1 23 Journal of Archaeological Method and Theory ISSN 1072-5369 J Archaeol Method Theory DOI 10.1007/s10816-014-9228-5 Reduced Population Variance in Strontium Isotope Ratios Informs Domesticated Turkey Use at Chaco Canyon, New Mexico, USA Deanna N. Grimstead, Amanda C. Reynolds, Adam M. Hudson, Nancy J. Akins & Julio L. Betancourt

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
Journal of Archaeological Methodand Theory ISSN 1072-5369 J Archaeol Method TheoryDOI 10.1007/s10816-014-9228-5
Reduced Population Variance in StrontiumIsotope Ratios Informs DomesticatedTurkey Use at Chaco Canyon, New Mexico,USA
Deanna N. Grimstead, AmandaC. Reynolds, Adam M. Hudson, NancyJ. Akins & Julio L. Betancourt

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media New York (outside
the USA). This e-offprint is for personal
use only and shall not be self-archived
in electronic repositories. If you wish to
self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Reduced Population Variance in Strontium IsotopeRatios Informs Domesticated Turkey Use at ChacoCanyon, New Mexico, USA
Deanna N. Grimstead & Amanda C. Reynolds &Adam M. Hudson & Nancy J. Akins &Julio L. Betancourt
# Springer Science+Business Media New York (outside the USA) 2014
Abstract Traditionally, strontium isotopes (87Sr/86Sr) have been used as a sourcingtool in numerous archaeological artifact classes. The research presented heredemonstrates that 87Sr/86Srbioapatite ratios also can be used at a population level toinvestigate the presence of domesticated animals and methods of management. Theproposed methodology combines ecology, isotope geochemistry, and behavioralecology to assess the presence and nature of turkey (Meleagris gallopavo)domestication. This case study utilizes 87Sr/86Srbioapatite ratios from teeth and bonesof archaeological turkey, deer (Odocoileus sp.), lagomorph (Lepus sp. and Sylvilagussp.), and prairie dog (Cynomys sp.) from Chaco Canyon, NM, USA (ca. A.D. 800–1250). Wild deer and turkey from the southwestern USA have much larger homeranges and dispersal behaviors (measured in kilometers) when compared to lagomorphsand prairie dogs (measured in meters). Hunted deer and wild turkey fromarchaeological contexts at Chaco Canyon are expected to have a higher variance intheir 87Sr/86Srbioapatite ratios, when compared to small range taxa (lagomorphs and
J Archaeol Method TheoryDOI 10.1007/s10816-014-9228-5
D. N. Grimstead (*)Department of Anthropology, The Ohio State University, 4034 Smith Laboratory, 174 W. 18th Ave.,Columbus, OH 43210-1106, USAe-mail: [email protected]
A. C. ReynoldsConocoPhillips, P.O. Box 2197, Houston, TX 77025-2197, USA
A. M. HudsonDepartment of Geosciences, The University of Arizona, Gould-Simpson Building, 1040 East 4th St.,Tucson, AZ 85721, USA
N. J. AkinsCenter for New Mexico Archaeology, 7 Old Cohitit Rd., Santa Fe, NM 87507, USA
J. L. BetancourtNational Research Program, Water Mission Area, U.S. Geological Survey, 12201 Sunrise Valley, Reston,VA 20192, USA
Author's personal copy

prairie dogs). Contrary to this expectation, 87Sr/86Srbioapatite values of turkey bones fromChacoan assemblages have a much lower variance than deer and are similar to that ofsmaller mammals. The sampled turkey values show variability most similar tolagomorphs and prairie dogs, suggesting the turkeys from Chaco Canyon wereconsuming a uniform diet and/or were constrained within a limited home range,indicating at least proto-domestication. The population approach has wide applicabilityfor evaluating the presence and nature of domestication when combined with paleo-ecology and behavioral ecology in a variety of animals and environments.
Keywords Domestication . Turkey . Home range . Strontium isotopes . Sourcing .
Southwestern U.S. . Chaco Canyon
Introduction
The prehistory of turkey domestication, with proposed independent origins in northernor central Mexico and the North American Southwest (Speller et al. 2010), is relativelycomplex, and many of the details are still being resolved using a battery of techniques.Domestication of the South Mexican turkey (Meleagris gallopavo gallopavo), nowendangered and ancestor to all domestic turkeys consumed in the world today, was longthought to have occurred around a thousand years ago in south-central Mexico(Flannery 1967). Current thinking, however, is that this domestication actually hap-pened much earlier. Ancient DNA (aDNA) and other evidence now show that the Mayaimported and raised the South Mexican turkey in captivity in the Petén region ofGuatemala, approximately 600 km south of its native distribution, during much of theLate Preclassic through the Late Classic Period (300 BC–AD 900) (Thornton et al.2012).
In the Southwestern U.S., (herein Southwest), phylogeographic analysis, includingmost domestic and North American wild turkey sequences along with aDNA analysesof turkey remains from numerous archaeological sites (200 BC–AD 1800), suggest twodifferent maternal lineages in the prehistoric Southwest, one less variable genetically(haplotype 1) than the other (haplotype 2) and both separate from domesticated andwild M. g. gallopavo (haplotype 3). The less genetically variable lineage (haplotypetermed ancient haplotype 1 or “aHap1”) probably represents a domesticated breed thatoriginated from an undetermined wild progenitor (Speller et al. 2010). This haplotypemore closely resembles both the Eastern U.S. (M. g. sylvestris) and Rio Grande (M. g.intermedia) subspecies (haplotype 1) than the Southwestern Merriam’s turkey (M. g.merriami; haplotype 2). About 15 % of the southwestern archaeological turkey bonesbelong to haplotype 2, suggesting that wild Merriam’s turkeys also were integrated intodomestic stocks. The initial area and history of domestication in the Southwest remainopen to question.
Turkey domestication in the Southwest could have happened as early as AD 200,based both on the identification of “aHap1” and the abundance of Zea mays pollen inturkey coprolites from both Basketmaker II and Pueblo Period layers at the Turkey PenSite in Grand Gulch, southeastern UT (Aasen 1984; Knott 2010; Speller et al. 2010;Lipe et al. 2011). Turkey husbandry appears to have intensified during the PuebloPeriods (AD 700–1300). For example, carbon isotope studies show that turkeys from
Grimstead et al.
Author's personal copy

Pueblo II (AD 900–1150) and Pueblo III sites (AD 1150–1300) in the San Juan Basinconsumed a diet high in C4 plants, most likely reflecting the maize foddering byhumans (Rawlings and Driver 2010; McCaffery et al. 2014). Scanning electronmicroscopy of eggshell suggests that, during the 12th century, Puebloans practiced insitu hatching and purposeful breeding of captive turkeys at Salmon Ruins (Fig. 1) onthe north bank of the San Juan River, northwestern NM (Beacham and Durand 2007).Intensified turkey husbandry might have been motivated by gradual declines in wildgame (Badenhorst and Driver 2009) and agricultural failures during prolongedmegadrought (e.g., 1130–1180) (Akins 1985, 1987). In the region, wild turkeysgenerally inhabit forested areas near running water, a habitat restricted mostly to themountains. To consume turkey, most Pueblos would have had to either hunt afar (tensof kilometers) or raise their own turkeys.
At Chaco Canyon, in the middle of the San Juan Basin, much of the timber used inconstruction included spruce (Picea engelmannii), fir (Abies sp.), ponderosa pine(Pinus ponderosa), and Douglas fir (Pseudotsuga menziesii) that had to be procuredfrom mountain ranges more than 75 km away (Betancourt et al. 1986). Strontium (Sr)isotopic analysis of possible source stands and the dated architectural timber suggestthat the spruce and fir came from the Chuska Mountains to the west and Mt. Taylor tothe south (English et al. 2001), while the ponderosa pine also could have originatedfrom the La Plata and San Juan Mountains and Lobo Mesa/Hosta Butte to the south(Reynolds et al. 2005). Drake et al. (2014) question the specificity of these and other Srisotopic sourcing studies at Chaco Canyon, but there is little denying that tens of
Fig. 1 Vegetation and wild turkey range in the Chaco Canyon region. The approximate range of modernMeleagris gallopavo is within the red line (GIS data from the National Wild Turkey Federation, NWTF; http://www.nwtf.org/NAWTMP/GIS.html). The distribution of plant communities is modified from Brown andLowe (1980). Names are abbreviated: ALP alpine tundra, SAG subalpine grassland, SAF subalpine coniferforest, MCF montane (mixed) conifer forest, PJW pinyon-juniper woodland, GBD Great Basin desert scrub,GBM Great Basin montane scrub, GPL plains and grassland, SDG semi-desert grassland. Ephemeral streamsand perennial streams are shown by dashed and solid blue lines, respectively. Sites and regions discussed intext are shown
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

