GENETICS | INVESTIGATION Regulation of CTP Synthase Filament Formation During DNA Endoreplication in Drosophila Pei-Yu Wang,* ,† Wei-Cheng Lin, † Yi-Cheng Tsai, ‡ Mei-Ling Cheng, §, ** Yu-Hung Lin, ‡ Shu-Heng Tseng, ‡ Archan Chakraborty, ‡ and Li-Mei Pai* ,†,‡,††,1 *Department of Biochemistry, † Molecular Medicine Research Center, ‡ Graduate Institute of Biomedical Sciences, College of Medicine, § Department of Biomedical Sciences, **Healthy Aging Research Center, Chang Gung University, and, †† Chang Gung Memorial Hospital, Kwei-Shan, Tao-Yuan, 333 Taiwan ORCID ID: 0000-0002-1417-6432 (L.-M.P.) ABSTRACT CTP synthase (CTPsyn) plays an essential role in DNA, RNA, and lipid synthesis. Recent studies in bacteria, yeast, and Drosophila all reveal a polymeric CTPsyn structure, which dynamically regulates its enzymatic activity. However, the molecular mechanism underlying the formation of CTPsyn polymers is not completely understood. In this study, we found that reversible ubiquitination regulates the dynamic assembly of the filamentous structures of Drosophila CTPsyn. We further determined that the proto-oncogene Cbl, an E3 ubiquitin ligase, controls CTPsyn filament formation in endocycles. While the E3 ligase activity of Cbl is required for CTPsyn filament formation, Cbl does not affect the protein levels of CTPsyn. It remains unclear whether the regulation of CTPsyn filaments by Cbl is through direct ubiquitination of CTPsyn. In the absence of Cbl or with knockdown of CTPsyn, the progression of the endocycle-associated S phase was impaired. Furthermore, overexpression of wild-type, but not enzymatically inactive CTPsyn, rescued the endocycle defect in Cbl mutant cells. Together, these results suggest that Cbl influences the nucleotide pool balance and controls CTPsyn filament formation in endocycles. This study links Cbl-mediated ubiquitination to the polymerization of a metabolic enzyme and reveals a role for Cbl in endocycles during Drosophila development. KEYWORDS Cbl; CTP synthase; cytoophidia; endocycle; Drosophila P ROVIDING the raw material for DNA/RNA replication is a key challenge faced by all organisms during rapid growth. One particularly interesting example of this challenge is poly- ploidy, which arises from endoreplication and is essential for the normal development and specific physiological conditions of many diploid organisms. Organisms often use endoreplica- tion, a type of cell cycle encompassing genomic replication without cell division, to provide nutrients to support the de- veloping egg/embryo and enlarged cell size. Examples of cells and tissues that endocycle include mammalian trophoblast giant cells, plant seeds/roots, and Drosophila egg chambers (Edgar and Orr-Weaver 2001; Lee et al. 2009). In Drosophila, embryogenesis requires the mass production of proteins and high metabolic activity, which are achieved through endore- plication in the terminally differentiated cells of the develop- ing egg chamber. Drosophila oogenesis provides an excellent system for ana- lyzing developmentally controlled endoreplication. Egg pro- duction takes place within 16-cell germline cysts, with the asymmetric and incomplete division of a germline stem cell (Calvi and Spradling 1999). After cyst formation, nurse cells immediately exit the mitotic cycle and begin a series of 10–12 endocycles to reach 512C DNA content to provide proteins and messenger RNAs (mRNAs) for the developing oocyte. Each germline cyst is enveloped by 15–20 somatic follicle cells that divide mitotically to form an epithelial monolayer of 1000 cells and then employ three endocycles to reach 16C DNA content during stages 7–10A, the so-called “endocycle stages” (Klusza and Deng 2011). Endoreplication in the follicular ep- ithelium ensures a large amount of eggshell protein produc- tion in ,24 h (Lilly and Spradling 1996; Calvi et al. 1998). The endocycle in the follicle cells ceases at stage 10B, but some specific genomic foci (e.g., chorion genes) continue to amplify Copyright © 2015 by the Genetics Society of America doi: 10.1534/genetics.115.180737 Manuscript received July 15, 2015; accepted for publication October 13, 2015; published Early Online October 19, 2015. Supporting information is available online at www.genetics.org/lookup/suppl/ doi:10.1534/genetics.115.180737/-/DC1. 1 Corresponding author: Chang Gung University, 259 Wen-Hwa 1st Rd., Kwei-Shan, Tao-Yuan, 333 Taiwan. E-mail:[email protected] Genetics, Vol. 201, 1511–1523 December 2015 1511

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GENETICS | INVESTIGATION
Regulation of CTP Synthase Filament FormationDuring DNA Endoreplication in Drosophila
Pei-Yu Wang,*,† Wei-Cheng Lin,† Yi-Cheng Tsai,‡ Mei-Ling Cheng,§,** Yu-Hung Lin,‡ Shu-Heng Tseng,‡
Archan Chakraborty,‡ and Li-Mei Pai*,†,‡,††,1
*Department of Biochemistry,†Molecular Medicine Research Center,‡Graduate Institute of Biomedical Sciences, College ofMedicine,§Department of Biomedical Sciences, **Healthy Aging Research Center, Chang Gung University, and, ††Chang Gung
Memorial Hospital, Kwei-Shan, Tao-Yuan, 333 Taiwan
ORCID ID: 0000-0002-1417-6432 (L.-M.P.)
ABSTRACT CTP synthase (CTPsyn) plays an essential role in DNA, RNA, and lipid synthesis. Recent studies in bacteria, yeast, andDrosophila all reveal a polymeric CTPsyn structure, which dynamically regulates its enzymatic activity. However, the molecular mechanismunderlying the formation of CTPsyn polymers is not completely understood. In this study, we found that reversible ubiquitination regulatesthe dynamic assembly of the filamentous structures of Drosophila CTPsyn. We further determined that the proto-oncogene Cbl, an E3ubiquitin ligase, controls CTPsyn filament formation in endocycles. While the E3 ligase activity of Cbl is required for CTPsyn filamentformation, Cbl does not affect the protein levels of CTPsyn. It remains unclear whether the regulation of CTPsyn filaments by Cbl isthrough direct ubiquitination of CTPsyn. In the absence of Cbl or with knockdown of CTPsyn, the progression of the endocycle-associatedS phase was impaired. Furthermore, overexpression of wild-type, but not enzymatically inactive CTPsyn, rescued the endocycle defect inCbl mutant cells. Together, these results suggest that Cbl influences the nucleotide pool balance and controls CTPsyn filament formationin endocycles. This study links Cbl-mediated ubiquitination to the polymerization of a metabolic enzyme and reveals a role for Cbl inendocycles during Drosophila development.
KEYWORDS Cbl; CTP synthase; cytoophidia; endocycle; Drosophila
PROVIDING the rawmaterial for DNA/RNA replication is akey challenge faced by all organisms during rapid growth.
One particularly interesting example of this challenge is poly-ploidy, which arises from endoreplication and is essential forthe normal development and specific physiological conditionsof many diploid organisms. Organisms often use endoreplica-tion, a type of cell cycle encompassing genomic replicationwithout cell division, to provide nutrients to support the de-veloping egg/embryo and enlarged cell size. Examples of cellsand tissues that endocycle include mammalian trophoblastgiant cells, plant seeds/roots, and Drosophila egg chambers(Edgar and Orr-Weaver 2001; Lee et al. 2009). In Drosophila,embryogenesis requires the mass production of proteins and
high metabolic activity, which are achieved through endore-plication in the terminally differentiated cells of the develop-ing egg chamber.
Drosophila oogenesis provides an excellent system for ana-lyzing developmentally controlled endoreplication. Egg pro-duction takes place within 16-cell germline cysts, with theasymmetric and incomplete division of a germline stem cell(Calvi and Spradling 1999). After cyst formation, nurse cellsimmediately exit the mitotic cycle and begin a series of 10–12endocycles to reach 512CDNA content to provide proteins andmessenger RNAs (mRNAs) for the developing oocyte. Eachgermline cyst is enveloped by 15–20 somatic follicle cells thatdivide mitotically to form an epithelial monolayer of �1000cells and then employ three endocycles to reach 16C DNAcontent during stages 7–10A, the so-called “endocycle stages”(Klusza and Deng 2011). Endoreplication in the follicular ep-ithelium ensures a large amount of eggshell protein produc-tion in,24 h (Lilly and Spradling 1996; Calvi et al. 1998). Theendocycle in the follicle cells ceases at stage 10B, but somespecific genomic foci (e.g., chorion genes) continue to amplify
Copyright © 2015 by the Genetics Society of Americadoi: 10.1534/genetics.115.180737Manuscript received July 15, 2015; accepted for publication October 13, 2015;published Early Online October 19, 2015.Supporting information is available online at www.genetics.org/lookup/suppl/doi:10.1534/genetics.115.180737/-/DC1.1Corresponding author: Chang Gung University, 259 Wen-Hwa 1st Rd., Kwei-Shan,Tao-Yuan, 333 Taiwan. E-mail:[email protected]
Genetics, Vol. 201, 1511–1523 December 2015 1511

synchronously, an event called gene amplification (Calvi et al.1998). Notch signaling is responsible for the mitotic cycle–endocycle transition of follicle cells (Deng et al. 2001; Lopez-Schier and St. Johnston 2001), which activates the CyclinE/Cyclin-Dependent Kinase 2 (CycE/Cdk2) complex to triggerthe endocycle transition (Shcherbata et al. 2004).
This rapid series of endoreplication events requires cells tohave sufficient stores of the raw materials for DNA synthesis.CTP synthase (CTPsyn) produces CTP to facilitate DNA andRNA synthesis. In both prokaryotes and eukaryotes, CTPsyn isallosterically bound to GTP, activating glutamine hydrolysis togenerate ammonia (Long and Pardee 1967; Long et al. 1970;Levitzki and Koshland 1972). Subsequently, CTPsyn catalyzesthe ATP-dependent transfer of ammonia from glutamine to theC-4 position of UTP to formCTP (Lieberman 1956; Chakrabortyand Hurlbert 1961; Levitzki and Koshland 1971; von der Saalet al. 1985; Endrizzi et al. 2004). Under low concentrations ofCTPsyn or in the absence of ATP/UTP/CTP, CTPsyn is presentas an inactive monomer. With an increasing concentration ofCTPsyn, CTPsyn initially forms inactive dimers and then formsactive tetramers in the presence of ATP/UTP/CTP (Anderson1983; von der Saal et al. 1985). Therefore, CTPsyn monitorscellular nucleotide pools through its four NTP-binding sites,allowing it to match its activity to the concentration of nucle-otides (Aronow and Ullman 1987).
Recently, filamentous CTPsyn structures were indepen-dently revealed in bacteria, budding yeast, Drosophila, andmammalian cell lines, representing a novel, evolutionarily con-served cellular structure (Ingerson-Mahar et al. 2010; Liu2010; Noree et al. 2010; Carcamo et al. 2011). In Caulobactercrescentus, the CTPsyn filaments (CtpS filaments) interact withintermediate filaments to regulate bacterial shape (Ingerson-Mahar et al. 2010). In budding yeast, CTP synthase filamentsare promoted under the condition of carbon source depletion(Noree et al. 2010). In Drosophila, this structure, termed the“cytoophidia,” is present in a number of tissues, including thesalivary gland, testis, mid-gut, and ovary (Buszczak et al. 2007;Liu 2010). Both in yeast and Drosophila, the formation of afilament composed of CTPsyn does not require active tetra-mers, arguing that the filament is composed of inactive CTPsyndimers or monomers (Aughey et al. 2014; Noree et al. 2014).However, CTPsyn filaments in bacteria are composed of aninactive form of tetramers (Barry et al. 2014). In mammals,this structure, termed “rods and rings” (RR), appears in boththe cytoplasm and the nucleus (Gou et al. 2014) and acts in acell cycle-independent manner (Carcamo et al. 2011). The RRstructure contains not only CTPsyn, but also inosine mono-phosphase dehydrogenase 2 (IMPDH2), a key enzyme inGTP biosynthesis (Carcamo et al. 2011). Recently, the RRstructure was recognized as reflecting the concentration ofglutamine, an essential amide nitrogen donor in the nucleotidebiosynthesis pathway. The depletion of glutamine forced theformation of the RR structure in mammalian cells (Calise et al.2014; Gou et al. 2014).
Despite this strikinglybroadevolutionaryconservation, thefunction of these filamentous structures and the regulation of
their assembly remain elusive. During Drosophila oogenesis,germline cells of the ovary contain two different sizes ofCTPsyn filaments in one cell; they can be classified into micro-cytoophidia (1–6 mm) and macro-cytoophidia (10–50 mm)(Liu 2010).While we know that increasing the level of CTPsyninDrosophila enhances the formation of cytoophidia structuresand that CTPsyn activity seems to correlate with the structurein yeast, bacteria, and Drosophila (Chen et al. 2011; Azzamand Liu 2013; Aughey et al. 2014; Barry et al. 2014; Noreeet al. 2014; Strochlic et al. 2014), othermechanisms regulatingtheir assembly remain unknown.
We previously analyzed the function of the proto-oncogeneCasitas B-lineage lymphoma (Cbl), which is conserved inCaenorhabditis elegans, Drosophila, and mammalian cells. Itencodes an E3 ligase that plays a negative role in the receptortyrosine kinase pathway through its ubiquitin ligase activity(Marmor and Yarden 2004). Ubiquitination of the epidermalgrowth factor receptor (EGFR) promotes endosomal sortingand lysosomal degradation of this protein (Chang et al. 2008;Wang and Pai 2011). In this role, Cbl determines the polarity ofthe developing egg by negatively regulating the EGFR in folliclecells (Pai et al. 2000). Cbl can also regulate Notch signaling, anactivity related to its short isoform, Cbl-S, during eye and wingdevelopment (Wang et al. 2010). Cbl has also been implicatedin regulating cell proliferation in the germline, although themechanism is not known (Epstein et al. 2009). Here, we de-scribe a novel role for Cbl in promoting the endocycle throughregulating CTPsyn activity. Moreover, we reveal that reversiblemodification of ubiquitin regulates the dynamic assembly ofCTPsyn structures, the cytoophidia. Together, our data demon-strate a potential role for cytoophidia during Drosophila devel-opment and begin to uncover the mechanisms regulatingCTPsyn filament assembly.
Materials and Methods
Fly stocks and genetics
The strainsused included the following:wild-type (Oregon-R);CblF165/TM3Ser (Pai et al. 2000); CblBB28/TM3Ser (Paiet al. 2006); UAS-l-top (Queenan et al. 1997); Prosb61/TM3Sb (Saville and Belote 1993); Hs83-D-Cbl-L-PRDrk-75(Wang and Pai 2011); Hs83-D-CblS-2 (Pai et al. 2006);UAS-HA-ubiquitin (a gift from C. T. Chien, Institute of Molec-ular Biology, Academia Sinica, Taiwan); UAS-Dicer2 (Bloom-ington Stock Center); UAS-CTP synthaseGD12759 (ViennaDrosophila RNAi Center); and GR1, CY2, and 55B-Gal4(Queenan et al. 1997). For the genetic interaction assay,OreR, Prosb61/ +, and CblF165/ Prosb61 were incubated for3 days at 18� and then at 29� for 2 days. e22cFLP/+;CblF165FRT80B (or CblBB28FRT80B) /hs-mycFRT80B and hs-FLPl22/X; CblF165FRT2A /ovoDFRT2A were used for generat-ing mosaic clones as described previously (Pai et al. 2000).hs-FLPl22/X; CblF165FRT80B/[ubi-nlsRFP]FRT80B was used togenerate mosaic clones in the salivary glands. Eggs, 2.5–3.5 hr old, were collected, heat-shocked at 37� for 1.5 hr,and developed to 96-hr third instar larvae.
1512 P.-Y. Wang et al.

