GASTROENTEROLOGY 1996;111:45–55 Reg Gene Expression Is Increased in Rat Gastric Enterochromaffin-like Cells Following Water Immersion Stress MASAKYO ASAHARA,* SOTARO MUSHIAKE, ‡ SHOICHI SHIMADA, § HIROKAZU FUKUI,* YOSHIKAZU KINOSHITA,* CHIHARU KAWANAMI,* TSUYOSHI WATANABE, x SATOSHI TANAKA, Ø ATSUSHI ICHIKAWA, Ø YASUO UCHIYAMA, x YOICHI NARUSHIMA, # SHIN TAKASAWA, # HIROSHI OKAMOTO, # MASAYA TOHYAMA, § and TSUTOMU CHIBA* *Division of Gerontology, Department of Internal Medicine, Kobe University School of Medicine, Kobe; Departments of ‡ Pediatrics, § Anatomy and Neuroscience, and x Cell Biology and Anatomy I, Osaka University Medical School, Osaka; Ø Department of Physiological Chemistry, Faculty of Pharmaceutical Sciences, Kyoto University, Kyoto; and # Department of Biochemistry, Tohoku University School of Medicine, Sendai, Japan Background & Aims: Reg gene has been isolated from tory granules of beta cells 5 and that administration of regenerating rat pancreatic islets, and subsequent Reg protein not only enhances DNA synthesis in isolated studies have shown a trophic effect of Reg protein on islets but also ameliorates diabetes in rats subjected to islet cells. However, little is known about the role of 90% depancreatomy, suggesting a trophic role for the Reg Reg protein in the stomach. The aim of this study was gene in pancreatic beta cells. 6 Interestingly, the predicted to clarify the localization of Reg messenger RNA three-dimensional structure of the protein shows homol- (mRNA) and its product in the stomach and to examine ogy with that of the lectins, a family of growth-promot- changes in the level of their expression during regener- ing molecules. 7–9 ation of gastric mucosal cells. Methods: Gastric le- REG gene has also been found in the stomach. 10 Al- sions were experimentally induced in Sprague – Dawley though the precise localization and the role of the REG rats by water immersion stress. Northern blot analysis gene in the stomach remains undetermined, it is interest- and in situ hybridization studies were performed to ex- ing to speculate that it is involved in the process of amine changes in mRNA levels. Immunohistochemical studies were performed to identify the cellular localiza- gastric mucosal repair, as has been suggested to occur in tion and to investigate the change in Reg protein level. the pancreas. In the pancreas, REG protein is primarily Results: Reg mRNA and its product were distributed in found in the exocrine gland under normal conditions, 11 the basal part of the oxyntic mucosa and were ex- whereas it is mainly present in the islets during regenera- pressed mainly in enterochromaffin-like cells. Levels of tion. 5 The present study was therefore undertaken to both Reg mRNA and its product were markedly in- clarify what type of cells in the stomach express Reg gene creased in the water immersion–induced gastric le- and its product and to investigate changes in the level sions. Conclusions: Reg mRNA and its product are ex- of their expressions in water immersion – induced gastric pressed in gastric enterochromaffin-like cells, and their lesions. levels are increased during the healing process of water immersion–induced gastric lesions. Materials and Methods Animals I Male Sprague – Dawley rats (250 – 300 g) were deprived n 1984, Yonemura et al. 1 found that administration of poly(ADP-ribose) synthetase inhibitors, such as nic- of food but allowed free access to water for 24 hours before otinamide, to rats subjected to 90% depancreatomy in- the experiments. Each study was performed with 5 animals in duced regeneration of pancreatic islets. Subsequent each group. To induce gastric mucosal lesions, the animals screening of the regenerating rat islet-derived comple- were placed in strain cages and immersed in water (23C) for mentary DNA (cDNA) library showed a novel gene that 6 hours as described previously. 12 Water-immersed rats were encodes a 165–amino acid protein with a 21–amino killed 0, 6, 12, 24, 48, and 72 hours after the beginning of acid signal peptide. 2 The gene has been termed Reg and is abundantly expressed in regenerating islets but not in Abbreviations used in this paper: ECL cell, enterochromaffin-like cell; GSH, glutathione-S-transferase; HDC, histidine decarboxylase; normal ones. 2 Further studies showed that enhanced Reg ISH, in situ hybridization; RT-PCR, reverse-transcription polymerase gene expression is linked to DNA synthesis in pancreatic chain reaction. islets both in vitro 3 and in vivo. 4 Moreover, it has been 1996 by the American Gastroenterological Association 0016-5085/96/$3.00 shown recently that Reg protein is present in the secre-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GASTROENTEROLOGY 1996;111:45–55
Reg Gene Expression Is Increased in Rat GastricEnterochromaffin-like Cells Following Water Immersion Stress
MASAKYO ASAHARA,* SOTARO MUSHIAKE,‡ SHOICHI SHIMADA,§ HIROKAZU FUKUI,*YOSHIKAZU KINOSHITA,* CHIHARU KAWANAMI,* TSUYOSHI WATANABE,x SATOSHI TANAKA,Ø
ATSUSHI ICHIKAWA,Ø YASUO UCHIYAMA,x YOICHI NARUSHIMA,# SHIN TAKASAWA,#
HIROSHI OKAMOTO,# MASAYA TOHYAMA,§ and TSUTOMU CHIBA**Division of Gerontology, Department of Internal Medicine, Kobe University School of Medicine, Kobe; Departments of ‡Pediatrics, §Anatomyand Neuroscience, and xCell Biology and Anatomy I, Osaka University Medical School, Osaka; ØDepartment of Physiological Chemistry,Faculty of Pharmaceutical Sciences, Kyoto University, Kyoto; and #Department of Biochemistry, Tohoku University School of Medicine,Sendai, Japan
Background & Aims: Reg gene has been isolated from tory granules of beta cells5 and that administration ofregenerating rat pancreatic islets, and subsequent Reg protein not only enhances DNA synthesis in isolatedstudies have shown a trophic effect of Reg protein on islets but also ameliorates diabetes in rats subjected toislet cells. However, little is known about the role of 90% depancreatomy, suggesting a trophic role for the RegReg protein in the stomach. The aim of this study was gene in pancreatic beta cells.6 Interestingly, the predictedto clarify the localization of Reg messenger RNA three-dimensional structure of the protein shows homol-(mRNA) and its product in the stomach and to examine ogy with that of the lectins, a family of growth-promot-changes in the level of their expression during regener-
ing molecules.7–9
ation of gastric mucosal cells. Methods: Gastric le-REG gene has also been found in the stomach.10 Al-sions were experimentally induced in Sprague–Dawley
