ORIGINAL INVESTIGATION Reduction in the latency of action of antidepressants by 17 β-estradiol in the forced swimming test E. Estrada-Camarena & N. M. Vega Rivera & C. Berlanga & A. Fernández-Guasti Received: 22 June 2007 / Accepted: 30 July 2008 # Springer-Verlag 2008 Abstract Rationale Antidepressants (ADs) are slow to produce their therapeutic effect. This long latency promotes the develop- ment of new strategies to short their onset of action. Previous reports indicated that 17β-estradiol (E 2 ) promotes the antidepressant-like activity of fluoxetine (FLX) and desipramine (DMI) in the forced swimming test (FST). Objective The aim of the present work was to analyze if E 2 reduces the antidepressant-like onset of action of venlafax- ine (VLX), FLX, and DMI. Materials and methods Independent groups of ovariecto- mized female Wistar rats were tested in the FST and in the open field after chronic (1 to 14 days) treatment with VLX (20 mg/kg/day), FLX (1.25 mg/kg/day), or DMI (1.25 mg/kg/day) alone or in combination with a single injection of E 2 (2.5 μg/rat sc, 8 h before FST). Results VLX, FLX, or DMI by themselves at these doses did not induce changes in the FST at short intervals after their injection (from 1 to 7 days). The addition of E 2 promoted the antidepressant-like effect of VLX and DMI as early as day 1. Such action was also evident after 3, for FLX, and 14 days for both FLX and DMI, but not for VLX. The behavioral actions of these ADs combined with E 2 were not accompanied by increases in general activity in the open-field test. Conclusion E 2 clearly reduced the latency to the onset of action for these ADs in the FST. These results represent an interesting therapeutic strategy for the treatment of depres- sion in perimenopausal women. Keywords 17β-Estradiol . Fluoxetine . Desipramine . Venlafaxine . Forced swimming test . Shorter onset of action Introduction Conventional antidepressants (ADs) have many limitations for the effective treatment of depression. In general, ADs are slow to produce their effect (around 2 to 3 weeks) and have a side-effect profile which limits compliance. There- fore, it is necessary to identify pharmacological strategies that offer a faster onset of action. In this line of research, the combination of ADs which act by inhibiting mono- amine reuptake with drugs like pindolol (Artigas et al. 1994; Blier and Bergeron 1995; Pérez et al. 1997; Shiah et al. 2000) or lithium (Austin et al. 1991; Bauer and Döpfmer 1999) has proved to shorten their latency of effect. On the other hand, several reports indicate that estrogens participate in the modulation of depression associated with the endocrine changes along the women’s life span (Genazzani et al. 1999; Robinson 2001; Stahl 1998). Although controversial, there are reports that therapy with some estrogens decrease depressive symptoms and improve the action of antidepressants like imipramine, sertraline, and fluoxetine (Amsterdam et al. 1999; Halbreich and Kahn 2001; Oppenheim 1983; Prange 1972; Robinson 2001; Psychopharmacology DOI 10.1007/s00213-008-1291-8 E. Estrada-Camarena : N. M. Vega Rivera Farmacología Conductual Dirección de Neurociencias, Instituto Nacional de Psiquiatría “Ramón de la Fuente Muñíz”, México City, Mexico C. Berlanga Dirección de Investigaciones Clínicas, Instituto Nacional de Psiquiatría “Ramón de la Fuente Muñíz”, México City, Mexico N. M. Vega Rivera : A. Fernández-Guasti (*) Departamento de Farmacobiología, Centro de Investigación y de Estudios Avanzados del IPN, Calzada de los Tenorios 235, Col Granjas Coapa, Tlalpan, México City, Mexico e-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL INVESTIGATION
Reduction in the latency of action of antidepressantsby 17 β-estradiol in the forced swimming test
E. Estrada-Camarena & N. M. Vega Rivera &
C. Berlanga & A. Fernández-Guasti
Received: 22 June 2007 /Accepted: 30 July 2008# Springer-Verlag 2008
AbstractRationale Antidepressants (ADs) are slow to produce theirtherapeutic effect. This long latency promotes the develop-ment of new strategies to short their onset of action.Previous reports indicated that 17β-estradiol (E2) promotesthe antidepressant-like activity of fluoxetine (FLX) anddesipramine (DMI) in the forced swimming test (FST).Objective The aim of the present work was to analyze if E2
reduces the antidepressant-like onset of action of venlafax-ine (VLX), FLX, and DMI.Materials and methods Independent groups of ovariecto-mized female Wistar rats were tested in the FST and inthe open field after chronic (1 to 14 days) treatment withVLX (20 mg/kg/day), FLX (1.25 mg/kg/day), or DMI(1.25 mg/kg/day) alone or in combination with a singleinjection of E2 (2.5 μg/rat sc, 8 h before FST).Results VLX, FLX, or DMI by themselves at these dosesdid not induce changes in the FST at short intervals aftertheir injection (from 1 to 7 days). The addition of E2
promoted the antidepressant-like effect of VLX and DMI asearly as day 1. Such action was also evident after 3, for
FLX, and 14 days for both FLX and DMI, but not for VLX.The behavioral actions of these ADs combined with E2
were not accompanied by increases in general activity inthe open-field test.Conclusion E2 clearly reduced the latency to the onset ofaction for these ADs in the FST. These results represent aninteresting therapeutic strategy for the treatment of depres-sion in perimenopausal women.
Keywords 17β-Estradiol . Fluoxetine . Desipramine .
Venlafaxine . Forced swimming test . Shorter onset of action
Introduction
Conventional antidepressants (ADs) have many limitationsfor the effective treatment of depression. In general, ADsare slow to produce their effect (around 2 to 3 weeks) andhave a side-effect profile which limits compliance. There-fore, it is necessary to identify pharmacological strategiesthat offer a faster onset of action. In this line of research,the combination of ADs which act by inhibiting mono-amine reuptake with drugs like pindolol (Artigas et al.1994; Blier and Bergeron 1995; Pérez et al. 1997; Shiah etal. 2000) or lithium (Austin et al. 1991; Bauer and Döpfmer1999) has proved to shorten their latency of effect.
On the other hand, several reports indicate that estrogensparticipate in the modulation of depression associated withthe endocrine changes along the women’s life span(Genazzani et al. 1999; Robinson 2001; Stahl 1998).Although controversial, there are reports that therapy withsome estrogens decrease depressive symptoms and improvethe action of antidepressants like imipramine, sertraline,and fluoxetine (Amsterdam et al. 1999; Halbreich and Kahn2001; Oppenheim 1983; Prange 1972; Robinson 2001;
PsychopharmacologyDOI 10.1007/s00213-008-1291-8
E. Estrada-Camarena :N. M. Vega RiveraFarmacología Conductual Dirección de Neurociencias,Instituto Nacional de Psiquiatría “Ramón de la Fuente Muñíz”,México City, Mexico
C. BerlangaDirección de Investigaciones Clínicas,Instituto Nacional de Psiquiatría “Ramón de la Fuente Muñíz”,México City, Mexico
N. M. Vega Rivera :A. Fernández-Guasti (*)Departamento de Farmacobiología,Centro de Investigación y de Estudios Avanzados del IPN,Calzada de los Tenorios 235, Col Granjas Coapa, Tlalpan,México City, Mexicoe-mail: [email protected]

Schneider et al. 1997, 2001; Shapira et al. 1985), particularlyduring perimenopause. Furthermore, Oppenheim (1983)suggested that estrogens facilitated that action of ADs byshortening their latency of effect.