Antibodies
The primary antibodies used for immunohistochemistry in-cluded rabbit anti-CTPsyn (1:300; y-88, sc-134457; SantaCruz), rat anti-HA (1:500; Roche 3F10), mouse anti-MPM2(1:200;Millipore), mouse anti-Cut (1:50; 2B10);mouse anti-Hnt (1:15 1G9); mouse anti-Flag (1:500; Sigma F1804), andrabbit anti-Myc (1:200; Santa Cruz), while the secondaryantibodies included Alexa Fluor 488 or 546 anti-mouse/rabbit/rat antibodies (Molecular Probes). To visualize DNA,0.5 mg/mL DAPI (Sigma) was added for 5 min at roomtemperature. To visualize actin, phalloidin (1:100;MolecularProbes) was used. The antibodies used for immunoblottingwere anti-D-Cbl 10F1 ascites (Pai et al. 2006), rabbit anti-dCTPsyn, and mouse anti-tubulin (Sigma).
Drug treatment
For MG132 treatment, ovaries were dissected in Schneider’sinsect medium (Gibco) and then were incubated in mediumcontaining 60 mMMG132 (20 mM stock dissolved in DMSO)or DMSO (as a control) for 2 hr at room temperature. ForPr619 plus MG132 treatment, ovaries were dissected inSchneider’s medium and then incubated in medium contain-ing 50 mM Pr619 (20 mM stock was dissolved in DMSO) orDMSO for 2 hr at room temperature. After this, the ovarieswere transferred to medium containing 60 mM MG132 orDMSO and incubated for another 2 hr at room temperature.After drug treatment, the ovaries were fixed with 4% para-formaldehyde and analyzed by immunostaining.
CTPsyn inhibitor treatment
DON (6-diazo-5-oxo-L-norleucine) (D2141, Sigma) was solu-bilized at 500 mg/ml in water. Adult females were yeasted for3–5 days and then fed onwet yeast with orwithout 500mg/mlof DON for 24 hr, as previously described (Chen et al. 2011).
Bromodeoxyuridine incorporation assays
Ovaries and salivary glands were collected in Schneider’smedium and then incubated in a medium containing30 mg/ml bromodeoxyuridine (BrdU) (Sigma) at room tem-perature for 1 h. The BrdU-incorporated tissues were fixed in200 ml of 4% formaldehyde containing 600 ml heptane and0.25% NP-40 for 20 min at room temperature. The sampleswere treated with 2 N HCl for 30 min, neutralized with100 mM sodium borate, and washed with PBT three times.For BrdU detection, the samples were incubated with PBTcontaining 10% horse serum and then with anti-BrdU anti-body (1:50, Becton Dickinson) at 4� overnight. Alexa Fluor488 goat anti-mouse (1:1000; Molecular Probes) was used asthe secondary antibody.
Quantification of BrdU-positive cells
For quantification of the percentage of BrdU-positive cells inthe Cbl clone, the number of BrdU-positive cells was dividedby the total cell number in the Cbl clone or its twin clone. Tocalculate the percentage of BrdU-positive cells for the geneticinteraction experiment, we counted BrdU-positive cells in the
first five rows of the anterior and posterior follicle cells peregg chamber in 55B-Gal4::UAS-CTPsynRNAi, 55B-Gal4::UAS-GFP, OreR,CY2-Gal4 in Cbl heterozygous mutants andCY2-Gal4::UAS-CTPsynRNA iwith or without Cbl heterozygousmutants and then divided this number by the total number ofcells in the five rows.
Transgenic line generation and induction
To generate Flag-CTPsyn, a complementary DNA (cDNA)fragment of CTPsyn isoform C was amplified by PCR froman ovarian cDNA library with the specific primers CTPsyn-C-F59-GTCTCGAGATGAAATACATCCTGGTAAC-39and CTPsyn-C-R 59-GTCTCGAGTTACTTATGGCCATTGGTAG-39. TheCTPsyn fragment was cloned into pUAST-Flag using XhoIand XbaI restriction enzymes and was in-frame fused to the39 end of the Flag-tag. For Flag-CTPsynC399G construction, weemployed a mutagenesis technique to generate the mutated-CTPsyn fragments using the following specific primers: dCTPSCt1195g C399G F 59-CCATTGCTTGGCATCGGCTTGGGTCTGCAAG -39 and dCTPSC t1195g C399G R 59-CTTGCAGACCCAAGCCGATGCCAAGCAATGG-39. The DNA sequence of theproduct was confirmed before microinjection. For wild-type ex-pression, hs-Gal4::UAS-Flag-CTPsyn transgenic flies were heat-shocked at 37� for 30 min and then incubated for 3 hr at 25�.Subsequently, ovaries were dissected and analyzed by immuno-staining. For expression in Cbl mosaic egg chambers, hsFlp/X;hs-Gal4/UAS-Flag-CTPsyn; CblFRT80B/MycFRP80Bwere heat-shocked for 30 min/day. After 2 days of induction and incuba-tion, the expression level of Flag-CTPsyn and occurrences ofthe S phase were examined by immunoblotting and by BrdU-incorporation assay, respectively.
Antibody generation
A total of 1–118 amino acids from cg45070 (dCTPsyn inFlybase) was amplified by KOD-plus- DNA polymerase usingthe paired primers 59-TAAGCAGAATTCATGAAATACATCCTGGTAACTG-39 and 59-ATTGCTCTCGAGAGTGATGTGTGGGACAACTTG-39 to clone the peptide into a pET32a vector, and 63His fusion protein was purified by Ni-NTA beads (Qiagen). An-tibodies were obtained by immunizing rabbit followed by affin-ity purification and then were used for Western blotting(1:1000).
Results
Ubiquitination is involved in the regulation of CTPsynfilamentous structure formation
An important insight into the developmental role of CTPsynfilaments can be derived from the identification of the bio-chemical switch that regulates its assembly. RNA silencing ofCTPsyn led to thedisassemblyofCTPsynfilaments, suggestingthat CTPsyn protein levels regulate the CTPsyn filamentstructure in a dose-dependent manner (Chen et al. 2011).To examine this hypothesis further, we treated Drosophilaovaries with MG132, a proteasome inhibitor that shouldblock the degradation of CTPsyn, leading to an increase in
Cbl Regulates CTPsyn Filament 1513

