal longitudinal muscles with a reduction and disappearance ofthe transverse and circular muscles; (c) simultaneous appearance of the dorsal hollownerve cord; (d) development of a postanal tail; and (e) appearance and specialization ofthe branchial basket with gill slits as a filter feeding apparatus. The primitive chordatewould be most similar to the lancelet (Acrania). Tunicates are advanced chordates spe-cialized for sessile life and lost most chordate features in the adult, but retained them inthe larvae as adaptations for active dispersal. Enteropneusts (acorn worms) are anotheradvanced group specialized for burrowing in fine sediments and that evolved the anteriorproboscis as a peristaltic burrowing organ. The notochord was lost as was the dorsal nervecord and segmented condition of the coelom. A collar originated as a means to preventdischarged water from re-entering the mouth. Pterobranchs arose from enteropneust-like forms; their major structural changes are reduction of the branchial basket and mod-ification of the collar into tentacles which are associated with life in a closed tube. Finally,echinoderms arose from a pterobranch-like ancestor by specializing for sessile life andfeeding with tentacles and by final loss of the branchial basket. Groups such as the tuni-cates, hemichordates and echinoderms could be eliminated as ancestral forms within thedeuterostome coelomates because the evolution of acraniates and vertebrates from eachof these groups would involve the appearance of gill slits before the notochord and/orthe evolution of a metameric coelom from an oligomeric one, both of which are exceed-ingly improbable. Central to the methods used to establish the transformation series offeatures and their direction of evolutionary change (Lesrichtung) are functional (me-chanical) analysis and adaptive interpretation of features; hence, functional-adaptive anal-yses are an integral and essential part of the methodology of phylogenetic investigation.
INTRODUCTION , , , ,, , , . . . . . _ , the hydrostatic skeleton and on the evo-
The phylogenetic relationship of the l u d o n o f t h e c o e l o m i n c l u d i n g t h e q u e s .phyla and subphyla of animals has been d o n o f t h e primitiveness of the metamericthe focus of active research ever since the a r r a n g e r r i en t of coelomic cavities, I under-pubhcationofthe Origin of Species, but t o o k a research project with the coopera-with almost no consensus of opinion de- d o n o f s e v e r a l colleagues at the Sencken-yeloping. Moreover, no methods existed . Museum (K. Bonik, T. L. Franzen, M.for choosing in any convincing way be- Gra
&sshof f ) D . Mollenhauer, D. S. Peters;
tween the various conflicting phylogenetic t h e r e s u k s a r e p r e s e n t e d i n Schafer, 1973,hypotheses for the invertebrate phyla. Us- 1 9 7 5 i g 7 6 , g 7 8 ) w k h t h e m a j o r ,s o f
ing ideas on the functional properties of e s t a b l i s h i n g m a j o r metazoan groups andelucidating their phylogenetic relation-ships. The basic assumption underlying
'From the Symposium on Functional-AdapUveAnal- ^ . { h • e v o l u t i o n a r y
ysts in Systematics presented at the Annual Meeting or r J • •• • /the American Society of Zoologists, 27-30 December changes are gradualists and adaptive as1979, at Tampa, Florida. advocated by Simpson (1953) and Bock
63
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

64 WOLFGANG FRIEDRICH GUTMANN
(1979) and that a functional-adaptive anal-ysis, as outlined by Bock (1981) in this sym-posium, is needed in addition to other per-tinent evolutionary analyses. Because themorphological systems used to inquire intothe phylogenetic relationships of the meta-zoan phyla operate as mechanical ones, itis essential to use physical laws of mechan-ics in the functional analysis. Soft bodiedinvertebrates possess a hydrostatic skele-ton; hence, the physics of hydrostatic sys-tems is needed for the functional analysisof the skeleto-muscular system of these an-imals. A last requirement, stemming fromthe demands of investigating hydrostaticskeletons and from general theoreticalconsiderations advanced by workers asDullemeijer (1974), is that the organismmust be studied as a functional whole.Complete, enclosed structural systemscomprise the functional units of hydro-static skeletons and are, therefore, the pre-requisite units of analysis. Conclusionsabout evolutionary changes in one featureshould be made against a background ofthe evolutionary modifications and func-tional properties of the entire complex.
With the use of these principles, my col-leagues in Frankfurt and I developed anew concept for the phylogeny of meta-zoan phyla and, more importantly, a meth-od for testing the validity of the varied hy-potheses available in the literature. In thispaper, I would like to present the centralideas of our approach using an abridgedanalysis of the phylogeny of the deutero-stome coelomates as an example. More de-tails my be found in Gutmann (1966, 1967,1969, 1971, 1972, 1975, 1977); Bonik1977/78); Gutmann and Bonik (1979);Bonik et al. (1976, 1977a); Bonik et al.(1978); Gutmann et al. (1978); Vogel andGutmann (unpublished).
BLOMECHANICAL PROPERTIES OF THEHYDROSTATIC SKELETON
All chemical and physical properties oflife are associated with and tied to liquidsolutions enclosed in membranes. Becauseof this inseparable concurrence, every liv-ing organism is inevitably a hydrostaticskeleton construction. In metazoans, themultitude of cells is aggregated into com-plex systems mechanically supported by
connective tissue formed in the interstitialspaces. The metazoan constructional sys-tem must not be interpreted as simple ag-gregations of cells whose functional prop-erties are the sum of the functionalproperties of the cells because the mechanQical intactness of the metazoan body is notprovided solely by cells. Every metazoanhas a complex configuration of extracel-lular mechanical structures (gels, fibers,etc.) secreted by cells. The organization ofthese extracellular materials and their in-teractions with the cells forms the basis forthe mechanical properties of the metazoanbody. In many (or all) primitive metazo-ans, the skeleton is built on a grid of con-nective tissue fibers surrounding the fluid-filled cells (Bonik et al., 1977a). The wholesystem may be braced further by internalmuscles or incorporated into a coatlikemuscular and connective tissue sheath.Muscles are always attached to the connec-tive tissue structures forming a continuouslattice throughout the entire body of themetazoan animal.
Unless deformed by internal or externalforces or supported by bracing structures,fluid filled constructions possess a strongtendency to assume a globular form. Thisshape is, if very few exceptions are per-mitted, inappropriate for the fulfillment ofuseful organismic activities, especially lo-comotion. For greater efficiency, the vastmajority of biological activities depend ona body shape other than that of a sphere.For most lower animals, movement de-pends on a controlled deformation of thebody. The network of braces formed bymuscular bands and/or of sheaths of mus-cular and connective tissue structures pro-vide the necessary control of body defor-mation and form an indispensableprecondition for almost all useful biologi-cal activities in soft-bodied metazoans.
All lower animals lacking a rigid skele-ton move by deformation of the wholebody; the only exception is those formspropelled by ciliary strokes. Locomotionby peristaltic waves and swimming by si-nusoidal movements are dependent on thehydrostatic skeleton and can be most effi-ciently executed by elongate, bilateral an-imals, e.g., worms. As a consequence of thebiomechanical properties of the hydro-
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 65
static skeleton, which was possessed by thefirst metazoans, and the demands of loco-motion in a fluid environment, a worm-like body shape evolved in the earliest free-living metazoans. This body construction
^presumably evolved several times indepen-dently if free-living metazoans arose sev-eral times. The evolution of a worm shapeis governed by the environmental de-mands and the structural (mechanical) andfunctional (physiological) requirements ofthe metazoan body construction.
Compared to the efficiency of typicalrigid skeleto-muscle systems, i.e., thosewith a hard, nondeforming skeleton foundin most higher animals, all hydraulic con-structions have a relatively poor locomo-tory performance. The low level of econ-omy results from the need to use activelyall muscular support of the body to pre-vent deformations that would hinder orprevent locomotion. Unlike the action ofmuscles in rigid skeletal systems in whichmuch of the generated force contributesdirectly to moving the body, a great pro-portion of the muscles in a hydraulic sys-tem cannot contribute directly to propul-sion. Movement in metazoans withhydrostatic skeletons is brought about bythe indirect activity of the entire body mus-culature which generates pressure on thefluid filling, thereby making it rigid andpreventing undesired deformations ofbody shape. The force produced by a smallpart of the active body musculature con-tributes directly to moving the body at anytime. Improvement of locomotory effi-ciency in organisms with rigid skeletonscompared to those with a hydrostatic skel-eton is usually achieved by the eliminationof those muscular components which donot contribute directly to locomotion, butare necessary for maintenance of the hy-drostatic skeleton. This reduces the massof muscular tissue in the organism {i.e.,lessens the total mass that has to be movedduring locomotion) and the amount ofmetabolic energy required by the muscularsystem during locomotion. The result is in-creased efficiency measured in terms ofamount of energy required to move simi-lar sized animals.
