REGULATORY ENZYMES OF FILARIAL PARASITES A DISSERTATION SUBMITTED TO THE ALIGARH MUSLIM UNIVERSITY, ALIGARH FOR THE DEGREE OF Master of Philosophtf IN Blochemfstrq BY HUNEZA HU5SAIN M. Sc. (Biochem.) DIVISION OF BIOCHEMISTRY CENTRAL DRUG RESEARCH INSTITUTE LUCKNOW-226001 December, 1988

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULATORY ENZYMES OF
FILARIAL PARASITES
A DISSERTATION SUBMITTED TO THE
ALIGARH MUSLIM UNIVERSITY, ALIGARH FOR THE DEGREE OF
Master of Philosophtf IN
Blochemfstrq
BY HUNEZA HU5SAIN
M. Sc. (Biochem.)
DIVISION OF BIOCHEMISTRY CENTRAL DRUG RESEARCH INSTITUTE
LUCKNOW-226001 December, 1988


DEDICATED
TO
APAJAN
AND
PAPAJI

Dr. s. N. ^mm EtiS^tU* Sct«ftti«^ H'-^^P ( Fvmef D»puiy timASt)
^ , /Vo.
CENTRAi DRUG RESEARCH INSTITUTE ( A Constftuont Establishment o( CSIfi)
Chattar Manzil, Post Box No. 173 LUCKNOW-226 001 ( I N D I A )
Date VQ-C. 29,19SS
CERTIFICATE
Thi^ 16 to (iQ.fiti{\q that the. wofik (imbodizd in thii, thd-il^i
eMt-iti&d "Regulators/ Enzifme.^ o^ FiiaJiiai Parasite.^" ha-6 been ca^^iad
out bij M-t4A Humza Hu4.Aain, M.Sc. [Biochzm] andzK mq ^ap^^vi^ion.
S/ie ha-i {sui{^illtd thz KzquiKe.rmnt6 oi thz . kliQaih Mu -f m
linivZK^^tij izqaKding thz pKZ^CKibzd pzfiiod o{) invz^t-tgationat wo fe
{\0>i thz avoaxd o^ H.Phil, dzgizz.
Thz woAk includzd in thii, thz-^i^ i-i, original unlzi6 -itatzd
uthzfiu)i.iz, and ha-^ not bzzn i>ubmittzd (s,oK anij othzn. dzgizz.
(S.W. GHATAIC)
,.^,„.^ ^ilut 1 aiM....9iW Tslssriim : CEWORUG Phone
Off. :32411-T8PABX Res. : 73490

ACKNOWLEVGEUENTS
It kai bo-zn my pfioad pfilvH^g^ io havz wo feed undzK tht lni,pi>i-
ing_guidance. _oi__my fL^^pgctzd tzacke.^, P^OI^ZMOH. A.M. SiddiQui, Hzad,
Vzpaitmznt Oj$ Bioch(iml-{>tfiy, MiQaih Muslim Unlve-fi^lty, Aliga^h, whoAZ
gzntfio-iity and anilagging intzud^t hai, hzZpdd me in achieving my
objZctiv&-i>.
I gmtzially acknouilQ-dgQ. Vi. S.N. Ghatak, Emznitai, Sciznti^t
0($ CSIR (J'ome.^ Hdad and VO-paty Vifizctofi), division o^ Biochzmi^tiy,
Czntn.al Vliig Rd^za^ck Jn^titutd (CVRI) iofi hii> kind i>apQ.>ivii>ion, untie
ing hzlp and valuablz ^uggz^tion^ in bfiinging up thd thu^i^ to iti
piz^iint ihapz.
I al-60 take, thii, opportunity to e.xpfie^^ mt/ de.ep ^zn^e o(, grati
tude. Vr(MA.^] hluzhat A. Kau^hal, Scizntiit, division 0)$ Biodi2.miUry
{^oK her untiring heZp, kind co-operation and vaiuabiz -i>ugge..6tion.6
(^or conducting my e.xperiment^ wihotLt which thi-i> dissertation would
not have been completed.
Words will not su^ice to e.xpre.ss my hzarty j ee- -cng-A and great-
{iUlne.ss to Vr. O.P. Shukla, Assistant Vire.ctor ior his valuable. he.lp
and suggestions and ^ruit^^ul criticism as well as interpretation oi
the data generated during the course o{^ these studie.s.
J am grateful to Vr. M.M. Vhar [Ex-Virzctor], Proizssor B.N.
Vhawan, director, CVRI and Vr. K.C. Saxzna, Head, Vivision o{^ Bio
chemistry ^or providing the neci-^bsary laboratory {facilities and to
the. Council o^ Scientific and Industrial Research, New Velhi ior {fin
ancial support.

I wouid like, to thank my collQ.agaQ.^, i,pzdially di. Bzchan Sha^ma,
Shalini, PooKnima, Hamta and {/ibha ion. tk2.lfi anc2.a-iing h^lp and co-
opziation. My ^-twce^e thanks to Shfii Shyam Sand^fi and Sii Uatadln
{,on. th^AK technical aMLitancz and SkAi K.L. Gupta ^ofi ^xcdlle^nt typing
oi my the.6lA.
HUNEZA HUSSAW

C O N T E N T S
Page No.
Abbreviat ions i
CHAPTER I
Introduction and Review of L i t e r a t u r e . . . , 1-17
CHAPTER II
Materials and Methods 18-25
CHAPTER III
Results and Discussion 26-61
CHAPTER IV
Summary and Conclusion. '. . 62-65
BIBLIOGRAPHY 66-72
* * * * *
• * •
*

A B B R E V I A T I O N S
A DP
BSA
DTT
GDP
LDH
MDH
I^ME
NAD
NADH
NADP
NADPH
OAA
PEP
PMSF
PFK
PK
SDH
Adenosine d iphospha t e
Bovine serum albumin
Di th io th r i to l
Guanidine d iphospha t e
Lactate dehydrogenase
Malate dehydrogenase
/ -mercaptoethanol
Nicotinamide adenine dinucleot ide
Reduced NAD
Nicotinamide adenine d inucleot ide
Reduced NADP
Oxaloacetate
Phosphoenol py ruva t e
Phenylmethyl su lphonyl f luor ide
Phosphofructokinase
Pyruva te kinase
Succinate dehydrogenase
phospha te

CHAPTER I
INTRODUCTION AND REVIEW OF LITERATURE

In recent years there has been a resurgence of interest
in research into various aspects of parasitic and other infectious
diseases with special emphasis on the search for new and improved
tools to combat those_ diseases for which the available techTiology
is inadequate. Helminthic infections like f i lar ias is , ascar iaasis ,
hookworm disease are the major health hazards to the mankind
in developing and under developed countries affecting millions of
human population. Although they do not cause acute mortality,
they sap the vi tal i ty of the nation already plagued by over popu
lation, food shortage, poor sanitation and lack of personal hygiene.
As compared to their hosts, these organisms generally have
primitive level of structural and biochemical refinement and some
of them survive under very selective ecological conditions, and
have to adapt thei r life cycle with the functioning of the a l te r
nate host system. They have developed intricate molecular mechan
isms for counteracting host defence system and to exploit their
metabolic machinery and regulatory molecules for thei r own survival .
F i la r ias i s , a disease caused by "thread like" worms of
the family Filaroidae of Phylum Nematoda is a chronic arthropod-
transmitted disease of great public health importance. Over 900
million people around the world are living in areas where the disease
is endemic. In India, the population exposed to the r isk of f i la r i
asis has increased from 25 million in 1955 to 304 million in 1982
and an estimated 22 million people are known to harbour circulating

microfilaria (mf) and another 16 million suffer from clinical mani
festations of filariasis (Ghatak et_ al^., 1987). Although the disease
is not fatal, it causes considerable morbidity, disability and social
stigma and the terminal stage of the disease is popularly known
as elephantiasis. The magnitude of suffering due to filariasis is
so enormous that world health organisation has included it in i ts
special programme of research and training for the control of tropical
diseases.
Filariasis is endemic all over India except few western,
northern and eastern s tates . It is prevalent in Assam, Kerala,
Andhra Pradesh, Uttar Pradesh, Madhya Pradesh and West Bengal.
Rapid industrialization and consequent migration of people has helped
in the spreading of the disease in areas where it did not exist
before. Hence filariasis has now been recognised as a disease
of national importance in this country.
Causative organisms
Several species of filarial parasi tes are known, each re la
tively specific for i ts host. Important pathogenic species of man
are Wuchereria bancrofti, Brugia malayi. Onchocerca volvulus and
Loa loa; while in animals and birds the disease is caused by
Setaria cervi (cat t le) , Dirofilaria immitis (dog), D.uniformis ( rabb i t ) ,
Litomosoides carinii (cotton r a t ) , Acanthocheilonema viteae (gerbil/
mastomys) and Chandlerella hawkingi (jungle crow). There are
three distinct phases in the development of f i larial parasites v i z . ,

microfilaria! (mf), infective larvae ( L , ) , and adult . The adult (male/
female) parasite resides in lymphatic/connective tissues and cavities
of the host and releases mf (sheath/unsheathed) in the peripheral
blood. These mf have a life span of 14-17 days and exhibit noctur
nal or diurnal periodici ty. The filarial parasite requires an al ter
nate host (mosquito/mite) for completing i t s life cycle. In India
Mansonella species of mosquito transmits B.malayi whereas Culex
quinquefasciatus spreads W.bancrofti. Adult worms have host speci
ficity and survive for 10-18 years . B.malayi and W.bancrofti are
lymph dwelling parasites whereas (Acanthocheilonema) and Onchocerca
species live in the tissues under the skin of the host .
Treatment of filariasis
Though filariasis was recognised as far back as 6th century
B.C. yet no effective chemical or immunological remedies could
be discovered for eradicating th is parasi t ic scourge. Few drugs
with limited efficacy and considerable side effects (sometimes fatal)
are available (Wang, 1982). Diethylcarbamazine (DEC) discovered
in mid forties, is essentially microfilaricidal and acts rather slowly
on adult worms. Hence it is not suitable for eradicating filarial
infections. Thus better drugs specially macrofilaricides are needed
for curing the disease. According to the existing l i terature some
compounds have been used as antifilarials l ike quinoline, metrifonate,
nitrofurantoin, levamisole, suramin, stibophen, DEC, centperazine,
flubendazole, mebendazole e tc .

The brief account presented above indicates the seriousness
of the disease and points towards making coordinated efforts to
control f i lar ias is . This could be achieved either by developing
an effective, nontoxic or least toxic chemotherapeutic agent which
can act both on microfilaria and adult worms or a suitable vaccine.
In either case it is necessary to have a sound knowledge of the
morphology, physiology and anatomy of the parasite especially i ts
biochemical machinery at the different life stages.
In recent years two main approaches have been made for
elucidating the mechanisms of action of anthelmintics. The first
approach is derived from the observation that several anthelmintics
have effect on neuromuscular system of the parasite leading to either
increase or decrease in their motil i ty. The other approach is
based on the knowledge that some compounds affect the energy generat
ing system of the parasite by selectively interfering with the funct
ional integrity of the parasite enzyme(s) or the uptake of carbo
hydrates (WHO, 1983). However, only meagre information is available
about the neuromuscular system as well as the energy metabolism
in filarial v/orms. Studies regarding these two systems may lead
to the development of specific drugs which can selectively inhibit
neurotransmission process or the enzymes of the parasite without
having adverse effect on the host .
The available information regarding the metabolic pattern
of filarial parasites in relation to drug action with special reference
to neurotransmission and energy metabolism published during the
last two decades are summarised below.

Biochemical analysis of filariids and their developmental stages
Glycoproteins have been identified as a structural component
of the cuticle in A.immitis, B.pahangi, and B.malayi (Cherlan^ et
al_., 1980; Furman and Ash, 1983; Kaushal et_ a l . , 1984). The sheath
present in the mf of W.bancrofti, Loa loa and B.malayi but absent
in A.perstans and 0.volvulus acts as a barr ier between the parasite
and the host defences. A better understanding of the chemistry
of filarial sheath would facilitate the development of procedure
for disruption of i ts biological function. Srivastava (1985) achieved
exsheathment of B.malayi mf by treatment with endopeptidases and
papain. Lectin binding studies revealed the presence of N-acetyl
glucosamine, glucose and mannose on the mature mf of B.pahangi,
while sheath of immature mf contain sialic acid, galactose, and
N-acetyl glucosamine (Furman and Ash, 1983).
Comparative chemical analysis of mf and adult have indicated
that dry matter accounts for 10 and 25% respectively of fresh body
weight in these two stages of S.cervi . The dry matter consisted
of carbohydrates, l i p ids , protein and traces of nucleic acids (Rathaur
et a l . , 1980). Filarial parasites do not store glycogen, the main
reserve food for providing energy under adverse conditions. Mf
of S.cervi contain very l i t t le glycogen as compared to adult, suggest
ing that this developmental stage of the parasite does not have
sufficient polysaccharide reserve (Rathaur et a l . , 1980). D.immitis
mf have, however, been reported to possess glycogen (Jaffe and
Doremus, 1970).