the number of CTPsyn structures. Surprisingly, a reduction inCTPsyn filament formation was found in both the germlinecells and the follicle cells of egg chambers treated withMG132, compared to that in the DMSO control (Figure 1, Aand B). The length of macro-cytoophidium in control eggchambers was 24.8 6 7 mm on average, and this lengthwas reduced to 11.6 6 3.9 mm in egg chambers treated withMG132 (Figure 1I). On average, almost every follicle cellcontained one CTPsyn filament (0.836 0.05/per cell, Figure1J), and a reduction in the number of filaments was observedwithMG132 treatment (0.246 0.14/cell, Figure 1J). Despitethis observation, not all egg chambers were affected (Sup-porting Information, Figure S1D). The phenotype, however,did occur in all stages of oogenesis. Moreover, the inhibitoryeffect of MG132 on CTPsyn filament formation occurred in adose-dependent manner with a higher dose resulting in astronger inhibitory response (Figure S1, D and E). A similar,but somewhat less severe phenotype was observed in eggchambers expressing exogenous Flag-CTPsyn induced byhs-Gal4 (Figure S1, A and B). Nevertheless, the CTPsyn pro-tein levels were comparable before and after MG132 treat-ment (Figure S1C).
We reasoned that MG132 treatment could result in twoconsequences: namely, blockage of protein degradation anddepletion of free ubiquitin (Melikova et al. 2006). Given thatCTPsyn protein levels remained largely unaffected, we nextinvestigated whether overexpression of ubiquitin could sup-press the effect of MG132. Indeed, selective expression ofexogenous ubiquitin in follicle cells driven by CY2-Gal4 sig-nificantly reduced, from 0.24 6 0.14/cell to 0.9 6 0.1/cell,the MG132 effect on CTPsyn filament numbers (Figure 1J),indicating that ubiquitination may play a positive role in theregulation of CTPsyn filaments. In this in vitro-cultured ovaryassay, the gradual disassembly of CTPsyn filaments (a “nil”phenotype that completely lacked filament) was observed in�20% of egg chambers at 2 hr and in�40% of egg chambersat 4 hr in medium containing DMSO (Figure 1K and FigureS1D). To further confirm that ubiquitin modification mayplay a positive role in filament formation, we next examinedthe effect of a deubiquitinase-inhibitor, Pr619. Indeed, theCTPsyn filament structures were preserved with Pr619 pre-treatment for 2 hr followed by MG132 treatment (Figure 1,E–H and K). Together, these results suggest that ubiquitina-tion is required to maintain CTPsyn filaments.
Since Cbl is an E3 ubiquitin ligase in follicle cells (Pai et al.2000), we next investigated the role of Cbl in ubiquitin-mediated regulation of CTPsyn filaments using a genetic in-teraction assay with the Prosb61 mutant. We used the alleleProsb61, a temperature-sensitive proteasome subunit mutant(Holden and Suzuki 1973; Saville and Belote 1993), and aCbl null mutant, CblF165 (Pai et al. 2000). At a nonpermissivetemperature (29�), this dominant-negative Prosb61 allele byitself did not show any significant difference from wild type(Figure 1L). However, the combination of CblF165 andProsb61 displayed a synergistic effect on the negative regu-lation of CTPsyn filament formation (Figure 1, M–P and L),
whereas the Cbl heterozygous mutant did not show signifi-cant effects. On average, every follicle cell contains 0.9, 1.0,and 0.9 CTPsyn filaments in OreR, CblF165/+, and Prosb61/+egg chambers, respectively, but the number of CTPsyn fila-ments was reduced to 0.5 filaments/cell in CblF165/Prosb61
egg chambers at a nonpermissive temperature (29�) (Figure1L). Collectively, these results suggest that ubiquitinationand Cbl may regulate the formation of CTPsyn filamentstructures.
Since disruption of Cbl E3 ligase activity enhances EGFRsignaling in follicle cells (Pai et al. 2000;Wang and Pai 2011),we further assessed whether Cbl regulates CTPsyn filamentstructures through EGFR signaling. The CTPsyn filament wasnot affected by the overexpression of constitutively activatedEGFR (l-top) (Figure 1, Q–T) nor by activated Ras (RasV12)(Figure S1, F–F99) in follicle cells. In summary, Cbl regulatesthe formation of filament structures composed of CTPsynthrough its E3 ligase activity and appears to do so indepen-dently of EGFR signaling.
Cbl regulates CTPsyn filaments without affectingCTPsyn protein levels
To elucidate the role of Cbl in regulating the formation ofCTPsyn filament structure, mosaic Cbl mutant clones weregenerated in the follicular epithelium. In wild-type egg cham-bers, at least one CTPsyn filament was detected in everyfollicle cell (Figure 2, A and B) of stages 9–10 egg chambers.In contrast, few CTPsyn filaments were seen in Cbl mutantfollicle cells (Figure 2, A–A99 and B).We quantified the effectson CTPsyn filament formation by classifying the structures ineach cell as (1) normal, (2) reduced in CTPsyn filamentlength (Figure 2D, “reduced”), or (3) the complete absenceof CTPsyn filaments (Figure 2E, “nil”). A reduced phenotypewas seen in 57% of Cblmutant follicle cells, while 18% lackedCTPsyn filaments at stage 9. The defects became more severeat stage 10, when 36% of Cbl mutant cells exhibited a “re-duced” phenotype and 58% showed a nil phenotype (Figure2F). Similar defects in CTPsyn filaments were also ob-served in follicle cell mutants with another null allele of Cbl(CblBB28) (Figure S2, A–A9) (Pai et al. 2006). However,CTPsyn filaments were maintained as normal in Cbl-mutantcells when these cells were still in the mitotic stages of eggchamber development (before stage 6) (Figure S2, B–B9). Todetermine if this effect was specific for follicle cells, we ex-amined another cell type that undergoes endocycles and thatshows visible CTPsyn filaments, i.e., the salivary glands (Fig-ure 2G). Strikingly, CTPsyn filaments were also absent in thesalivary glands of the Cbl mutant (Figure 2, H–H9). In con-trast, the structure of the macro-cytoophidia in germline cells(nurse cells and the oocyte) was not obviously affected by theloss of Cbl (Figure S2D). The effects of Cbl on CTPsyn fila-ment were confirmed by rescue experiments in which weectopically expressed one of the two Cbl isoforms, CblL orCblS, in Cbl mutant follicle cells. Because overexpression ofCblL induces lethality during development (Pai et al. 2006),we used D-CblL-PRDrk, a full-length CblL carrying a point
1514 P.-Y. Wang et al.
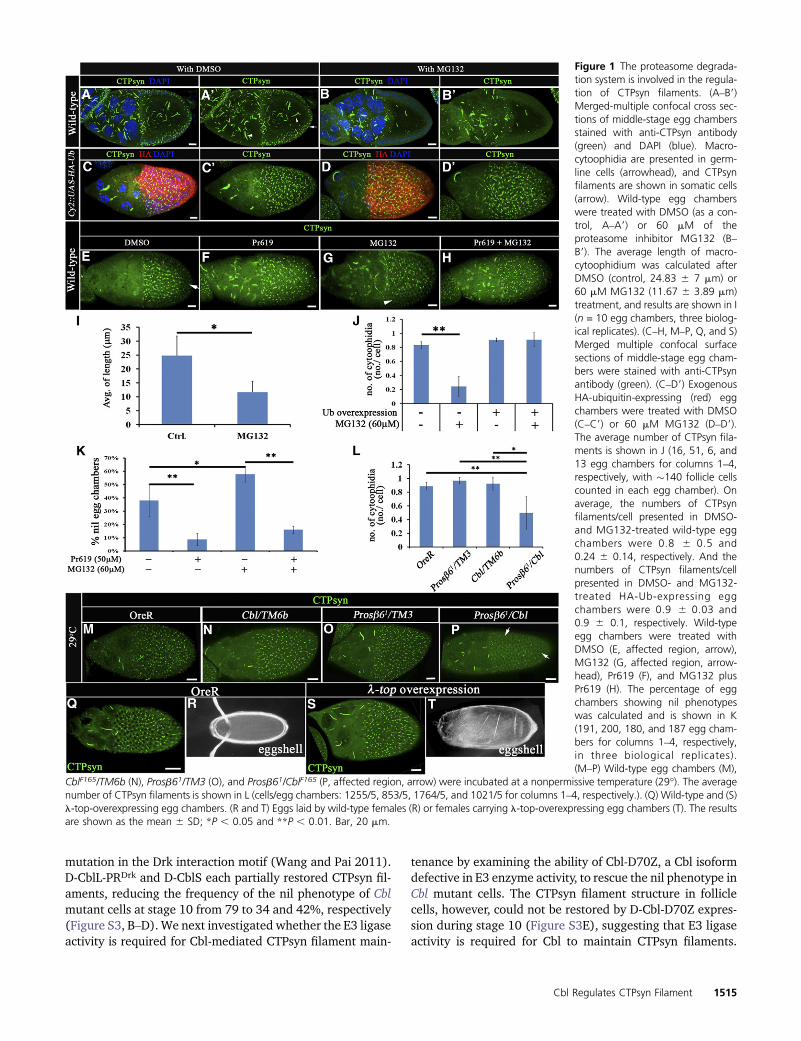
mutation in the Drk interaction motif (Wang and Pai 2011).D-CblL-PRDrk and D-CblS each partially restored CTPsyn fil-aments, reducing the frequency of the nil phenotype of Cblmutant cells at stage 10 from 79 to 34 and 42%, respectively(Figure S3, B–D). We next investigated whether the E3 ligaseactivity is required for Cbl-mediated CTPsyn filament main-
tenance by examining the ability of Cbl-D70Z, a Cbl isoformdefective in E3 enzyme activity, to rescue the nil phenotype inCbl mutant cells. The CTPsyn filament structure in folliclecells, however, could not be restored by D-Cbl-D70Z expres-sion during stage 10 (Figure S3E), suggesting that E3 ligaseactivity is required for Cbl to maintain CTPsyn filaments.
Figure 1 The proteasome degrada-tion system is involved in the regula-tion of CTPsyn filaments. (A–B9)Merged-multiple confocal cross sec-tions of middle-stage egg chambersstained with anti-CTPsyn antibody(green) and DAPI (blue). Macro-cytoophidia are presented in germ-line cells (arrowhead), and CTPsynfilaments are shown in somatic cells(arrow). Wild-type egg chamberswere treated with DMSO (as a con-trol, A–A9) or 60 mM of theproteasome inhibitor MG132 (B–B9). The average length of macro-cytoophidium was calculated afterDMSO (control, 24.83 6 7 mm) or60 mM MG132 (11.67 6 3.89 mm)treatment, and results are shown in I(n = 10 egg chambers, three biolog-ical replicates). (C–H, M–P, Q, and S)Merged multiple confocal surfacesections of middle-stage egg cham-bers were stained with anti-CTPsynantibody (green). (C–D9) ExogenousHA-ubiquitin-expressing (red) eggchambers were treated with DMSO(C–C9) or 60 mM MG132 (D–D9).The average number of CTPsyn fila-ments is shown in J (16, 51, 6, and13 egg chambers for columns 1–4,respectively, with �140 follicle cellscounted in each egg chamber). Onaverage, the numbers of CTPsynfilaments/cell presented in DMSO-and MG132-treated wild-type eggchambers were 0.8 6 0.5 and0.24 6 0.14, respectively. And thenumbers of CTPsyn filaments/cellpresented in DMSO- and MG132-treated HA-Ub-expressing eggchambers were 0.9 6 0.03 and0.9 6 0.1, respectively. Wild-typeegg chambers were treated withDMSO (E, affected region, arrow),MG132 (G, affected region, arrow-head), Pr619 (F), and MG132 plusPr619 (H). The percentage of eggchambers showing nil phenotypeswas calculated and is shown in K(191, 200, 180, and 187 egg cham-bers for columns 1–4, respectively,in three biological replicates).(M–P) Wild-type egg chambers (M),
CblF165/TM6b (N), Prosb61/TM3 (O), and Prosb61/CblF165 (P, affected region, arrow) were incubated at a nonpermissive temperature (29�). The averagenumber of CTPsyn filaments is shown in L (cells/egg chambers: 1255/5, 853/5, 1764/5, and 1021/5 for columns 1–4, respectively.). (Q) Wild-type and (S)l-top-overexpressing egg chambers. (R and T) Eggs laid by wild-type females (R) or females carrying l-top-overexpressing egg chambers (T). The resultsare shown as the mean 6 SD; *P , 0.05 and **P , 0.01. Bar, 20 mm.
Cbl Regulates CTPsyn Filament 1515
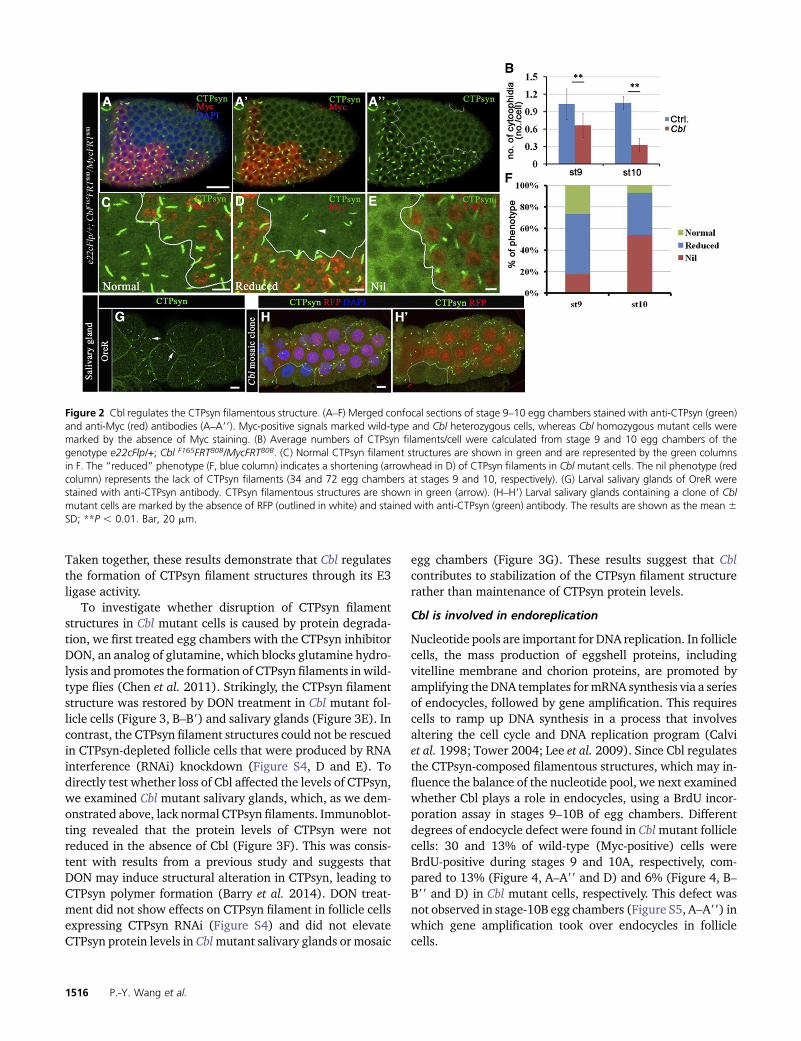
Taken together, these results demonstrate that Cbl regulatesthe formation of CTPsyn filament structures through its E3ligase activity.
To investigate whether disruption of CTPsyn filamentstructures in Cbl mutant cells is caused by protein degrada-tion, we first treated egg chambers with the CTPsyn inhibitorDON, an analog of glutamine, which blocks glutamine hydro-lysis and promotes the formation of CTPsyn filaments in wild-type flies (Chen et al. 2011). Strikingly, the CTPsyn filamentstructure was restored by DON treatment in Cbl mutant fol-licle cells (Figure 3, B–B9) and salivary glands (Figure 3E). Incontrast, the CTPsyn filament structures could not be rescuedin CTPsyn-depleted follicle cells that were produced by RNAinterference (RNAi) knockdown (Figure S4, D and E). Todirectly test whether loss of Cbl affected the levels of CTPsyn,we examined Cbl mutant salivary glands, which, as we dem-onstrated above, lack normal CTPsyn filaments. Immunoblot-ting revealed that the protein levels of CTPsyn were notreduced in the absence of Cbl (Figure 3F). This was consis-tent with results from a previous study and suggests thatDON may induce structural alteration in CTPsyn, leading toCTPsyn polymer formation (Barry et al. 2014). DON treat-ment did not show effects on CTPsyn filament in follicle cellsexpressing CTPsyn RNAi (Figure S4) and did not elevateCTPsyn protein levels in Cblmutant salivary glands or mosaic
egg chambers (Figure 3G). These results suggest that Cblcontributes to stabilization of the CTPsyn filament structurerather than maintenance of CTPsyn protein levels.
Cbl is involved in endoreplication
Nucleotide pools are important for DNA replication. In folliclecells, the mass production of eggshell proteins, includingvitelline membrane and chorion proteins, are promoted byamplifying the DNA templates formRNA synthesis via a seriesof endocycles, followed by gene amplification. This requirescells to ramp up DNA synthesis in a process that involvesaltering the cell cycle and DNA replication program (Calviet al. 1998; Tower 2004; Lee et al. 2009). Since Cbl regulatesthe CTPsyn-composed filamentous structures, which may in-fluence the balance of the nucleotide pool, we next examinedwhether Cbl plays a role in endocycles, using a BrdU incor-poration assay in stages 9–10B of egg chambers. Differentdegrees of endocycle defect were found in Cblmutant folliclecells: 30 and 13% of wild-type (Myc-positive) cells wereBrdU-positive during stages 9 and 10A, respectively, com-pared to 13% (Figure 4, A–A99 and D) and 6% (Figure 4, B–B99 and D) in Cbl mutant cells, respectively. This defect wasnot observed in stage-10B egg chambers (Figure S5, A–A99) inwhich gene amplification took over endocycles in folliclecells.
Figure 2 Cbl regulates the CTPsyn filamentous structure. (A–F) Merged confocal sections of stage 9–10 egg chambers stained with anti-CTPsyn (green)and anti-Myc (red) antibodies (A–A99). Myc-positive signals marked wild-type and Cbl heterozygous cells, whereas Cbl homozygous mutant cells weremarked by the absence of Myc staining. (B) Average numbers of CTPsyn filaments/cell were calculated from stage 9 and 10 egg chambers of thegenotype e22cFlp/+; Cbl F165FRT80B/MycFRT80B. (C) Normal CTPsyn filament structures are shown in green and are represented by the green columnsin F. The “reduced” phenotype (F, blue column) indicates a shortening (arrowhead in D) of CTPsyn filaments in Cbl mutant cells. The nil phenotype (redcolumn) represents the lack of CTPsyn filaments (34 and 72 egg chambers at stages 9 and 10, respectively). (G) Larval salivary glands of OreR werestained with anti-CTPsyn antibody. CTPsyn filamentous structures are shown in green (arrow). (H–H9) Larval salivary glands containing a clone of Cblmutant cells are marked by the absence of RFP (outlined in white) and stained with anti-CTPsyn (green) antibody. The results are shown as the mean 6SD; **P , 0.01. Bar, 20 mm.
1516 P.-Y. Wang et al.
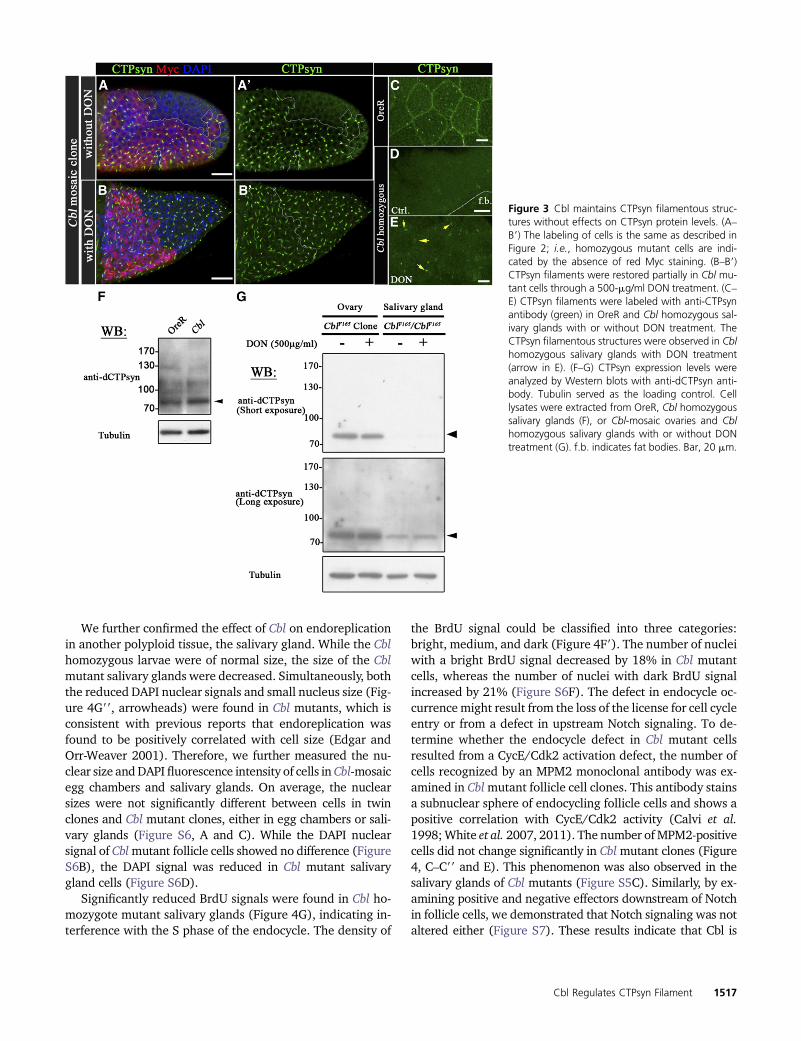
We further confirmed the effect of Cbl on endoreplicationin another polyploid tissue, the salivary gland. While the Cblhomozygous larvae were of normal size, the size of the Cblmutant salivary glands were decreased. Simultaneously, boththe reduced DAPI nuclear signals and small nucleus size (Fig-ure 4G99, arrowheads) were found in Cbl mutants, which isconsistent with previous reports that endoreplication wasfound to be positively correlated with cell size (Edgar andOrr-Weaver 2001). Therefore, we further measured the nu-clear size andDAPI fluorescence intensity of cells in Cbl-mosaicegg chambers and salivary glands. On average, the nuclearsizes were not significantly different between cells in twinclones and Cbl mutant clones, either in egg chambers or sali-vary glands (Figure S6, A and C). While the DAPI nuclearsignal of Cblmutant follicle cells showed no difference (FigureS6B), the DAPI signal was reduced in Cbl mutant salivarygland cells (Figure S6D).
Significantly reduced BrdU signals were found in Cbl ho-mozygote mutant salivary glands (Figure 4G), indicating in-terference with the S phase of the endocycle. The density of
the BrdU signal could be classified into three categories:bright, medium, and dark (Figure 4F9). The number of nucleiwith a bright BrdU signal decreased by 18% in Cbl mutantcells, whereas the number of nuclei with dark BrdU signalincreased by 21% (Figure S6F). The defect in endocycle oc-currence might result from the loss of the license for cell cycleentry or from a defect in upstream Notch signaling. To de-termine whether the endocycle defect in Cbl mutant cellsresulted from a CycE/Cdk2 activation defect, the number ofcells recognized by an MPM2 monoclonal antibody was ex-amined in Cblmutant follicle cell clones. This antibody stainsa subnuclear sphere of endocycling follicle cells and shows apositive correlation with CycE/Cdk2 activity (Calvi et al.1998;White et al. 2007, 2011). The number ofMPM2-positivecells did not change significantly in Cblmutant clones (Figure4, C–C99 and E). This phenomenon was also observed in thesalivary glands of Cbl mutants (Figure S5C). Similarly, by ex-amining positive and negative effectors downstream of Notchin follicle cells, we demonstrated that Notch signaling was notaltered either (Figure S7). These results indicate that Cbl is
Figure 3 Cbl maintains CTPsyn filamentous struc-tures without effects on CTPsyn protein levels. (A–B9) The labeling of cells is the same as described inFigure 2; i.e., homozygous mutant cells are indi-cated by the absence of red Myc staining. (B–B9)CTPsyn filaments were restored partially in Cbl mu-tant cells through a 500-mg/ml DON treatment. (C–E) CTPsyn filaments were labeled with anti-CTPsynantibody (green) in OreR and Cbl homozygous sal-ivary glands with or without DON treatment. TheCTPsyn filamentous structures were observed in Cblhomozygous salivary glands with DON treatment(arrow in E). (F–G) CTPsyn expression levels wereanalyzed by Western blots with anti-dCTPsyn anti-body. Tubulin served as the loading control. Celllysates were extracted from OreR, Cbl homozygoussalivary glands (F), or Cbl-mosaic ovaries and Cblhomozygous salivary glands with or without DONtreatment (G). f.b. indicates fat bodies. Bar, 20 mm.
Cbl Regulates CTPsyn Filament 1517