though the precise localization and the role of the REGrats by water immersion stress. Northern blot analysisgene in the stomach remains undetermined, it is interest-and in situ hybridization studies were performed to ex-ing to speculate that it is involved in the process ofamine changes in mRNA levels. Immunohistochemical
studies were performed to identify the cellular localiza- gastric mucosal repair, as has been suggested to occur intion and to investigate the change in Reg protein level. the pancreas. In the pancreas, REG protein is primarilyResults: Reg mRNA and its product were distributed in found in the exocrine gland under normal conditions,11
the basal part of the oxyntic mucosa and were ex- whereas it is mainly present in the islets during regenera-pressed mainly in enterochromaffin-like cells. Levels of tion.5 The present study was therefore undertaken toboth Reg mRNA and its product were markedly in- clarify what type of cells in the stomach express Reg genecreased in the water immersion–induced gastric le- and its product and to investigate changes in the levelsions. Conclusions: Reg mRNA and its product are ex-
of their expressions in water immersion–induced gastricpressed in gastric enterochromaffin-like cells, and theirlesions.levels are increased during the healing process of water
immersion–induced gastric lesions. Materials and Methods
Animals
I Male Sprague–Dawley rats (250–300 g) were deprivedn 1984, Yonemura et al.1 found that administrationof poly(ADP-ribose) synthetase inhibitors, such as nic-
of food but allowed free access to water for 24 hours beforeotinamide, to rats subjected to 90% depancreatomy in- the experiments. Each study was performed with 5 animals induced regeneration of pancreatic islets. Subsequent each group. To induce gastric mucosal lesions, the animalsscreening of the regenerating rat islet-derived comple- were placed in strain cages and immersed in water (23�C) formentary DNA (cDNA) library showed a novel gene that 6 hours as described previously.12 Water-immersed rats wereencodes a 165–amino acid protein with a 21–amino killed 0, 6, 12, 24, 48, and 72 hours after the beginning ofacid signal peptide.2 The gene has been termed Reg andis abundantly expressed in regenerating islets but not in Abbreviations used in this paper: ECL cell, enterochromaffin-like
cell; GSH, glutathione-S-transferase; HDC, histidine decarboxylase;normal ones.2 Further studies showed that enhanced RegISH, in situ hybridization; RT-PCR, reverse-transcription polymerasegene expression is linked to DNA synthesis in pancreaticchain reaction.
islets both in vitro3 and in vivo.4 Moreover, it has been � 1996 by the American Gastroenterological Association0016-5085/96/$3.00shown recently that Reg protein is present in the secre-

46 ASAHARA ET AL. GASTROENTEROLOGY Vol. 111, No. 1
the stress period for Northern blot analysis and 0 and 24 hours oligonucleotide A (5�-GTGGATCCATGATGGAGCCCT-GTGAATAC-3�) and oligonucleotide B (5�-GTGGATCCT-after the beginning of the stress period for in situ hybridizationCAAATCAAGCCAGCCTTCT-3�), which contain BamHI(ISH) and immunohistochemical studies. All experiments weresites at their 5� ends. Amplified fragments were digested withreviewed and approved by the Kobe University Ethical Com-BamHI and then subcloned into the BamHI site of pGEX-2T.mittee for Animal Experiments.The resulting fusion protein combined the 210 amino acids
Tissue Preparation of the N-terminal region of mouse HDC with the C terminusof GST.The rats were anesthetized with sodium pentobarbital
Overnight cultures of Escherichia coli strain HB101, trans-(Nembutal, 55 mg/kg; Abott Laboratories, North Chicago, IL)formed with the expression vector, were diluted 1:10, and theand perfused transcardinally with ice-cold Zamboni’s solutioncells were grown for 1.5 hours at 37�C before adding isopropyl-(4% paraformaldehyde and 0.2% picrinic acid in 0.1 mol/L1-thio-b-D-galactoside (0.1 mmol/L). After an additional 3-phosphate buffer, pH 7.4; 2 mL/g body wt). The stomachs werehour growth period, bacteria were plated, resuspended, andexcised, immersed in fixative for 2–3 hours, and incubatedsonicated in PBS containing 1% sarcosyl and 0.2 mmol/Lin phosphate-buffered saline (PBS) containing 30% (wt/vol)phenylmethylsulfonyl fluoride and then centrifuged at 10,000gsucrose at 4�C until they sank. The glandular stomachs werefor 5 minutes at 4�C. The fusion protein in the lysate was thendivided into fundus (body) and antrum. They were thenpurified by glutathione-coupled Sepharose 4B (Pharmacia) af-quickly frozen in a dry ice plus 2-methylbutane mixture, andfinity chromatography in 50 mmol/L Tris-HCl (pH 8.0) con-5-mm sections were cut on a cryostat and thaw-mounted on 3-taining 20 mmol/L reduced glutathione.aminopropyl-triethoxysilane–coated slides. The sections were
To raise antisera, male Japanese white rabbits were immu-rinsed with 0.1 mol/L PBS and then treated with 10 mg/mLnized by intradermal injections of 200 mg purified GST-HDCproteinase K (Boehringer Mannheim, Mannheim, Germany)fusion protein mixed with complete Freund’s adjuvant (1:1)in 50 mmol/L Tris-HCl and 5 mmol/L ethylenediaminetetra-at 2-week intervals. After the fourth booster, the rabbits wereacetic acid (EDTA) (pH 8.0) for 10 minutes. After treatmentbled and antiserum was obtained. To purify anti–GST-HDCwith 4% paraformaldehyde, they were rinsed with PBS, acet-antibody, the antiserum was precipitated with ammonium sul-ylated with 0.25% (vol/vol) acetic anhydride in 0.1 mol/Lfate (0%–50%) and then adsorbed to Sepharose 4B gel coupledtriethanolamine, rinsed with PBS, dehydrated in an ethanolwith purified GST-HDC fusion protein. The antibody wasseries, and air dried.eluted with glycine-HCl (pH 2.5) and desalted using a PD-10 column (Pharmacia) preequilibrated with PBS.Preparation of Reg RNA Probes
The specificity of the anti-HDC antibody was checked byThe rat Reg cDNA used for ISH study and Northern
Western blotting and immunohistological study. An absorp-blot analysis was obtained by reverse-transcription polymerase
tion test was also performed, in which absorption of the antise-chain reaction (RT-PCR) amplification of the total RNA ex-
rum by excess GST-HDC fusion protein completely eliminatedtracted from rat pancreas2 using the synthetic oligonucleotide
the immunostaining.primers 5�-TGCCAGAACATGAATTC-3� and 5�-TTGAAC-
The antiserum against somatostatin was obtained from Ja-TTGCAGACAAA-3� under the following conditions: 94�C
pan Immunoresearch Laboratory Co. Ltd. (Tokyo, Japan). Thefor 30 seconds, 60�C for 60 seconds, and 72�C for 60 seconds
characterization of this antiserum has been described else-for 35 cycles. The PCR product was subcloned into the pCR-
where.13 In the present study, absorption of this antiserumII vector using a TA cloning kit (Invitrogen, San Diego, CA).