Reducing sugars account for the major portion of the total
carbohydrate in both mf (52%) and adult (67%) of S.cervi . Glucose
was the main constituent of the reducing sugars while concentration
of fructose was very low in adults and undetectable in mf (Rathaur
13 et a l . , 1980). Using C-glucose and NMR spectroscopy, Powell
et a l . (1986) showed the presence of trehalose in B.pahangi and
B.malayi. Glucose, galactose, mannose, myoinositol, ribose and
xylose were found in D.immitis (Ueda and Sawada, 1968), while
significant amounts of glucosamine and glucuronic acid were detected
both in adult and mf of S.cervi (Rathaur et_ al_., 1980).
Neuromuscular transmission
Neurotransmitter system is fairly well developed in various
helminths including filarial worms. They have underneath the cuticle
a single layer of muscles consisting of longitudinal f ibres . The
movement of these muscles are controlled by nervous system consist
ing of nerve f ibres , which operate through release of neurotrans
mit ters . The presence of a number of putative neurotransmitters
v i z . , catecholamines (norepinephrine, dopamine ) 5-hydroxytryptartiine
(5-HT) and acetylcholine with excitatory and inhibitory propert ies
have been established in filarial worms (Saxena et_ al^., 1977; Rathaur
et a l . , 1985; Singhal et_ a]_., 1975). In addition to thei r role in
neurotransmission and behavioural coordination these neurotransmitters
have been shown to be involved in regulating many metablic r e
actions like stimulation of phosphofructokinase and phosphorylase

by 5-HT resulting in enhancement of carbohydrate metabolism. Neuro
transmitters execute their physiological responses through specific
membrane bound receptors , the properties and significance of which
had been well documented in vertebrate t issues . Although thei r
presence have been speculated in filarial worms, no reports are
available regarding the nature of the receptors.
Energy metabolism
Considerable evidence has accumulated showing that carbo
hydrate is the primary source for the energy production in para
sitic helminths. These organisms assimilate carbohydrates, but
the processes involved in carbohydrate utilization are not identical
with those established in vertebrate tissues as none of these para
sites oxidise carbohydrate completely to C0_ and H_0. The in
complete oxidation leads to the accumulation and excretion of a
variety of part ial ly oxidised metabolic end products (Saz, 1970;
von Brand, 1973). The biological significance of incomplete oxidation
of carbohydrates is not properly understood. McManus (1986) while
reviewing part ial oxidation of carbohydrates reempKasized the out
standing feature of carbohydrate catabolism in these organisms v i z . ,
production of reduced organic end products even under aerobic condi
t ions.
Utilization of hexoses
The fi larial parasites derive their energy mainly through
carbohydrates and remain motile in a mineral medium fortified with

glucose or mannose. However, when the sugar concentration goes
down the worms start degrading their polysaccharide reserves for
remaining a l ive . The nature of the metabolic processes by which
filarial parasites obtain energy have been examined by several
investigators (von Brand, 1973). Incubation of L.carinii and S.cervi
adults with glucose, mannose, fructose or galactose produced lactic
and pyruvic acid. Glucose and mannose were utilized at a rate
much faster than that with galactose and fructose (Anwar et a l . ,
1975). L.carinii adult was found to be aerobic but l ike all other
parasit ic helminths it accumulates the end products of carbohydrate
utilization consisting of acetate or lactate and C0_ (Saz, 1981).
A.viteae and B.pahangi, however, differ markedly from L.carinii
as former two parasites survive for extended periods anaerobically
whereas L.carinii loses motility rapidly in absence of a i r . In
addition, isotope and aerobic fermentation balance studies indicate
that A.viteae and B.pahangi are homolactate fermenting anaerobes.
L.carinii on the other hand accumulate acetate in addition to lactate
and tr icarboxylic acid (TCA) cycle is not operative as an energy
generating pathway in this paras i te . Acomplete carbon balance
was not obtained in L.carinii and other products are presumed to
be present which must be identified before more definite statements
on i ts 0- requirement can be made (Dunn et_ a]^., 1988).
13 Recent studies using C glucose and NMR have detected
1-2% and 2-5% succinate among the fermentation products in B.pahangi

and A.viteae suggesting minor involvement of alternate pattern of
glucose catabolism (Powell et a l . , 1986).
Enzymes of carbohydrate metabolism
Evidence for the functioning of different metabolic pathways
in filarial worms has been adduced mainly by the demonstration
of the enzymatic steps or the identification of the intermediates
of the pathway. The carbohydrate metabolism of a few filarial
parasites (C.hawkingi, L.carinii and S.cervi) have been studied
in detail in this Institute. These filarial worms have been shown
to be equipped with many of the enzymes of anaerobic glycolysis,
PEP-succinate pathway and TCA cycle (Ghatak et al •, 1987). Distri
bution studies on m.any of the glycolytic enzymes and the demons
tration of phosphorylated glycolytic intermediates in filarial worms
clearly indicate the functioning of typical glycolytic sequence until
phosphoenol pyruvate (PEP) or pyruvate is reached (Barret, 1983;
McManus, 1986). The operation of a full glycolytic pathway for
conversion of glucose to lactic acid has been demonstrated
in C.hawkingi (Srivastava et_ a l . , 1968; Srivastava and Ghatak,
1971), L.carinii (Srivastava et_ al_. , 1970) and D.immitis (Hutchison
and McNeill, 1970). The enzymes of Embden Meyerhof scheme were
localized in the cytosol showing a resemblance with the mammalian
system. The activity of some of the key enzymes were higher
than the corresponding values reported for other f i l a r i ids .
Although most of the enzymes of the TCA cycle have been
documented in a few filarial worms (McNeill and Hutchison, 1971;

10
Agarwal et a l . , 1986; Dunn et_ a l . , 1988), the functioning of a true
TCA cycle in filarial parasi tes is st i l l a debatable point. Similarly
the presence of low or negligible activit ies of glucose 6-phosphate
dehydrogenase and 6-phosphogluconate dehydrogenase in S.cervi
(Anwar et_ aJ_., 1977), O.gutturosa (Dunn et_ a]_., 1988), C.hawking!
(Srivastava, 1969) cast a doubt about the functioning of a pentose
phosphate pathway in fi larial paras i tes . So it is presumed that
S.cervi mainly gains energy from the glycolytic pathway for i ts
survival . Presence of comparatively low activity of phosphofructo-
kinase (PFK) in S.cervi suggests that th is key enzyme may be play-,
ing a regulatory role in controlling the, operation of glycolytic path
way. Under aerobic conditions L.carinii produces acetate which
is derived from decarboxylation of pyruvate, probably via pyruvate
dehydrogenase. This enzyme is present in significant amount in
L.carinii but in lower amount in B.pahangi and A .viteae (Wang and
Saz, 1974; Meddulin and Saz, 1979).
The balance between pyruvate kinase (PK) and PEP-carboxy-
kinase (PEPCK) and their affinity for their common substrate (PEP)
are the factors that determine whether metabolic products are
channeled to succinate or to lactate (von Brand, 1973). S.cervi
which converts 25% of metabolised glucose to lactic acid and possesses
low level of PK and lactate dehydrogenase (LDH) but high activities
of PEPCK and malate dehydrogenase (MDH) has a functional PEP-
succinate pathway. On the other hand, typical lactic acid producers,
like C.hawkingi (Srivastava et a l . , 1968; Srivastava and Ghatak
1971) which convert 80-90% glucose into lactic acid, resemble

11
v e r t e b r a t e t i s sues in possess ing high l eve l s of PK and LDH and
low l eve l s of PEPCK and MDH. According to Bueding and Saz (1968)
PK/PEPCK ra t io g r e a t e r than one indicate tha t the metabolic pathway-
leads to lac ta te accumulation while the values l e s s than one suggest
the operat ion of C0_ fixing pathway and probable product ion of
succ ina te . Both succinate and lac ta te have been found to accumulate
in ra t io dependent on the age of the worms (Watts and F a i r b a i r n ,
1974) and the amount of 0- in the medium (von Brand, 1973). Both
mf and adu l t s of S . ce rv i have a PK/PEPCK ra t io l e s s than one (Anwar
et a l . , 1977; Rathaur et_ al_., 1982). However, in adul t D. immit is
(Braz ie r and Jaffe , 1973), C.hawkingi (Sr ivas tava and Ghatak, 1971)
and L . ca r in i i (S r ivas tava iet_ a l . , 1970a) the ra t io was h ighe r than
one, suppor t ing the p re fe ren t i a l formation of l a c t a t e . Among t h e
o the r carbon d iox ide fixing enzymes, malic enzyme seems to be
of some importance in a few he lmin ths . Malate en te r s the mi to
chondrion and undergoes a dismutation to pyruva te (under the influence
of NADP l inked malic enzyme) and to succinate (under the action
of fumarase and t h e subsequent reduction of fumarate by a fumarate
reduc tase c o m p l e x ) . The nature of terminal ox idase in f i l a r i a l
worms i s uncer ta in . F a i r l y normal looking c r i s t a t e mitochondria
have been r e p o r t e d in adu l t s of B.pahangi , L . ca r i n i i and D.immitif;
(Lee and Mi l l e r , 1969; Johnson and Bemrick, 1969; Middleton and
Saz, 1969). However, ne i t he r cytochrome-C nor cy tochrome-C-oxidase
could be de t ec t ed in adu l t s of L . c a r i n i i , B.pahangi or A .v i t eae
(Wang and Saz, 1974). In con t ras t , Rew and Saz (1977) de tec ted
cytochrome ox idase in mf of B .pahangi . S . ce rv i adu l t s contained

12
GUYCOGEH
GLUCOSE;
LACTATE
OXALOACETATE
NADH^
NAD MALATE
Id < a: U S D
• U . -HOH
FUMARATE
FADH,
SUCCINATE
Fig. 1: Phosphoenol pyruvate-succinate pathway in bovine f i larial
parasi te Setaria ce rv i .

13
cytochrome b , and in spite of several attempts presence of cytochrome
P.j.^ could not be detected in this parasite (Saxena e^ al_. in p re s s ) .
Lack of cytochrome P^cn in O.gibsoni and A, viteae was reported
by Mendis (1985).
Succinate is excreted in exceeding amounts under conditions
of low glucose availability (Powell et_ al_., 1986) and provided that
succinate formation is important or v i ta l , under these conditions
inhibition of PEP carboxykinase would provide a novel target for
filarial chemotherapy. PEPCK has been shown to be very active
and i ts properties have been determined in several paras i tes .
The enzyme occurs both in cytosol and mitochondria of l iver although
the distribution of the enzyme between these two cell compartments
varies from one species to another. Another important branch point
enzyme of PEP-succinate pathway is pyruvate kinase. Two types
of pyruvate kinase (L and M type) have been reported in mammals
as well as in paras i tes . L-type enzyme seems to be predominant
in fi larial worms - D.immitis and L.carini i . The other important
enzymes are fumarase, malic enzyme, lactate dehydrogenase, malate
dehydrogenase, succinate dehydrogenase and fumarate reductase (a
flavoprotein containing covalently bound FAD and iron sulphur groups).
(Fig. 1).
Metabolic targets for chemotherapeutic attack
The aim of specific chemotherapy is the removal of invading
organism without causing injury to the host. In order to achieve

14
t h i s , we must define the biochemical structure and metabolic pathways
of the parasite and its various developmental stages and synthesize
a selective agent which can knock off the parasite witKout showing
adverse effect on the host harbouring them. The identification
of sensitive molecular targets can provide a more rational approach
for the chemotherapy of f i lar ias is .
As mentioned ear l ier , most anthelmintic agents act directly
or indirectly by inhibiting ei ther neuromuscular transmission or
energy generation. The general sites of action of two well known
antifilarial compounds, DEC and suramin and antinematodal compound,
levamisole have been discussed by van den Bossche (1981).
DEC has been found to be an effective filaricide ir]_ vivo
for W.bancrofti, 0.volvulus, A. viteae and B.malayi. It acts p r i
marily as a microfilaricide, although some action against adults
of W.bancrofti, B.malayi (Otteson, 1984; Goodwin, 1984) and L
larvae of B.malayi (Ewert and Emerson, 1975) has also been demons
t ra ted . DEC, however, has no action against L_ larvae of
W.bancrofti (Jorden, 1958) and exhibi ts poor and impredictable
response in A. viteae and B.malayi infection in j i rds (Tanaka e^
a l . , 1981; Nogami et al_., 1984). In the studies by Langham et_
a l . (1980) all 0 . volvulus died at 0.1 mg per cent concentration
of DEC which is in the plasma concentration range of DEC found
in patients infected with onchocerca (Ree et_ a l . , 1977). Thus,
it appears that death of 0.volvulus mf may be related to an effect

15
on the neuromuscular sys tem. DEC also affects mot i l i ty in A .v i t eae
(Cavier et a l . , 1971) and Breinlia sergent i (Natarajan et a l . , 1973).
Levamisole at 100 ng/ml concentration completely s tops moti
l i t y of f i l a r i a l p a r a s i t e s such as L . c a r i n i i , A.vi teae and B.pahangi
(Wang and Saz, 1974; Rew and Saz, 1977). Levamisole ac ts by s t imu
lation of ganglion s t ruc tu res followed by a depolar i s ing type of
neuromuscular inhibi t ion (van Nueten, 1972; Coles et_ a l . , 1974).
Energy metabolism is a lso affected with decreased uti l izat ion of
glucose and shift to homolactate fermentation (Saz, 1981).
Suramin, an ant i t rypanosomal compound was found to act
upon 0 .vo lvu lus (van Hoof et_ al_., 1947). Due to the tendency of
suramin to combine with p r o t e i n s , i t i n h i b i t s enzymes involved
in energy metabolism, calcium t r a n s p o r t and var ious enzymes con
cerned with phospho ry l a t i on /dephosphory l a t i on r e a c t i o n s .
Centperazine , CDRI compound 72/70, levamisole and DEC
signif icant ly inh ib i t glucose u t i l iza t ion and syn the s i s of glycogen
in S .ce rv i (Anwar et_ a]_., 1978).
Avermectins are effect ive in des t roy ing mf of D. immit i s .
The drug b locks t ransmiss ion of command from vent ra l interneurons
to motor neurons in Ascaris and enhances Y-aminobutyr ic ac id (GABA)
re l ease and i t s binding to post synap t i c GABA recep to r (Pong et^
a l . , 1980; Pong and Wang, 1982).

16
SCOPE AND PLAN OF WORK
The information on the metabolic aspects of filarial parasites
presented above reveals fascinating mosaic of biochemical reactions
employed by the organisms for the i r survival and for their adaptat
ions to different hos ts . The filarial worms differ widely from
the respective hosts with regard to different metabolic reactions
and biochemical proper t ies . Further , the parasite differs consider
ably from each other reaffirming the suggestion of Saz (1981) that
each organism must be examined as a biochemical entity before
any reasonable understanding of i t s metabolism can be attained.
The filarial worms studied so far employ predominantly
anaerobic metabolism of carbohydrate as the major energy yielding
pathway and utilize limited quantity of 0^ (if avai lable) . However,
they do not possess the abi l i ty of bringing about complete oxidation
of substrates to C0_. Electron transport chain is rudimentary and
the parasites catalyse only limited terminal oxidation with generation
of l i t t le energy. Since PEP-succinate pathway has a crucial regu
latory function in channelling of carbon, regeneration of pyridine
nucleotides as well as generation of energy in filarial parasi tes ,
a study of the status of the enzymes of this pathway with special
reference to the effect of anthelmintic/antifilarial drugs would provide
an attractive area of research . The basic problem encountered
in studying f i lariasis is the non-availability of real human pathogen
(W.bancrofti/B.malayi) in sufficient amounts. S.cervi a bovine
filarial worm resembles human parasite in microfilarial periodicity,

17
antigenic pattern (Malhotra e^ a]_. , 1986; Kaushal e^ aj^., 1987) and
sensitivity towards known antif i lar ials . The parasite is available
in sufficient quantity^ from local slaughter house and hence it i s
ideal for conducting biochemical investigation.
Attempts have been made for studying the following aspects
in the present dissertat ion.
1. Subcellular localization of various enzymes of PEP-succinate
pathway and effect of ant if i lar ials .
2. Isolation and propert ies of PEP-carboxykinase from S.cervi .

