Reduction of CMP-N-acetylneuraminic acid hydroxylase activity in engineered Chinese hamster ovary cells using an antisense-RNA strategy Stephane Chenu a , Anne Gre ´goire a , Yanina Malykh b , Athanase Visvikis c , Lucia Monaco d,1 , Lee Shaw b , Roland Schauer b , Annie Marc a , Jean-Louis Goergen a, * a Laboratoire des Sciences du Ge ´nie Chimique, CNRS-ENSAIA, 2, av. de la Fore ˆt de Haye, F-54505 Vandoeuvre-le `s-Nancy, France b Biochemisches Institut, Christian-Albrechts-Universita ¨t zu Kiel, Olshausenstr. 40, D-24098 Kiel, Germany c Faculte ´ de Pharmacie, Univ. H. Poincare ´, 30 rue Lionnois, F-54000 Nancy, France d DIBIT – San Raffaele Scientific Institute, I-20132 Milan, Italy Received 2 April 2003; received in revised form 5 June 2003; accepted 18 June 2003 Abstract Rodent cells, widely used for the industrial production of recombinant human glycoproteins, possess CMP-N-acetylneuraminic acid hydroxylase (CMP-Neu5Ac hydroxylase; EC 1.14.13.45) which is the key enzyme in the formation of the sialic acid, N-glycolylneuraminic acid (Neu5Gc). This enzyme is not expressed in an active form in man and evidence suggests that the presence of Neu5Gc in recombinant therapeutic glycoproteins may elicit an immune response. The aim of this work was, therefore, to reduce CMP-Neu5Ac hydroxylase activity in a Chinese Hamster Ovary (CHO) cell line, and thus the Neu5Gc content of the resulting glycoconjugates, using a rational antisense RNA approach. For this purpose, the cDNA of the hamster hydroxylase was partially cloned and sequenced. Based on the sequence of the mouse and hamster cDNAs, optimal antisense RNA fragments were selected from preliminary in vitro translation tests. Compared to the parental cell line, the new strain (CHO-AsUH2), which was transfected with a 199-bp antisense fragment derived from the mouse CMP-Neu5Ac hydroxylase cDNA, showed an 80% reduction in hydroxylase activity. An analysis of the sialic acids present in the cells’ own glycoconjugates revealed a decrease in the percentage of Neu5Gc residues from 4% in the parental cells to less than 1% in the CHO-AsUH2 cell line. D 2003 Elsevier B.V. All rights reserved. Keywords: CMP-Neu5Ac hydroxylase; Sialylation; N-glycolylneuraminic acid; CHO cell; Antisense RNA 1. Introduction Chinese Hamster Ovary (CHO) cells are widely used for the production of recombinant glycoproteins employed in human therapy. Although the posttranslational modifica- tions of recombinant glycoproteins generated in CHO cells are very similar to that present on human proteins, these rodent cells do not perfectly mimic the human sialylation pattern required for bioactivity, stability, and absence of antigenicity [1,2]. One of the most significant differences between rodent and human sialylation is the incorporation of N-glycolylneuraminic acid (Neu5Gc). This sialic acid is present in the glycoconjugates of virtually all vertebrates, with the notable exception of man and chicken. In animals, Neu5Gc is formed as the sugar-nucleotide cytidine mono- phosphate-N-glycolylneuraminic acid (CMP-Neu5Gc) by the action of CMP-Neu5Ac hydroxylase (EC 1.14.13.45) [3]. In man, the gene encoding this enzyme lacks an exon, resulting in an incomplete mRNA with a premature termi- nation signal [4,5]. Due to this mutation, Neu5Gc cannot be formed by CMP-Neu5Ac hydroxylase in man and is prac- tically undetectable in healthy human tissues [6], though small amounts of Neu5Gc are found in gangliosides and glycoproteins of certain human tumors [7]. Glycoconjugates containing this common sialic acid are therefore foreign to humans and can induce an immune response, causing the formation of so-called Hanganutziu– Deicher antibodies [8,9]. The appearance of these ‘‘serum sickness’’ antibodies was observed after inoculation of patients with animal 0304-4165/03/$ - see front matter D 2003 Elsevier B.V. All rights reserved. doi:10.1016/S0304-4165(03)00137-5 * Corresponding author. Tel.: +33-383-59-5844; fax: +33-383-59- 5804. E-mail address: [email protected] (J.-L. Goergen). 1 Present address: Keryos SpA, via Maritano 26, I-20097 San Donato Milanese, Italy. www.bba-direct.com Biochimica et Biophysica Acta 1622 (2003) 133 – 144

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.bba-direct.com
Biochimica et Biophysica Acta 1622 (2003) 133–144
Reduction of CMP-N-acetylneuraminic acid hydroxylase activity in
engineered Chinese hamster ovary cells using an antisense-RNA strategy
Stephane Chenua, Anne Gregoirea, Yanina Malykhb, Athanase Visvikisc, Lucia Monacod,1,Lee Shawb, Roland Schauerb, Annie Marca, Jean-Louis Goergena,*
aLaboratoire des Sciences du Genie Chimique, CNRS-ENSAIA, 2, av. de la Foret de Haye, F-54505 Vandoeuvre-les-Nancy, FrancebBiochemisches Institut, Christian-Albrechts-Universitat zu Kiel, Olshausenstr. 40, D-24098 Kiel, Germany
cFaculte de Pharmacie, Univ. H. Poincare, 30 rue Lionnois, F-54000 Nancy, FrancedDIBIT–San Raffaele Scientific Institute, I-20132 Milan, Italy
Received 2 April 2003; received in revised form 5 June 2003; accepted 18 June 2003
Abstract
Rodent cells, widely used for the industrial production of recombinant human glycoproteins, possess CMP-N-acetylneuraminic acid
hydroxylase (CMP-Neu5Ac hydroxylase; EC 1.14.13.45) which is the key enzyme in the formation of the sialic acid, N-glycolylneuraminic
acid (Neu5Gc). This enzyme is not expressed in an active form in man and evidence suggests that the presence of Neu5Gc in recombinant
therapeutic glycoproteins may elicit an immune response. The aim of this work was, therefore, to reduce CMP-Neu5Ac hydroxylase activity
in a Chinese Hamster Ovary (CHO) cell line, and thus the Neu5Gc content of the resulting glycoconjugates, using a rational antisense RNA
approach. For this purpose, the cDNA of the hamster hydroxylase was partially cloned and sequenced. Based on the sequence of the mouse
and hamster cDNAs, optimal antisense RNA fragments were selected from preliminary in vitro translation tests. Compared to the parental
cell line, the new strain (CHO-AsUH2), which was transfected with a 199-bp antisense fragment derived from the mouse CMP-Neu5Ac
hydroxylase cDNA, showed an 80% reduction in hydroxylase activity. An analysis of the sialic acids present in the cells’ own
glycoconjugates revealed a decrease in the percentage of Neu5Gc residues from 4% in the parental cells to less than 1% in the CHO-AsUH2
cell line.
D 2003 Elsevier B.V. All rights reserved.
Keywords: CMP-Neu5Ac hydroxylase; Sialylation; N-glycolylneuraminic acid; CHO cell; Antisense RNA
1. Introduction of N-glycolylneuraminic acid (Neu5Gc). This sialic acid is
Chinese Hamster Ovary (CHO) cells are widely used for
the production of recombinant glycoproteins employed in
human therapy. Although the posttranslational modifica-
tions of recombinant glycoproteins generated in CHO cells
are very similar to that present on human proteins, these
rodent cells do not perfectly mimic the human sialylation
pattern required for bioactivity, stability, and absence of
antigenicity [1,2]. One of the most significant differences
between rodent and human sialylation is the incorporation
0304-4165/03/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0304-4165(03)00137-5
* Corresponding author. Tel.: +33-383-59-5844; fax: +33-383-59-
5804.
E-mail address: [email protected]
(J.-L. Goergen).1 Present address: Keryos SpA, via Maritano 26, I-20097 San Donato
Milanese, Italy.
present in the glycoconjugates of virtually all vertebrates,
with the notable exception of man and chicken. In animals,
Neu5Gc is formed as the sugar-nucleotide cytidine mono-
phosphate-N-glycolylneuraminic acid (CMP-Neu5Gc) by
the action of CMP-Neu5Ac hydroxylase (EC 1.14.13.45)
[3]. In man, the gene encoding this enzyme lacks an exon,
resulting in an incomplete mRNA with a premature termi-
nation signal [4,5]. Due to this mutation, Neu5Gc cannot be
formed by CMP-Neu5Ac hydroxylase in man and is prac-
tically undetectable in healthy human tissues [6], though
small amounts of Neu5Gc are found in gangliosides and
glycoproteins of certain human tumors [7]. Glycoconjugates
containing this common sialic acid are therefore foreign to
humans and can induce an immune response, causing the
formation of so-called Hanganutziu–Deicher antibodies
[8,9]. The appearance of these ‘‘serum sickness’’ antibodies
was observed after inoculation of patients with animal

S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144134
antisera containing glycoconjugates sialylated with Neu5Gc
[10,11]. The titer of the Hanganutziu–Deicher antibodies
seems to be dependent on the presentation of the antigen.
