Ecological Engineering 49 (2012) 65–72 Contents lists available at SciVerse ScienceDirect Ecological Engineering j ourna l ho me page: www.elsevier.com/locate/ecoleng Relationship between various physiological and biochemical parameters activated by cadmium in Sinapis alba L. and Hordeum vulgare L. Marianna Molnárová ∗ , Agáta Fargaˇ sová 1 Department of Ecosozology and Physiotactics, Faculty of Natural Sciences, Comenius University in Bratislava, Mlynská dolina B2, SK-842 15 Bratislava, Slovak Republic a r t i c l e i n f o Article history: Received 26 April 2012 Received in revised form 12 July 2012 Accepted 10 August 2012 Available online 28 September 2012 Keywords: Cadmium phytotoxicity Regression analyses Photosynthetic pigments content Cadmium accumulation Oxidative stress parameters Dry and fresh mass a b s t r a c t The aim of this study was to examine the phytotoxicity of cadmium (Cd) on commonly measured physiological parameters [root and shoot growth inhibition, dry mass (DM) and fresh mass (FM) pro- duction, water content (WC), Cd accumulation] and biochemical parameters [photosynthetic pigments (chlorophyll a, b, total carotenoids), proteins, thiobarbituric acid reactive substances (TBARS), and protein sulfhydryl group content] using regression analyses. Biochemical parameters were evaluated to estimate oxidative stress in seedlings. For all parameters, the sensitivity of Hordeum vulgare L. and Sinapis alba L. plants was studied as representative of widely cultivated crop plants. Cd showed a greater inhibitory effect on root than shoot growth, predominantly in S. alba. In addition to growth inhibition, Cd reduced biomass production (FM, DM), mainly in the shoots. Although Cd was accumulated chiefly in the roots, the accumulation of Cd in S. alba shoots exceeded that in H. vulgare. While Cd resulted a greater reduction of chlorophyll b than chlorophyll a, carotenoids content in S. alba increased when metal concentration reached 71 and 143 mg Cd l −1 . The adverse effect of Cd was also confirmed by a significant reduction in protein sulfhydryl groups. In a regression analysis, positive correlations were observed between proteins and sulfhydryl groups in shoots of S. alba and TBARS and sulfhydryl groups in H. vulgare, while negative correlations were found between Cd concentrations in S. alba roots and proteins, TBARS, and WC contents. Cd concentration in H. vulgare roots was also negatively correlated with TBARS and shoot length. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Although the environmental impact assessment of cadmium (Cd) has gained increasing attention (e.g. Chaturvedi, 2004; Heged ˝ usová et al., 2006; Hiller et al., 2010; Kalin et al., 2006), the mechanisms of its toxicity are currently not completely under- stood (Benavides et al., 2005). Recent laboratory research has paid preferential attention to understanding the mechanism of metal tolerance/toxicity. Many publications deal with only Cd toxicity and omit to estimate relations between physiological, production and biochemical parameters using regression analyses. Anthropogenic sources are the most common cause for the widespread Cd contamination of soil, releasing an estimated 30 000 tons per year (Sanitá di Toppi and Gabbrielli, 1999). Except for the Cd-carbonic anhydrase of marine diatoms, Cd is not an essen- tial element for plants (Lane and Morel, 2000). Its absorption by roots and also leaves (Hendry et al., 1992) results in root and shoot ∗ Corresponding author. Tel.: +421 2 602 96 575; fax: +421 2 602 96 704. E-mail addresses: [email protected] (M. Molnárová), [email protected] (A. Fargaˇ sová). 1 Tel.: +421 2 602 96 575; fax: +421 2 602 96 704. growth, water balance, nutrient uptake and homeostasis, and net photosynthesis inhibition coupled with changes in chloroplasts structure and chlorophyll content depression (Gallego et al., 1996). Moreover, chlorophyll content is an indicator of critical levels of Cd in plants (Burton et al., 1986). All these parameters may be used as Cd phytotoxicity indicators (Drazic and Mihailovic, 2005; Sanitá di Toppi and Gabbrielli, 1999; Wang and Zhou, 2005). Roucoux and Dabin (1977), as well as Sharma and Dietz (2006), observed that Cd specifically induced sulfhydrylated aminoacid (cysteine and methione) synthesis. The most important bio- chemical characteristic of Cd ions is their strong affinity to sulfhydryl groups of several compounds (e.g. Cd complexes with metallothionein-like protein). In the presence of Cd, toxic effects were observed very early and oxidative stress was enhanced (Stohs and Bagchi, 1995). During stress and H 2 O 2 production, the presence of Cd resulted in a decrease of chlorophyll and GSH content and an increase of superoxide radical production and lipoperoxidation (Gallego et al., 1996; Maksymiec and Krupa, 2006). Moreover, Cd frequently accumulates in important crops consumed by humans and animals, and, in addition to the associated health risks, may also have an economic impact associated with decreased crop yield. Consequently, we directed our attention in this study to evalu- ating the effect of Cd on plant seedlings from two aspects: (1) 0925-8574/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.ecoleng.2012.08.025

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ra
MD
a
ARRAA
KCRPCOD
1
(Htsptaa
wtttr
f
0h
Ecological Engineering 49 (2012) 65– 72
Contents lists available at SciVerse ScienceDirect
Ecological Engineering
j ourna l ho me page: www.elsev ier .com/ locate /eco leng
elationship between various physiological and biochemical parametersctivated by cadmium in Sinapis alba L. and Hordeum vulgare L.