Recently, using an animal model for the screening ofantidepressant drugs, the forced swimming test (FST; Porsoltet al. 1977; Porsolt and Lenègre 1992; Lucki 1997), it wasfound that some estrogenic compounds were able to facilitatethe antidepressant-like action of various ADs. In this case, asuboptimal dose of 17 β-estradiol (E2), ethynil-estradiol, anddiethyl-stilbestrol synergize with subthreshold doses of twotypes of ADs: the selective serotonin reuptake inhibitor(SSRI), fluoxetine (FLX), and the noradrenaline reuptakeinhibitor (NaRI), desipramine (DMI) (Estrada-Camarena etal. 2004). The mechanisms that underlie the action of theseestrogens are still unknown, but most likely involve changesin the serotonergic—(Estrada-Camarena et al. 2006a) andnoradrenergic—(López-Rubalcava et al. 2007; Vega-Riveraet al., unpublished observations) transmission in addition toestrogen receptors (Estrada-Camarena et al. 2006b). Indeed,there are several reports indicating that estrogens inducechanges in serotonergic targets like the serotonin transporter(SERT) or 5-HT receptors such as the 5-HT1A, 5-HT1B, and5-HT2 as well as on the synthesis and release of 5-HT (forreview Amin et al. 2005; Bethea et al. 1998; Mize and Alper2000). Noradrenergic neurotransmission is also modulatedby estrogens at several levels, for example, noradrenaline(NA) synthesis is decreased by OVX (Currant-Rauhut andPetersen 2003), whereas the expression and sensitivity of α2-and β-receptors change depending on estrogen levels(Ansonoff and Etgen 2000, 2001; Etgen et al. 2001).
Estrogens synergy with antidepressants in clinics andanimal models, modulate various neurotransmitter systemswhich are target of antidepressants like monoamines,glutamate, and GABA, among others, and affect signalingpathways and transcription factors (for review Bethea et al.1998; McEwen 2001; Scharfman and MacLusky 2005,2006) that are activated by antidepressants (Berton andNestler 2006; Sanacora et al. 2008; Schmidt and Duman2007). Since estrogens produce some of these actions witha shorter latency than antidepressants (Carrasco et al. 2004;Mize and Alper 2000, 2002), it is feasible to propose thatestrogens could shorten their onset of action.
Therefore, the aim of the present work was to evaluate ifestrogens facilitate the antidepressant-like action of venlafax-ine (VLX), FLX, and DMI by reducing their onset of actionand/or by enhancing their antidepressant-like activity in theFST. These antidepressants were selected on the bases of theirmechanism of action, FLX as a SSRI, DMI as a NaRI, andVLX as a mixed 5-HT/NA reuptake inhibitor. In addition,VLX has been proposed to have a shorter latency of actionand is a useful drug for the treatment of clinical depression inwomen (Carrol 2006; Dias et al. 2006).
Material and methods
Animals
Ovariectomized female Wistar rats (250–300 g, 3 monthsold), bred in the animal facilities of the “National Instituteof Psychiatry ‘Ramón de la Fuente’ (Mexico City, Mexico),were housed in groups of six in polycarbonate cages andmaintained on a 12 h light–dark cycle (lights on at 22:00hours) in a temperature-controlled (22°C) room with freeaccess to food and water. The rats were handled 3–5 daysprior to behavioral testing. All experimental procedureswere performed in accordance with the Mexican officialnorm for animal care and handling (NOM-062-ZOO-1999)and approved by the Institutional Ethics Committee of theCINVESTAV-IPN and National Institute of Psychiatry‘Ramón de la Fuente’
Surgical procedures
Ovariectomy was performed 3 weeks before the behavioraltests following the procedures described in Estrada-Camarenaet al. (2003, 2004).
Forced swimming test
The FST was conducted as previously described (Detke etal. 1995, 1997; Estrada-Camarena et al. 2003, 2004) withthe exception that the two swimming sessions wereseparated by different intervals: an initial 15-min pretestfollowed 1, 3, 7 or 14 days later by a 5-min test. Thisexperimental design was chosen because it has been shownthat the pretest-induced immobility behavior remains atleast for 14 days (Detke et al. 1997). Test sessions were runbetween 15:00 and 16:00 hours and videotaped for laterscoring. The behaviors scored were immobility, swimming,and climbing according to pervious reports (Detke et al.1995, 1997; Estrada Camarena et al. 2003, 2004).
Open-field test
In order to evaluate nonspecific actions of the drug treatmentson general activity, six to seven rats of each group were testedin the open-field test following the same procedure describedelsewhere (Estrada-Camarena et al. 2003, 2004).
Drugs
The estrogenic compound 17β-estradiol (E2, SIGMA-Aldrich, St Louis, MO., EUA) was dissolved in corn oiland injected s.c. (0.2 ml/rat). Fluoxetine hydrochloride(FLX, Eli-Lilly, México), desipramine hydrochloride (DMI,SIGMA-Aldrich, St Louis, MO., EUA), and venlafaxine
Psychopharmacology
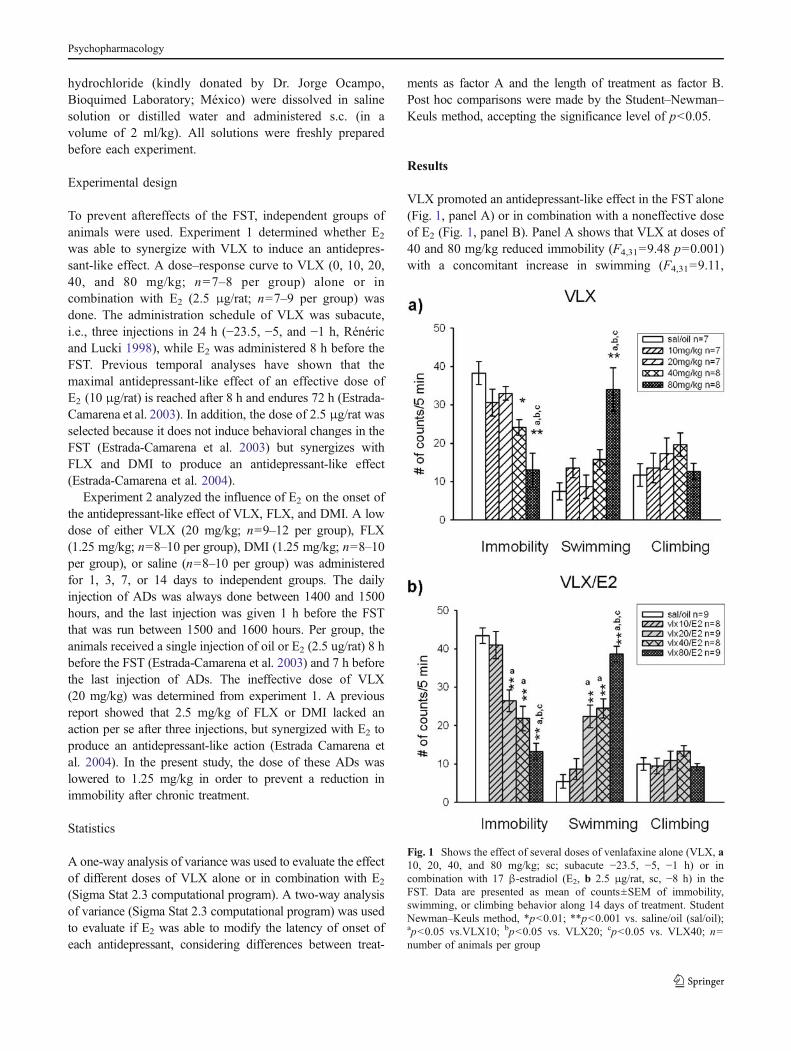
hydrochloride (kindly donated by Dr. Jorge Ocampo,Bioquimed Laboratory; México) were dissolved in salinesolution or distilled water and administered s.c. (in avolume of 2 ml/kg). All solutions were freshly preparedbefore each experiment.
