Relationship between freshwater input to the coastal zone and the historical landings of the benthic/demersal fish Eleginops maclovinus in central-south Chile R. A. QUIN ˜ ONES * AND R. M. MONTES Departamento de Oceanografı a, Universidad de Concepcio ´ n, Casilla 160-C, Concepcio ´ n, Chile ABSTRACT The ecosystem off central-south Chile is one of the most productive marine systems in the world, provi- ding approximately 4% of worldwide fish captures. We analysed the effect of freshwater input (river runoff and rainfall) to the coastal zone in central-south Chile (36°00¢–37°30¢ lat. S) on the landings of the Ro ´balo (Eleginops maclovinus) using the following data: Ro ´balo annual landing statistics, monthly mean and annual mean runoff of Itata (1975–91) and Bio-Bio rivers (1975–94), and cumulative monthly rainfall and annual mean rainfall (1975–94). Time series were smoothed using three-point equally weighted running means. To analyse the relationship between Ro ´balo’s landing and river runoff, as well as rainfall, Pearson’s product-moment correlation analyses were conducted at relevant lag times. The statistical significance of correlation coefficients was calculated taking into account intraseries autocorrelation. Our results showed that Ro ´balo’s landings were significantly cor- related to freshwater input to the coastal zone in central-south Chile. Thus, significant correlation coefficients were found between annual Ro ´balo’s landings and: (i) annual mean runoff of the Itata (r )0.94) and Bio-Bio (r )0.80) rivers lagged 3 and 4 years, respectively, and (ii) annual mean rainfall lagged 4 years (r )0.75). The lag time at which highest correlation coefficients between annual mean environmental and annual landing time series took place (i.e. 3–4 years) corresponded to the age at which Ro ´balo reaches the minimum exploitation size. Accordingly, we hypothesize that river runoff and rainfall strongly influence the survival rate of Ro ´balo’s early stages during the first year of life. Key words: Chile, Eleginops maclovinus, freshwater input, Humboldt Current System, rainfall, river runoff, Ro ´balo INTRODUCTION Environmental variability on different time and space scales can strongly influence population dynamics (e.g. Sharp and Csirke, 1983; Cury and Roy, 1989; Kawasaki et al., 1991; Mullin, 1993). Interannual variation in oceanographic and atmo- spheric conditions are able to produce important fluctuations in fish recruitment and, consequently, in population biomass (e.g. Quinn II and Niebauer, 1995; Arnstz and Fahrbach, 1996). The environ- ment/resource relationship is especially important in the coastal and estuarine zone where freshwater input can affect the abundance and distribution of fish populations (Pati, 1984; Drinkwater, 1986; Gargett, 1997; Runge et al., 1999). Thus, significant relationships have been found between the runoff of the St. Lawrence (e.g. Sutcliffe, 1972, 1973; Nilo et al., 1997), Miramichi (Gibson and Myers, 1988) and Zambesi (Gammelsrod, 1992) rivers, among others, and the landings, catches or abundance indices of several commercial species. On the other hand, rainfall has also been considered an important factor in prerecruit survival of fish stocks (Harvey and Davies, 1970; Ulanowicz et al., 1982; Power et al., 2000). Most studies confronting the relationship between abundance/catches of marine resources and freshwater input to ecosystems have been based on correlation analysis. As such they cannot prove a cause–effect relationship but only an association between variables. Correlation studies on historical time series of envi- ronmental and fisheries’ variables have been criticised due to the low success rate of most predictive models and the possibility of presenting spurious correlation (e.g. Walters and Collie, 1988; Baumann, 1998). *Correspondence. e-mail: [email protected] Received 24 September 1998 Revised version accepted 15 November 1999 FISHERIES OCEANOGRAPHY Fish. Oceanogr. 10:4, 311–328, 2001 Ó 2001 Blackwell Science Ltd. 311

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Relationship between freshwater input to the coastal zoneand the historical landings of the benthic/demersal ®shEleginops maclovinus in central-south Chile
R. A. QUINÄ ONES* AND R. M. MONTES
Departamento de Oceanografõ�a, Universidad de Concepcio n,Casilla 160-C, Concepcio n, Chile
ABSTRACT
The ecosystem off central-south Chile is one of themost productive marine systems in the world, provi-ding approximately 4% of worldwide ®sh captures. Weanalysed the effect of freshwater input (river runoffand rainfall) to the coastal zone in central-south Chile(36°00¢±37°30¢ lat. S) on the landings of the RoÂbalo(Eleginops maclovinus) using the following data: RoÂbaloannual landing statistics, monthly mean and annualmean runoff of Itata (1975±91) and Bio-Bio rivers(1975±94), and cumulative monthly rainfall andannual mean rainfall (1975±94). Time series weresmoothed using three-point equally weighted runningmeans. To analyse the relationship between RoÂbalo'slanding and river runoff, as well as rainfall, Pearson'sproduct-moment correlation analyses were conductedat relevant lag times. The statistical signi®cance ofcorrelation coef®cients was calculated taking intoaccount intraseries autocorrelation. Our resultsshowed that RoÂbalo's landings were signi®cantly cor-related to freshwater input to the coastal zone incentral-south Chile. Thus, signi®cant correlationcoef®cients were found between annual RoÂbalo'slandings and: (i) annual mean runoff of the Itata(r � )0.94) and Bio-Bio (r � )0.80) rivers lagged 3and 4 years, respectively, and (ii) annual mean rainfalllagged 4 years (r � )0.75). The lag time at whichhighest correlation coef®cients between annual meanenvironmental and annual landing time series tookplace (i.e. 3±4 years) corresponded to the age at whichRoÂbalo reaches the minimum exploitation size.Accordingly, we hypothesize that river runoff and
rainfall strongly in¯uence the survival rate of RoÂbalo'searly stages during the ®rst year of life.
Key words: Chile, Eleginops maclovinus, freshwaterinput, Humboldt Current System, rainfall, river runoff,RoÂbalo
INTRODUCTION
Environmental variability on different time andspace scales can strongly in¯uence populationdynamics (e.g. Sharp and Csirke, 1983; Cury andRoy, 1989; Kawasaki et al., 1991; Mullin, 1993).Interannual variation in oceanographic and atmo-spheric conditions are able to produce important¯uctuations in ®sh recruitment and, consequently, inpopulation biomass (e.g. Quinn II and Niebauer,1995; Arnstz and Fahrbach, 1996). The environ-ment/resource relationship is especially important inthe coastal and estuarine zone where freshwaterinput can affect the abundance and distribution of®sh populations (Pati, 1984; Drinkwater, 1986;Gargett, 1997; Runge et al., 1999).
Thus, signi®cant relationships have been foundbetween the runoff of the St. Lawrence (e.g. Sutcliffe,1972, 1973; Nilo et al., 1997), Miramichi (Gibson andMyers, 1988) and Zambesi (Gammelsrod, 1992) rivers,among others, and the landings, catches or abundanceindices of several commercial species. On the otherhand, rainfall has also been considered an importantfactor in prerecruit survival of ®sh stocks (Harvey andDavies, 1970; Ulanowicz et al., 1982; Power et al.,2000).
Most studies confronting the relationship betweenabundance/catches of marine resources and freshwaterinput to ecosystems have been based on correlationanalysis. As such they cannot prove a cause±effectrelationship but only an association between variables.Correlation studies on historical time series of envi-ronmental and ®sheries' variables have been criticiseddue to the low success rate of most predictive modelsand the possibility of presenting spurious correlation(e.g. Walters and Collie, 1988; Baumann, 1998).
*Correspondence. e-mail: [email protected]
Received 24 September 1998
Revised version accepted 15 November 1999
FISHERIES OCEANOGRAPHY Fish. Oceanogr. 10:4, 311±328, 2001
Ó 2001 Blackwell Science Ltd. 311
However, when correlation studies are conductedwithin a sound conceptual and methodologicalframework, they can provide important insights aboutthe relationship between the environment and thedistribution and abundance of marine resources (Tyler,1992).
The marine ecosystem off central-south Chile is avery productive system with an average annual primaryproductivity of 2.54 g C m±2 day±1 (Daneri et al., 2000)and daily values as high as 9.9 g C m±2 day±1 (Fossinget al., 1995), which are among the highest ever reportedin the literature. This ecosystem also provides around3.5 million tonnes of ®sh capture per year, approxi-mately 4% of the worldwide total (FAO, 1995). Theproductivity is fuelled mostly by wind-driven upwelling(Arcos and Navarro, 1986), although Kelvin wave-associated mixing events have also been postulated asan important factor in triggering and maintaining thisproductivity (Djurfeldt, 1989). The continental shelf incentral-south Chile is also characterized by the pres-
ence of extensive benthic mats of the nitrate-reducingbacteria Thioploca (Gallardo, 1977; Fossing et al.,1995), and signi®cant freshwater and nutrient inputs(Della Croce et al., 1992). Moreover, the central-southshelf has been recognized as a spawning and nurseryarea for many ®sh species (Rojas et al., 1983; Castilloet al., 1991; Arcos et al., 1996; Castro et al., 1997).
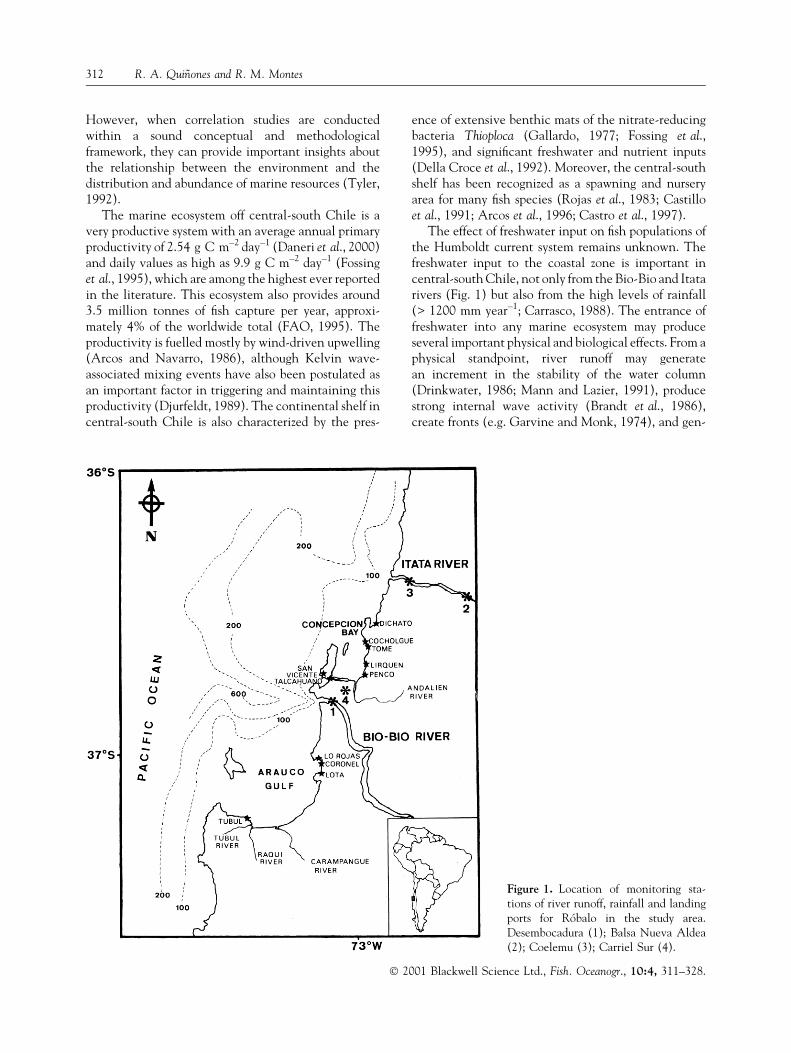
The effect of freshwater input on ®sh populations ofthe Humboldt current system remains unknown. Thefreshwater input to the coastal zone is important incentral-south Chile, not only from the Bio-Bio and Itatarivers (Fig. 1) but also from the high levels of rainfall(> 1200 mm year±1; Carrasco, 1988). The entrance offreshwater into any marine ecosystem may produceseveral important physical and biological effects. From aphysical standpoint, river runoff may generatean increment in the stability of the water column(Drinkwater, 1986; Mann and Lazier, 1991), producestrong internal wave activity (Brandt et al., 1986),create fronts (e.g. Garvine and Monk, 1974), and gen-
Figure 1. Location of monitoring sta-tions of river runoff, rainfall and landingports for RoÂbalo in the study area.Desembocadura (1); Balsa Nueva Aldea(2); Coelemu (3); Carriel Sur (4).
312 R. A. Quin~ones and R. M. Montes
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.