with synthetic somatostatin completely eliminated the immu-The recombinant plasmid was linearized by cutting at a single
nostaining in the stomach.site with a restriction enzyme (Xho I for the antisense probe For the generation of anti–chromogranin A antiserum, aand with BamHI for the sense probe). In vitro transcription 14–amino acid residue peptide corresponding to position 57–was performed using the appropriate RNA polymerase (SP6 70 of the rat chromogranin A deduced from the cDNA se-RNA polymerase for the antisense probe, T7 RNA polymerase quence14 was synthesized by a peptide synthesizer (430A; Per-for the sense probe) and [a-35S]uridine triphosphate (Amer- kin–Elmer, Foster City, CA). This peptide was conjugatedsham, Little Chalfont, Buckinghamshire, England). The 35S- with keyhole limpet hemocyanin (Sigma Chemical Co., St.labeled antisense riboprobe was complementary to part of the Louis, MO), and a rabbit anti–chromogranin A polyclonalcloned Reg gene (nucleotide positions 187–492).2 antiserum was raised against it. The characteristics of the anti-
serum were examined by immunoblotting and immunocyto-Preparation of Anti–histidinechemistry: the anti–chromogranin A antiserum properly rec-decarboxylase, Anti-somatostatin, andognized an 85-kilodalton band of native chromogranin AAnti–chromogranin A Antibodiesprotein in the blot of rat adrenal gland extract and stained
A glutathione-S-transferase (GST)–histidine decarbox- the secretory granules of adrenal chromaffin cells (unpublishedylase (HDC) fusion protein was produced by inserting a 630– data). In addition, the immunostaining in the stomach in thebase pair fragment of mouse HDC cDNA (nucleotides 1–630) present study was completely eliminated when the antiseruminto the expression vector pGEX-2T (Pharmacia LKB Biotech, was absorbed with the synthetic peptide described above before
use.Piscataway, NJ). This fragment was prepared by PCR with

Reg EXPRESSION IN GASTRIC ECL CELLS 47July 1996
ity and Reg messenger RNA (mRNA) was quantified by count-ISH Histochemistrying two randomly selected visual fields for each region in
Two protocols were used for the ISH studies. Onetwo sections from 5 animals under a light microscope (201
provided a single label using ISH histochemistry, and the otherobjective, magnification 2001). In the present study, we de-
protocol combined ISH and immunohistochemistry.fined erosions deeper than half of the mucosal height as the
The procedure used for ISH and detection have been de-erosive lesions and defined an area located more than 1 cm
scribed previously in detail.15 In brief, 200 mL of hybridizationdistant from the erosion as the nonerosive region. For each
mixture containing 50% formamide, 10% dextran sulfate, 0.3rat, the total number of Reg mRNA–positive cells, HDC- or
mol/L NaCl, 20 mmol/L Tris-HCl, 5 mmol/L EDTA, 0.1 mol/somatostatin-immunoreactive cells, and double-labeled cells
L NaPO4, 0.2% sarcosyl, 200 mg/mL salmon sperm DNA,were recorded. Using the ISH plus immunohistochemistry
11 Denhardt’s solution, 500 mg/mL yeast transfer RNA, andmethod, cells with grain densities at least five times higher
100 mmol/L dithiothreitol and 6.01 105 cpm of 35S-labeledthan the background densities were considered to be positively
probes were denatured for 2 minutes at 80�C, applied to eachlabeled for Reg mRNA, and a distinctive brown chromogen in
section, and covered with a siliconized coverslip. Hybridizationthe cell cytoplasm indicated HDC or somatostatin immunore-
was performed in a sealed humid chamber for 18 hours at 55�C.activity. Data are expressed as means { SE. A nonpaired Stu-
After hybridization, the slides were immersed in a mixture ofdent’s t test was used for comparison of the data. A P value
51 SSC (11 SSC is 150 mmol/L NaCl and 15 mmol/L sodiumof õ0.05 was considered significantly different.
citrate) and 1% mercaptoethanol at room temperature, and thecoverslips were allowed to fall off. The sections were incubated Northern Blot Analysisin 50% formamide, 21 SSC, and 10% mercaptoethanol at
Total RNAs were obtained from the excised glandular65�C for 30 minutes and equilibrated with ribonuclease bufferstomachs by extraction with guanidine thiocyanate, followed(10 mmol/L Tris-HCl, 1 mmol/L EDTA, and 0.5 mmol/Lby cesium chloride centrifugation as described previously.18
NaCl) three times for 10 minutes each at 37�C. They wereThe total RNAs were separated using electrophoresis on 0.66then treated with ribonuclease A (1 mg/mL) in the same buffermol/L formaldehyde plus 1% agarose gel with 0.4 mol/L 3-for 30 minutes; incubated in 50% formamide, 21 SSC, and(N-monopolyno)-propane sulfate, 0.1 mol/L sodium acetate,10% mercaptoethanol at 65�C for 30 minutes; and rinsed inand 0.02 mol/L EDTA. The nucleic acid was transferred to21 SSC and in 0.21 SSC at room temperature for 10 minutesnitrocellulose membranes (Schleicher & Schuell, Dassel, Ger-each. After washing, the sections were dehydrated in a gradedmany) and was fixed to the membrane using UV cross-linking.alcohol series and air dried.The rat Reg cDNA probe used for the Northern blot analysisFor autoradiography, the tissue sections were coated withwas obtained by RT-PCR amplification of the total RNAKodak NTB3 emulsion (diluted 1:1 with distilled water atextracted from rat pancreas (see section Preparation of Reg42�C; Eastman Kodak, Rochester, NY) and exposed for 3RNA Probes), and the HDC cDNA probe was also obtainedweeks in light-tight boxes at 4�C. After development in Kodakby RT-PCR of the total RNA from the stomach using oligonu-D-19 and fixing in Fujifix (Fuji Film, Tokyo, Japan), the sec-cleotide primers 5�-CTGATGCCATCAACTGCTTG-3� andtions were rinsed in distilled water, counterstained with5�-CCCTAAGGTTGCACAGACAA-3�.19 The plasmids wereH&E, rinsed in tap water, dehydrated in a graded alcoholdigested by EcoRI, and 32P-radiolabeled DNA probes wereseries, cleared in xylene, and coverslipped.synthesized using a random primer labeling kit (BoehringerIn the second protocol, tissue sections were first stainedMannheim). Hybridization was performed at 42�C for 16immunohistochemically using the avidin-biotin complexhours, and the filters were washed twice for 20 minutes each atmethod following a hybridization procedure described pre-55�C in 0.11 SSC/0.1% sodium dodecyl sulfate, as describedviously.16,17 After pretreatment with 1% normal goat serumpreviously.18 The radiolabeled DNA probes were detected byin PBS, the tissue sections were incubated with rabbit anti-exposure to Kodak XAR-5 film at 080�C for 24 hours. Themouse HDC antibody (1:600) or rabbit anti-somatostatin anti-signal intensity of the gene expression was quantified usingbody (1:600) overnight at 4�C in a humidified environment.the bioimaging analyzer BAS2000 (Fujix, Tokyo, Japan).20
Endogenous peroxidase activity was blocked with 1.5% H2O2
in 99% methanol for 30 minutes. The slides were washed,Immunohistochemistry for Reg Proteinand biotinylated goat anti-rabbit immunoglobulin (Gibco–
Bethesda Research Laboratories, Gaithersburg, MD; 1:400) A recombinant DNA corresponding to amino acid po-sitions 22–165 of the deduced rat Reg protein2 was synthe-was applied overnight at 4�C, followed by washing with PBS.