Experimental design
To prevent aftereffects of the FST, independent groups ofanimals were used. Experiment 1 determined whether E2
was able to synergize with VLX to induce an antidepres-sant-like effect. A dose–response curve to VLX (0, 10, 20,40, and 80 mg/kg; n=7–8 per group) alone or incombination with E2 (2.5 μg/rat; n=7–9 per group) wasdone. The administration schedule of VLX was subacute,i.e., three injections in 24 h (−23.5, −5, and −1 h, Rénéricand Lucki 1998), while E2 was administered 8 h before theFST. Previous temporal analyses have shown that themaximal antidepressant-like effect of an effective dose ofE2 (10 μg/rat) is reached after 8 h and endures 72 h (Estrada-Camarena et al. 2003). In addition, the dose of 2.5 μg/rat wasselected because it does not induce behavioral changes in theFST (Estrada-Camarena et al. 2003) but synergizes withFLX and DMI to produce an antidepressant-like effect(Estrada-Camarena et al. 2004).
Experiment 2 analyzed the influence of E2 on the onset ofthe antidepressant-like effect of VLX, FLX, and DMI. A lowdose of either VLX (20 mg/kg; n=9–12 per group), FLX(1.25 mg/kg; n=8–10 per group), DMI (1.25 mg/kg; n=8–10per group), or saline (n=8–10 per group) was administeredfor 1, 3, 7, or 14 days to independent groups. The dailyinjection of ADs was always done between 1400 and 1500hours, and the last injection was given 1 h before the FSTthat was run between 1500 and 1600 hours. Per group, theanimals received a single injection of oil or E2 (2.5 ug/rat) 8 hbefore the FST (Estrada-Camarena et al. 2003) and 7 h beforethe last injection of ADs. The ineffective dose of VLX(20 mg/kg) was determined from experiment 1. A previousreport showed that 2.5 mg/kg of FLX or DMI lacked anaction per se after three injections, but synergized with E2 toproduce an antidepressant-like action (Estrada Camarena etal. 2004). In the present study, the dose of these ADs waslowered to 1.25 mg/kg in order to prevent a reduction inimmobility after chronic treatment.
Statistics
A one-way analysis of variance was used to evaluate the effectof different doses of VLX alone or in combination with E2(Sigma Stat 2.3 computational program). A two-way analysisof variance (Sigma Stat 2.3 computational program) was usedto evaluate if E2 was able to modify the latency of onset ofeach antidepressant, considering differences between treat-
ments as factor A and the length of treatment as factor B.Post hoc comparisons were made by the Student–Newman–Keuls method, accepting the significance level of p<0.05.
Results
VLX promoted an antidepressant-like effect in the FST alone(Fig. 1, panel A) or in combination with a noneffective doseof E2 (Fig. 1, panel B). Panel A shows that VLX at doses of40 and 80 mg/kg reduced immobility (F4,31=9.48 p=0.001)with a concomitant increase in swimming (F4,31=9.11,
Fig. 1 Shows the effect of several doses of venlafaxine alone (VLX, a10, 20, 40, and 80 mg/kg; sc; subacute −23.5, −5, −1 h) or incombination with 17 β-estradiol (E2, b 2.5 μg/rat, sc, −8 h) in theFST. Data are presented as mean of counts±SEM of immobility,swimming, or climbing behavior along 14 days of treatment. StudentNewman–Keuls method, *p<0.01; **p<0.001 vs. saline/oil (sal/oil);ap<0.05 vs.VLX10; bp<0.05 vs. VLX20; cp<0.05 vs. VLX40; n=number of animals per group
Psychopharmacology

p=0.001) without changes in climbing (F4,31=1.15, ns).Interestingly, when a noneffective dose of E2 was added, 20mg/kg of VLX—which lacked an effect per se—produced areduction in immobility (F4,38=20.89, p=0.001) and anenhancement in swimming (F4,38=27.87, p=0.001). Thehigher doses of VLX retain their similar antidepressant-likeaction even if combined with E2. The different combinationsof VLX plus E2 did not induce changes in climbing(F4,38=0.74, ns).
Remarkably, E2 reduced the latency to decreased immo-bility for all three antidepressants (Figs. 2, 3, and 4, panel A).As can be observed in Fig. 2, VLX alone at 20 mg/kg did
not reduce immobility even after 14 days of treatment.Interestingly, the addition of E2 decreased immobility after 1or 3 days from the beginning of treatment (p=0.01);however, after 14 days of VLX treatment, E2 produced theopposite effect. Thus, the two-way analysis of variance(ANOVA) test showed differences in the immobilitydependent of treatment (F3,155=2.84, p=0.040), of timing(F3,155=6.95, p=0.001), and in the interaction between thesefactors (F9,155=2.14, p=0.02). In relation to swimming(Fig. 2, panel B), an increase was observed after 1 and 3days from the beginning of treatment with VLX/E2 (p=0.01), while after 14 days, a drastic decrease in swimmingwas observed. The two-way ANOVA test showed that thiseffect is dependent of treatment (F3,155=7.26, p=0.001), of
Fig. 2 Shows the effect of venlafaxine (VLX; 20 mg/kg, sc, once/day) alone or in combination with 17 β-estradiol (E2; 2.5 μg/rat, sc,−8 h) along time in the FST. Data are presented as mean of counts±SEM of immobility (a), swimming (b), or climbing (c) behaviorregistered every 5 s during a 5-min test period. Student Newman–Keuls method, *p<0.01 vs. 1 day; ap<0.01 vs. saline/oil (sal/oil); bp<0.01 vs. sal/E2;
cp<0.01 vs. VLX/oil; n=number of animals per group
Fig. 3 Shows the effect of fluoxetine (FLX; 1.25 mg/kg, sc, once/day) alone or in combination with 17 β-estradiol (E2; 2.5 μg/rat, sc,−8 h) along time in the FST. Data are presented as mean of counts±SEM of immobility (a), swimming (b), or climbing (c) behaviorregistered every 5 s during a 5-min test period. Student Newman–Keuls method, *p<0.01 vs. 1 day; ap<0.01 vs. saline/oil (sal/oil); bp<0.01 vs. sal/E2;
cp<0.01 vs. FLX/oil; n=number of animals per group
Psychopharmacology

treatment duration (F3,155=2.68, p=0.04), and their interac-tion (F9,155=2.39, p=0.01). Finally, differences in climbing(Fig. 2, panel C), dependent on treatment (F3,155=13.50, p=0.001), duration of treatment (F3,155=5.30, p=0.002), andtheir interaction (F9,155=2.05, p=0.03) were also observed.As it can be seen, the lowest value of climbing was observedin the VLX/oil group at 3 and 7 days after treatment (p=0.01), whereas the highest value corresponds to the groupthat was treated with saline/E2 (p=0.05). VLX/E2 did notshow differences along time.