Thus, the injection of recombinant human erythropoietin,
containing about 1% of Neu5Gc in total sialic acids, failed
to produce immune response in humans [12,13], while small
amounts of GM1 (Neu5Gc) introduced with bovine brain
gangliosides are immunogenic in man [14].
Most of CHO cell lines produce recombinant glycopro-
teins with low levels of Neu5Gc comprising approximately
3% of total sialic acid [15]. However, the amount of
Neu5Gc in recombinant glycoproteins may be dependent
on the nature of the protein [16], the place of the insertion of
the recombinant gene in the host genome [17], and the
cultivation conditions [18]. In some CHO clones, Neu5Ac is
almost completely replaced by Neu5Gc genome [17]. All
these facts lead to the possibility that the injection of
Neu5Gc-containing recombinant glycoproteins produced
in CHO-cells could lead to an immunogenic reaction in
man. The generation of a CHO cell line lacking hydroxylase
activity and hence Neu5Gc would therefore be desirable in
order to abolish this potential hazard.
The relatively recent cloning of the mouse CMP-Neu5Ac
hydroxylase cDNA [19] has provided the necessary infor-
mation for a rational genetic approach to suppress or abro-
gate the expression of this gene in biotechnologically
important mammalian cell lines. Gene knock-out, random
mutagenesis and antisense oligonucleotide strategies can be
envisaged to inhibit the expression of CMP-Neu5Ac hy-
droxylase [20]. Due to its selectivity and ease of handling,
antisense RNA provides an accessible approach to this
problem. Antisense oligonucleotides can be introduced into
the cell by transcription of a transfected cDNA fragment or
by exogenous antisense oligonucleotides. The latter method
has the advantage of allowing rapid testing of the antisense
efficiency, but remains transient. However, the expression of
antisense RNAs within the cell can lead to the generation of a
cell line of industrial interest with stable modified properties.
Although several examples of the manipulation of gly-
cosylation in CHO cells using antisense technology have
been published [21–24], only empirical approaches, or the
generation of unstable cell lines were reported. Since the
main difficulty of this strategy is in the choice of antisense
sequences, an in vitro translation assay is necessary for
selecting an antisense fragment that will efficiently hybrid-
ize to its target mRNA [25,26].
In this paper, we describe the generation of a stable CHO
cell line with significantly reduced levels of CMP-Neu5Ac
hydroxylase activity and Neu5Gc amounts, using a rational
antisense RNA strategy. The first part of the paper presents in
vitro assays where different antisense fragments were tested
for their efficacy to inhibit the hydroxylase translation. The
efficiency of a selected antisense fragment was then assessed
in cultured CHO cells by measuring the resulting intracellu-
lar CMP-Neu5Ac hydroxylase activity and the amount of
Neu5Gc present on cellular glycoconjugates.
2. Materials and methods
2.1. Partial cloning and sequencing of cDNA for CMP-
Neu5Ac hydroxylase from CHO-cells
All DNA and RNA manipulations were performed
according to Ref. [27]. Total RNA was prepared from
5� 106 CHO-UH cells (SV Total RNA isolation system,
Promega). RNA integrity was checked by electrophoresis
on a 1% agarose gel. Total cellular RNA (1 Ag) was
reverse-transcribed (Thermoscript-RT, Gibco-BRL) with
random hexamer primers. Two microliters of the reverse-
transcribed mixture was used as a template for PCR
amplification of hamster cDNAs in a mixture containing:
5 Al of 10� PCR buffer, 1 Al of 10 AM dNTPs (Euro-
gentec, Belgium), 20 pmol of each primer (Eurogentec, for
sequences see below) and 5 units of AmpliTaq polymerase
(Perkin-Elmer) in a final volume of 50 Al. Oligonucleotideprimers for amplification of CHO CMP-Neu5Ac hydroxy-
lase were designed from the published mouse CMP-
Neu5Ac hydroxylase cDNA sequence [19]. Sense primer
(5V-GAAACAGACAGCTGAGACCCTGTTGTC-3V) and an-
tisense primer (5V-GGGTGGTGTTAGAGGGAGTTT-
TATCTG-3V) were used to amplify a 1691-bp fragment of
the hamster sequence. Amplification was carried out with
35 cycles of 94 jC for 1 min, 58 jC for 1 min and 72 jCfor 2 min. The PCR products were purified by electropho-
resis on a 1.5% agarose gel (QIAEX, Qiagen), prior to
subcloning into pCR2.1 TOPO (Invitrogen). The plasmid
containing the hamster hydroxylase cDNA cloned in
pCR2.1 TOPO was termed pCRCNAH. These constructs
were introduced in DH5a E. coli cells by electroporation
and three clones were isolated and sequenced (MWG-
Biotech, Germany). The hamster CMP-Neu5Ac hydroxy-
lase cDNA partial sequence reported in this paper has been
submitted to the EMBL Data bank with the accession
number AJ242835.
2.2. Plasmid constructs
For in vitro translation tests, mouse or hamster antisense
fragments were generated from pBSCNAH (complete
mouse CMP-Neu5Ac hydroxylase cDNA sequence cloned
into the pBluescript II K + vector) [19] and pCRCNAH
(hamster hydroxylase cDNA partial sequence cloned into
the pCR2.1 TOPO vector), respectively. The fragments were
subcloned either into pGEM3Z (Promega) or pCR2.1
TOPO. The mouse 403 bp antisense cDNA is a XbaI/SacI
pBSCNAH fragment (nucleotides 1–403 according to Fig.
1) inserted into pGEM3Z. The mouse 714 bp antisense
cDNA (nucleotides 99–812 according to Fig. 1) was
amplified by PCR using pBSCNAH as the template with
sense (5V-GATGATGGACAGGAAACAGACAGC-3V) and
antisense (5V-ACCTGGATC-AGAGTGGCGTCGGGT-3V)primers, and cloned into pCR2.1 TOPO. The mouse and
hamster 199 bp fragments (nucleotides 112–309 according

50 100 mouse ggcagtctaa agtcatcctc gccatcctcg ccttcctggt gtgattcgca gaagtagaaa gaacaccagc agctgctttg aaataccctg gagctggcag hamster ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ 101 150 200 mouse ATGATGGACA GGAAACAGAC AGCTGAGACC CTGCTGACCC TGTCTCCTGC TGAAGTTGCC AACCTCAAGG AAGGGATCAA TTTTTTTCGA AATAAGACTA hamster ~~~~~~~~~~ ~GAAACAGAC AGCTGAGACC CTGTTGTCTC TGTCACCTGC TGAAACTGCC AACCTCAAGG AAGGAATCAA TTTTTTTCGA AATAAGACTA 201 250 300 mouse CTGGGAAAGA GTACATTTTA TACAAGGAGA AGGACCATCT AAAGGCATGC AAGAACCTCT GCAAGCACCA GGGAGGCCTG TTCATGAAAG ACATCGAGGA hamster CTGGCAAAGA GTACATTTTA TACAAGGAGA AAAACCACCT AAAGGCATGC AAGAACCTCT GCAAGCACCA AGGAGGCCTG TTCATAAAAG ACATTGAGGA 301 350 400 mouse TTTAGATGGA AGGTCCGTTA AATGCACAAA GCACAACTGG AAGTTAGACG TGAGCACCAT GAAATATATC AACCCTCCAG GGAGCTTCTG TCAAGACGAG hamster TTTAGACGGA AGGTCCGTTA AATGTACAAA GCACAACTGG AAGTTAGATG TGAGCACCAT GAAGTACATC AACCCTCCAG GGAGCTTCTG TCAGGACGAG 401 450 500 mouse CTCGTTATTG AAATGGATGA AAACAATGGG CTTTCCCTGG TAGAACTGAA CCCTCCTAAC CCCTGGGACT CTGATCCCAG GTCTCCTGAA GAATTAGCTT hamster CTGGTTGTCG AAATGGATGG AAACGATGGG CTTTTCCTTA TAGAATTGAA TCCTCCTAAC CCTTGGGACT CTGATCCCAG GACTCCTGAA GAGTTGGCTT 501 550 600 mouse TTGGGGAAGT ACAGATAACA TATCTCACTC ATGCCTGCAT GGACCTCAAG TTGGGAGACA AGCGAATGGT ATTTGACCCT TGGTTAATTG GCCCTGCTTT hamster TTGGGGAAGT ACAGATAACA TATCTCACTC ATGCCTGCAT GGACCTCAAG TTGGGAGACA AGCGGATGGT GTTTGACCCT TGGTTAATTG GCCCTGCTTT 601 650 700 mouse TGCCCGAGGA TGGTGGTTGC TACATGAGCC TCCATCTGAC TGGTTGGAGA