CHAPTER II
MATERIALS AND METHODS

18
Biochemicals and enzymes
The biochemicals obtained from Sigma Chemical Company,
St . Louis , USA were ADP, BSA, DTT, GDP, L D H . ' M D H , ^-UE, OAk,
PMSF, PEP and sodium p y r u v a t e . Malic acid from E.Merck, West
Germany; sodium fumarate and sodium succinate from BDH, UK were
used for t h i s s t u d y . Sephadex G-25 and Blue Sepharose CL-6B
used for the pur i f icat ion s tud ies were procured from Pharmac ia ,
Uppsala , Sweden. NADH and NADP were suppl ied by CSIR Centre
for Biochemicals , Delhi , Ind ia . Among the anthelmint ics u sed ,
suramin was from Hoechst, AG, West Germany; levamisole was a
gift from Late Prof. G. Lammler, FRG and DEC c i t r a t e were p u r
chased from Cyanamide Ind ia . Pure base of DEC was p r e p a r e d
in Medicinal Chemis t ry Division of t h i s Inst i tute by Column Chromato-
'g raphy of DEC c i t r a t e . Centperazine and compound 72/70 were
supp l ied from Medicinal Chemis t ry Division of CDRI.
All o the r chemicals used were of ana ly t ica l grade and t r i p l e
g lass d i s t i l l e d water was used for a l l biochemical e x p e r i m e n t s .
Parasite
Motile adul t females (average length 6.0 ± 1.0 cm, average
weight 35 ± 6 mg) of Setar ia c e r v i , the bovine f i l a r i a l p a r a s i t e
were col lec ted from the per i tonea l cavi ty of f r e sh ly s l augh te red
, buffaloes (Bubalus buba l i s L inn) . The motile worms were brought
to the l a b o r a t o r y in Ringers solution (Singhal et aJ_, , 1973) wi th in

19
2 hours of s laughter ing from the local a b a t t o i r . The worms were
thoroughly washed with luke warm isotonic sa l ine for removing
t h e adher ing contaminants and used for enzymic s tud i e s within
2-3 hours of t h e i r removal from the c a d a v e r s , or immediately ch i l l ed
in ice and homogenized.
Preparation of medium
Ringers solution : 9 g of sodium c h l o r i d e , 420 mg of potassium
c h l o r i d e and 500 mg of glucose were weighed and d i s s o l v e d in mini
mum amount of w a t e r . After ra i s ing the volume to approx imate ly
900 ml , s e p a r a t e l y d i s so lved 250 mg sodium bicarbonate and 420
mg of calcium ch lo r ide were added and the volume made up to
one l i t r e .
Buffers; The buffers used for the s tudies were made with Tr i s
and t h e pH was adjusted with HCl ( l .ON). Sodium-potassium buffer
was made by adding the two sa l t s at such a concentrat ion tha t
r e q u i r e d pH was ob ta ined .
Preparation of homogenate
The homogenate of the f i l a r i a l pa r a s i t e Se tar ia c e r v i used
for t h e enzymic s tud ies was made in KCl (150 mM; 1:10 w / v ) .
F r e sh intact worms p rev ious ly ch i l l ed in ice were homogenised
using a high speed t ight f i t t ing Pot ter -Elvehjem homogeniser . The
homogenate t hus obtained was centrifuged under cold condit ions
at 800xg for 10 min at 12,000xg for 30 min and at 105,000xg for

20
60 min for the separat ion of cell d e b r i s , mi tochondr ia l , post mi to
chondr ia l and microsomal f rac t ions ,
I . Enzyme assay:
Details of the reaction mixtures used for the measurement
of PK, PEPCK, LDH, MDH, FRD, SDH, fumarase and malic enzyme
a r e d e s c r i b e d under each assay sy s t em. Spect rophotometr ic measure
ments were made in a final volume of 3 ml using a Beckman DU
spec t rophotometer or spec t ronic-21 UVD model with s i l i ca cuvet tes
of 1 cm l ight p a t h . All assay mix tures were p re incuba ted for
3 min at room tempera ture (28±2 C) before s t a r t ing the react ion
by t h e addi t ion of the s u b s t r a t e s . Change in absorbance at 340
nm was noted p r i o r to the addi t ion of the s u b s t r a t e s as well as
for t h e pe r iod of actual enzymic react ion following the addi t ion
of the s u b s t r a t e .
Pyruvate kinase (ATP, Pyruva te phospho t r ans fe ra se EC 2 . 7 . 1 . 4 0 ) :
Assay was based on Bucher and P f l e i d e r e r (1955) modified by the
use of Tr i s -buf fe r ins tead of Triethanolamine buffer . Assay mixture
contained: Tr i s HCl (pH 7.4) 200 mM; KCl, 100 mM; MgCl2, 10 mM;
ADP, 5 mM; PEP, 5 mM; LDH, 4 un i t s ; NADH-; 0,24 mM and enzymic
pro te in 100-200 ug.
Phosphoenol pyruvate carboxykinase (GTP: oxaloaceta te ca rboxy lase
(Phosphory la t ing ) EC 4.1.1 .32) was a s sayed according to Ward et^
a l . (1969) , as modified by the use of Tr i s -buf fe r (pH 6.5) ins tead
of T r i s buffer pH 7.4 200 mM, MnCl2lO mM, NaHCO^ 50 mM, GDP,

21
1 mM; PEP, 5 mM; MDH 4 uni t s ; NADH^ 0.24 mM and enzyme protein
20-50 ug.
Lactate dehydrogenase (L-lactate:NAD oxido r e d u c t a s e , EC 1.1.1.27)
-was assayed according to Kornberg (1955). Assay mixture contained
Tr i s HCl (pH 7.4) 200 mM; KCl, 100 mM; sodium p y r u v a t e , 5 mM,
NADH_ 0.24 mM and enzyme protein 5-15 ug.
Malate dehydrogenase (L-malate:NAD oxido r educ ta se EC 1.1.1.37)
was assayed according to Shonk and Boxer (1964) . Assay mixture
contained Tr i s HCl (pH 8.0) 200 mM; MgCl_ 10 mM; oxa loace ta te ,
10 mM, NADH_, 0.24 mM and enzyme prote in 0 .6-1 .5 ug.
Malic enzyme (L-mala te : NADP oxido r educ tase (deca rboxy la t ing)
EC 1.1.1.40) was assayed according to Ochoa (1955) . Assay mixture
contained Tr i s HCl (pH 7.4) 20D mM, MnCl- 10 mM; L malate 10
mM; NADP 0.24 mM and enzyme pro te in 0 .5-1 .5 mg.
Fumarase (L-malate hyd ro lyase EC 4 .2 .1 .2 ) was a s sayed according
to the method of Racker (1950). The react ion mixture consis ted
of potassium phospha te buffer (pH 7 . 4 ) , 150 mM; L-mala te , 30 mM
and enzyme protein 40-80 ug. Increase in abso rbancy was monitored
for 3 minutes at 240 nm.
Fumarate reductase (Succinate: ( accep to r ) oxido r e d u c t a s e , EC 1.3.99.1);
The a s say based on tha t d e s c r i b e d by P r i c h a r d (1973) was performed
with mitochondria l p repa ra t ions . - Reaction mix ture contained Tr i s
buffer (pH 7 . 2 ) , 100 mM; MgCl2, 10 mM; NADH, 0.3 mM; K" fumarate,

22
10 umol; CaCl^, 1.5 umol and enzyme protein 0.5-1.5 mg.
Succinate dehydrogenase (Succinate: (acceptor) oxido r e d u c t a s e ,
EC 1.3.99.1): It was assayed according to Metzger and Duwel (1973)
and was performed with mitochondria l p r e p a r a t i o n . The reactipn
mixture contained sodium potassium phospha te buffer (pH 7 . 4 ) ,
EDTA 1 mg/ml; 2,6 d ich lo ro phenol indophenol (Na DCIP 15 mg/100
ml ) ; sodium succinate (4.05 g/?5 ml) and enzyme prote in 0.5-1.5
mg. The decrease in ext inct ion was followed at 600 nm.
Defination of enzyme unit and speci f ic act iv i ty
One unit of enzyme a c t i v i t y i s the amount of enzyme r equ i r ed
to ca ta lyse the transformation of one umol subs t r a t e or the forma
tion of one umol product pe r minute under speci f ied exper imenta l
condi t ions . Specific a c t i v i t y i s unit/mg of enzyme p r o t e i n . The
extinction for oxidat ion or reduct ion of p y r i d i n e nucleot ides was
measured at 340 nm whi le increase in op t ica l dens i ty in fumarase
assay was r ecorded at 240 nm.
6 ? An ext inct ion coefficient of 6.22 xlO cm /mole (Horecker
and Kornberg, 1948) at 340 nm was used for NAD or NADP.
For fumarase the molar ext inct ion coefficient used was
3 -1 -1 2.44 X 10 x m M at 240 nm (Mahler et_ al_., 1958). For succinate
6 ? dehydrogenase the molar ext inct ion coefficient was 2.06x10 cm /molar
at 600 nm.
The following general equation was a p p l i e d for calculat ing
enzyme a c t i v i t y :

23
lU = OD/min X TRMV X DF Extinction coefficient
lU = Internat ional Unit
OD/min = Change in op t i ca l dens i ty /min
TRMV = Total react ion mixture volume (ml)
DF = Dilution factor of enzyme used
Protein estimation: Protein content in different subce l lu la r f rac t ions ,
column eluates (obta ined during pur i f ica t ion) was es t imated s p e c t r o -
photomet r ica l ly according to Layne (1957) or ca lo r imet r i ca l ly by
the method of Lowry et_ a l . (1951) using bovine serum albumin as
s t anda rd .
II. Purification of PEPCK
Adult females of S . ce rv i were homogenised in ice cold Tr i s
HCl buffer (50 mM pH 6 . 5 , 1:10 w / v ) containing DTT (1.5 mM) and
PMSF (0.1 mM) using a t igh t f i t t ing Po t te r Elvehjam homogeniser
for 2-j min. Centrifugation was done at 800xg for 10 min and at
12,000xg for 30 min. The post mi tochondr ia l supernatant thus o b t
ained was used for the pur i f ica t ion of PEPCK.
Step I: Ammonium sulphate fractionation
The supernatant from t h e p rev ious s t ep was brought to
301 saturat ion with r e spec t to ammonium su lpha te and the mixture
was gently s t i r r e d for 1 hou r . After centrifugation at l l ,000xg
for 30 min the p r e c i p i t a t e was d i s c a r d e d . To t h e supernatant

24
ammonium sulphate was added at constant stirring to 70% saturation.
The pH was adjusted to 7.0 with dilute NH_ after each addition
of (NH.)_SO. . The mixture was allowed to stand for 2 hours and
then centrifuged. The supernatant was discarded and the p rec ip i
tate was dissolved in a small volume of 50 mM Tris HCl containing'
1.5 mM DTT and 0.1. mM PMSF. The solution was further passed
through the Sephadex G-25 column.
Step II: Desalting on sephadex G-25 column
2 gm of Sephadex G-25 was soaked in about 500 ml of
double dis t i l led water and kept at boiling temperature for 2 hr
with intermittent gentle s t i r r ing. The upper layer of water contain
ing the fine part icles of Sephadex was decanted. Fresh volume
of dis t i l led water was added and the contents were left overnight
for complete soaking. The sephadex beads were equilibrated with
the suspension buffer and packed in a glass tube for making column
size of 11x1.4 cm. The protein fraction obtained after 70% ammonium
sulphate saturation was layered over this column and washed with
equilibration buffer, 3 ml eluates were collected at a flow rate
of 3 ml/5 min. The protein content of different eluates was moni
tored at 280 nm and the PEPCK rich fractions were pooled and
used for affinity chromatography.
Step III: Blue sepharose CL-6B affinity chromatography
1 g of Blue Sepharose CL-6B was soaked in dis t i l led water
for swelling of the beads . For the regeneration of Blue Sepharose

25
the gel was washed with regenerat ing buffers I and II followed
by washing with the equ i l ib ra t ing buffer of following composi t ion.
The regenera t ing buffers v i z . , Buffer I contained Tr i s HCl, pH
8.5 (100 mM)and NaCl (0 .5 M); whi le Buffer IT contained sodium
a c e t a t e , pH 4.5 (100 mM) and NaCl (0 .5 M). The equ i l i b r a t i ng
buffer , Buffer III contained Tr i s HCl, pH 6.5 (50 mM) , PMSF (0 .1
mM) and DTT (1.5 mM). These buffers were f r e sh ly p r e p a r e d . The
beads of Blue sepharose were washed a l t e rna t e ly with Buffer I ,
Buffer II and then again with Buffer I and f inal ly with Buffer I I I .
The washed beads were uniformly packed in a small g lass column
(5x0.5 cm) and desa l t ed fraction from Step I I , r i c h in PEPCK a c t i
v i t y , was s lowly l aye red over the column. Now equ i l ib ra t ion buffer
(Buffer III) was passed through the column for removing unadsorbed
p r o t e i n s . The dehydrogenases bound to Blue Sepharose were removed
by pass ing Buffer III containing NADH (1 mM) (equiva lent to 4 v o l
umes of loaded f r ac t ion ) . Then the adso rbed enzyme was e lu ted
from the column by passing Buffer III containing 0.5 M, 0.75 M,
1.0 M and 2.0 M NaCl in a s tepwise manner. The fraction conta in
ing maximum PEPCK ac t i v i t y was e luted with IM NaCL.