erate horizontal circulation due to horizontal densitydifferences (Drinkwater, 1986). At the same time, sev-eral important biological effects of river runoff havebeen proposed, such as: (i) changes in phytoplanktonbiomass and/or productivity of estuaries (e.g. Neu, 1975,1976; Funicelli, 1984; Levasseur et al., 1984; Therriaultand Levasseur, 1985; Smetacek, 1986); (ii) changes infood abundance for ®sh larvae (Skreslet, 1976); (iii)changes in the distribution of species (Mitchel, 1976;Brandt et al., 1986); (iv) changes in the natural mor-tality rates of early stages of invertebrate and vertebratemarine species (Sutcliffe, 1972, 1973; Wilber, 1992);and (v) alteration of population parameters such asgrowth, recruitment, spawning time, predation rates,etc. (e.g. Sutcliffe, 1973; Sheldon et al., 1982; Da Silva,1986; Donn, 1987).
The coastal and shelf ecosystem off central-southChile supports approximately 59% of the nationallandings. Part of the life cycle of most of theseresources takes place in the neritic pelagic system thatis potentially in¯uenced by freshwater input in thisregion (Montes and QuinÄones, 1999). Therefore wehypothesize that the population dynamics of some ofthese resources inhabiting the coast of central-southChile may be associated with the variations of fresh-water input into the coastal zone.
Here, we conduct an exploratory analysis to testwhether there is an association between freshwaterinput to the coastal zone in central-south Chile andthe landings of the benthic/demersal ®sh RoÂbalo(Eleginops maclovinus; Cuvier and Valenciennes, 1830)during the 1975±94 period.
MATERIALS AND METHODS
Study area
The study was conducted in the coastal zone of cen-tral-south Chile (36°00¢±37°30¢ lat; S Fig. 1). The Bio-Bio and the Itata rivers are the two major rivers ofcentral-south Chile, with typical annual average run-off at their mouths of 900 m3 s±1 and 300 m3 s±1,respectively (Chilean General Water Directorate,unpublished data).
Distribution and biology of Eleginops maclovinus
Eleginops maclovinus is one of the three genera of thefamily Nototheniidae and the only species of the genusEleginops (PequenÄo, 1989). It is thought to be a phy-logenetically primitive form related to Dissostichus(Balushkin, 1984; Johnston et al., 1998). The speciesis euryhaline and eurythermic (PequenÄo, 1989), andinhabits the coastal area of the southern zone of South
America. In the Paci®c Ocean its habitat extends fromthe Beagle Channel (54°S) to ValparaõÂso (33°S)(GuzmaÂn and CampodoÂnico, 1973; PequenÄo, 1989).In the Atlantic Ocean, the RoÂbalo lives from theBeagle Channel to the coast of Uruguay (35°S; East-man, 1993), including the Falkland/Malvinas Islands(Gosztonyi, 1974).
The RoÂbalo is commonly found at the mouth ofsouthern-Chile rivers (PequenÄo, 1979), and it hasbeen observed several kilometres upstream (Mann,1954; Ruiz, 1993). It is classi®ed as a neritic specieswith demersal to benthic characteristics (PequenÄo,1979) and it has not been detected at depths deeperthan 40 m, con®rming its preference for neritic andlittoral habitats (Trujillo, 1972; PequenÄo, 1979, 1981).The RoÂbalo preys mainly on benthic organisms(e.g. polychaetes, crustaceans) of estuarine areasin¯uenced by tides, and it is de®ned as omnivorouswith secondary carnivore predominance (PequenÄo,1979). Important predation on algae has been reportedfor specimens larger than 16 cm (GuzmaÂn andCampodoÂnico, 1973).
The RoÂbalo is a sequential protandrous herma-phrodite (Calvo et al., 1992) with partial spawningtypically occurring in June in central-south Chile(Panozo, 1996). According to Ruiz (1993) thespawning takes place in the estuary, and the plank-tonic early stages remain mostly there until reachingthe juvenile stage when they begin making incursionsinto the adjacent coastal waters. The juveniles per-manently move between the coastal and estuarinewaters, although they can also live a few kilometresupstream in river waters (Ruiz, 1993).
Time series
The main landings of E. maclovinus take place in thefour major ports of the study area: Talcahuano, SanVicente, Coronel, and Tome (Fig. 1). However,occasional landings also take place in some smallerports and villages such as Penco, Lo Rojas, Lota, andTubul (Fig. 1; SERNAPESCA, unpublished data).The landing statistics were obtained from the AnnualReports of the Chilean National Fisheries Service(SERNAPESCA) and cover every year from 1975 to1994 (SERNAP, 1975±94; SERNAPESCA, 1995).
During the analysed period, the RoÂbalo was notunder strong ®shing pressure and is considered a minor®shery (approximately less than 0.002% of the total®sh catch) in central-south Chile. The history andcharacteristics of this artisanal ®shery in central-southChile shows that ®shing effort has not changed sig-ni®cantly during the study period. Unfortunately,there is insuf®cient information to standardize
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 313

historical ®shing effort, precluding any attempt togenerate more precise relative indexes of abundancethan landing levels.
The river runoff data of the Bio-Bio and Itata riverswas obtained from the monthly mean records collectedby the Chilean General Water Service from 1975 to1994 at the sampling stations Desembocadura(36°50¢S; 73°05¢W) and Balsa Nueva Aldea (36°39¢S;72°27¢W), respectively (Fig. 1). The sampling stationBalsa Nueva Aldea is located more upstream (approx.25 km) than the Coelemu station (Fig. 1), butunfortunately the latter only has records for the period1986±88. In order to test the validity of using the datacoming from Balsa Nueva Aldea, we performed acorrelation analysis between monthly runoff datacoming from this and the Coelemu stations during1986±88 obtaining a correlation coef®cient of 0.92(P < 0.01). In consequence the data from Balsa NuevaAldea was considered an adequate approximation ofthe runoff temporal dynamics at the mouth of the ItataRiver.
The joint discharge of the Bio-Bio and Itata riversis operationally called RIVSUM. Since these riverscorrespond to more than 90% of the runoff of thestudy area, RIVSUM is an indicator of total riverfreshwater input to the coastal zone of central-southChile.
The rainfall data (1975±94) were obtained from thedaily monitoring by the Chilean MeteorologicalDirectorate at the Carriel Sur station (36°46¢S;73°03¢W; Fig. 1). The rainfall data were aggregated ona monthly basis as cumulative monthly rainfall (i.e.the sum of daily rainfall during a speci®c month) andannually as annual mean rainfall (i.e. the average ofcumulative monthly rainfall during one year).
Statistical methods
The intra-annual variability of river runoff and rainfallwas characterized by determining, from the raw timeseries, the average value for each month during theyears covered in this study, i.e. 20 years for the Bio-Biorunoff, 17 years for the Itata runoff and 20 years forthe rainfall data at Carriel Sur.
The landing data were smoothed using 3-yearrunning means. This procedure improves the detectionof long-term trends in the interannual abundance of®sheries resources (e.g. Sutcliffe et al., 1977; Welch,1986; Parker et al., 1995), and reduces the high-fre-quency variability in the time series (Drinkwater et al.,1991; Pyper and Peterman, 1998). Furthermore, itdiminishes the effects of the age-structure in theannual landing data (Sutcliffe et al., 1976; Hollowedet al., 1987; Drinkwater et al., 1991; Parker et al.,
1995). The smoothing of the annual environmentaltime series (i.e. river runoff and rainfall) was alsoconducted using 3-year running means. In the case ofthe monthly environmental time series three-pointrunning means were used; in other words, the mean ofthe same month in three successive years (e.g. March1985, March 1986, March 1987).
To investigate the relationship (if any) betweenRoÂbalo landings and freshwater input (i.e. river runoffand rainfall), the lag correlation method was used(e.g. Sutcliffe, 1973; Sutcliffe et al., 1977; Ulanowiczet al., 1982; Drinkwater et al., 1991; Wilber, 1992).The main objective of our manuscript is to establishwhether there is any historical association betweenriver runoff/rainfall and the landings of the RoÂbalo incentral-south Chile. In the case of the RoÂbalo ®shery,the annual scale (i.e. annual landing) seems to beadequate to observe possible associations with any ofthe environmental variables because, at least in thesouthern zone of Chile (lat. 39.15±40°S), it presents acertain degree of seasonal trends due to the effect ofclimatic conditions on artisanal ®shermen's behaviour(PequenÄo, 1987). In contrast, the daily, weekly, andmonthly landings rely on the short-term dynamics ofthe ®shery, which depend on several external factorssuch as meteorological conditions and market pres-sures.
On the other hand, it is critical to analyse theassociation between the independent variables(i.e. rainfall, runoff) on a monthly scale with theRoÂbalo annual landings because basic characteristicsof its life cycle (e.g. spawning, recruitment, etc.) couldbe affected by environmental variability at shorterthan annual time scales. In addition, freshwater inputat the annual scale is meaningful in analysing thecumulative effect of the independent variables onRoÂbalo landings.
It is important to highlight that a month-to-monthcomparison between freshwater input and RoÂbalolandings does not have any biological meaning in thecase of this ®shery, because the increment (or decre-ment) of the survival rate of any early stage (includingjuveniles) occurring in a given month is not neces-sarily re¯ected on the catch level in a given monthone or several years later.
After smoothing the time series, Pearson correla-tion coef®cients were calculated between (i) the timeseries of annual RoÂbalo landings and the time series ofannual mean environmental data with differentannual lag times (i.e. 1¼5 years), and (ii) the timeseries of annual RoÂbalo landings and the time series ofthe environmental variable for each month of the year
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
314 R. A. Quin~ones and R. M. Montes

(i.e. monthly mean runoff, cumulative monthly rain-fall) with different annual lag times (i.e. 1¼5 years).
It is known that running means increase the auto-correlation and consequently reduce the effectivenumber of independent data points in both time series(Drinkwater and Myers, 1987). One way to estimateintraseries autocorrelation in a highly autocorrelatedtime series is to calculate the effective number ofindependent data points (N*) (Drinkwater and Myers,1987; Peterman et al., 1998; Pyper and Peterman,1998; Zheng and Kruse, 2000). An adjusted number ofdegrees of freedom (i.e. N* ) 2) was used in testingthe statistical signi®cance of correlation coef®cientsbetween landings and environmental time series.Following Pyper and Peterman (1998), for a time seriesof N points, N* is given by Eq. 1.
1
N�� 1
N� 2
N
XN=5
j�1
rXX�j�rYY�j� �1�
where N is the number of sample pairs and rxx (j) andryy (j) denote the sample autocorrelation at lag j of thetwo time series being correlated. Simulation analyses(Pyper and Peterman, 1998) have shown that this testprocedure performs well at maintaining type I errorrates at the speci®ed alpha when time series datacontain positive autocorrelation typical of AR(1)(®rst-order autoregressive model) processes. We usedthe autocorrelation estimator recommended byChat®eld (1989) and Pyper and Peterman (1998)which is very robust to estimate N* when time seriesare short. The estimator is calculated as shown in Eq. 2:
rXX�j� � N
Nÿ j�PNÿj
t�1
�Xt ÿ X��Xt�j ÿ X�PNt�1
�Xt ÿ X�2�2�
The signi®cance level of the correlation coef®cientsbetween the time series of landing and environmentalvariables were evaluated using both P and P*, wherethe latter is the level of signi®cance for N*. In order toavoid spurious correlations, we accepted as valid onlythose correlation coef®cients with P* £ 0.05. Never-theless, the correlation coef®cients of every pair ofvariables tested both at P and P* level were reported.
Relationships between landing, runoff, and rainfallwere calculated using single linear regression models,but only in those cases where the correlation analysishad previously shown signi®cant coef®cients at P*level. We used this approach instead of the time series
models (i.e. transfer function) since the dependentvariable (annual RoÂbalo landings) in our time serieshas a maximum length of 20 years, which is much lessthan the minimum necessary (i.e. approximately 50±60data points in time) to properly use Box±Jenkinsapproaches (i.e. AR, ARIMA and transfer functionsmodels, e.g. Box and Jenkins, 1976; Prager andHoenig, 1992; Rothschild et al., 1996). Finally, astepwise regression analysis was conducted to estimatethe contribution of each environmental variable tothe observed variance in RoÂbalo annual landings(Draper and Smith, 1998).