GGCTGTGCAA AGCAGACCTC ATTTATATCA GCCACATGCA CTCAGACCAC hamster TGCCCGAGGA TGGTGGTTGC TACATGAGCC TCCATCTGAT TGGCTGGAGA GGCTGTGCAA AGCAGACCTC ATTTACATCA GTCACATGCA CTCAGACCAC 701 750 800 mouse CTGAGCTACC CTACCCTGAA GCAGCTTTCC CAGAGACGAC CAGACATTCC CATTTATGTT GGCGACACAG AAAGGCCTGT GTTTTGGAAC CTGGATCAGA hamster CTGAGTTACC CCACTCTGAA GCAGCTTTCC CAGAGACGAC CAGACATTCC CATATATGTT GGTGACACGG AAAGGCCTGT GTTTTGGAAT CTGGACCAGA 801 850 900 mouse GTGGCGTCGG GTTAACTAAC ATCAACGTGG TTCCATTTGG AATATGGCAA CAGGTAGACA AAAGTCTGCG GTTCATGATC TTGATGGACG GCGTTCATCC hamster GCGGTGTTCA GCTAACTAAC ATCAATGTGG TACCATTTGG AGTGTGGCAA CAGGTAGACA AAAATCTCCG ATTCATGATC TTGATGGATG GCGTTCATCC 901 950 1000 mouse TGAGATGGAC ACATGCATTA TCGTGGAGTA CAAAGGTCAT AAAATACTCA ACACAGTGGA CTGCACCAGA CCCAATGGGG GAAGGCTTCC TGAGAAAGTT hamster TGAAATGGAC ACATGCATTA TCGTAGAGTA CAAAGGTCAT AAAATACTCA ACACAGTGGA TTGCACCAGA CCCAATGGGG GAAGGCTGCC TGAGAAAGCT 1001 1050 1100 mouse GCTCTAATGA TGAGTGATTT CGCAGGAGGT GCATCAGGCT TTCCAATGAC TTTCAGTGGT GGAAAATTTA CTGAGGAATG GAAAGCCCAG TTCATTAAGG hamster GCTCTAATGA TGAGTGATTT TGCTGGAGGA GCATCAGGCT TTCCAATGAC TTTCAGCGGT GGAAAATTTA CTGAGGAATG GAAGGCCCAG TTCATTAAGG 1101 1150 1200 mouse CTGAAAGAAG AAAGCTTCTG AATTACAAAG CTCAGCTGGT GAAGGACCTG CAGCCCCGAA TCTACTGTCC GTTTGCTGGG TACTTTGTGG AGTCTCACCC hamster CGGAAAGAAG AAAGCTTCTG AATTACAAAG CTCAGCTCGT GAAGGACCTC CAGCCTCGAA TCTACTGTCC CTTTGCTGGG TATTTTGTGG AATCTCATCC 1201 1250 1300 mouse ATCTGACAAG TACATTAAGG AAACAAACAC CAAAAATGAC CCAAATCAGC TCAACAATCT TATCAGGAAA AACTCTGACG TGGTGACATG GACCCCACGA hamster ATCTGACAAG TATATTAAGG AAACAAACAT CAAAAATGAC CCGATTCAAC TCAACAATCT CATCAAGAAA AACTGTGATG TGGTGACATG GACCCCACGA 1301 1350 1400 mouse CCTGGCGCTG TCCTCGACCT TGGCAGGATG CTGAAGGACC CAACAGACAG CAAGGGCATT GTGGAGCCTC CAGAGGGGAC AAAGATTTAC AAGGATTCCT hamster CCTGGAGCTA CTCTTGACCT GGGCAGGATG CTGAAGGACC CAACAGACAG CCAGGGCATC ATAGAGCCTC CAGAAGGGAC AAAAATTTAC AAGGATTCAT 1401 1450 1500 mouse GGGACTTTGG CCCGTACCTG GAGATCTTGA ATTCTGCTGT CAGAGATGAA ATCTTCTGTC ATTCATCCTG GATTAAAGAG TACTTCACGT GGGCTGGATT hamster GGGACTTCGG CCCATACCTG AGCACCTTGC ACTCTGCTGT AGGAGATGAA ATCTTCCTTC ACTCGTCCTG GATAAAAGAG TACTTCACTT GGGCTGGATT 1501 1550 1600 mouse TAAGAATTAC AACCTGGTGG TCAGGATGAT TGAAACAGAT GAAGATTTCA GCCCTTTTCC TGGAGGGTAC GACTATCTGG TGGACTTTCT AGATTTATCC hamster TAAGAGTTAC AACTTGGTGG TCAGGATGAT TGAAACAGAT GAAGACTTCA ACCCTTTTCC TGGAGGGTAT GACTATCTGG TGGACTTTCT AGATTTGTCT 1601 1650 1700 mouse TTTCCGAAAG AAAGACCCAG CCGGGAGCAT CCTTATGAAG AAATCCATAG CCGGGTGGAT GTCATCAGGT ACGTGGTGAA GAACGGCCTG CTGTGGGATG hamster TTTCCAAAAG AAAGACCAAG CAGGGAGCAT CCCTATGAAG AAATCCGTAG CCGTGTGGAT GTCGTCAGGT ACGTGGTGAA GCACGGTCTG CTGTGGGATG 1701 1750 1800 mouse ATCTGTATAT TGGATTCCAG ACCCGATTGC TGCGGGACCC TGATATATAC CATCATCTGT TTTGGAATCA TTTTCAGATA AAACTCCCTC TAACACCACC hamster ACCTGTACAT TGGCTTCCAG ACCCGGTTGC AGCGGGACCC TGACATATAC CATCATCTGT TTTGGAATCA TTTTCAGATA AAACTCCCTC TAACACCACC 1801 1850 mouse CAACTGGAAG TCGTTCCTAA TGCACTGTGA TTAGtctgga cctggtaagt ccca hamster CA~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~
Fig. 1. Hamster and mouse CMP-Neu5Ac hydroxylase cDNA sequence alignment: shaded letters show differences between the mouse and hamster sequences.
Start and stop codons are boxed.
S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144 135
to Fig. 1) were amplified by PCR using identical sense (5V-TTTCTAGAGAAACAGACAGCTGAGACCC-3V) and anti-
sense (5V-TTTGAGCTCC CGTCTAAATCCTCAATGTC-3V)primers. The XbaI/SacI digested PCR products were then
subcloned into pGEM3Z. For cell transfections, the mouse
199, 403 and 714 bp antisense cDNAs were subcloned into
the EcoRI/XbaI-cleaved pClSfiT [28] vector and the ham-
ster 199 bp antisense was subcloned into the same vector
digested with EcoRI and SalI. For selection of stable clones,
plasmid pSfiSVdhfr [29], carrying the dehydrofolate reduc-
tase selectable marker, was used.
2.3. In vitro transcription and translation assays
In vitro transcription reactions were performed using the
‘‘Riboprobe in vitro Transcription System’’ (Promega). T7
RNA polymerase was used for transcription of the mouse
CMP-Neu5Ac hydroxylase cDNA from pBSCNAH linear-
ized with XhoI, as well as the 714, 403 and 199 bp antisense
DNAs generated by cleavage of the appropriate plasmid with
BamHI,XbaI and SalI, respectively. T3 RNApolymerase was
used to produce a full-length antisense mouse CMP-Neu5Ac
hydroxylase RNA from pBSCNAH linearized with NotI.

S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144136
In vitro translation reactions were performed using the
‘‘Rabbit Reticulocyte Lysate System’’ (Promega) and
chemiluminescent detection of the translation products
was performed using the ‘‘Transcendk non-radioactive
Translation Detection System’’ (Promega).
2.4. Generation of stable CHO-AsUH2 cells expressing
CMP-Neu5Ac hydroxylase antisense RNA
The cell line (CHO-UH cell line) used for expression of
antisense fragments results from the co-transfection of a
plasmid bearing the rat a2,6-sialyltransferase cDNA and a
plasmid carrying the neomycin selectable marker into the
Dux-B11 CHO (dhfr-) cell line [2]. CHO-UH cells were
used to generate a second stable cell line producing CMP-
Neu5Ac hydroxylase antisense RNAs. CHO-UH cells were
transfected using a method derived from the in vitro
amplification system described previously [30], in which
lipofection was replaced by electroporation (1400 V, 25 AF)with 10 Ag of a concatenamer composed of the cDNA
coding for the tested antisense RNA and the cDNA coding
for dhfr at a ratio of 10:1. The selective medium used for
CHO cells was aMEM (without ribo- and deoxyribonucleo-
sides) (Gibco-BRL) + 5% dialyzed heat inactivated fetal
calf serum (FCS) (ATGC, France). Methotrexate (MTX)
was used for gene amplification at a starting concentration
of 50 nM and was increased progressively to a final
concentration of 500 nM. Clones were isolated after a 9-
week period in the selection medium. For sialic acid
analyses, cells were maintained in the same selection
medium except that calf serum was replaced by 4% human
serum.
2.5. Preparation of high-speed supernatants from different
cell lines for CMP-Neu5Ac hydroxylase assays
Cell pellets (about 108 cells) were thawed, resuspended
in one volume (ml/g wet mass) of ice-cold 50 mM HEPES/
NaOH, pH 7.4 containing Triton X-100 (5% by mass) and
incubated for 20 min at 4 jC with permanent agitation. The
resulting homogenates were centrifuged for 15 min at
100000� g and 4 jC and the clear high-speed supernatants
were removed and used for determination of the hydroxy-
lase activity.
2.6. Assay of CMP-Neu5Ac hydroxylase activity
The activity of CMP-Neu5Ac hydroxylase was detected
by incubation of 15 Al of the high-speed supernatants with 1
mM NADH, 0.5 mM FeSO4, pig liver microsomes (60 Agprotein solubilised in a final concentration of 0.5% w/v
Triton X-100), and CMP-[4,5,6,7,8,9-14C]Neu5Ac (Amer-
sham) in 50 mM HEPES/NaOH, pH 7.4 at 37 jC, the finalvolume of the assay being 25 Al. The concentration of CMP-
Neu5Ac in the test varied from 2.1 to 18 AM and the
quantity of the radioactivity was 12.5–15.6 nCi per assay.