Hydrostatic skeletons may be achievedby several different morphological sys-
tems. Their basic structure is a muscularand/or fibrous sheath surrounding com-pletely a fluid filling. The outer coat maycontain circular and longitudinal musclesor longitudinal muscles alone. Additionalbracings of transverse muscles may bepresent. The fluid filling may be composedof a gel from soft (jelly) to stiff consistency(Bonik et al., 1977a), a liquid-filled sac(e.g., a coelom) (Chapman, 1958; Gut-mann, 1960, 1966, 1967; Clark, 1964,1979; Trueman, 1975), muscles (e.g., themammalian tongue or the cephalopodarm), etc. These systems vary in propertiessuch as stiffness and in overall efficiency.Liquid filled sacs have several advantagesover other hydrostatic skeletal structures,e.g., soft gels. Evolution of internal cavitiesin animals (e.g., coeloms, pseudocoels) hasalmost certainly occurred under the actionof selection for a more efficient hydrostaticskeleton associated with better abilities oflocomotion.
SEGMENTATION OF THE COELOM
Fluid-filled body cavities (coeloms, pseu-docoels) occur in two basic forms, the me-tameric condition and the oligomeric con-dition. A metameric coelom is one in whichthe body, and hence the coelom is subdi-vided into a large number of units by mus-cle-bearing transverse septa—the dissepi-ments—which meet the mesenteries in thesagittal plane and suspend the gut. Otherorgans, e.g., the nephridia, are associatedwith the transverse septa. An oligomericcoelom is one in which the body cavity isundivided or divided into two or threelarge cavities. Transverse septa are lack-ing; and hence the gut is suspended onlyby midsagittal mesenteries. A major dif-ference between the metameric coelomand the oligomeric coelom is that in theformer, the hydrostatic skeleton in the dif-ferent body segments can act indepen-dently of one another while in the latter,the hydrostatic skeleton of the whole ani-mal acts as a single unit. Furthermore thetransverse bracing allows additional con-trol of the cross section of the body whichis automatically circular in hydraulic sys-tems lacking an internal bracing.
A major question in invertebrate phy-logeny is whether the metameric coelom
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

66 WOLFGANG FRIEDRICH GUTMANN
or the oligomeric coelom represents theprimitive condition in the evolution ofbody cavities. Most workers (Siewing,1972; Reisinger, 1973; Remane, 1973)have postulated that the oligomeric con-dition is primitive because it has a simplermorphology in comparison to the morpho-logically more complex metameric condi-tion. Evolution from the primitive oligo-meric coelom to the advanced metamericcondition would involve the appearanceand gradual enlargement of transversesepta subdividing the coelom and the ap-pearance of transverse muscles in thesesepta. Additional features would have toevolve in a segmented arrangement, suchas the nephridia which are associated withthe dissepiments and whose tubules usu-ally extend from one body cavity to thenext through the wall of the dissepiment.
The conflicting theory is that the meta-meric coelom is the primitive condition.The transverse septa with their transversemuscles and structures such as the ne-phridia are present in the acoelomate jellysupported animal. The internal cavitiesappear between the septa and enlarge un-til successive cavities meet at the preexist-ing septa, transforming them into muscu-lar dissepiments. Evolution to the advancedoligomeric condition would occur by thegradual reduction and disappearance ofthe transverse muscles and the dissepi-ments.
Both hypotheses must be tested by pro-viding, among other arguments, a func-tional-adaptive analysis for the postulatedmorphological changes from the suggest-ed primitive condition to the advancedone, using the assumption that the evolu-tionary changes are gradualistic ones (Pe-ters and Gutmann, 1971; Schafer, 1973,1975, 1976, 1978; Bock, 1979). Possiblereversal of evolutionary change must alsobe considered (Gutmann, 1976; Bonik etal., 1977ft; Bock, 1979).
Origin of the metameric coelom fromthe acoelomate condition is postulated tobe by a segmental series of body cavitieswithin the gridwork of muscles graduallyreplacing the jelly and/or parenchymal fill-ing of the body. The existing transversemuscular bands were condensed between
neighboring coelomic cavities and becameassociated with the double-walled trans-verse mesentery formed when the epithe-lium of adjoining cavities pressed together.These transverse muscles continued theirrole of maintaining body shape (GutmannjJ1972; Bonik et al., 1977a). They nowserved, in addition, to isolate the individ-ual parts of the segmented hydrostaticskeleton characteristic of the metamericcoelom (Clark, 1964, 1979). Evolutionfrom the metameric to the oligomeric coe-lom involves the disappearance of the dis-sepiments and their transverse muscles.This can occur in gradual steps with theappearance of holes in the transverse septaby which fluid can move from one coelo-mic cavity to another. Enlargement ofthese holes would increase movement offluid from one cavity to another and equal-ize pressure throughout the entire coelom.Evolutionary modification from a com-plete metameric condition to a fully oligo-meric coelom can easily be conceived as aseries of gradualistic steps as the change isa loss of existing features. An oligomericcoelom is characteristic of metazoans inwhich locomotion is reduced or in whichthe hydrostatic skeleton had been replacedby a rigid skeleton; in both cases a hydro-static skeleton consisting of a series of sep-arate liquid filled sacs is no longer advan-tageous.
Origin of an oligomeric coelom wouldbe by the appearance of fluid-filled sacs,but these would enlarge and merge into asingle large cavity within the body. The lin-ing epithelium of individual coelomic sacswould break down wherever they mergedresulting in the joining of separate cavitiesupon meeting. Existing bands of trans-verse muscles in the acoelomate ancestordisappear in the primitive oligomericmetazoan. Although an oligomeric coelomcould serve as a hydrostatic skeleton, it isless advantageous than that formed by ametameric coelom because the entire bodyacts as a single unit. Although the fullydeveloped oligomeric coelom can serve asa hydrostatic skeleton, the intermediarystages in its evolution cannot be reasonablyexplained. Locomotion in worm-like or-ganisms depends on a homogeneous body
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 67
structure over the whole length. Emer-gence of few fluid-filled cavities wouldhave produced very heterogeneous seg-ments which would have functioned to-gether as an integral unit with difficulty,
win addition, evolution from the oligomericto the metameric coelom requires the for-mation of segmental transverse septa,transverse muscles and associated segmen-tal features as the nephridia which have acomplex relationship with the transversedissepiments. The segmental arrangementof the septa must be regular for greatestefficiency. Considering only the septa andtransverse muscles, serious problems existfor a gradualistic explanation of their ap-pearance in an oligomeric coelom and oftheir evolution to the metameric condition(Clark, 1964, 1979; Mettam, 1971; Schafer,1973). The important function of thetransverse septa is to subdivide the coelominto separate hydraulic chambers; thisworks only when the septa are complete.No functions and adaptive advantageshave been suggested for incomplete septaand for the adaptive evolution from ru-dimentary septa to complete ones. Trans-verse muscles must evolve from existingsets of muscles and it is difficult to explainthe origin of transverse muscles from ex-isting longitudinal or circular muscles.Moreover transverse muscles must extendacross the body to function properly, i.e.,they must attach to resisting structures ateither end and actively contract during thelife of the individual organism. Nonfunc-tioning muscles atrophy rapidly during thelife of an individual. Transverse muscleswould possess little or no functional oradaptive significance in a rudimentary sep-tum. Lastly, the set of transverse musclesmust be well developed in the fully formeddissepiments if the transverse septa are toisolate the coelomic cavities into indepen-dent hydraulic units.
The transverse dissepiments are com-plex morphological systems of coelomicepithelium, transverse muscles, nephridia,etc. which had to evolve as a unit. Theycould not evolve adaptively and graduallyunder the control of selection forces asso-ciated with properties of the metamericcoelom because these functional proper-
ties operate only when the septa are com-plete. I know of no other functional prop-erties and associated selection forces whichcould be responsible for a gradual evolu-tion of the dissepiments from an ancestraloligomeric condition. Unless one is willingto accept the possibility of a single step sal-tatory evolution of complete dissepiments,an adaptive gradual evolution of the me-tameric coelom from the oligomeric coe-lom appears highly improbable.
Several conclusions may be offered:
a) The metameric coelom is primitive andthe oligomeric coelom is advanced incoelomates.
b) The oligomeric coelom can evolvegradually and adaptively from themetameric coelom by loss of the dissep-iments and transverse muscles, but thereverse change is highly improbable.
c) The oligomeric coelom probablyevolved independently in several groupsof metazoans.
EVOLUTION OF CHORDATE FEATURES
Among the coelomate metazoans is acomplex of groups known as the deutero-stomes which include the hemichordates,tunicates, acrania, vertebrates and echi-noderms. A strong consensus exists thatthese taxa form a monophyletic groupwithin animals, but little agreement existson the phylogeny within the deutero-stomes. I would like to present a model forthe evolutionary origin and subsequentchange for the major features in thisgroup and to compare this model with oth-er hypotheses. Because I believe that thechordates are the primitive stock of thesedeuterostomes, I will be concerned withthe evolution of the major chordate fea-tures. These will be discussed in the se-quence of their presumed evolutionary or-igin.