Bound antibody was detected using the avidin-biotin peroxi- sized in Saccharomyces cerevisae strain AH22, and a mouse anti–Reg protein monoclonal antibody was raised against it. Thedase method (ABC Elite Kit; Vector Laboratories, Burlingame,
CA). Peroxidase activity was subsequently revealed by incubat- antibody was found to be monospecific for Reg protein byWestern blot analysis.5ing with 3,3�-diaminobenzidine (Sigma Chemical Co.) in 0.05
mol/L Tris-HCl for 20 minutes at room temperature. After Three micrometers of paraffin-embedded sections was pre-pared from each animal. The tissue sections were stained im-dehydration in a graded alcohol series, the sections were cov-
ered with emulsion for autoradiography as described above. munohistochemically using the avidin-biotin complexmethod. After pretreatment with 1% normal goat serum inThe colocalization of HDC or somatostatin immunoreactiv-

48 ASAHARA ET AL. GASTROENTEROLOGY Vol. 111, No. 1
PBS, the tissue sections were incubated with mouse anti-rat cells positive for HDC immunoreactivity had a brownReg monoclonal antibody (1:500) for overnight at 4�C in a reaction product in the cytoplasm and were present inhumidified environment. Endogenous peroxidase activity was the basal part of the oxyntic gland both in normal andblocked with 1.5% H2O2 in 99% methanol for 30 minutes. water-immersed rat stomachs. Under lower-power mag-The slides were washed, and biotinylated goat anti-mouse im- nification, the cells positive for HDC immunoreactivitymunoglobulin (ABC Elite Kit, diluted 1:200; Vector Labora- in the basal part of the oxyntic gland were clearly visibletories) was applied for 2 hours at room temperature, followed
in the bright-field image (Figure 3A ) and the Regby washing with PBS. Bound antibody was detected usingmRNA–positive cells showed a similar distribution inthe avidin-biotin peroxidase method (ABC Elite Kit; Vectordark-field images (Figure 3B ), suggesting colocalizationLaboratories). Peroxidase activity was subsequently shown byof HDC immunoreactivity and Reg mRNA. Under high-incubating with 3,3�-diaminobenzidine (Sigma Chemical Co.)power magnification, most of the cells expressing Regin 0.05 mol/L Tris-HCl for 10 minutes at room temperature,
followed by hematoxylin staining. After dehydration in a mRNA were positive for HDC immunoreactivity (Figuregraded alcohol series, the sections were cleared in xylene. 3C ); 42.6% { 3.2% of HDC-immunoreactive cells
expressed the Reg gene and 68.9% { 4.8% of RegImmunohistochemical Double StainingmRNA–positive cells showed HDC immunoreactivity
Immunohistochemical double staining for Reg protein in the normal stomach (Table 1). In contrast, somato-and chromogranin A in the stomachs was performed 24 hours statin-immunoreactive cells were completely devoid ofafter the beginning of the stress. The sections were incubated silver grain accumulations, indicating an absence of Regwith mouse anti-Reg monoclonal antibody (1:500) and rabbit gene expression in D cells (Figure 4).anti–chromogranin A polyclonal antibody (1:500). Then, flu-orescein isothiocyanate–conjugated anti-mouse immunoglob- Changes of Reg Gene Expression inulin (Silenus Laboratories, Hawthorn, Australia; dilution, Water-Immersed Rat Stomachs1:200) and biotinylated sheep anti-rabbit immunoglobulin(ABC Elite Kit, Vector Laboratories) were applied for 2 hours Northern blot analysis showed that Reg gene ex-at room temperature, followed by incubation with strepto- pression was relatively low in the normal stomach. How-avidin–Texas Red (Amersham) for 2 hours. After washing with ever, Reg mRNA signals in the stomachs with waterPBS, the sections were observed by confocal laser microscopy immersion–induced gastric lesions gradually increased(Olympus, Tokyo, Japan). up to 24 hours and returned to almost normal levels at
72 hours (Figure 5A ). When the signal intensities wereResultsquantified by BAS 2000, a bioimaging analyzer, Reg gene
Identification of Cells Expressing Reg expression increased to approximately three times controlGene 6 hours after the start of the stress period, reaching its
maximum at 24 hours, with a 10-fold increase in itsISH with a 35S-labeled Reg riboprobe detectedpositive cells in the oxyntic mucosa, but not in the pylo- signal intensity compared with normal levels. In
agreement with the Northern blot data, ISH with a 35S-ric gland mucosa, both in normal and water-immersedrat stomachs (data not shown). Under low-power magni- labeled Reg riboprobe showed that both the number of
Reg mRNA-positive cells and signal intensity in eachfication, most of the cells expressing Reg mRNA, identi-fied by silver grain accumulation, were localized in the cell were obviously increased throughout the oxyntic mu-
cosa of water-immersed rat stomachs and that the increasebasal part of the oxyntic gland in normal rats (Figure1A and B ). Under higher magnification, distinct hybrid- was more prominent in the erosive lesions (Figure 1C