RESULTS
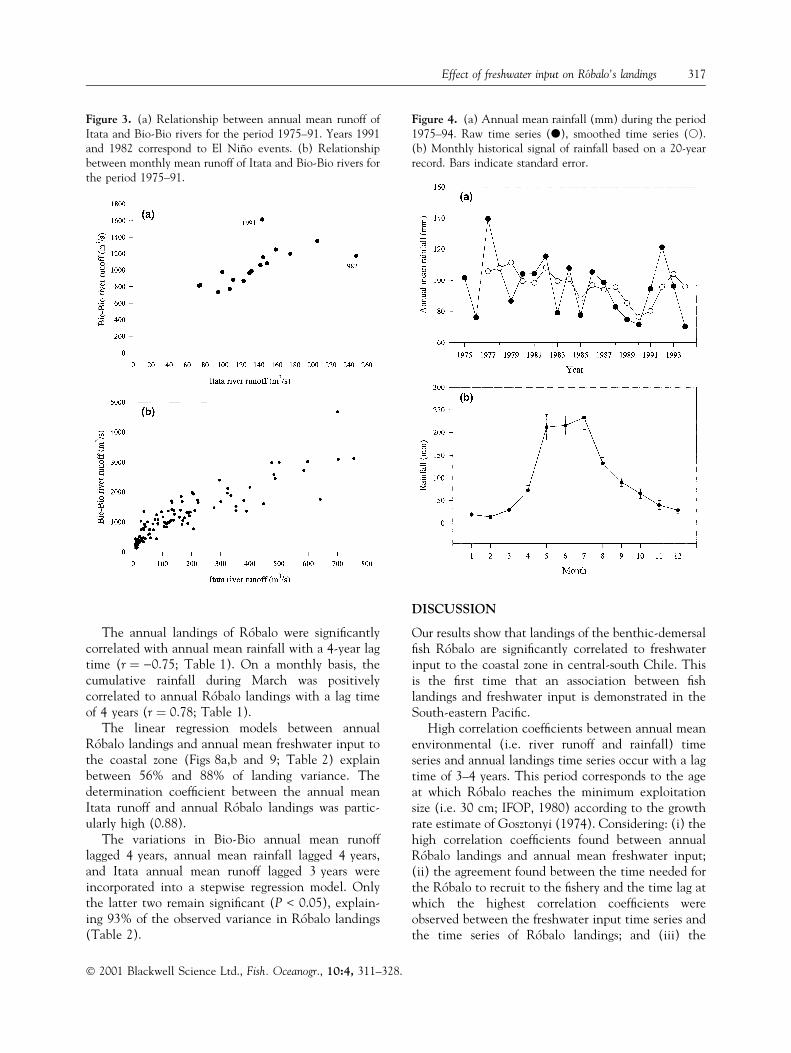
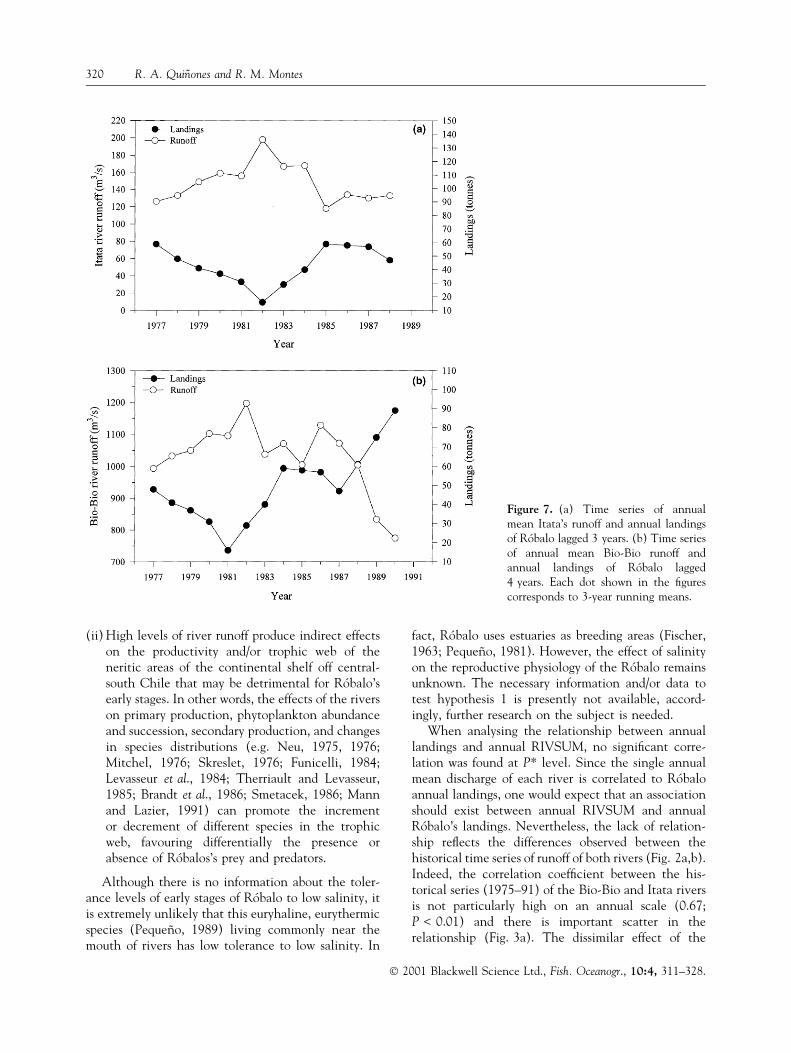
River runoff time series are shown in Fig. 2. Themaximum annual runoff of each series took place indifferent years, 1991 and 1982 being the years ofhighest runoff for the Bio-Bio and the Itata rivers,respectively (Fig. 2a,b; Fig. 3a). The historical timeseries of Itata and Bio-Bio river runoffs (1975±91) aresigni®cantly correlated at the annual level (r � 0.67;P < 0.01), but they present important differences(Fig. 3a). High precipitation levels are observed incentral-south Chile during El NinÄo events (Rutllantand Fuenzalida, 1991; Rutllant, 1997). During El NinÄo1982, the Itata had the highest annual runoff of itstime series but the Bio-Bio runoff was only slightlyhigher than the average historical level (Fig. 3a). Incontrast, during El NinÄo 1991 the Bio-Bio presentedthe highest annual runoff of its time series, while theItata annual runoff was close to the historical mean(Fig. 3a). At the monthly time scale there are alsosigni®cant differences in the simultaneous runoff levelsbetween both rivers (Fig. 3b). Both rivers have dissi-milar hydrological dynamics due to the differentialeffect of factors such as total surface and shape of eachbasin (Bio-Bio � 24360 km2; Itata � 11385 km2),river lengths (Bio-Bio � 380 km; Itata � 230 km),nival in¯uences, spatial and temporal dynamics ofrainfall level, and river slope (SantibanÄez and Uribe,1993; Parra, 1996; Parra and Habit, 1998; EULA,1999).
The RIVSUM time series (Fig. 2c) resembles, to acertain degree, the Bio-Bio time series because thehistorical annual average of the time series of the Bio-Bio river runoff (Fig. 2a) is almost three times largerthan that of the Itata (Fig. 2b). The intra-annualseasonal signal is similar for both rivers, with maxi-mum runoff taking place during austral winter andminimum runoff during austral summer (Fig. 2d). Thehighest peaks of annual mean rainfall took place in1977 and 1992 (Fig. 4a). The seasonal runoff signal
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 315
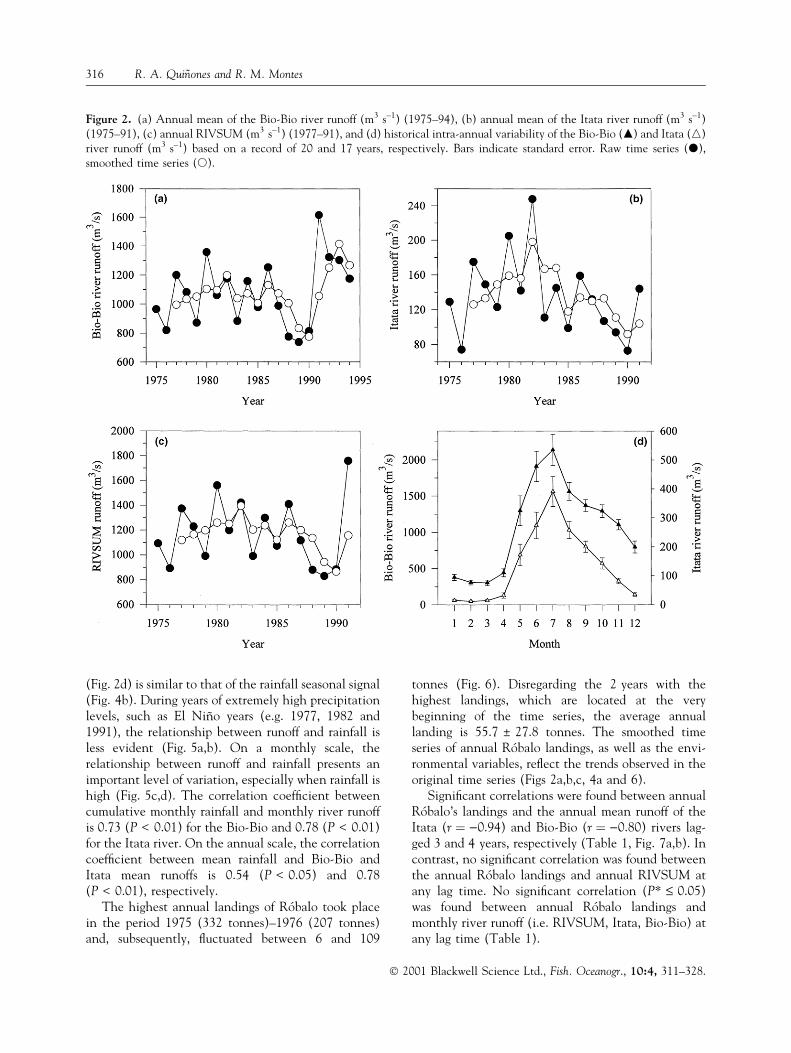
(Fig. 2d) is similar to that of the rainfall seasonal signal(Fig. 4b). During years of extremely high precipitationlevels, such as El NinÄo years (e.g. 1977, 1982 and1991), the relationship between runoff and rainfall isless evident (Fig. 5a,b). On a monthly scale, therelationship between runoff and rainfall presents animportant level of variation, especially when rainfall ishigh (Fig. 5c,d). The correlation coef®cient betweencumulative monthly rainfall and monthly river runoffis 0.73 (P < 0.01) for the Bio-Bio and 0.78 (P < 0.01)for the Itata river. On the annual scale, the correlationcoef®cient between mean rainfall and Bio-Bio andItata mean runoffs is 0.54 (P < 0.05) and 0.78(P < 0.01), respectively.
The highest annual landings of RoÂbalo took placein the period 1975 (332 tonnes)±1976 (207 tonnes)and, subsequently, ¯uctuated between 6 and 109
tonnes (Fig. 6). Disregarding the 2 years with thehighest landings, which are located at the verybeginning of the time series, the average annuallanding is 55.7 � 27.8 tonnes. The smoothed timeseries of annual RoÂbalo landings, as well as the envi-ronmental variables, re¯ect the trends observed in theoriginal time series (Figs 2a,b,c, 4a and 6).