The deuterostome coelomates evolvedfrom a worm-like acoelomate ancestor pos-sessing a hydrostatic skeleton consisting ofa complex, dense muscular grid (longitu-dinal, circular and transverse) surround-ing a gel filling. The metameric coelomdeveloped from inflated regions of a canalsystem with the newly developed coelomic
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022
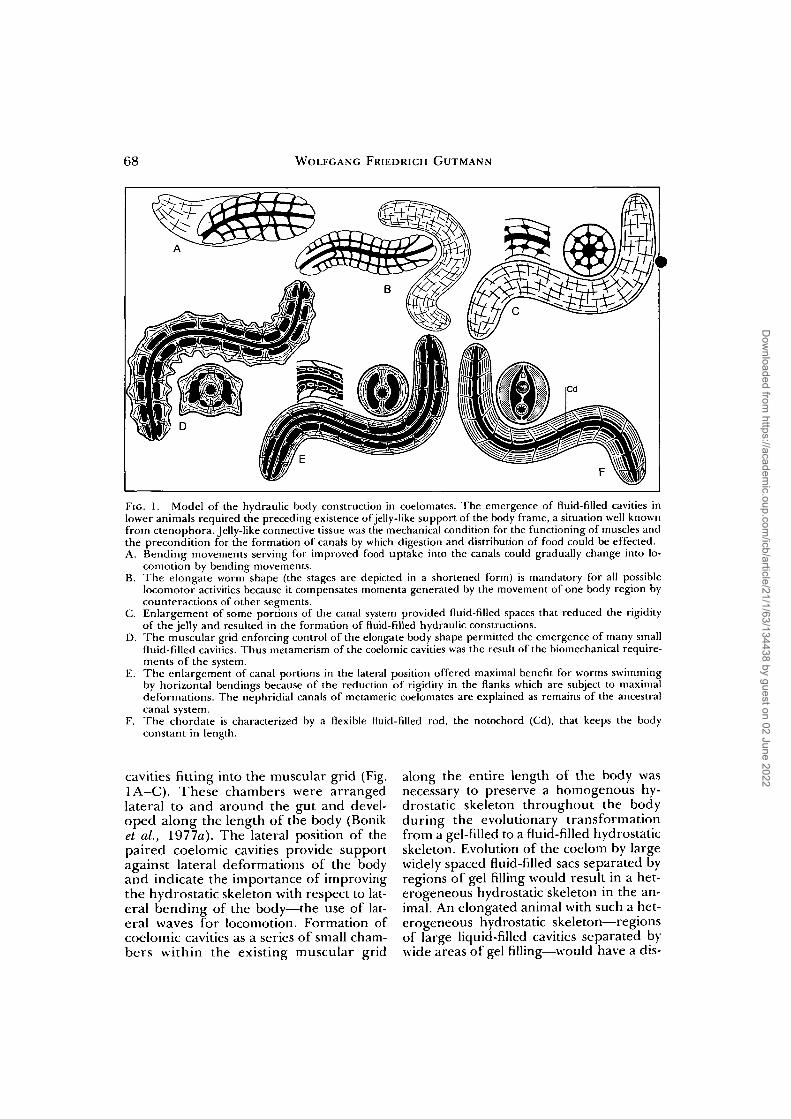
68 WOLFGANG FRIEDRICH GUTMANN
FIG. 1. Model of the hydraulic body construction in coelomates. The emergence of fluid-filled cavities inlower animals required the preceding existence of jelly-like support of the body frame, a situation well knownfrom ctenophora. Jelly-like connective tissue was the mechanical condition for the functioning of muscles andthe precondition for the formation of canals by which digestion and distribution of food could be effected.A. Bending movements serving for improved food uptake into the canals could gradually change into lo-
comotion by bending movements.B. The elongate worm shape (the stages are depicted in a shortened form) is mandatory for all possible
locomotor activities because it compensates momenta generated by the movement of one body region bycounteractions of other segments.
C. Enlargement of some portions of the canal system provided fluid-filled spaces that reduced the rigidityof the jelly and resulted in the formation of fluid-filled hydraulic constructions.
D. The muscular grid enforcing control of the elongate body shape permitted the emergence of many smallfluid-filled cavities. Thus metamerism of the coelomic cavities was the result of the biomechanical require-ments of the system.
E. The enlargement of canal portions in the lateral position offered maximal benefit for worms swimmingby horizontal bendings because of the reduction of rigidity in the flanks which are subject to maximaldeformations. The nephridial canals of metameric coelomates are explained as remains of the ancestralcanal system.
F. The chordate is characterized by a flexible fluid-filled rod, the notochord (Cd), that keeps the bodyconstant in length.
cavities fitting into the muscular grid (Fig.1A-C). These chambers were arrangedlateral to and around the gut and devel-oped along the length of the body (Boniket ai, 1977a). The lateral position of thepaired coelomic cavities provide supportagainst lateral deformations of the bodyand indicate the importance of improvingthe hydrostatic skeleton with respect to lat-eral bending of the body—the use of lat-eral waves for locomotion. Formation ofcoelomic cavities as a series of small cham-bers within the existing muscular grid
along the entire length of the body wasnecessary to preserve a homogenous hy-drostatic skeleton throughout the bodyduring the evolutionary transformationfrom a gel-filled to a fluid-filled hydrostaticskeleton. Evolution of the coelom by largewidely spaced fluid-filled sacs separated byregions of gel filling would result in a het-erogeneous hydrostatic skeleton in the an-imal. An elongated animal with such a het-erogeneous hydrostatic skeleton—regionsof large liquid-filled cavities separated bywide areas of gel filling—would have a dis-
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 69
advantage because the properties of thesetwo systems differ greatly and it wouldbe difficult to integrate them into a singlefunctional unit. Consequently the coelomevolves in a metameric fashion leaving in-
Wact the muscular control of the body.The primary advantage obtained by the
formation of the metameric coelom is thedecrease in stiffness in the body construc-tion when the gel filling is replaced by seg-mental fluid chambers. The skeleton re-mains a hydrostatic one dependent uponthe action of the entire body musculaturefor maintaining the skeleton during loco-motion and hence remains at a low level ofefficiency. A gel-filled hydrostatic skeletonpermits the animal to crawl on the sub-strate or swim, by sinusoidal waves, in afluid; however, burrowing in soft, fine-grained sediments is not possible. The lat-eral position of the coelom indicates itsadaptive advantage in an animal locomot-ing with lateral bends of the body. Oncethe coelom became fully developed, theanimals could burrow in soft, fine-grainedsediments. So the evolution of the coelomand its metamerism in burrowing organ-isms as sometimes suggested (Clark, 1964,1979) is highly improbable.
Evolution of the coelom was the firststep in the evolution of features in the deu-terostome coelomates from their acoelo-mate ancestors (Fig. 1D-E). The animal atthis point would be an elongated wormwith a metameric coelom and a completeset of body musculature (i.e., transverse,circular and longitudinal). Although themain locomotory performance might havebeen swimming, these forms could burrowas well as creep on the substrate. Reduc-tion of stiffness in the lateral directionleading to more efficient locomotion ap-pears to be the adaptive significance of thisstep.
The second step is a continuation of thefirst in that the adaptive significance is fur-ther increase in efficiency of locomotion.The important change was the evolutionof a stiff rod—the notochord—dorsal tothe gut as the major axis of support for thebody (Fig. IF). Although the notochordoperates as a hydrostatic skeleton, it is in-termediate between hydrostatic skeletons
FIG. 2. Upper stage: biomechanical preconditionsfor the development of the notochord lie in the ex-istence of transverse dissepimental bracings of a me-tameric worm. Dissepiments prevent bending of thebody axis within the body cavity. Lower stage: theconsequences of lacking dissepiments is shown in theworm-like organism in which the notochord wouldbe useless because it could not contribute to the main-tenance of the body length.
formed by fluid filled sacs and rigid skel-etons formed of cartilage or bone. It hasseveral advantages over a coelom as a bodyskeleton. First, it is smaller and requiresless space within the body, being composedof cells filled with a stiff gel and surround-ed by a heavy fibrous sheath. Second, itdoes not require the action of the bodymusculature to maintain its constancy ofbody length.
Prior to the evolution of the notochord,the animal could locomote by sinusoidalundulations in which the body is held con-stant in length by the circular and trans-verse muscles and by peristalsis in whichthe length of the body changes by the al-ternate contraction of the longitudinal andcircular muscles. With the evolution of thenotochord, body length is held constant bythis rod and movement by peristalsis is en-tirely eliminated (Fig. 2).
Evolution of the notochord is dependentupon the presence of the transverse dis-sepiments. These structures support thenotochord in its position dorsal to the gutand prevent it from bending within theanimal. Basically the dissepiments anchorthe notochord to the system of body mus-culature which is an essential requirementif the notochord serves as the central, non-compressive support of the body. If the
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

70 WOLFGANG FRIEDRICH GUTMANN
FIG. 3. Phylogenetic model for the evolution of the chordate body construction starting with a worm-likemetameric organism.A. Metameric coelomate equipped with paired coelomic cavities (C) which are integrated into the three
dimensional muscular grid. The whole system works as a hydrostatic skeleton. Shape control is exertedby the muscular bracings. The gut (D) is suspended in the mesenterium (Me) and held in its position bytransverse muscles bearing tissue sheets (Dis).