arianna Molnárová ∗, Agáta Fargasová1
epartment of Ecosozology and Physiotactics, Faculty of Natural Sciences, Comenius University in Bratislava, Mlynská dolina B2, SK-842 15 Bratislava, Slovak Republic
r t i c l e i n f o
rticle history:eceived 26 April 2012eceived in revised form 12 July 2012ccepted 10 August 2012vailable online 28 September 2012
eywords:admium phytotoxicityegression analyseshotosynthetic pigments contentadmium accumulation
a b s t r a c t
The aim of this study was to examine the phytotoxicity of cadmium (Cd) on commonly measuredphysiological parameters [root and shoot growth inhibition, dry mass (DM) and fresh mass (FM) pro-duction, water content (WC), Cd accumulation] and biochemical parameters [photosynthetic pigments(chlorophyll a, b, total carotenoids), proteins, thiobarbituric acid reactive substances (TBARS), and proteinsulfhydryl group content] using regression analyses. Biochemical parameters were evaluated to estimateoxidative stress in seedlings. For all parameters, the sensitivity of Hordeum vulgare L. and Sinapis albaL. plants was studied as representative of widely cultivated crop plants. Cd showed a greater inhibitoryeffect on root than shoot growth, predominantly in S. alba. In addition to growth inhibition, Cd reducedbiomass production (FM, DM), mainly in the shoots. Although Cd was accumulated chiefly in the roots,the accumulation of Cd in S. alba shoots exceeded that in H. vulgare. While Cd resulted a greater reduction
xidative stress parametersry and fresh mass
of chlorophyll b than chlorophyll a, carotenoids content in S. alba increased when metal concentrationreached 71 and 143 mg Cd l−1. The adverse effect of Cd was also confirmed by a significant reduction inprotein sulfhydryl groups. In a regression analysis, positive correlations were observed between proteinsand sulfhydryl groups in shoots of S. alba and TBARS and sulfhydryl groups in H. vulgare, while negativecorrelations were found between Cd concentrations in S. alba roots and proteins, TBARS, and WC contents.Cd concentration in H. vulgare roots was also negatively correlated with TBARS and shoot length.
gpsMiCT
o(csmw
. Introduction
Although the environmental impact assessment of cadmiumCd) has gained increasing attention (e.g. Chaturvedi, 2004;egedusová et al., 2006; Hiller et al., 2010; Kalin et al., 2006),
he mechanisms of its toxicity are currently not completely under-tood (Benavides et al., 2005). Recent laboratory research has paidreferential attention to understanding the mechanism of metalolerance/toxicity. Many publications deal with only Cd toxicitynd omit to estimate relations between physiological, productionnd biochemical parameters using regression analyses.
Anthropogenic sources are the most common cause for theidespread Cd contamination of soil, releasing an estimated 30 000
ons per year (Sanitá di Toppi and Gabbrielli, 1999). Except for
he Cd-carbonic anhydrase of marine diatoms, Cd is not an essen-ial element for plants (Lane and Morel, 2000). Its absorption byoots and also leaves (Hendry et al., 1992) results in root and shoot∗ Corresponding author. Tel.: +421 2 602 96 575; fax: +421 2 602 96 704.E-mail addresses: [email protected] (M. Molnárová),
[email protected] (A. Fargasová).1 Tel.: +421 2 602 96 575; fax: +421 2 602 96 704.
aoa(faaCa
925-8574/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.ecoleng.2012.08.025
© 2012 Elsevier B.V. All rights reserved.
rowth, water balance, nutrient uptake and homeostasis, and nethotosynthesis inhibition coupled with changes in chloroplaststructure and chlorophyll content depression (Gallego et al., 1996).oreover, chlorophyll content is an indicator of critical levels of Cd
n plants (Burton et al., 1986). All these parameters may be used asd phytotoxicity indicators (Drazic and Mihailovic, 2005; Sanitá dioppi and Gabbrielli, 1999; Wang and Zhou, 2005).
Roucoux and Dabin (1977), as well as Sharma and Dietz (2006),bserved that Cd specifically induced sulfhydrylated aminoacidcysteine and methione) synthesis. The most important bio-hemical characteristic of Cd ions is their strong affinity toulfhydryl groups of several compounds (e.g. Cd complexes withetallothionein-like protein). In the presence of Cd, toxic effectsere observed very early and oxidative stress was enhanced (Stohs
nd Bagchi, 1995). During stress and H2O2 production, the presencef Cd resulted in a decrease of chlorophyll and GSH content andn increase of superoxide radical production and lipoperoxidationGallego et al., 1996; Maksymiec and Krupa, 2006). Moreover, Cdrequently accumulates in important crops consumed by humans
nd animals, and, in addition to the associated health risks, maylso have an economic impact associated with decreased crop yield.onsequently, we directed our attention in this study to evalu-ting the effect of Cd on plant seedlings from two aspects: (1)
6 cologi
pdbeabtr
2
2
b(w
2
2
twp(sf43TTpnaa8
wl12s
2
smcd
W
W
2
tm1iCtTt
a0iw
2
2
d(cAwsl
c
c
c
cc
2g
ped
t0wlrbc
(twhica1
1(sr4w
2
6 M. Molnárová, A. Fargasová / E
hysiological and production (roots and shoots growth, fresh andry biomass production, water content, Cd accumulation); and (2)iochemical (leaf chlorophyll and carotenoids content, protein lev-ls, TBARS, and protein sulfhydryls groups). Two widely growinggricultural plants, white mustard (Sinapis alba L.) and commonarley (Hordeum vulgare L.), were used in our studies and the rela-ionships among the observed parameters were evaluated usingegression analyses.
. Materials and methods
.1. Plant materials and chemicals
Seeds of white mustard (S. alba L., cv. Severka) and commonarley (H. vulgare L., cv. Orthega) were achieved from Chepo, Ltd.Unhost’ – Fialka, Czech Republic). Analytical grade CdCl2·2.5H2Oas obtained from Lachema, Brno, Czech Republic.