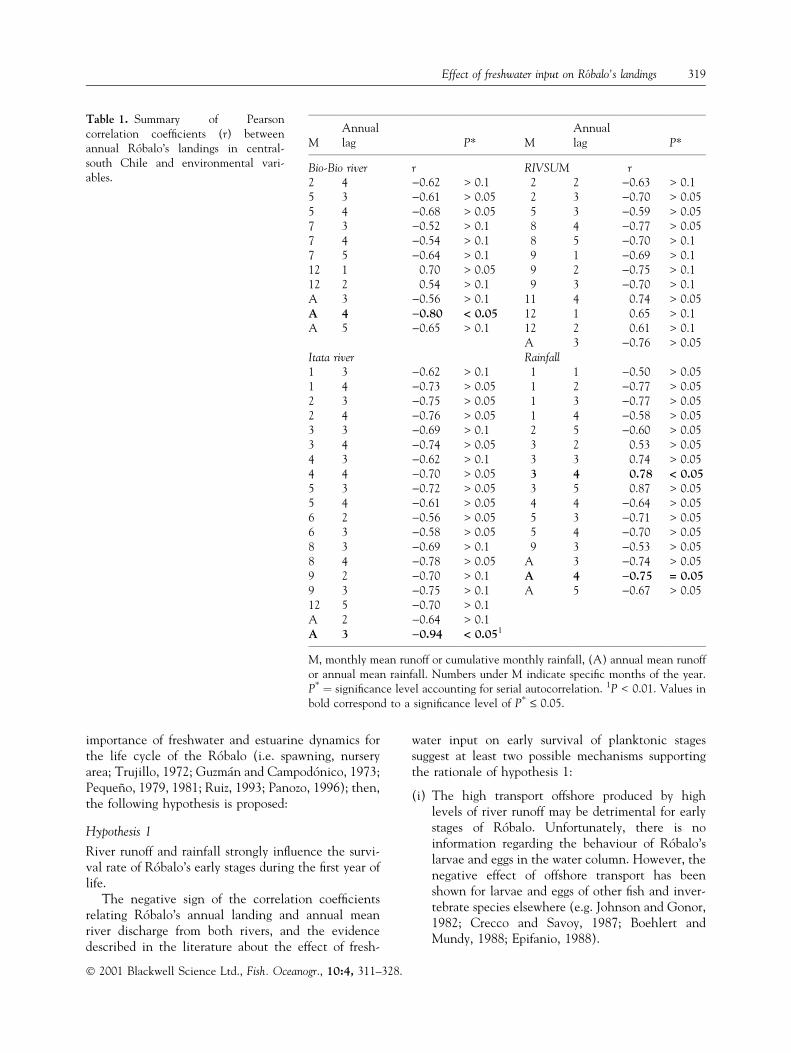
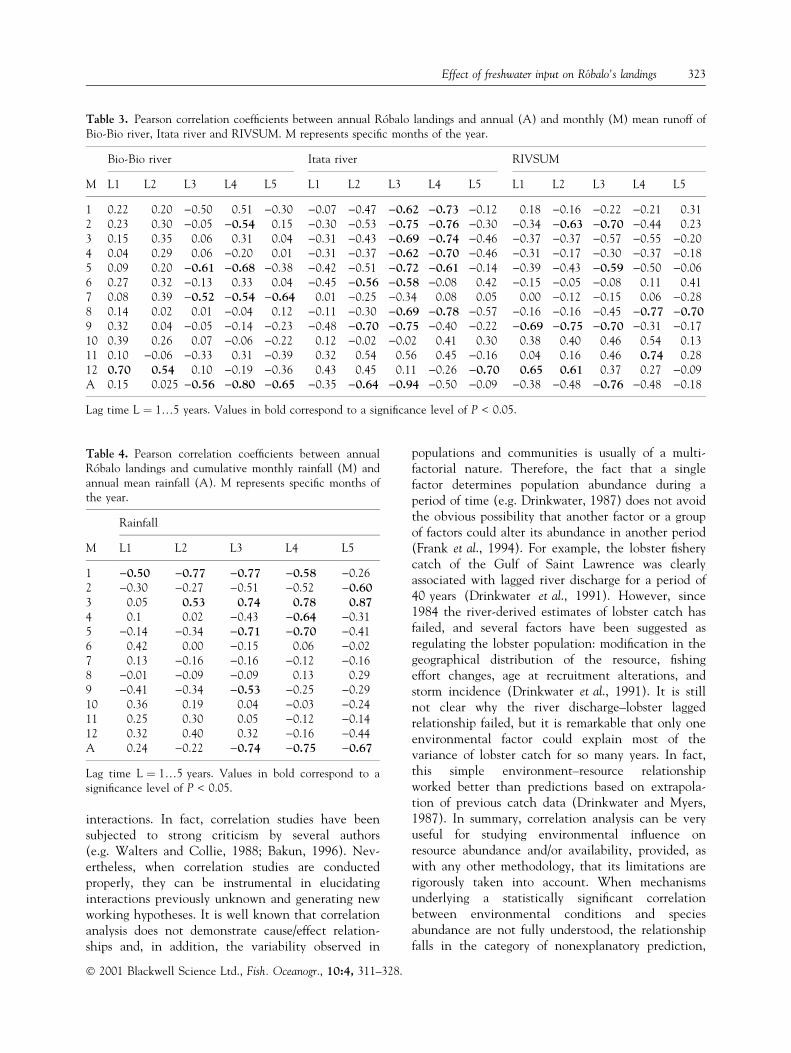
Signi®cant correlations were found between annualRoÂbalo's landings and the annual mean runoff of theItata (r � )0.94) and Bio-Bio (r � )0.80) rivers lag-ged 3 and 4 years, respectively (Table 1, Fig. 7a,b). Incontrast, no signi®cant correlation was found betweenthe annual RoÂbalo landings and annual RIVSUM atany lag time. No signi®cant correlation (P* £ 0.05)was found between annual RoÂbalo landings andmonthly river runoff (i.e. RIVSUM, Itata, Bio-Bio) atany lag time (Table 1).
Figure 2. (a) Annual mean of the Bio-Bio river runoff (m3 s±1) (1975±94), (b) annual mean of the Itata river runoff (m3 s±1)(1975±91), (c) annual RIVSUM (m3 s±1) (1977±91), and (d) historical intra-annual variability of the Bio-Bio (m) and Itata (n)river runoff (m3 s±1) based on a record of 20 and 17 years, respectively. Bars indicate standard error. Raw time series (d),smoothed time series (s).
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
316 R. A. Quin~ones and R. M. Montes
The annual landings of RoÂbalo were signi®cantlycorrelated with annual mean rainfall with a 4-year lagtime (r � )0.75; Table 1). On a monthly basis, thecumulative rainfall during March was positivelycorrelated to annual RoÂbalo landings with a lag timeof 4 years (r � 0.78; Table 1).
The linear regression models between annualRoÂbalo landings and annual mean freshwater input tothe coastal zone (Figs 8a,b and 9; Table 2) explainbetween 56% and 88% of landing variance. Thedetermination coef®cient between the annual meanItata runoff and annual RoÂbalo landings was partic-ularly high (0.88).
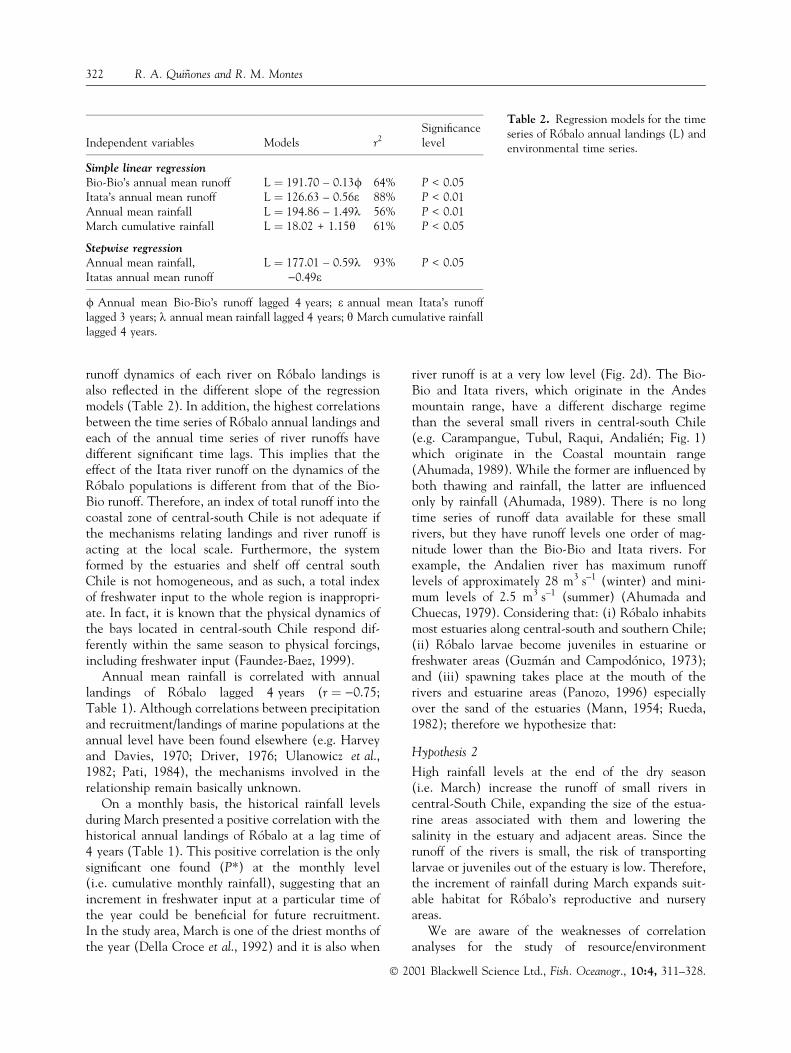
The variations in Bio-Bio annual mean runofflagged 4 years, annual mean rainfall lagged 4 years,and Itata annual mean runoff lagged 3 years wereincorporated into a stepwise regression model. Onlythe latter two remain signi®cant (P < 0.05), explain-ing 93% of the observed variance in RoÂbalo landings(Table 2).
DISCUSSION
Our results show that landings of the benthic-demersal®sh RoÂbalo are signi®cantly correlated to freshwaterinput to the coastal zone in central-south Chile. Thisis the ®rst time that an association between ®shlandings and freshwater input is demonstrated in theSouth-eastern Paci®c.
High correlation coef®cients between annual meanenvironmental (i.e. river runoff and rainfall) timeseries and annual landings time series occur with a lagtime of 3±4 years. This period corresponds to the ageat which RoÂbalo reaches the minimum exploitationsize (i.e. 30 cm; IFOP, 1980) according to the growthrate estimate of Gosztonyi (1974). Considering: (i) thehigh correlation coef®cients found between annualRoÂbalo landings and annual mean freshwater input;(ii) the agreement found between the time needed forthe RoÂbalo to recruit to the ®shery and the time lag atwhich the highest correlation coef®cients wereobserved between the freshwater input time series andthe time series of RoÂbalo landings; and (iii) the
Figure 3. (a) Relationship between annual mean runoff ofItata and Bio-Bio rivers for the period 1975±91. Years 1991and 1982 correspond to El NinÄo events. (b) Relationshipbetween monthly mean runoff of Itata and Bio-Bio rivers forthe period 1975±91.
Figure 4. (a) Annual mean rainfall (mm) during the period1975±94. Raw time series (d), smoothed time series (s).(b) Monthly historical signal of rainfall based on a 20-yearrecord. Bars indicate standard error.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 317
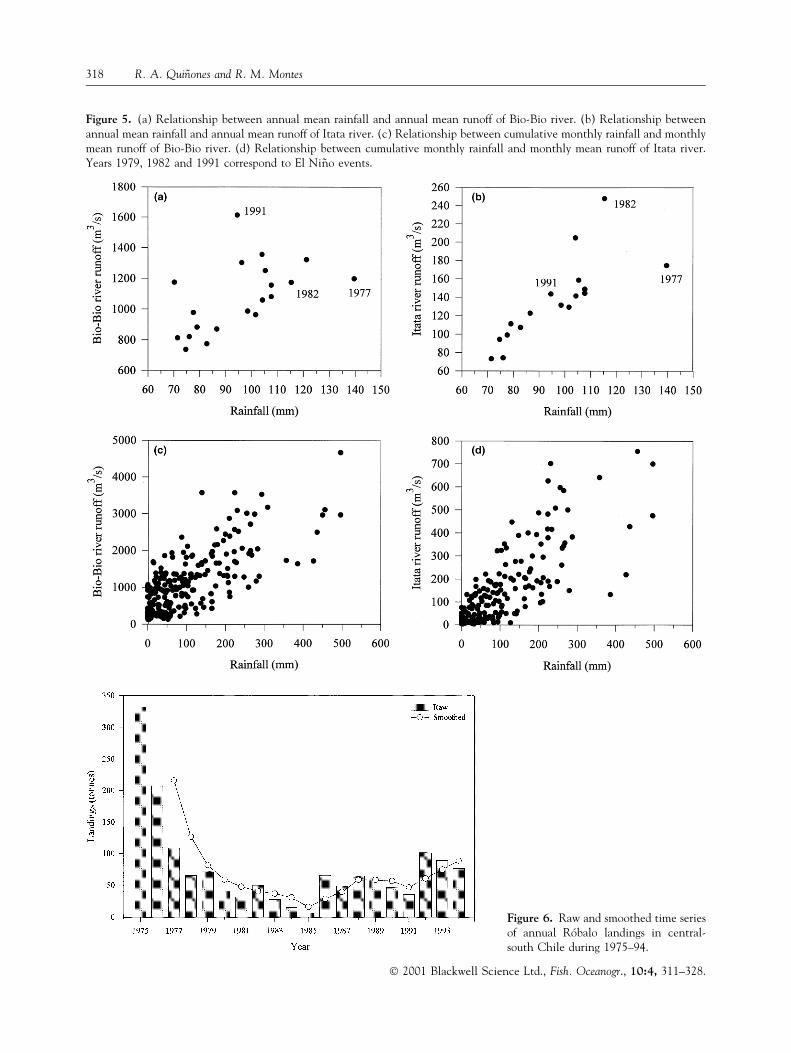
Figure 5. (a) Relationship between annual mean rainfall and annual mean runoff of Bio-Bio river. (b) Relationship betweenannual mean rainfall and annual mean runoff of Itata river. (c) Relationship between cumulative monthly rainfall and monthlymean runoff of Bio-Bio river. (d) Relationship between cumulative monthly rainfall and monthly mean runoff of Itata river.Years 1979, 1982 and 1991 correspond to El Ni~no events.
Figure 6. Raw and smoothed time seriesof annual RoÂbalo landings in central-south Chile during 1975±94.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
318 R. A. Quin~ones and R. M. Montes
importance of freshwater and estuarine dynamics forthe life cycle of the RoÂbalo (i.e. spawning, nurseryarea; Trujillo, 1972; GuzmaÂn and CampodoÂnico, 1973;PequenÄo, 1979, 1981; Ruiz, 1993; Panozo, 1996); then,the following hypothesis is proposed:
Hypothesis 1
River runoff and rainfall strongly in¯uence the survi-val rate of RoÂbalo's early stages during the ®rst year oflife.