To obtain substrate concentrations above 12 AM the re-
quired amount of non-radioactive CMP-Neu5Ac was added.
Incubation times are as indicated in Results and all enzyme
tests were performed in duplicate. The assays were stopped
by the addition of 5 Al 1 M trichloroacetic acid, and
precipitated proteins were removed by centrifugation for 3
min at 14000� g. Released [14C]Neu5Gc and [14C]Neu5Ac
were separated and quantified by radio thin-layer chroma-
tography, as described previously [31]. Minute amounts of
activity due to traces of hydroxylase in the pig microsomes,
were subtracted from the sample activity. With this method,
the minimum detectable conversion of CMP-Neu5Ac to
CMP-Neu5Gc was about 1–2%. The activity of each
sample was estimated using two incubation times, both of
which were within the linear region of the reaction time
course (see Results). The apparent KM for CMP-Neu5Ac of
the enzyme in the extracts of CHO-UH cells was determined
using the non-linear regression program Enzfitter (Elsevier
Biosoft, UK).
2.7. RNA analysis by RT-PCR
Total RNA was prepared from CHO-UH and CHO-
AsUH2 cells (SV Total RNA isolation system, Promega)
and the first cDNA strand synthesis was performed using
the AMV reverse transcriptase and random nonamers.
Oligonucleotides specific for the hamster CMP-Neu5Ac
hydroxylase and the expression vector pCISfiT sequences
were used to amplify either the CMP-Neu5Ac hydroxylase
cDNAs or the 199 bp fragments present in the cell lines.
PCR amplification was performed in a final volume of 50
Al, using 0.8 units of AccuTaq polymerase (Sigma) and 1 Agof template cDNA. Amplification was carried out using 35
cycles with a temperature profile of: 40 s at 94 jC, 1 min at
58 jC and 1.5 min at 68 jC. The PCR products were
analysed by electrophoresis on a 1.5% agarose gel. A
similar protocol was used to amplify a h-actin sequence,
with the help of standard oligonucleotides, except that the
temperature of the annealing step was increased to 68 jC.
2.8. Sialic acid analysis
Cells (6–10� 107) were thawed, resuspended in two
volumes (ml/g wet mass) of distilled water and homoge-
nised on ice for 3� 10 s using a Branson B-12 sonicator (40
W power) fitted with a microtip, allowing 15 s cooling
between bursts. To release sialic acids, the resulting homog-
enate (0.1 ml) was mixed with the same volume of 0.2 M
HCl and incubated for 2 h at 80 jC. The samples were
centrifuged for 20 min at 14000� g and the resulting pellets
were washed with 0.1 ml ice-cold water followed by
centrifugation at 14000� g. After both centrifugation steps,
the supernatants were pooled and filtered with a Centricon
10 ultrafiltration unit (Amicon). The retained material was
washed twice with 0.2 ml water followed by ultrafiltration.
The filtrates were lyophilised, and the released sialic acids

S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144 137
were purified by passage through 2 ml cation- and anion-
exchange columns as described by Ref. [32]. The adsorbed
sialic acids were eluted from the latter resin with seven
column volumes of 1 M formic acid, lyophilised and
resuspended in 1 ml water. The enriched sialic acids were
derivatized with 1,2-diamino-4,5-methylene dioxybenzene
(DMB) and the resulting derivatives were analysed by
reversed-phase HPLC with fluorimetric detection [33].
Quantification of sialic acids was performed by integration
of the peaks in the chromatogram using a calibration curve
obtained by DMB-derivatization of known Neu5Ac
amounts.
2.9. Protein concentration
Protein concentrations were measured using the BCA
protein assay reagent (Pierce) with bovine serum albumin as
a standard.
3. Results
3.1. PCR amplification and cloning of the partial CMP-
Neu5Ac hydroxylase cDNA from CHO-cells
Using primers designed from the nucleotide sequence of
the mouse CMP-Neu5Ac hydroxylase cDNA [19], two
products of 1691 and 1250 bp were amplified by RT-PCR
using total RNA from CHO-UH cells as a template (data not
shown). These fragments were subcloned into pCR2.1
TOPO and transformed into E. coli. Three colonies from
each transformation were randomly selected, their plasmids
isolated and the DNA inserts sequenced. The amino acid
sequence deduced from the ORF of the 1250-bp product
exhibited a 98% similarity to fibronectin from CHO, a
protein which is abundantly expressed in CHO cells. The
nucleotide sequence of the 1691-bp product was 94%
identical to that of the mouse CMP-Neu5Ac hydroxylase
cDNA. The deduced sequence of the 563 amino acids was
97% homologous to the mouse enzyme, indicating that
Fig. 2. Schematic representation of the mouse CMP-Neu5Ac hydroxylase cDNA cl
translation inhibition tests.
several nucleotide exchanges are silent (Fig. 1). Due to
the fact that oligonucleotides from the mouse hydroxylase
open reading frame were used to amplify the PCR frag-
ments, the given sequence is only partial, covering 98% of
the mouse CMP-Neu5Ac hydroxylase cDNA. The high
degree of homology between the sequence presented here
and the primary structure of CMP-Neu5Ac hydroxylase
from mouse and other sources [34] confirms its assignment
as CMP-Neu5Ac hydroxylase, presumably the main en-
zyme responsible for the biosynthesis of Neu5Gc in CHO
cells.
3.2. Selection of CMP-Neu5Ac hydroxylase antisense
constructs
Since initial experiments [35,36] showed that artificial
antisense oligonucleotides may be used to interfere with
gene expression, there has been a growing body of literature
on antisense nucleic acids [22,23,37–39]. Due to the lack of
generally applicable rules for the design of efficient anti-
sense RNAs, the ability of different antisense RNAs to
inhibit the translation of the mouse CMP-Neu5Ac hydrox-
ylase mRNA was tested in vitro. The approach adopted
involved examining the efficiency of antisense RNAs of
different lengths by generating fragments from the mouse
CMP-Neu5Ac hydroxylase cDNA. Four different antisense
RNAs of 714, 403, and 199 nucleotides (nt) plus the full-
length of the mouse cDNA were generated (for exact
position on the hydroxylase cDNA see materials and meth-
ods). In addition, a 199 nt fragment from the hamster CMP-
Neu5Ac hydroxylase cDNA spanning the same region as
the mouse 199 nt fragment was tested (Fig. 2).
The mouse CMP-Neu5Ac hydroxylase cDNA and the
antisense fragments were transcribed in vitro with either T7
or T3 RNA polymerases and the formation of the respective
RNA was confirmed by agarose gel electrophoresis by
comparison with four standard RNAs of 250, 1065, 1525,
and 2346 nt (data not shown). The band intensity reflected
the amount of RNA obtained, allowing an estimation of the
sense and antisense RNA quantities used in the in vitro
oned into pGEM3Z and of the sense and antisense RNAs used in the in vitro
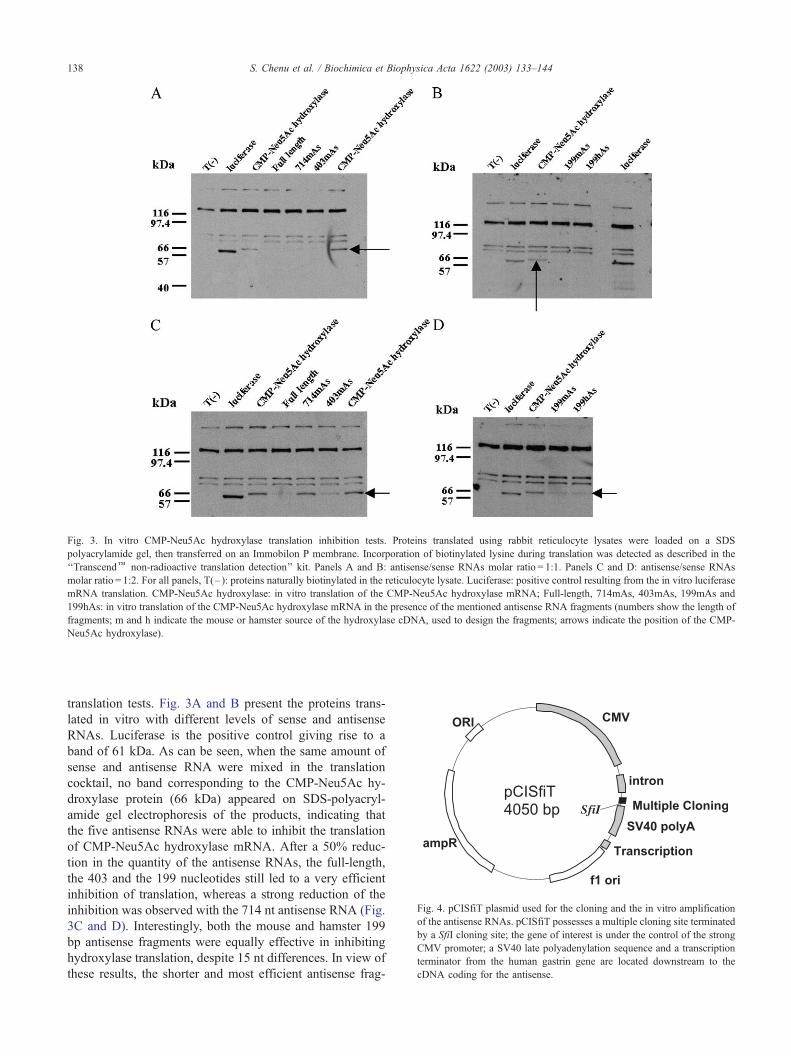
Fig. 4. pCISfiT plasmid used for the cloning and the in vitro amplification
of the antisense RNAs. pCISfiT possesses a multiple cloning site terminated
by a SfiI cloning site; the gene of interest is under the control of the strong
CMV promoter; a SV40 late polyadenylation sequence and a transcription
terminator from the human gastrin gene are located downstream to the
cDNA coding for the antisense.