Interestingly, E2 drastically reduced the time to dimin-ished immobility for FLX (Fig. 3, panel A). Thus, the two-way ANOVA test denoted differences in immobility that aredependent of treatment (F3,127=8.48, p=0.001), of treat-ment duration (F3,127=4.76, p=0.004), and an interaction
between these factors (F9,127=2.25, p=0.02). As it can beseen in panel A, FLX (1.25 mg/kg) alone did not reduceimmobility before 14 days of treatment. In contrast, theaddition of E2 promoted an anti-immobility effect of FLXalready, 3 days after the beginning of treatment. Comparedwith the respective controls, such reduction remained after7 and 14 days of treatment. E2 promoted the increase inswimming classically produced by FLX in the FST (two-way ANOVA values, for treatment, F3,127=9.02, p=0.001;for timing, F3.127=1.21, ns, and for interaction betweenthese factors, F9,127=2.33, p=0.01). The control groupdenoted the lowest level of climbing after 3 days oftreatment, while the FLX/oil group showed an increase ofclimbing after 14 days of treatment (Fig. 3, panel C, two-way ANOVA, treatment, F3,127=1.09, ns, timing F3,127=7.62, p=0.001, and the interaction F9,127=2.52, p=0.01).
Figure 4 shows the effect of adding E2 to the treatmentwith DMI in the FST. Clearly, E2 also drasticallyshortened the time to the reduction in immobility forDMI (Fig. 4, panel A). Thus, the two-way ANOVA testshowed a reduction of immobility dependent of thetreatment (F3,136=29.0, p=0.001) and of the duration ofthe treatment (F3,136=3.6, p=0.01) without significantvalues in the interaction (F9,136=1.24, ns). DMI alonedecreased the immobility after 14 days (p=0.01). As canbe observed in panel A, when E2 was added, the reductionof immobility was evident even after the first injection of
Fig. 4 Shows the effect of desipramine (DMI;1.25 mg/kg, sc, once/day) alone or in combination with 17 β-estradiol (E2; 2.5 μg/rat, sc,−8 h) along time in the FST. Data are presented as mean of counts±SEM of immobility (a), swimming (b), or climbing (c) behaviorregistered every 5 s during a 5-min test period. Student Newman–Keuls method, *p<0.01 vs. 1 day; ap<0.01 vs. saline/oil (sal/oil); bp<0.01 vs. sal/E2;
cp<0.01 vs. DMI/oil; n=number of animals per group
Fig. 5 Shows the effect of venlafaxine (VLX; 20 mg/kg, sc, once/day), fluoxetine (FLX; 1.25 mg/kg, sc, once/day), or desipramine(DMI;1.25 mg/kg, sc, once/day) in combination with 17 β-estradiol(E2; 2.5 μg/rat, sc, −8 h) along time in the FST. The dashed areacorresponds to the immobility values shown by all animals treatedwith saline/oil Data are presented as mean of counts±SEM ofimmobility behavior registered every 5 s during a 5-min test period.Student Newman–Keuls method #p<0.05 vs FLX/E2;
&p<0.001 vsVLX/E2
Psychopharmacology

DMI (p=0.009). The anti-immobility action of DMI plusE2 reached its maximal effect after 3 days of treatmentwith DMI and remained without changes after 7 or 14 days oftreatment, compared with the control groups (p=0.01). Asmentioned before, the reduction in immobility was accom-panied by increases in climbing (Fig. 4, panel C). The two-way ANOVA showed differences due to the treatment(F3,136=36.41, p=0.001) and due to the timing (F3,136=5.013, p=0.003) without significant values for the interac-tion (F9,136=1.69, ns). No change in swimming was inducedby these treatments. Two-way ANOVA values for swimmingwere: for treatment (F3,136=1.28, ns); for timing (F3,136=0.84, ns); and for interaction (F9,136=0.86, ns).
Figure 5 compares the effect of E2 combined with eachantidepressant on the immobility behavior in the FST. Thedashed area corresponds to the immobility values shown byall animals treated with saline/oil. This figure is composedof data derived from Figs. 2, 3, and 4. The combinationsDMI/E2 and FLX/E2 were more effective to reduce theimmobility behavior than VLX/E2 after 3 days; such aneffect remains until 14 days of treatment. Interestingly, theDMI/E2 combination was more effective to induce areduction of immobility behavior than FLX/E2 at 1 and14 days (p<0.05). Two-way ANOVA values for immobilitybehavior were: for treatment F2,100=47.23, p=0.001; fortiming F3,100=5.86, p=0.001 and for the interactionbetween factors F6,100=8.40, p=0.001.
The results of the locomotor activity test are shown inTable 1. As it can be seen, the control group showedrelatively similar values of activity along the first 7 days oftesting; however, a reduced locomotion was observed at 14days. E2/saline decreased the number of squares crossed at3, 7, and 14 days compared with 1 day after treatment andthe control group (p=0.05). The FLX/oil group showed an
increase in locomotor activity after 14 days of treatment,while VLX/oil and DMI/oil groups did not show changes inlocomotion along time.
The combination of FLX or DMI with E2 reducedambulation (p=0.05) after 3, 7, or 14 days of treatment,whereas the combination with VLX remained withoutsignificant changes compared with their respective controlgroups. The two-way ANOVA values were: F7,176=5.8, p=0.001 for treatment; F3,176=6.17; p=0.001 for timing; andF21,176=3.76; p=0.001 for the interaction.
Discussion
The main findings of the present study are: E2 synergizeswith DMI, FLX, and VLX in the FST to shorten theirlatency of action, and similarly to DMI and FLX (Estrada-Camarena et al. 2004), a subacute treatment with VLXreduced immobility behavior, an action that was potentiatedby E2.