.2. Physiological and production parameters
.2.1. Growth inhibition testsFor seed cultivation, 21 cm × 15.5 cm vertical cultivation con-
ainers (Phytotoxkit, MicroBioTests Inc., Nazareth, Belgium) linedith cellulose and filter paper soaked with 24 ml of freshly pre-ared Cd solutions were used (OECD, 2006). For dicotyledonaeS. alba) and monocotyledonae (H. vulgare) plants, 15 and 8eeds per container, respectively, were used. The IC50 valuesrom more than four different Cd concentrations ranging from9 to 219 mg l−1 (438–1949 �mol dm−3) for S. alba and 94 to94 mg l−1 (832–3504 �mol dm−3) for H. vulgare were estimated.he pH of the applied Cd solutions was between 6.35 and 7.51.ap water (72.6 mg l−1 Ca, 17.7 mg l−1 Mg; free Cl < 0.02 mg l−1;H = 7.06 ± 0.05) was used as the control. For IC50 value determi-ation, at least 48 and 90 seeds were used for monocotyledonaend dicotyledonae plants, respectively. Containers were placed in
dark temperature-controlled chamber (t = 25 ± 1 ◦C; air humidity0%) and root and shoot lengths were measured after 72 h.
After root and shoot lengths measuring, the containersere placed vertically in the laboratory with natural day-
ight [photosynthetic photon-flux density (PPFD) = approximately00 �mol m−2 s−1] with 12 h photoperiod and a temperature of3 ± 1 ◦C. The containers were shielded from direct sunlight andeeds were left to cultive for next four days.
.2.2. Biomass production and water content determinationAfter seven days (3 + 4), plants were divided into roots and
hoots and the fresh mass was immediately weighed. The plantaterial was then oven-dried (55 ◦C) to constant weight. The water
ontent of the plants was determined on the basis of the fresh andry mass as follows (Drazic and Mihailovic, 2005):
C = FM − DMDM
(g g−1 DM)
C = water content, FM = fresh mass, and DM = dry mass.
.2.3. Accumulation of cadmium in roots and shootsAccumulation of Cd was determined after seven days’ cul-
ivation. A minimum of 11 mg of root or shoot dry mass wasineralized in 5 ml of HNO3:H2O2 mixture (4:1) for 60 min at
80 ◦C in a ZA-1 autoclave (Czech Republic). After cooling, mineral-zed samples were diluted up to 25 ml with distilled water, and the
d content was determined by galvanostatic dissolved chronopo-enciometry on an EcaFlow 150 GLP (Istran, Slovak Republic).his electrochemical method is comparable to the atomic absorp-ion spectrophotometric (AAS) method in precision, accuracy,wdp
cal Engineering 49 (2012) 65– 72
nd sensitivity of the measured results with concentration range.5–1000 �g Cd dm−3 and reproducibility 1.5% at 50 �g dm−3 metal
on in the measured solution. Three samples of each concentrationere determined.
.3. Biochemical characteristics
.3.1. Photosynthetic pigment determinationThe pigment contents (chlorophyll a, b, total carotenoids) were
etermined in fresh leaves after extraction in 95% ethanol (w/v)60 mg of fresh leaves per 6 ml of ethanol). Pigment extractionontinued until all of the homogenized plant mass was white.fter brief centrifugation (2 min at 2900 × g), the pigment contentas measured spectrophotometrically at 665, 649, and 470 nm in
upernatant. The pigment contents were calculated using the fol-owing equations (Lichtenthaler and Wellburn, 1983):
hl a = 13.95 (A665) − 6.88 (A649)
hl b = 24.96 (A649) − 7.32 (A665)
ar = 1000 (A470) − 2.05 (chl a) − 114.8 (chl b)245
hl a = chlorophyll a, chl b = chlorophyll b, and car = totalarotenoids; in �g mg−1 DM; DM = dry mass.
.3.2. Determination of proteins, TBARS, and protein sulfhydrylroups
Proteins, thiobarbituric acid reactive substances (TBARS), androtein sulfhydryl groups were determined in fresh leaf mass afterthanol extraction as described above for photosynthetic pigmentetermination.
For determination of protein concentration, the Bradford spec-rophotometric method was used (Bradford, 1976), in which 1 ml of.01% (w/v) Coomassie Brilliant Blue G-250 [4.7% (w/v) of ethanolith 8.5% (w/v) of H3PO4] was added to 0.1 ml of 1% (w/v) of fresh
eaf homogenate and then vortexed. After 5 min of incubation atoom temperature, the sample was measured at 595 nm against alank of 95% ethanol. Bovine serum albumin was used as a standardalibration curve.
TBARS were measured as an indicator of lipoperoxidationGallego et al., 1996): 1 ml of solution containing 20% ofrichloroacetic acid and 0.5% (w/v) thiobarbituric acid was mixedith 2 ml of 1% (w/v) fresh leaf homogenate. Then 0.1 ml of butyl-ydroxytoluene (4% in ethanol) was added, briefly vortexed, and
ncubated for 30 min at 90 ◦C in a water bath. Samples wereooled in water with ice and spectrophotometrically measuredt 532 nm against destilated water. An extinction coefficient of55 (mmol dm−3) −1 cm−1 was used.
Protein sulfhydryl groups were estimated as follows (Viner et al.,996): 0.1 ml of 20 mmol dm−3 5,5′-dithiobis-(2-nitrobenzoic acid)DTNB) was added to 0.5 ml of 1% (w/v) fresh leaf homogenate. Theample was briefly vortexed, incubated for 45 min at laboratoryoom temperature and then spectrophotometrically measured at12 nm against 95% ethanol. The extinction coefficient for DTNBas 1.36 × 104 (mol dm−3)−1 cm−1.
All three parameters were calculated per 1 mg of dry mass (DM).
.4. Statistical analysis
All phytotoxic tests were carried out in six parallels. Resultsere evaluated by determining IC50 values (concentrations pro-ucing 50% inhibition) and their 95% confidence intervals (CI) byrobit analysis or as average values with their standard deviations

M. Molnárová, A. Fargasová / Ecologic
Table 1IC50 values and their 95% confidence intervals (CI) for roots and shoots growth inhi-bition of S. alba and H. vulgare after 3 days exposure to Cd in the dark (IC – inhibitoryconcentration).