B. The notochord (Cd) arises above the gut in the muscle-connective tissue grid of the mesenterium. Thefluid filling in the emerging notochord is held in its position. The muscular and connective tissues con-trolling the shape of the notochord aid the muscular bracing in ensuring constancy of length while theworm swims by lateral undulations.
C and D. As the function of the notochord to ensure constancy of length is improved, the circular musclesof the muscle coat of the body wall and the transverse muscles of the dissepiments are reduced. Thedissepiments persist as connective tissue sheets (My) which connect the strengthened longitudinal musclepackages to the body axis.
E, F and G. Deepening mouth slits allow water to flow out of the mouth after being used for filtration. Thegill slits of the branchial basket (Ks) develop by subdivision of the slits by dorsoventral bars. This isnecessitated by the weakening of the body flanks.
H. Typical chordate construction as represented in the acrania. All characteristic organs of the chordates aredepicted: notochord (Cd), metameric muscle packages (My), branchial basket (Kd), dorsal nerve chord(Nr). Whereas the body cavity containing the gut is devoid of subdivisions, the coelomic cavity showssegmental vestiges in the form of fluid filled compartments in the niyotomes (St).
notochord bends internally in the body asthe longitudinal muscles contract, it will nolonger maintain the constant length of thebody. Constant length of the body is anessential requirement for locomotion bylateral undulations.
Subsequent to the evolution of the no-tochord, the transverse and circular mus-cles could disappear as they are no longeressential for the integrity of the hydrostaticskeleton and because peristalsis is no long-er possible (Fig. 3A-D). Reduction of thesemuscles was advantageous as it reducedthe bulk of the animal and the amount ofenergy needed for muscle contraction andmaintenance. Loss of these circular andtransverse muscles presumably occurredsoon after the evolution of the notochord.
Together with the reduction of the cir-cular and transverse muscles, the dorsalpart of the longitudinal muscles enlargedand became the major set of locomotorymuscles. These muscles and only thesemuscles could enlarge because they arepositioned next to the notochord which isnow the only anticompressive structuralelement in the body. The notochord canresist effectively the compressive forcesplaced on the body axis by the locomotorymuscles only if these muscles lie close to it.With the enlargement of the dorsal longi-tudinal muscles, development of the cen-tral nervous system in a dorsal position isadvantageous because it reduces thelength of the peripheral nerves to the mus-cles. Thus the dorsal nerve cord evolved
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 71
in conjunction with the dorsal longitudinalmusculature. Shift of the dorsal nerve cordfrom a superficial position to the presentposition just dorsal to the notochord pro-vided greater mechanical protection (Fig.WH).
Metamerism of the body was retained atthe chordate level because the thick pack-age of segmental longitudinal muscles(myotomes) had to remain connected tothe body axis. The dissepiments evolvedinto myosepta with the loss of transversemuscles. The myosepta subdivide the lon-gitudinal muscles into metameric unitswhich are retained throughout the verte-brates.
The metameric arrangement of the pe-ripheral spinal nerves was a consequenceof the mechanical system of muscles andtransverse septa. The peripheral nervescould fit readily into a segmental subdivi-sion of myotomes and myosepta.
After the evolution of the notochord,enlargement of the dorsal longitudinalmuscles and the specialization of the cen-tral nervous system as a dorsal nerve cord,this complex of features could be extendedbeyond the end of the digestive tract as apostanal tail (Fig. 3H). This locomotorysystem, which is unique to chordates, isdependent upon the evolution of the no-tochord as the central support of the bodyand the subsequent development of thedorsal musculature and nerve chord. Apostanal tail could not evolve so long asbody support was provided by a coelom-based hydrostatic skeleton.
The last major chordate feature to ap-pear was the branchial basket. This filterfeeding structure is dependent on a seriesof gill slits through the body wall connect-ing the anterior end of the digestive systemwith the outside (Fig. 3E-G). These slits inthe body wall could only appear and be-come specialized after the notochordevolved and assumed the full role of axialsupport. At an earlier stage in deutero-stome evolution when body support wasprovided by a hydrostatic skeleton, regard-less of whether it was a gel-filled system ora metameric coelom, the slits in the lateralbody wall would have completely disrupt-ed the hydraulic system in the anterior endof the body. The body wall has to remain
FIG. 4. Model for the evolution of the craniote con-struction. The two stages continue the model of Fig-ure 1, H, into the craniote level of organization. Themajor changes consist of the formation of a cephalicregion (K) equipped with large sense organs and thebrain in which bending movements are suppressedby stiffening skeletal structures. The efficiency of themotoric apparatus, the precondition for the emer-gence of a stiff head region, is increased by the de-velopment of stabilizing osseous scales (S) which inthe next stage of vertebrate evolution are replaced bya more economical internal grid of skeletal elements.
turgid throughout the entire hydrostaticskeleton containing fluid filled cavities un-der pressure. Either the gill slits would beforced closed by the turgid hydrostaticskeleton, or the open gill slits would dis-rupt the turgidity of the hydraulic systemand the anterior end of the body would belimp. Thus only after the establishment ofthe notochord and the myotomes had pro-vided the needed support in the dorsolat-eral part of the body, could the branchialbasket evolve. The incipient stages of thisfilter feeding system consisted of slit-likedeepenings of the mouth which graduallybecame subdivided into separate gill clefts.
To summarize: The evolution of themajor chordate features from a worm-likemetameric coelomate ancestor are in thefollowing order:
a) notochord;b) specialization of dorsal longitudinal
muscles together with reduction of thetransverse and circular muscles;
c) a simultaneous specialization of the
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

72 WOLFGANG FRIEDRICH GUTMANN
FIG. 5. Phylogenetic model for the evolution of hemichordates and echinoderms from acrania-like chordateancestors.A. Chordate construction which can live and filter feed in the sediment because the water current is driven
through the branchial basket (Ks) by the cilial lining. In compensation for the locomotor activity thebranchial basket is enlarged by multiplication of the gill slits.
B. Formation of muscular proboscis (R) allows the intermediate stage to move in the sediment. The notochord(Cd) and the spinal chord (Nr) are reduced in the posterior part of the body. The cross section shows thereduction and opening of the atrium from the atrioporus.
C. Enteropneust-like construction showing vestiges of notochord (Cd), nerve chord (Nr), and longitudinalmuscles (Lm) in the hind part of the body. Metamerism is entirely reduced because the suspensorystructures for the notochord are no longer required. The body became divided into three compartments:the proboscis (R), the collar (Kr), and the posterior part.
D. The pterobranch-construction arose by transformation of the collar region into a tentacular apparatus(T) which was protruded from the burrow. Its emergence was accompanied by the almost total reductionof the branchial basket (Ks—vestigial gill slits).
E. Transitory stage to echinoderms. The tentacular apparatus that corresponds to the collar region is ar-ranged in a radial way. Some (Ks) gill slits are still present. The proboscis becomes reduced.
F. Primitive echinoderm construction. The body frame is supported by a calcareous skeleton (Cs), the ten-tacular apparatus (T) continues to function as a hydraulic system.Whereas the primitive echinoderms were sedentary organisms, the advanced eleutherozoans lost con-tact with the substrate and became freely moving forms: G. Asteroid; H. Echinoid.
central nervous system as a dorsalnerve cord;
d) development of a postanal tail;e) origin and specialization of the bran-
chial basket with numerous gill slits asa filter feeding apparatus (follows "c").
The chordate group which has the struc-ture seen at this stage of chordate evolu-
tion is the lancelet (Acrania) which can beconsidered as representing the most prim-itive group of known chordates and thestock from which other deuterostome coe-lomates have evolved. I will not trace theevolution of vertebrates from their acraniaancestor (Fig. 4) as this evolution is mainlya further specialization of the featurespresent in the lancets and little disagree-
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 73
ment exists about this aspect of chordatephylogeny (Gutmann, 1972, 1975, 1977).
FROM ACRANIA TO HEMICHORDATES
Typical chordates (e.g., lancelets) might€bt be considered as real candidates forburrowing in the mud. The notochord andthe longitudinal muscles arranged in myo-tomes do not enable these animals to pen-etrate into fine sediment because only lat-eral undulations are possible. Thesemovements are, of course, sufficiently ef-fective to allow wriggling into coarse sedi-ment (sand) in which filter-feeding couldbe continued. The coarse sediments allowthe water currents to pass through its in-terstices. Organisms such as Branchiosto-ma, which are fully equipped for active lo-comotion, have acquired a sessile way oflife in coarse sediments. Energy used forlocomotion could be saved when trappingof food was improved by enlargement ofthe branchial basket. The long extensionof the branchial basket and the large num-ber of gill slits have been considered to bean adaptation to a sedentary mode of life.These features and others not mentionedhere are not typical for the chordate con-struction ancestral to the other branches ofdeuterostomes (Gutmann, 1966, 1967,1972; Bonik and Gutmann, 1977; Gut-mann and Bonik, 1979).