Fig. 3. In vitro CMP-Neu5Ac hydroxylase translation inhibition tests. Proteins translated using rabbit reticulocyte lysates were loaded on a SDS
polyacrylamide gel, then transferred on an Immobilon P membrane. Incorporation of biotinylated lysine during translation was detected as described in the
‘‘Transcendk non-radioactive translation detection’’ kit. Panels A and B: antisense/sense RNAs molar ratio = 1:1. Panels C and D: antisense/sense RNAs
molar ratio = 1:2. For all panels, T(– ): proteins naturally biotinylated in the reticulocyte lysate. Luciferase: positive control resulting from the in vitro luciferase
mRNA translation. CMP-Neu5Ac hydroxylase: in vitro translation of the CMP-Neu5Ac hydroxylase mRNA; Full-length, 714mAs, 403mAs, 199mAs and
199hAs: in vitro translation of the CMP-Neu5Ac hydroxylase mRNA in the presence of the mentioned antisense RNA fragments (numbers show the length of
fragments; m and h indicate the mouse or hamster source of the hydroxylase cDNA, used to design the fragments; arrows indicate the position of the CMP-
Neu5Ac hydroxylase).
S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144138
translation tests. Fig. 3A and B present the proteins trans-
lated in vitro with different levels of sense and antisense
RNAs. Luciferase is the positive control giving rise to a
band of 61 kDa. As can be seen, when the same amount of
sense and antisense RNA were mixed in the translation
cocktail, no band corresponding to the CMP-Neu5Ac hy-
droxylase protein (66 kDa) appeared on SDS-polyacryl-
amide gel electrophoresis of the products, indicating that
the five antisense RNAs were able to inhibit the translation
of CMP-Neu5Ac hydroxylase mRNA. After a 50% reduc-
tion in the quantity of the antisense RNAs, the full-length,
the 403 and the 199 nucleotides still led to a very efficient
inhibition of translation, whereas a strong reduction of the
inhibition was observed with the 714 nt antisense RNA (Fig.
3C and D). Interestingly, both the mouse and hamster 199
bp antisense fragments were equally effective in inhibiting
hydroxylase translation, despite 15 nt differences. In view of
these results, the shorter and most efficient antisense frag-
S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144 139
ments were selected and cloned into eucaryotic expression
vectors for the in vivo inhibition of CMP-Neu5Ac hydrox-
ylase translation.
3.3. Generation of CMP-Neu5Ac hydroxylase depleted cell
lines
EcoRI–XbaI antisense fragments of 199 and 403 bp
were cloned into the pCISfiT vector (Fig. 4) and expressed
under the control of the CMV promoter. Each of these
constructs was combined with the pSfiSVdhfr plasmid,
which bears the DHFR selectable marker, according to the
in vitro amplification method [30], for the insertion of
multiple copies of the antisense fragment into the genome
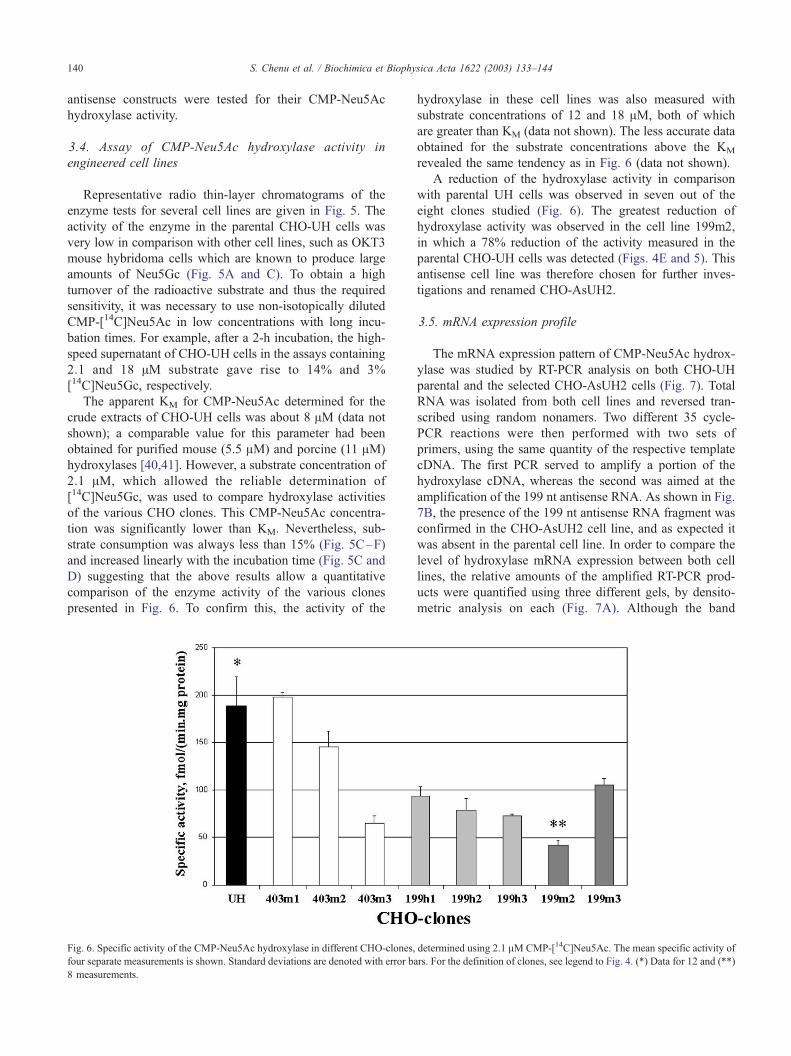
Fig. 5. Radio thin-layer chromatograms of [14C] sialic acids released by acid trea
lines with 2.1 AM CMP-[14C]Neu5Ac and cofactors, as described in Materials and
Al of high-speed supernatant of OKT3 mouse hybridoma cells. Panel B (negative c
supernatant. Panels C and D: analysis of the parental CHO-UH cell line for 60 and
and 403m2 cell lines, respectively. The numbers above the peaks identify the nat
of CHO-UH cells. DHFR-positive transformants were
obtained after 9 weeks by initial selection in aMEM
medium lacking ribo- and deoxyribonucleosides, followed
by exposure to progressively increasing concentrations of
MTX up to 500 nM. One of the main advantages of the in
vitro amplification system is the high rate of good producer
clones generated and then a reduction of the clones to be
screened. The combined in vitro/in vivo amplification
procedures was already used for the production of granulo-
cyte colony-stimulating factor (G-CSF) [29]: a 30-fold
improvement of the protein production was obtained com-
pared to the in vitro amplification protocol alone.
A total of approximately 15 clones was obtained, of
which two to three stable clones from each of the three
tment of incubation mixtures of high-speed supernatants of the various cell
methods. The time of incubation is indicated. Panel A (positive control): 15
ontrol): 15 Al of the 50 mM HEPES/NaOH was used instead of high-speed
120 min, respectively. Panels E and F: analysis for 120 min of the 199m2
ure of the sialic acid ((1) Neu5Gc; (2) Neu5Ac).
S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144140
antisense constructs were tested for their CMP-Neu5Ac
hydroxylase activity.
3.4. Assay of CMP-Neu5Ac hydroxylase activity in
engineered cell lines
Representative radio thin-layer chromatograms of the
enzyme tests for several cell lines are given in Fig. 5. The
activity of the enzyme in the parental CHO-UH cells was
very low in comparison with other cell lines, such as OKT3
mouse hybridoma cells which are known to produce large
amounts of Neu5Gc (Fig. 5A and C). To obtain a high
turnover of the radioactive substrate and thus the required
sensitivity, it was necessary to use non-isotopically diluted
CMP-[14C]Neu5Ac in low concentrations with long incu-
bation times. For example, after a 2-h incubation, the high-
speed supernatant of CHO-UH cells in the assays containing
2.1 and 18 AM substrate gave rise to 14% and 3%
[14C]Neu5Gc, respectively.