thousands of constructional timbers originated from far away, regardless of whichmountain range.
The motivation for our current study was the hypothesis that, if it was procured aswild game and not from domesticated animals, the turkey bone recovered from pastexcavations at Chaco Canyon probably originated from similar places as the architec-tural timber. As part of the timber sourcing study, in 2005, we ran 87Sr/86Srbioapatiteanalyses on 28 archaeological bones or teeth of turkey, deer (Odocoileus sp.), andcottontail (Sylvilagus sp.), spanning the period from before AD 800 to after AD 1250.Our initial 87Sr/86Sr turkey ratios clustered around local values for soil and plants,suggesting that the turkeys foraged locally. The tightness of the cluster compared todeer bone further suggested a surprisingly limited foraging range for turkey, whichmade us think that the birds were possibly penned or tethered. To enhance comparisonsacross animals with different home ranges, we ran 87Sr/86Srbioapatite analyses onadditional 34 archaeological bones of deer, cottontail, jackrabbit (Lepus sp.), andprairie dog (Cynomys sp.). Both wild deer and turkey from the Southwest have muchlarger home ranges, exhibit different dispersal behaviors, and should exhibit a widerrange in 87Sr/86Srbioapatite than cottontail, jackrabbit, and prairie dog. We now use thesedata to introduce a novel application that relies on both the absolute values andpopulation variance of 87Sr/86Srbioapatite isotopes, as well as comparative isotopicstudies of animals with different home ranges, to discriminate between wild anddomesticated animals in the archaeological record.
Archaeology and Environment
Chaco Canyon, listed as a world heritage site by the United Nations in 1987, hascommanded the attention of archaeologists and the public owing to its conspicuousmonumental architecture, centrality within an extensive regional exchange network, andbrief history of existence. Cultural habitation within Chaco Canyon dates as far back asthe Paleo-Indian period, but the Basketmaker III through Pueblo II periods (approxi-mately AD 500–1100) represents a time of flourishing human populations (Reed 2000),before the abandonment around AD 1130 with the beginning of the mid-twelfth centurydrought. Chacoans commanded vast regional exchange networks in support of theirlarge but short-lived community located in the San Juan Basin of northwestern NM(Fig. 1). This cultural trajectory is marked by intensification of agricultural practices, thedevelopment of extensive trade networks, large community building projects, andprobably hierarchical social structures (see individual chapters in Lekson 2006).
Chaco Canyon (2073 masl) lies in the middle of the broad basin that is surroundedby mountain ranges with distinct lithologies and vegetation regimes, including theChuska Mountains (highest elevation at 2982 masl), the La Plata Mountains (4035masl), the San Juan Mountains (4361 masl), the San Pedro Mountains (3232 masl), theZuni Mountains (2743 masl), and Mt. Taylor (3444 masl) (Fig. 1). Several springslocated within or near Chaco Canyon provide the only stable water supply, as noperennial rivers run close by (Fig. 1). Chaco Canyon is covered by shrub-grass steppe,with modern rainfall averaging 22.4 cm/year. The complex surficial geology in ChacoCanyon, the broader San Juan Basin, and surrounding mountain ranges is ideal forsourcing studies using Sr isotopes.
Grimstead et al.
Author's personal copy

The distribution of modern and paleovegetation communities in the Chaco Canyonregion is determined by the effects of elevation and slope aspect on temperature andprecipitation. Elevations above approximately 2500 masl support subalpine coniferforests characterized by Engelmann spruce and subalpine fir (Abies lasiocarpa)(Fig. 1). Aspen (Populus tremuloides) is common at elevations below 2900 masl. Atthe lowest elevations, the spruce-fir forest grades into montane conifer forest consistingof ponderosa pine on drier southern slopes and mixed conifer communities of Douglasfir, white fir (Abies concolor), limber pine (Pinus flexilis), and aspen at higher elevationand northern slopes. Gambel oak (Quercus gambelii) becomes common near the lowerlimits (approximately 2300 masl) of ponderosa pine. Pinyon-juniper woodland,consisting of Colorado pinyon (Pinus edulis) and one-seed juniper (Juniperusmonosperma), extend down to approximately 2000 m in the San Juan Basin and coversmuch of Chacra Mesa (2165 masl) bordering the upper reaches of Chaco Canyon to thesouth (Fig. 1). North-facing slopes and shady alcoves on Chacra Mesa harbor restrictedstands of Douglas fir and ponderosa pine (Betancourt and Van Devender 1981;Betancourt et al. 1983).
Fossil packrat middens indicate there was significant elevation lowering of subalpineforests during the late Pleistocene, with midden assemblages dated to approximately13,000–12,000 calYBP at Chaco Canyon (approximately 1900 masl) dominated bylimber pine (Pinus flexilis), Douglas fir, and blue spruce (Picea pungens). Moderndistributions of spruce fir, mixed conifer, and ponderosa pine forests were established inthe San Juan Basin and surrounding mountains during the Early Holocene (Betancourtand Van Devender 1981; Betancourt et al. 1983; Betancourt 1990) and have been stablefor at least the last 2500 years (Petersen and Mehringer 1976). Pollen reconstructionsfrom high elevations in the mountains of NM indicate considerable stability in moun-tain forest communities around the time of Pueblo occupation (Anderson et al. 2008;Jimenez-Moreno et al. 2008; Petersen 1994). Local reduction in the pinyon-juniperwoodland in the immediate vicinity of the Great Houses at Chaco Canyon happenedbetween 1200 and 500 years ago, which could reflect fuel or timber overharvesting bythe Pueblos (Betancourt and Van Devender 1981; Betancourt et al. 1983; Samuels andBetancourt 1982) or possibly drought-induced tree die offs near the woodland’s lowerlimits at approximately 1900 masl, akin to what happened elsewhere in the Southwestduring the 1950s or 2002–2003 droughts (Allen and Breshears 1998; Swetnam andBetancourt 1998). Except pinyon-juniper woodland in Chaco Canyon proper, regionalvegetation was probably the same as today during Puebloan occupation.
Strontium Isotopes
Strontium (Sr) isotope values have been broadly used to source human, animal, andplant remains in the archaeological record (Bentley 2006). This works because87Sr/86Sr values in teeth, bone, and plant cellulose generally reflect the surficialgeology of their source areas, which can vary considerably in complex terrain. Inregions where bedrock lithology and age is variable and distinct, 87Sr/86Sr ratios areuseful as a sourcing tool because they are dependent on both time and lithology. That is,bedrock lithologies of varying age have unique 87Sr/86Sr ratios due to the radiogenicdecay of 87Rb to 87Sr and the initial ratio of 87Sr/86Sr ratio at the time of mineralformation. Plants metabolize local bedrock and dust derived strontium from the soil and
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

groundwater in which they live, so that the ratios in vegetation reflect geographicalvariations in the 87Sr/86Sr of soil, dust, and water (e.g., Graustein and Armstrong 1983;Capo et al. 1998; Reynolds et al. 2012). Because there is no biological fractionation ofthe 87Sr/86Sr ratios, animal bone and teeth chemically reflect the 87Sr/86Sr ratio of localsoils, consumed plants, meteoric waters, and other animals within their home range.Plants provide the bulk of available strontium, especially in non-obligate and partiallyobligate drinkers (Sillen and Kavanagh 1982; Hoppe et al. 2003). Within the ChacoCanyon region, strontium isotope ratios have been used to source architectural logs inAnasazi Great Houses (English et al. 2001; Reynolds et al. 2005), agricultural products(Benson et al. 2003), and animal remains (Grimstead 2011) to geochemically distinctterrains.
The isotopic composition of discrete resource patches with known distances fromChaco Canyon have been clearly identified in previous work (Benson et al. 2003;English et al. 2001; Grimstead 2011; Reynolds et al. 2005). Chaco Canyon is situatedat the boundary between Cretaceous marine deposits and Cretaceous through Neogenefluvial deposits, producing 87Sr/86Sr range of 0.70897–0.70964 (Grimstead 2011).Sandstone, shale, and mudstone of various age and composition make up the broaderSan Juan Basin (87Sr/86Sr range=0.70871–0.71219). The Chuska Mountains west ofChaco Canyon are underlain mainly by Oligo-Miocene sandstone, locally overlain byandesitic basalt (87Sr/86Sr range=0.70706–0.71022). The San Mateo Mountains arecomposed of Pliocene lava and ash flows (87Sr/86Sr range=0.70550–70868). The SanPedro/Sierra Nacimientos are composed of Paleozoic sedimentary rocks overlyingProterozoic granitoids (87Sr/86Sr range =0.71191–72791). The La Plata and San JuanMountains are highly varied geologically and composed of Precambrian metamorphicrocks, Paleozoic/Mesozoic sedimentary rocks, and Cenozoic intrusive igneous andvolcanic rocks (87Sr/86Sr range =0.70865–0.71399).
Animal Ecology
87Sr/86Sr ratios derived from consumption are highly localized for animals with smallhome ranges and dispersal distances and more generalized for wide-ranging taxa (cf.Feranec et al. 2007). Our isotopic analysis of archaeological faunas focused on fourprey groups recovered from Chacoan middens: deer (Odocoileus sp.) and turkey(Meleagris gallopavo) and animals with small home ranges: prairie dogs (Cynomyssp.) and lagomorphs: jackrabbits (Lepus sp.) and cottontails (Sylvilagus sp.). These taxawere selected for their utility in understanding the 87Sr/86Sr ratio variation presentwithin animals having variable-sized home ranges and dispersal behaviors. The homeranges (Burt 1943), dispersal behaviors (Greenwood 1980), and habitat preferences ofthese groups are important for interpreting strontium isotope ratios to source areas ofprocurement.
Prairie Dog Ecology
The Gunnison’s Prairie Dog (Cynomys gunnisoni) (Whitaker 1996) is the most likelycandidate to be found in Chacoan assemblages, but the black-tailed prairie dog(Cynomys ludovicianus) also could have been present in the past (Merriam 1902;
Grimstead et al.
Author's personal copy