In some acrania-like group, the animalsstarted to burrow in finer and finer sedi-ments in which lateral undulations wereless suitable for movement. Moreover asthe sediments became finer, the animalhad to form a tube for water flow (Fig. 5A).In one of these groups, the anteriormostend of the body (anterior to the mouth andbranchial basket) must have modified intoa hydrostatic organ that allowed the ani-mal to burrow in muddy sediments byperistaltic movements (Fig. 5B). The orig-inal function of the rudimentary proboscismay have been the opening of a hole inthe sediment through which the inhalentcurrent of water could enter the mouth.With further development of the probos-cis, penetration into mud and active loco-motion through it were achieved gradu-ally. A burrow is formed by the animal inwhich it lives and through which the ciliaof the branchial basket drive a water cur-
rent. The proboscis has a hydrostatic skel-eton with a complex arrangement of mus-cular braces. It lacks a notochord-like axialsupport and hence can move by peristalticaction. It permits active but slow locomo-tion in muddy sediments of a worm-likeanimal.
The notochord was reduced because itsstiffness and its function to preserve con-stancy of body length were disadvantageousin the new environment. The animalevolved a worm-like body which is suitableto life in an U-shaped tube. Longitudinalmuscles were retained as they serve toshorten and pull along the elongated pos-terior part of the body which drags behindas the anterior end moves forward by ac-tive burrowing action of the proboscis.Muscles are not needed to stretch the bodywhich is pulled out passively as it contactsthe walls of the burrow during locomotion.Metamerism of the longitudinal muscleswas lost and the metameric coelom evolvedinto an oligomeric one as the function ofthe transverse septa to suspend the noto-chord in the body became obsolete as aconsequence of reduction of the noto-chord and the septa. Vestiges of thenotochord persist as the stomochord (Fig.5C). In addition to this vestige of thenotochord, a short piece of neural tube re-mained in the collar region of the body.The preponderance of longitudinal mus-cles, the vestiges of the notochord andneural cord are strong evidence for thechordate origin of the hemichordates.Their advanced position relative to prim-itive acrania is shown by the oligomericcoelom, the unsegmented longitudinalmuscles and the vestiges of the notochordand neural cord.
These worm-like chordates are the en-teropneusts (acorn worms) which are usu-ally placed in a distinct group—the hemi-chordates—which retained the filterfeeding branchial basket possessed by typ-ical chordates. The profound modificationof the body architecture is an adaptationto active burrowing in muddy sediments.
The three body regions of the enterop-neusts fulfill specific functions in theseburrowing animals. The proboscis is theactive hydraulic system for burrowing.The collar maintains contact with the wall
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

74 WOLFGANG FRIEDRICH GUTMANN
of the tube and prevents the water dis-charged from the gills from reentering themouth (Fig. 5C). The hind part comprisesmost of the body and houses the branchialbasket and the gut; it is dragged along pas-sively when the animal burrows with itsproboscis.
The oligomeric coelom of the enterop-neusts corresponds to the reduction of thenotochord and of the muscular system ofthese animals and evolved with the bur-rowing mode of living. It still serves as ahydrostatic skeleton but is a much poorerone than the metameric one of its ances-tors. In contrast to other burrowing wormsthat possess a hydrostatic skeleton, such asystem could not have arisen de novo in theenteropneusts because the gill clefts per-forate the body wall and because the nec-essary circular muscles for an efficient sys-tem could not develop gradually from thelongitudinal ones. Therefore the morpho-logical and functional properties of theirchordate forerunners (acrania-like forms)restricted the possibilities of of the enter-opneusts.
Enteropneust-like organisms gave rise tothe pterobranchs which still possess char-acteristic chordate features. They have anoligomeric coelom, only longitudinal mus-cles in the posterior region of the body,and vestiges of the notochord in the an-terior part of the body. These sessile formslive in a permanent tube secreted by theanimal. The proboscis is the only part thatcan actively move in this tube and pulls theanimal out for feeding. The hind regionis passively stretched and can only shortenactively to pull the pterobranch back intoits tube. This feature supports the hypoth-esis of the origin of pterobranchs fromchordate ancestors possessing only longi-tudinal muscles.
Whereas in the enteropneusts the chor-date branchial basket still functions in es-sentially the same way as in the typicalchordates, the gill clefts of the ptero-branchs are reduced to a single pair. Eventhis pair of gill slits is closed in some gen-era. In exchange for the gills, filter-feed-ing is performed by tentacles which havedeveloped from the collar of the enterop-neust-like ancestors (Fig. 5D). In the
acorn-worms, the inner lining of the collaris able to collect food particles on its sur-face. When the collar was enlarged inpterobranchs it was gradually transformedinto a tentacular apparatus which becamethe primary feeding apparatus with redu0tion of the gills. The branchial basket canfunction as a filter feeding apparatus foranimals living in a tube open at both ends,but it cannot operate in an organism livingin a closed tube. Change from the bran-chial basket to the tentacular feeding ap-paratus took place with the change in lifefrom a burrowing animal in an open tubeto a sessile one in a closed tube.
THE ECHINODERMS
Although the body construction of adultechinoderms is radically different fromthat of chordates, these phyla share a num-ber of embryological and biochemical fea-tures, and most workers agree that theyare closely related. A reasonable transfor-mation series was indicated by Grobben(1924), but was never described in detail;only a very sketchy model touching on themajor points will be presented here.
The evidence for the existence of gillclefts in primitive echinoderms was pre-sented by Jefferies (1975). Based on thisevidence, it may be concluded that ances-tral echinoderms possessed a branchialbasket inherited from their chordateancestors, but that this filter feeding ap-paratus, together with the gill slits, was lostearly in the phylogeny of the echidoderms.Because the adult morphology of echino-derms is so profoundly modified from thatof typical acrania-like ancestors, it is diffi-cult to suggest a reasonable transformationseries and to link possible intermediatestages with known groups. Most evidencesuggests a very close link between echino-derms and hemichordates so that I will as-sume that the pterobranchs or at least theenteropneusts represent the chordate con-struction from which echinoderms evolved.
The ancestors of echinoderms left thesediment and sedentary life in tubes inwhich their forerunners lived and attachedthemselves onto firm surfaces. Specializa-tion to sessile life resulted in transforma-tion of bilateral symmetry, still seen in
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 75
pterobranchs, to radial symmetry. In ad-dition, a stiff framework of superficial cal-careous skeletal plates evolved for me-chanical protection, another adaptationfor sessile life. These calcareous platesfirmed the basis of the rigid skeleton ofechinoderms by which they were able toovercome the mechanical limitation inher-ent in the possession of the oligomeric coe-lom and only longitudinal muscles in thehind region of the body in their hemi-chordate ancestors (Gutmann, 1972; Gut-mann and Bonik, 1979).
The tentacles that had evolved in thepterobranch-like forerunners continued tofunction as filter-feeding organs and wereheld upwards into the water current totrap small particles. The tentacles were re-organized into a pentaradial pattern incorrespondence with the radial symmetryof the body (Fig. 5E). The hydrostaticmechanism of the tentacles reflects thepentaradial configuration and is present inall echinoderms. It is connected to the col-lar coelom in which hydrostatic pressureis developed that expands and stiffens thetentacles. This coelomic system is separat-ed from the body wall and is, therefore,able to contract freely and independentlyof the stiff body wall when pumping fluidinto the tentacles.
Primitive echinoderms were stationaryanimals of the crinoid type (Fig. 5F), butthe advanced echinoderms, the eleuther-ozoa, evolved the ability to move activelyon their tentacles (Fig. 5G-H). Thischange was achieved when sedentary, ra-dial forms turned over onto their oral sur-face, whereby the tentacles contacted theground and were able to perform loco-motory movements. This astonishingtransformation could occur graduallywhen crinoid-like forms with flexible stalksbent their tentacle crown down to collectfood from the bottom. Walking on the sub-strate with the tentacles improved the ef-ficiency of feeding and was continued afterthe stalk lost contact with the substrate.
The evolution of echinoderms from anancestral pterobranch-like chordate stockcan be summarized with the followinglarge groups from most primitive to mostadvanced:
a) The branchio-echinoderms (Bonik etal., 1978) which are groups still pos-sessing a fully developed branchial bas-ket, presumably possessing tentacles,which were secondarily reduced. Thestalk which is still present in someforms has lost contact with the sub-strate;
b) the pelmatozoans which are sessilegroups with a stalk, pentaradial sym-metry, tentacles, but have lost the bran-chial basket including the gill slits;
c) the eleutherozoans which are free-liv-ing groups that have lost the stalk andhave turned over onto the oral surface.