The negative sign of the correlation coef®cientsrelating RoÂbalo's annual landing and annual meanriver discharge from both rivers, and the evidencedescribed in the literature about the effect of fresh-
water input on early survival of planktonic stagessuggest at least two possible mechanisms supportingthe rationale of hypothesis 1:
(i) The high transport offshore produced by highlevels of river runoff may be detrimental for earlystages of RoÂbalo. Unfortunately, there is noinformation regarding the behaviour of RoÂbalo'slarvae and eggs in the water column. However, thenegative effect of offshore transport has beenshown for larvae and eggs of other ®sh and inver-tebrate species elsewhere (e.g. Johnson and Gonor,1982; Crecco and Savoy, 1987; Boehlert andMundy, 1988; Epifanio, 1988).
Table 1. Summary of Pearsoncorrelation coef®cients (r) betweenannual RoÂbalo's landings in central-south Chile and environmental vari-ables.
MAnnuallag P* M
Annuallag P*
Bio-Bio river r RIVSUM r2 4 )0.62 > 0.1 2 2 )0.63 > 0.15 3 )0.61 > 0.05 2 3 )0.70 > 0.055 4 )0.68 > 0.05 5 3 )0.59 > 0.057 3 )0.52 > 0.1 8 4 )0.77 > 0.057 4 )0.54 > 0.1 8 5 )0.70 > 0.17 5 )0.64 > 0.1 9 1 )0.69 > 0.112 1 0.70 > 0.05 9 2 )0.75 > 0.112 2 0.54 > 0.1 9 3 )0.70 > 0.1A 3 )0.56 > 0.1 11 4 0.74 > 0.05A 4 )0.80 < 0.05 12 1 0.65 > 0.1A 5 )0.65 > 0.1 12 2 0.61 > 0.1
A 3 )0.76 > 0.05Itata river Rainfall1 3 )0.62 > 0.1 1 1 )0.50 > 0.051 4 )0.73 > 0.05 1 2 )0.77 > 0.052 3 )0.75 > 0.05 1 3 )0.77 > 0.052 4 )0.76 > 0.05 1 4 )0.58 > 0.053 3 )0.69 > 0.1 2 5 )0.60 > 0.053 4 )0.74 > 0.05 3 2 0.53 > 0.054 3 )0.62 > 0.1 3 3 0.74 > 0.054 4 )0.70 > 0.05 3 4 0.78 < 0.055 3 )0.72 > 0.05 3 5 0.87 > 0.055 4 )0.61 > 0.05 4 4 )0.64 > 0.056 2 )0.56 > 0.05 5 3 )0.71 > 0.056 3 )0.58 > 0.05 5 4 )0.70 > 0.058 3 )0.69 > 0.1 9 3 )0.53 > 0.058 4 )0.78 > 0.05 A 3 )0.74 > 0.059 2 )0.70 > 0.1 A 4 )0.75 = 0.059 3 )0.75 > 0.1 A 5 )0.67 > 0.0512 5 )0.70 > 0.1A 2 )0.64 > 0.1A 3 )0.94 < 0.051
M, monthly mean runoff or cumulative monthly rainfall, (A) annual mean runoffor annual mean rainfall. Numbers under M indicate speci®c months of the year.P* � signi®cance level accounting for serial autocorrelation. 1P < 0.01. Values inbold correspond to a signi®cance level of P* £ 0.05.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 319
(ii) High levels of river runoff produce indirect effectson the productivity and/or trophic web of theneritic areas of the continental shelf off central-south Chile that may be detrimental for RoÂbalo'searly stages. In other words, the effects of the riverson primary production, phytoplankton abundanceand succession, secondary production, and changesin species distributions (e.g. Neu, 1975, 1976;Mitchel, 1976; Skreslet, 1976; Funicelli, 1984;Levasseur et al., 1984; Therriault and Levasseur,1985; Brandt et al., 1986; Smetacek, 1986; Mannand Lazier, 1991) can promote the incrementor decrement of different species in the trophicweb, favouring differentially the presence orabsence of RoÂbalos's prey and predators.
Although there is no information about the toler-ance levels of early stages of RoÂbalo to low salinity, itis extremely unlikely that this euryhaline, eurythermicspecies (PequenÄo, 1989) living commonly near themouth of rivers has low tolerance to low salinity. In
fact, RoÂbalo uses estuaries as breeding areas (Fischer,1963; PequenÄo, 1981). However, the effect of salinityon the reproductive physiology of the RoÂbalo remainsunknown. The necessary information and/or data totest hypothesis 1 is presently not available, accord-ingly, further research on the subject is needed.
When analysing the relationship between annuallandings and annual RIVSUM, no signi®cant corre-lation was found at P* level. Since the single annualmean discharge of each river is correlated to RoÂbaloannual landings, one would expect that an associationshould exist between annual RIVSUM and annualRoÂbalo's landings. Nevertheless, the lack of relation-ship re¯ects the differences observed between thehistorical time series of runoff of both rivers (Fig. 2a,b).Indeed, the correlation coef®cient between the his-torical series (1975±91) of the Bio-Bio and Itata riversis not particularly high on an annual scale (0.67;P < 0.01) and there is important scatter in therelationship (Fig. 3a). The dissimilar effect of the
Figure 7. (a) Time series of annualmean Itata's runoff and annual landingsof RoÂbalo lagged 3 years. (b) Time seriesof annual mean Bio-Bio runoff andannual landings of RoÂbalo lagged4 years. Each dot shown in the ®gurescorresponds to 3-year running means.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
320 R. A. Quin~ones and R. M. Montes
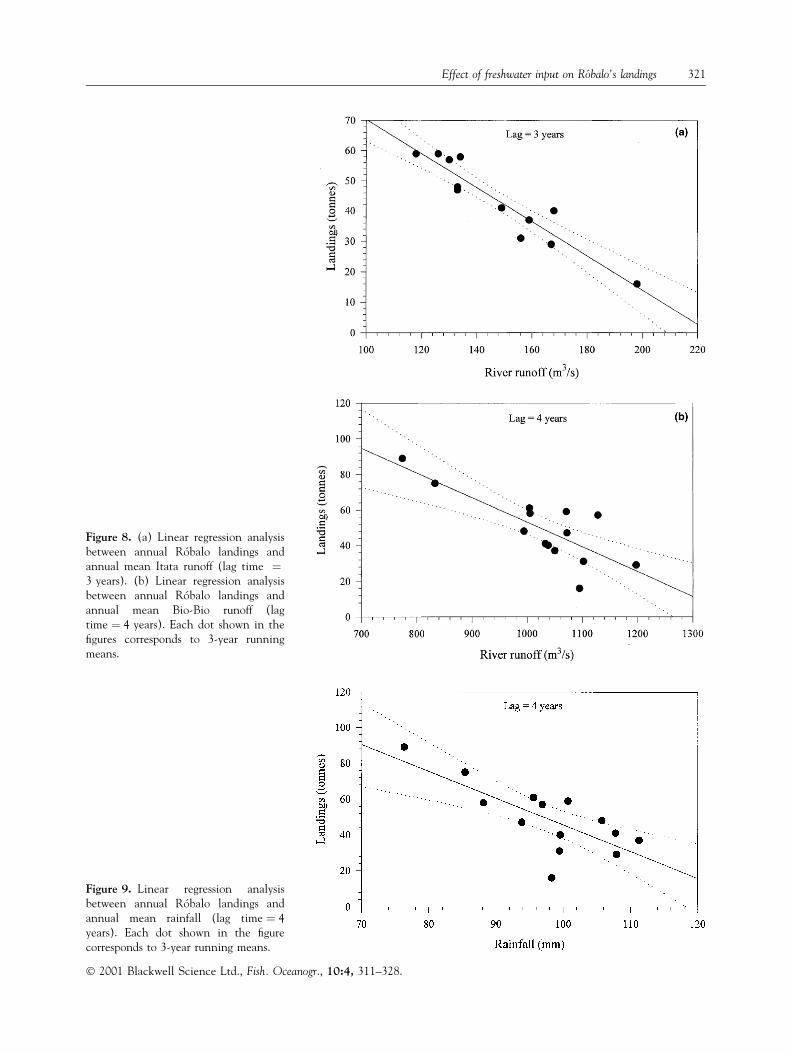
Figure 8. (a) Linear regression analysisbetween annual RoÂbalo landings andannual mean Itata runoff (lag time �3 years). (b) Linear regression analysisbetween annual RoÂbalo landings andannual mean Bio-Bio runoff (lagtime � 4 years). Each dot shown in the®gures corresponds to 3-year runningmeans.
Figure 9. Linear regression analysisbetween annual RoÂbalo landings andannual mean rainfall (lag time � 4years). Each dot shown in the ®gurecorresponds to 3-year running means.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 321
runoff dynamics of each river on RoÂbalo landings isalso re¯ected in the different slope of the regressionmodels (Table 2). In addition, the highest correlationsbetween the time series of RoÂbalo annual landings andeach of the annual time series of river runoffs havedifferent signi®cant time lags. This implies that theeffect of the Itata river runoff on the dynamics of theRoÂbalo populations is different from that of the Bio-Bio runoff. Therefore, an index of total runoff into thecoastal zone of central-south Chile is not adequate ifthe mechanisms relating landings and river runoff isacting at the local scale. Furthermore, the systemformed by the estuaries and shelf off central southChile is not homogeneous, and as such, a total indexof freshwater input to the whole region is inappropri-ate. In fact, it is known that the physical dynamics ofthe bays located in central-south Chile respond dif-ferently within the same season to physical forcings,including freshwater input (Faundez-Baez, 1999).
Annual mean rainfall is correlated with annuallandings of RoÂbalo lagged 4 years (r � )0.75;Table 1). Although correlations between precipitationand recruitment/landings of marine populations at theannual level have been found elsewhere (e.g. Harveyand Davies, 1970; Driver, 1976; Ulanowicz et al.,1982; Pati, 1984), the mechanisms involved in therelationship remain basically unknown.
On a monthly basis, the historical rainfall levelsduring March presented a positive correlation with thehistorical annual landings of RoÂbalo at a lag time of4 years (Table 1). This positive correlation is the onlysigni®cant one found (P*) at the monthly level(i.e. cumulative monthly rainfall), suggesting that anincrement in freshwater input at a particular time ofthe year could be bene®cial for future recruitment.In the study area, March is one of the driest months ofthe year (Della Croce et al., 1992) and it is also when
river runoff is at a very low level (Fig. 2d). The Bio-Bio and Itata rivers, which originate in the Andesmountain range, have a different discharge regimethan the several small rivers in central-south Chile(e.g. Carampangue, Tubul, Raqui, AndalieÂn; Fig. 1)which originate in the Coastal mountain range(Ahumada, 1989). While the former are in¯uenced byboth thawing and rainfall, the latter are in¯uencedonly by rainfall (Ahumada, 1989). There is no longtime series of runoff data available for these smallrivers, but they have runoff levels one order of mag-nitude lower than the Bio-Bio and Itata rivers. Forexample, the Andalien river has maximum runofflevels of approximately 28 m3 s±1 (winter) and mini-mum levels of 2.5 m3 s±1 (summer) (Ahumada andChuecas, 1979). Considering that: (i) RoÂbalo inhabitsmost estuaries along central-south and southern Chile;(ii) RoÂbalo larvae become juveniles in estuarine orfreshwater areas (GuzmaÂn and CampodoÂnico, 1973);and (iii) spawning takes place at the mouth of therivers and estuarine areas (Panozo, 1996) especiallyover the sand of the estuaries (Mann, 1954; Rueda,1982); therefore we hypothesize that:
Hypothesis 2
High rainfall levels at the end of the dry season(i.e. March) increase the runoff of small rivers incentral-South Chile, expanding the size of the estua-rine areas associated with them and lowering thesalinity in the estuary and adjacent areas. Since therunoff of the rivers is small, the risk of transportinglarvae or juveniles out of the estuary is low. Therefore,the increment of rainfall during March expands suit-able habitat for RoÂbalo's reproductive and nurseryareas.
We are aware of the weaknesses of correlationanalyses for the study of resource/environment
Independent variables Models r2Signi®cancelevel
Simple linear regressionBio-Bio's annual mean runoff L � 191.70 ± 0.13/ 64% P < 0.05Itata's annual mean runoff L � 126.63 ± 0.56e 88% P < 0.01Annual mean rainfall L � 194.86 ± 1.49k 56% P < 0.01March cumulative rainfall L � 18.02 + 1.15h 61% P < 0.05
Stepwise regressionAnnual mean rainfall,Itatas annual mean runoff
L � 177.01 ± 0.59k)0.49e
93% P < 0.05
/ Annual mean Bio-Bio's runoff lagged 4 years; e annual mean Itata's runofflagged 3 years; k annual mean rainfall lagged 4 years; h March cumulative rainfalllagged 4 years.