Species Roots Shoots−1 −1
(dmt*f
3
3
3
gaahI2S
3
i
iisiibfitp
3
irrdCwftr
3
3
ert
Fo
F(
IC50 (mg l ) (95% CI) IC50 (mg l ) (95% CI)
Sinapis alba 127.6 (111.7–145.7) 207.0 (175.2–244.5)Hordeum vulgare 215.5 (179.9–257.4) 218.2 (193.7–245.9)
SD), and were plotted with Microsoft Excel 2003 software. Theata were subjected to analysis of variance (ANOVA) and treat-ent means were compared by Scheffe’s multiple-range test using
he following levels of statistical significance: *p < 0.05; **p < 0.01;**p < 0.001. Regression analyses and ANOVA were calculated withormulas inserted in Microsoft Excel.
. Results
.1. Physiological and production parameters
.1.1. Plant growth inhibitionThe deleterious effects of Cd were expressed as root and shoot
rowth inhibition and evaluated by probit analysis as IC50 valuesnd their 95% confidence intervals (CI) (Table 1). The roots of S.lba seedlings were more sensitive to Cd than those of H. vulgare;owever, sensitivity of shoots was nearly the same for both species.
C50 values for S. alba and H. vulgare shoot growth were 207.0 and18.2 mg l−1, respectively and this suggests that the dicotyledonae. alba is more sensitive to Cd than the monocotyledonae H. vulgare.
.1.2. Biomass production and water content in roots and shootsA major precursor for increasing plant yield is an increase
n biomass production in terms of dry weight mass. Our results
tita
ig. 1. Roots and shoots fresh (FM) and dry mass (DM) production in S. alba and H. vulgareo p < 0.01, ooo p < 0.001; ### p < 0.001;* for FM-root, o for FM-shoot and # for DM-root).
ig. 2. Water contents (WC) in the roots and shoots of S. alba and H. vulgare with thei**p < 0.01, ***p < 0.001; analogically oo p < 0.01, ooo p < 0.001; * for root, o for shoot).
al Engineering 49 (2012) 65– 72 67
ndicate that Cd dramatically reduced both FM and DM productionn plant roots and shoots (Fig. 1). The only exception was root andhoot FM production in S. alba at concentrations of 71 mg Cd l−1,n which we did not find any significant differences in compar-son with the control. Cd reduced water content in the roots ofoth plant species (Fig. 2); however, the reduction was only con-rmed in the shoots of S. alba. In H. vulgare, the water content inhe shoots increased and surpassed the water content in controllants by about 27%.
.1.3. Cadmium accumulation in roots and shootsCd accumulation in both plants was higher in the roots than
n the shoots (Fig. 3). While Cd concentration in S. alba shootsose with its remaining concentration in the solution, the oppositeesults were found for H. vulgare. However, in this monocotyle-onae plant Cd content in the roots increased in accordance withd concentration in the solution, its accumulation in the shootsas reversed. That indicate that while for S. alba Cd translocation
rom roots to shoots depends on the Cd concentration in the solu-ion, its translocation from roots to shoots in H. vulgare is intact andeduced with increasing Cd concentration in the solution (Fig. 3).
.2. Biochemical characteristics
.2.1. Photosynthetic pigmentsPhotosynthetic pigment contents in the shoots of plants
xposed to different Cd concentrations are shown in Fig. 4. Theesults suggest that chlorophyll b content was reduced more thanhat of chlorophyll a in both plant species. All tested Cd concen-
rations reduced chlorophyll a and b content only slightly, mainlyn H. vulgare, and their contents did not fall below 50% of the con-rol. An exception was observed for chlorophyll b content in S. albat concentrations of 219 mg Cd l−1. Carotenoids, which were lessafter 7 days exposure to Cd (*p < 0.05, **p < 0.01, ***p < 0.001; analogically o p < 0.05;
r standard deviation (SD) and polynomic trend lines after 7 days exposure to Cd

68 M. Molnárová, A. Fargasová / Ecological Engineering 49 (2012) 65– 72
Fig. 3. Accumulation of Cd in the roots and shoots of S. alba and H. vulgare after 7 days exposure to different Cd concentrations. Average values with their standard deviations(SD) are plotted.
F S. alb( *p < 0c
sric
3s
Tifrsdm
3
r
spmceclbapwTidgr
TAe
ig. 4. Photosynthetic pigment content after 7 days exposure to Cd in shoots of
chl a = chlorophyll a, chl b = chlorophyll b, car = total carotenoids, DM = dry mass;arotenoids).
ensitive to Cd than other pigments in both plant species, wereeduced only at the highest Cd concentration (Fig. 4). The resultsndicate considerable differences in the effect of Cd on pigmentsontent in S. alba and H. vulgare, especially for chlorophylls.
.2.2. Parameters of oxidative stress (TBARS and proteinulfhydryl groups)
Cd decreased protein content in both plant species (Table 2).he adverse effect of Cd was stronger for S. alba, when a reductionn protein content reached 34–53%. The same trend was foundor concentrations of protein sulfhydryl groups (Table 2). Theseesults reconfirm the greater sensitivity of S. alba to Cd. Although Cdlightly reduced TBARS concentration in both plants, no significantifferences to control were confirmed. Reduction reached maxi-um 18 and 23% for S. alba and H. vulgare, respectively (Table 2).
.3. Regression analyses
Advantage of regression analysis is in their capability toeveal reciprocal relations among two studied parameters under
gwa(
able 2verage concentrations of proteins, proteins’ sulfhydryl groups ( SH) and thiobarbituricxposure to Cd (SD – standard deviation).