Longhurst 1944; Wagner et al. 2006). Prairie dog natal and breeding dispersal occursover less than a few hundred meters, and foraging areas can be measured in squaremeters (Fitzgerald and Lechleitner 1974; Longhurst 1944). The Gunnison’s Prairie Dogprefers short grass prairies in upper elevation (1800–3600 masl) mountain valleys andplateaus. It is a non-obligate drinker and therefore does not need a permanent standingwater supply (Whitaker 1996; Knowles 2002).
Jackrabbit Ecology
Whereas the black-tailed jackrabbit (Lepus californicus) is the most likely species in theChacoan assemblages, the white-tailed jackrabbit (Lepus townsendii) and snowshoehare (Lepus americanus) may have found their way into the Chacoan menu (Frey andMalaney 2006; Whitaker 1996). Jackrabbits eat a variety of foods ranging fromsucculents to grasses to shrubs (Best 1996; Flinders and Hansen 1975; 1972; Hansenand Flinders 1969; Mohammad et al. 1988), fungi (Maser et al. 1988), and in rare casescarrion along with some gravel and sand (Vorhies and Taylor 1933). Their home rangesvary from 0.2 to 1.4 km2, with seasonal and natal dispersals of usually less than 2 km(Lechleitner 1958; French et al. 1965; Harestad and Bunnel 1979). There is noevidence that jackrabbits require standing water on a daily basis, rather they obtainmost of their water from plants (Best 1996; Nagy et al. 1976; Vorhies 1945).
Cottontail Ecology
The desert cottontail (Sylvilagus audubonii), mountain cottontail (Sylvilagus nuttallii),and the eastern cottontail (Sylvilagus floridanus) were all potentially available toprehistoric hunters in or around Chaco Canyon. Desert cottontails tolerate diversehabitats, but most can be found at or below 2000 masl, leaving the upper elevationsto the mountain cottontail (Chapman and Willner 1978; Whitaker 1996). Althoughcottontails will forage in open areas, they prefer areas with cover (Orr 1940; Ingles1941). All species feed on a variety of grasses, shrubs, forbs, bark, and succulents(Turkowski 1975). Home ranges are measured in square meters, with males usinglarger ranges than females (Fitch 1947; Ingles 1941). Cottontails do not requirestanding water, meeting their daily water requirements with their food (Vorhies 1945).
Deer Ecology
Most white-tailed deer (Odocoileus virginianus) and black-tailed deer (Odocoileushemionus) are philopatric (Dasmann and Taber 1956; Longhurst et al. 1952; Rue1988; Tierson et al. 1985; Zwickel et al. 1953). Home ranges are established by theage of 1 or 2 years, and individuals generally stay within that range for life (Dasmannand Taber 1956; Longhurst et al. 1952). Home ranges are smaller where forage isabundant and evenly distributed (Heffelfinger 2006). Bucks may have home rangestwice the size of does, and the breeding season can bring bucks well outside of theirhome ranges (up to 12 km) (Koerth and Bryant 1982; Ragotzkie and Bailey 1991;Relyea et al. 2000; Rodgers 1978; Taber and Dasmann 1958). Home ranges can varyfrom 1 to 2 km2 to approximately 60 km2 (Dasmann and Taber 1956; Dickinson andGarner 1979; Gallina et al. 1997; Horejsi et al. 1988; Krausman 1985; Taber and
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