involved in endocycle regulation and may act downstream ofNotch signaling.
Depletion of CTPsyn disrupts endocycles in follicle cells
To further investigate whether CTPsyn activity is involved inthe endocycle process, we examined DNA replication under
CTPsyn depletion conditions. Dicer 2 (Dcr2) CTPsyn RNAi orGFP were driven by 55B-Gal4, which was expressed only inanterior follicle cells, allowing posterior follicle cells to serveas an internal control. Of the GFP-expressing control cells,41% were BrdU-positive (Figure 5, A–A9 and C), while inCTPsyn-depleted cells the percentage of BrdU-positive cells
Figure 4 Cbl is involved in theendocycle. (A–C99) Mosaic eggchambers containing Cbl mutantfollicle cell clones. Cells are labeledas described in Figure 2. (A–B99)BrdU incorporation is shown in thefollicular epithelium containing mo-saic clones for Cbl (Myc-negativecells, outlined in white) of stage 9(A–A99) and stage 10A (B–B99) eggchambers. The BrdU signal is ingreen; nuclei were stained withDAPI (blue). (C–C99) The MPM2 sig-nal (in green) was detected in Cblmutant cells and wild-type folliclecells in stage-9 egg chambers. (D)Percentages of BrdU-positive vs.Myc-negative cells (Cbl, light graycolumn) and Myc-positive twinclone cells with wild-type Cbl genes(Ctrl, dark gray column). Results areshown as the mean 6 SEM; *P ,0.05. There were 16 and 18 eggchambers at stages 9 and 10, re-spectively. N = 3. (E) MPM2 positiv-ity was assessed as described above;N.S. indicates not significant. (F–G99) BrdU incorporation (shown ingreen) in wild-type (F–F99) and inCbl homozygous (G–G99) salivaryglands. In F9, BrdU signals were clas-sified as bright (B), medium (M), ordark (D) (arrow). (F99 and G99) DNAwas stained with DAPI (blue), and areduction in nuclear size in Cbl mu-tant (arrowhead in G99) was ob-served. f.b. indicates fat bodies.Bar, 20 mm.
1518 P.-Y. Wang et al.

was decreased significantly to 2% (Figure 5, B–B9 and C). Thenumber of MPM2-positive cells did not decrease underCTPsyn-depleted conditions (Figure S5E), suggesting thatCTPsyn depletion could reduce DNA replication, but not en-try into the S phase. The depletion of CTPsyn could lead to animbalance in nucleotide pools, which could disrupt the endo-cycle event in Drosophila follicle cells. We further examinedthe relationship between Cbl and CTPsyn in endocycle regu-lation by investigating the endocycle in CTPsyn-depleted fol-licle cells in Cbl +/null mutants. The Cbl heterozygous andwild-type egg chambers showed similar percentages of BrdU-positive cells situated in the anterior follicular epithelium(Figure 5F). Silencing of CTPsyn without Dcr2 led to a re-duction in BrdU-positive cells from 33 to 22% (Figure 5, D–D9and F). The percentage of BrdU-positive cells decreased fur-ther to 16% (Figure 5, E–E9 and F) in a Cbl heterozygotebackground. In addition, the nuclear size of follicle cellswas reduced from 31.2 to 24.0 and 19.9 mm in CY2-Gal4/+;CTPsynRNAi/+ and CY2-Gal4/+;CTPsynRNAi /CblF165 stage-10A egg chambers, respectively (Figure S6E). Whereas Cblheterozygous females generally produced normal eggs (Fig-ure 5G), 21% of collapsed eggs were laid by females carryinga Cbl heterozygous mutation with CTPsyn depletion (Figure5H). Therefore, Cbl interacts genetically with CTPsyn to reg-ulate the endocycle. Taken together, Cbl may control theendocycle by maintaining the nucleotide pool balance inDro-sophila follicle cells.
Cbl regulates endocycle by modulating CTPsyn activity
If the endoreplication defect in the Cblmutant cell was due toan imbalance in nucleotides during the S phase, overexpres-sion of CTPsyn might increase the activity of CTPsyn, rescu-ing the endoreplication defect. We found that exogenouslyexpressed Flag-CTPsyn driven by hS-Gal4 could be incorpo-rated into the CTPsyn filament structure in wild-type cells(Figure 6A), but that a CTPsyn filament structure was notseen in Cbl mutant cells despite overexpression of CTPsyn(Figure 6C). Nevertheless, the BrdU assay showed that over-expression of Flag-CTPsyn restored DNA replication from 16to 25% in Cbl mutant cells (Figure 6, E–F99 and Figure S8F).These results suggest that in Cblmutant cells CTPsyn activitymight not be adequate and that overexpression of CTPsyncould rescue this defect. If true, the catalytically defectivemutant should not overcome the defect in Cblmutant. There-fore, we generated a UAS-Flag-CTPsynC399G mutant, a muta-tion homologous to the C404G Ura7p mutant in yeast, whichcannot bind its glutamine substrate (Noree et al. 2014). Inagreement with our prediction, we found that this mutantcould not restore the replication defect in Cbl mutant. Inter-estingly, this mutant also played a dominant-negative role inwild-type follicle cells (Figure 6G and Figure S8G), and inwild-type follicle cells, this Flag-CTPsynC399G mutant proteincould be incorporated into a filament structure that containsendogenous wild-type CTPsyn (Figure 6B). Moreover, theFlag- CTPsynC399G mutant still formed filament structuresin Cbl mutant follicle cells at similar expression levels to the
wild-type CTPsyn (Figure 6D and Figure S8A), suggestingthat the Flag-CTPsynC399G mutant protein escapes Cbl-mediated regulation in the process of CTPsyn filament for-mation. Glutamine depletion has been shown to induce theformation of CTPsyn polymers in a cell culture system (Caliseet al. 2014). A similar effect on CTPsyn filaments was foundin Drosophila S2 cells (Figure S8B9). To our surprise, thismutant could form filament structure in S2 cells in completeSchneider’s Drosophila medium (Figure S8B9), indicatingthat the polymerization process of this mutant may differfrom that of wild-type CTPsyn. In sum, Cbl influences endo-cycles by affecting the nucleotide pool balance and is re-quired for wild-type CTPsyn to form filaments.
Next, we investigated whether Cbl-mediated regulation offilament formation is throughdirect ubiquitination of CTPsyn.The lysates of Drosophila S2 cells expressing HA-ubiquitinand Flag-CTPsyn were subjected to immunoprecipitation us-ing anti-Flag antibody, and HA signals were detected in theimmunoprecipitation products (Figure S8C). Furthermore,lysates of ovarian and salivary gland cells expressing HA-ubiquitin and Flag-CTPsyn were analyzed under similar con-ditions. However, no ubiquitination signal on CTPsyn wasdetected in either ovaries or salivary glands (Figure S8, Dand E). Collectively, these results suggest that Cbl-mediatedregulation of CTPsyn filament formation in follicle cells orsalivary glands is not through direct ubiquitination of theCTPsyn protein itself.
Discussion
In this study, we revealed ubiquitin modification as a regula-torymechanism for the assembly of CTPsyn filaments through-out all stages of oogenesis (Figure 1 and Figure S1). Moreover,we identified Cbl as a specific E3 ubiquitin ligase that main-tains these structures during endocycles (Figure 2) and showedthat Cbl mutation resulted in a endoreplication-associateddefect in the endocycle S phase (Figure 4). We also demon-strated that CTPsyn activity is essential for endoreplication(Figure 5) and that overexpression of wild-type CTPsyn, butnot the catalytically defective mutant, is capable of rescuingthe endocycle defect in Cbl mutant cells (Figure 6, E–G, andFigure S8). These results suggest that the activity of CTPsyn isinsufficient to provide nucleotides for rapid endoreplicationin Cbl mutant cells. Disruption of the CTPsyn filamentousstructure in the absence of Cbl (Figure 2) may account forthe defects in endocycles.
The ubiquitin modification of proteins by polyubiquitina-tionormulti-monoubiquitination serves as a posttranslationalregulatory mechanism to control protein stability or protein–protein interactions, respectively (Sadowski et al. 2012). Inthe salivary glands of Cbl null larvae, the CTPsyn protein levelwas not diminished (Figure 3F), suggesting that Cbl andCTPsyn filament structures are not required for stabilizationof CTPsyn. However, the fact that D-Cbl-D70Z could not re-store CTPsyn filament structures in Cbl mutant cells indi-cates that the E3 ligase activity of Cbl is required for the
Cbl Regulates CTPsyn Filament 1519