Species Proteins
c (mg Cd l−1) c ± SD (�g mg−1 DM)
Sinapis alba0 70.1 ± 41.1
71 46.3 ± 25.0
143 37.8 ± 26.0
219 32.8 ± 23.9
Hordeum vulgare0 119.3 ± 16.1
94 101.4 ± 14.4
187 113.4 ± 20.7
281 91.9 ± 10.3**
* p < 0.05.** p < 0.01.
a and H. vulgare. Average values with their standard deviations (SD) are plotted.05, **p < 0.01; analogically oo p < 0.01, ooo p < 0.001; * for chlorophyll a, o for total
ame conditions (e.g. same cadmium concentrations for botharameters) which is common for selected parameters and esti-ate the significance (relevance) of this relationship. Negative
orrelation means that with increase of one selected param-ter the second one is decrease in relation to changed Cdoncentration. Equal findings should be done for positive corre-ation when with progressive cadmium concentration increasesoth parameters. Regression analysis showed a significant neg-tive correlation between Cd concentration in the roots androtein or TBARS content in S. alba (Fig. 5). Similar resultsere found for H. vulgare between Cd root concentration and
BARS content, shoot length, and DM (Fig. 6). The remain-ng Cd concentration in the shoots of S. alba significantlyecreased water content, chlorophyll a, and proteins sulfhydrylroups level or shoot length (Fig. 5). A significant positive cor-elation was found between proteins and proteins sulfhydryl
roups of S. alba (Fig. 5). In H. vulgare, positive correlationsere confirmed for carotenoids and chlorophyll a, proteinsnd chlorophyll b, and TBARS and proteins sulfhydryl groupsFig. 6).
acid reactive substances (TBARS) in the shoots of S. alba and H. vulgare after 7 days
SH groups TBARSc ± SD (nmol mg−1 DM) c ± SD (mmol mg−1 DM)
56.6 ± 23.8 2.98 ± 1.4245.2 ± 17.5 2.53 ± 1.3738.4 ± 21.9 2.59 ± 0.6527.7 ± 10.6* 2.47 ± 0.47
89.6 ± 31.3 1.25 ± 0.0977.9 ± 13.7 1.17 ± 0.3377.4 ± 24.5 1.13 ± 0.2660.7 ± 16.1* 0.97 ± 0.27

M. Molnárová, A. Fargasová / Ecological Engineering 49 (2012) 65– 72 69
Pears
4
mHh
eo
Fig. 5. Regression analyses for selected parameters of S. alba, where r is
. Discussion
Cadmium is one of the most widely studied nonessential toxicetals/metalloids which is always found in association with zinc.owever, zinc is essential micronutrient in living cells, cadmiumas not known useful biological function (Vymazal, 1987). Tosner
poie
on’s coefficient and p is statistical significance of Pearson’s coefficient.
t al. (2012) observed that type of soil also influenced the resultsf the ecotoxicity tests with seeds of Lactuca sativa. Cd uptake in
lant cells (in the form of Cd2+, the most common oxidative state)ccurs via Zn2+, Fe2+, and Ca2+ transporters (Clemens, 2006) andts ions compete for uptake by the same transmembrane carri-rs as various nutrients, such as calcium (Ca), magnesium (Mg),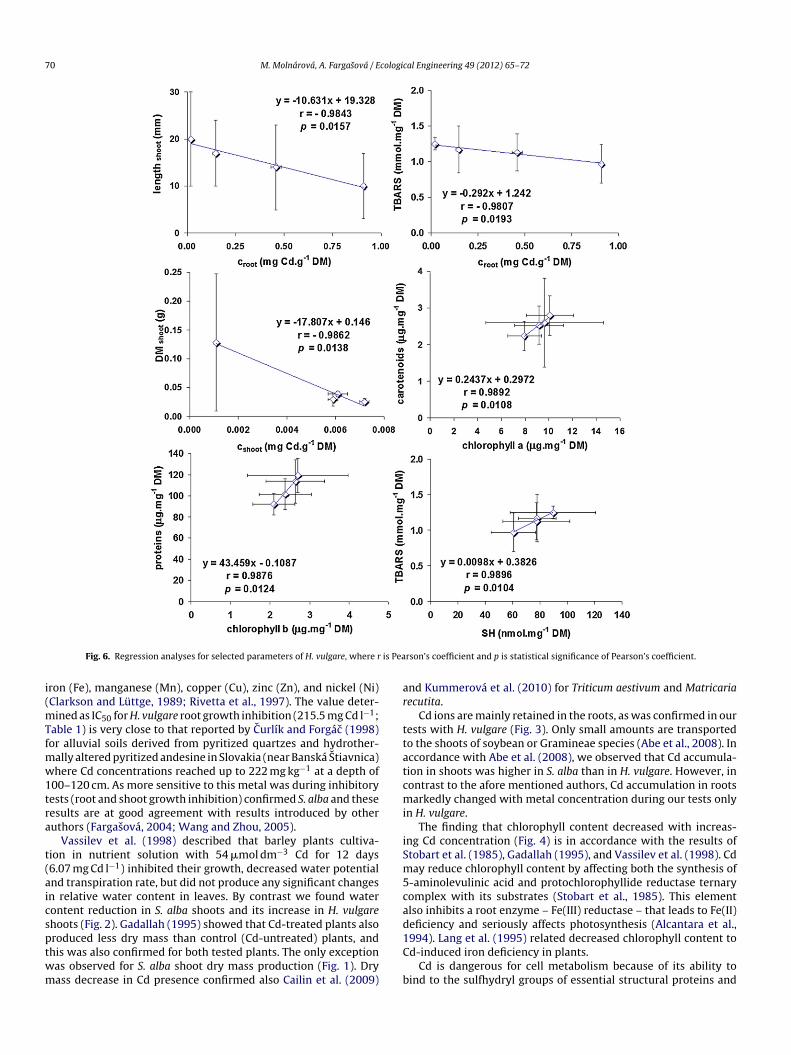
70 M. Molnárová, A. Fargasová / Ecological Engineering 49 (2012) 65– 72
is Pea
i(mTfmw1tra
t(aicsptwm
ar
ttatcmi
iSm5cad
Fig. 6. Regression analyses for selected parameters of H. vulgare, where r
ron (Fe), manganese (Mn), copper (Cu), zinc (Zn), and nickel (Ni)Clarkson and Lüttge, 1989; Rivetta et al., 1997). The value deter-
ined as IC50 for H. vulgare root growth inhibition (215.5 mg Cd l−1;able 1) is very close to that reported by Curlík and Forgác (1998)or alluvial soils derived from pyritized quartzes and hydrother-
ally altered pyritized andesine in Slovakia (near Banská Stiavnica)here Cd concentrations reached up to 222 mg kg−1 at a depth of
00–120 cm. As more sensitive to this metal was during inhibitoryests (root and shoot growth inhibition) confirmed S. alba and theseesults are at good agreement with results introduced by otheruthors (Fargasová, 2004; Wang and Zhou, 2005).