Dasmann 1958), but the majority of studies indicate home ranges at or below 10 km2
(Fox and Krausman 1994; Haywood et al. 1987; Heffelfinger 2006; Horejsi et al. 1988;Koerth et al. 1985; Lawrence et al. 1994; Ragotzkie and Bailey 1991; Raught 1967;Relyea et al. 2000; Rodgers 1978; Wallmo 1981). Deer herds may disperse when foodis seasonally scarce, traveling only as far as needed (5–30 km) to find forage (Haywoodet al. 1987; Heffelfinger 2006; Mierau and Schmidt 1981). Even in these conditions,deer display seasonal philopatry, returning every year to the same summer and winterhome ranges (Heffelfinger 2006; Longhurst et al. 1952).
Deer eat a variety of forbs, grasses, shrubs, and succulents, depending upon avail-ability (Gill et al. 1983; Heffelfinger 2006). They are able to satisfy most of their waterneeds from eating plants with high water content during the cooler months(Heffelfinger 2006). When summer temperatures soar and plant moisture content islow, deer must drink standing (free) water to survive (Heffelfinger 2006; O’Brien et al.2006; Rosenstock et al. 2004). Herds that include both sexes may form in the winterbut rarely in great numbers. Females may form small groups during rutting season, butthey disperse with their fawns the rest of the year.
Wild Turkey Ecology
According to modern distributions of wild turkey subspecies, two varieties would havebeen available to the Chacoans: the Rio Grande (Meleagris gallopavo intermedia) andMerriam’s (Meleagris gallopavo merriami) (National Wild Turkey Federation 2012).Rio Grande gobblers’ and hens’ primary habitats occur in brushy areas near streamsand rivers, generally in mesquite, pine, and scrub oak forests (NRCS 1999). Merriam’sturkeys require trees for roosting, thus they prefer ponderosa pine forests, pinyon-juniper woodlands, and other conifer- forests (Hoffman 1968; Scott and Boecker 1975).Wild turkeys in the Southwest are partially obligate drinkers, requiring a stable watersupply during the warm summer months (Scott and Boecker 1975; Spicer 1959).
Wild turkeys in the Southwest have much larger home ranges and greater distancesof seasonal dispersal (Logan 1973; MacDonald 1963; Scott and Boecker 1975; Walker1951) when compared to flocks from habitats with greater primary productivity (e.g.,Barwick and Speake 1973; Ellis and Lewis 1967; Paisley et al. 2000; Wigley et al.1986). Birds fromMinnesota were documented to have home ranges averaging 0.1 km2
in the winter and 20 km2 during the spring (Porter 1978). Turkeys in upland desertecosystems occupy home range areas from 5 to 50 km2, with sub-adults having largerhome ranges (Hoffman 1991; Scott and Boecker 1975). Seasonal dispersals find thesebirds traveling from 5 to 29 km (Hoffman 1991; MacDonald 1963; Scott and Boecker1975; Spicer 1959; Walker 1951), and birds have been known to occupy high elevationhabitats above 3000 masl (Spicer 1959). Important for understanding prehistoric turkeybehavioral ecology is that hunting pressure has been shown to increase home range sizeand the distance the bird will travel from roosting sites (Hoffman 1991).
Results
The 87Sr/86Srbioapatite ratios of turkey, prairie dog, and cottontail and jackrabbit teeth andbone are tightly constrained (0.70912–0.70951) (Table 1). Deer show a much wider
Grimstead et al.
Author's personal copy
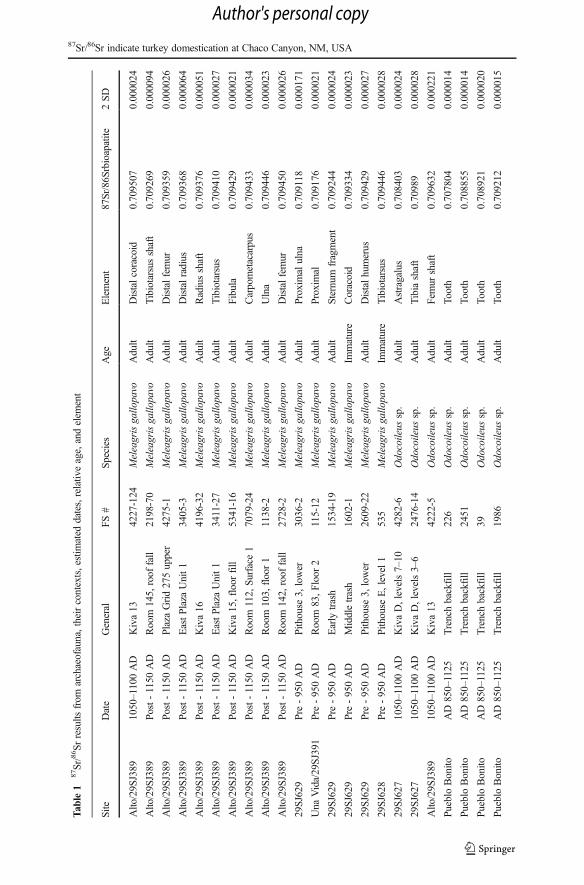
Tab
le1
87Sr/86Sr
results
from
archaeofauna,theircontexts,estim
ated
dates,relativ
eage,andelem
ent
Site
Date
General
FS#
Species
Age
Element
87Sr/86S
rbioapatite
2SD
Alto/29S
J389
1050–1100AD
Kiva13
4227-124
Meleagrisgallo
pavo
Adult
Distalcoracoid
0.709507
0.000024
Alto/29S
J389
Post-1150
AD
Room
145,
roof
fall
2198-70
Meleagrisgallo
pavo
Adult
Tibiotarsus
shaft
0.709269
0.000094
Alto/29S
J389
Post-1150
AD
PlazaGrid275upper
4275-1
Meleagrisgallo
pavo
Adult
Distalfemur
0.709359
0.000026
Alto/29S
J389
Post-1150
AD
EastPlazaUnit1
3405-3
Meleagrisgallo
pavo
Adult
Distalradius
0.709368
0.000064
Alto/29S
J389
Post-1150
AD
Kiva16
4196-32
Meleagrisgallo
pavo
Adult
Radiusshaft
0.709376
0.000051
Alto/29S
J389
Post-1150
AD
EastPlazaUnit1
3411-27
Meleagrisgallo
pavo
Adult
Tibiotarsus
0.709410
0.000027
Alto/29S
J389
Post-1150
AD
Kiva15,floor
fill
5341-16
Meleagrisgallo
pavo
Adult
Fibula
0.709429
0.000021
Alto/29S
J389
Post-1150
AD
Room
112,
Surface1
7079-24
Meleagrisgallo
pavo
Adult
Carpometacarpus
0.709433
0.000034
Alto/29S
J389
Post-1150
AD
Room
103,
floor1
1138-2
Meleagrisgallo
pavo
Adult
Ulna
0.709446
0.000023
Alto/29S
J389
Post-1150
AD
Room
142,
roof
fall
2728-2
Meleagrisgallo
pavo
Adult
Distalfemur
0.709450
0.000026
29SJ629
Pre-950AD
Pithouse
3,lower
3036-2
Meleagrisgallo
pavo
Adult
Proxim
alulna
0.709118
0.000171
Una
Vida/29SJ391
Pre-950AD
Room
83,F
loor
2115-12
Meleagrisgallo
pavo
Adult
Proxim
al0.709176
0.000021
29SJ629
Pre-950AD
Early
trash
1534-19
Meleagrisgallo
pavo
Adult
Sternum
fragment
0.709244
0.000024
29SJ629
Pre-950AD
Middletrash
1602-1
Meleagrisgallo
pavo
Immature
Coracoid
0.709334
0.000023
29SJ629
Pre-950AD
Pithouse
3,lower
2609-22
Meleagrisgallo
pavo
Adult
Distalhumerus
0.709429
0.000027
29SJ628
Pre-950AD
Pithouse
E,level1
535
Meleagrisgallo
pavo
Immature
Tibiotarsus
0.709446
0.000028
29SJ627
1050–1100AD
KivaD,levels7–10
4282-6
Odocoileus
sp.
Adult
Astragalus
0.708403
0.000024
29SJ627
1050–1100AD
KivaD,levels3–6
2476-14
Odocoileus
sp.
Adult
Tibiashaft
0.70989
0.000028
Alto/29S
J389
1050–1100AD
Kiva13
4222-5
Odocoileus
sp.
Adult
Femur
shaft
0.709632
0.000221
Pueblo
Bonito
AD
850–1125
Trenchbackfill
226
Odocoileus
sp.
Adult
Tooth
0.707804
0.000014
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2451
Odocoileus
sp.
Adult
Tooth
0.708855
0.000014
Pueblo
Bonito
AD
850–1125
Trenchbackfill
39Odocoileus
sp.
Adult
Tooth
0.708921
0.000020
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1986
Odocoileus
sp.
Adult
Tooth
0.709212
0.000015
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy
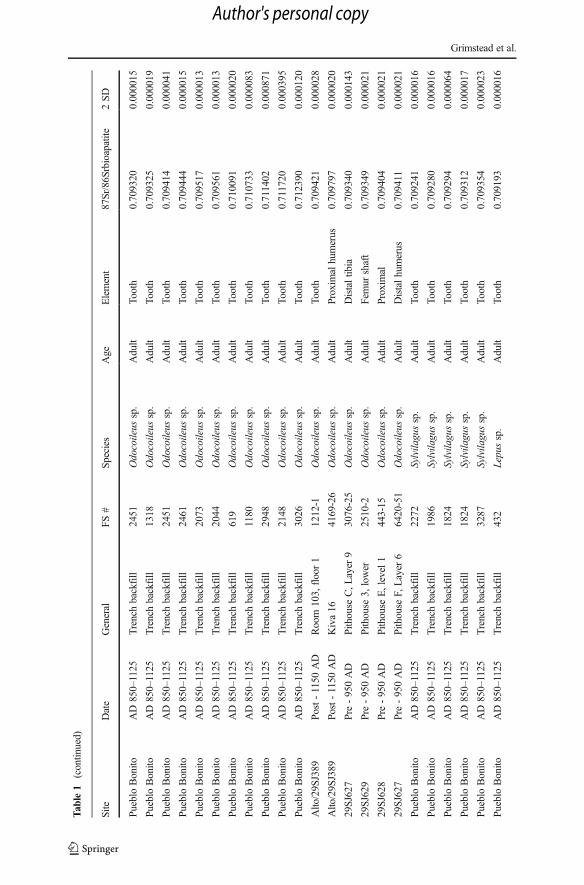
Tab
le1
(contin
ued)
Site
Date
General
FS#
Species
Age
Element
87Sr/86S
rbioapatite
2SD
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2451
Odocoileus
sp.
Adult
Tooth
0.709320
0.000015
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1318
Odocoileus
sp.
Adult
Tooth
0.709325
0.000019
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2451
Odocoileus
sp.
Adult
Tooth
0.709414
0.000041
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2461
Odocoileus
sp.
Adult
Tooth
0.709444
0.000015
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2073
Odocoileus
sp.
Adult
Tooth
0.709517
0.000013
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2044
Odocoileus
sp.
Adult
Tooth
0.709561
0.000013
Pueblo
Bonito
AD
850–1125
Trenchbackfill
619
Odocoileus
sp.
Adult
Tooth
0.710091
0.000020
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1180
Odocoileus
sp.
Adult
Tooth
0.710733
0.000083
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2948
Odocoileus
sp.
Adult
Tooth
0.711402
0.000871
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2148
Odocoileus
sp.
Adult
Tooth
0.711720
0.000395
Pueblo
Bonito
AD
850–1125
Trenchbackfill
3026
Odocoileus
sp.
Adult
Tooth
0.712390
0.000120
Alto/29S
J389
Post-1150
AD
Room
103,
floor1
1212-1
Odocoileus
sp.
Adult
Tooth
0.709421
0.000028
Alto/29S
J389
Post-1150
AD
Kiva16
4169-26
Odocoileus
sp.
Adult
Proxim
alhumerus
0.709797
0.000020
29SJ627
Pre-950AD
Pithouse
C,L
ayer
93076-25
Odocoileus
sp.
Adult
Distaltib
ia0.709340
0.000143
29SJ629
Pre-950AD
Pithouse
3,lower
2510-2
Odocoileus
sp.
Adult
Femur
shaft
0.709349
0.000021
29SJ628
Pre-950AD
Pithouse
E,level1
443-15
Odocoileus
sp.
Adult
Proxim
al0.709404
0.000021
29SJ627
Pre-950AD
Pithouse
F,Layer
66420-51
Odocoileus
sp.
Adult
Distalhumerus
0.709411
0.000021
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2272
Sylvila
gussp.
Adult
Tooth
0.709241
0.000016
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1986
Sylvila
gussp.
Adult
Tooth
0.709280
0.000016
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1824
Sylvila
gussp.
Adult
Tooth
0.709294
0.000064
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1824
Sylvila
gussp.
Adult
Tooth
0.709312
0.000017
Pueblo
Bonito
AD
850–1125
Trenchbackfill
3287
Sylvila
gussp.
Adult
Tooth
0.709354
0.000023
Pueblo
Bonito
AD
850–1125
Trenchbackfill
432
Lepussp.
Adult
Tooth
0.709193
0.000016
Grimstead et al.
Author's personal copy
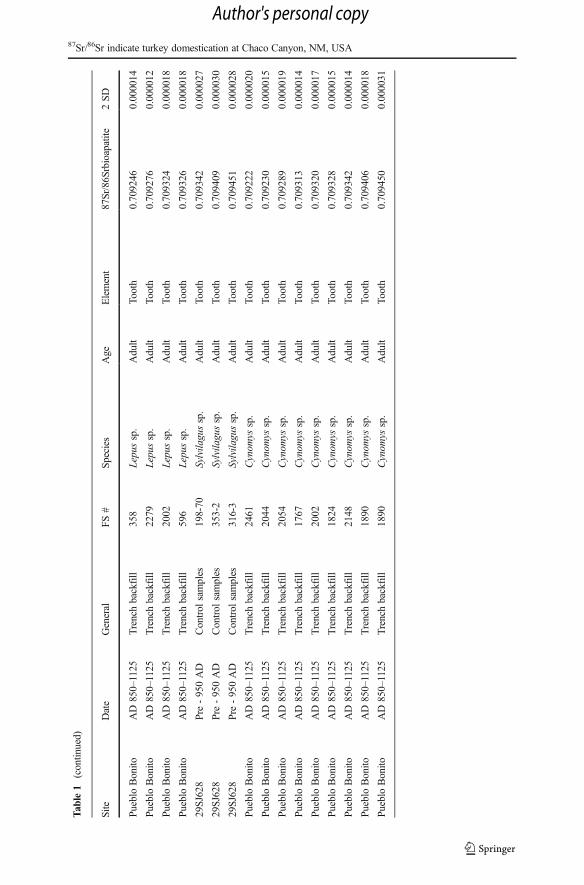
Tab
le1
(contin
ued)
Site
Date
General
FS#
Species
Age
Element
87Sr/86S
rbioapatite
2SD
Pueblo
Bonito
AD
850–1125
Trenchbackfill
358
Lepussp.
Adult
Tooth
0.709246
0.000014
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2279
Lepussp.
Adult
Tooth
0.709276
0.000012
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2002
Lepussp.
Adult
Tooth
0.709324
0.000018
Pueblo
Bonito
AD
850–1125
Trenchbackfill
596
Lepussp.
Adult
Tooth
0.709326
0.000018
29SJ628
Pre-950AD
Control
samples
198-70
Sylvila
gussp.
Adult
Tooth
0.709342
0.000027
29SJ628
Pre-950AD
Control
samples
353-2
Sylvila
gussp.
Adult
Tooth
0.709409
0.000030
29SJ628
Pre-950AD
Control
samples
316-3
Sylvila
gussp.
Adult
Tooth
0.709451
0.000028
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2461
Cynom
yssp.
Adult
Tooth
0.709222
0.000020
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2044
Cynom
yssp.
Adult
Tooth
0.709230
0.000015
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2054
Cynom
yssp.
Adult
Tooth
0.709289
0.000019
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1767
Cynom
yssp.
Adult
Tooth
0.709313
0.000014
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2002
Cynom
yssp.
Adult
Tooth
0.709320
0.000017
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1824
Cynom
yssp.
Adult
Tooth
0.709328
0.000015
Pueblo
Bonito
AD
850–1125
Trenchbackfill
2148
Cynom
yssp.
Adult
Tooth
0.709342
0.000014
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1890
Cynom
yssp.
Adult
Tooth
0.709406
0.000018
Pueblo
Bonito
AD
850–1125
Trenchbackfill
1890
Cynom
yssp.
Adult
Tooth
0.709450
0.000031
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy
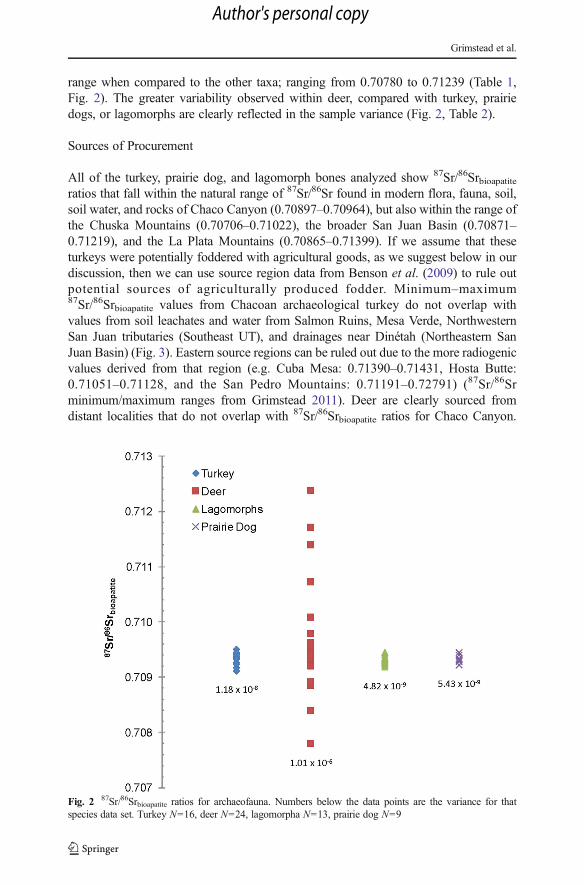
range when compared to the other taxa; ranging from 0.70780 to 0.71239 (Table 1,Fig. 2). The greater variability observed within deer, compared with turkey, prairiedogs, or lagomorphs are clearly reflected in the sample variance (Fig. 2, Table 2).
Sources of Procurement
All of the turkey, prairie dog, and lagomorph bones analyzed show 87Sr/86Srbioapatiteratios that fall within the natural range of 87Sr/86Sr found in modern flora, fauna, soil,soil water, and rocks of Chaco Canyon (0.70897–0.70964), but also within the range ofthe Chuska Mountains (0.70706–0.71022), the broader San Juan Basin (0.70871–0.71219), and the La Plata Mountains (0.70865–0.71399). If we assume that theseturkeys were potentially foddered with agricultural goods, as we suggest below in ourdiscussion, then we can use source region data from Benson et al. (2009) to rule outpotential sources of agriculturally produced fodder. Minimum–maximum87Sr/86Srbioapatite values from Chacoan archaeological turkey do not overlap withvalues from soil leachates and water from Salmon Ruins, Mesa Verde, NorthwesternSan Juan tributaries (Southeast UT), and drainages near Dinétah (Northeastern SanJuan Basin) (Fig. 3). Eastern source regions can be ruled out due to the more radiogenicvalues derived from that region (e.g. Cuba Mesa: 0.71390–0.71431, Hosta Butte:0.71051–0.71128, and the San Pedro Mountains: 0.71191–0.72791) (87Sr/86Srminimum/maximum ranges from Grimstead 2011). Deer are clearly sourced fromdistant localities that do not overlap with 87Sr/86Srbioapatite ratios for Chaco Canyon.
Fig. 2 87Sr/86Srbioapatite ratios for archaeofauna. Numbers below the data points are the variance for thatspecies data set. Turkey N=16, deer N=24, lagomorpha N=13, prairie dog N=9
Grimstead et al.
Author's personal copy
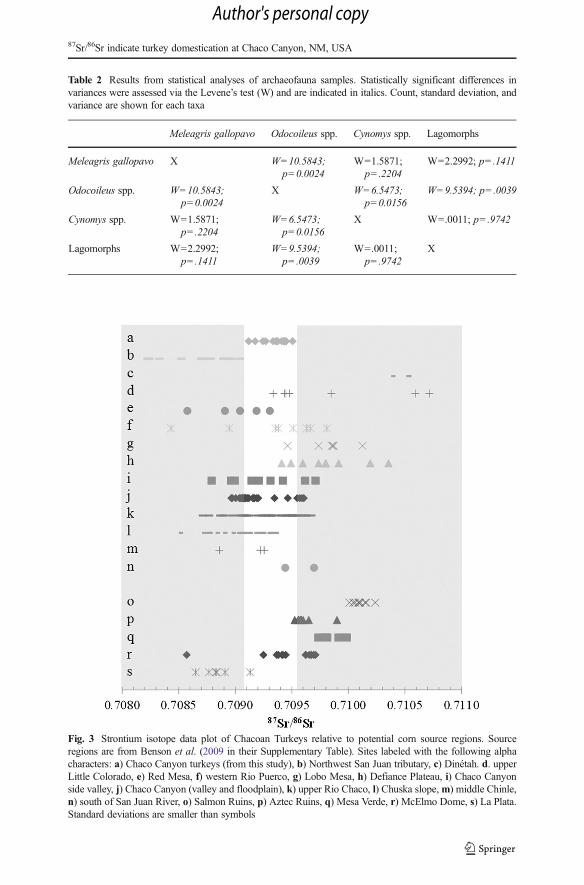
Table 2 Results from statistical analyses of archaeofauna samples. Statistically significant differences invariances were assessed via the Levene’s test (W) and are indicated in italics. Count, standard deviation, andvariance are shown for each taxa
Meleagris gallopavo Odocoileus spp. Cynomys spp. Lagomorphs
Meleagris gallopavo X W=10.5843;p=0.0024
W=1.5871;p=.2204
W=2.2992; p=.1411
Odocoileus spp. W=10.5843;p=0.0024
X W=6.5473;p=0.0156
W=9.5394; p=.0039
Cynomys spp. W=1.5871;p=.2204
W=6.5473;p=0.0156
X W=.0011; p=.9742
Lagomorphs W=2.2992;p=.1411
W=9.5394;p=.0039
W=.0011;p=.9742
X
Fig. 3 Strontium isotope data plot of Chacoan Turkeys relative to potential corn source regions. Sourceregions are from Benson et al. (2009 in their Supplementary Table). Sites labeled with the following alphacharacters: a) Chaco Canyon turkeys (from this study), b) Northwest San Juan tributary, c) Dinétah. d. upperLittle Colorado, e) Red Mesa, f) western Rio Puerco, g) Lobo Mesa, h) Defiance Plateau, i) Chaco Canyonside valley, j) Chaco Canyon (valley and floodplain), k) upper Rio Chaco, l) Chuska slope,m) middle Chinle,n) south of San Juan River, o) Salmon Ruins, p) Aztec Ruins, q) Mesa Verde, r) McElmo Dome, s) La Plata.Standard deviations are smaller than symbols
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