This sketch of echinoderm evolutionfrom chordate ancestors is admittedlybrief as I am currently undertaking a de-tailed analysis of this question in coopera-tion with paleontologists. The resultswhich will be based on a functional-adap-tive analysis of living and fossil forms, willbe presented in future papers. We are es-pecially interested in determining whethersome of the soft-bodied fossils of the Bur-gess shale formation belong to the trans-formation series leading from the chor-dates to the echinoderms (Conway Morrisand Whittington, 1979).
THE TUNICATES
The phylogeny of the deuterostomeswould be incomplete without mentioningthe tunicates. The transformation seriesleading to these animals will be sketchedonly briefly herein; details can be found inGutmann (1975). Tunicates can be derivedeasily from typical chordates; they are ad-vanced, not primitive members of this phy-lum. The acrania-like forerunners of tu-nicates became stationary and anchoredtheir body to the ground. By avoiding lo-comotion, they economized on energyneeded to maintain a locomotory systemand for active locomotion. The whole lo-comotory apparatus, including the noto-chord, metameric coelom and most of thecentral nervous system, was lost in theadult organism (Fig. 6). The lack of loco-motion was compensated for by an enor-mous enlargement of the branchial basketand a more effective pumping mechanism.
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022
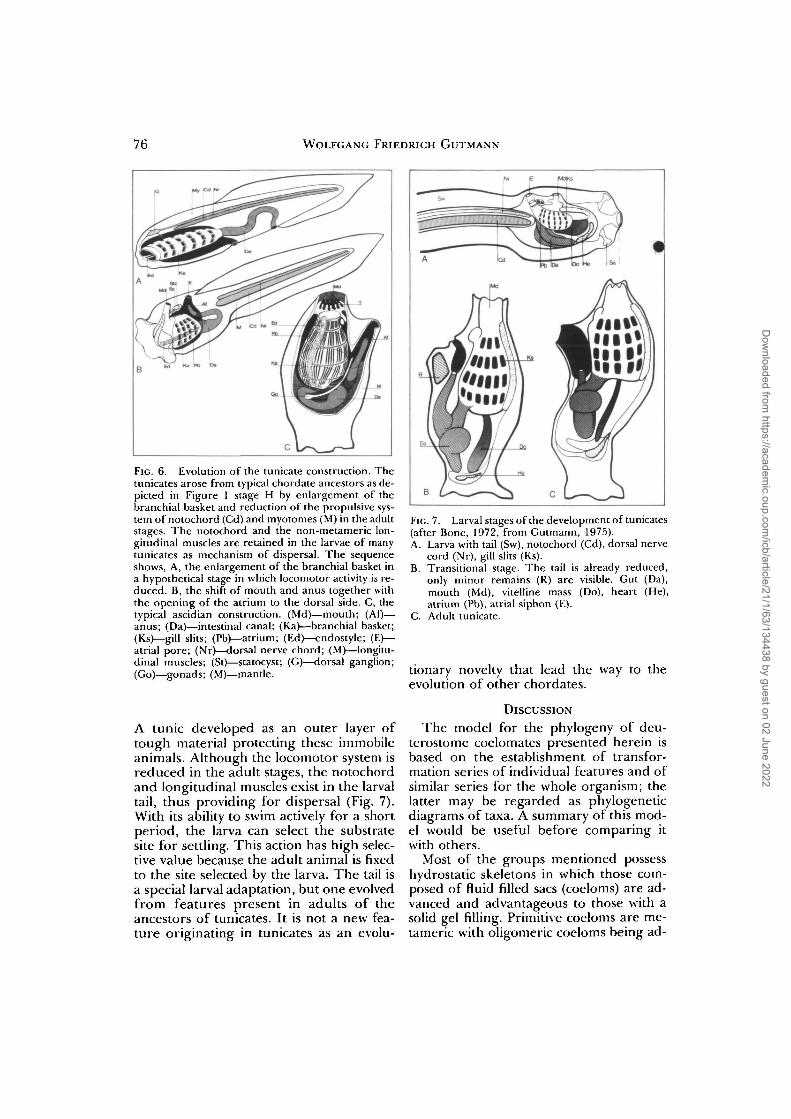
76 WOLFGANG FRIEDRICH GUTMANN
FIG. 6. Evolution of the tunicate construction. Thetunicates arose from typical chordate ancestors as de-picted in Figure 1 stage H by enlargement of thebranchial basket and reduction of the propulsive sys-tem of notochord (Cd) and myotomes (M) in the adultstages. The notochord and the non-metameric lon-gitudinal muscles are retained in the larvae of manytunicates as mechanism of dispersal. The sequenceshows, A, the enlargement of the branchial basket ina hypothetical stage in which locomotor activity is re-duced. B, the shift of mouth and anus together withthe opening of the atrium to the dorsal side. C, thetypical ascidian construction. (Md)—mouth; (Af)—anus; (Da)—intestinal canal; (Ka)—branchial basket;(Ks)—gill slits; (Pb)—atrium; (Ed)—endostyle; (E)—atrial pore; (Nr)—dorsal nerve chord; (M)—longitu-dinal muscles; (St)—statocyst; (G)—dorsal ganglion;(Go)—gonads; (M)—mantle.
A tunic developed as an outer layer oftough material protecting these immobileanimals. Although the locomotor system isreduced in the adult stages, the notochordand longitudinal muscles exist in the larvaltail, thus providing for dispersal (Fig. 7).With its ability to swim actively for a shortperiod, the larva can select the substratesite for settling. This action has high selec-tive value because the adult animal is fixedto the site selected by the larva. The tail isa special larval adaptation, but one evolvedfrom features present in adults of theancestors of tunicates. It is not a new fea-ture originating in tunicates as an evolu-
FIG. 7. Larval stages of the development of tunicates(after Bone, 1972, from Gutmann, 1975).A. Larva with tail (Sw), notochord (Cd), dorsal nerve
cord (Nr), gill slits (Ks).B. Transitional stage. The tail is already reduced,
only minor remains (R) are visible. Gut (Da),mouth (Md), vitelline mass (Do), heart (He),atrium (Pb), atrial siphon (E).
C. Adult tunicate.
tionary novelty that lead the way to theevolution of other chordates.
DISCUSSION
The model for the phylogeny of deu-terostome coelomates presented herein isbased on the establishment of transfor-mation series of individual features and ofsimilar series for the whole organism; thelatter may be regarded as phylogeneticdiagrams of taxa. A summary of this mod-el would be useful before comparing itwith others.
Most of the groups mentioned possesshydrostatic skeletons in which those com-posed of fluid filled sacs (coeloms) are ad-vanced and advantageous to those with asolid gel filling. Primitive coeloms are me-tameric with oligomeric coeloms being ad-
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 77
vanced. The ancestral stock of the deu-terostomes is postulated to be a worm-likeanimal with a metameric coelom.
Evolution of the basic chordate featuresoccurred in the following order:
T?) the notochord;b) reduction of the transverse and circular
muscles and specialization of the dorsallongitudinal muscles in a metameric ar-rangement;
c) specialization of the nervous systeminto a dorsal nerve cord and segmentalperipheral nerves;
d) evolution of a postanal tail; ande) origin and specialization of a branchial
basket with gill slits.
The last feature could have evolved onlyafter the evolution of the notochord whichreplaced the coelom as the axial supportand the specialization of the dorsal longi-tudinal muscles. The primitive chordatestock possessing these features would bean animal similar to the lancelet (acrania).
Vertebrates evolved from acrania byfurther specialization of the traits in thelatter group and the evolution of rigidskeletal structures.
Hemichordates evolved from acrania-like ancestors by specializing for bur-rowing in soft sediments. A peristalticstructure—the proboscis—evolved forpenetration and burrowing in mud. Thenotochord became reduced, as did thedorsal nerve cord, the coelom evolved toan oligomeric one with loss of transversesepta and the body muscles became elon-gated, nonsegmented longitudinal mus-cles. The pterobranchs became specializedfor life in closed tubes in which the bran-chial basket became reduced to a vestigeand replaced by tentacles evolved from theenteropneust collar.
Tunicates evolved from acrania-likeancestors by attaching to the substrate andabandoning active locomotion in the adult.These animals lost the notochord, most ofthe central nervous system, most of themusculature and the metameric conditionof the coelom. The muscular tail in larvaeis an adaptation for dispersal, but is de-rived from structures present in the acran-ia-like forerunners.
Echinoderms are considered to haveevolved from chordate ancestors from apterobranch-like stage. Intermediate formspossessed a branchial basket and tentacles,but the former were lost and the latter spe-cialized early in echinoderm evolution. Asuperficial layer of calcareous platesevolved for protection, and subsequentlybecame a rigid skeleton compensating forthe poor mechanical system of an oligo-meric coelom and only longitudinal mus-cles in the ancestor. Radial symmetryevolved in the early sessile echinoderms,and lastly the advanced echinoderms be-came free-living by turning over onto theoral surface and walking with the tentacles.