Vassilev et al. (1998) described that barley plants cultiva-ion in nutrient solution with 54 �mol dm−3 Cd for 12 days6.07 mg Cd l−1) inhibited their growth, decreased water potentialnd transpiration rate, but did not produce any significant changesn relative water content in leaves. By contrast we found waterontent reduction in S. alba shoots and its increase in H. vulgarehoots (Fig. 2). Gadallah (1995) showed that Cd-treated plants also
roduced less dry mass than control (Cd-untreated) plants, andhis was also confirmed for both tested plants. The only exceptionas observed for S. alba shoot dry mass production (Fig. 1). Dryass decrease in Cd presence confirmed also Cailin et al. (2009)1C
b
rson’s coefficient and p is statistical significance of Pearson’s coefficient.
nd Kummerová et al. (2010) for Triticum aestivum and Matricariaecutita.
Cd ions are mainly retained in the roots, as was confirmed in ourests with H. vulgare (Fig. 3). Only small amounts are transportedo the shoots of soybean or Gramineae species (Abe et al., 2008). Inccordance with Abe et al. (2008), we observed that Cd accumula-ion in shoots was higher in S. alba than in H. vulgare. However, inontrast to the afore mentioned authors, Cd accumulation in rootsarkedly changed with metal concentration during our tests only
n H. vulgare.The finding that chlorophyll content decreased with increas-
ng Cd concentration (Fig. 4) is in accordance with the results oftobart et al. (1985), Gadallah (1995), and Vassilev et al. (1998). Cday reduce chlorophyll content by affecting both the synthesis of
-aminolevulinic acid and protochlorophyllide reductase ternaryomplex with its substrates (Stobart et al., 1985). This elementlso inhibits a root enzyme – Fe(III) reductase – that leads to Fe(II)eficiency and seriously affects photosynthesis (Alcantara et al.,
994). Lang et al. (1995) related decreased chlorophyll content tod-induced iron deficiency in plants.Cd is dangerous for cell metabolism because of its ability toind to the sulfhydryl groups of essential structural proteins and

cologic
eelisp(asertaflCtoihetoibtppdipocte
posTih
eciacremsusps
5
otga
ssIdicCtsticgvbcw
A
t1
R
A
A
B
B
B
C
C
C
C
C
C
D
D
F
F
F
G
G
M. Molnárová, A. Fargasová / E
nzymes, which can lead to misfolding of proteins, inhibition ofnzyme activity, and/or interference with redox-enzymatic regu-ation (Hall, 2002). Constituents of the cell wall can immobilize Cdons and prevent cytosolic uptake. Once Cd enters into the cyto-ol, it stimulates the synthesis of sulfhydryl reach molecules likehytochelatins (PC), glutathione, and possibly metallothioneinsSharma and Dietz, 2006; DalCorso et al., 2008). Higher plants,lgae, and some fungi synthesize phytochelatins by the enzyme PCynthase, which is a constituent of the cytoplasm of plant cells (Stoltt al., 2003). Cd binds primarily to the sulfhydryl group of cysteineesidues in the PC peptide, and the Cd–PC complex is up to 1000imes less toxic to many plant enzymes than the free Cd ion (Kneernd Zenk, 1992). Vacuolar compartmentalization of Cd prevents theree circulation of metal ions in the cytosol and forces them into aimited area (Sanitá di Toppi and Gabbrielli, 1999). In presence ofd, S. alba showed a greater decrease in sulfhydryl group levelshan H. vulgare (Table 2). This can be explained by the synthesisf type 1 metallothioneins (MT), which was previously confirmedn barley genes (Cobbett and Goldsbrough, 2002). Moreover, theighest Cd concentrations in our study significantly decreased lev-ls of sulfhydryl groups not only in shoots of S. alba, but also inhose of H. vulgare (Table 2). The reduction in sulfhydryl groups isne of the most important oxidative stress parameters. Since Cds not considered a redox active element, it does not take part inasic oxidative stress reactions like Fenton and Haber–Weiss reac-ions. Metals without redox capacity (such as Cd2+) can enhance therooxidant status by reducing the antioxidant glutathione (GSH)ool (Pinto et al., 2003). General imbalances in ROS generation andetoxication, and consequately oxidative stress, may be apparent
n our experiments by the significantly lower concentrations ofrotein sulfhydryl groups (Table 2). However, it is not clear if thever-production of ROS during Cd treatment is the cause of redoxellular imbalance or if it is a specific stress mechanism activated byhe plant cell to cope with the heavy metal ions (Romero-Puertast al., 2004).
Cd changes the functionality of membranes by inducing lipideroxidation (Fodor et al., 1995). The final product of lipoper-xidation is malondialdehyde, which we determined by thepectrophotometric method TBARS. No significant changes inBARS levels (Table 2) were observed in our experiments, whichs in accordance with the results of Kumar et al. (2008) for Arachisypogaea seedlings.