Table 2. Regression models for the timeseries of RoÂbalo annual landings (L) andenvironmental time series.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
322 R. A. Quin~ones and R. M. Montes
interactions. In fact, correlation studies have beensubjected to strong criticism by several authors(e.g. Walters and Collie, 1988; Bakun, 1996). Nev-ertheless, when correlation studies are conductedproperly, they can be instrumental in elucidatinginteractions previously unknown and generating newworking hypotheses. It is well known that correlationanalysis does not demonstrate cause/effect relation-ships and, in addition, the variability observed in
populations and communities is usually of a multi-factorial nature. Therefore, the fact that a singlefactor determines population abundance during aperiod of time (e.g. Drinkwater, 1987) does not avoidthe obvious possibility that another factor or a groupof factors could alter its abundance in another period(Frank et al., 1994). For example, the lobster ®sherycatch of the Gulf of Saint Lawrence was clearlyassociated with lagged river discharge for a period of40 years (Drinkwater et al., 1991). However, since1984 the river-derived estimates of lobster catch hasfailed, and several factors have been suggested asregulating the lobster population: modi®cation in thegeographical distribution of the resource, ®shingeffort changes, age at recruitment alterations, andstorm incidence (Drinkwater et al., 1991). It is stillnot clear why the river discharge±lobster laggedrelationship failed, but it is remarkable that only oneenvironmental factor could explain most of thevariance of lobster catch for so many years. In fact,this simple environment±resource relationshipworked better than predictions based on extrapola-tion of previous catch data (Drinkwater and Myers,1987). In summary, correlation analysis can be veryuseful for studying environmental in¯uence onresource abundance and/or availability, provided, aswith any other methodology, that its limitations arerigorously taken into account. When mechanismsunderlying a statistically signi®cant correlationbetween environmental conditions and speciesabundance are not fully understood, the relationshipfalls in the category of nonexplanatory prediction,
Table 4. Pearson correlation coef®cients between annualRoÂbalo landings and cumulative monthly rainfall (M) andannual mean rainfall (A). M represents speci®c months ofthe year.
Rainfall
M L1 L2 L3 L4 L5
1 )0.50 )0.77 )0.77 )0.58 )0.262 )0.30 )0.27 )0.51 )0.52 )0.603 0.05 0.53 0.74 0.78 0.874 0.1 0.02 )0.43 )0.64 )0.315 )0.14 )0.34 )0.71 )0.70 )0.416 0.42 0.00 )0.15 0.06 )0.027 0.13 )0.16 )0.16 )0.12 )0.168 )0.01 )0.09 )0.09 0.13 0.299 )0.41 )0.34 )0.53 )0.25 )0.2910 0.36 0.19 0.04 )0.03 )0.2411 0.25 0.30 0.05 )0.12 )0.1412 0.32 0.40 0.32 )0.16 )0.44A 0.24 )0.22 )0.74 )0.75 )0.67
Lag time L � 1¼5 years. Values in bold correspond to asigni®cance level of P < 0.05.
Table 3. Pearson correlation coef®cients between annual RoÂbalo landings and annual (A) and monthly (M) mean runoff ofBio-Bio river, Itata river and RIVSUM. M represents speci®c months of the year.
Bio-Bio river Itata river RIVSUM
M L1 L2 L3 L4 L5 L1 L2 L3 L4 L5 L1 L2 L3 L4 L5
1 0.22 0.20 )0.50 0.51 )0.30 )0.07 )0.47 )0.62 )0.73 )0.12 0.18 )0.16 )0.22 )0.21 0.312 0.23 0.30 )0.05 )0.54 0.15 )0.30 )0.53 )0.75 )0.76 )0.30 )0.34 )0.63 )0.70 )0.44 0.233 0.15 0.35 0.06 0.31 0.04 )0.31 )0.43 )0.69 )0.74 )0.46 )0.37 )0.37 )0.57 )0.55 )0.204 0.04 0.29 0.06 )0.20 0.01 )0.31 )0.37 )0.62 )0.70 )0.46 )0.31 )0.17 )0.30 )0.37 )0.185 0.09 0.20 )0.61 )0.68 )0.38 )0.42 )0.51 )0.72 )0.61 )0.14 )0.39 )0.43 )0.59 )0.50 )0.066 0.27 0.32 )0.13 0.33 0.04 )0.45 )0.56 )0.58 )0.08 0.42 )0.15 )0.05 )0.08 0.11 0.417 0.08 0.39 )0.52 )0.54 )0.64 0.01 )0.25 )0.34 0.08 0.05 0.00 )0.12 )0.15 0.06 )0.288 0.14 0.02 0.01 )0.04 0.12 )0.11 )0.30 )0.69 )0.78 )0.57 )0.16 )0.16 )0.45 )0.77 )0.709 0.32 0.04 )0.05 )0.14 )0.23 )0.48 )0.70 )0.75 )0.40 )0.22 )0.69 )0.75 )0.70 )0.31 )0.1710 0.39 0.26 0.07 )0.06 )0.22 0.12 )0.02 )0.02 0.41 0.30 0.38 0.40 0.46 0.54 0.1311 0.10 )0.06 )0.33 0.31 )0.39 0.32 0.54 0.56 0.45 )0.16 0.04 0.16 0.46 0.74 0.2812 0.70 0.54 0.10 )0.19 )0.36 0.43 0.45 0.11 )0.26 )0.70 0.65 0.61 0.37 0.27 )0.09A 0.15 0.025 )0.56 )0.80 )0.65 )0.35 )0.64 )0.94 )0.50 )0.09 )0.38 )0.48 )0.76 )0.48 )0.18
Lag time L � 1¼5 years. Values in bold correspond to a signi®cance level of P < 0.05.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 323

which is also a valid goal of science (as is prediction,explanation and nonpredictive explanation; Sayer,1984; Schumm, 1991).
The use of running means in the time series ofRoÂbalo landings is justi®ed by the fact that catch and/or abundance time series correspond to the sum overage classes which is a weighted sum over recruitment(Sutcliffe et al., 1977; Kope and Botsford, 1990; Parkeret al., 1995). Obviously, the use of running meansincreases the autocorrelation of the time series,diminishing the number of independent data pointsand degrees of freedom (Sutcliffe et al., 1977; Kopeand Botsford, 1990). We opted for the estimation ofthe level of signi®cance of the correlation as describedby Pyper and Peterman (1998), by taking into accountthe number of independent observations. In order toavoid spurious correlations we only accepted as validcorrelation coef®cients those with P* £ 0.05. How-ever, we report in Tables 3 and 4 all correlationcoef®cients obtained, as has been suggested by Waltersand Collie (1988) for studies on the relationshipbetween environmental factors and resources based oncorrelation analysis.
The continental shelf off central-south Chile is oneof the most productive ecosystems of the world, butthe mechanisms explaining this high productivity arestill poorly understood. Several oceanographic featuresare interacting on the shelf: (i) wind-driven upwelling(e.g. Ahumada and Chuecas, 1979; Kelly and Blanco,1984; Arcos and Navarro, 1986); (ii) topographicallyinduced upwelling by the shelf break and river sub-marine canyons (Sobarzo, 1999; Sobarzo et al., 2001);(iii) intrusion of low oxygen and high nutrient waters(Ahumada et al., 1983); (iv) presence of mesoscalephysical structures (e.g. ®laments, eddies, fronts;CaÂceres and Arcos, 1991; CaÂceres, 1992); (v) Kelvinwave-associated mixing events (Djurfeldt, 1989);(vi) topographic factors (CaÂceres and Arcos, 1991);(vii) re-suspension of particulate carbon and nitrogenfrom sediments (FarõÂas et al., 1994); (viii) tides(Blanco, 1984; Urrutia et al., 1993); (ix) freshwaterinputs (Djurfeldt, 1989; Sobarzo et al., 1993; Urrutia,1993); and (x) a high level of organic wastes fromurban areas and industry (Chuecas and Salamanca,1985; Ahumada et al., 1989).
Our research has highlighted the potentialimportance of freshwater input for neritic ®sh popu-lations in central-south Chile. Further research will benecessary to test the hypotheses emerging from thisinvestigation. Previously, Montes and QuinÄones(1999) had shown an association between freshwaterinput (Itata river runoff and rainfall) and landings oftwo clam populations (Venus antiqua and Protothaca
thaca) in the same study area. Our ®ndings, and thoseof Montes and QuinÄones (1999), emphasize the needto regard freshwater input as a potential factor indetermining the variability of other neritic populationsin central-south Chile. In addition, the associationfound between benthic/demersal populations andfreshwater in¯ow reinforces the hypothesis that thehigh biological productivity of the continental shelfoff central-south Chile is the outcome of severalsimultaneous factors.
ACKNOWLEDGEMENTS
We wish to thank the DireccioÂn General de AguasVIII RegioÂn and the DireccioÂn MeteoroloÂgica deChile for facilitating, respectively, the river runoff andrainfall data used in this study. This research wasfunded by the FONDAP Program on AdvancedStudies in the Humboldt Current System `Circulationand Physical±Biological Interactions in the HumboldtCurrent System (HCS) and their Impact uponRegional Biogeochemical Cycling' (FONDAP-Hum-boldt, CONICYT, Chile). We are very grateful toBernard A. Megrey, Michael M. Mullin and oneanonymous reviewer for their valuable comments andcriticisms to an earlier version of this manuscript.
REFERENCES
Ahumada, R. (1989) ProduccioÂn y destino de la biomasa ®to-planctoÂnica en un sistema de bahõÂas en Chile central: unahipoÂtesis. Biol. Pesq. (Chile) 18:53±66.
Ahumada, R. and Chuecas, L. (1979) Algunas caracterõÂsticashidrogra®cas de la Bahia de ConcepcioÂn (36°40¢S; 73°02¢W)y aÂreas adyacentes. Gayana Misc. 8:1±56.
Ahumada, R., Rudolph, A. and Martinez, V. (1983) Circulationand fertility of waters in ConcepcioÂn Bay. Est. Coast. Shelf.Sci. 16:95±105.
Ahumada, R., Rudolph, A., Madariaga, S. and Carrasco, F.(1989) DescripcioÂn de las condiciones oceanogra®cas de laBahõÂa San Vicente y antecedentes sobre los efectos de lacontaminacioÂn. Biol. Pesq. (Chile) 18:37±52.
Arcos, D. and Navarro, N. (1986) AnaÂlisis de un õÂndice desurgencia para la zona de Talcahuano, Chile (lat. 37°S).Invest. Pesq. (Chile) 33:91±98.
Arcos, D., NunÄez, S. and AcunÄa, A. (1996) Variabilidad depequenÄa escala en la zona neritica del sistema de surgenciade Talcahuano (Chile central): identi®cacioÂn y dinaÂmica deaÂeas de retencioÂn larval. Gayana. Oceanol. 4:21±58.
Arnstz, W.E. and Fahrbach, E. (1996) El NinÄo, ExperimentoClimaÂtico de la Naturaleza Causas FõÂsicas Efectos BioloÂgicos.Mexico: Fondo de Cultura EconoÂmica.
Bakun, A. (1996) Patterns in the Ocean. Ocean Processes andMarine Populations Dynamics. California Sea Grant CollegeSystem, La Jolla, CA, USA.
Balushkin, A.V. (1984) Morphological basis of the systematicsand phylogeny of the nototheniid ®shes. Proceedings Zoological
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
324 R. A. Quin~ones and R. M. Montes

Institute, Leningrad, 151:133 (in Russian; available in Eng-lish as Russian Translation Series No. 73, 1990 from A. A.Balkema, Rotterdam).
Baumann, M. (1998) The fallacy of the missing middle: physics® ¼ ® ®sheries. Fish. Oceanogr. 7:63±65.
Blanco, J.L. (1984) CaracterõÂsticas de la circulacioÂn sobre laplataforma continental de Talcahuano. Thesis, CatholicUniversity of ValparaõÂso, Chile.
Boehlert, G.W. and Mundy, B.C. (1988) Roles of behav-ioral and physical factors in larval and juvenile ®shrecruitment to estuarine nursery areas. Am. Fish. Soc.Symp. 3:51±67.
Box, G.E.P. and Jenkins, G.M. (1976) Time series analysis,forecasting and control. San Francisco: Holden-Day, Inc.
Brandt, A., Sarabun, C.C., Seliger, H.H. and Tyler, M.A.(1986) The effects of a broad spectrum of physical activityon the biological processes in the Chaseapeake Bay.In: Marine Interfaces Ecohydrodynamics. J.C.L. Nihoul (ed.).Amsterdam: Elsevier, pp. 361±384.