Ten of the 24 deer specimens produced 87Sr/86Srbioapatite ratios that fall outside thenatural range for Chaco Canyon, meaning these individuals are non-local, likelycoming from 87Sr/86Sr ratio source regions associated with montane habitats(Grimstead 2011).
Levene’s Test of Equality of Variance
The Levene’s test assesses homogeneity of variance between two samples (Gastwirthet al. 2009). Generally, the Levene’s test is used to evaluate whether the variance of twopopulations are equal as a precursor to additional statistical procedures. Here, weapplied it to test the hypothesis that the 87Sr/86Sr ratios of domesticated turkeypopulations would reflect a restricted diet (low variance) due to their restricted homeranges, when compared to deer with large variance with exposure to larger homeranges, dispersal behavior, and greater human hunting ranges. Domesticated turkey87Sr/86Srbioapatite ratios should be statistically equivalent to lagomorphs and prairie dog87Sr/86Sr ratios, reflecting these animal’s small home ranges, restricted dispersaldistances, and localized human management patterns. Our results show that turkey,lagomorph, and prairie dog samples have uniformly low variance compared to deer(Table 2, Fig. 2). There is no difference in the turkey 87Sr/86Srbioapatite ratio variancecompared to lagomorphs and prairie dogs (Table 2, Fig. 2).
Discussion
The presence of domesticated turkeys in the Four Corners region has been previouslyinferred from genetic and embryonic development studies (Beacham and Durand 2007;Speller et al. 2010), and carbon isotopic data suggest human foddering of these birds(McCaffery et al. 2014; Rawlings and Driver 2010). Akins (1985) suggested thatturkey domestication took place relatively late during the occupational history of ChacoCanyon, to fulfill a need for resources above what could be supplied within the localhabitat. This study demonstrates that sampled Chacoan turkey 87Sr/86Srbioapatite ratios,as a whole, have a degree of low variance when compared to archaeological deer. Theobserved variability in the turkey population is most similar to locally sourced smallmammals with small home ranges. This pattern indicates that the Chacoan turkeys livedwithin a constricted home range and were consuming uniform diets (from a strontiumisotope perspective), as opposed to modern wild turkeys, which display large homeranges, significant dispersal behavior, and preferences for tree roosting sites.Furthermore, this appears to be the case even for early contexts, although sample sizeprevents us from having a high degree of certainty for the time period as a whole.
Because they prefer forest roosting sites, modern wild turkey home ranges do notoccur in or within approximately 30 km of Chaco Canyon (Fig. 1). If the archaeologicalbone at Chaco Canyon originated from the hunting of wild birds, we would certainlyexpect them to exhibit highly variable and even extralocal 87Sr/86Srbioapatite ratiossimilar to deer, which has a similar home range and dispersal behavior. In contrast,the results from 16 turkey specimens of varying times and contexts show that ChacoCanyon turkey have very narrow 87Sr/86Srbioapatite ratios and low variability whencompared to deer, but similar to values from small mammals with small, local home
Grimstead et al.
Author's personal copy