CHAPTER III
RESULTS AND DISCUSSION

26
The distribution pattern of the enzymes of PEP-succinate
pathway was analysed in different subcellular fractions v i z . , cel l-
free, post mitochondrial and post microsomal fractions of the bovine
filarial worm S.servi and is presented in Table I. Pyruvate kinase,
phosphoenolpyruvate carboxykinase, lactate dehydrogenase, malate
dehydrogenase, malic enzyme and fumarase were localized in 105,000xg
cytosolic fraction of the worm homogenate, while succinate dehydro
genase and fumarate reductase were found to be particulate bound.
Among the soluble enzymes, malate dehydrogenase was most active
exhibiting 313 units act ivi ty/g wet worms with specific activity
of 17.08. Significant levels of LDH (27 units/g wet worms) fumarase
(14.9 units/g wet worms) and PEPCK (15.0 units/g wet worms) were
observed while the levels of pyruvate kinase and malic enzyme
were very low. Decrease in the level of pyruvate kinase from
4.72 units (12,000xg) to 2.41 units (105,000xg) suggest the removal
of some factor from the cytosol required for enzyme act iv i ty .
The mitochondrial pellet (12,000xg sediment) showed the presence
of succinate dehydrogenase (4.07 units/g wet worms, specific act i
vity 0.067) and fumarate reductase (1.35 units/g wet worms; specific
activity 0.02).
Effect of antifilarials on different enzymes of PEP-succinate pathway
The action of a few antifi larial agents v i z . , DEC.centperazine
compound 72/70, levamisole and suramin on the different enzymes
of PEP-succinate pathway was studied by preincubating the enzyme

t/5
X o o o
o
60 X
o
C3
a 3
CO
X o o o
& 3
en
X o o 00
w
E N n
W
u >
to u —^ <Ti
be
< 3
> s
O >
> be •H - ^
o "
^^ c 3
a t
^ •
be
<
o >
at;
> w E o p O
< g S
00 CD
Q
i n
C
tt)
> D
O IS
^ o 1—1
' ^ IT)
O ^ CO O fS]
m oo
> 3 U >- O p . CO
' ^ rt 2 c <u ^ 2 >-a o 2 ^
(1< o
Q 2
sO 00
t n
o
OJ
00
o
00 t — '
o o
vD in t>
o O Z
Q 2
o
CO I—1
CO
O
o
a 2
o
CD (X)
O
OO
CD
I o u
Xi > <u
Ti
0)
rt in
rt (1) 1-^ bC
CD CD • i n i n
I O U
<D
0) OJ
2 bC
" 1 r J
0) E > N
c
o • H r—I
E 3
r--o
CO 1—1
o o
vO vO in
CD (-J 2
o 2
i n i n •^ CD O^ 1—' CD (NJ O
CM ro
I O
0)
c •H
(D .. w
o c 3 0)
to be
Q 2
Q 2
Q 2
Q Z
Q 2
Q 2
Q 2
O
O
(—I (NJ CD
CD
i n CO
Q 2
Q 2
i n o o
fO (NJ
eg r-j
3 Q)
O -t- "+-1 +J C O
bC g C c E o
• H nJ - H
27
(fl
(U
x:
^ 1
o (U
T3
« .5 ai 4)
(U +-' (fl 5H w "J -{^
!* C <" +^ 3 X) iJ
m 3 Ti
og (U X ! -(-> • <D
c X! 2 •
5 2 I S >2 (u -
Ti -2 c z i
r-H (ti O
!i£| ,—I C («
•H -H C
E ^ i J o S o 0)
^ c S >< c ^
(U 0)
— be + j u 6 0)
en (u P-o\o nJ W W CD >-^
& t; '^ >- . (13 J-
13 N U 3 <D C .2 ^ O C C ^ ^ -^ (U -C be u o (S E <i o
• ^ • ^
S E OJ OJ
w en
E
Tl (I) ^1 •H
3 CT (U V4
• 1 - 1
u (U U-
-t-i
o 3
TJ 0
^ 1
o
o
00 (fl • -t->
i-< U)
o
E o >- E N 3 C <U (U
C +H O
o o

28
prepara t ion with the t e s t compound for 15 min at room tempera ture
(28±2 C) in the d e s i r e d buffer and then determining the r e s idua l
ac t i v i t y in the usual way .
Pyruvate kinase: Among the f ive f i l a r i c ida l compounds screened
the enzyme was most affected by suramin exhib i t ing 52% inhib i t ion
at 1 yuM concentration (F ig . 2 ) . Higher concentration of the drug
(100 /iM) caused 76% i n h i b i t i o n . DEC, levamisole and centperazine
could reduce the a c t i v i t y of the enzyme to 50% at 10 jaM concentrat ion.
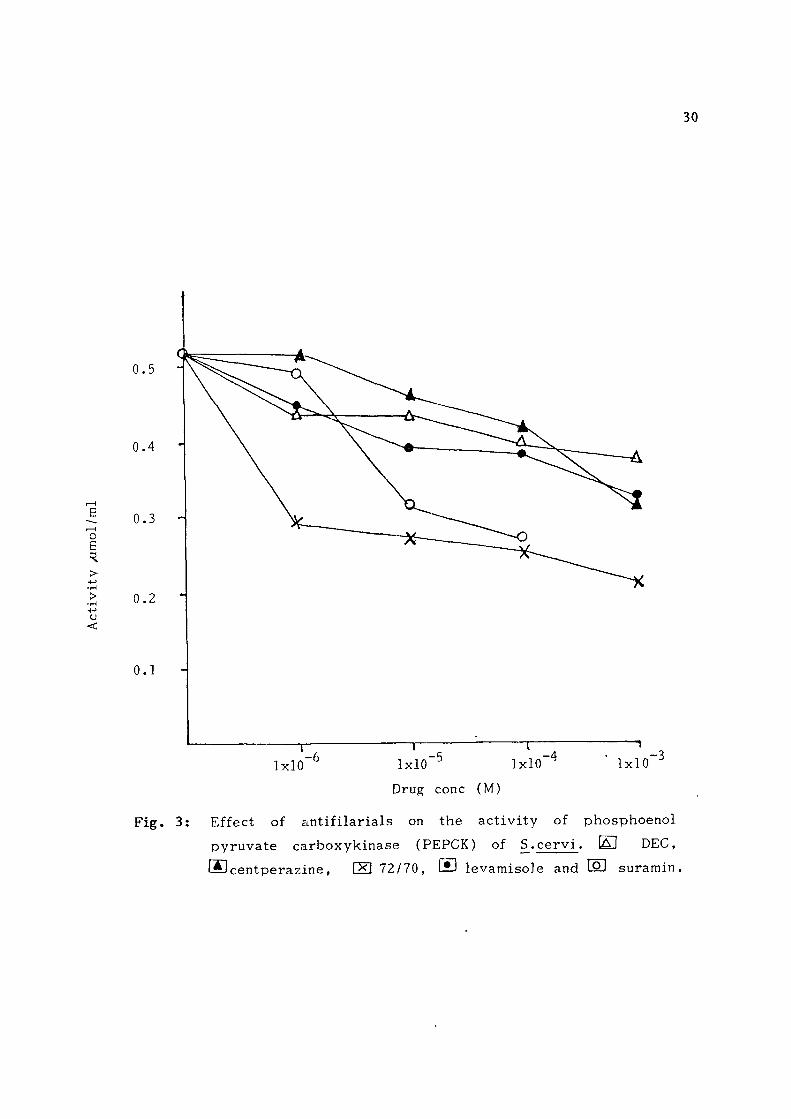
Phosphoenol pyruvate carboxykinase : CDRI compound 72/70 and s u r a
min were found to be more effect ive in reducing the ac t i v i t y of
PEPCK than t h e remaining t h r e e f i l a r i c i d a l compounds. The i n h i
bition by former two drugs was 45-50% at 100 /uM concentrat ion.
Compound 72/70 showed 41% inh ib i t ion o f the enzyme at 1 yuM con
centrat ion whi le l evamiso le , DEC and centperazine d id not show
much effect at 1/iM concentrat ion (F ig . 3 ) .
Lactate dehydrogenase: Suramin e x h i b i t e d maximum inh ib i t o ry action
lowering the LDH a c t i v i t y by 40% at 10 ^ M concentration while com
pound 72/70, l evamiso le , cen tperaz ine and DEC showed 33, 31, 29
and 27 percen t inh ib i t ion at 100 /aM concentration (F ig . 4 ) .
Malate dehydrogenase: Maximum inh ib i t o ry effect was obse rved
with suramin inh ib i t ing MDH a c t i v i t y by 33% at 10/uM concentrat ion,
while centperaz ine and levamiso le showed only 19% and 7% i n h i b i -

29
o e
0.3 -
0.2 -
0.1
1x10 -6 1x10 " 1x10
Drug cone. (M)
1x10 -3
Fig . 2: Effect of an t i f i l a r i a l s on the ac t i v i t y of py ruva t e k inase of S . ce rv i E DEC, ® cen tperaz ine , E ! 72/70, nri levamisole and [oj suramin.
30
o E
(J
<
0.5 -
0.4
0.3 -
0.2
0.1
1x10 -5 X
1x10 " 1x10
Drug cone (M)
-4 1x10
-3
Fig. 3: Effect of antifilarials on the activity of phosphoenol
pyruvate carboxykinase (PEPCK) of S.cervi . E ] DEC,
fflcentperaTiine, [ 3 72/70, S levamisole and E ) suramin.

31
1.0 -1
0.8
0.6 -
o g
u 0.4
0.2 -
1x10 1x10 1x10
Drug cone (M)
1x10
F i g . 4 : Effect of an t i f i l a r i a l s on the a c t i v i t y of l ac ta te d e h y d r o
genase (LDH) of S . c e r v i . K l DEC, '--' cen tperaz ine ,
\Y} 72/70, [ f i 1 evamisole and El suramin.

32
tion at 100 AIM concentration and compound 72/70 was ineffective
(F ig . 5 ) .
Fumaj-ase: The enzyme was i n h i b i t e d by very low concentration
of suramin recording 72% decl ine at 1 AIM concentration while cent -
pe r az ine , DEC and 72/70 showed 32%, 32% and 20% inhibi t ion r e s
pec t ive ly at 10 yuM concentration (F ig . 6 ) .
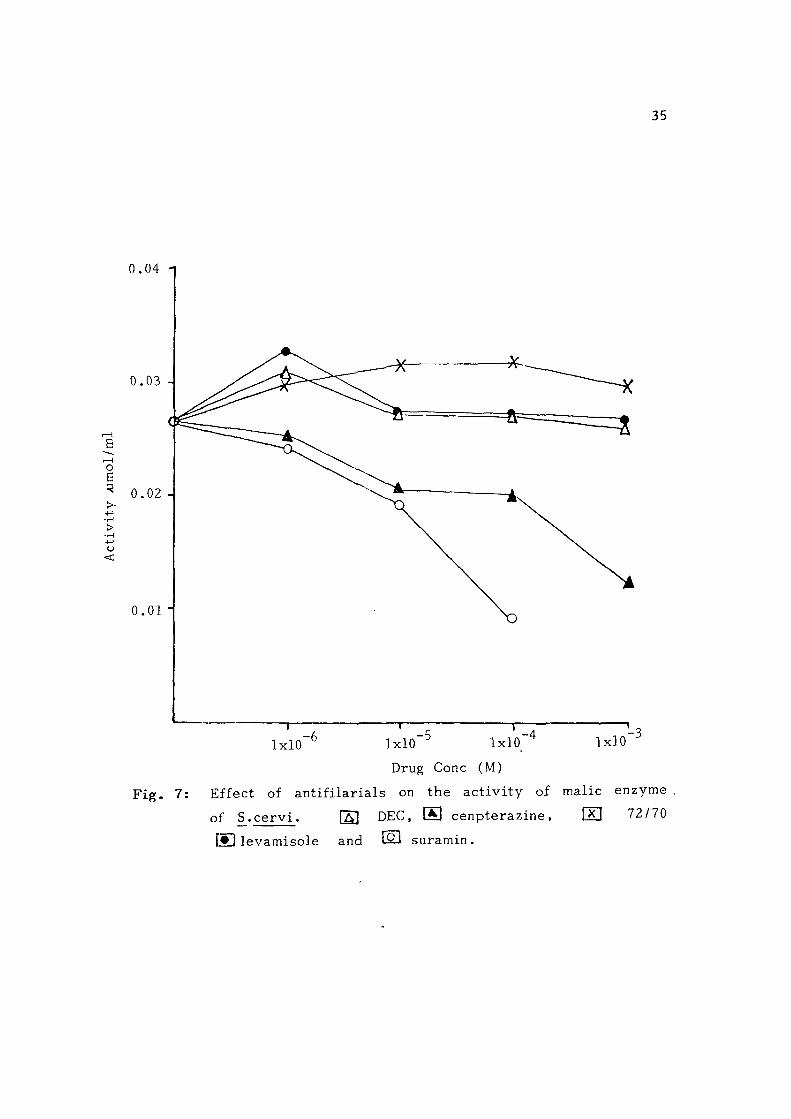
Malic enzyme: Centperazine and suramin were found to be effect ive
to some extent at h i g h e r concent ra t ions . Suramin caused 60% i n h i b i
tion at 0.1 mM concentration whi le 1 mM centperazine was r equ i r ed
to get 51% inhib i t ion of the enzyme. Levamisole , DEC and 72/70
were ineffective even at h ighe r concentration (F ig . 7 ) .
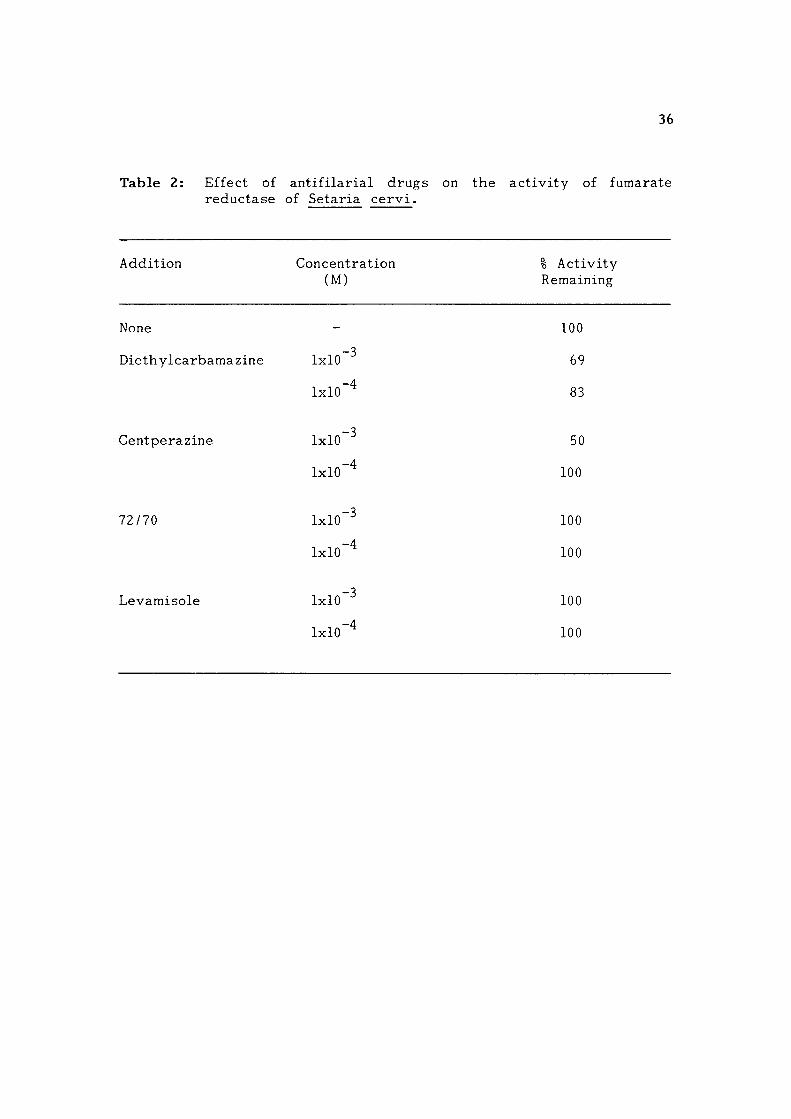
Fumarate reductase: The enzyme was inh ib i t ed by 50% and 31%
r e s p e c t i v e l y in presence of 1 mM centperaz ine and DEC. Levamisole
and compound 72/70 were ineffective (Table 2 ) .
Succinate dehydrogenase: About 75% inhibi t ion of SDH ac t i v i t y was
obse rved at 100 /JM concentration of suramin whi le centperazine
levamisole and compound 72/70 showed 50-57% inhib i t ion at t h i s
concentration (Table 3 ) .
Since suramin was found to be a potent i nh ib i t o r of most
of t h e enzymes of PEP-succinate p a t h w a y , t h e va lues of the i n h i
bi t ion constant (K.) was determined from the Dixon p lo t s and r e
corded in Table 4 . The K. va lues for PK, LDH and fumarase were

33
10.0 -
o E
>
<
IxlO -6 1x10 ^ 1x10
Drug cone (M)
1x10
F ig . 5: Effect of a n t i f i l a r i a l s on the a c t i v i t y of malate d e h y d r o
genase (MDH) of S . c e r v i . (AI DEC, E cen tperaz ine ,
t S 72 /70 , I S levamisole and \9} suramin.