The apparent KM for CMP-Neu5Ac determined for the
crude extracts of CHO-UH cells was about 8 AM (data not
shown); a comparable value for this parameter had been
obtained for purified mouse (5.5 AM) and porcine (11 AM)
hydroxylases [40,41]. However, a substrate concentration of
2.1 AM, which allowed the reliable determination of
[14C]Neu5Gc, was used to compare hydroxylase activities
of the various CHO clones. This CMP-Neu5Ac concentra-
tion was significantly lower than KM. Nevertheless, sub-
strate consumption was always less than 15% (Fig. 5C–F)
and increased linearly with the incubation time (Fig. 5C and
D) suggesting that the above results allow a quantitative
comparison of the enzyme activity of the various clones
presented in Fig. 6. To confirm this, the activity of the
Fig. 6. Specific activity of the CMP-Neu5Ac hydroxylase in different CHO-clones,
four separate measurements is shown. Standard deviations are denoted with error ba
8 measurements.
hydroxylase in these cell lines was also measured with
substrate concentrations of 12 and 18 AM, both of which
are greater than KM (data not shown). The less accurate data
obtained for the substrate concentrations above the KM
revealed the same tendency as in Fig. 6 (data not shown).
A reduction of the hydroxylase activity in comparison
with parental UH cells was observed in seven out of the
eight clones studied (Fig. 6). The greatest reduction of
hydroxylase activity was observed in the cell line 199m2,
in which a 78% reduction of the activity measured in the
parental CHO-UH cells was detected (Figs. 4E and 5). This
antisense cell line was therefore chosen for further inves-
tigations and renamed CHO-AsUH2.
3.5. mRNA expression profile
The mRNA expression pattern of CMP-Neu5Ac hydrox-
ylase was studied by RT-PCR analysis on both CHO-UH
parental and the selected CHO-AsUH2 cells (Fig. 7). Total
RNA was isolated from both cell lines and reversed tran-
scribed using random nonamers. Two different 35 cycle-
PCR reactions were then performed with two sets of
primers, using the same quantity of the respective template
cDNA. The first PCR served to amplify a portion of the
hydroxylase cDNA, whereas the second was aimed at the
amplification of the 199 nt antisense RNA. As shown in Fig.
7B, the presence of the 199 nt antisense RNA fragment was
confirmed in the CHO-AsUH2 cell line, and as expected it
was absent in the parental cell line. In order to compare the
level of hydroxylase mRNA expression between both cell
lines, the relative amounts of the amplified RT-PCR prod-
ucts were quantified using three different gels, by densito-
metric analysis on each (Fig. 7A). Although the band
determined using 2.1 AM CMP-[14C]Neu5Ac. The mean specific activity of
rs. For the definition of clones, see legend to Fig. 4. (*) Data for 12 and (**)
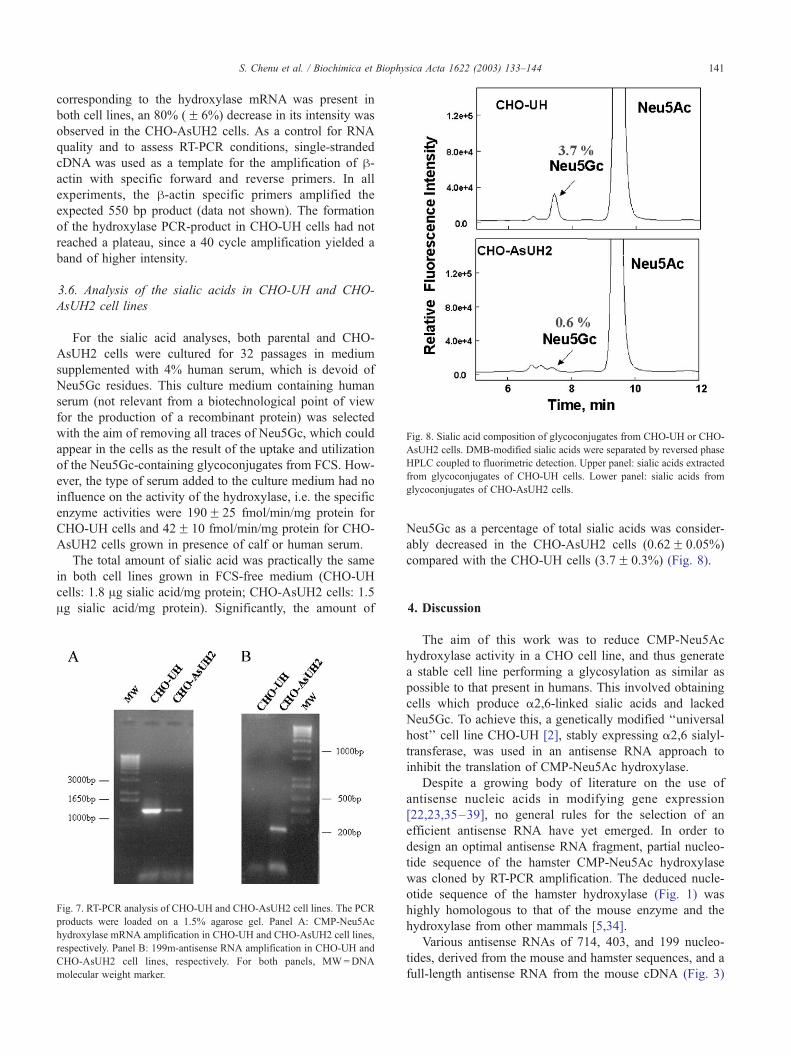
Fig. 8. Sialic acid composition of glycoconjugates from CHO-UH or CHO-
AsUH2 cells. DMB-modified sialic acids were separated by reversed phase
HPLC coupled to fluorimetric detection. Upper panel: sialic acids extracted
from glycoconjugates of CHO-UH cells. Lower panel: sialic acids from
glycoconjugates of CHO-AsUH2 cells.
S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144 141
corresponding to the hydroxylase mRNA was present in
both cell lines, an 80% (F 6%) decrease in its intensity was
observed in the CHO-AsUH2 cells. As a control for RNA
quality and to assess RT-PCR conditions, single-stranded
cDNA was used as a template for the amplification of h-actin with specific forward and reverse primers. In all
experiments, the h-actin specific primers amplified the
expected 550 bp product (data not shown). The formation
of the hydroxylase PCR-product in CHO-UH cells had not
reached a plateau, since a 40 cycle amplification yielded a
band of higher intensity.
3.6. Analysis of the sialic acids in CHO-UH and CHO-
AsUH2 cell lines
For the sialic acid analyses, both parental and CHO-
AsUH2 cells were cultured for 32 passages in medium
supplemented with 4% human serum, which is devoid of
Neu5Gc residues. This culture medium containing human
serum (not relevant from a biotechnological point of view
for the production of a recombinant protein) was selected
with the aim of removing all traces of Neu5Gc, which could
appear in the cells as the result of the uptake and utilization
of the Neu5Gc-containing glycoconjugates from FCS. How-
ever, the type of serum added to the culture medium had no
influence on the activity of the hydroxylase, i.e. the specific
enzyme activities were 190F 25 fmol/min/mg protein for
CHO-UH cells and 42F 10 fmol/min/mg protein for CHO-
AsUH2 cells grown in presence of calf or human serum.
The total amount of sialic acid was practically the same
in both cell lines grown in FCS-free medium (CHO-UH
cells: 1.8 Ag sialic acid/mg protein; CHO-AsUH2 cells: 1.5
Ag sialic acid/mg protein). Significantly, the amount of
Fig. 7. RT-PCR analysis of CHO-UH and CHO-AsUH2 cell lines. The PCR
products were loaded on a 1.5% agarose gel. Panel A: CMP-Neu5Ac
hydroxylase mRNA amplification in CHO-UH and CHO-AsUH2 cell lines,
respectively. Panel B: 199m-antisense RNA amplification in CHO-UH and
CHO-AsUH2 cell lines, respectively. For both panels, MW=DNA
molecular weight marker.
Neu5Gc as a percentage of total sialic acids was consider-
ably decreased in the CHO-AsUH2 cells (0.62F 0.05%)
compared with the CHO-UH cells (3.7F 0.3%) (Fig. 8).
4. Discussion
The aim of this work was to reduce CMP-Neu5Ac
hydroxylase activity in a CHO cell line, and thus generate
a stable cell line performing a glycosylation as similar as
possible to that present in humans. This involved obtaining
cells which produce a2,6-linked sialic acids and lacked
Neu5Gc. To achieve this, a genetically modified ‘‘universal
host’’ cell line CHO-UH [2], stably expressing a2,6 sialyl-
transferase, was used in an antisense RNA approach to
inhibit the translation of CMP-Neu5Ac hydroxylase.
Despite a growing body of literature on the use of
antisense nucleic acids in modifying gene expression
[22,23,35–39], no general rules for the selection of an
efficient antisense RNA have yet emerged. In order to
design an optimal antisense RNA fragment, partial nucleo-
tide sequence of the hamster CMP-Neu5Ac hydroxylase
was cloned by RT-PCR amplification. The deduced nucle-
otide sequence of the hamster hydroxylase (Fig. 1) was
highly homologous to that of the mouse enzyme and the
hydroxylase from other mammals [5,34].
Various antisense RNAs of 714, 403, and 199 nucleo-
tides, derived from the mouse and hamster sequences, and a
full-length antisense RNA from the mouse cDNA (Fig. 3)

S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144142
were preliminary tested in hydroxylase translation inhibition
in vitro tests with different amounts of antisense and sense
RNAs. All antisense RNAs, independent of the animal
source, were able to inhibit the translation of the hydroxy-
lase in vitro, though the 714 nt fragment was markedly less
effective, probably due to the formation of stable secondary
structures. The shorter antisense RNAs, which appeared to
be most efficient, were therefore used to generate stable
transfectants of the CHO-UH cells.