CaÂceres, M.M. (1992) VoÂrtices y ®lamentos observados enimaÂgenes de sateÂlite frente al aÂrea de surgencia de Tal-cahuano, Chile central. Invest. Pesq. (Chile) 37:55±66.
CaÂceres, M.M. and Arcos, D. (1991) Variabilidad en la estruc-tura espacio-temporal de un aÂrea de surgencia frente a lacosta de ConcepcioÂn, Chile. Invest. Pesq. (Chile) 36:27±38.
Calvo, J., Morriconi, E., Rae, G.A. and San Roman, N.A.(1992) Evidence of protandry in a subantarctic notothenid,Eleginops maclovinus (Cuv. & Val., 1830) from the BeagleChannel, Argentina. J. Fish Biol. 40:157±164.
Carrasco, P. (1988) Problemas de manejo de la cuenca del rõÂoBio-Bio: Uso, manejo y desarrollo de la hoya hidrogra®ca delrõÂo Bio-Bio. In: Programa Cuenca Del Bio-Bio. C. Murcia(ed.). Chile: Universidad de ConcepcioÂn, ed. ConcepcioÂn,pp. 41±46.
Castillo, G., MunÄoz, H., Gonzalez, H. and Bernal, P. (1991)Daily analysis of abundance and size variability of ®sh larvaein relation to oceanic water intrusions in coastal areas. Biol.Pesq. 20:21±35.
Castro, L., QuinÄones, R.A., Arancibia, H., Figueroa, D., Roa, R.,Sobarzo, M. and Retamal, M. (1997) Areas de desove deanchoveta y sardina comuÂn en la zona central. Informe ®nal,Fondo de InvestigacioÂn Pesquera, Subsecretaria de Pesca,Ministerio de EconomõÂa, ValparaõÂso, Chile, p. 114.
Chat®eld, C. (1989) The Analyses of Time Series. An Introduction,4th edn. London, UK: Chapman & Hall.
Chuecas, L. and Salamanca, M. (1985) Primer Informe `Pro-grama de vigilancia de contaminantes en BahõÂa ConcepcioÂn,VIII RegioÂn, Chile. In: Plan de AccioÂn para la ProteccioÂn delMedio Marino y aÂrera costera del Pacõ®co Sudeste. Chile:PNUMAS/CPPS, pp. 76.
Crecco, V. and Savoy, T. (1987) Effects of climatic and density-dependent factors on intra-annual mortality of larvalAmerican Shad. Am. Fish. Soc. Symp 2:69±81.
Cury, P. and Roy, C. (1989) Optimal environmental windowand pelagic ®sh recruitment success in upwelling areas. Can.J. Fish. Aquat. Sci. 46:670±680.
Cuvier, G. and Valenciennes, A. (1830) Histoire Naturelle DesPoissons, 5. ParõÂs: pp. 499.
Da Silva, J.A. (1986) River runoff and shrimp abundance in atropical coastal ecosystem ± the example of the Sofala Bank(Central Mozambique). In: The Role of Freshwater Out¯ow inCoastal Marine Ecosystems. NATO ASI Series, Vol. G7.S. Skreslet (ed.). Berlin: Springer-Verlag, pp. 329±344.
Daneri, G., Dellarossa, V., QuinÄones, R., Jacob, B., Montero, P.and Ulloa, O. (2000) Primary production and communityrespiration in the Humboldt Current System off Chile andassociated oceanic areas. Mar. Ecol. Prog. Series 197:41±49.
Della Croce, N., Parra, O., Stuardo, J., Arrizaga, A., Ahumada,R., Chong, J. and OyarzuÂn, C. (1992) El rõÂo BioBio y el maradyacente como unidad ambiental. In: Serie Monogra®caCientõ®ca. O. Parra and F. Faranda (eds). Proyecto EULA-Chile, Universidad de ConcepcioÂn, ConcepcioÂn, pp. 12±61.
Djurfeldt, L. (1989) Circulation and mixing in a coastalupwelling embayment; Gulf of Arauco, Chile, 1989. Cont.Shelf Res. 9:1003±1016.
Donn, T. (1987) Longshore distribution of Donax serra in twolog-spiral bays in the eastern Cape, South Africa. Mar. Ecol.Prog. Series 35:217±222.
Draper, N.R. and Smith, H. (1998) Applied Regression Analysis.Wiley Series in Probability and Statistics, 3rd edn. New York:John Wiley and Sons.
Drinkwater, K.F. (1986) On the role of freshwater out¯ow oncoastal marine ecosystems ± a workshop summary. S. Skreslet(ed.). Berlin: Springer-Verlag, pp. 429±438.
Drinkwater, K.F. (1987) Sutcliffe revisited. Previously publishedcorrelations between ®sh stocks and enviromental indicesand their recent performance, In: Enviromental EffectsRecruitment to Canadian Fish Stocks. R.I. Perry and K.T.Frank (eds). Can. Tech. Rep. Fish. Aquat. Sci. 1556:41±61.
Drinkwater, K.F. and Myers, R.A. (1987) Testing predictions ofmarine ®sh and shell®sh landings from environmental vari-ables. Can. J. Fish. Aquat. Sci. 44:1568±1573.
Drinkwater, K.F. and Frank, K. (1994) Effects of river regulationand diversion on marine ®sh and invertebrates. Aquat.Conservation: Freshwater Mar. Ecosys. 4:135±151.
Drinkwater, K.F., Harding, G.C., Vass, W.P. and Gauthier, D.(1991) The relationship of Quebec lobster landings tofreshwater runoff and wind storms. In: The Gulf of St. Law-rence: Small Ocean or Big Estuary? J.C. Therriault (ed.). Can.Spec. Publ. Fish. Aquat Sci. 113:179±187.
Driver, P.A. (1976) Prediction of ¯uctuations in the landings ofbrown shrimp (Crangon crangon) in the Lancashire andwestern sea ®sheries district. Est. Coast. Shelf Sci. 4:567±573.
Eastman, J.T. (1993) Antarctic ®sh biology. Evolution in aUnique Environment. New York: Academic Press.
Epifanio, C.E. (1988) Transport of invertebrate larvae betweenestuaries and the continental shelf. Am. Fish. Soc. Symp3:104±114.
EULA (1999) Estudio de impacto ambiental Proyecto ComplejoForestal Industrial Itata (Celulosa Arauco y ConstitucioÂn S.A.),Vol. 1±6. Chile: Centro Universitario Internacional Europa-AmeÂrica Latina de InvestigacioÂn y FormacioÂn de CienciasAmbientales EULA-Chile Center, Universidad de Concep-cioÂn.
FAO (1995) El estado mundial de la pesca y acuicultura. Roma:OrganizacioÂn de las Naciones Unidas para la Agricultura y laAlimentacioÂn.
FarõÂas, L., Salamanca, M.A. and Chuecas, L.A. (1994) Variaci-ones estacionalesdel ¯ujo de partõÂculas y contenido de materiaorgaÂnica en la interfase agua-sedimento en BahõÂa ConcepcioÂn,Chile central. Cienc. Tec. Mar., CONA 17:15±31.
Faundez-Baez, C.P. (1999) CaracterizacioÂn hidrogra®ca invernalde las BahõÂas ConcepcioÂn, San Vicente y Coronel (Chile,Centro-Sur). Thesis, Department of Oceanography, Uni-versity of ConcepcioÂn, Chile.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 325

Fischer, W. (1963) Die Fishe des Brackwassergebietes Lenga beiConcepcioÂn (Chile). Int. Revue Ges. Hydrobiol. 48:419±511.
Fossing, H., Gallardo, V.A., Joergensen, B.B., Huettel, M.,Nielsen, L.P., Schultz, H., Can®eld, D.E., Forster, S., Glud,R.N., Gundersen, J.K., Kuever, J., Ramsing, N.B., Teske, A.,Thamdrup, B. and Ulloa, O. (1995) Concentration andtransport of nitrate by the mat-forming sulphur bacteriumThioploca. Nature 374:713±715.
Frank, K.T., Drinkwater, K.F. and Page, F.H. (1994) Possiblecauses of recent trends and ¯uctuations in Scotian Shelf/Gulf of Maine cod stocks. ICES Mar. Sci. Symp198:110±120.
Funicelli, N.A. (1984) Assessing and managing effects ofreduced freshwater in¯ow to two Texas estuaries. In: TheEstuary as a Filter. V.S. Kennedy (ed.). pp. 435±446.
Gallardo, V.A. (1977) Large benthic microbial communities insulphide biota under Peru±Chile Subsurface Countercurrent.Nature 268:331±332.
Gammelsrod, T. (1992) Variation in shrimp abundance on theSofala Bank, Mozambique, and its relation to the Zambesiriver runoff. Est. Coast. Shelf Sci. 35:91±103.
Gargett, A.E. (1997) The optimal stability `window': a mech-anism underlying decadal ¯uctuations in North Paci®c sal-mon stocks?. Fish. Oceanogr. 6:109±117.
Garvine, R.W. and Monk, J.D. (1974) Frontal structure of ariver plume. J. Geophys. Res. 79:2251±2259.
Gibson, R.J. and Myers, R.A. (1988) In¯uence of seasonal riverdischarge on survival of juvenile Atlantic salmon SalmoSalar. Can. J. Fish. Aquat. Sci. 45:344±348.
Gosztonyi, A.E. (1974) Edad y crecimiento del `roÂbalo', Elegin-ops maclovinus (Osteichthyes, Nototheniidae) en aguas de larõÂa Deseado y sus adyacencias. Physis Secc. A. 33:1±8.
GuzmaÂn, L. and CampodoÂnico, I. (1973) Algunos aspectos de labiologõÂa de Eleginops maclovinus (Cuv. y Val.) 1830, conespecial referencia a su morfometrõÂa, caracteres merõÂsticos yalimentacioÂn. An. Inst. Patagonia 4:343±371.
Harvey, K.A. and Davies, R.M. (1970) Factors in¯uencingstanding crops and survival of juvenile salmon at BarrowsStream, Maine. Trans. Am. Fish. Soc. 99:297±311.
Hollowed, A.B., Bailey, K.M. and Wooster, W.S. (1987) Patternin recruitment of marine ®shes in the northeast Paci®cOcean. Biol. Oceanogr. 5:99±131.
IFOP (1980) CataÂlogo de Recursos Pesqueros, Chile. Santiago,Chile: CorporacioÂn de Fomento de la ProduccioÂn, Institutode Fomento Pesquero, Chile.
Johnson, G.E. and Gonor, J.J. (1982) The tidal exchange ofCallianassa californiensis (Crustacea, Decapoda) larvaebetween the ocean and the Salmo River Estuary, Oregon.Est. Coast. Shelf Sci. 14:501±516.
Johnston, I.A., Calvo, J., Guderley, H., FernaÂndez, D. andPalmer, L. (1998) Latitudinal variation in the abundanceand oxidative capacities of muscle mithocondria in perci-form ®shes. J. Exp. Biol. 201:1±12.
Kawasaki, T., Tanaka, S., Toba, Y. and Taniguchi, A., eds.(1991) Long Term Variability of Pelagic Fish Populations andTheir Environment. Oxford: Pergamon Press.
Kelly, R.J. and Blanco, J.G. (1984) Proceso de surgencia enPunta Nugurne, Chile (Lat. 36°S), marzo 1983. Invest. Pesq.(Chile) 31:89±94.
Kope, R.G. and Botsford, L.W. (1990) Determination of factorsaffecting recruitment of Chinook salmon Oncorhynchustshawytscha in Central California. Fish. Bull. 88:257±269.
Levasseur, M., Therriault, J.-C. and Legendre, L. (1984) Hier-archichal control of phytoplankton succesion by physicalfactors. Mar. Ecol. Prog., Series 19:211±222.
Mann, G. (1954) Vida de los Peces en Aguas Chilenas. Santiago,Chile: Ministerio de Agricultura-Universidad de Chile,Chile.
Mann, K.H. and Lazier, J.R.N. (1991) Dynamics of MarineEcosystems. Oxford: Blackwell Scienti®c Publications.
Mitchel, R. (1976) A possible relationship between rate of river¯ow and recruitment in an estuarine bivalve population.In: Fresh Water on the Sea. S. Skreslet, R. Leinebo, J.B.Matthews and E. Sakshaug (eds). Oslo: Association ofNorwegian Oceanographers, pp. 203±209.
Montes, R.M. and QuinÄones, R.A. (1999) Efecto de la pluvi-osidad y del caudal de los rõÂos Bio-Bio e Itata sobre losdesembarques de las almejas Protothaca thaca y Venus antiquaen la zona centro-sur de Chile. Rev. Chil. Hist. Nat. 72:13±30.
Mullin, M.M. (1993) Webs and scales. Physical and EcologicalProcesses in Marine Fish Recruitment. Washington: Washing-ton Sea Grant Program, University of Washington Press.