ranges. In addition to the low isotope variability, no turkey specimen can be sourced toa geological region outside of the Late Cretaceous marine sandstone and shales ofChaco Canyon. Because modern wild birds require tree stands for roosting sites, whichwere likely sparse during Pueblo occupation, we can conclude that our data represents aturkey population whose diet and ranging behavior was controlled in some way byhumans, rather than hunted across broad landscapes. Undoubtedly, our results could befurther bolstered by aDNA analysis, zooarchaeological methods, eggshell ontologicalstudies, carbon and nitrogen isotopes, and evidence of concentrated turkey dungaccumulation. Presently, we cannot say for certain whether Chacoan turkey homeranges were constrained by some sort of barricade, such as pueblo walls, or if thebirds were penned.
Akins (a co-author of this paper) recalls turkeys from a variety of Chacoan assem-blages having an unusually high frequency of healed limb fractures compared to allother faunal classes, which has been observed at other Puebloan sites in the region (e.g.Durand and Durand 2008; Gillespie 1991; Lang and Harris 1984). Furthermore, thefirst author observed one potential cause of these fractures, while conducting fieldworkin Azerbaijan. While their chicks foraged freely, domesticated turkey hens weretethered by the leg with a piece of rope to a restricted foraging area that was movedperiodically. The hatchlings would remain close to the mother, but the mother wouldaggressively attempt to free herself from the tether (contact primary author for a videoof this phenomenon). The ferocious pulling and jerking from the tether most certainlycaused stress fracturing that may not completely heal between clutches, perhapseventually resulting in a fracture. Several sites in the region display similar patternsof healed limb fractures in Turkeys including: Salmon Ruins, Bluff Great House, andthe Eleventh Hour site (Fothergill 2012). Additional archaeological and experimentalresearch should be conducted to elucidate the management practices behind theseabnormally high frequencies of limb fractures among domesticated turkeys.
Turkey pens are another potential explanation for constrained home ranges. Severalsites within the broader Four Corners region have turkey pen features, often withassociated turkey dung deposits, including Mug House (Rohn 1971), Big JuniperHouse (Swannack 1969), Johnson Canyon (Nickens 1981), Balcony House Mesa inMesa Verde, Tularosa Cave (Schorger 1966), and Salmon Ruins (Reed 2006). To date,Great Houses within Chaco Canyon have shown no archaeological evidence of pens,which might suggest barricaded containment within pueblo walls or abandoned roomsor perhaps even physical tethering. Alternatively, Chacoan birds may have beenimported from nearby communities actively raising turkeys, but not from SalmonRuins, Mesa Verde, Northwestern San Juan tributaries (Southeast UT), and drainagesnear Dinétah, which would have yielded very different 87Sr/86Srbioapatite ratios.
Carbon isotope data from turkey remains in the Northern San Juan Basin regionhave shown approximately 50 % or more of the diet resulted from C4 plant consump-tion, consistent with intensive maize foddering (McCaffery et al. 2014). Human carbonisotope values from the McCaffery and colleagues (2014) study overlap significantlywith the turkey values; thus, it is difficult to discern if these birds were simply freeranging and eating table scraps or confined and directly foddered by humans. Adifferent animal husbandry practice is indicated by carbon data from Shields Pueblo,a site in Southwest Colorado. Rawlings and Driver’s (2010) data indicate a more mixeddiet consistent with free ranging in an environment where C3, CAM, and C4 plants are
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

available. Some foddering may have occurred at Shields Pueblo, but the carbon dataoverlaps with values from jackrabbits and cottontails at the site, obscuringinterpretation.
The 87Sr/86Srbioapatite ratio results from this study clearly indicate that all of theChacoan turkeys, from a variety of time periods and contexts, were eating anisotopically uniform diet, which is an unlikely result if these large-bodied birds werehunted in the wild. This uniform diet could have been directly fed to penned birds or byconfining the birds, thereby forcing them to consume a uniform fare of local insects,plants, and table scraps. Compelling evidence for turkey foddering in the Southwestcomes from Schorger’s (1961) analysis of a penned desiccated turkey from TularosaCave, NM, where turkey dung recovered from the breast feathers contained legumestarch and the crop contained 190 cm3 of corn kernels. Schorger (1961: 142) alsocollected turkey dung samples that contained both legume and corn starch from pens atBalcony Mesa.
Owing to the methodology applied in this paper, we can say with certainty that thesebirds were neither imported from potential northern San Juan localities (i.e. Dinétah,Salmon Ruins, Southwestern UT, or Mesa Verde), nor were they being foddered withagricultural goods from these regions. It is possible these birds were raised locally atChaco, scavenging and foraging locally produced goods or being foddered withagricultural goods from the Rio Chaco Corridor. It is also possible that these birdswere being raised or foddered with agriculture goods from areas to the west (Chuskanslopes), or to the south (e.g. Lobo Mesa, Rio Puerco, and Red Mesa). These results areconsistent with interpretations from Benson (2012), which suggests corn cobs fromChaco Canyon were being locally produced in the Rio Chaco corridor pre-AD 1130.They argue that after AD 1180, corn came from either the Zuni or Totah region (Benson2012). Our study concludes that the Totah region did not contribute significantquantities of agricultural goods to Chaco Canyon turkey fodder, if indeed the fodderwas non-local during this time. Akins (1985) has suggested that the lack of immatureturkey bones during the late periods in the Chacoan sequence suggests these birds wereimports. If this is the case, then these birds likely came from the Chuskan slopes or theZuni region.
Our results suggest that the Chacoan turkeys lived within a restricted home rangeand most likely were domesticated and confined within pueblo walls, either by penningor tethering. We acknowledge four general blindspots in our interpretation that could beresolved with further study. First, due to different initial objectives and limited sampleat the time of the Sr isotopic study, we did not analyze carbon isotopes from thearchaeological bone collagen. Together, strontium and carbon isotopic ratios couldhave provided a much stronger case. Second, the tight clustering of 87Sr/86Srbioapatitevalues from the archaeological turkey bones around local soil values could haveresulted from diagenesis. This is not borne out, however, by the large spread of thearchaeological deer bones, which were exposed to the same burial environment. Thereare no known differential effects of diagenesis based on the different size and structureof the crystalline lattice in turkey or bird hydroxyapatite versus mammals. Third,87Sr/86Srbioapatite in wild turkey populations has not been studied and could be muchnarrower than we assumed in this study. And fourth, reconstructions of pastenvironments or our uniformitarian assumptions based on the current habits ofMerriam’s wild turkey could be incorrect, and wild turkeys were previously more
Grimstead et al.
Author's personal copy

common and occupied small home ranges at low elevations in the San Juan Basin, notalways in near proximity to running water.
Our comparative methods could have wide applicability to the question of animalhusbandry practices beyond Chaco Canyon and for other domesticated animals. In theright geological setting, population-level variance in Sr isotopes could be used to detectthe subtleties of herd animals that were still being hunted from the wild versus proto-domesticates whose home ranges were under some human control. Sr isotopic variancecould help identify proto-domesticates in the archaeological record and perhaps informhow and why herd animals were domesticated.
Conclusions
This study demonstrates the potential of utilizing 87Sr/86Srbioapatite ratio variability toexamine ranging behavior of an entire animal population, and how this information canbe used in studies of animal domestication. Here, we relied on the relative variance of87Sr/86Srbioapatite ratios in bones of turkeys and other animals to help discriminate wildversus domesticated turkeys in the archaeological record at Chaco Canyon, NM, USA.If wild turkeys had been hunted from their preferred habitats, then we should have seensignificant variability in 87Sr/86Srbioapatite ratio values of turkey bones recovered fromarchaeological contexts. The small variance observed within the Chacoan turkeysample was instead similar to locally sourced taxa with very small home ranges, suchas lagomorphs and rodents. The turkeys could have been raised locally on forage orimported from areas near the Chuska or Zuni Mountains. Another possibility is that theturkeys could have been raised locally, but fed with fodder from these distant localities.The 87Sr/86Srbioapatite ratios rule out bird or fodder importation from source areas to thenorth or east. The methodology extends our understanding of turkey domesticationbeyond a simple presence-absence of domesticates. The use of isotopic data suggeststhat these birds probably were confined, not free ranging, and were not wild animalshunted within the local habitat. We hope that future studies across many regions canfurther extend our use of Sr isotopes to population level assessment of domesticationhistory and process in conjunction with interdisciplinary approaches, such as aDNA,zooarchaeology, and ontogeny.
Methods and Sampling
Bone and teeth from deer [number of identified specimens (NISP=24)], turkey (NISP=16), jackrabbits and cottontails (NISP=13), and prairie dogs (NISP=9) were analyzedas part of this study. Nancy Akins selected the non-Pueblo Bonito samples fromarchaeofaunal assemblages associated with the National Park Service’s excavationsof several contexts within Chaco Canyon during the 1970s (Akins 1985; for dating ofcontexts see Mathien 1985). Pueblo Bonito samples were derived from a 2007 exca-vation conducted by W. Wills (University of New Mexico) and were selected byJonathan Driver (Simon Frasier University). The Wills excavation occurred withinthe backfill of a previously excavated unit. For this reason the dates for the PuebloBonito sample can only be assigned to the entire occupational span of the Great House
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

(approximately A.D. 850–1125). In both samples care was taken to avoid collection ofspecimens from a single individual.
Samples were mechanically cleaned with Ultrapure™ (herein ultrapure) water,ultrasonicated in ultrapure water, then allowed to dry thoroughly. Samples were scrapedto expose fresh surfaces, then powdered via agate mortar and pestle. Samples were thenpre-treated to isolate and purify carbonyl-apatite for analysis. Powdered samples wereplaced in a 15 mL centrifuge tube with 3 % pure NaOCl, ultrasonicated for approxi-mately 60 min, centrifuged, followed by removal of the 3 % NaOCl solution viapipette. The samples were subjected to three successive stages of ultrapure water rinses.Acetic acid (1.0 M, CH3COOH) was added to the sample, ultrasonicated for 1 h, andthen allowed to sit for 12 h. The sample was centrifuged and the acetic acid wasremoved via pipette, followed by three ultrapure water rinses, and then allowed to dry.The resulting powder was dissolved in approximately 10 % solution of ultrapure waterand 14 M nitric acid (HNO3) in a Teflon beaker. The samples were dried on a hotplate,and re-dissolved in 3.5 M nitric acid. The solution was then run through Eichron Sr-specific resin columns to isolate strontium. Samples were analyzed for 87Sr/86Sr on aMicromass Sector 54 thermal ionization mass spectrometer (TIMS) at the University ofArizona Geochronology Laboratory. NBS-987 standards were analyzed on a per runbasis for normalization, yielding an 87Sr/86Sr ratio of 0.710258±0.000021 (n=13). Allsample preparations and analyses were conducted by the primary and second author.
Acknowledgments We thank Larry Benson, W. Wills, John Driver, Wendy Bustard, and the Museum ofSouthwestern Biology and the NPS Chaco Museum Collection for samples or data used in our paper. We alsothank Scott P. Lehrich, Senior Regional Biologist for the National Wild Turkey Federation, for providingaccess to GIS data for modern turkey distributions and Selin Nugent for capturing the video of the Azerbaijanitethered turkey. Funding was provided by the Jacob K. Javits Fellowship, NSF IGERT in ArchaeologicalScience, the International Society of the P.E.O., William G. McGinnis Arid Lands Scholarship, Joe BenWheatResearch Award, and the Fred Plog Memorial Fellowship.
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsementby the U.S. Government.
References
Aasen, D.K. (now Rylander, D.K. Aasen) (1984). Pollen, macrofossil and charcoal analysis of basketmakercoprolites from Turkey Pen Ruin, Cedar Mesa, Utah. Unpublished M.S. thesis, Pullman: WashingtonState University.
Akins, N. J. (1985). Prehistoric faunal utilization in Chaco Canyon: Basketmaker III through Pueblo III. In F.J. Mathien (Ed.), Environment and subsistence of Chaco Canyon, New Mexico, publication in archae-ology 18E (pp. 305–446). New Mexico: National Park Service.
Akins, N. J. (1987). Faunal remains fromPuebloAlto. In F. J.Mathien&T. C.Windes (Eds.), Investigations at thePueblo Alto complex, Chaco Canyon, New Mexico, 1975–1979, Vol. 3. publications in archaeology 18F,Chaco Canyon studies. National park service (pp. 445–650). Santa Fe: U.S. Department of the Interior.
Allen, C. D., & Breshears, D. D. (1998). Drought-induced shift of a forest–woodland ecotone: rapid landscaperesponse to climate variation. Proceedings of the National Academy of Science, 95(25), 14839–14842.
Anderson, R. S., Jass, R. B., Toney, J. L., Allen, C. D., Ciseros-Dozal, L. M., Hess, M., Heikoop, J., &Fessenden, J. (2008). Development of mixed conifer forest in northern New Mexico, and its relationshipto Holocene environmental change. Quaternary Research, 69, 263–275.
Badenhorst, S., & Driver, J. C. (2009). Faunal changes in farming communities from Basketmaker II to PuebloIII (A.D. 1–1300) in the San Juan Basin of the American Southwest. Journal of Archaeological Science,36, 1832–1841.
Grimstead et al.
Author's personal copy