Several approaches were tested to analyse which of the
cell lines expressing different antisense fragments were most
efficient at suppressing hydroxylase production. However,
due to the very low level of enzyme in the parental CHO-
UH cells, the sensitivity of the detection methods was
generally insufficient. For example, the screening of a
CHO cDNA library using the full-length mouse CMP-
Neu5Ac hydroxylase cDNA as a probe, failed to detect
any hydroxylase-specific clones (data not shown). Accord-
ingly, a RT-PCR analysis showed that mRNA coding for
CMP-Neu5Ac hydroxylase is expressed at a very low level
in CHO-UH cells (Fig. 7A). Furthermore, a sensitive
Western blot analysis with a CMP-Neu5Ac hydroxylase-
specific antibody, at a detection limit of about 20 ng
enzyme, failed to reveal the hydroxylase in 90 Ag of total
protein from high-speed supernatants of CHO-UH cells
resolved by SDS-PAGE (data not shown). Determining
Neu5Gc was also a misleading measure of cellular hydrox-
ylase, due to the small amount of this sialic acid (3.7% of
total sialic acid) in the parental CHO-UH cell line. This
latter analysis is further complicated by the fact that cells in
culture incorporate Neu5Gc from FCS present in the medi-
um [42,43]. For this reason, screening the clones by sialic
acid analysis was only meaningful after cultivating cells for
a long period of time in FCS-free medium. Despite the fact
that sub-KM CMP-[14C]Neu5Ac concentrations had to be
used, the sensitive radio-TLC-based assay provided a quan-
titative measure of the relative hydroxylase content of the
various cell lines.
Using this enzyme assay, it was first shown that most of
the tested clones displayed a reduced CMP-Neu5Ac hy-
droxylase activity; in fact, this is probably due to the use of
the combined in vitro/in vivo amplification procedures
yielding a high rate of strong producer clones. Second, the
efficiency of the antisense fragments in vivo was indepen-
dent of the animal source, and the 199 nt fragments were
more effective than the 403 nt antisense RNAs at suppress-
ing hydroxylase expression (Fig. 6). The cell line trans-
fected with the 199 nt fragment derived from the cDNA of
the mouse enzyme exhibited the greatest (78%) reduction in
the hydroxylase activity and was termed CHO-AsUH2.
Although the latter antisense fragment differed from the
corresponding hamster fragment by 15 nt, both were effi-
cient at inhibiting the translation of the hydroxylase mRNA
in vivo. In fact, these sequence differences could be advan-
tageous, since natural antisense RNAs do not exhibit com-
plete complementary to the target mRNA and thus are less
prone to intracellular RNases [44,45]. Two different mech-
anisms could account for the inhibition of translation by
antisense fragments [37]: (i) steric blocking of translation
initiation, and (ii) degradation of the antisense RNA-mRNA
duplex by specific RNases. However, since none of the 199
nt antisense fragments was located upstream the AUG
initiator codon, the reduction of CMP-Neu5Ac hydroxylase
activity in CHO-AsUH2 is probably mainly due to the
degradation of mRNAs. Indeed, the semi-quantitative esti-
mation of the amounts of mRNA by RT-PCR showed a
significant reduction (approximately 80%) in CMP-Neu5Ac
hydroxylase mRNA in the CHO-AsUH2 cell line as com-
pared to CHO-UH cells (Fig. 7A). This result is also in a
good agreement with previous findings demonstrating that
(antisense RNA-sense mRNA) hybrids are rapidly degraded
by RNases specific for RNA duplexes, leading to a decrease
in the endogenous message levels of a targeted gene [46,47].
An analysis of Neu5Gc, the product of the hydroxylase
reaction, in total cellular glycoconjugates produced by cells
cultivated for 32 passages in the presence of human serum,
revealed an 80% decrease in the amount of Neu5Gc in the
CHO-AsUH2 cell line relative to the parental cells (Fig. 8).
Since a reduction in CMP-Neu5Ac hydroxylase activity of
about 80% led to the same decrease in the amount of
Neu5Gc residues present on the glycoconjugates in the
new CHO-AsUH2 cell line, the amount of hydroxylase
protein appears to be governing the incorporation of
Neu5Gc into glycoconjugates. This is in agreement with
previous studies on various mammalian tissues [33,41,48] in
which the amount of Neu5Gc was found to be regulated by
the level of hydroxylase activity. This, in turn, generally
correlates with the quantity of hydroxylase protein. More-
over, the estimated 80% reduction in the levels of hydrox-
ylase mRNA found in CHO-AsUH2 cells suggests that in
this cell line, the observed decrease in the CMP-Neu5Ac
hydroxylase activity and the amount of Neu5Gc are due to
the presence of antisense RNA.
Although we were able to significantly reduce the CMP-
Neu5Ac hydroxylase activity in CHO-UH cells, the anti-
sense RNA approach does not yield a complete suppression
of the enzyme activity. This agrees with previous reports on
the inhibition of sialidase activity in CHO cells [23]. Despite
the fact that the CMP-Neu5Ac hydroxylase mRNA only
seems to be present in a low copy number, and the antisense
fragment was amplified by means of DHFR-MTX selection
and in vitro concatenamer amplification systems, the CHO-
AsUH2 cells still exhibited 20% of the parental enzyme
activity. Nevertheless, compared to previous studies [21,23,
38], the approach described here enabled the generation of a
stable cell line which retained a significant reduction of the
target enzyme activity for more than 30 passages. Though
the overproduction of a recombinant protein by this cell line
might require the use of a selection/amplification marker
different from DHFR/MTX (for instance glutamine syn-
thase/methionine sulfoximine) [49], these new CHO-
AsUH2 cells are suitable for industrial purposes and further

S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144 143
studies will focus on the assessment of the reduction of
Neu5Gc levels in recombinant glycoproteins produced by
these cells.
Acknowledgements
This work was partially supported by the European
community, grant No. 960767. The authors thank Dr.
Kawano for providing the pBSCNAH vector containing the
mouse CMP-Neu5Ac hydroxylase cDNA.
References
[1] N. Jenkins, E.M. Curling, Glycosylation of recombinant proteins: prob-
lems and prospects. Enzyme Microb. Technol. 16 (1994) 354–364.
[2] A. Bragonzi, G. Distefano, L.D. Buckberry, G. Acerbis, C. Foglieni,
D. Lamotte, G. Campi, A. Marc, M.R. Soria, N. Jenkins, L. Monaco,
A new Chinese hamster ovary cell line expressing a2,6-sialyltransfer-
ase used as universal host for the production of human-like sialylated
recombinant glycoproteins, Biochim. Biophys. Acta 1474 (2000)
273–282.
[3] L. Shaw, R. Schauer, The biosynthesis of N-glycoloylneuraminic acid
occurs by hydroxylation of the CMP-glycoside of N-acetylneuraminic
acid, Biol. Chem. Hoppe-Seyler 369 (1988) 477–486.
[4] A. Irie, A. Suzuki, CMP-N-acetylneuraminic acid hydroxylase is ex-
clusively inactive in humans. Biochem. Biophys. Res. Commun. 248
(1998) 330–333.
[5] H.H. Chou, H. Takematsu, S. Diaz, J. Iber, E. Nickerson, K.L.Whright,
E.A. Muchmore, D.L. Nelson, S.T. Warren, A. Varki, A mutation in
human CMP-sialic acid hydroxylase occurred after the homo-Pan di-
vergence, Proc. Natl. Acad. Sci. U. S. A. 95 (1998) 11751–11756.
[6] E.A. Muchmore, S. Diaz, A. Varki, A structural difference between
the cell surfaces of humans and the great apes, Am. J. Phys. Anthro-
pol. 107 (1998) 187–198.
[7] H. Higashi, N-glycolylneuraminic acid-containing glycoconjugates
as tumor-associated antigen, Trends Glycosci. Glycotech. 2 (1990)
7–15.
[8] H. Higashi, M. Naiki, S. Matuo, K. Okouchi, Antigen of ‘‘serum
sickness’’ type of heterophile antibodies in human sera: identification
as gangliosides with N-glycolylneuraminic acid, Biochem. Biophys.
Res. Commun. 79 (1977) 388–395.
[9] J.M. Merrick, K. Zadarlik, F. Milgrom, Characterization of the Hang-
anutziu–Deicher (serum-sickness) antigen as gangliosides containing
N-glycolylneuraminic acid, Int. Arch. Allergy Appl. Immunol. 57
(1978) 477–480.
[10] M. Hanganutziu, Hemagglutinines heterogenetiques apres injection
de serum de Cheval, C.R. Seances-Soc. Biol. 91 (1924) 1457–1459.
[11] H. Deicher, Uber die Erzeugung heterospezifischer Hamagglutinine
durch Injektion artfremden Serums, Z. Hyg. 106 (1926) 561–579.
[12] A. Noguchi, C.J. Mukuria, E. Suzuki, M. Naiki, Immunogenicity of
N-glycolylneuraminic acid-containing carbohydrate chains of re-
combinant human erythropoietin expressed in CHO cells, J. Biochem.
117 (1995) 59–62.