maintenance of the CTPsyn filament structure. In addition,we found that reversible ubiquitination regulates the dy-namic assembly of both macro-and micro-cytoophidia inegg chambers. These results suggest that ubiquitinationmay provide a signal for polymerization of CTPsyn. Whilethe activities of several enzymes have been reported to becontrolled by ubiquitination-mediated protein degradation(Mattiroli and Sixma 2014) and by enzymatic inactivationthrough conformational changes (Sagar et al. 2007), theCTPsyn filament represents the first case in which an enzymepolymeric structure is regulated by ubiquitination.
Ubiquitin modification has been found in human CTPS1using a proteomic approach (Udeshi et al. 2013). Our dataindicate that Cbl-related ubiquitination of the CTPsyn proteinitself might not occur in Drosophila follicle cells or salivaryglands, suggesting that Cbl might act on an unknown proteinthat coordinates the formation of CTPsyn filament structuresin an ubiquitination-dependent manner (Figure S8, D and E).We initially detected a CTPsyn filament-like structure usingan antibody that was raised to recognize human phosphory-lated PKD peptide. This structurewas also recognized by anti-CTPsyn antibodies. Furthermore, even though DrosophilaPKD has been excluded from the CTPsyn complex, we believe
that CTPsyn is a complex that includes many types of en-zymes, such as inosine-59-monophosphate dehydrogenaseand other proteins. This notion is supported by the recentfinding that DAck colocalizes with CTPsyn filaments in germ-line cells (Strochlic et al. 2014).
Even thoughCbl regulates CTPsyn filament structures onlyin follicle cells and not in germline cells, our inhibitor treat-ment assays and other genetic approaches in ovaries andother tissues demonstrated that CTPsyn filaments in germ-line cells, follicle cells, and salivary glands are sensitive toubiquitin modification, suggesting that ubiquitination maybe a general mechanism for the regulation of CTPsyn fila-ment structures. Therefore, one possible explanation for thedifferent effects of Cbl is that different cells may controlCTPsynfilaments by different types of E3-mediated ubiquitina-tion. Furthermore, the role of Cbl in CTPsyn filament forma-tion seems to be crucial only in polyploid cells (Figure 2),such as follicle cells during the late endocycle stages (i.e.,stages 9–10A) and the salivary glands. In contrast, in diploidcells, such as follicle cells at the mitotic stage, CTPsyn fila-ments are normally maintained in Cbl mutant cells (FigureS2, B–B9). One possibility is that the endocycle places addi-tional regulations on CTPsyn filament formation in follicle
Figure 5 Cbl genetically inter-acts with CTPsyn. (A–B9) BrdU in-corporation (green) was seen instage-9 egg chambers expressingGFP (A–A9) or CTPsyndsRNA (B–B9)in their anterior follicle cells. (C)Quantification of BrdU-positivecells in the anterior and posteriorregions of GFP-expressing (bluebars) or CTPsyndsRNA-expressing(red bars) egg chambers. (D–E9)BrdU incorporation was seen infollicle cells with CTP depletionin wild-type (D and D9) and Cblheterozygous (E and E9) back-grounds. (F) Statistics for the per-centage of BrdU-positive cells inthe first five rows of anterior fol-licle cells/egg chamber with andwithout CTPsyndsRNA expressionin wild-type and Cbl heterozy-gotes. (G) Wild-type eggshellsand (H) collapsed eggshells laidby females with CTPsyn deple-tion in a Cbl heterozygous back-ground. DNA was stained withDAPI (blue). Results are shownas the mean 6 SEM; *P , 0.05and **P , 0.01; N.S. indicatesnot significant. The numbers inC and F indicate the numbers ofcells/egg chamber. Bar, 20 mm.
1520 P.-Y. Wang et al.
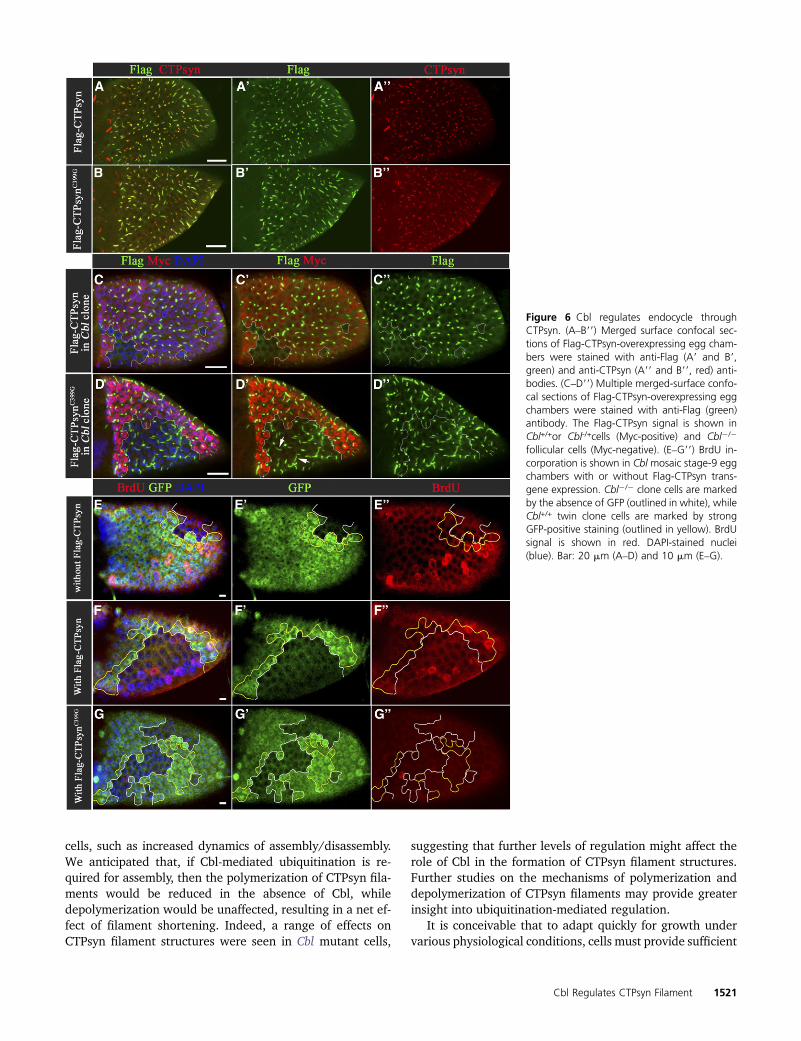
cells, such as increased dynamics of assembly/disassembly.We anticipated that, if Cbl-mediated ubiquitination is re-quired for assembly, then the polymerization of CTPsyn fila-ments would be reduced in the absence of Cbl, whiledepolymerization would be unaffected, resulting in a net ef-fect of filament shortening. Indeed, a range of effects onCTPsyn filament structures were seen in Cbl mutant cells,
suggesting that further levels of regulation might affect therole of Cbl in the formation of CTPsyn filament structures.Further studies on the mechanisms of polymerization anddepolymerization of CTPsyn filaments may provide greaterinsight into ubiquitination-mediated regulation.
It is conceivable that to adapt quickly for growth undervarious physiological conditions, cells must provide sufficient
Figure 6 Cbl regulates endocycle throughCTPsyn. (A–B99) Merged surface confocal sec-tions of Flag-CTPsyn-overexpressing egg cham-bers were stained with anti-Flag (A9 and B9,green) and anti-CTPsyn (A99 and B99, red) anti-bodies. (C–D99) Multiple merged-surface confo-cal sections of Flag-CTPsyn-overexpressing eggchambers were stained with anti-Flag (green)antibody. The Flag-CTPsyn signal is shown inCbl+/+or Cbl-/+cells (Myc-positive) and Cbl2/2
follicular cells (Myc-negative). (E–G99) BrdU in-corporation is shown in Cbl mosaic stage-9 eggchambers with or without Flag-CTPsyn trans-gene expression. Cbl2/2 clone cells are markedby the absence of GFP (outlined in white), whileCbl+/+ twin clone cells are marked by strongGFP-positive staining (outlined in yellow). BrdUsignal is shown in red. DAPI-stained nuclei(blue). Bar: 20 mm (A–D) and 10 mm (E–G).
Cbl Regulates CTPsyn Filament 1521