The present experiments show a clear reduction inimmobility (reflecting antidepressant-like actions) 3 daysafter a low dose of FLX, DMI, or VLX when combined witha low dose of E2 (2.5 μg/rat). It is worth noting that none ofthe ADs nor E2 at these doses and intervals produced aneffect per se. Higher doses of E2 (10 μg/rat) produceantidepressant-like actions in the FST evidenced as adecreased immobility and an increased swimming (Rachmanet al. 1998; Estrada-Camarena et al. 2003; Galea et al. 2001;Walf and Frye 2005), thus it can be proposed that thesynergism observed between E2 and the ADs is theconsequence of the estrogen’s effect rather than that ofthe ADs. Against this idea, however, it has been found thatafter a 5-day treatment, 2.5 μg/rat of E2 fails to induce an
Table 1 Effect of E2 in combination with FLX, DMI, or VLX on locomotor activity test
Treatments Days of treatment
1 3 7 14
Sal/oil 57.0±4.4 n=10 66.7±3.8 n=10 70±3.4 n=7 43.7±4.24a,b,c n=10Sal/E2 70.5±6.6 n=6 40.6±6.9*,a n=6 30.8±4.7*,a n=6 46.6±2.3a n=6FLX/oil 58.3±5.4 n=6 40±4.5*,a,c n=6 64.5±6.0 n=6 71.8±6.9*,** n=6DMI/oil 66.1±12 n=6 44.3±5.2*,a n=6 58.0±3.3 n=6 56.6±8.0 n=6VLX/oil 56.5±2.7 n=6 51.3±6.4 n=6 55.1±7.0 n=6 55.0±7.3 n=6FLX/E2 65.0±6.3 n=7 40.0±4.5*a n=6 41.8±6.1*,a,*** n=6 46.2±8.0a,*** n=6DMI/E2 50.8±5.1 n=7 44.3±5.2* n=6 33.8±3.2*,*** n=6 35.8±3.4 n=6VLX/E2 55.1±4.2 n=6 60.3±4.9 n=6 64.8±5.8** n=6 49.1±7.5 n=6
Data are shown as mean±SEM of the number of squares crossed in 5 min of locomotor activity test.*p=0.05 vs. saline/oil (sal/oil)** p=0.05 vs. sal/E2
***p=0.05 vs. antidepressant/oila,b,c p=0.05 vs. 1, 3, or 7 days of administration of antidepressant drug. 17 β-estradiol (E2; 2.5 μg/rat, s.c.) was applied 8 h before FST whilefluoxetine (FLX; 1.25 mg/kg, s.c.), desipramine (DMI; 1.25 mg/kg, s.c.), or venlafaxine (VLX; 20 mg/kg, s.c) were administered once/day
Psychopharmacology

antidepressant-like effect (immobility counts: OVX-oil=31.2±1.39 vs OVX-E2=37.7±2.0; Student t test, NS).Additionally, if the ADs would be facilitating E2’s action, itcould be expected that their combination at any time wouldpromote the antidepressant-like effect. Interestingly, this wasnot the case since exclusively at day 3 of treatment (but notat days 1, 7, or 14), the combination of each AD with E2
produced the same decrease in immobility with, however,important differences in the expression of active behaviors(see below). From these results, it seems clear that theantidepressant is not promoting the antidepressant-like actionof the estrogen.
In the FST, the main effect of ADs consists in reducingthe immobility behavior (Detke et al. 1995, 1997; Lucki1997; Cryan et al. 2005a, b); such reduction is accompaniedby increases in the active behaviors. Interestingly, anincreased climbing indicates drug actions on the catechol-aminergic systems while increased swimming seems todenote the activation of the serotonergic transmission. Inagreement with previous reports (Detke et al. 1997; Cryanet al. 2005a), the present study showed that E2 preserves thebehavioral profile of DMI and FLX (Estrada Camarena etal. 2004), further stressing that estradiol is promoting theaction of the AD. Accordingly, VLX, a mix serotonergic/noradrenergic antidepressant, should promote a reduction inimmobility with a concomitant increase in both swimmingand climbing. In line, Rénéric and Lucki 1998; Rénéric etal. 2002 showed in male rats that VLX produced anincrease in swimming at doses varying from 20–40 mg/kgwhile at the highest dose (80 mg/kg) also climbing wasincreased. These data agree with a higher effectiveness ofVLX to inhibit 5-HT than NA reuptake (Béïque et al. 1998,2000; Owens et al. 1997). In the present study, no dose ofVLX alone or in combination with estradiol produced anincrease in climbing, suggesting that in ovx females thisantidepressant produces a behavioral profile reflectingprimarily a serotonergic action. From these data, it couldbe proposed that females are more sensitive than males tothe serotonergic actions of this compound. Future studiesare required to analyze this proposition.
The present report does not offer experimental evidenceto establish a clear mechanism of action for the effect ofestrogens on the latency of onset of ADs. Severalpossibilities may be proposed including neurotransmitterreceptors, signaling pathways, and/or neurogenesis whichare similarly modified by AD and estrogens. Interestingly,most of the estrogens’ actions appear with shorter latenciesthan those required for ADs (Alves et al. 2002; Banasr et al.2001 Berton and Nestler 2006; Brake et al. 2001; Carrascoet al. 2004; Galea 2008; Jackson and Uphouse 1998;Mize and Alper 2000; Ormerod et al. 2003; Santarelli et al.2003; Schmidt and Duman 2007). Based on present andprevious reports, mechanisms involving 5-HT and NA
transmission may be proposed (Estrada-Camarena et al.2006a, b; López-Rubalcava et al. 2007). In this regard, ithas been reported that an acute low dose of estrogensinhibits SERT activity similarly to SSRIs (Michel et al.1987; Mendelson et al. 1993). Additionally, the destructionof serotoninergic fibers only partially cancels the action ofE2 in the FST (López-Rubalcava et al. 2005), suggestingthat postsynaptic components also participate in theantidepressant and/or facilitatory actions of E2. Interesting-ly, ADs and estrogens both promote the desensitization ofthe 5-HT1A receptor, one of various processes that mayunderlie the latency of ADs action (for review Duman1999). However, estrogens desensitize this receptor inseveral hours, whereas ADs require at least 14 days(Carrasco et al. 2004; Li et al. 1996; Jackson and Uphouse1998; Mize and Alper 2000; Mize et al. 2001; Raap et al.2000). Additionally, for the desensitization of 5-HT1A
induced by E2, activation of the ER plus PKA, PKC, andRGZ proteins is required (Mize and Alper 2002; Carrascoet al. 2004). It is important to mention that at least PKA andPKC are target for antidepressants (Berton and Nestler2006).
Estrogenic compounds are able to modulate the activityof noradrenergic receptors (Ansonoff and Etgen 2000,2001) and noradrenaline transporter (Ghraf et al. 1983).Idazoxan, an α2- antagonist, blocks the antidepressant-likeeffect of ethynil-estradiol alone or combined with DMI inthe FST (López-Rubalcava et al. 2007; Vega Rivera,unpublished observations). In addition, in vitro studieshave shown that estradiol promotes noradrenaline release inthe hypothalamus by stabilizing α2 adrenoceptor phosphor-ylation (Ansonoff and Etgen 2001) and induces desensiti-zation of β-adrenoceptors (Ansonoff and Etgen 2000;Karkanias and Etgen 1993). Changes in the sensitivity ofthese receptors are required to observe an antidepressanteffect (for review, see Duman 1999; Schmidt and Duman2007). Thus, it is possible that E2 induces a rapid change inthe sensitivity of these receptors in order to shorten theaction of DMI.
As previously reported for DMI and FLX (Estrada-Camarena et al. 2004), three injections of VLX produced areduction in immobility that was potentiated by E2. Theseresults are in agreement with those previously reported byNowakowska and Kus (2005). Accordingly, the addition ofa low dose of E2 also shortened the latency for the VLXeffect. Contrary to expected, the addition of E2 to VLXafter 7 or 14 days increased, rather than decreased, theimmobility behavior. Commonly, after chronic treatment (atleast for 14 days) with most ADs, brain neurochemistrychanges in order to promote clearer antidepressant-likeeffects. VLX seems to be an exception, since it lacks actionin the FST and in other animal models of depression evenafter prolonged treatments (Connor et al. 2000; Gur et al.
Psychopharmacology

1999). The neuropharmacological mechanism proposed forVLX action involves an initial blockade of 5-HT and NAuptake in a dose-dependent manner (Bëique et al. 1998,2000; Owens et al. 1997), followed by changes in thesensitivity of monoaminergic receptors (Bëique et al. 1998;Rossi et al. 2006; Weikop et al. 2004) and transcriptionfactors (Gur et al. 1999; Larsen et al. 2008; Xu et al. 2003).Interestingly, Gur et al. (1999) found that an acute treatmentwith VLX—at a dose that enhances 5-HT levels in thehippocampus—is no longer effective after 4 weeks oftreatment (Connor et al. 2000; Gur et al. 1999; Rossi et al.2006). Moreover, the neuroprotective action of VLX in thehippocampus after chronic treatment is evident with a low(5 mg/kg) dose but not with a high (10 mg/kg) dose (Xu et al.2003). These data suggest a dose adjustment during chronictreatment with VLX alone or in combination with E2.