and D ). It is noteworthy that in the water-immersed ratization signals of Reg mRNA could be detected in normalstomach neither in parietal cells, easily identified by their stomach, Reg gene expression was observed not only in
the lower segment but also in the upper segment of thelarge size and clear or acidophilic cytoplasm, nor in chiefcells, which were identified by the basal localization of oxyntic gland (Figure 1C and D ). However, as with
normal stomachs, neither parietal cells nor chief cellstheir nuclei and their basophilic cytoplasm. Instead, ac-cumulations of silver grains were found over relatively were positive for any hybridization signals, even in the
treated animals (Figure 2B ). In contrast to the Regsmall cells localized primarily in the basal part of theoxyntic gland (Figure 2A ). In contrast, no hybridization mRNA–positive cells, the number of HDC-immunore-
active cells was not changed by water immersion stresssignals were detectable above background for radiola-beled sense probe (data not shown). (Table 1). Consequently, the number of HDC-immuno-
reactive cells positive for Reg mRNA signal increasedTo clarify the type of cells expressing Reg mRNA, ISHcombined with immunohistochemistry was performed from 42.6% { 3.2% to 72.4% { 5.1% in the erosive
lesion by water immersion stress (Table 1).using anti-HDC or anti-somatostatin antibodies. The
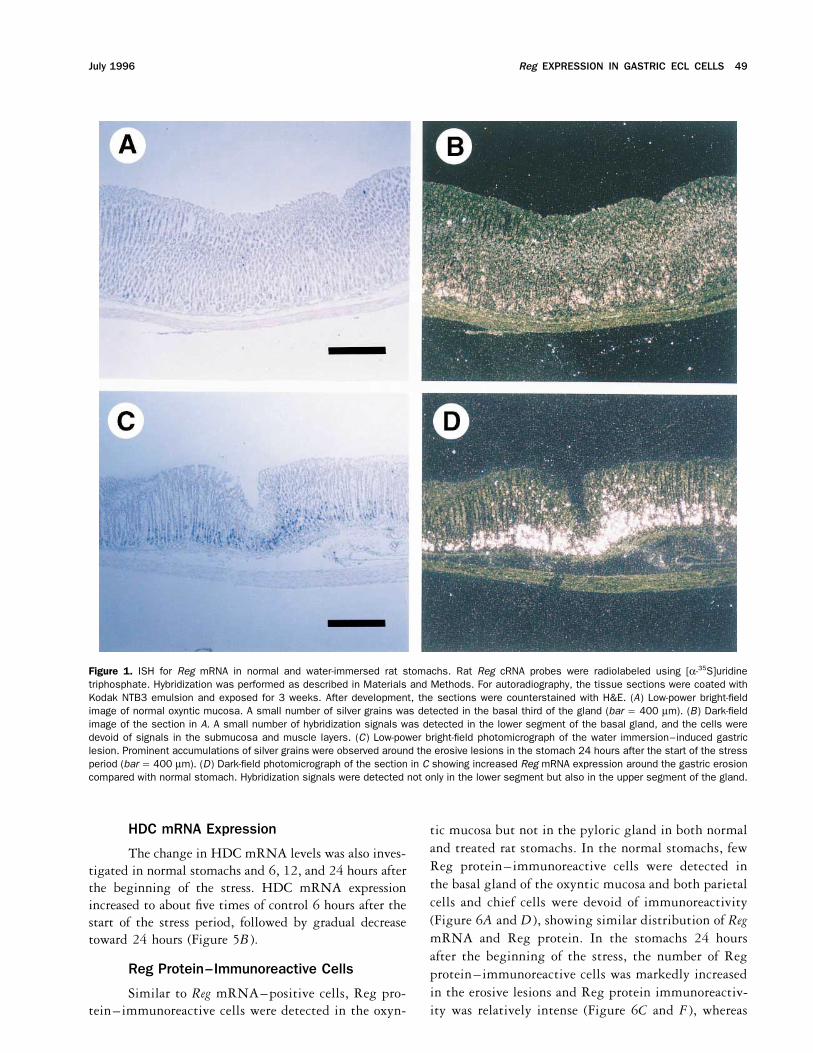
Reg EXPRESSION IN GASTRIC ECL CELLS 49July 1996
Figure 1. ISH for Reg mRNA in normal and water-immersed rat stomachs. Rat Reg cRNA probes were radiolabeled using [a-35S]uridinetriphosphate. Hybridization was performed as described in Materials and Methods. For autoradiography, the tissue sections were coated withKodak NTB3 emulsion and exposed for 3 weeks. After development, the sections were counterstained with H&E. (A) Low-power bright-fieldimage of normal oxyntic mucosa. A small number of silver grains was detected in the basal third of the gland (bar Å 400 mm). (B) Dark-fieldimage of the section in A. A small number of hybridization signals was detected in the lower segment of the basal gland, and the cells weredevoid of signals in the submucosa and muscle layers. (C) Low-power bright-field photomicrograph of the water immersion–induced gastriclesion. Prominent accumulations of silver grains were observed around the erosive lesions in the stomach 24 hours after the start of the stressperiod (bar Å 400 mm). (D) Dark-field photomicrograph of the section in C showing increased Reg mRNA expression around the gastric erosioncompared with normal stomach. Hybridization signals were detected not only in the lower segment but also in the upper segment of the gland.
HDC mRNA Expression tic mucosa but not in the pyloric gland in both normaland treated rat stomachs. In the normal stomachs, fewThe change in HDC mRNA levels was also inves-Reg protein–immunoreactive cells were detected intigated in normal stomachs and 6, 12, and 24 hours afterthe basal gland of the oxyntic mucosa and both parietalthe beginning of the stress. HDC mRNA expressioncells and chief cells were devoid of immunoreactivityincreased to about five times of control 6 hours after the(Figure 6A and D ), showing similar distribution of Regstart of the stress period, followed by gradual decreasemRNA and Reg protein. In the stomachs 24 hourstoward 24 hours (Figure 5B ).after the beginning of the stress, the number of Reg
Reg Protein–Immunoreactive Cells protein–immunoreactive cells was markedly increasedin the erosive lesions and Reg protein immunoreactiv-Similar to Reg mRNA–positive cells, Reg pro-ity was relatively intense (Figure 6C and F ), whereastein–immunoreactive cells were detected in the oxyn-

50 ASAHARA ET AL. GASTROENTEROLOGY Vol. 111, No. 1
Figure 2. ISH for Reg mRNA in normal and water-immersed rat stomachs. Higher-magnification photomicrographs corresponding to Figure 1Aand C. (A) Normal oxyntic mucosa. Distinct hybridization signals cannot be observed in either parietal cells (easily identified by their large sizeand clear or acidophilic cytoplasm) or chief cells (identified by basal localization of their nuclei and their basophilic cytoplasm). Accumulationsof silver grains were found over the relatively small cells localized primarily in the basal part of the oxyntic gland (bar Å 20 mm). (B) Waterimmersion–induced gastric lesions. Although both the number of positive cells expressing Reg mRNA and the signal intensity of Reg mRNA ina single cell were increased, parietal and chief cells were devoid of any hybridization signals (bar Å 20 mm).
only a slight increase was observed in nonerosive re- cells were positive for chromogranin A immunoreactiv-ity. We subsequently performed a combined ISH andgions (Figure 6B and E ).