Barwick, L. H., & Speake, D. W. (1973). Seasonal movements and activities of wild turkey gobblers inAlabama. Proceedings of the National Wild Turkey Symposium, 2, 125–133.
Beacham, E. B., & Durand, S. R. (2007). Eggshell and the archaeological record: new insights into turkeyhusbandry in the American Southwest. Journal of Archaeological Science, 34, 1610–1621.
Benson, L. (2012). Development and application of methods used to source prehistoric Southwestern maize: areview. Journal of Archaeological Science, 39, 791–807.
Benson, L., Cordell, L., Vincent, K., Taylor, H., Stein, J., Farmer, G. L., & Futa, K. (2003). Ancient maizefrom chacoan great houses: where was it grown? Proceedings of the National Academy of Science, 100,13111–13115.
Benson, L., Stein, J. R., & Taylor, H. E. (2009). Possible sources of archaeological maize found in ChacoCanyon and Aztec Ruin, New Mexico. Journal of Archaeological Science, 36, 387–407.
Bentley, R. A. (2006). Strontium isotopes from the earth to the archaeological skeleton: a review. JournalArchaeological Method and Theory, 13, 135–187.
Best, T. L. (1996). Lepus californicus. Mammalian Species, 530, 1–10.Betancourt, J. L. (1990). Late quaternary biogeography of the Colorado plateau. In J. L. Betancourt, T. R. Van
Devender, & P. S. Martin (Eds.), Packrat middens: The last 40,000 years of biotic change (pp. 259–292).Tucson: University of Arizona Press.
Betancourt, J. L., & Van Devender, T. R. (1981). Holocene vegetation in Chaco Canyon, New Mexico.Science, 214, 658–660.
Betancourt, J. L., Van Devender, T. R., & Martin, P. S. (1983). Fossil packrat middens from Chaco Canyon,New Mexico: Cultural and ecological significance. In S. G. Wells, D. W. Love, & T. W. Gardner (Eds.),Chaco Canyon country, a field guide to the geomorphology, quaternary geology, paleoecology, andenvironmental geology of northwestern New Mexico (pp. 207–217). Albuquerque: AmericanGeomorphological Field Group.
Betancourt, J. L., Dean, J. S., & Hull, H. (1986). Prehistoric long-distance transport of construction beams,Chaco Canyon, New Mexico. American Antiquity, 51, 370–375.
Brown, D. E., & Lowe, C. H. (1980). Biotic communities of the Southwest, scale 1:1,000,000. Generaltechnical report of the rocky mountain forest and range experiment station. Washington D.C.: USDAForest Service.
Burt, W. H. (1943). Territoriality and home range concepts as applied to mammals. Journal of Mammalogy,24, 346–352.
Capo, R. C., Stewart, B. W., & Chadwick, O. A. (1998). Strontium isotopes as tracers of ecosystem processes:theory and methods. Geoderma, 82, 197–225.
Chapman, J. A., & Willner, G. R. (1978). Sylvilagus audubonii. Mammalian Species, 106, 1–4.Dasmann, R. F., & Taber, R. D. (1956). Behavior of Columbian black-tailed deer with reference to population
ecology. Journal of Mammology, 37(2), 143–164.Dickinson, T. G., & Garner, W. (1979). Home range and movements of desert mule deer in Southwestern
Texas. Proceedings of the Annual Conference of Southeastern Fish and Wildlife Agencies, 33,267–278.
Drake, B. L., Wills, W. H., Hamilton, M. I., & Dorshow, W. (2014). Strontium isotopes and the reconstructionof the Chaco regional system: evaluating uncertainty with Bayesian mixing models. PLoS ONE, 9(5),e95580. doi:10.1371/journal.pone.0095580.
Durand, K. R., & Durand, S. R. (2008). Animal bone from Salmon Ruins and other great houses: Faunalexploitation in the Chaco world. In P. F. Reed (Ed.), Chaco’s northern prodigies: Salmon, Aztec, and theascendancy of the middle San Juan region after AD 1100 (pp. 96–112). Salt Lake City: University of UtahPress.
Ellis, J. E., & Lewis, J. B. (1967). Mobility and annual range of wild turkeys in Missouri. The Journal ofWildlife Management, 31, 568–581.
English, N., Betancourt, J. L., Dean, J. S., & Quade, J. (2001). Strontium isotopes reveal distant sources ofarchitectural timber in Chaco Canyon, NewMexico. Proceedings of the National Academy of Science, 98,11891–11896.
Feranec, R. S., Hadly, E. A., & Paytan, A. (2007). Determining landscape use of Holocene mammals usingstrontium isotopes. Oecologia, 153, 943–950.
Fitch, H. S. (1947). Ecology of a cottontail rabbit (Sylvilagus audubonii) population in central California.California Fish and Game, 3, 159–184.
Fitzgerald, J. P., & Lechleitner, R. R. (1974). Observations on the biology of Gunnison’s prairie dog in CentralColorado. American Midland Naturalist, 92, 146–163.
Flannery, K. V. (1967). Vertebrate fauna and hunting practices. In D. S. Byers (Ed.), Prehistory of the tehuacanvalley, Vol 1: Environment and subsistence (pp. 132–177). Austin: Univ of Texas Press.
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

Flinders, J. T., & Hansen, R. M. (1972). Diets and habits of jackrabbits in northeastern Colorado. ColoradoState University, Range Science Department Science Series, 12, 1–29.
Flinders, J. T., & Hansen, R. M. (1975). Spring population responses of cottontails and jackrabbits to cattlegrazing shortgrass prairie. Journal Range Management, 28, 290–293.
Fothergill, B. (2012). The Bird of the next dawn: the husbandry, translocation, and transformation of theturkey. Ph.D. Dissertation, Leicester, UK: University of Leicester.
Fox, K. B., & Krausman, P. R. (1994). Fawning habitat of desert mule deer. Southwestern Naturalist, 39, 269–275.
French, N. R., McBride, R., & Detmer, J. (1965). Fertility and population density of the black-tailedjackrabbit. The Journal of Wildlife Management, 29, 14–26.
Frey, J. K., & Malaney, J. L. (2006). Snowshoe hare (Lepus americanus) and mountain cottontail (Sylvilagusnuttallii) biogeography at their southern range limit. Journal of Mammalogy, 87, 1175–1182.
Gallina, S., Mandujano, S., Bello, J., & Delfin, C. (1997). Home-range size of White-tailed Deer innortheastern Mexico. Proceedings of the 1997 Deer/Elk workshop, Rio Rico, Arizona, edited by J. C.deVos Jr., pp. 47–50. Phoenix: Arizona Game and Fish Department.
Gastwirth, J. L., Gel, Y. R., & Miao, W. (2009). The impact of Levene’s Test of equality of variance onstatistical theory and practice. Statistical Science, 24, 343–360.
Gill, R. B., Carpenter, L. H., Bartmann, R. M., Baker, D. L., & Schoonveld, G. G. (1983). Fecal analysis toestimate mule deer diets. The Journal of Wildlife Management, 47(4), 902–915.
Gillespie, W. B. (1991). Faunal remains from 29SJ 633. In F. J. Mathien (Ed.), Excavations at 29SJ 633: Theeleventh hour site Chaco Canyon, New Mexico, reports of the Chaco center 10 (pp. 243–315). Santa Fe:Branch of Cultural Research, National Park Service.
Graustein, W. C., & Armstrong, R. L. (1983). The use of87Sr/86Sr to measure atmospheric transport intoforested watershed. Science, 219, 2890–292.
Greenwood, P. J. (1980). Mating systems, philopatry and dispersal in birds and mammals. Animal Behaviour,28, 1140–1162.
Grimstead, D.N. (2011). Applications of evolutionary ecology and isotope geochemistry shed light on NorthAmerican prehistoric human behavior and regional procurement systems. Ph.D. Dissertation, Tucson:University of Arizona.
Hansen, R. M., & Flinders, J. T. (1969). Food habits of North American hares. Range Science DepartmentScience Series, 1, 1–18.
Harestad, A. S., & Bunnel, F. L. (1979). Home range and body weight—a reevaluation. Ecology,60, 389–402.
Haywood, D. D., Brown, R.L., Smith, R.H., & McCulloch, C.Y. (1987). Migration patterns and habitatutilization by Kaibab Mule Deer. Federal Aid Project W-78-R, WP2, Job 18. Phoenix, Arizona Game andFish Department.
Heffelfinger, J. (2006). Deer of the Southwest: A complete guide to the natural history, biology, andmanagement of Southwestern mule deer and white-tailed deer. College Station: Texas A&M UniversityPress.
Hoffman, D. M. (1968). Roosting sites and habits of Merriam’s Turkeys in Colorado. The Journal of WildlifeManagement, 32, 859–866.
Hoffman, D. M. (1991). Spring movements, roosting activities, and home-range characteristics of maleMerriam’s wild turkey. Southwestern Naturalist, 36, 332–337.
Hoppe, K. A., Koch, P. L., & Furutani, T. T. (2003). Assessing the preservation of biogenic strontium in fossilbones and tooth enamel. International Journal of Osteoarchaeology, 13, 20–28.
Horejsi, R. G., Haywood, D. D., & Smith, R. H. (1988). The effects of hunting of desert mule deerpopulations. Federal Aid Project W-78-R, WP2, Job 27. Phoenix: Arizona Game and Fish Department.
Ingles, L. G. (1941). Natural history observations on the Audubon cottontail. Journal of Mammalogy, 22,227–250.
Jimenez-Moreno, G., Fawcett, P. J., & Anderson, R. S. (2008). Millennial- and centennial-scale vegetation andclimate chance during the late Pleistocene and Holocene from Northern New Mexico (USA). QuaternaryScience Reviews, 27, 1442–1452.
Knott, B. (2010) Documenting domestication: molecular and palynological analysis of ancient turkeycoprolites from the American Southwest. Unpublished M.S. thesis, Pullman: Washington StateUniversity.
Knowles, C. (2002). Status of white-tailed and Gunnison’s prairie dogs. Washington: National WildlifeFederation.
Koerth, B. H., & Bryant, F. C. (1982). Home ranges of mule deer bucks in the Texas panhandle. PrairieNaturalist, 30, 579–587.
Grimstead et al.
Author's personal copy