Some authors (e.g. Tudoreanu and Phillips, 2004) analyzed lin-ar regression of published values only for soil parameters andadmium concentrations in plant tissue and try to develop empir-cal models with calculated of uptake coefficients that can bepplied in a wide range of circumstances. Prediction of metal bio-oncentration in different organisms and estimation of exposureisks can better assess dynamics of trace metals in organisms andcosystems (Fränzle et al., 2007). Revealing of relations amongeasured parameters can be one of the first steps for under-
tanding of Cd function/role in the plants through developing ofptake coefficient. This article contributes to mentioned area ofcientific interest not only with regression analysis of standardarameters but also by analysis of biochemical characteristics inemi-hydroponic conditions without soil particle interactions.
. Conclusions
The aim of this study was analysis of the relationships among
bserved parameters by using regression analyses. Based onhe results from physiological and production (root and shootrowth inhibition, dry and fresh mass, Cd accumulation), as wells biochemical (chlorophylls and carotenoids, proteins, proteinH
H
al Engineering 49 (2012) 65– 72 71
ulfhydryl groups, and TBARS) parameters, the following conclu-ions about Cd effects on S. alba and H. vulgare plants can be done.nhibition of root and shoot growth and fresh and dry biomass pro-uction were confirmed in accordance with published data. Decline
n photosynthetic pigment contents coupled with cadmium con-entration increase was observed mainly in S. alba shoots. Whiled concentration rose with its remaining concentration in the solu-ion mainly in roots of both plants, their translocation into thehoots was observed only for S. alba. Significant reduction of pro-ein sulfhydryl groups as well as decrease of TBARS were measuredn presence of Cd for both studied plants. Positive correlations wereonfirmed by regression analyses between proteins and sulfhydrylroups in S. alba shoots and TBARS and sulfhydryl groups in H.ulgare. On the contrary, negative correlations were determinedetween Cd concentrations in S. alba roots and proteins and TBARSontent as well as between Cd concentration in S. alba shoots andater content and chlorophyll a and sulfhydryl groups contents.
cknowledgments
This study was supported by the Grant of Ministry of Educa-ion, Science, Research and Sport of the Slovak Republic VEGA/1139/11.
eferences
be, T., Fukami, M., Ogasawara, M., 2008. Cadmium accumulation in the shoots androot of 93 weed species. Soil Sci. Plant Nutr. 54, 566–573.
lcantara, E., Romera, F.J., Canete, M., De La Guardia, M.D., 1994. Effects of heavymetals on both induction and function of root Fe(III)reductase in Fe-deficientcucumber (Cucumis sativus L.) plants. J. Exp. Bot. 45, 1893–1898.
enavides, M.P., Gallego, S.M., Tomaro, M.L., 2005. Cadmium toxicity in plants. Braz.J. Plant Physiol. 17, 21–34.
radford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein–dye binding. Anal.Biochem. 72, 248–254.
urton, K.W., King, J.B., Morgan, E., 1986. Chlorophyll as an indicator of the uppercritical tissue concentration of cadmium in plants. Water Air Soil Pollut. 27,147–154.
ailin, G.E., Ding, Y., Wang, Z., Wan, D., Wang, Y., Shang, Q., et al., 2009. Responses ofwheat seedlings to cadmium, mercury and trichlorobenzene stresses. J. Environ.Sci. 21, 806–813.
haturvedi, I., 2004. Phytotoxicity of cadmium and its effect on two genotypes ofBrassica juncea L. Emir. J. Agric. Sci. 16, 1–8.
larkson, D.T., Lüttge, U., 1989. Mineral nutrition. Divalent cations, transport andcompartmentalization. Prog. Bot. 51, 93–112.
lemens, S., 2006. Toxic metal accumulation, responses to exposure and mecha-nisms of tolerance in plants. Biochimie 88, 1707–1719.
obbett, C., Goldsbrough, P., 2002. Phytochelatins and metallothioneins: role inheavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 53, 159–182.
ˇurlík, L., Forgác, J., 1998. Acid sulfate weathering and its impact on soil acidifica-tion and pollution in area of Banská Stiavnica. In: Vedecké práce VÚPÚ, vol. 21,Bratislava, pp. 103–118.
alCorso, G., Farinati, S., Maistri, S., Furini, A., 2008. How plants cope with cad-mium: staking all on metabolism and gene expression. J. Integr. Plant Biol. 50,1268–1280.
razic, G., Mihailovic, N., 2005. Modification of cadmium toxicity in soybeanseedlings by salicylic acid. Plant Sci. 168, 511–517.
argasová, A., 2004. Toxicity comparison of some possible toxic metals (Cd, Cu, Pb,Se, Zn) on young seedlings of Sinapis alba L. Plant Soil Environ. 50, 33–38.
odor, A., Szabó-Nagy, A., Erdei, L., 1995. The effects of cadmium on the fluidityand H+-ATPase activity of plasma membrane from sunflower and wheat roots.J. Plant Physiol. 14, 787–792.
ränzle, S., Markert, B., Wünschmann, S., 2007. Dynamics of trace metals inorganisms and ecosystems: prediction of metal bioconcentration in differentorganisms and estimation of exposure risks. Environ. Pollut. 150, 23–33.
adallah, M.A.A., 1995. Effects of cadmium and kinetin on chlorophyll content,saccharides and dry mater accumulation in sunflower plants. Biol. Plant. 37,233–240.
allego, S.M., Benavídes, M.P., Tomaro, M.L., 1996. Effect of heavy metal ion excesson sunflower leaves: evidence for involvement of oxidative stress. Plant Sci. 121,151–159.
all, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J.Exp. Bot. 53, 1–11.
egedusová, A., Hegedus, O., Musilová, J., 2006. Riziká kontaminácie pôd kadmiom.Faculty of Natural Sciences, Constantine The Philosopher University in Nitra,Nitra.