materials to synthesize DNA, RNA, and lipids. Consistent withthis interpretation, a special requirement for human CTPsynthase1 (CTPS1) activity in DNA replication was revealedrecently in receptor-mediated activation of T cells by anti-CD3and anti-CD28 antibodies (Martin et al. 2014). The synthesis ofphospholipids for membrane expansion is controlled by thelevels of CTP, and the research group of Jeffrey R. Petersonat Fox Chase Cancer Center, Philadelphia, has demonstrated arole for CTPsyn in regulating the phospholipid composition ofDrosophila germline cells (Chang and Carman 2008; Strochlicet al. 2014). Our results demonstrated that supplying the en-zymatic activity of CTPsyn can restore endoreplication defectsin Cbl mutant cells, indicating that Cbl may be involved inmaintaining the nucleotide balance. During endocycles DNAreplication reaches 16Cwithin 24 hr in follicle cells and 1024Cwithin 120 hr in the salivary glands. Loss of Cbl regulationresulted in a 50% reduction in DNA replication function (Fig-ure 4D and Figure S6F). This defect could not be detected bymeasurement of nuclear size (Figure S6, A and C); however, itwas reflected in the DAPI fluorescence signal in Cbl mutantsalivary gland cells (Figure S6D). Given that in Cbl mutantcells CTPsyn filament structures are reduced, one possible ex-planation for these results is that CTPsyn filament structuresare required to allow CTPsyn activity to meet the demand fornucleotide pools during endocycle. However, we still cannotexclude the possibility that Cbl regulates CTPsyn filamentstructures and CTPsyn activity independently.
Interestingly, CTPsynfilamentswere observed undermanyspecific physiological conditions. Nutritiondepletion seems tobe a common factor in the induction of this structure in yeast,brains of Drosophila larva, and mammalian cells (Augheyet al. 2014; Barry et al. 2014; Noree et al. 2010, 2014). InDrosophila S2 cells, we found that glutamine depletion in-duced filament formation, whereas normal Schneider’s me-dium allowed very little filament formation. Interestingly,also, the glutamine-binding defect mutant C399G, anenzymatically inactive CTPsyn in vivo, seemed to mimic thewild-type CTPsyn in glutamine-depleted conditions so thatfilaments were formed in normal medium (Figure S8, B9 andB99). Indeed, this mutant was not regulated by Cbl andformed filament structures in Cbl mutant cells. Given thatthis inactive mutant mingled with endogenous wild-typeCTPsyn to form filaments, we hypothesize that CTPsyn fila-ments may be composed of different types of CTPsyn, includ-ing active, inactive, or a mixture of both forms.
CTP biosynthesis is one of several biosynthesis pathwaysimplicated in liver malignancy and kidney carcinoma, andCTPsyn activity is increased in hepatomas (Williams et al.1978). As the transition from tetraploidy to aneuploidy isconsidered to be an important step in cancer development(Davoli and de Lange 2011), the role of the proto-oncogeneCbl in the regulation of CTPsyn activity during endocyclesimplies a new mechanism through which Cbl promotes can-cer formation. In this study, we defined a new role for theproto-oncogene Cbl in endocycles. We demonstrated thatCbl-mediated ubiquitination controls the assembly of CTPsyn
filaments and may regulate the nucleotide supply for rapidendoreplication in Drosophila.
Acknowledgments
We thank Hai-Wei Pi, Fly Core Taiwan, the Bloomington StockCenter, and the Vienna Stock Center for providing the reagentsand fly stocks; Trudi Schüpbach, Ji-Long Liu, Mark Peifer, JörgGrosshans, Cheng-Ting Chien, Hai-Wei Pi, Bertrand Tan, Scott C.Schuyler, Tzu-Chien Van Wang, Chien-Kuo Lee, and Shu-YuanYang for commenting on the manuscript; and members of thePai laboratory for their comments and helpful discussions. Thiswork was supported by grants from the National Science Councilof the Republic of China (NSC 101-2811-B-182-017 to P.-Y.W.and MOST103-2311-B-182-004-MY3 to L.-M.P.); the ChangGung Memorial Hospital (CMRPD-2B0061-3 to L.-M.P.); andthe Ministry of Education, Taiwan (EMRPD1E1421 to L.-M.P.).
Literature Cited
Anderson, P. M., 1983 CTP synthetase from Escherichia coli: animproved purification procedure and characterization of hyster-etic and enzyme concentration effects on kinetic properties. Bio-chemistry 22: 3285–3292.
Aronow, B., and B. Ullman, 1987 In situ regulation of mammalianCTP synthetase by allosteric inhibition. J. Biol. Chem. 262:5106–5112.
Aughey, G. N., S. J. Grice, Q. J. Shen, Y. Xu, C. C. Chang et al.,2014 Nucleotide synthesis is regulated by cytoophidium for-mation during neurodevelopment and adaptive metabolism.Biol. Open 3: 1045–1056.
Azzam, G., and J. L. Liu, 2013 Only one isoform of Drosophilamelanogaster CTP synthase forms the cytoophidium. PLoSGenet. 9: e1003256.
Barry, R. M., A. F. Bitbol, A. Lorestani, E. J. Charles, C. H. Habrianet al., 2014 Large-scale filament formation inhibits the activityof CTP synthetase. eLife 3: e03638.
Buszczak, M., S. Paterno, D. Lighthouse, J. Bachman, J. Planck et al.,2007 The carnegie protein trap library: a versatile tool for Dro-sophila developmental studies. Genetics 175: 1505–1531.
Calise, S. J., W. C. Carcamo, C. Krueger, J. D. Yin, D. L. Purich et al.,2014 Glutamine deprivation initiates reversible assembly ofmammalian rods and rings. Cell. Mol. Life Sci. 71: 2963–2973.
Calvi, B. R., and A. C. Spradling, 1999 Chorion gene amplificationin Drosophila: a model for metazoan origins of DNA replicationand S-phase control. Methods 18: 407–417.
Calvi, B. R., M. A. Lilly, and A. C. Spradling, 1998 Cell cyclecontrol of chorion gene amplification. Genes Dev. 12: 734–744.
Carcamo, W. C., M. Satoh, H. Kasahara, N. Terada, T. Hamazakiet al., 2011 Induction of cytoplasmic rods and rings structuresby inhibition of the CTP and GTP synthetic pathway in mam-malian cells. PLoS One 6: e29690.
Chakraborty, K. P., and R. B. Hurlbert, 1961 Role of glutamine inthe biosynthesis of cytidine nucleotides in Escherichia coli. Biochim.Biophys. Acta 47: 607–609.
Chang, W. L., W. Liou, H. C. Pen, H. Y. Chou, Y. W. Chang et al.,2008 The gradient of Gurken, a long-range morphogen, is di-rectly regulated by Cbl-mediated endocytosis. Development135: 1923–1933.
Chang, Y. F., and G. M. Carman, 2008 CTP synthetase and its rolein phospholipid synthesis in the yeast Saccharomyces cerevisiae.Prog. Lipid Res. 47: 333–339.
1522 P.-Y. Wang et al.

Chen, K., J. Zhang, O. Y. Tastan, Z. A. Deussen, M. Y. Siswick et al.,2011 Glutamine analogs promote cytoophidium assembly inhuman and Drosophila cells. J. Genet. Genomics 38: 391–402.
Davoli, T., and T. de Lange, 2011 The causes and consequences ofpolyploidy in normal development and cancer. Annu. Rev. CellDev. Biol. 27: 585–610.
Deng, W. M., C. Althauser, and H. Ruohola-Baker, 2001 Notch-Deltasignaling induces a transition from mitotic cell cycle to endocycle inDrosophila follicle cells. Development 128: 4737–4746.
Edgar, B. A., and T. L. Orr-Weaver, 2001 Endoreplication cell cy-cles: more for less. Cell 105: 297–306.
Endrizzi, J. A., H. Kim, P. M. Anderson, and E. P. Baldwin,2004 Crystal structure of Escherichia coli cytidine triphosphatesynthetase, a nucleotide-regulated glutamine amidotransferase/ATP-dependent amidoligase fusion protein and homologue of an-ticancer and antiparasitic drug targets. Biochemistry 43: 6447–6463.
Epstein, A. M., C. R. Bauer, A. Ho, G. Bosco, and D. C. Zarnescu,2009 Drosophila Fragile X protein controls cellular prolifera-tion by regulating cbl levels in the ovary. Dev. Biol. 330: 83–92.
Gou, K. M., C. C. Chang, Q. J. Shen, L. Y. Sung, and J. L. Liu,2014 CTP synthase forms cytoophidia in the cytoplasm andnucleus. Exp. Cell Res. 323: 242–253.
Holden, J. J., and D. T. Suzuki, 1973 Temperature-sensitive mu-tations in Drosophila melanogaster. XII. The genetic and devel-opmental effects of dominant lethals on chromosome 3.Genetics 73: 445–458.
Ingerson-Mahar, M., A. Briegel, J. N. Werner, G. J. Jensen, and Z.Gitai, 2010 The metabolic enzyme CTP synthase forms cyto-skeletal filaments. Nat. Cell Biol. 12: 739–746.
Klusza, S., and W. M. Deng, 2011 At the crossroads of differenti-ation and proliferation: precise control of cell-cycle changes bymultiple signaling pathways in Drosophila follicle cells. BioEs-says 33: 124–134.
Lee, H. O., J. M. Davidson, and R. J. Duronio, 2009 Endoreplication:polyploidy with purpose. Genes Dev. 23: 2461–2477.
Levitzki, A., and D. E. Koshland, Jr., 1971 Cytidine triphosphatesynthetase. Covalent intermediates and mechanisms of action.Biochemistry 10: 3365–3371.
Levitzki, A., and D. E. Koshland, Jr., 1972 Role of an allostericeffector. Guanosine triphosphate activation in cytosine triphos-phate synthetase. Biochemistry 11: 241–246.
Lieberman, I., 1956 Enzymatic amination of uridine triphosphateto cytidine triphosphate. J. Biol. Chem. 222: 765–775.
Lilly, M. A., and A. C. Spradling, 1996 The Drosophila endocycle iscontrolled by Cyclin E and lacks a checkpoint ensuring S-phasecompletion. Genes Dev. 10: 2514–2526.
Liu, J. L., 2010 Intracellular compartmentation of CTP synthase inDrosophila. J. Genet. Genomics 37: 281–296.
Long, C. W., and A. B. Pardee, 1967 Cytidine triphosphate syn-thetase of Escherichia coli B. I. Purification and kinetics. J. Biol.Chem. 242: 4715–4721.
Long, C. W., A. Levitzki, and D. E. Koshland, Jr., 1970 The sub-unit structure and subunit interactions of cytidine triphosphatesynthetase. J. Biol. Chem. 245: 80–87.
Lopez-Schier, H., and D. St. Johnston, 2001 Delta signaling from thegerm line controls the proliferation and differentiation of the somaticfollicle cells during Drosophila oogenesis. Genes Dev. 15: 1393–1405.
Marmor, M. D., and Y. Yarden, 2004 Role of protein ubiquityla-tion in regulating endocytosis of receptor tyrosine kinases. On-cogene 23: 2057–2070.
Martin, E., N. Palmic, S. Sanquer, C. Lenoir, F. Hauck et al.,2014 CTP synthase 1 deficiency in humans reveals its centralrole in lymphocyte proliferation. Nature 510: 288–292.
Mattiroli, F., and T. K. Sixma, 2014 Lysine-targeting specificity inubiquitin and ubiquitin-like modification pathways. Nat. Struct.Mol. Biol. 21: 308–316.
Melikova, M. S., K. A. Kondratov, and E. S. Kornilova, 2006 Twodifferent stages of epidermal growth factor (EGF) receptor en-docytosis are sensitive to free ubiquitin depletion produced byproteasome inhibitor MG132. Cell Biol. Int. 30: 31–43.
Noree, C., B. K. Sato, R. M. Broyer, and J. E. Wilhelm,2010 Identification of novel filament-forming proteins in Sac-charomyces cerevisiae and Drosophila melanogaster. J. Cell Biol.190: 541–551.
Noree, C., E. Monfort, A. K. Shiau, and J. E. Wilhelm,2014 Common regulatory control of CTP synthase enzyme ac-tivity and filament formation. Mol. Biol. Cell 25: 2282–2290.
Pai, L. M., G. Barcelo, and T. Schupbach, 2000 D-cbl, a negativeregulator of the Egfr pathway, is required for dorsoventral pat-terning in Drosophila oogenesis. Cell 103: 51–61.
Pai, L. M., P. Y. Wang, S. R. Chen, G. Barcelo, W. L. Chang et al.,2006 Differential effects of Cbl isoforms on Egfr signaling inDrosophila. Mech. Dev. 123: 450–462.
Queenan, A. M., A. Ghabrial, and T. Schupbach, 1997 Ectopic activa-tion of torpedo/Egfr, a Drosophila receptor tyrosine kinase, dorsalizesboth the eggshell and the embryo. Development 124: 3871–3880.
Sadowski, M., R. Suryadinata, A. R. Tan, S. N. Roesley, and B.Sarcevic, 2012 Protein monoubiquitination and polyubiquiti-nation generate structural diversity to control distinct biologicalprocesses. IUBMB Life 64: 136–142.
Sagar, G. D., B. Gereben, I. Callebaut, J. P. Mornon, A. Zeold et al.,2007 Ubiquitination-induced conformational change withinthe deiodinase dimer is a switch regulating enzyme activity.Mol. Cell. Biol. 27: 4774–4783.
Saville, K. J., and J. M. Belote, 1993 Identification of an essentialgene, l(3)73Ai, with a dominant temperature-sensitive lethalallele, encoding a Drosophila proteasome subunit. Proc. Natl.Acad. Sci. USA 90: 8842–8846.
Shcherbata, H. R., C. Althauser, S. D. Findley, and H. Ruohola-Baker,2004 The mitotic-to-endocycle switch in Drosophila follicle cellsis executed by Notch-dependent regulation of G1/S, G2/M andM/G1 cell-cycle transitions. Development 131: 3169–3181.
Strochlic, T. I., K. P. Stavrides, S. V. Thomas, E. Nicolas, A. M.O’Reilly et al., 2014 Ack kinase regulates CTP synthase fila-ments during Drosophila oogenesis. EMBO Rep. 15: 1184–1191.
Tower, J., 2004 Developmental gene amplification and origin reg-ulation. Annu. Rev. Genet. 38: 273–304.
Udeshi, N. D., T. Svinkina, P. Mertins, E. Kuhn, D. R. Mani et al.,2013 Refined preparation and use of anti-diglycine remnant(K-epsilon-GG) antibody enables routine quantification of10,000s of ubiquitination sites in single proteomics experiments.Mol. Cell. Proteomics 12: 825–831.
von der Saal, W., P. M. Anderson, and J. J. Villafranca,1985 Mechanistic investigations of Escherichia coli cytidine-59-triphosphate synthetase. Detection of an intermediate by po-sitional isotope exchange experiments. J. Biol. Chem. 260:14993–14997.
Wang, P. Y., and L. M. Pai, 2011 D-Cbl binding to Drk leads todose-dependent down-regulation of EGFR signaling and in-creases receptor-ligand endocytosis. PLoS One 6: e17097.
Wang, Y., Z. Chen, and A. Bergmann, 2010 Regulation of EGFRand Notch signaling by distinct isoforms of D-cbl during Dro-sophila development. Dev. Biol. 342: 1–10.
White, A. E., M. E. Leslie, B. R. Calvi, W. F. Marzluff, and R. J.Duronio, 2007 Developmental and cell cycle regulation of theDrosophila histone locus body. Mol. Biol. Cell 18: 2491–2502.
White, A. E., B. D. Burch, X. C. Yang, P. Y. Gasdaska, Z. Dominskiet al., 2011 Drosophila histone locus bodies form by hierarchi-cal recruitment of components. J. Cell Biol. 193: 677–694.
Williams, J. C., H. Kizaki, G. Weber, and H. P. Morris, 1978 IncreasedCTP synthetase activity in cancer cells. Nature 271: 71–73.
Communicating editor: R. J. Duronio
Cbl Regulates CTPsyn Filament 1523

GENETICSSupporting Information
www.genetics.org/lookup/suppl/doi:10.1534/genetics.115.180737/-/DC1
Regulation of CTP Synthase Filament FormationDuring DNA Endoreplication in Drosophila
Pei-Yu Wang, Wei-Cheng Lin, Yi-Cheng Tsai, Mei-Ling Cheng, Yu-Hung Lin, Shu-Heng Tseng,Archan Chakraborty, and Li-Mei Pai
Copyright © 2015 by the Genetics Society of AmericaDOI: 10.1534/genetics.115.180737