34
^ >-
<
1.6
1.4
1.2
0.8
0.4
1x10 -4 1x10
-3 xlO -' 1x10
Drug cone (M)
F ig . 6: Effect of a n t i f i l a r i a l s on the a c t i v i t y of fumarase- of S . c e r v i .
LSj DEC, |A] cen tperaz ine , t S 72/70, CS le vamisole and
suramin .
35
0 . 0 4 - 1
5
<
0.03 -
0.02 -
0.01
1x10 -6 1x10
F i g . 7:
1x10 - 1x10
Drug Cone (M)
Effect of an t i f i l a r i a l s on the a c t i v i t y of malic enzyme
of S . c e r v i . {^ DEC, I S cenp te raz ine , CE] 72/70
f S levamisole and tSJ suramin.
36
Table 2: Effect of an t i f i l a r i a l d rugs on the ac t i v i t y of fumarate reductase of Setaria c e r v i .
Addition Concentration % Act ivi ty (M) Remaining
None - 100
-3 Diethylcarbamazine 1x10 69
IxlO"'^ 83
-3 Centperazine 1x10 50
IxlO"^ 100
72/70 1x10 ^ 100
Ixio"'^ 100
-3 Levamisole 1x10 100
IxlO"'^ 100

37
Table 3: Effect of a n t i f i l a r i a l drugs on the a c t i v i t y succinate dehydrogenase of Setar ia c e r v i .
Addition Concentration (M)
% Act ivi ty remaining
None
Centperazine 1x10 -3
1x10 -4
100
25
43
72/70 1x10
1x10
40
50
Levamisole 1x10
1x10 -4
25
43
Suramin 1x0 25
/ 'A -:a.kO Lixx :-4^^
I ^ f
[ D3/3/7 .-3

38
Table 4: K. value of suramin for the enzymes of PEP-succinate 1
pathway of Setar ia cervi_.
Enzymes K. (M)
-7 Pyruva te kinase 6.5x10
-5 Phosphoenol p y r u v a t e - 2.5x10 carboxy kinase
-7 Lactate dehydrogenase 8.75x10
Malate dehydrogenase 1.00x10
-7 Fumarase 7.00x10
Malic enzyme 1.4x10

39
-7 -7 -7 recorded as 6.5x10 , 8.75x10 and 7.0x10 r e s p e c t i v e l y . There
were lower than the values for PEPCK, MDH and malic enzyme.
Effect of 2J1 v i tro treatment of S. cervi adults on the enzymes of
PEP succinate pathway
Since suramin, centperazine and levamisole showed i n h i b i t o r y
effect on the enzymes of PEP-succinate pa thway , a t t empts were
a lso made for s tudying the effects of these drugs on l i ve worms
during Jji v i t r o incubation for 3 h r in R inger ' s so lut ion. Significant
inh ib i t ion was obse rved when the concentration of t h e s e drugs was
5 mM. At t h i s concentration suramin inh ib i t ed the mi tochondr ia l
enzymes (SDH and FRD) by 75% and 79% and the cy tosol ic enzymes
(PEPCK, PK and LDH) by 44%, 35% and 41% r e s p e c t i v e l y (Table
5 ) . Centperazine (5 mM) caused 62%, 63% inhib i t ion of SDH and
FRD and 51% and 34% inhib i t ion of MDH and malic enzyme (Table
6 ) . Levamisole at 5 mM concentration inh ib i t ed SDH and FRD by
65% and 33% whi le the a c t i v i t y of PK and MDH were lowered by
49% and 30% r e s p e c t i v e l y (Table 7 ) .
Isolation and propert ies of phosphoenol pyruvate carboxykinase
Phosphoenol py ruva te carboxykinase (PEPCK) from S. c e rv i
adul t females was ve ry unstable in nat ive s ta te and the enzyme
was inac t iva ted within 24 hours when kept at 4-5 C. A s tudy
of the effect of a few th io l compounds (y^-mercaptoe thanol , ME
and d i t h i o t h r i t o l , DTT) on the ac t i v i t y of PEPCK ind ica te t h a t

40
Table 5: Effect of ui v i t r o t reatment of l ive Setaria ce rv i with suramin on the a c t i v i t y of the enzymes of PEP-succinate pa thway .
Enzyme Control (/umol/ml)
Exper imenta l (/umol/ml)
Inhibi t ion ( I )
Pyruvate kinase
Phosphoenol py ruva te ca rboxykinase
Lactate dehydrogenase
Malate dehydrogenase
Malic enzyme
0.279±0.010
0.939±0.070
2.450±0.001
31.800±0.001
0.032±0.000
0.182±0.013
0.626+0.040
1.460±0.110
23.30+1.690
0.023±0.001
35
44
41
27
29
Fumarase
Fumarate reduc tase
Succinate dehydrogenase
0.614±0.000
0.062+0.000
0.174±0.001
1,780±0.084
0.016±0.003
0.036+0.009
189(act i -vation)
75
79
Adult motile S .ce rv i were incubated with suramin (5 mM) in R inge r ' s solution at 37 C for 3 h r . A 10% ( w / v ) homogenate of the worms was made in 150 mM KCl. The a c t i v i t i e s of the cytosol ic enzymes were efjtimated in post mi tochondr ia l f rac t ion , whereas the a c t i v i t i e s of fumarate reduc tase and succinate dehydrogenase were determined in the mi tochondr ia l f rac t ion , A control batch was also incubated along with e x p e r i m e n t a l .

41
Table 6: Effect of in v i t r o t rea tment of l ive Setaria cerv i with centperazine on the a c t i v i t y of the enzymes of PEP-succinate pathway enzymes .
Enzyme
Pyruvate kinase
Phosphoenol pyruva te carboxykinase
Lactate dehydrogenase
Malate dehydrogenase
Malic enzyme
Fumarase
Fumarate reductase
Succinate dehydrogenase
Adult motile S.
Control (>umol/ml)
0.616±0.05
1.123±0.10
2.670±0.24
31.50±0.35
• 0.027±0.01
0 .76U0.07
0.122+0.02
0.269±0.03
ce rv i were n
Exper imenta l Oumol/ml)
0.481±0.09
0.984+0.09
2.21010.41
15.91±2.05
O.OlSiO.OO
0.49210.04
0.045+0.01
0.10110.06
incubated wi th centperaz ine
Inhibit ion (%)
22
13
17
51
34
35
63
62
(5 mM) in Ringer ' s solution at 37 C for 3 h r . A 10% (w/v ) homogenate of the worms who made in 150 mM KCl. The a c t i v i t i e s of the c y t o -solic enzymes were estimated in pos t mi tochondr ia l f rac t ion , whereas the a c t i v i t i e s of fumarate r educ ta se and succinate dehydrogenase were determined in the mi tochondr ia l f rac t ion . A control pa tch was also incubated along with the e x p e r i m e n t a l .
42
Table 7: Effect of iii v i t r o t reatment of l ive Setar ia ce rv i with levamisole on the a c t i v i t y of the enzymes of PEP-succinate pathway enzymes
Enzyme
Pyruvate k inase
Phosphoenol py ruva te carboxy kinase
Malate dehydrogenase
Lactate dehydrogenase
Malic enzyme
Fumarase
Fumarate reduc tase
Succinate dehydrogenase
Adult motile S,
Control Oumol/ml)
0.56±0.06
1.24±0.25
24.50±0.10
1.80±0.10
0.03±0.01
1.63±0.12
0.03±0.00
0.15±0.04
,cerv i were O
Experimental Inhibi t ion (/umol/ml) (%)
0.29±0.03
1.21+0.22
17.21±0.18
1.61±0.02
0.03+0.01
1.28±0.51
0.0210.00
0.05±0.00
incubated with levamisole (5
49
4
30
11
17
22
33
65
mM) in R inge r ' s solution at 37 C for 3 h r . A 10% (w/v ) homogenate of the worms was made in 150 mM KCl. The a c t i v i t i e s of the c y t o -sol ic enzymes were es t imated in pos t mi tochondr ia l f rac t ion , whereas the a c t i v i t i e s of fumarate reduc tase and succinate dehydrogenase were determined in t h e mi tochondr ia l f rac t ion. A control batch was also incubated along with expe r imen ta l .