Several reports indicate that chronic treatment withestrogens (Holschneider et al. 1998; Ma et al. 1995) ortheir use at very high doses (Somani and Khurana 1973)affect the metabolism of antidepressants by interacting withthe monoamino-oxidase (Holschneider et al. 1998; Ma et al.1995) or the cytochrome P450 system (Wang and Strobel1997). Therefore, it could be proposed that the synergisticaction of E2 involves changes in the metabolism ofantidepressants. This mechanism is unlikely since theadministration of E2 at the dose used in the present study(2.5 µg/kg/rat) is low to produce this effect. On the otherhand, both E2 and ADs alter the levels of pregnaneneurosteroids by affecting enzymes involved in the synthe-sis of allopregnanolone (Griffin and Mellon 1999; Soma etal. 2005), a steroid associated with the relief of depressivesymptoms during the premenstrual syndrome and theperimenopausal period (for review Eser et al. 2006; Marxet al. 2006; Uzunova et al. 2006). Thus, it is possible thatone of the mechanisms involved in the synergistic action ofE2 in conjunction with ADs is related to an increased levelof neuroactive progestins.
It is well-known that a nonspecific action of someantidepressants consists in reducing locomotor activity(Borsini and Meli 1988). It is important to mention that adrug treatment that decreases immobility and increasesgeneral activity is interpreted as a false positive in the FST(Borsini and Meli 1988; Cryan et al. 2005a, b; Porsolt et al.1978), as was the case after 14 days of FLX alone.Inversely, reduced locomotion is not associated withdecreased immobility (Porsolt et al. 1978). The presentresults showed a reduction in both immobility andlocomotion after the combination of E2 plus the antide-pressants, ruling out the possibility that motor effects mightmediate the antidepressant-like behaviors observed.
Important limitations should be addressed from presentresults: first, the fact that estradiol levels determination aftera single injection of 2.5 μg/rat is missing; and second,
further experimental evidence is needed to support anintegrative mechanism of action. However on the basis ofpresent data, it could be proposed that estrogens mayrepresent a therapeutic alternative for shortening the latencyof action of ADs.
In conclusion, a low dose E2 shortened FLX and DMI inthe FST, representing a putative therapeutic strategy.
Acknowledgements The current study has been carried out withfinancial support from CONACyT. EE-C thanks Eder Gómez andSergio Márquez for animal care and technical assistance. Authorsthank Dr. Bryan Phillips for language checking. All experimentalprocedures were performed in accordance with the Mexican officialnorm for animal care and handling (NOM-062-ZOO-1999) andapproved by the Institutional Ethics Committee of the CINVESTAV-IPN and National Institute of Psychiatry ‘Ramón de la Fuente’. Thecurrent study has been carried out with financial support fromCONACyT (to A.F.-G., F1 61187).
References
Alves SE, Hoskin E, Lee SJ, Brake WG, Ferguson D, Luine V, AllenPB, Greengard P, McEwen BS (2002) Serotonin mediates CA1spine density but is not crucial for ovarian steroid regulation ofsynaptic plasticity in the adult rat dorsal hippocampus. Synapse45:143–151
Amin Z, Camli T, Epperson NC (2005) Effect of estrogen–serotonininteractions on mood and cognition. Behav & Cog Neurosci Rev4:43–58
Amsterdam J, García-España F, Fawett J, Quitkin F, Reimherr F,Rosenbaum J, Beasley C (1999) Fluoxetine efficacy in meno-pausal women with and without estrogen replacement. J AffecDisord 55:11–17
Ansonoff MA, Etgen AM (2000) Evidence that oestradiol attenuatesbeta-adrenoceptor function in the hypothalamus of female rats byaltering receptor phosphorylation and sequestration. J Neuro-endocrinol 12:1060–1066
Ansonoff MA, Etgen AM (2001) Receptor phosphorylation mediatesestradiol reduction of alpha2-adrenoceptor coupling to G proteinin the hypothalamus of female rats. Endocrine 14:165–174
Artigas F, Pérez V, Alvarez E (1994) Pindolol induces a rapidimprovement of depressed patients treated with serotonin reup-take inhibitors. Arch Gen Psychiatr 51:248–251
Austin MP, Souza FG, Goodwin GM (1991) Lithium augmentation inantidepressant-resistant patients. A quantitative analysis. Br JPsychiatr 159:510–514
Banasr M, Hery M, Brezun JM, Daszuta A (2001) Serotonin mediatesoestrogen stimulation of cell proliferation in the adult dentategyrus. Eur J Pharmacol 14:1417–1424
Bauer M, Döpfmer S (1999) Lithium augmentation in treatment-resistant depression: meta-analysis of placebo controlled studies.J Clin Psychopharmacol 19:427–434
Bëique J, de Montigny C, Blier P, Debonnel G (1998) Blockade of 5-hydroxytryptamine and noradrenaline uptake by venlafaxine: acomparative study with paroxetine and desipramine. Br JPharmacol 125:526–532
Bëique J, de Montigny C, Blier P, Debonnel G (2000) Effects ofsustained administration of serotonin and norepinephrine reup-take inhibitor venlafaxine: I. In vivo electrophysiological studiesin the rat. Neuropharmacology 39:1800–1812
Psychopharmacology

Berton O, Nestler EJ (2006) New approaches to antidepressant drugdiscovery: beyond monoamines. Nat Rev Neurosci 7:137–151
Bethea CL, Pecins-Thompson M, Schutzer W, Gundlah C, Lu Z(1998) Ovarian steroids and serotonin neural function. MolNeurobiol 18:87–122
Blier P, Bergeron R (1995) Effectiveness of pindolol with selectedanti-depressant drugs in the treatment of major depression. J ClinPsychopharmacol 15:217–222
Borsini F, Meli A (1988) Is the forced swimming test a suitable modelfor revealing antidepressant activity? Psychopharmacology (Berl)94:127–160
Brake W, Alves S, Dunlop J, Lee S, Bulloch K, Allen P, Greendard P,McEwen B (2001) Novel target sites for estrogen action in thedorsal hippocampus: an examination of synaptic proteins.Endocrinology 142:1284–1289
Carrasco GA, Barker SA, Zhang Y, Damjanoska KJ, Sullivan N,Garcia F, D’Souza DN, Muma N, Van de Kar LD (2004)Estrogen treatment increases the levels of regulator of G proteinsignaling-Z1 in the hypothalamic paraventricular nucleus: possi-ble role in desensitization of 5-Hydroxytryptamine1A receptors.Neuroscience 127:261–267
Carrol D (2006) Non-hormonal therapies for hot flashes in meno-pause. Am Fam Physician 73:457–464
Connor TJ, Kelliher P, Shen Y, Harkin A, Kelly JP, Leonard BE(2000) Effect of sub-chronic antidepressant treatments onbehavioral, neurochemical, and endocrine changes in theforced-swim test. Pharmacol Biochem Behav 65:591–597