Figure S1. Inhibition of CTPsyn filament structures without altering CTPsyn
protein levels. (A–B’) Multiple merged-surface confocal sections of Flag-CTPsyn
overexpressing egg chambers treated with DMSO (control, A–A”) or 60 µM MG132

(B–B’) were stained with anti-Flag (green), anti-CTPsyn (red) and DAPI (blue,
nuclei). (C) Western blot analysis of Flag-CTPsyn protein expression by anti-Flag
antibody (arrow). Ovarian cell lysates were extracted from hsGal4::Flag-CTPsyn
after heat shock for 2 h or 4 h, and treated with DMSO (-) or increasing doses of
MG132. Tubulin was used as a loading control. Relative Flag-CTPsyn expression
levels were analyzed with a densitometer and normalized to the level at 2 h. (D)
Wild-type egg chambers were treated with DMSO, or 20 µM or 60 µM MG132. The
percentage of nil phenotypes (red column) was calculated for DMSO (Ctrl. 16.6%),
20 µM MG132 (28.3%), and 60 µM MG132 (42.7%) treatments. (E) The percentages
of nil phenotypes in stages 8–10A egg chambers with each indicated drug treatment
were calculated. (F–F”) Multiple merged-surface confocal sections of stage 9 RasV12
overexpressing (green) egg chambers stained with anti-CTPsyn (red) and DAPI (blue).
The results are shown as the mean ± s.d.; *p < 0.05 and **p < 0.01. Scale bar: 20 µm.
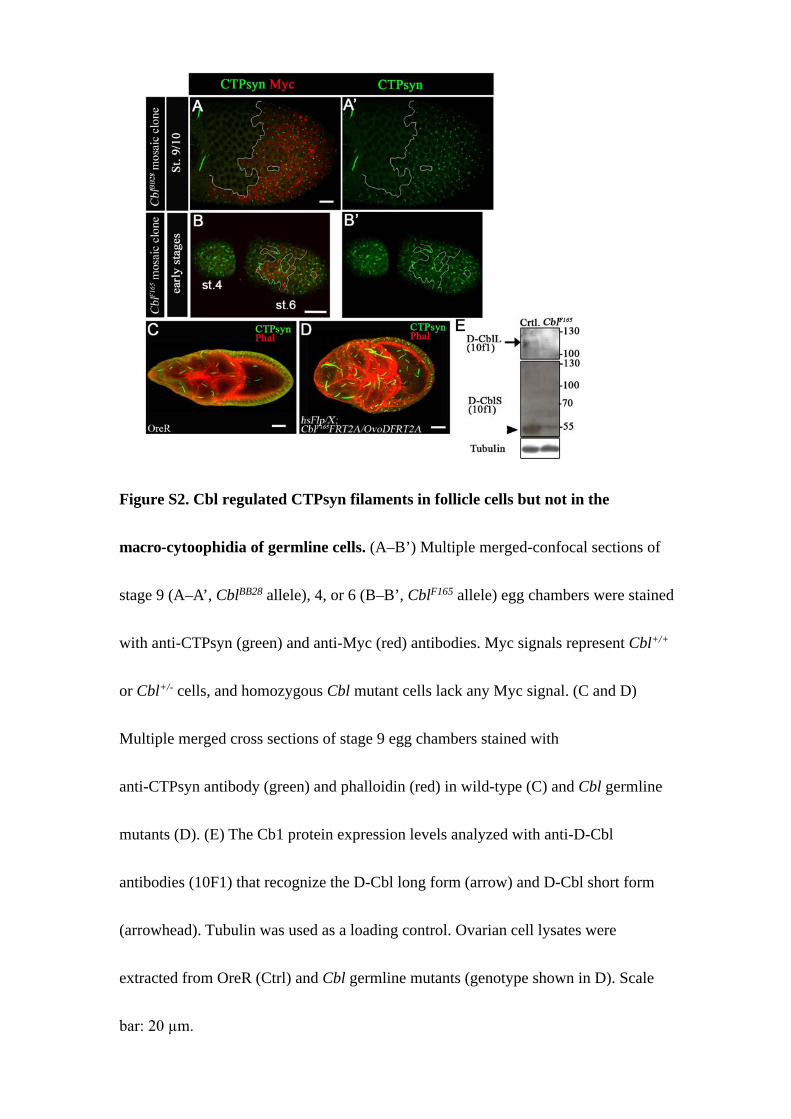
Figure S2. Cbl regulated CTPsyn filaments in follicle cells but not in the
macro-cytoophidia of germline cells. (A–B’) Multiple merged-confocal sections of
stage 9 (A–A’, CblBB28 allele), 4, or 6 (B–B’, CblF165 allele) egg chambers were stained
with anti-CTPsyn (green) and anti-Myc (red) antibodies. Myc signals represent Cbl+/+
or Cbl+/- cells, and homozygous Cbl mutant cells lack any Myc signal. (C and D)
Multiple merged cross sections of stage 9 egg chambers stained with
anti-CTPsyn antibody (green) and phalloidin (red) in wild-type (C) and Cbl germline
mutants (D). (E) The Cb1 protein expression levels analyzed with anti-D-Cbl
antibodies (10F1) that recognize the D-Cbl long form (arrow) and D-Cbl short form
(arrowhead). Tubulin was used as a loading control. Ovarian cell lysates were
extracted from OreR (Ctrl) and Cbl germline mutants (genotype shown in D). Scale
bar: 20 µm.
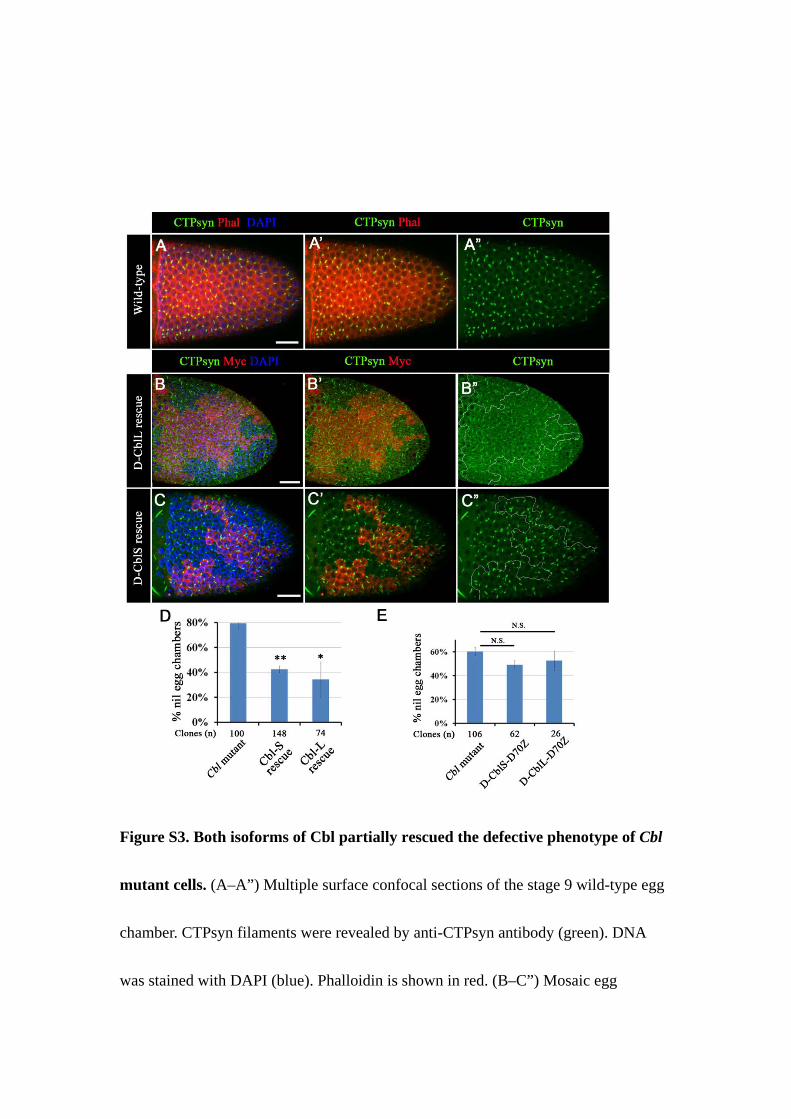
Figure S3. Both isoforms of Cbl partially rescued the defective phenotype of Cbl
mutant cells. (A–A”) Multiple surface confocal sections of the stage 9 wild-type egg
chamber. CTPsyn filaments were revealed by anti-CTPsyn antibody (green). DNA
was stained with DAPI (blue). Phalloidin is shown in red. (B–C”) Mosaic egg

chambers containing CblF165 mutant follicle cell clones were labeled as described in
Figure S2A. CTPsyn filaments were restored by constitutive expression of
CblL-PRDrk (B-B”) and CblS (C-C”) in Cbl mutant cells (Myc-negative). (D) The nil
phenotype was calculated in Cbl mutant cells with or without CblL-PRDrk or CblS
expression in stage 10 egg chambers. (E) The nil phenotype was calculated in Cbl
mutant cells with or without D-CblS-D70Z and D-CblL-D70Z rescued in stage 10.
Results are shown as the mean ± s.e.m; *p < 0.05 and **p < 0.01. N.S indicates not
significant. Scale bar: 20 µm.

Figure S4. CTPsyn filament formation depends on CTPsyn protein levels in
follicle cells. (A–E) Multiple confocal cross sections (A–A’, B–B’, D–D’) and
surface-sections (C, E) of stage 9 egg chambers were immunostained with
anti-CTPsyn antibody (green) and DAPI (blue). (A–A’) In the wild-type egg chamber,
macro-cytoophidia were presented in the germline cells (arrowhead) and cytoophidia
were presented in the main-body follicle cells (arrow). (B–C) Dcr2 and CTPsyn RNAi
were co-overexpressed in the main-body follicle cells to reduce the level of CTPsyn.
Macro-cytoophidia were not affected in the germline (arrowhead). (D–E)
CTPsyn-depleted egg chambers were treated with 500 µg/ml DON. Scale bar: 20 µm.
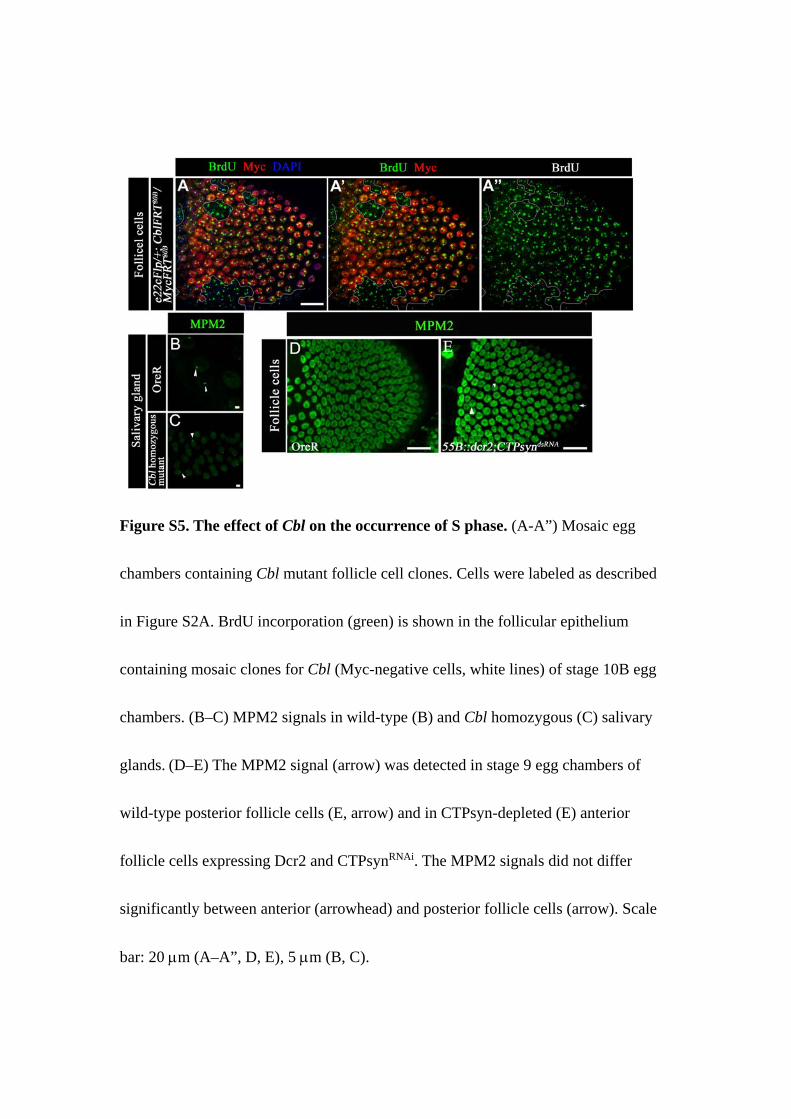
Figure S5. The effect of Cbl on the occurrence of S phase. (A-A”) Mosaic egg
chambers containing Cbl mutant follicle cell clones. Cells were labeled as described
in Figure S2A. BrdU incorporation (green) is shown in the follicular epithelium
containing mosaic clones for Cbl (Myc-negative cells, white lines) of stage 10B egg
chambers. (B–C) MPM2 signals in wild-type (B) and Cbl homozygous (C) salivary
glands. (D–E) The MPM2 signal (arrow) was detected in stage 9 egg chambers of
wild-type posterior follicle cells (E, arrow) and in CTPsyn-depleted (E) anterior
follicle cells expressing Dcr2 and CTPsynRNAi. The MPM2 signals did not differ
significantly between anterior (arrowhead) and posterior follicle cells (arrow). Scale
bar: 20 µm (A–A”, D, E), 5 µm (B, C).
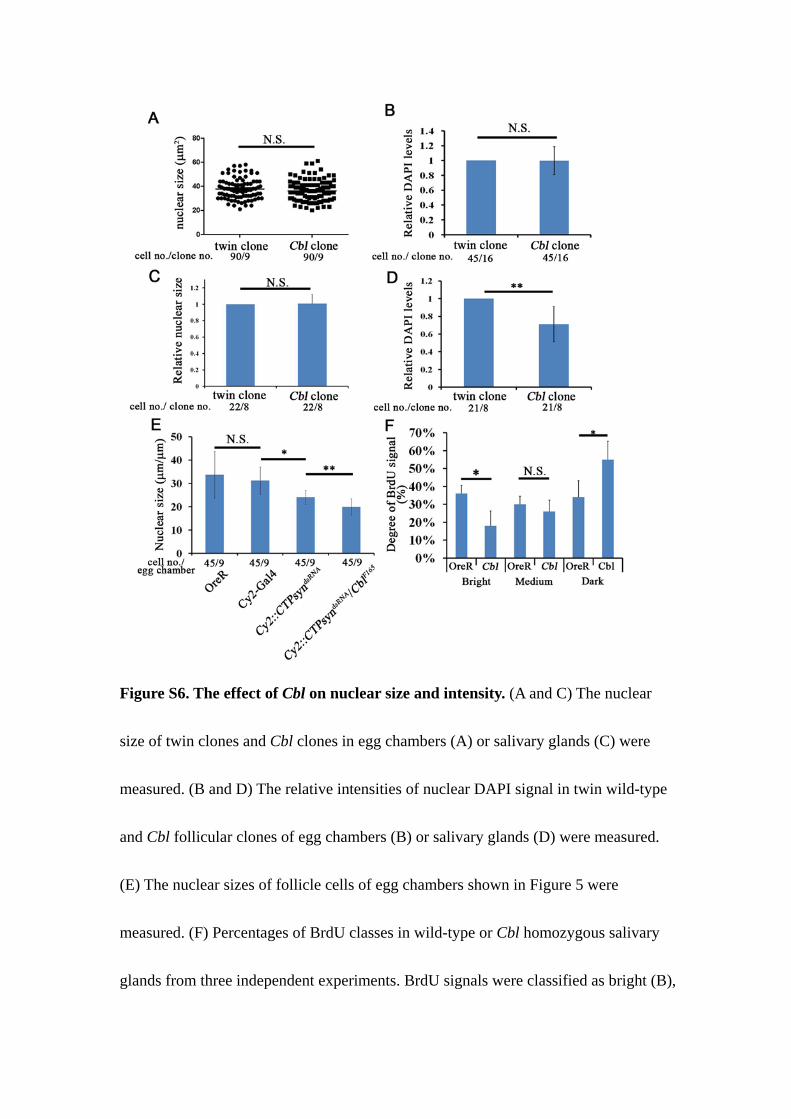
Figure S6. The effect of Cbl on nuclear size and intensity. (A and C) The nuclear
size of twin clones and Cbl clones in egg chambers (A) or salivary glands (C) were
measured. (B and D) The relative intensities of nuclear DAPI signal in twin wild-type
and Cbl follicular clones of egg chambers (B) or salivary glands (D) were measured.
(E) The nuclear sizes of follicle cells of egg chambers shown in Figure 5 were
measured. (F) Percentages of BrdU classes in wild-type or Cbl homozygous salivary
glands from three independent experiments. BrdU signals were classified as bright (B),