43
both DTT and /3-UE a c t i va t ed the enzyme suggesting t h e r e b y the
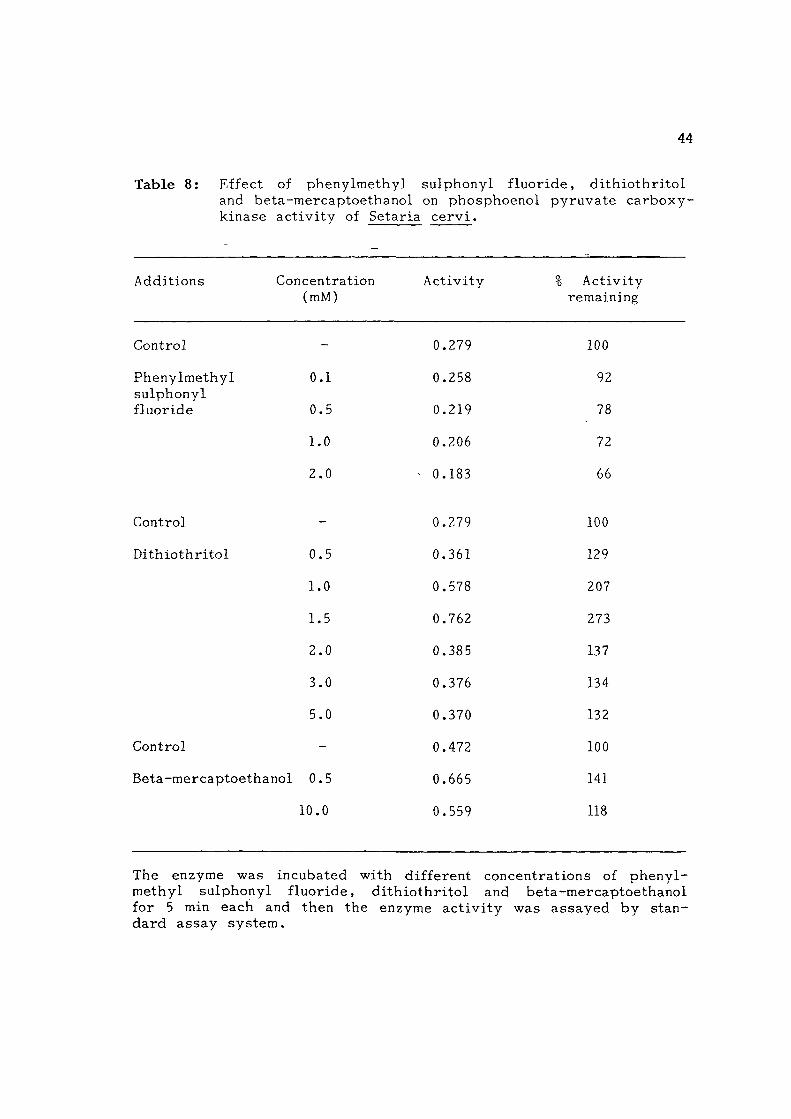
involvement of -SH grcSups in the ca ta ly t i c reaction (Table 8 ) . Add i
tion of p ro tease i n h i b i t o r (phenyl methyl sulphonyl f luor ide , PMSF)
showed s t ab i l i z ing effect on the enzyme during s to rage .
PEPCK from adul t female S .cerv i was p a r t i a l l y pur i f i ed
using a combined p rocedure of ammonium sulphate f rac t ionat ion,
desal t ing through Sephadex G-25 column and affinity chromatography
on Blue Sepharose CL-6B, d e t a i l s of which have been d e s c r i b e d
under Mater ia ls and Methods sec t ion . The post mi tochondr ia l s u p e r
natant (12,000xg) p r e p a r e d by homogenising 7 g motile worms in
suspension buffer containing Tr i s HCl (50 mM), DTT (1.5 mM) and
PMSF(0.1 mM) was subjec ted to (NH.)_SO. f ract ionat ion. The p ro t e in s
p rec ip i t a t i ng between 30-70% saturat ion of (NH.)_SO. contained most
of the or ig ina l PEPCK a c t i v i t y . The p r e c i p i t a t e was so lub i l i s ed
in small volume of suspension buffer and passed through a f r e sh ly
p r e p a r e d Sephadex G-25 column (14x1.4 cm) for d e s a l t i n g . The
fract ions r i c h in PEPCK were pooled and app l i ed over a f r e sh ly
p r e p a r e d Blue Sepharose column. The p ro te ins adso rbed on t h e
affinity column were e lu ted with increasing concentrat ions of NaCl
(0 .5 -2 .0 M) in a s t epwise manner. The summary of the pur i f ica t ion
protocol i s shown in Table 9. Over 20 fold pur i f ica t ion of the
enzyme was a c h i e v e d .
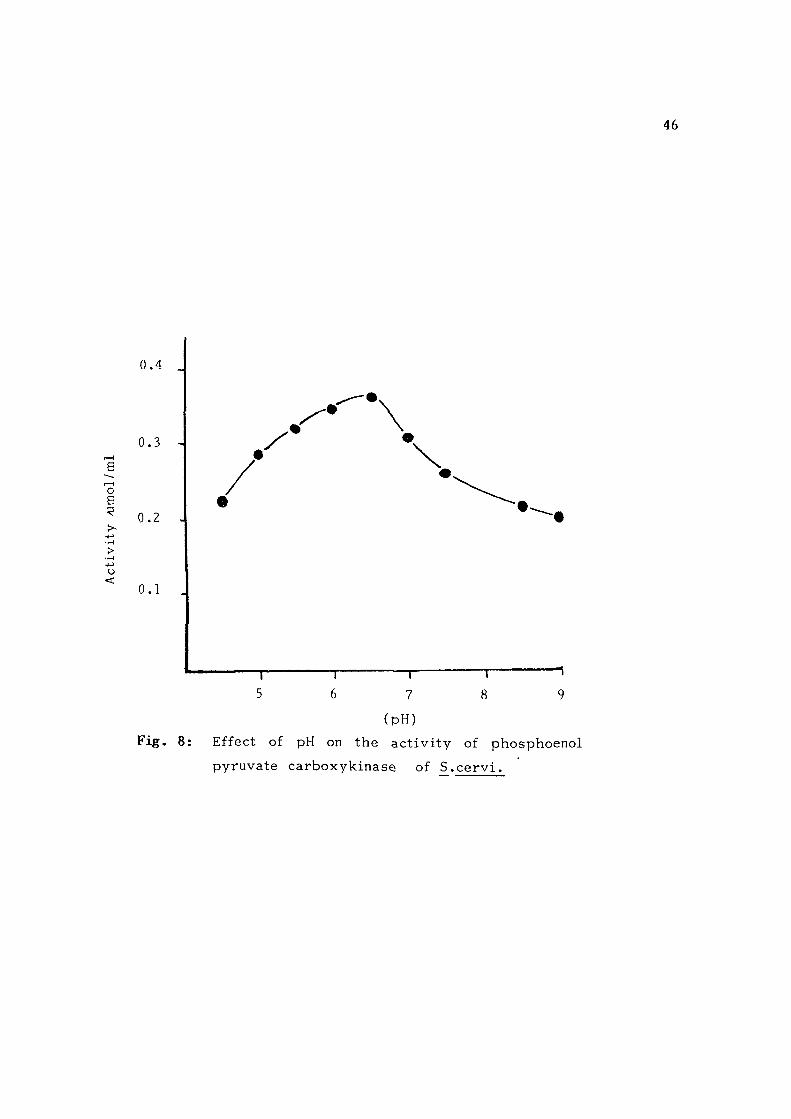
Properties of the enzyme: F ig . 8 shows the pH a c t i v i t y curve
of PEPCK using 2 buffer sys tems v i z . , sodium potassium phospha t e
and T r i s HCl. The enzyme showed maximal a c t i v i t y at pH 6 . 5 .
44
Table 8: Effect of phenylmethyl sulphonyl f luo r ide , d i t h i o t h r i t o l and beta-mercaptoethanol on phosphoenol py ruva t e c a r b o x y -kinase ac t i v i t y of Setaria c e r v i .
-
Additions
Control
Phenylmethyl sulphonyl f luor ide
Control
Di th io th r i to l
Control
Beta-mercaptoetha
Concentration (mM)
-
0.1
0.5
1.0
2.0
-
0.5
1.0
1.5
2.0
3.0
5.0
-
nol 0.5
10.0
Act ivi ty
0.279
0.258
0.219
0.206
- 0.183
0.279
0.361
0.578
0.762
0.385
0.376
0.370
0.472
0.665
0.559
% Act ivi ty remaining
100
92
78
72
66
100
129
207
273
137
134
132
100
141
118
The enzyme was incubated with different concentra t ions of p h e n y l methyl sulphonyl f luo r ide , d i t h i o t h r i t o l and be ta -mercap toe thanol for 5 min each and then the enzyme a c t i v i t y was a s sayed by s t an da rd assay sys t em.

- fH
" >-^ +^
0) Tj
at) tn nj
> U
> ^^ O ° ° o - (U Di
C
o +->
O
<o <o
• .—1
1—1
o^ o
• o
o o 1—1
i n (Nl
• (NJ
un o tNl
• o
00 1—1 r—(
CO
'—' " i n
i n vO '^f
• o
i n
I — (
o o •
(NJ
a r-• 1—i
oo
•>-' 2 o T3
>^: • f H
> • H + J O
<
> U <U > o o (D oi
_E (n
-i-> • H C D '
o\o
^"^
c •H
-(-> O O U
H a
c •r^
-i->
o u P-.
1—1
<i + J
o H
1—1
G " • " ^
bO E
o E ^ 3 ^ -5 E > "^
c o
• H
a •H !H O W <U
Q
CO
(N!
1—1
o i n
0 0
0 0
(NJ
CM CO
OO
ro
^ o •vD
0 0 r-•
CO
o ^ * 1-H
CO CD
o
00
o
rt Vi
+ j
X <u
<u <u ^ m
o 0) o u 00 ^
be
o o o
*•
(M 1—1
1 o -t-" r-t
V <« E -H
^
o o a j :
v ^ O
z * ^
c o
•H
o rt ?H
m
c o
o U
o ^
• ^ I
Z CO
tn O u
r-H , _ | CQ 0)
CO
E
H H D
he c
•H C
•H rt c o u .
c ^ o i n -H
vO u -H
P- u 3
. P-
E o
i n Ti w 0)
tn
m rt
•- 5 ^ ^
C " In .,-1 (U
a D
•5 <u +^ 0) 3
> I E
IS D
u 5-1
OJ P-
tn
tsi • iH
0) bC O E o
J2
u <L)
tn E
o •'^ •H X
< T3 2 1—1
O MH O O
u o •H E , D 5-1
0)
o-B >
o S E
E ^
c 3
<U
E N
c
45
46
0.4
0.3 A / '
• 0.2 J
0.1
-^
(pH)
Fig. 8: Effect of pH on the activi ty of phosphoenol
pyruvate carboxykinase of S.cervi .

47
Since GDP is necessary for the enzyme, i t s effect on PEPCK has
been s tud i ed . Maximum a c t i v i t y was obse rved when 0.5 mM GDP
was added to the react ion mixture (F ig . 9 ) . K value determined m
by studying the r a t e s of the PEPCK ca ta lysed reaction at different
concentration of subs t r a t e (PEP) was found to be 2.0 mM (F ig .
10). The enzyme e x h i b i t e d h y p e r b o l i c sa tura t ion curve with r e spec t
to s u b s t r a t e .
Since PEPCK from seve ra l sources has been shown to r e q u i r e
Mn and HCO, for i t s a c t i v i t y , exper iments were conducted using
different concentrations of anions and ca t ions . The enzyme from
S .cerv i r equ i r e s 10 mM MnCl-, and 50 mM NaHCO, for showing optimal
a c t i v i t y (F ig . 11a,b) MgCl^ showed i n h i b i t o r y effect as 28% ac t i v i t y
was lost when 2 mM MgCl_ was added to t h e react ion mixture (Fig .12)
The effect of FDP and ATP on PEPCK of S . ce rv i was also s t u d i e d .
ATP showed i n h i b i t o r y effect lowering the a c t i v i t y by 69% at 4
mM concentration (F ig . 13a), FDP, howeve r , showed ac t iva t ing effect
when the enzyme was incubated for 15 min in presence of the buffer
(F ig .13b) .

48
0.4 H
o e
•H >
• H +-> O
<
0.2
F i g . 9:
0.5 1.0 1.5
GDP (mM)
Effect of GDP on the ac t i v i t y of p h o s p h o -
enol py ruva te ca rboxykinase of S . c e r v i .

49
L. vO
w 0
h •*
o c (L) O
X a VI o x; a
MH O
• ( - ' • H
> • H 4-> U
I )
c o
(U 4 - ' n) Js
4-1 U)
X I 3 [ f i
4- (
o
c o
• H +->
(5) o c o u
M
c • r - (
<t) 0)
u c
•H H-l
o
u
H H
W
.—1
X I 3 o
TJ
X u 3
0)
> (D ^ <U c
x; >->
o x: tn
+ j (D m n
1—1
> ! H
<D CJ
c/5
O
(D V)
a c
•rH 4«i > X O
X ^ 1
u
0)
> 3 U
>-a
• 0) 3
1—1
n]
> E
fc-i
0) -C 4 J
• H
<ti • — (
3 O
1—1
(D O
U O
"+H
o 1—1
r—1 (TJ o o a
•H
o 0) ^ 1
•H
-[Ui/xouirv A;TA :: ^V

0.8 1 50
o E ? >
+-> • i - t
> •r-i +->
u <
0.6
0.4 -
0.2
10 20
MnCl^ (mM)
F i g . 11a: Effect of Mn on the ac t i v i t y
of phosphoenol py ruva te c a r b o x y -
kinase of S . ce rv i -
o e
NaHCO^(mM)
F i g . llb^: Effect of HCO: on the . , .^i. uix^ enzyme
phosphoenol py ruva t e carboxykinase
of S . c e r v i .

51
0.3 -
o E
0.2 -
OA -
T 4
2 +
10
Mg (mM)
F i g . 12: Effect of Mg''' on the ac t i v i t y of Tshospho-
enol p y r u v a t e ca rboxykinase of S . c e r v i .

0.2 T 52
o E ?
>^ +->
> •H +->
u
0.1 -
"T 4 1 2 3
ATP (mM)
Fig. 13a: Effect of ATP on part ial ly purified
phosphoenol pyruvate carboxykinase
of S.cervi .
< 0.4 -
0.2 _
— 1 ~ 0.2
— r 0.4
T — r 0.8
Fig. 13b:
0.6
FDP (mM)
Effect of FDP on part ial ly purified
phosphoenol pyruvate carboxykinase
of S.cervi .

53
DI SOBS ion
The in te rmedia ry metabolism of p a r a s i t i c nematodes has
r ece ived wide attention during the las t two decades and significant
cont r ibut ions have been made in dec ipher ing different metabolic
pa thways opera t ing in f i l a r i a l and o the r in tes t ina l he lmin ths as
well a s s tudying the p r o p e r t i e s of the regula tory and terminal
enzymes of c a rbohyd ra t e metabolism. These s tud ies enable us
in improving our understanding of the biology of t he se organ isms .
S . ce rv i a p p e a r s to be equipped with a l l t he enzymes of
PEP-succinate p a t h w a y . The a c t i v i t y of some of the key enzymes
in S . ce rv i a p p e a r s to be h ighe r than the corresponding values
r e p o r t e d for o the r f i l a r i i d s ( L . c a r i n i i , S r ivas tava et_ al^., 1970;
C .hawking i , S r ivas tava and Ghatak, 1971) t rematodes (Fasciola
h e p a t i c a , P r i c h a r d and Schofield, 1968) and nematodes (Haemonchus
contortus l a r v a e , Ward and Schofield, 1967; Ascar i id ia g a l l i , S r i v a s t a v a
et a l . , 1970). Invest igat ion on g lycoly t ic and carbon d iox ide fixing
enzymes have shown tha t p a r a s i t e s producing succinic and vo la t i l e
fa t ty ac ids in preference to lac t ic acid as end produc t of c a r b o
h y d r a t e u t i l iza t ion have a modified scheme of g lyco lys i s d iverging
at t h e PEP l e v e l . These p a r a s i t e s usual ly pos ses s high a c t i v i t e s
of PEPCK and MDH and low l eve l s of PK and LDH, and the g lyco
l y t i c pa thway p roceeds only up to the formation of PEP which
i s then ca rboxy la t ed to oxaloacetate by an ac t ive PEP-carboxykinase
(von Brand, 1973). The worms coming under t h i s group a re Ascaris
lumbr ico ides (Bueding and Saz, 1968). H. contortus l a r v a e (Ward
et a l . , 1968a ,b) , Hymenolepis diminuta (Bueding and Saz, 1968),

54
F.hepatica (Prichard and Schofield, 1968b) and T.spiral is larvae
(Agosin and Aravena, 1959) which convert negligible amount of sugars
to lactic acid. On the other hand, typical lactic acid producers
like S.mansoni {Buoding and Saz, 1968) and C.hawkingi (Srivastava
et a l . , 1968; Srivastava and Ghatak, 1971) converting 80-90% glucose
into lactic acid resemble vertebrate tissue in possessing high levels
of PK and LDH and low levels of PEPCK and MDH. The intermediate
case may also occur as in Dictyocaulus viviparous, a worm depending
essentially on aerobic life where the activit ies of PK and PEPCK
are more or less of the same order (von Brand, 1973).
S.cervi aerobically converts about 28% glucose into lactic
acid (Anwar ei_ aL., 1975) and possess a PK/PEPCK ratio of around
0.4. Hence it appears that both glycolytic and PEP-succinate path
ways are operating in the bovine filarial parasite and thus it differs
metabolically from other filarial worms (L.carinii and C.hawkingi)
and resemble more closely the intestinal parasite specially with
regard to the PEP metabolising enzymes. The difference in the
metabolic activity can be explained on the basis of the location
of the parasite in the host . S.cervi resides on the peritoneal
folds of the intestine having relat ively high oxygen tension while
C.hawkingi and L.carinii thr ive in heart and pleural cavity of
crow and cotton rat respectively under a rich supply of oxygen.
Studies on the subcellular localization of the enzymes of
PEP-succinate pathway in S.cervi showed that PK, PEPCK, MDH,