[13] A. Noguchi, C.J. Mukuria, E. Suzuki, M. Naiki, Failure of human
immunoresponse to N-glycolylneuraminic acid epitope contained in
recombinant human erythropoietin, Nephron 72 (1996) 599–603.
[14] M. Odaka, N. Yuki, H. Yoshino, T. Kasama, S. Handa, F. Irie, Y.
Hirabayashi, A. Suzuki, K. Hirata, N-glycolylneuraminic acid-con-
taining GM1 is a new molecule for serum antibody in Guillain–Barre
syndrome, Ann. Neurol. 43 (1998) 829–834.
[15] C.H. Hokke, A.A. Bergwerff, D.W. van Dedem, J. van Oostrum, J.P.
Kamerling, J.F. Vliegenthart, Sialylated carbohydrate chains of re-
combinant human glycoproteins expressed in Chinese hamster ovary
cells contain traces of N-glycolylneuraminic acid, FEBS Lett. 275
(1990) 9–14.
[16] C.P. Quan, J. Cacia, J. Frenz, J.V. O’Conner, RhDNase is a well
characterized protein, Pharm. Sci. 3 (1997) 53–57.
[17] S.C. Hubbard, L. Walls, H.E. Ruley, E.A. Muchmore, Generation of
CHO cell glycosylation mutant by retroviral insertional mutagenesis.
Integration into a discrete locus generates mutants expressing high
levels of N-glycolylneuraminic acid, J. Biol. Chem. 269 (1994)
3717–3724.
[18] R. Kimura, W.M. Miller, Glycosylation of CHO-derived recombinant
tPA produced under elevated pCO2, Biotechnol. Prog. 13 (1997)
311–317.
[19] T. Kawano, S. Koyama, H. Takematsu, Y. Kozutsumi, H. Kawasaki,
S. Kawashima, T. Kawasaki, A. Suzuki, Molecular cloning of cytidine
monophospho-N-acetylneuraminic acid hydroxylase, J. Biol. Chem.
270 (1995) 16458–16463.
[20] T.G. Warner, Enhancing therapeutic glycoprotein production in CHO
cells by metabolic engineering endogenous gene control with anti-
sense DNA and gene targeting, Glycobiology 9 (1999) 841–850.
[21] E.G.P. Prati, P. Scheidegger, A.R. Sburlati, J.E. Bailey, Antisense
strategies for glycosylation engineering of Chinese Hamster Ovary
(CHO) cells, Biotechnol. Bioeng. 59 (1998) 445–450.
[22] E.G. Prati, M. Matasci, T.B. Suter, A.R. Sburlati, J.E. Bailey, Engi-
neering of coordinated up- and down-regulation of two glycosyltrans-
ferases of the O-glycosylation pathway in Chinese hamster ovary
(CHO) cells, Biotechnol. Bioeng. 68 (2000) 239–244.
[23] J. Ferrari, J. Gunson, J. Lofgren, L. Krummen, T.G. Warner, Chinese
Hamster Ovary cells with constitutively expressed sialidase antisense
RNA produce recombinant DNase in batch culture with increased
sialic acid, Biotechnol. Bioeng. 60 (1998) 589–595.
[24] N. Hiraiwa, T. Dohi, N. Kawakami-Kimura, M. Yumen, K. Ohmori,
M. Maeda, R. Kannagi, Suppression of sialyl Lewis X expression and
E-selectin-mediated cell adhesion in cultured human lymphoid cells
by transfection of antisense cDNA of an (1 –3 fucosyltransferase
(Fuc-T VII), J. Biol. Chem. 271 (1996) 31556–31561.
[25] N. Milner, K.U. Mir, E.M. Southern, Selecting effective antisense
reagents on combinatorial oligonucleotide arrays, Nat. Biotechnol.
15 (1997) 537–541.
[26] J. Wu, M.A. Gerber, The inhibitory effects of antisense RNA on
hepatitis B virus surface antigen synthesis, J. Gen. Virol. 78 (1997)
641–647.
[27] J. Sambrook, E.F. Fritsch, T. Maniatis, Molecular Cloning, A Labo-
ratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press, Cold
Spring Harbor, NY, 1989.
[28] L. Monaco, Optimizing gene expression in mammalian cells, Meth-
ods Biotechnol. 8 (1999) 39–48.
[29] L. Monaco, R. Tagliabue, S. Giovanazzi, A. Bragonzi, M. Soria,
Expression of human granulocyte colony-stimulating factor in CHO
dhfr-cells: new insights into the in vitro amplification expression
system, Gene 180 (1996) 145–150.
[30] L. Monaco, R. Tagliabue, M.R. Soria, M. Uhlen, An in vitro ampli-
fication approach for the expression of recombinant proteins in mam-
malian cells, Biotechnol. Appl. Biochem. 20 (1994) 157–171.
[31] L. Shaw, R. Schauer, Detection of CNP-N-acetylneuraminic acid hy-
droxylase activity in fractionated mouse liver, Biochem. J. 263 (1989)
355–363.
[32] G. Reuter, R. Schauer, Determination of sialic acids, Methods Enzy-
mol. 230 (1994) 168–199.
[33] Y.N. Malykh, L. Shaw, R. Schauer, The role of CMP-N-acetylneur-
aminic acid hydroxylase in determining the level of N-glycolylneur-
aminic acid in porcine tissues, Glycoconj. J. 15 (1998) 885–893.
[34] W. Schlenzka, L. Shaw, S. Kelm, C.L. Schmidt, E. Bill, A.X. Traut-
wein, F. Lottspeich, R. Schauer, CMP-N-acetylneuraminic acid hy-
droxylase: the first cytosolic Rieske iron– sulphur protein to be
described in Eukarya, FEBS Lett. 385 (1996) 197–200.
[35] P.C. Zamecnik, M.L. Stephenson, Inhibition of Rous sarcoma virus

S. Chenu et al. / Biochimica et Biophysica Acta 1622 (2003) 133–144144
replication and cell transformation by specific oligodeoxynucleotides,
Proc. Natl. Acad. Sci. U. S. A. 75 (1978) 280–284.
[36] B.M. Paterson, B.E. Roberts, E.L. Kuff, Structural gene identification
and mapping by DNA–mRNA hybrid-arrested cell-free translation,
Proc. Natl. Acad. Sci. U. S. A. 74 (1977) 4370–4374.
[37] H. Helene, J.J. Toulme, Specific regulation of gene expression by
antisense, sense and antigene nucleic acids, Biochim. Biophys. Acta
1049 (1990) 99–125.
[38] C. Parades, E. Prats, J.J. Cairo, F. Azorin, Ll. Cornudella, F. Godia,
Modification of glucose and glutamine metabolism in hybridoma cells
through metabolic engineering, Cytotechnology 30 (1999) 85–93.
[39] H. Kirschke, R. Eerola, V.K. Hopsu-Havu, D. Bromme, E. Vuorio,
Antisense RNA inhibition of cathepsin L expression reduces tumor-
igenicity of malignant cells, Eur. J. Cancer 36 (2000) 787–795.
[40] P. Schneckenburger, L. Shaw, R. Schauer, Purification, characteriza-
tion and reconstitution of CMP-N-acetylneuraminate hydroxylase
from mouse liver, Glycoconj. J. 11 (1994) 194–203.
[41] W. Schlenzka, L. Shaw, P. Schneckenburger, R. Schauer, Purification
and characterization of CMP-N-acetylneuraminic acid hydroxylase
from pig submandibular glands, Glycobiology 4 (1994) 675–683.
[42] K. Furukawa, H. Yamaguchi, H.F. Oettgen, L.J. Old, K.O. Lloyd,
Analysis of the expression of N-glycolylneuraminic acid-containing
gangliosides in cells and tissues using two human monoclonal anti-
bodies, J. Biol. Chem. 263 (1988) 18507–18512.
[43] J.A. Ecsedy, K.A. Holthaus, H.C. Yohe, T.N. Seyfried, Expression of
mouse sialic acid on gangliosides of a human glioma grown as a
xenograft in SCID mice, J. Neurochem. 73 (1999) 254–259.
[44] W. Nellen, G. Sczakiel, In vitro and in vivo action of antisense RNA,
Mol. Biotechnol. 6 (1996) 7–15.
[45] N. Delihas, S.E. Rokita, P. Zheng, Natural antisense RNA/target RNA
interactions: possible models for antisense oligonucleotide drug de-
sign, Nat. Biotechnol. 15 (1997) 751–754.
[46] J.T. Stout, C.T. Caskey, Antisense RNA inhibition of endogenous
genes, in: Methods in Enzymology, vol. 151, Academic Press, New
York, 1987, pp. 519–530.
[47] H. Wu, A.R. MacLeod, W.F. Lima, S.T. Crooke, Identification and
partial purification of human double strand RNase activity. A novel
terminating mechanism for oligonucleotide antisense drugs, J. Biol.
Chem. 273 (1998) 2532–2542.
[48] T. Kawano, Y. Kozutsumi, T. Kawasaki, A. Suzuki, Biosynthesis of N-
glycolylneuraminic acid-containing glycoconjugates, J. Biol. Chem.
269 (1994) 9024–9029.
[49] T. Omasa, Gene amplification and its application in cell and tissue
engineering, J. Biosci. Bioeng. 94 (2002) 600–605.
Related Documents