This concept of deuterostome phyloge-ny is very different from many others pre-sented earlier in the literature in whichgroups such as the tunicates (Berill, 1955),the hemichordates (Siewing, 1972; Re-mane, 1973) and the echinoderms (Jef-feries, 1975) are regarded as the primitivestock within the deuterostomes or thechordates rather than advanced members.Presentation of models or hypotheses fordeuterostome phylogeny is only part of theanalysis. More important is the develop-ment of methods by which the several the-ories can be tested and thereby falsified orfurther verified. The importance of thefunctional-adaptive analysis developed to-gether with studies of hydrostatic skeletonsis that it provides one way to test these con-flicting theories.
The functional study includes a mechan-ical analysis of complex sets of morpholog-ical structures which must be consistentwith the laws of physics (e.g., mechanics).The advantage of this approach is that atleast part of the test is based on laws out-side of biology and to that extent is inde-pendent. The adaptive and evolutionarysegment of the method is based on theconcept of gradual macroevolutionarymodification (e.g., Simpson, 1953; Bock,1979) in distinction to concepts of saltationas postulated by earlier workers (e.g.,Goldschmidt, 1940; Schindewolf, 1950). Itis necessary to postulate, in detail, the se-quence of change in features, to providean adaptive explanation for their evolu-tion, to show that the organisms at all in-
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

78 WOLFGANG FRIEDRICH GUTMANN
termediate stages are functional and adap-tive to their environments, and to postulatea series of environmental interactions thatcould be responsible for the adaptive evo-lutionary changes.
The demands required in the function-al-adaptive method of testing have beenmet for the hypothesis of deuterostomephylogeny proposed herein for the evo-lution of chordates from a metamericworm-shaped coelomate with the sequenceof evolutionary changes being the originof the notochord, myotomes of dorsal lon-gitudinal muscles, and myosepta, loss oftransverse and circular muscles, the dorsalnerve cord, the postanal tail and lastly thebranchial basket and gill slits. Morpholog-ical construction of the most primitivechordate is shown by the acrania. Subse-quent loss of most of the notochord anddorsal nerve cord, loss of metamerism inthe coelom and longitudinal muscles, andappearance of the proboscis occurred inthe hemichordates with their invasion ofsoft substrates and adaptation to life intubes open at each end. Appearance oftentacles from the enteropneust collar andeventual loss of the branchial basket tookplace in the evolution of pterobranchswhich left the soft substrate for a sessilelife in closed tubes. Similar changes oc-curred in the evolution of echinodermsfrom a hemichordate-like ancestor withthe addition of an armor of calcareousplates which became a rigid skeleton. Lossof the notochord, the locomotory muscu-lature and the dorsal nerve cord occurredin the evolution of tunicates as they be-came sessile animals attached to hard sur-faces. These features were retained as lar-val specializations in some tunicates andbecame reincorporated in the adult formof some derived, free-living groups of tu-nicates by neoteny.
Proponents of alternative hypothesesfor deuterostome or chordate phylogenyhave not attempted to test these ideas bymeans of a functional-adaptive analysis. Iwould like to outline each of these alter-native models giving attention only to thecritical evolutionary changes to be testedfunctionally and adaptively.
a) The tunicate model. The commonesttheory places the tunicates as the prim-itive group of chordates. The branchialbasket and gill slits are considered to bethe first chordate features to appear inan animal possessing an oligomeric c o ^lorn. Subsequently, the notochord, dor-sal nerve cord and segmented longitu-dinal muscles appeared as larvaladaptations for active swimming need-ed for dispersion. These features wereincorporated into the adult stage of de-scendent forms (acrania, vertebrates)by neoteny.
b) The hemichordate model. This theoryplaces the hemichordates (enterop-neusts) as the primitive group of chor-dates. The branchial basket and gill slitsare again considered to be the firstchordate features to appear in an ani-mal possessing an oligomeric coelomwith unsegmented longitudinal mus-cles. Subsequently the notochord, dor-sal nerve cord and metamerism, bothin the muscles and the coelom, evolvedas features for improved locomotion.
c) The echinoderm theory. This theoryconsideres the echinoderms to be theprimitive members of the deutero-stome coelomates. Again a branchialbasket and gill slits are believed to haveappeared first in an animal with anoligomeric coelom, possibly lackingeven longitudinal body muscles (as seenin the chordates), but having a rigidexoskeleton and associated muscula-ture attached to the skeletal plates. Theearliest group would be a branchoechi-noderm. Later the exoskeleton wouldbe lost and replaced by a hydroskeletonand body musculature, which wouldlater be segmented by transverse septasubdividing both the coelom and lon-gitudinal musculature. A dorsal nervechord evolved, presumably with themuscles and finally the notochord ap-peared and acquired the role of thebody support. Many of these points arenot mentioned by Jefferies (1975) butthey are a definite consequence of amodel for the evolution of chordatesfrom echinoderm ancestors.
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 79
Although these theories differ in theconclusion on the primitive group and inmany details on the phylogeny and the se-quence of evolution of features, they allinclude two postulated evolutionary steps
0rrhich cannot be explained by a functional-adaptive analysis within gradual macro-evolutionary change. These are:
a) The postulated origin of an oligomericcoelom as the primitive one and theevolution of a metameric coelom fromthe oligomeric condition with the evo-lution of dissepiments and segmenta-tion of the longitudinal muscles and
b) the evolution of a branchial basket withgill slits in the body wall in an animalpossessing a hydrostatic skeleton andprior to the evolution of the notochordand specialization of the dorsal longi-tudinal muscles.
These changes which are essential toeach of the alternate models mentionedabove could occur only by saltations. Thisis at variance with generally accepted evo-lutionary theory, including that acceptedby most proponents of these alternatemodels. It may be possible with futureknowledge to provide a gradual function-al-adaptive analysis for these changes withgreater knowledge about possible systemsof biological construction and relation-ships between these animals and their en-vironments. However, the organisms arestill subject to the laws of mechanics whichare not likely to change radically. At thispoint in our knowledge of the functionalmorphology of the deuterostomes, I canonly conclude that a vanishingly smallprobability exists for the occurrence of thecritical evolutionary changes, gradualisticones, in these alternate models for thephylogeny of the deuterostomes, andhence I would conclude these models havebeen falsified strongly.
Special mention should be made of theneoteny idea for the origin of higher chor-dates from ancestral tunicates because thismodel has been presented most persua-sively (Garstang, 1928; Berill, 1955; Bone,1960) and has been widely accepted. No
question exists that neoteny has occurredin the evolution of several groups of free-swimming tunicates which have lost thesessile adult stage in their life history. Andit is entirely possible that existing chordategroups, such as the vertebrates, evolvedfrom tunicate ancestors by neoteny. Theproblem is how the chordate charactersevolved in the evolution of the tunicates.
A hypothesis for the appearance of newfeatures as larval specializations and theirsubsequent incorporation into the adultstage of descendent groups by neoteny.does not eliminate the requirement to pro-vide a functional-adaptive analysis forthese changes. Not only has this neverbeen attempted for the tunicate neotenyhypothesis for higher chordates, but thismodel does not rest on solid morphologi-cal description of the tunicate larvae.
The larval tail of tunicates is a connec-tive tissue sheath filled with cells and fluid,and constitutes a hydraulic apparatus. Thefew muscle cells are not arranged segmen-tally, but are attached directly to the con-nective tissue sheath and generate thrustby bending the axial support. The hydrau-lic organ and longitudinal muscles requiredefinite preconditions for their evolution-ary origin as discussed above. These pre-conditions have not been mentioned byadvocates of the neotenic tunicate modeland it appears unlikely that they could ex-ist.
The neotenic tunicate model suffers,moreover, from the problem of the originof the branchial basket in an animal witha hydrostatic skeleton and need not bementioned again here.
Evolution of a tail in a larva, includingall the specialized features of the tail lo-comotory system of tunicates, appearshighly improbable. The use of a tail as apropulsory organ depends on a certaindegree of perfection and the attainment ofa minimal length. A functional explana-tion, other than locomotion, would have tobe postulated for the adaptive significanceof the incipient stages in the de novo evo-lution of a propulsive larval tail. Such ex-planations have not been offered, and rea-sonable nonlocomotory arguments for the
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

80 WOLFGANG FRIEDRICH GUTMANN
de novo origin of the tunicate tail are notapparent.
The neotenic tunicate larval model failsto mention that not all sessile tunicateshave a tailed larva, but some have one thatswims by cilia covering the body. Are theseprimitive forms advanced degenerateforms or ones on a side branch? Moreoverthe tailed larva in sessile tunicates have ashort existence, just long enough to dis-perse and locate a suitable site for attach-ment. It does not feed in all species pos-sessing a tailed larva. This model mustpostulate two steps, each with adaptive ex-planations. First, is the evolution of feed-ing in the tailed larva, and the second isneoteny with elimination of the sessileadult form.