7 cologi
H
H
K
K
K
K
L
L
L
M
O
P
R
R
R
S
S
S
S
S
T
T
V
V
2 M. Molnárová, A. Fargasová / E
endry, G.A.F., Baker, A.J.M., Ewart, C.F., 1992. Cadmium tolerance and toxic-ity, oxygen radical processes and molecular damage in cadmium-tolerantand cadmium-sensitive clones of Holcus lanatus. Acta Bot. Neerl. 41,271–281.
iller, E., Jurkovic, L’., Sutriepka, M., 2010. Metals in the surface sediments of selectedwater reservoirs, Slovakia. Bull. Environ. Contam. Toxicol. 84, 635–640.
alin, M., Wheeler, W.N., Olaveson, M.M., 2006. Response of phytoplankton to eco-logical engineering remediation of a Canadian Shield Lake affected by acid minedrainage. Ecol. Eng. 28, 296–310.
neer, R., Zenk, M.H., 1992. Phytochelatins protect plant enzymes from heavy metalpoisoning. Phytochemistry 31, 2663–2667.
umar, S., Mehta, U.J., Hazra, S., 2008. Accumulation of cadmium in growing peanut(Arachis hypogaea L.) seedlings – Its effect on lipid peroxidation and on theantioxidative enzymes catalase and guaiacol peroxidase. J. Plant Nutr. Soil Sci.171, 440–447.
ummerová, M., Zezulka, S., Král’ová, K., Masarovicová, E., 2010. Effect of zinc andcadmium on physiological and production characteristics in Matricaria recutita.Biol. Plant. 54, 308–314.
ane, T.W., Morel, F.M., 2000. A biological function for cadmium in marinediatomeas. Proc. Natl. Acad. Sci. U.S.A. 97, 4627–4631.
ang, F., Sarvari, E., Szigeti, Z., Fodor, F., Cseh, E., 1995. Effects of heavy metalson the photosynthetic apparatus in cucumber. In: Mathis, P. (Ed.), Photo-synthesis: From Light to Biosphere, vol. IV. Kluwer Academic Publishers,Dordrecht/Boston/London, pp. 533–536.
ichtenthaler, H.K., Wellburn, A.R., 1983. Determination of total carotenoids andchlorophyls a and b of leaf extracts in different solvents. Biochem. Soc. Trans.603, 591–592.
aksymiec, W., Krupa, Z., 2006. The effects of short-term exposition to Cd, excessCu ions and jasmonate on oxidative stress appearing in Arabidopsis thaliana.Environ. Exp. Bot. 57, 187–194.
ECD (Organization for Economic Co-operation and Development), 2006. Guide-
lines for the Testing of Chemicals. Terrestrial Plant Test: Seedling Emergenceand Seedling Growth Test, Test 208. OECD, Paris.into, E., Sigaud-Kutner, T.C.S., Leitão, M.A.S., Okamoto, O.K., Morse, D., Colepi-colo, P., 2003. Heavy metal-induced oxidative stress in algae. J. Phycol. 39,1008–1018.
V
W
cal Engineering 49 (2012) 65– 72
ivetta, A., Negrini, N., Cocucci, M., 1997. Involvement of Ca2+-calmodulin in Cd2+
toxicity during the early phases of radish (Raphanus sativus L.) seed germination.Plant Cell Environ. 20, 600–608.
omero-Puertas, M.C., Rodriguez-Serrano, M., Corpas, F.J., Gomez, M., del Rio, L.A.,Sandalio, L.M., 2004. Cadmium-induced subcellular accumulation of O2− andH2O2 in pea leaves. Plant Cell Environ. 27, 1122–1134.
oucoux, P., Dabin, P., 1977. The effect of cadmium on the nitrogen fixation. Paperpresented at Semin. Carbohydrate and Protein Synthesis, Giessen, September 7,1977, p. 215.
anitá di Toppi, L., Gabbrielli, R., 1999. Response to cadmium in higher plants. Envi-ron. Exp. Bot. 41, 105–130.
harma, S.S., Dietz, K.J., 2006. The significance of amino acids and amino acid-derivedmolecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot.57, 711–726.
tobart, A.K., Griffiths, W.T., Ameen-Bukhari, I., Sherwood, R.P., 1985. The effect ofCd2+ on the biosynthesis of chlorophyll in leaves of barley. Physiol. Plant. 63,293–298.
tohs, S.J., Bagchi, D., 1995. Oxidative mechanisms in the toxicity of metal ions. FreeRadic. Biol. Med. 18, 321–336.
tolt, J.P., Sneller, F.E.C., Bryngelsson, T., Lundborg, T., Schat, H., 2003. Phytochelatinand cadmium accumulation in wheat. Environ. Exp. Bot. 49, 21–28.
osner, J., Parízek, O., Kocí, V., 2012. Vliv vlastností pud na vysledky kontaktníchtestu ekotoxicity (Influence of soil properties on results of contact ecotoxicitytests). Acta Environ. Univ. Comenianae (Bratislava) 20, 89–93.
udoreanu, L., Phillips, C.J.C., 2004. Empirical models of cadmium accumulation inmaize, rye grass and soya bean plants. J. Sci. Food Agric. 84, 845–852.
assilev, A., Berova, M., Zlatev, Z., 1998. Influence of Cd2+ on growth, chloro-phyll content, and water relations in young barley plants. Biol. Plant. 41, 601–606.
iner, R.I., Hühmer, A.F.R., Bigelow, D.J., Schöneich, C., 1996. The oxidative inacti-vation of sarcoplasmic reticulum Ca2+-ATPase by peroxynitrite. Free Radic. Res.
24, 243–259.ymazal, J., 1987. Toxicity and accumulation of cadmium with respect to algae andcyanobacteriea: a review. Environ. Toxicol. Water Qual. 2, 387–415.
ang, X.-F., Zhou, Q.-X., 2005. Ecotoxicological effects of cadmium on three orna-mental plants. Chemosphere 60, 16–21.
Related Documents