medium (M), or dark (D). Results are shown as the mean ± s.d.; *p < 0.05; ** p <
0.01; N.S.: not significant.


Figure S7. Notch signaling was not altered in Cbl mutant follicle cells.
(A–F”) Cut (white) and (G–L”) Hnt (white) staining in wild-type (A–A’, C–C’, E–E’,
G–G’, I–I’, and K–K’) or Cbl mosaic egg chambers (B–B”, D–D”, F–F”, H–H”, J–J”,
and L–L”). Cbl+/+ or Cbl+/- cells were Myc-tagged. (A–B”) Expression of Cut in
follicle cells of early-stage egg chambers (stages 1–6); (C–D”) at stage 9/10A; and
(E–F”) at stages 10B/11. (G–J”) Expression of Hnt at stages 7–10A when Notch is
activated and (K–L”) at stages 10B/11. Scale bar: 20 µm.
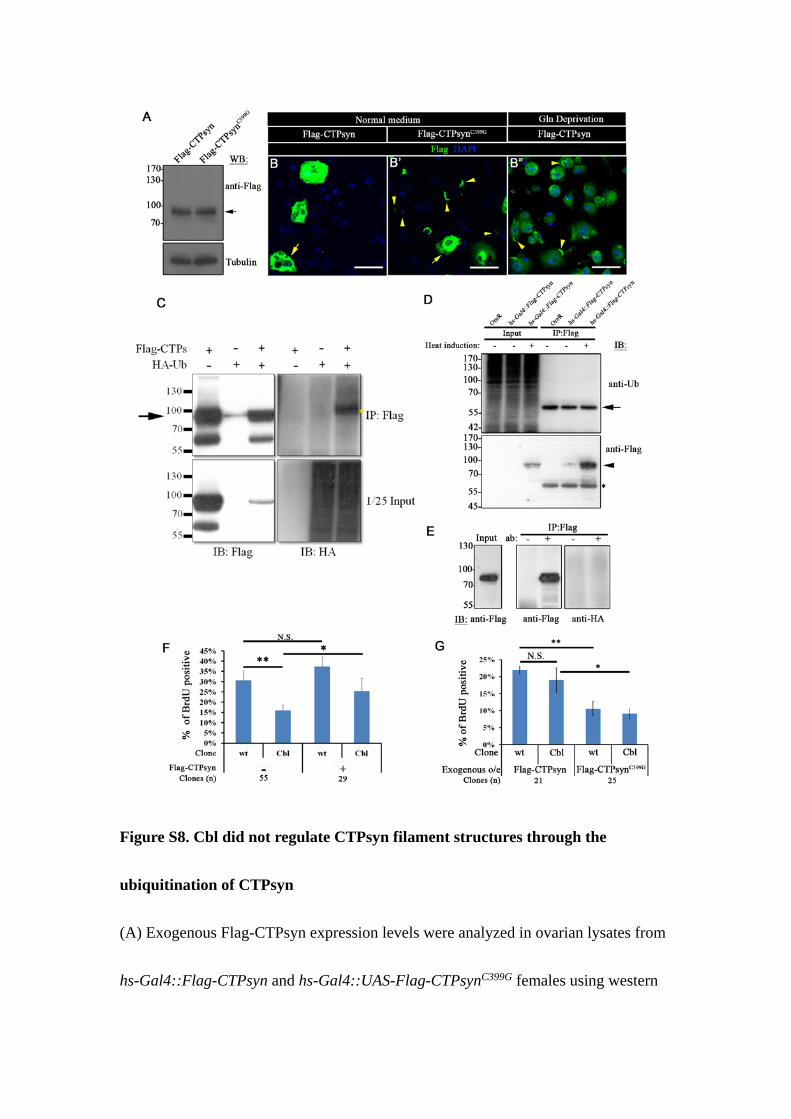
Figure S8. Cbl did not regulate CTPsyn filament structures through the
ubiquitination of CTPsyn
(A) Exogenous Flag-CTPsyn expression levels were analyzed in ovarian lysates from
hs-Gal4::Flag-CTPsyn and hs-Gal4::UAS-Flag-CTPsynC399G females using western

blot. Hs-Gal4 was first expressed at 37°C for 30 minutes and then switched to 25°C to
express UAS-Flag- CTPsyn for an additional 3 h before lysing the cells for western
blotting using anti-Flag antibody. Tubulin was used as a loading control. (B–B”)
Multiple merged-confocal sections of S2 cells incubated in complete (B–B’) or
glutamine-deficient (B”) Schneider’s medium were stained with anti-Flag (green) and
DAPI (blue). The CTPsyn filament structures were observed in Flag-CTPsynC399G-
transfected cells (arrowhead in B’) in complete Schneider’s medium, and in
Flag-CTPsyn-transfected cells in glutamine-deficient medium (arrowhead in B”).
Arrows indicate highly expressed cells. (C) Flag-CTPsyn was immunoprecipitated
from transfected S2 cells that were co-transfected with HA-ubiquitin using anti-Flag
antibody. The immunoprecipitated product was subjected to immunoblotting using
anti-HA (right) or anti-Flag antibodies (left). The arrow indicates Flag-CTPsyn. The
ubiquitinated protein signal is presented at the right (yellow star). (D and E)
Flag-CTPsyn was immunoprecipitated from ovarian (D) or salivary gland (E) cell
lysates by anti-Flag antibody. In this assay, ovaries expressed Flag-CTPsyn and
salivary glands co-expresses HA-ubiquitin and Flag-CTPsyn. The products were
subjected to immunoblotting using anti-Flag antibody (arrowhead in D), anti-ubiquitin
(D), or anti-HA (E) antibody. The arrow in D indicates a non-specific band, and the
heavy chains of antibodies are labeled by *. (F and G) The percentage of

BrdU-positive versus GFP-negative cells (white line in Figure 7), and GFP-positive
cells (yellow line in Figure 7) was quantified per egg chamber. The egg chambers
overexpressing Flag-CTPsyn or Flag-CTPsynC399G in main-body follicle cells. (n = 3
biological repeats). Scale bar: 20 µm. Results are shown as the mean ± s.d.; *p < 0.05;
**p < 0.01, and N.S.: not significant.

File S1. Supplemental Experimental Procedures
Cell culture and transfection
S2 cells were transfected with UAS-Flag-dCTPsyn C form and UAS-HA-Ubiquitin
using Effectine reagent (Qiagen), then cultured for 1 day in Schneider’s Drosophila
medium (Genaxxon) supplemented with 10% fetal bovine serum (FBS) and 1%
antibiotic-antimycotic (Invitrogen), with or without L-glutamine.
Immunoprecipitation
S2 cells were transfected with UAS-Flag-dCTPsyn C form and UAS-HA-Ubiquitin
using Effectine reagent (Qiagen), and then cultured in Schneider’s Drosophila
medium (Genaxxon) supplemented with 10% fetal bovine serum (FBS) and 1%
antibiotic-antimycotic (Invitrogen). A total of 1 x 107 S2 cells were homogenized
with lysis buffer. The ovarian lysates were extracted from ovaries of OreR and
hs-Gal4::CTPsyn females, which were heat-shocked at 37°C for 30 min, and then
incubated at 25°C for an additional 2 h before dissection. Salivary gland samples, 180
pairs, from 96h CY2-Gal4::HA-Ub; Flag-CTPsyn larvae were dissected in
Schneider’s Drosophila medium each time, and then stored at -80°C until reaching

1000 pairs (1 pair contains about 1 µg of proteins). Lysates were fractionated by
centrifugation at 12000 g for 10 min at 4°C. The supernatants were incubated with
anti-Flag antibody (1 µg/500 µg lysate) in lysis buffer (50 mM HEPES, 50 mM NaCl,
0.5% Doc, 1% Triton-100, 10 mM NEM, 5 mM EDTA, 1 mM EGTA, 10 mM NaF, 1
mM NaVO3, and 1× protease inhibitor) for 2 h at 4°C. Then, protein G sepharose
beads were added to bring down the immunoprecipitates for 1 h at 4°C. Pellets were
dissolved in 2× sample buffer that was boiled for 10 min. The samples were resolved
by SDS-PAGE, transferred to an NC membrane, and immunoblotted with anti-Flag
(Sigma), anti-ubiquitin (P4D1, Santa Cruz) and anti-HA (Roche) antibodies.
Quantification of nuclear size and DAPI intensity
The quantification results of follicle cell/salivary gland nuclear sizes and the DAPI
intensity of Cbl clone are shown in Figures S6A-D. In figure S6A, 90 pairs of cells of
twin clone and of Cbl follicular mutant clone were analyzed. The stage 10A egg
chamber confocal images came from 4 independent experiments. For DAPI intensity
in follicle cells, 90 nuclei of sixteen Cbl clones from 7 independent experiments were
analyzed. The relative intensity of DAPI signal was measured through dividing the
signal of Cbl mutant cells by that of adjacent wild type cells. The intensity was
determined by total intensity in a defined area divided by pixels inside the area. Since

the nucleus size of salivary gland was heterogenous along the proximal and distal axis,
the relative nuclear size was measured through dividing the size of Cbl mutant cell
nuclei by that of adjacent wild type cell nuclei. There are 22 pairs of cells in 8 clones
from 3 independent experiments were analyzed. Twenty-one nuclei of 8 salivary
glands from 4 independent experiments were analyzed on DAPI intensity. For nuclear
sizes in figure 5, the average nuclear size of 45 cells from each genotype (random 5
cells in each egg chamber were selected from 3 independent experiments) were
analyzed. The images were quantified using ZEISS LSM Image Browser software.
Related Documents