Immunohistochemical double staining was also per- immunohistochemistry study to identify what type ofendocrine cells express the Reg gene using anti-HDCformed to characterize Reg protein–immunoreactive
cells more precisely. As shown in Figure 7, all Reg pro- and anti-somatostatin antibodies. Interestingly, we foundthat approximately 70% of the cells expressing Reg genetein–immunoreactive cells showed chromogranin A im-
munoreactivity, confirming that Reg protein is present were positive for immunoreactive HDC but that no im-munoreactive somatostatin cells had hybridization sig-exclusively in endocrine cells.nals. Because we and other investigators have previously
Discussion shown that HDC is exclusively present in enterochromaf-fin-like (ECL) cells on the basis of immunohistochemicalReg mRNA has been shown to be present in thestudy and electronmicroscopic examination,21–23 ourgastric wall,10 but its precise localization in the stomachpresent data indicate that Reg gene is primarily expressedhas not been elucidated. The present study, using ISHin ECL cells. However, approximately 30% of Regand immunohistochemistry, showed that both RegmRNA–positive cells did not show either HDC or so-mRNA and Reg protein are expressed in the basal partmatostatin immunoreactivity. It is possible that theseof the oxyntic mucosa but not in the pyloric mucosa.are also ECL cells, in which the HDC content is too smallMoreover, parietal and zymogenic cells were completelyto be detected by immunohistochemistry. However, itdevoid of any Reg mRNA hybridization signals or Regis well known that the oxyntic mucosa in the rat harborsprotein immunoreactivity, and most of the silver grainsat least three ultrastructurally distinct endocrine cells:in ISH were detected over small cells in the basal partECL cells, which constitute Ç65% of all endocrine cells;of the gland, suggesting that fundic endocrine cells ex-A-like (or X) cells, which constitute Ç25%; and somato-press Reg mRNA and its product. Confirming these data,
we also showed that all the Reg protein–immunoreactive statin D cells, which constitute Ç10%.24 Thus, it may be

Reg EXPRESSION IN GASTRIC ECL CELLS 51July 1996
considered that, in addition to ECL cells, other endocrinecells, such as A-like cells, express the Reg gene. In anycase, the present observation that Reg gene is present inendocrine cells in the gastric mucosa is consistent withthe fact that pancreatic endocrine cells express RegmRNA.2,5
The significant increase in both Reg gene expressionand Reg protein immunoreactivity in water immersionstress–induced gastric mucosal injury are importantfindings. In Northern blot analysis, Reg gene expressionincreased by 10-fold 24 hours after the start of the stress.Moreover, the ISH study showed that not only the num-ber of Reg mRNA–positive cells but also the signal in-tensity of Reg mRNA in each cell was increased. Simi-larly, HDC mRNA level was also increased by waterimmersion stress. However, in spite of the increasednumber of Reg mRNA–positive cells, the number ofthe cells immunoreactive for HDC was not altered. Thenumber of HDC-immunoreactive cells positive for RegmRNA signals increased from 42% to 72% by waterimmersion stress. Thus, it appears that water immersionstress induces enhancement of Reg and HDC mRNAexpression in ECL cells without affecting their total num-ber. These data suggest a possible specific role of Reggene expression in ECL cells in the process of gastricmucosal damage. Whether the enhanced Reg gene expres-sion, with resulting increase of Reg protein, is directlyrelated to water immersion stress or nonspecifically togastric mucosal damage is an interesting question. Inthis study, we clearly show that the Reg gene was mostprominently expressed in the proximity of the erosivelesions. In addition, we found that the changes of Regprotein immunoreactivity paralleled those of Reg geneexpression. Indeed, in good agreement with the ISHstudy, the immunohistochemical study showed that thenumber of Reg protein–immunoreactive cells is in-
�Figure 3. ISH immunohistochemical (HDC) study of the stomach 24hours after the stress period. After the hybridization procedure, thesections were immunohistochemically stained for HDC with polyclonalantibody and the standard avidin-biotin complex method. (A) Bright-field photomicrograph of the water-immersed rat stomach showed thatHDC-immunoreactive cells were distributed in the lower segment ofthe oxyntic gland, similar to cells expressing Reg mRNA, as shown inB (bar Å 400 mm). (B) Dark-field photomicrograph of the section inA. The distribution of cells expressing Reg mRNA is similar to that ofHDC-immunoreactive cells. (C) Under high-power field, the double-labeled cells (arrowheads) showed a brown reaction product in thecytoplasm, indicating HDC immunoreactivity, and an accumulation ofsilver grains overlying the cells, indicating the presence of Reg mRNA.Approximately 70% of HDC-immunoreactive cells became positive forReg mRNA 24 hours after the start of water-immersion stress,whereas 40% of the cells were positive for Reg gene expression innormal stomach (bar Å 15 mm).
52 ASAHARA ET AL. GASTROENTEROLOGY Vol. 111, No. 1
Table 1. Changes in Number of HDC-Immunoreactive Cells and Reg mRNA–Positive Cells by Water Immersion Stress
HDC-positive cells Reg mRNA–positive cells
Total Reg-positive (%) Total HDC-positive (%)
Normal rats 48.4 { 4.3 42.6 { 3.2 29.4 { 2.8 68.9 { 4.8Water-immersed rats
Nonerosive region 51.3 { 4.1 52.9 { 4.7 38.7 { 3.4a 71.2 { 4.4Erosive lesion 51.9 { 4.2 72.4 { 5.1b,c 54.7 { 4.1b,d 68.1 { 4.6
NOTE. Results are expressed as mean { SE.aP õ 0.05, bP õ 0.01 significantly different from respective values in normal rats.cP õ 0.05, dP õ 0.01 significantly different from nonerosive region of the water-immersed rats.
creased in the erosive lesions. Furthermore, Reg gene andits product have been shown to be overexpressed in thegastric mucosa of indomethacin-treated rats and in anacetic acid–induced gastric ulcer model (unpublisheddata). Therefore, the enhancement of Reg gene expressionmay be a generalized phenomenon with damaged gastricmucosa.
Recently, Francis et al.3 showed that increased isletcell replication is paralleled by enhanced Reg gene expres-sion in isolated pancreatic islets of rats. Miyaura et al.4
showed that an increase in Reg gene expression was associ-ated with an increase of beta cell volume in animal mod-els. Watanabe et al.6 reported that recombinant rat Regprotein not only stimulates [3H]thymidine incorporationinto isolated pancreatic islets of rats in vitro but alsoameliorates surgically induced diabetes in vivo. On theother hand, studies predicting the three-dimensional
Figure 5. Northern blot analysis in the normal and water-immersedrat stomachs. Total RNAs were hybridized with the 32P-labeled cDNAprobes. (A) Northern blotting for Reg mRNA. The signals for Reg mRNAgradually increased up to 24 hours after the start of water immersion(top). The nitrocellulose membranes were rehybridized with the probefor b-actin mRNA (bottom). Lane 1, normal stomach; lanes 2–6, stom-Figure 4. ISH immunohistochemical (somatostatin) study of the
stomach 24 hours after the beginning of the stress. After the hybridiza- ach 6, 12, 24, 48, and 72 hours after the start of water immersionstress, respectively. (B) Northern blotting for HDC mRNA. The signalstion procedure, the sections were immunohistochemically stained for
somatostatin and were subjected to the standard avidin-biotin com- were most increased 6 hours after the beginning of the stress andgradually decreased toward 24 hours. Lane 1, normal stomach; lanesplex method. Under high-power field, somatostatin-immunoreactive
cells (arrowheads) did not show any hybridization signal (bar Å 30 2–4, stomach 6, 12, and 24 hours after the start of water immersionstress, respectively.mm).