Cryan JF, Page M, Lucki I (2005a) Differential behavioral effects ofthe antidepressants reboxetine, fluoxetine and moclobemide in amodified forced swim test following chronic treatment. Psycho-pharmacology 182:335–344
Cryan JF, Valentino R, Lucki I (2005b) Assessing substrates underlyingthe behavioral effects of antidepressants using the modified ratforced swimming test. Neurosci Biobehav Rev 29:547–569
Curran-Rauhut MA, Petersen SL (2003) Oestradiol-dependent and -independent modulation of tyrosine hydroxylase mRNA levels insubpopulations of A1 and A2 neurones with oestrogen receptor(ER)alpha and ER beta gene expression. J Neuroendocrinol15:296–303
Detke M, Rickels M, Lucki I (1995) Active behaviors in the rat forcedswimming test differentially activated by serotonergic andnoradrenergic antidepressants. Psychopharmacology 121:66–72
Detke M, Johnson J, Lucki I (1997) Acute and chronic antidepressantdrug treatment in the rat forced swimming test model ofdepression. Exp Clin Psychopharmacol 5:107–112
Dias R, Kerr-Correa F, Moreno R, Trinca LA, Pontes A, Halbe HW,Gianfaldoni A, Dalben I (2006) Efficacy of hormone therapywith and without methyltestosterone augmentation of venlafaxinein the treatment of postmenopausal depression: a double-blindcontrolled pilot study. Menopause 13:202–211
Duman R (1999) The neurochemistry of mood disorders: preclinicalstudies. In: Charney D, Nestler E, Bunney B (eds) Neurobiology ofmental illness,. 1st edn. Oxford University Press, Oxford, pp 333–347
Eser E, Romeo E, Baghai TC, DiMichele F, Schüle C, Pasini A, ZwanzgerP, Padberg F, Rupprecht R (2006) Neuroactive steroids as modulatorsof depression and anxiety. Neuroscience 138:1041–1048
Estrada-Camarena E, Fernandez-Guasti A, Lopez-Rubalcava L (2003)Antidepressant-like effect of different estrogenic compounds in theforced swimming test. Neuropsychopharmacology 28:830–838
Estrada-Camarena E, Fernández-Guasti A, López-Rubalcava C (2004)Interaction between estrogens and antidepressants in the FST inrats. Psychopharmacology 173:139–145
Estrada-Camarena E, Fernández Guasti A, López-Rubalcava C(2006a) Participation of the 5-HT1A receptors in the antidepres-sant-like action of estrogens in the forced swimming test.Neuropsychopharmacology 31:247–255
Estrada-Camarena E, López-Rubalcava C, Fernández-Guasti A(2006b) Facilitating antidepressant-like actions of estrogens aremediated by 5-HT1A and estrogen receptors in the forcedswimming test. Psychoneuroendocrinology 31:905–14
Etgen AM, Ansonoff MA, Quesada A (2001) Mechanisms of ovariansteroid regulation of norepinephrine receptor-mediated signaltransduction in the hypothalamus: implications for femalereproductive physiology. Horm Behav 40:169–177
Galea LA (2008) Gonadal hormone modulation of neurogenesis in thedentate gyrus of adult male and female rodents. Brain ResReviews 57:332–341
Galea L, Wide J, Barr A (2001) Estradiol alleviates depressive-likesymptoms in a novel animal model of post-partum depression.Behav Brain Res 122:1–9
Genazzani A, Spinetti A, Gallo R, Bernardi F (1999) Menopause and thecentral nervous system: intervention options. Maturitas 31:103–110
Ghraf R, Michel M, Hiemke C, Knuppen R (1983) Competition bymonophenolic estrogens and catecholestrogens for high-affinityuptake of [3H] (-)-norepinephrine into synaptosomes from ratcerebral cortex and hypothalamus. Brain Res 277:163–168
Griffin LD,Mellon S (1999) Selective serotonin reuptake inhibitors directlyalter activity of neurosteroidogenic enzymes. PNAS 96:13512–13517
Gur E, Dremencov E, Lerer B, Newman ME (1999) Venlafaxine:acute and chronic effects on 5-hydrytryptamine levels in rat brainin vivo. Eur J Pharmacol 372:17–24
Halbreich U, Kahn L (2001) Role of estrogen in the aetiology andtreatment of mood disorders. CNS Drugs 15:797–817
Holschneider D, Kumazawa T, Chen K, Shih J (1998) Tissue specificeffects of estrogen on monoamino oxidase A and B in the rat.Life Sci 63:155–160
Jackson A, Uphouse L (1998) Dose dependent effects of estradiolbenzoate on the 5-HT1A receptor agonist action. Brain Res796:299–302
Karkanias G, Etgen A (1993) Estradiol attenuates α2-adrenoceptor-mediated inhibition of hypothalamic norepinephrine release. JNeurosc 13:3448–3455
Larsen MH, Hay-Schmidt A, Ronn LC, Mikkelsen JD (2008)Temporal expression of brain-derived neurotrophic factor(BDNF) mRNA in the rat hippocampus after treatment withselective and mixed monoaminergic antidepressants. Eur JPharmacol 578:144–122
Li Q,Muma A, Van de Kar L (1996) Chronic fluoxetine induces a gradualdesensitization of 5-HT1A receptors: reductions in hypothalamicandmidbrain Gi and G0 proteins and in neuroendocrine responses to5-HT1A agonist. J Pharmacol Exp Ther 279:1035–1042
López-Rubalcava C, Oikawa-Sala J, Chávez-Álvarez K, Estrada-Camarena E (2005) Analysis of the participation of theserotonergic system in the antidepressant-like action of 17beta -estradiol in the forced swimming test (FST): presynaptic orpostsynaptic actions Program No. 567.12. 2005 Abstract Viewer/Itinerary Planner. Washington, DC: Society for Neuroscience,2005. Online
López-Rubalcava C, Vega Rivera NM, Cruz-Martínez JJ, Estrada-Camarena E (2007) Participation of both estrogen and alpha2-receptors in the antidepressant-like actions of ethynil-estradiol inrats tested in the forced swimming test. 12th Biennial meeting ofthe European Behavioral Pharmacology Society
Lucki I (1997) The forced swimming test as a model for core andcomponent behavioral effects of antidepressant drugs. BehavPharmacol 8:523–532
Lucki I (1998) The spectrum of behaviors influenced by serotonin.Biol Psychiatry 44:151–162
Ma Z-Q, Violani E, Villa F, Picotti G, Maggi A (1995) Estrogeniccontrol of monoamine oxidase A activity in human neuroblasto-ma cells expressing physiological concentrations of estrogenreceptor. Eur J Pharmacol 284:171–176
Psychopharmacology

Marx C, Stevens R, Shampine LJ, Uzunova V, Trost WT, ButterfieldMI, Massing MW, Hamer R, Morrow AL, Lieberman JA (2006)Neuroactive steroids are altered in schizophrenia and bipolardisorder: relevance to pathophysiology and therapeutics. Neuro-psychopharmacology 31:1249–1263
McEwen BS (2001) Genome and hormones: gender differences inphysiology invited review: estrogen effects on the brain: multiplesites and molecular mechanisms. J Appl Physiol 91:2785–2801
Mendelson S, McKittrick C, McEwen B (1993) Autoradiographicanalyses of the effects of estradiol benzoate on [ 3H]paroxetinebinding in the cerebral cortex and dorsal hippocampus ofgonadectomized male and female rats. Brain Res 601:299–302