The tunicate tail is, indeed, a specializedlarval adaptation, but it is dependent onall of the essential structures being presentalready in the ancestors of the tunicates. Itis not necessary to postulate the de novoappearance of any features in the evolu-tion of the specialized larvae as they simplyretained features present in the adult of itsancestors.
In conclusion, if the entire set of chor-date features is present in the ancestorsof the tunicates, then it is equally reason-able to postulate that the higher chordates(vertebrates) evolved directly from anacrania-like ancestor than from a special-ized tunicate larva through neoteny.
The conclusion to be drawn from thiscase study of deuterostome phylogeny isthat a functional-adaptive analysis is an es-sential part of the testing of phylogeneticmodels. This approach provides a strongefficient method for the falsification andrejection of alternate models. Such a meth-od had been lacking in earlier discussionsof the validity of diverse models of inver-tebrate phylogeny. It is my belief that withthe application of proper functional-adap-tive analysis our understanding of inver-tebrate phylogeny will improve rapidly.
ACKNOWLEDGMENTS
The illustrations were skilfully executedby Mrs. Renate Klein-Rodder to whom Iam very grateful. The English manuscriptwas virtually rewritten bv Professor Bock.
In addition to this time and energy con-suming help I owe to him a careful checkof the arguments and valuable advice forthe presentation of the phylogenetic re-construction.
REFERENCES *
Berrill, N.J. 1955. The origin of vertebrates. ClarendonPress, Oxford.
Bock, W. 1979. The synthetic explanation of macro-evolutionary change—a reductionistic approach.Bull. Carnegie Mus. Nat. Hist. 13:20-69.
Bock, W. 1981. Functional-adaptive analysis in evo-lutionary classification. Amer. Zool. 21:5-20.
Bone, Q. 1960. The origin of chordates. ]. Linn.Soc. Zool. 44:252-269.
Bonik, K. 1977/1978. Quantitative Aspekte hydrau-lischer Systeme in Metazoen-Konstruktionen. ICourier Forschungs-Institut Senckenberg 23:1 —79. II Courier Forschungs-Institut Senckenberg32:1-57.
Bonik, K., M. Grasshoff, and W. F. Gutmann. 1977a.Funktion bestimmt Evolution. Umschau Natur-wissenschaft Technik 77:657-668.
Bonik, K. and W. F. Gutmann. 1977. Die Biotechnikder Doppelhydraulik (Chorda-Sklerocoelen-Myomeren-System) bei den Acraniern. Sencken-bergiana Biol. 58:275-286.
Bonik, K., W. F. Gutmann, M. Grasshoff, and R.Haude. 1978. Stachelhauter mit Kiemen-Ap-parat: Der Beleg fur die Ableitung der Echino-dermen von Chordatieren. Natur u. Museum108:211-214.
Bonik, K., W. F. Gutmann, and D. S. Peters. 19776.Optimierung und Okonomisierung im Kontextvon Evolutionstheorie und phylogenetischer Re-konstruktion. Acta Biotheor. 26:75-119.
Chapman, E. 1958. The hydrostatic skeleton in theinvertebrates. Biol. Rev. Soc. Cambridge 33:339-357.
Clark, R. B. 1964. Dynamics in metazoan evolution.Clarendon Press, Oxford.
Clark, R. B. 1979. Radiation of the Metazoa. In M.R. House (ed.), The origin of major invertebrategroups. The Systematic Assoc, Special Vol. 12,pp. 55-102. Academic Press, London, New York,San Francisco.
Conway Morris, S. and H. B. Whittington. 1979.The animals of the Burgess Shale. Sci. Amer.241:110-121.
Dullemeijer, P. 1974. Concepts and approaches in ani-mal morphology. Van Gercum Co., Assen.
Garstang, W. 1928. The morphology of the Tuni-cata and its bearing on the phylogeny of theChordata. Quarterl. J. Microsc. Sci. 72:51-187.
Goldschmidt, R. 1940. The material basis of evolution.Yale Univ. Press, New Haven.
Grobbcn, K. 1924. Theoretische Erorterungen be-treffend die phylogenetische Ableitung der Echi-nodermen. Sitzber. Akad. VViss. VVien (Math.-Naturw. Kl.) 132:262-290.
Gutmann, W. F. 1960. Funktionelle Morphologie vonBalanus balanaides. Abhandlungen Senckenberg.
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

PHYLOGENY OF DEUTEROSTOME COELOMATES 81
Naturf. Ges. 500. Woldemar Kramer, Frankfurtam Main.
Gutmann, W. F. 1966. Coelomgliederung, Myo-merie und die Frage der Vertebraten-Antezed-enten. Z. Zool. Syst. Evolutionsf. 4:13-57.
Gutmann, W. F. 1967. Die Entstehung des Coeloms9 und seine phylogenetische Abwandlung im Deu-
terostomier-Stamm. Zool. Anz. 179:109-131.Gutmann, W. F. 1969. Acranier und Hemichorda-
ten, ein Seitenast der Chordaten. Zool. Anz.182:1-26.
Gutmann, W. F. 1971. DerbiomechanischeGehaltderWurmtheorie. Z. Wiss. Zool. 182:229-262.
Gutmann, W. F. 1972. Die Hydroskelett-Theorie.Aufsatze u. Reden Senckenberg. Naturf. Ges. 21.Woldemar Kramer, Frankfurt am Main.
Gutmann, W. F. 1973. Diskussions-Beitrag zur Coe-lom-Problematik. In W. Schafer (ed.), Das Archi-coelomaten-Problem. Aufsatze u. Reden Senckenb.Naturf. Ges. 22:51-101.
Gutmann, W. F. 1975. Das Tunicaten-Modell. Zool.Beitr. (N.F.) 21:279-303.
Gutmann, W. F. 1976. Aspekte einer konstruktivis-tischen Phylogenetik: Postulate und ein Exem-pel. In W. Schafer (ed.), Evoluierende Systeme Iund II. Aufsatze u. Reden Senckenb. Naturf. Ges.28:165-183.
Gutmann, W. F. 1977. Phylogenetic reconstruction:Theory, methodology and application. In M.Hecht, P. Goody, and B. Hecht (eds.), Major pat-terns in vertebrate evolution, pp. 645-670. PlenumPress, New York, London.
Gutmann, W. F. and K. Bonik. 1979. Detaillierungdes Acranier- und Enteropneusten-Modells.Senckenbergiana Biol. 59:325-363.
Gutmann, W. F., K. Vogel, and H. Zorn. 1978. Bra-chiopods. Biomechanical interdependences gov-erning their origin and phylogeny. Science199:890-893.
Jefferies, R. P. S. 1975. Fossil evidence concerningthe origin of the chordates. Symp. Zool. Soc.London 36:253-318.
Mettam, C. 1971. Functional design and evolutionof the polychaete Aphrodite aculeata. J. Zool.,London 153:245-267.
Peters, D. S. and W. F. Gutmann. 1971. Ober dieLesrichtung von Merkmals- und Konstruktions-reihen. Z. Zool. Syst. Evolutionsf. 9:237-263.
Reisinger, E. 1973. SchluBwort. In W. Schafer (ed.),Das Archicoelomatenproblem. Aufs. RedenSenckenberg. Naturf. Ges. 22:109-111. Wolde-mar Kramer, Frankfurt am Main.
Remane, A. 1973. Stellungnahme. In W. Schafer(ed.), Das Archicoelomaten-Problem. Aufs. RedenSenckenberg. Naturf. Ges. 22:105-108. Wolde-mar Kramer, Frankfurt am Main.
Schafer, W. (ed.) 1973. Phylogenetische Rekon-struktionen—Theorie und Praxis. Aufsatze u.Reden Senckenb. Naturf. Ges. 24. WoldemarKramer, Frankfurt am Main.
Schafer, W. (ed.) 1975. Ontogenetische und kon-struktive Gesichtspunkte bei phylogenetischenRekonstruktionen. Aufsatze u. Reden Senckenb.Naturf. Ges. 27. Woldemar Kramer, Frankfurtam Main.
Schafer, W. (ed.) 1976. Evoluierende Systeme 1 undII. Aufsatze u. Reden Senckenb. Naturf. Ges. 28.Woldemar Kramer, Frankfurt am Main.
Schafer, W. (ed.) 1978. Evoluierende Systeme III.Aufsatze u. Reden Senckenb. Naturf. Ges. 29.Woldemar Kramer, Frankfurt am Main.
Schindewolf, O. H. 1950. Grundfragen der Paldonto-logie. E. Schweizerbart, Stuttgart.
Siewing, R. 1972. Zur Deszendenz der Chordaten—Erwiderung und Versuch einer Geschichte derArchicoelomaten. Z. Zool. System. Evolutions-forsch. 10:267-291.
Simpson, G. G. 1953. The major features of evolution.Columbia University Press.
Trueman, E. R. 1975. The locomotion of soft-bodied an-imals. Edward Arnold, London.
Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022

Dow
nloaded from https://academ
ic.oup.com/icb/article/21/1/63/134438 by guest on 02 June 2022
Related Documents