Reg EXPRESSION IN GASTRIC ECL CELLS 53July 1996
Figure 6. Immunohistochemical staining for Reg protein in (A and D) normal stomachs, (C and F) erosive lesions, and (B and E) areas 1 cmdistant from the erosions 24 hours after the beginning of the stress. (A) Reg protein immunoreactivity in the normal oxyntic mucosa. A smallnumber of cells are positive for Reg protein, and most of the cells are distributed in the bottom of the fundic gland (bar Å 400 mm). (B) Regprotein immunoreactivity in the area 1 cm distant from the erosion. Number of Reg protein–immunoreactive cells is slightly increased comparedwith the mucosa of the normal rats (bar Å 400 mm). (C) Reg protein immunoreactivity in the erosive lesion. Number of Reg protein–immunoreac-tive cells is significantly increased (bar Å 400 mm). (D) High-power view of A (bar Å 30 mm). (E) High-power view of B (bar Å 30 mm). (F) High-power view of C. Arrowheads indicate the margin of the erosion (bar Å 30 mm).
structure of Reg protein suggest homology with lectins, tric mucosal injury.25–27 In the present study, the expres-sion of Reg gene and its product were markedly increaseda family of proteins with diverse roles involved in growth
promotion.7,8 Based on these data, the Reg gene is consid- in the erosive lesions. The direct effect of Reg proteinon the gastric mucosal cell growth should therefore beered to play a role in the regeneration of pancreatic islets.
It is also interesting to speculate that increased Reg gene examined in future studies.On the other hand, it has been proven that the deducedexpression participates in the healing of gastric mucosal
injury through a direct effect of Reg protein on the amino acid sequence of human REG protein is identicalto that of pancreatic stone protein2,28,29 and that REGgastric mucosal cells. Several investigators have shown,
using [3H]thymidine or bromodeoxyuridine labeling, protein and pancreatic stone protein are derived from thesame gene.10,30 Pancreatic stone protein was originallythat the number of proliferative cells increased around
gastric erosions and ulcers in the healing process of gas- discovered as a protein that comprises up to 10% of the

54 ASAHARA ET AL. GASTROENTEROLOGY Vol. 111, No. 1
increased expression of Reg gene and its product in ECLcells suggests that ECL cells play important roles in thehealing process of gastric mucosal injury.
References1. Yonemura Y, Takashima T, Miwa K, Miyazaki I, Yamamoto H,
Okamoto H. Amelioration of diabetes mellitus in partially depan-creatised rats by poly(ADP-ribose) synthetase inhibitors. Diabe-tes 1984;33:401–404.
2. Terazono K, Yamamoto H, Takasawa S, Shiga K, Yonemura Y,Tochino Y, Okamoto H. A novel gene activated in regeneratingislets. J Biol Chem 1988;263:2111–2114.
3. Francis PJ, Southgate JL, Wilkin TJ, Bone AJ. Expression of anislet regenerating (reg) gene in isolated rat islets: effects of nutri-ent and non-nutrient growth factors. Diabetologia 1992;35:238–242.
4. Miyaura C, Ling C, Appel M. Expression of reg gene/PSP, a pan-creatic exocrine gene: relationship to changes in islet beta-cellFigure 7. Immunohistochemical double staining for Reg protein andmass. Mol Endocrinol 1991;5:226–234.chromogranin A in the erosive lesions of the stomach 24 hours after
5. Terazono K, Uchiyama Y, Ide M, Watanabe T, Yonekura H, Yama-the stress. The sections were incubated with anti-Reg monoclonalmoto H, Okamoto H. Expression of reg protein in rat regeneratingantibody and anti–chromogranin A polyclonal antibody. Fluoresceinislets and its co-localization with insulin in the beta cell secretoryisothiocyanate–conjugated anti-mouse immunoglobulin and biotinyl-granules. Diabetologia 1990;33:250–252.ated anti-rabbit immunoglobulin were applied, followed by incubation
6. Watanabe T, Yonemura Y, Yonekura H, Suzuki Y, Miyashita H,with streptavidin–Texas Red. The sections were observed using confo-Sugiyama K, Moriizumi S, Unno M, Tanaka O, Kondo H, Bonecal laser microscopy. Reg protein immunoreactivity was visualized byAJ, Takasawa S, Okamoto H. Pancreatic beta-cell replication andfluorescence (fluorescein isothiocyanate) (left), and chromogranin Aamelioration of surgical diabetes by reg protein. Proc Natl Acadimmunoreactivity was visualized by fluorescence (Texas Red) (right).Sci USA 1994;91:3589–3592.In the middle, yellow staining represents colocalization of Reg protein
7. Patty L. Homology of pancreatic stone protein with animal lectin.and chromogranin A immunoreactivity (bar Å 10 mm).Biochem J 1988;253:309–311.
8. Peterson TE. The amino terminal domain of thrombomodulin andPSP are homologous with lectin. FEBS Lett 1988;231:51–53.protein in pancreatic juice, maintaining it in a supersatu-
9. Lassere C, Simon MT, Ichikawa H, Dirong S, Nguyen VC, Christarated state with respect to calcium carbonate.31 It wasL, Vernier P, Brechot C. Structural organization and chromosomal
therefore speculated that pancreatic stone protein has a localization of a human gene (HIP/PAP) encoding a C-type lectinoverexpressed in primary liver cancer. Eur J Biochem 1994;224:role in stabilizing pancreatic juice and preventing pancre-29–38.atic stone formation. Indeed, in addition to the preferen-
10. Watanabe T, Yonekura H, Terazono K, Yamamoto H, Okamototial expression of Reg gene in pancreatic islets during H. Complete nucleotide sequence of human reg gene and its
expression in normal and tumoral tissues: the reg protein, pan-regeneration of the pancreas, some investigators have re-creatic stone protein, and pancreatic thread protein are one andported that both REG mRNA and its product were local-the same product of the gene. J Biol Chem 1990;265:7432–ized in the acinar cells of the human adult pancreas,11,32,337439.