55
LDH, fumarase and malic enzyme were present in the soluble fract
ion of the homogenate. This is similar to the cytosolic localization
of these enzymes observed in vertebrate tissues (Lehninger, 1984).
As in vertebrate tissues and other intestinal parasites SDH and
FRD were localized in mitochondrial pellet of S.cervi .
^" S.cervi the level of PEPCK is more than that of PK,
which is in contrast to the observation made in S.mansoni (Brazier,
1973) and a few other nematode paras i tes . This is possibly due
to the different habitat of the parasi tes as S.cervi is found in
a high oxygen tension. According to Prichard (1976) in F.hepatica,
the observed NADH stimulation of PEPCK under conditions of lower
oxygen avai labi l i ty , higher concentration of NADH and PEP due
to a stimulation of PFK and overall rate of glycolysis may facilitate
PEPCK activity to increase the supply of fumarate for regeneration
of NAD and to maintain pyruvate production via MDH and MDH (decar-
boxylating ) should the PK activity be insufficient due to a lower
concentration of FDP. Thus, the net effect of metabolic control
on the act ivi t ies of PK and PEPCK would be to allow the enzymes
to complement each other for maintaining a steady supply of pyru
vate in accordance with the demands for NADH to generate energy
while allowing the parasite to adjust the flow of carbon to fumarate
in accordance with supply of NADH and oxygen.
Although the detailed mechanism for controlling fermentation
in different helminths has not been resolved, it is clear from

56
the s tudies made tha t PK and PEPCK play a key role in de te rmin
ing the fermentation p roduc t s produced by different helminths and
pe rhaps in determining whe the r the products can change in response
to a l tera t ion in environmental demands on the organism.
In most of the helminth p a r a s i t e s PEPCK se rves in the
carboxylat ion of PEP leading to t h e syn thes i s of OAA and thus
regulates g l y c o l y s i s . The enzyme in mammalian t i s sues b r ings
about the syn thes i s of PEP from OAA thus playing an important
role in the regulation of gluconeogenesis .
PEPCK pur i f i ed from S . ce rv i has a r ecovery g rea te r than
100% in post ' mi tochondria l and ammonium sulphate (30-70% sa tura t ion)
fraction (Table 9 ) . Presence of an inh ib i to r has e a r l i e r been
repor ted by Tar tora et_ a l . (1985) in DEAE sephace l fraction from
yeast PEPCK, which was ident i f ied as adenyla te k i n a s e . The enzyme
s tudied in S .ce rv i i s cy toso l ic resembling the analogous enzyme
isola ted from A. lumbr ico ides muscle (van den Bossche , 1969). In
general the p r o p e r t i e s of PEPCK from bovine f i l a r i a l worm d id
not differ s ignif icant ly from t h e analogous enzyme obtained from
some helminths and v e r t e b r a t e t i s s u e s . The enzyme from S .ce rv i
showed maximal a c t i v i t y around pH 6.5 while PEPCK from F .hepa t i ca
(Behm and Bryant , 1982) and H.diminuta (Prescot t and Campbel l ,
1965) were opt imal ly ac t i ve between 6 . 0 - 6 . 5 . PEPCK from S .ce rv i
has speci f ic requirement for Mn which could not be r ep laced
t>y Mg showing s i m i l a r i t y with the enzyme from chicken l i v e r

57
(Kurahashi et_ al_., 1957), E.granulosus (Agosin and Repet to , 1965)
and F . h e p a t i c a (Behm and Bryant , 1982). Among the anions , HCO_
st imulated the carboxyla t ion of PEP in S .ce rv i while PEPCK from
pig l i v e r was i nh ib i t ed by HCO- (Chang et_ a l . , 1966). Among
the s eve ra l nucleot ides inves t iga ted GDP was most effect ive cofactor
for S . ce rv i PEPCK, followed by IDP while negl igible a c t i v i t y was
obse rved with ADP. Similar pa t t e rn was obse rved in M.expansa
(Behm and Bryant , 1982). However, IDP was more ac t ive for the
enzyme from M.s imi l i s (McManus and James , 1975), A.suum muscle
(van den Bossche , 1969), F . h e p a t i c a ( P r i c h a r d and Schof ie ld , 1968)
and H.diminuta (Presco t t and Campbel l , 1965).
PEPCK from S .cerv i d i s p l a y e d t y p i c a l h y p e r b o l i c k ine t i c s
with r ega rd to PEP and nucleot ides (GDP) resembling the enzyme
from many o the r sou rce s . ATP e x e r t s i nh ib i t o ry effect on PEPCK
of S . ce rv i s imi la r to the obse rva t ions made in M.expansa (Behm
and Bryant , 1982). However, t he enzyme from A.suum muscle (van
den Bossche , 1969) and o the r sources were unaffected by ATP.
Since ATP also i n h i b i t s PK in S .ce rv i (Anwar, N. P h . D . t h e s i s ,
1976) i t i s un l ike ly tha t t h i s nucleot ide i s a major regula tor of
PEP/OAA bronchpoint in the forward d i rec t ion to succ ina te . Stimu
lation of PEPCK a c t i v i t y in S . ce rv i by th io l compounds suggest
the p resence of s u l p h y d r y l (-SH) groups at t h e ac t ive s i t e of the
enzyme molecule . Wilkes et_ a l . (1981) also r epo r t ed t h e presence
of --SH groups in the enzyme of A.suum.

58
The jji vitro effect of a few filaricides on the enzymes
of PEP-succinate pathway was studied for identifying sensitive
molecular targets to provide a more rational approach for the chemo
therapy of f i lar ias is . Among the different filaricides tested,suramin
was most effective, inhibiting PK and fumarase activity of S.cervi
at 1 /uM concentration, while centperazine and DEC exerted inhibitory
effect on these enzymes at ten times higher concentration. Maximum
inhibition of PEPCK was achieved with CDRI compound 72/70 which
lowered the enzyme activity by 41% at 1 ^M concentration, while
suramin was effective at higher concentration (100 pm). Levamisole,
DEC and centperazine, however, could inhibit the enzyme at 1 mM
concentration. LDH was inhibited by suramin (10 jaM), centperazine,
levamisole, compound 72/70 and DEC at 100 /uM concentration; while
MDH was inhibited by suramin (10 /uM), and levamisole and cent
perazine (100 /uM). Malic enzyme was inhibited by 100 uM suramin
and 1 mM centperazine respect ively. Among the mitochondrial enzymes,
SDH was inhibited by suramin, centperazine, 72/70 and levamisole
at 100 /uM concentration, while 1 mM centperazine was needed for
exhibiting inhibitory effect on FRD .
A study of the effect of these drugs on live worms during
in vitro incubation indicated that suramin, centperazine and leva
misole could exhibit inhibitory effect on most of the enzymes of
PEP-succinate pathway only when motile worms were incubated at
37 C for 3 hours at 5 mM concentration. It is quite possible that
the cuticle of the worm is serving as a bar r ie r for the transport

59
of these drugs and hence high concentrations are needed for showing
inhibitory effect on the enzymes of living worms.
The biochemical mode of action of suramin (hexasodium
3,3 ' ureylene-bis-8(3 benzamido-p-toluido)-l ,3 ,5-naphthalene sulpho-
nate) against the filarial worms, hookworms and ascaris has been
the subject of research in this laboratory as well as elsewhere.
Studies conducted so far have indicated that suramin inhibits LDH
from D.immitis at 6 /UM (Walter, 1979); LDH and MDH from 0.volvulus
at 5 yuM (Walter and Shulzkey, 1980), protein kinase I from
0.volvulus at I jaU (Walter and Shulzkey, 1980), phosvitin phosphory-
lating protein kinase from B.malayi, A.viteae and S.cervi at 30
uM (Saxena et_ a3 . , 1984), folate dehydrogenase from D.immitis
and B.pahangi at 5 ^M (Jaffe, 1980); malic enzyme from 0.volvulus
at 1"0 nM (Walter, 1982); phosphofructokinase from S.cervi at 5
nM (Sharma et a l . , 1987); N-acetyl-^-D-glucosamindase from S.cervi
at 30 /UM (Singh et_ al_., 1986) and phosphatidylglycerolphosphate
synthetase, the key enzyme in the biosynthesis of phosphatidyl
glycerol, from 0.volvulus (Srivastava et a l . , 1987). However,suramin
had no inhibitory effect on AChE from S.cervi even at 1 mM con
centration (Sharma, 1988 Ph.D. t h e s i s ) . All these studies were
conducted with adult stage of these paras i tes .
For onchocerciasis suramin remains the only acceptable
microfilaricidal drug. It also acts on adult worms of L.carinii
(Lammler e^ a l . , 1971) and W.bancrofti (Hawking, 1978). After

60
intravenous injection it combines with serum proteins (histones,
serum globulins , albumin and casein) and persists for long periods
in the blood stream.
Diethylcarbamazine, the currently used filaricide in t r ea t
ment of fi lariasis does not exhibit significant effect on the enzymes
of PEP-succinate pathway from adult S.cervi . It is a microfilari-
cidal agent active against W.bancrofti and B.malayi (Peters, 1979)
as well as 0.volvulus (Hawking, 1978). Sufficient evidence exists
to show that piperazine induces revers ible paralysis by acting
on neuromuscular system (van den Bossche, 1976). Electron micros
copic studies on the mf of A.viteae and L.carinii isolated from
infected hosts indicted cellular damage on treatment with DEC which
could be the direct effect of the drug or secondary manifestation
of its action (Melhorn et_ a^. , 1981).
DEC along with other drugs (centperazine, levamisole and
CDRI compound 72/70) significantly inhibit glucose utilization, syn
thesis of glycogen, protein synthesizing capacity and release of
mf from adult S.cervi females (Anwar et a l . , 1978; Srivastava,
1980).
Comley et_ al_. (1981) observed significant inhibition of
SDH and FRD activi t ies from A.tetraptera and A.suum during in
vitro incubation with 5 mM levamisole, which is in agreement with
our findings showing that 5 mM levamisole was needed for exhibi t
ing inhibitory effect on SDH and FRD of S.cervi .

61
Thus success has been achieved in purifying and character is
ing PEPCK from S.cervi, and studying the effect of a few filaricides
on the enzymes of PEP-succinat^ pathway. Strong inhibition _of
LDH and MDH by low concentration of suramin would interfere with
the reoxidation of NADH generated by glyceraldehyde 3-phosphate
dehydrogenase and lead to the eventual arrest of glycolysis of
the parasi te . Further, attempts would now be made for studying
the kinetic properties of the nea^ homogenous preparation of PEPCK
(free of dehydrogenases) in relation to the effect to act ivators ,
inhibitors and anthelmintic agents.

CHAPTER IV
SUMMARY AND CONCLUSION

62
Filariasis is a major public health problem affecting millions
of people in the tropical and subtropical regions of the world.
Though the disease is not fatal. Taut it is responsible for consider
able disabi l i ty and social stigma due to elephantiasis caused by
immunopathological reactions of parasite with the host . Effective
chemical or immunological remedies against filariasis are s t i l l lacking
and this is mainly because of poor understanding about the parasite
biochemistry. It i s , therefore, necessary to have a sound knowledge
of the morphology, physiology, sepcially of the biochemistry of
these parasi tes for developing drugs specifically inhibiting the
metabolic processes of the parasi te .
The filarial parasites derive energy for their survival*
mainly through carbohydrate utilization, however, the processes
involved in carbohydrate degradation are not identical with those
established in vertebrate tissues as none of these parasites possesses
the abil i ty of oxidising the carbohydrate completely to COp and
H_0. PEP-succinate pathway has been shown to have a crucial
role in the channeling of carbon as well as in the regeneration
of energy in many helminth paras i tes . Therefore, a study of PEP-
succinate pathway enzymes with special reference to the action
of antifilarials may provide useful leads for identifying paras i te-
specific enzyme targets for chemotherapy. In view of the non
availabil i ty of human filarial parasites (W. bancrof t i / B. malayi) in
sufficient amounts, Setaria cervi , a bovine filarial paras i te , resembl-

63
ing the human filarial worm in microfilarial periodicity and antigenic
pattern has been used in this investigation.
The present stu^y shows the subcellular localization of
the enzymes of PEP-succinate pathway. The enzymes-pyruvate
kinase, phosphoenol pyruvate carboxykinase, lactate dehydrogenase,
malate dehydrogenase, malic enzyme and fumarase were localized
in the cytosol whereas succinate dehydrogenase and fumarate reduct
ase were found to be particulate bound. Malate dehydrogenase
was found to be most active among the soluble enzymes.
Action of antifilarials on the enzymes of PEP-succinate
pathway wa"! also studied by incubating the enzyme extract or
live S.cervi with different concentrations of ant i f i lar ia ls . Among
the various compounds tested suramin at low concentration could
cause a marked inhibition of the activit ies of most of the enzymes.
Due to the large size of suramin (hexasodiumS ,3 ' -ureylene-bis -8(3-
benzamido-p-toluido)-l ,3,5-naphthalene sulphonate), it is likely
that an extensive biochemical interaction occurs between different
enzymes and the drug. Centperazine also showed significant inhi
bitory action on pyruvate kinase, lactate dehydrogenase fumarase
and succinate dehydrogenase. Incubation of live worms with th is
drug caused 62%, 63% and 51% inhibition of succinate dehydrogenase,
fumarate reductase and malate dehydrogenase respect ive ly . Diethyl-
carbamazine and levamisole were found to be more or less ineffective
at lower concentration. However, they could inhibit some of these

64
enzymes when used at h i g h e r concentra t ion. Compound 72/70 e x h i
b i t ed significant i nh ib i to ry effect only on phosphoenol pyruva te
ca rboxyk inase .
The most important regula tory branchpoint enzymes of PEP-
succinate pathway in S .ce rv i i s phosphoenol py ruva t e carboxykinase
which has been p a r t i a l l y pur i f ied and i t s p r o p e r t i e s were s tud i ed .
The p a r t i a l l y pur i f ied enzyme was v e r y unstable in nature but
i t could be s t ab i l i zed by the addi t ion of 0.1 mM phenylmethyl
sulphonyl f luoride (a p ro tease i n h i b i t o r ) and 1.5 mM d i t h i o t h r i t o l
( th io l compound) when added in the ex t rac t ion buffer during homo-
genisa t ion. These also prevented the inact ivat ion of the enzyme
during purif icat ion p r o c e d u r e .
Purif icat ion was done by a t h r e e s t ep p rocedure employing
(NH.)-SO^ fract ionat ion, gel f i l t ra t ion on Sephadex G-25 and Blue
Sepharose affinity chromatography which r e su l t ed in 20 fold p u r i f i
cat ion. The enzyme was found to be opt imal ly ac t ive at pH 6.5
and had a K value at 2.0 niM for s u b s t r a t e ( P E P ) . MnCl^ (10 m L
mM)and NaHCO-(50 mM) a re r e q u i r e d for maximal enzyme a c t i v i t y ,
2+ whi le Mg ions (2 mM) exe r t ed i n h i b i t o r y effect on the enzyme.
FDP was found to ac t iva te while ATP i n h i b i t e d the enzyme.
These s tud ies indicate the s e n s i t i v i t y of some enzymes
of PEP-succinate pathway in f i l a r i a l p a r a s i t e s to known drugs and
some candidate drugs being deve loped at t he i n s t i t u t e . They may.