Michel M, Rother A, Hiemke C, Ghraf R (1987) Inhibition ofsynaptosomal high-affinity uptake of dopamine and serotonin byestrogen agonists and antagonists. BiochemPharmacol 36:3175–3180
Mize A, Alper R (2000) Acute and long-term effects of 17b-estradiolon Gi/0 coupled neurotransmitter receptor function in female ratbrain as assessed by agonist-stimulated [35S] GTPgS binding.Brain Res 859:326–333
Mize A, Alper R (2002) Rapid uncoupling of serotonin-1A receptorsby 17-b estradiol in vitro requires protein kinases A and C.Neuroendocrinology 76:339–347
Mize A, Poisner A, Alper R (2001) Estrogens act in rat hippocampusand frontal cortex to produce rapid, receptor mediated decreasesin serotonin 5-HT1A receptor function. Neuroendocrinology73:166–174
Nowakowska E, Kus K (2005) Antidepressant and memory affectinginfluence of estrogen and venlafaxine in ovariectomized rats.Arzneim-Forsch/Drug Res 55:153–159
Oppenheim G (1983) Estrogen in the treatment of depression:neuropharmacological mechanisms. Biol Psychiatry 18:721–725
Ormerod BK, Falconer EM, Galea LA (2003) Estradiol initiallyenhances but subsequently suppresses (via adrenal steroids)granule cell proliferation in the dentate gyrus of adult femalerats. J Neurobiol 55:247–260
Owens MJ, Morgan WN, Plott SJ, Nemeroff CB (1997) Neurotrans-mitter receptor and transporter binding profile of antidepressantsand their metabolites. J Pharmacol Exp Ther 283:1305–1322
Pérez V, Gilaberte I, Faries D, Alvarez E, Artigas F (1997)Randomised, doble-blind, placebo-controlled trial of pindolol incombination with fluoxetine
Porsolt R, Lenègre A (1992) Behavioural models of depression. In:Elliot J, Heal D, Maisden C (eds) Experimental approaches toanxiety and depression. Wiley, New York, pp 73–85
Porsolt R, Le PichonM, JalfreM (1977) Depression: a new animal modelsensitive to antidepressant treatments. Nature 266:730–732
Porsolt R, Anton G, Blavet N, Jalfre M (1978) Behavioral despair inrats: a new model sensitive to antidepressant treatments. Eur JPharmacol 47:379–391
Prange A (1972) Estrogen may well affect response to antidepressant.JAMA 219:143–144
Raap D, Don Carlos L, Garcia F, Muma N, Wolf W, Battaglia A, Vande Kar L (2000) Estrogen desensitizes 5-HT1A receptors andreduces levels of Gz,Gi1 and Gi3 proteins in the hypothalamus.Neuropharmacology 39:1823–1832
Rachman I, Unnerstall J, Pfaff D, Cohen R (1998) Estrogen alters behaviorand forebrain c-fos expression in ovariectomized rats subjected to theforced swim test. Proc Natl Acad Sci USA 95:13941–13946
Rénèric JP, Lucki I (1998) Antidepressant behavioral effects by dualinhibition of monoamine reuptake in the rat forced swimmingtest. Psychopharmacology 136:190–197
Rénèric JP, Bouvard M, Stinus L (2002) In the rat forced swimmingtest, NA-system mediated interactions may prevent 5-HTproperties of some subacute antidepressant treatments beingexpressed. Eur Neuropsychopharmacol 12:159–171
Robinson G (2001) Psychotic and mood disorders associated with theperimenopausal period. Epidemiology, aetiology and manage-ment. CNS Drugs 15:175–184
Rossi DV, Valdes M, Gould GG, Hensler JG (2006) Chronicadministration of venlafaxine fails to atenúate 5-HT1A receptorfunction at level of receptor-g protein interaction. Int J Neuro-psychopharmacol 9:393–406
Sanacora G, Zarate C, Krystal J, Manji H (2008) Targeting theglutamatergic system to develop novel, improved therapeutics formood disorders. Nat Rev Drugs Dis 7:426–437
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S,WeisstaubN, Lee J, Duman R,Arancio O, Belzung C,Hen R (2003)Requirement of hippocampal neurogenesis for the behavioral effectsof antidepressants. Science 301:805–809 Aug 8
Scharfman HE, MacLusky NJ (2005) Similarities between action ofestrogen and BDNF in the hippocampus: coincidence or clue?Trends Neurosci 28(2):79–85
Scharfman HE, MacLusky NJ (2006) Estrogen and brain-derivedneurotrophic factor (BDNF) in hippocampus: complexity ofsteroid hormone-growth factor interactions in the adult CNS.Front Neuroendocrinol 27(4):415–435
Schmidt HD, Duman RS (2007) The role of neurotrophic factors inadult hippocampal neurogénesis, antidepressant treatments andanimal models of depressive-like behavior. Behav Pharmacol18:391–418
Schneider M, Small G, Hamilton S, Bystrisky A, Nemeroff C, MeyersB (1997) Estrogen replacement and response to fluoxetine insurgically menopausal women. Am J Geriatr Psychiatr 5:97–106
Schneider L, Small G, Clary CM (2001) Estrogen replacement therapyand antidepressant response to sertraline in older depressedwomen. Am J Geriatr Psychiatr 9:393–399
Shapira B, Oppenheim G, Zohar J, Segal M, Malach D, Belmaker R(1985) Lack of efficacy of estrogen supplementation to imipra-mine in resistant female depressives. Biol Psychiatry 20:570–583
Shiah IS, Yatman L, Srisurapanont M, Lam R, Tam E, Zis A (2000)Does the addition of pindolol accelerate the response toelectroconvulsive therapy in patients with major depression? Adouble-blind, placebo-controlled pilot study. J Clin Psychophar-macol 20(3):373–378
SomaKK, Lakhter SA, Schilinger BA,Micevych PE (2005) Neurosteroidsand female reproduction: estrogen increases 3beta-HSD mRNA andactivity in rat hypothalamus. Endocrinology 146:4386–4390
Somani S, Khurana R (1973) Mechanism of estrogen–imipramineinteraction. JAMA 23:560
Stahl S (1998) Augmentation of antidepressants by estrogen.Psychopharmacol Bull 34:319–321
Uzunova V, Sampson L, Uzunov DP (2006) Relevance of endogenous3α-reduced neurosteroids to depression and antidepressantaction. Psychopharmacology 186:3351–361
Walf AA, Frye CA (2005) Antianxiety and antidepressive behaviorproduced by physiological estradiol regimen may be modulatedby hypothalamic–pituitary–adrenal axis activity. Neuropsycho-pharmacology 30:1288–1301
Wang H, Strobel HW (1997) Regulation of CYP3A9 gene expressionby estrogen and catalytic studies using P450 3A9 expressed inEscherichia coli. Arc Biochem Biophys 344:365–372
Weikop P, Kehr J, Scheel-Kruger J (2004) The role of alpha1- andalpha2-adrenoreceptors on venlafaxine-induced elevation ofextracellular serotonin, noradrenaline and dopamine levels inthe rat prefrontal cortex and hippocampus. J Psychopharmacol18:395–403
Xu H, Richardson S, Li XM (2003) Dose-related effects of chronicantidepressants on neuroprotectíve proteins BDNF, Bcl-2 and Cu/Zn-SOD in rat hippocampus. Neuropsychopharmacology 28:53–62
Psychopharmacology
Related Documents