Neu, H.J.A. (1975) Runoff regulation for hydropower and itseffect on the ocean environment. Can. J. Civ. Eng. 2:583±591.
Neu, H.J.A. (1976) Runoff regulation for hydro-power and itseffect on the ocean environment. Hydrol. Sci. 21:433±444.
Nilo, P., Dumont, P. and Fortin, R. (1997) Climatic andhydrological determinants of year-class strenght ofSt. Lawrence River lake sturgeon (Acipenser fulvescens).Can. J. Fish. Aquat. Sci. 54:774±780.
Panozo, R. (1996) Evidencias del hermafroditismo secuencial enel RoÂbalo Eleginops maclovinus, Valenciennes 1830 (Perci-formes-Eleginopsidae). ConcepcioÂn: Thesis, Universidad deConcepcioÂn, ConcepcioÂn, Chile.
Parker, K.S., Royer, T.C. and Deriso, R.B. (1995) High-latitudeclimate forcing and tidal mixing by the 18.6-year lunar nodalcycle and low frequency recruitment trends in Paci®c halibut(Hippoglossus stenolepis). In: Climate Change and Northern FishPopulations. R.J. Beamish (ed.). Can. Spec. Publ. Fish. Aquat.Sci. 121:447±459.
Parra, O. (1996) El rõÂo Bio Bio: El recurso natural base deldesarrollo de la RegioÂn. Cuadernos del Bio Bio. ConcepcioÂn:Ediciones Universidad de ConcepcioÂn, Chile.
Parra, O. and Habit, E. (1998) Documento de SõÂntesis. Estudiode LõÂnea base para la evaluacioÂn de impacto ambiental delcomplejo forestal industrial Itata (Celulosa Arauco y Con-stitucioÂn S. A.). ConcepcioÂn: EULA Center, Universidad deConcepcioÂn, Chile.
Pati, S. (1984) Observations on the relation between rainfalland the inshore ®shery along the Orissa coast. J. Cons. Int.Explor. Mer. 41:145±148.
PequenÄo, G.H. (1979) Antecedentes alimentarios de Eleginopsmaclovinus (Valenciennes, 1830) (Teleostomi: Notothenii-dae), en Mehuin, Chile. Acta Zool. Lilloana 35:207±230.
PequenÄo, G. (1981) Peces de las riberas estuariales del rio Lin-gue. Chile. Biol. Mar. Roscoff 22:141±163.
PequenÄo, G. (1987) Aspectos antropogeÂnicos en la variacioÂnestacional de las pesquerõÂas artesanales de Valdivia.In: Manejo y Desarrollo Pesquero. P. Arana (ed.). ValparaõÂso,Chile: Escuela de Ciencias del Mar., Universidad CatoÂlica deValparaõÂso, pp. 183±194.
PequenÄo, G.H. (1989) The geographical distribution and taxo-nomic arrangement of South American notothenidae ®shes
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
326 R. A. Quin~ones and R. M. Montes

(Osteichthyes, Notothenidae). Bol. Soc. Biol. ConcepcioÂn,Chile 60:183±200.
Peterman, R.M., Pyper, B.J., Lapointe, M.F., Adkison, M.D. andWalters, C.J. (1998) Patterns of covariation in survival ratesof British Columbian and Alaskan sockeye salmon(Oncorhynchus nerka) stocks. Can. J. Fish. Aquat. Sci. 55:2503±2517.
Power, M., Dempson, J.B., Power, G. and Reist, J.D. (2000)Environmental in¯uences on an exploited anadromousArctic charr stock in Labrador. Fish Biol. 57:82±98.
Prager, M.H. and Hoenig, J.M. (1992) Can we determine thesigni®cance of key-event effects on a recruitment time series?A power study of superposed epoch analysis. Trans. Am.Fish. Soc. 121:123±131.
Pyper, B.J. and Peterman, R.M. (1998) Comparison of methodsto account for autocorrelation in correlation analyses of ®shdata. Can. J. Fish. Aquat. Sci. 55:2127±2140.
Quinn, II, T.J. and Niebauer, H.J. (1995) Relation of easternBering Sea walleye pollock (Theragra chalcogramma)recruitment to environmental and oceanographic variables.In: Climate Change and Northern Fish Populations. R.J. Beamish(ed.). Can. Spec. Publ. Fish. Aquat. Sci. 121:497±507.
Rojas, O.J., Mujica, A.R., Labra, M.C., Lederman, G.H. andMiles, H.U. (1983) EstimacioÂn de la abundancia relativa dehuevos y larvas de peces. ValparaõÂso, Chile: Instituto deFomento Pesquero, Chile, AP 83±31.
Rothschild, B.J., Smith, S.G. and Li, H. (1996) The applicationof time series analysis to ®sheries populations assessment andmodeling. In: Stock Assessment: Quantitative Methods andApplications for Small-Scale Fisheries. V.F. Gallucci, S.B. Saila,D.J. Gustafson and B.J. Rothschild (eds). Florida: CRC Press,pp. 354±402.
Rueda, T.R. (1982) ContribucioÂn al conocimiento de la biologõÂadel `RoÂbalo' (Eleginops maclovinus Cuv. y Val., 1830): Zonade los canales (XI RegioÂn). Thesis, Department of Ocean-ography, University of ConcepcioÂn, Chile.
Ruiz, V.H. (1993) Ictiofauna del RõÂo AndalieÂn (ConcepcioÂn,Chile). Gayana Zool. 57:109±278.
Runge, J.A., Castonguay, M., De Lafontaine, Y., Ringuette, M.and Beaulieu, J.L. (1999) Covariation in climate,zooplankton biomass, and mackerel recruitment in thesouthern Gulf of St. Lawrence. Fish. Oceanogr. 8:139±149.
Rutllant, J. (1997) Variabilidad interanual del clima en la costaoccidental arida de sudamerica: consideraciones para uncambio climaÂtico global. In: GestioÂn de Sistemas OceanograÂf-icos Del Pacõ®co Oriental. E. TarifenÄo (ed.). ComisioÂnOceanogra®ca Intergubernamental de la UNESCO, IOC/INF-1046, pp. 281±287.
Rutllant, J. and Fuenzalida, H. (1991) Synoptic aspects of cen-tral Chile rainfall variability associated with the SouthernOscillation. Int. J. Climatol. 11:63±76.
SantibanÄez, F. and Uribe, J. M. (1993) Atlas AgroclimaÂtico deChile de la VIII y IX Regiones. Santiago, Chile: Facultad deCiencias Agrarias y Forestales, Universidad de Chile, Min-isterio de Agricultura and CorporacioÂn de Fomento de laProduccioÂn, pp. 105.
Sayer, A. (1984) Method in Social Science. London: Hutchinson,pp. 271.
Schumm, S.A. (1991) To Interpret the Earth. Ten Ways to BeWrong. Cambridge: Cambridge University Press.
SERNAP (1975±94) Anuario EstadõÂstico de Pesca. Santiago,Chile: Servicio Nacional de Pesca (SERNAP), Ministerio deEconomõÂa, Fomento y ReconstruccioÂn.
SERNAPESCA (1995) Anuario Estadõ�stico de Pesca. Santiago,Chile: Servicio Nacional de Pesca (SERNAPESCA), Min-isterio de EconomõÂa, Fomento y ReconstruccioÂn.
Sharp, G.D. and Csirke, J., eds (1983) Proceedings of the ExpertConsultation to Examine Changes in Abundance and SpeciesComposition of Neritic Fish Resources. FAO Fish Report 291(3rd vol.), pp. 1066. Rome: FAO, United Nations.
Sheldon, R.W., Sutcliffe, W.H. Jr and Drinkwater, K.F. (1982)Fish production in multispecies ®sheries. In: MultispeciesApproaches to Fisheries Management Advice. M.C. Mercer,(ed.). Can. Spec. Publ. Fish. Aquat. Sci. 59, 28±34.
Skreslet, S. (1976) In¯uence of freshwater out¯ow from Norwayon recruitment to the stock of Arcto-Norwegian cod (GadusMorhua). In: Fresh Water on the Sea. S. Skreslet, R. Leinebo,J.B. Matthews and E. Sakshaug (eds). Oslo: The Associationof Norwegian Oceanographers.
Smetacek, V.S. (1986) Impact of freshwater discharge onproduction and transfer of materials in the marine environ-ment. In: The Role of Freshwater Out¯ow in Coastal MarineEcosystems. S. Skreslet (ed.). Berlin: Springer-Verlag,pp. 85±106.
Sobarzo, M. (1999) Surgencia costera sobre una platafor-ma continental limitada por canÄones submarinos, Concep-cioÂn, Chile central (36°40¢ S; 73°15¢ W). Doctorate Thesis,Department of Oceanography, Universidad de ConcepcioÂn,ConcepcioÂn, Chile.
Sobarzo, M., Sansone, E., De Maio, A., Arcos, D., Salamanca,M. and HenrõÂquez, J. (1993) OceanografõÂa FõÂsica del Golfode Arauco. Primera Parte: Variabilidad espacio temporal dela estructura hidrogra®ca de las aguas del Golfo de Arauco.In: Gestio n de los Recursos Hõ�dricos de la Cuenca del Rõ�o Bõ�oBõÂo y del Area Marina Costera Adyacente. F. Faranda andO. Parra (eds). ConcepcioÂn: Universidad de ConcepcioÂn,Serie MonografõÂas Cientõ®cas, EULA Centre, Vol. 4.
Sobarzo, M., Figueroa, D. and Djurfeldt, L. (2001) Upwelling ofsubsurface water into the rim of the BiobõÂo submarine can-yon as a response to surface winds. Cont. Shelf Res. 21:279±299.
Sutcliffe, W.H. Jr (1972) Some relations of land drainage,nutrients, particulate materials and ®sh catch in two easternCanadian bays. J. Fish. Res. Board Can. 29:357±362.
Sutcliffe, W.H. Jr (1973) Correlations between seasonal riverdischarge and local landings of American lobster (Homarusamericanus) and Atlantic halibut (Hippoglossus hippoglossus)in the Gulf of St. Lawrence. J. Fish. Res. Board Can. 30:856±859.
Sutcliffe, W.H. Jr, Loucks, R.H. and Drinkwater, K.F. (1976)Coastal circulation and physical oceanography of the Sco-tian Shelf and the Gulf of Maine. J. Fish. Res. Board Can.33:98±115.
Sutcliffe, W.H. Jr, Drinkwater, K. and Muir, B.S. (1977)Correlations of ®sh catch and enviromental factors in theGulf of Maine. J. Fish. Res. Board Can. 34:19±30.
Therriault, J.-C. and Levasseur, M. (1985) Control of phyto-plankton production in the lower St. Lawrence estuary: lightand freshwater runoff. Naturaliste Can. 112:77±96.
Trujillo, M. (1972) DistribucioÂn y abundancia de recursosdemersales capturados durante pescas exploratorias realizadasa lo largo de la costa chilena 1964±69. Bol. Inst. Fom. Pesq.Santiago 17:1±64.
Tyler, A.V. (1992) A context for recruitment correlations: whymarine ®sheries biologists should still look for them. Fish.Oceanogr. 1:97±107.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
Effect of freshwater input on Ro balo's landings 327

Ulanowicz, R.E., Ali, M.L., Vivian, A., Heinle, D.R., Richkus,W.A. and Summers, J.K. (1982) Identifying climatic factorsin¯uencing commercial ®sh and shell®sh landings in Mary-land. Fish. Bull. 80:611±619.
Urrutia, A.H. (1993) AnaÂlisis del comportamiento de la plumadel RõÂo Bio Bio en base a un estudio de sus campos de sal-inidad, temperatura, densidad y de velocidad. DoctorateThesis, EULA Center, Universidad de ConcepcioÂn, Con-cepcioÂn, Chile.
Urrutia, A., Figueroa, D. and Soto, L. (1993) Difusividades en lapluma del RõÂo BioBõÂo para un perõÂodo de alta descarga. Cien.Tec. Mar., CONA 16:21±32.
Walters, C.J. and Collie, J.S. (1988) Is research on environ-mental factors useful to ®sheries management? Can. J. Fish.Aquat. Sci. 45:1848±1854.
Welch, D.W. (1986) Identifying the stock-recruitment rela-tionship for age structured populations using time-invariantmatched linear ®lters. Can. J. Fish. Aquat. Sci. 43:108±123.
Wilber, D. (1992) Associations between freshwater in¯ows andoyster productivity in Apalachicola Bay, Florida. Est. Coast.Shelf Sci. 35:179±190.
Zheng, J. and Kruse, G.H. (2000) Recruitment patterns ofAlaskan Crabs in relation to decadal shifts in climate andphysical oceanography. ICES J. Mar. Sci. 57:438±451.
Ó 2001 Blackwell Science Ltd., Fish. Oceanogr., 10:4, 311±328.
328 R. A. Quin~ones and R. M. Montes
Related Documents