65
therefore, serve as a potential metabolic targets for antifilarial
drug design. Further attempts would be made for purifying the
enzyme (PEP-carboxykinase) to homogeneity for studying the kinetic
properties in relation to act ivators, inhibitors and anthelmintic
agents. Oxidative activit ies in mitochondria like particles from
S.cervi would also be examined. The sensitive molecular targets
identified during this study are expected to provide a more rational
approach to filarial chemotherapy.

B I B L I O G R A P H Y

66
Agarwcil, A . , Saxena, J .K. and Ghatak, S. (1986). Ind . J . Med.
R e s . , 84, 260.
Agosin, M. and R^pet to, V. (1965). Comp. Biochem. P h y s i o l . ,
14, 299.
Anwar, N. (1976) . P h . D . t h e s i s submitted to Kanpur Un ive r s i ty ,
Kanpur.
Anwar, N . , Ansar i , A.A. and Ghatak, S.N. (1975). P r o c . Indian
Nat. Sc i . Acad. 41 , 550-558.
Anwar, N . , Ansar i , A.A. , Ghatak, S. and Krishna Murti , C .R . (1977) .
Zei tschr i f t fur Paras i tenkunde 51 , 275-288.
Anwar, N . , S r i va s t ava , A.K. and Ghatak, S. (1978). Ind . J . P a r a -
s i t o l . 2, 101-105.
B a r r e t t , J . (1983). Helminthol. Abs . 52, 1.
Behm, C.A. and Bryant , C. (1982) . In t . J . P a r a s i t o l . 12, 271 -
278.
Braz i e r , J . G . and Jaffe , J . J . (1973) . Comp. Biochem. P h y s i o l . ,
44B, 145.
Bucher , T. and P f l e i d e r e r , G. (1955) . Methods in Enzymology
New York: Academic P re s s 1, 435-440.
Bueding, E. and Saz, H . J . (1968) . Comp. Biochem. P h y s i o l . ,
24, 511.
Cav ie r , R . , Leger, N . , Har ichaux, J .M. and L o m e , M.C. (1971) .
Annals P a r a s i t o l . Hum Comp. 46, 497-501.
Chang, H . C . , Maruyama, H. , Mil ler , R .S . and Lane, M.D. (1966) .
J . Bio l . Chem. , 241, 2421.

67
Cher ian , S .N. , Stromberg, B . E . , Weiner, O .J . and Soulsky , E . J . L .
(1980) . In t . J . P a r a s i t o l . , 10, 227.
Coles , G . C . , Eas t , J .M. and Jenk ins , S.N. (1974) . E x p e r i e n t i a ,
30, 1265.
Comley, C.W. and Wright, D.W. (1981). In t . J . P a r a s i t o l . 11,
79-84.
Dunn, T . S . , Rovines , P . S . , Ba r re t t , J . and Bu t t e rwor th , P . E . (1988) .
In t . J . P a r a s i t o l . , 18, 21-26.
Ewer t , A. and Emerson, G.A. (1975). Am. J . Trop .Med . Hyg. ,
24, 71 .
Furman, A. and Ash, L.R. (1983). Acta Tropica 40, 45 .
Ghatak , S .N. , Bha t t achar j i , S. and Dhar , M.M. (1987) c i t ed in:
"CDRI Researches on Pa ra s i t i c Diseases" p . 6 3 .
Goodwin, L.G. (1984) . Trans . Roy. Soc. T r o p . Med. Hyg. ,78 (Supp l ) ,
1.
Hawking, F . (1978) . Wld. Hlth. Org. ONCHO 78, 143, 1-47.
Horecker , B . L . and Romberg , A. (1948) . J . Biol . Chem. 175,
385-390.
Hutchison, W.F. and McNeill,, K.M. (1970) . Comp. Biochem. P h y s i o l .
62B, 71-73 .
Jaffa , J . J . and Doremus, H.M. (1970) . J . P a r a s i t o l . 56, 254-260.
Jaf fe , J . J . , Chr in , L.R. and Smith, R . B . (1980) . J . P a r a s i t o l .
66, 428-433.
Johnson, K.H. and Bemrick, W.J. (1969) . Am. J . Vet . R e s . , 30,
1443.

68
Jo rden , P . (1958) . Br i t . J . Pha rmaco l . , 13, 318.
Kaushal , N.A. , Simpson, A . J . G . , Hussain, R. and Otteson, E.A.
(1984). E x p . P a r a s i t o l . 58, 182.
Kaushal , N.A. , Kaushal, D.C. and Ghatak , S. (1987) . Immunol.
Inves t . 16, 139-149.
Kornberg, A. (1955) . Methods in Enzymology New York: Academic
P re s s 1, 441-443.
Kurahash i , K., Pennigton, R . J . and Ut te r , M.F. (1957) . J . Biol .
Chem. 226, 1059.
Langham, M.E. and Kramer, T.R. (1980) . Tropenmed P a r a s i t . , 31,
59-66.
Lammler, G. , Herzgog, H. and Shutze, H.R. (1971) . Bul l . Wld.
Hl th . Org . , 44, 765-770.
Layne, E. (1957). Methods in Enzymology ( S . P . Colowick and N.O.
Kaplan e d s . ) HI , 451-454 New York: Academic P r e s s .
Lee, C.C. and Mil ler , J .H . (1969) . J . P a r a s i t o l . 55, 1035.
Lehninger , A.L. (1984). "Biochemistry" Worth P u b l i s h e r s .
Lowry, O.H. , Rosebrough, N . J . , F a r r , A.L. and Randal l , R . J .
(1951) . J . Biol . Chem. 193, 265-275.
Mahler , H.R.", Wit tenberger , M.H. and Brand, L. (1958) . J . B i o l .
Chem. 233, 770-782.
Malhotra , A . , Kaushal, N.A. and Ghatak, S. (1986) Immunol. Inves t ig .
15, 505.
McManus.D.P. (1986) . Int . J . P a r a s i t o l . , 17, 79-95.
McManus, D .P . and James, B . L . " ( 1 9 7 5 ) . Comp. Biochem. P h y s i o l .
518, 299.

69
Melhorn, H. , S te inhors t , U. , L indenba t t , B . , Gruentzig, J . and
Thomas, H, (1981), Zent ra l . B a k t e r i o l . Mikrobia l . Hyg.
Abst . 251, 263-272.
Mendis, A.H.W. (1985). T r o p . Med. P a r a s i t o l . 36, 15.
Metzger and Duwel, D. (1973). Int . J . Bioch. 4, 133-143.
Middleton, K.R. and Saz, H . J . (1979) . J . P a r a s i t o l . 65, 1.
Natarajan, P . N . , Zaman, V. and Yesh, T . S . (1973) . In t . J . P a r a s i t o l .
3 , 803-807.
Nelson, G.S. (1987). Tropenmed. P a r a s i t o l . 38, 57.
Nogami, S . , Nakamura, M. and Tanaka, M. (1984) . Kesei-chugoku-
zoss l i , 13, 141.
Ochoa, S. (1955). Methods in enzymology ( S . P . Colowick and N.O.
Kaplan, e d s . ) 1, 739-759.
Otteson, E.A. (1984). T rans . Roy. Soc. T r o p . Med. Hyg. , 78, 91.
P e t e r s , W. (1979). Pharmaceut ical for Developing Count r ies , p . 5 9 . "
Pong, S . S . , Wang, C.C. and F r i t z , L . C . (1980) . J . Neurochem.,
34, 351.
Pong, S.S. and Wang, C.C. (1982) . J . Neurochem., 38, 375.
Powell , J .W. , S tab les , J . N . and Watt, R.A. (1986) . Mol. Biochem.
P a r a s i t o l . , 18, 171.
P re sco t t , L.M. and Campbel l , J .W. (1965) . Comp. Biochem. P h y s i o l .
14, 491.
P r i c h a r d . L.M. and Schof ie ld , P . J . (1968a) . Comp. Biochem.
P h y s i o l . , 24, 697.
P r i c h a r d , L.M. and Schofie ld , P . J . (1968b) . Comp. Biochem.
P h y s i o l . , 24, 773-785.

70
P r i c h a r d , R.K. (1973) . Ind . J . P a r a s i t o l . 3 , 409-417.
P r i c h a r d , R.K. (1976) . Int . J . P a r a s i t o l . . 6, 227-233.
Racker, . E.- (1950) . Bipchem. B i o p h y s . Acta 4, 211-214.
Rathaur , S . , Anwar, N. and Ghatak , S. (1980). Z. Pa ras i t enkunde ,
62, 85 .
Rathaur , S . , Anwar, N . , Saxena, J .K . and Ghatak, S. (1982) . Z.
Pa ras i t enkunde , 68, 331.
Rathaur , S . , Kaushal , N.A. and Ghatak , S. (1985). Ind. J . P a r a
s i t o l . 9, 203-206.
Ree, G.H. , Hal l , A . P . , Hutchison, D.B.A. and Weather ley , B .C.
(1977) . T r a n s . R. Soc. T r o p . Med. Hyg. 71 , 542-543.
Rew, R . S . and Saz, H . J . (1977) . J . P a r a s i t o l . , 63, 123.
Saxena, J . K . , Bose, . S .K. , Sen, R . , Cha t t e r j i , R .K. , Sen, A.B.
and Ghatak , S. (1977) . E x p . P a r a s i t o l . 43 , 239-243.
Saxena, J . K . , S r i v a s t a v a , A.K. , K'alpanamurti, P . , C h a t t e r j i , R .K. ,
Ghatak , S. and Walter , R.D. (1984) . Tropenmed. P a r a s i t o l . ,
35, 174-176.
Saxena, J . K . , Singh, R . P . , Shukla , O .P . and Ghatak, S. (1988) .
(In p r e s s ) .
Saz, H . J , (1970) . J . P a r a s i t o l . 56, 634.
Saz, H . J . (1981) . Trends in Biochemical Science 6, 117-119.
Sharma, B . , Kaushal , N.A. and Ghatak , S. (1987). Ind . J . P a r a
s i t o l . , 11, 5-8 .
Sharma, B. (1988) P h . D . t h e s i s submit ted to Banaras Hindu Uni
v e r s i t y , Varanas i .

71
Shonk, C .E . and Boxer, G.E. (1964) . Can. Res . 24, 709-721.
Singh, R . P . , Saxena, J .K . and Ghatak, S. (1986) P r o c . Asian
Congress of Pa ras i to logy , 13-16 F e b r u a r y , Lucknow, I n d i a , p . 6 5 .
Singhal, K . C , Madan, B.R. and Saxena, P .N . (1973), J a p . J .
Pharmacol . 23 , 793-797.
Singhal, K . C , Madan, B .R . , Saxena, P .N. and J o h r i , M.B.L. (1975) .
Indian J . Pharmacol . 7, 22.
S r i va s t a va , A.K. (1985) . J . P a r a s i t o l . , 71 , 257.
S r iva s t ava , A.K. and Walter , R.D. (1987) . Med. Sc i . R e s . , 15,
435-436.
S r iva s t ava , V.M.L. (1969) . P h . D . t h e s i s , Agra Unive r s i ty , Agra.
S r i va s t ava , V.M.L. and Ghatak , S. (1971) . Ind. J . E x p . B i o l . ,
12, 472.
S r i va s t a va , V .M.L . , Ghatak, S. and Krishna Murti , C.R. (1968)
Exper imenta l P a r a s i t o l . 28, 176-185.
S r i va s t av a , V .M.L . , C h a t t e r j i , R .K. , Sen, A . B . , Ghatak, S . , Krishna
Murt i , C.R. (1970a). E x p . P a r a s i t o l . 28, 176-185.
S r i v a s t a v a , , V.M.L. , Ghatak, S. and Krishna Murt i , C.R. (1970b) .
Paras i to logy 60, 157-180.
Tanaka, H. , Takaoka, M., Matsuda, H. and Hayashi , Y. (1981) .
J a p . J . E x p . Med. 51 , 195.
Ta r to ra , P . , Hanozet, G.M. and Andrica , G. (1985) . Anal. Biochem.
144, 179-185.
Ueda, T. and Sawada, T. (1968) . J a p . J . E x p . Med. , 38, 145.
Van den Bossche , H. (1969) . Comp. Biochem. P h y s i o l . 3 1 , 789-
797.

72
Van den Bossche, H. (1976). Biochemistry of P a r a s i t e s and Host
Pa ras i t Re la t ionsh ip , 553-572-
Van den Bossche , H. (1981). Ann. Soc. b e i g e . Med. t r o p . , 61 ,
287-296.
Van Hof, L . , Henrard , C , Pee l , £ . and Wanson, M. (1947) . Annals
Soc. beige Med. t r o p . , 27, 173-177.
Van Nueten, J .M. (1972). Compjirative Biochemist ry of P a r a s i t e s
Ed . by H. Van den Bossche, 101-115 Academic P r e s s , N.Y.
Von Brand, T. (1973) Cited in "Biochemistry of P a r a s i t e s " , 2nd
E d n . , Academic P r e s s , New York.
Walter, R.D. (1979) . Tropenmed and P a r a s i t o l .
Walter, R.D. and Shulzkey, H. (1980). T r o p . Med. P a r a s i t o l . ,
3 1 , 55-58.
Wang, C.C. (1982) . Trends in Biochem. Sc i . 7, 354.
Wang, E . J . and Saz, H.J . (1974). J . P a r a s i t o l . , 60, 316.
Ward, C.W., Cas t ro , C.A. and F a i r b a i r n , D. (1969) . J . P a r a s i t o l .
55, 67 -71 .
Ward, C.W. and Schofield, P . J . (1967) . Comp. Biochem. P h y s i o l . ,
23, 356.
Ward, C.W., Schofie ld , P . J . and Johns tone , I . L . (1968a) . Comp.
Biochem. P h y s i o l . 24, 643.
Ward, C.W., Schofield, P . J . and Johns tone , I . L . (1968b) . Comp.
Biochem. P h y s i o l . , 26,
Watts, S.D.M. and Fa i rba i rn , F . (1974) . J . Exp .Zoo l . ,183,365-374.
Wilkes , J . , Corn i sh , R.A. and Met t r ick , D . F . (1981) . J . P a r a s i t o l . ,67 ,
832-840.
Related Documents