suggesting exocrine secretion of REG protein. Interest- 11. Kimura N, Yonekura H, Okamoto H, Nagura H. Expression ofhuman regenerating gene mRNA and its product in normal andingly, the REG gene contains an amino acid sequenceneoplastic human pancreas. Cancer 1992;70:1857–1863.corresponding to that of signal peptide, suggesting a
12. Takeuchi K, Okabe S. Importance of pepsin and stomach disten-characteristic feature of a secretory protein.2 Moreover, sion in morphological alteration of stress-induced gastric lesion
in pylorus-ligated rats. Dig Dis Sci 1988;33:52–59.REG protein possesses three disulfide bonds in its mole-13. Tohyama M, Siosaka S, Takagi H, Inagaki S, Takatsuki K, Saka-cule, indicating that, similar to trefoil peptides, it may
naka M, Senba E, Kawai Y, Minagawa H. Somatostatin-like immu-be resistant to enzymatic digestion.34 Thus, although noreactivity in the facial, glossopharyngeal and vagal lobes of
the carp. Neurosci Lett 1981;24:233–236.neither signal for Reg mRNA nor Reg protein immunore-14. Iacangelo A, Okayama H, Eiden ED. Primary structure of rat chro-activity in gastric exocrine cells, it is an interesting possi-
mogranin A and distribution of its mRNA. FEBS Lett 1988;227:bility that Reg protein is secreted into the gastric lumen, 115–121.influencing the stability of the mucous layers. 15. Yoshimuara R, Kiyama H, Kimura T, Araki T, Maeno H, Tnaizawa
O, Tohyama M. Localization of oxytocin receptor messenger ribo-In conclusion, we show that the expressions of Regnucleic acid in the rat brain. Endocrinology 1993;133:1239–gene and its product in rat ECL cells are increased in1246.
water immersion–induced gastric lesions. It has recently 16. Noguchi K, Kowalski K, Solodkin A, Iadarola MJ, Ruda MA. Dynor-phin expression and Fos-like immunoreactivity following inflam-been reported that nitric oxide synthesis is increased inmation induced hyperalgesia are colocalized in spinal cord neu-gastric ECL cells during the healing process of aceticrons. Mol Brain Res 1991;10:227–233.
acid–induced gastric ulcers.35 Although the number of 17. Noguchi K, Dubner R, Ruda A. Preprokephalin mRNA in spinaldorsal horn neurons is induced by peripheral inflammation andECL cells was not increased in our present study, the

Reg EXPRESSION IN GASTRIC ECL CELLS 55July 1996
is co-localized with Fos and Fos-related proteins. Neuroscience 28. Roquier S, Verdier JM, Iovanna J, Dagorn JC, Giorgi D. Rat pancre-atic stone protein messenger RNA. Abundant expression in ma-1992;46:561–570.
18. Nakata H, Matsui T, Ito M, Taniguchi T, Naribayashi Y, Arima N, ture exocrine cells, regulation by food content and sequenceidentity with the endocrine Reg transcript. J Biol Chem 1991;Nakamura A, Kinoshita Y, Chihara K, Hosoda S, Chiba T. Cloning
and characterization of gastrin receptor from ECL carcinoid tumor 266:786–791.29. Giorge D, Bernard JP, Roquier S, Iovanna J, Sarles H, Dagorn JC.of Mastomys natalensis. Biochem Biophys Res Commun 1992;
187:1151–1157. Secretary pancreatic stone protein messenger RNA: nucleotidesequence and expression in chronic calcifying pancreatitis. J Clin19. Joseph DR, Sullivan P, Wang YM, Kozak CA, Fenstermacher DA,
Behrendsen ME, Zahnow CA. Characterization and expression of Invest 1989;84:100–106.30. Stewart TA. The human reg gene encodes pancreatic stone pro-the cDNA encoding rat histidine decarboxylase. Proc Natl Acad
Sci USA 1990;87:733–777. tein. Biochem J 1989;260:622–623.31. De Caro A, Multigner L, Lafont H, Lombardo D, Sarles H. The20. Asahara M, Kinoshita Y, Nakata H, Matsushima Y, Naribayashi
Y, Nakamura A, Matsuil T, Chihara K, Yamamoto J, Ichikawa A, molecular characteristics of a human pancreatic acidic phospho-protein that inhibits calcium carbonate crystal growth. BiochemChiba T. Gastrin receptor genes are expressed in gastric parietal
cells and enterochromaffin-like cells of Mastomys natalensis. Dig J 1984;222:669–677.32. De La Porte PP, De Caro A, Lafont H, Sarles H. Immunocytochemi-Dis Sci 1994;39:2149–2156.
21. Rubin W, Schwartz B. An electron microscopic radioautographic cal localization of pancreatic stone protein in the human digestivetract. Pancreas 1986;1:301–308.identification of the ECL cell as the histamine-synthesizing endo-
crine cell in the rat stomach. Gastroenterology 1979;77:458– 33. Hoorens A, Lemoine NR, McLellan E, Morohoshi T, Kamisawa T,Heitz PU, Stamm B, Ruschoff J, Wiedemann B, Kloppel G. Pancre-4567.
22. Soll AH, Lewin KJ, Beaven MA. Isolation of histamine-containing atic acinar cell carcinoma. An analysis of cell lineage marker,p53 expression, and Ki-ras mutation. Am J Pathol 1993;143:cells from rat gastric mucosa: biochemical and morphological
differences from mast cells. Gastroenterology 1981;80:717– 685–698.34. De A, Brown DG, Gorman MA, Carr M, Sanderson MR, Freemont727.
23. Kubota H, Taguchi Y, Tohyama M, Matsuura N, Shiosaka S, Ishi- PS. Crystal structure of a disulfide-linked ‘‘trefoil’’ motif found ina large family of putative growth factors. Proc Natl Acad Sci USAhara T, Watanabe T, Shiotani Y, Wada H. Electron microscopic
identification of histidine decarboxylase–containing endocrine 1994;91:1084–1088.35. Akiba Y, Nakamura M, Oda M, Kimura H, Miura S, Tsuchiya M,cells of the rat gastric mucosa: an immunohistochemical analy-
sis. Gastroenterology 1984;87:496–502. Ishii H. Nitric oxide synthesis in activated enterochromaffin-likecell in the healing process of gastric ulcer and hypergastrinemia24. Simonsson M, Eriksson S, Hakanson R, Lind T, Lonroth H, Lun-
dell L, O’Connor DT, Sundler F. Endocrine cells in the human (abstr). Digestion 1994;55:1–62.oxyntic mucosa. A histochemical study. Scand J Gastroenterol1988;3:1089–1099.
25. Yeomans ND, St John DJB, de Boer WGRM. Regeneration of Received August 7, 1995. Accepted March 29, 1996.Address requests for reprints to: Tsutomu Chiba, M.D., Ph.D., Divi-gastric mucosa after aspirin-induced injury in the rat. Am J Dig
Dis 1973;18:773–780. sion of Gerontology, Department of Internal Medicine, Kobe Univer-sity School of Medicine, Kusunoki-cho 7-5-2, Chuo-ku, Kobe 650,26. Ohning GV, Guth PH. Time course of mucosal cell proliferation
following acute aspirin injury in rat stomach. Dig Dis Sci 1995; Japan. Fax: (81) 78-361-7524.Supported by a grant-in-aid for scientific research from the Ministry40:1351–1356.
27. Helpap B, Hattori T, Gedigk P. Repair of gastric ulcer. Virchow of Education, Science and Culture and a grant from the Ministry ofHealth and Welfare, Japan.Arch A Pathol Anat Histopathol 1981;392:159–170.
Related Documents