Koerth, B. H., Sowell, B. F., Bryant, F. C., &Wiggers, E. P. (1985). Habitat relations of mule deer in the Texaspanhandle. Southwestern Naturalist, 30, 579–587.
Krausman, P. R. (1985). Impacts of the central Arizona project on desert mule deer and bighorn sheep, finalreport. Tucson: U. S. Bureau of Reclamation.
Lang, R., & Harris, A. H. (1984). The faunal remains from Arroyo Hondo Pueblo, New Mexico. Santa Fe:School of American Research Press.
Lawrence, R. K., Relyea, R. A., Demarais, S., & Lutz, R. S. (1994). Population dynamics and habitatpreferences of desert mule deer in the trans-Pecos, federal Aid project W-127-R, Job 94. Austin: TexasParks and Wildlife Department.
Lechleitner, R. R. (1958). Movements, density, and mortality in a black-tailed jackrabbit population. TheJournal of Wildlife Management, 22, 371–384.
Lekson, S. (2006). The archaeology of Chaco Canyon: An eleventh-century pueblo regional center. Santa Fe:School of American Research Press.
Lipe, W. D., Matson, R. G., & Kemp, B. M. (2011). New insights from old collections: Cedar Mesa, Utah,revisited. Southwestern Lore, 77, 103–111.
Logan, T. H. (1973). Seasonal behavior of Rio Grande wild turkeys in western Oklahoma. Proceedings of theSouth-eastern Association Game and Fish Communication, 26, 74–91.
Longhurst, W. (1944). Observations on the ecology of the Gunnison prairie dog in Colorado. Journal ofMammalogy, 25, 24–36.
Longhurst, W. M., Leopold, A. S., & Dasmann, R. F. (1952). A survey of California deer herds: Their rangesand management problems, game bulletin no. 6. Sacramento: California Department of Fish and Game.
MacDonald, D. (1963). Trapping and marking Merriam’s wild turkeys. Proceedings of the WesternAssociation of State Game Fish, 43, 196–201.
Maser, C., Maser, Z., & Molina, R. (1988). Small-mammal mycophagy in rangelands of central andsoutheastern Oregon. Journal Range Management, 41, 309–312.
Mathien, F. J. (Ed.). (1985). Environment and subsistence of Chaco canyon, New Mexico, publication inarchaeology 18E. National Park Service: New Mexico.
McCaffery, H., Tykot, R. H., Gore, K. D., & DeBoer, B. R. (2014). Stable isotope analysis of turkey(Meleagris gallopavo) diet from pueblo II and pueblo III sites, middle San Juan region, northwest NewMexico. American Antiquity, 79, 337–352.
Merriam, C. H. (1902). The prairie dog of the great plains. Yearbook of the Department Agriculture, 1901,257–270.
Mierau, G. W., & Schmidt, J. L. (1981). The mule deer of the Mesa Verde national park (Mesa Verde researchseries, paper no. 2). Mesa Verde National Park, Colorado: Mesa Verde Museum Association.
Mohammad, F., Pieper, R. D., & Beck, R. F. (1988). Seasonal food habits of blacktailed jackrabbits (Lepuscalifornicus) in southern New Mexico. Southwestern Naturalist, 33, 367–370.
Nagy, K. A., Shoemaker, V. H., & Costa, W. R. (1976). Water, electrolyte, and nitrogen budgets of jackrabbits(Lepus californicus) in the Mojave Desert. Physiological Zoology, 49, 351–363.
National Wild Turkey Federation (2012). 2011 wild turkey range. Turkey Country March/April, 122–123.Nickens, P. R. (1981). Pueblo III communities in transition: Environment and adaptation in Johnson Canyon.
Boulder: The Colorado Archaeological Society in cooperation with the University of Colorado Boulder.NRCS (Natural Resources Conservation Service). (1999). Wild turkey (Meleagris gallopavo). Fish and
Wildlife Habitat Management Leaflet, 12, 1–11.O’Brien, C. S., Waddel, R. B., Rosenstock, S. S., & Rabe, M. J. (2006). Wildlife use of water catchments in
Southwestern Arizona. Wildlife Society Bulletin, 34(3), 582–592.Orr, R. T. (1940). The rabbits of California. California Academy of Sciences Occasional Papers No. 19. 227 pp.Paisley, R.N., Conrad, P.J., Denk, D.D. & Kubisiak, J.F. (2000). Home range characteristics of eastern wild
turkey gobblers in Wisconsin’s Driftless Region. Wisconsin Department of Natural Resources ResearchManagement Findings, No. 43.
Petersen, K. L. (1994). A warm and wet little climatic optimum and a cold dry little ice age in the southernrocky mountains. Climate Change, 26, 243–269.
Petersen, K. L., & Mehringer, P. J. (1976). Postglacial timberline fluctuations, La Plata Mountains,Southwestern Colorado. Arctic and Alpine Research, 8(3), 275–288.
Porter, W. F. (1978). Home range dynamics of wild turkeys in southeastern Minnesota. The Journal of WildlifeManagement, 41, 434–437.
Ragotzkie, K. E., & Bailey, J. A. (1991). Desert mule deer use of grazed and ungrazed habitats. Journal RangeManagement, 44, 487–490.
Raught, R. W. (1967). White-tailed deer. In W. S. Hue (Ed.), New Mexico Wildlife Management (pp. 52–60).Santa Fe: New Mexico Department of Game and Fish.
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy

Rawlings, T. A., & Driver, J. C. (2010). Paleodiet of domestic turkey, shields pueblo (5MT3807), Colorado:isotopic analysis and its implications for care of a household domesticate. Journal of ArchaeologicalScience, 37, 2433–2441.
Reed, P. F. (2000). Foundations of Anasazi culture: The basketmaker pueblo transition. Salt Lake City:University of Utah Press.
Reed, P.F. (2006). Salmon Pueblo: Room by Room. In P.F. Reed (Ed.) Thirty-Five Years of ArchaeologicalResearch at Salmon Ruins, Vol. 1: Introduction, Architecture, Chronology, and Conclusions, Vol. 1 (pp.41–55)., Tucson: Center for Desert Archaeology & Bloomfield: Salmon Ruins Museum.
Relyea, R. A., Lawrence, R. K., & Demaris, S. (2000). Home range of desert mule deer: testing the body-sizeand habitat-productivity hypothesis. The Journal of Wildlife Management, 64, 146–153.
Reynolds, A. C., Betancourt, J. L., Quade, J., Patchett, P. J., Dean, J. S., & Stein, J. (2005). 87Sr/86Sr sourcingof ponderosa pine used in Anasazi great house construction at Chaco Canyon, New Mexico. Journal ofArchaeological Science, 32, 1061–1075.
Reynolds, A. C., Quade, J., & Betancourt, J. L. (2012). Strontium isotope and nutrient sourcing in a semi-aridwoodland. Geoderma, 189–190, 574–584.
Rodgers, K.J. (1978). Seasonal movements of mule deer on the Santa Rita experimental range. Master’sThesis, Department of Renewable and Natural Resources. Tucson: University of Arizona.
Rohn, A.H. (1971). Mug House, Mesa Verde National Park, Colorado. National Park Service ArchaeologicalResearch Series, 7D. Washington, D.C.: United States Department of the Interior, National Park Service.
Rosenstock, S. S., O’Brien, C. S., Waddell, R. B., & Rabe, M. J. (2004). Studies of wildlife waterdevelopments in Southwestern Arizona: Wildlife use, water quality, wildlife diseases, wildlife mortalities,and influences on native pollinators, technical guide bulletin no. 8. Phoenix: Arizona Game and FishDepartment, Research Branch.
Rue, L. L. R., III. (1988). The deer of north America. Danbury: Grolier Book Clubs Inc.Samuels, M., & Betancourt, J. L. (1982). Modeling the long-term effects of fuelwood harvests on pinyon-
juniper woodlands. Environmental Management, 6, 505–515.Schorger, A. W. (1961). An ancient Pueblo turkey. Auk, 78, 138–144.Schorger, A. W. (1966). The wild turkey. Norman: University of Oklahoma Press.Scott, V. E., & Boecker, E. L. (1975). Ecology of the Merriam’s wild turkey on the Fort Apache Indian
reservation. Proceedings of the National Wild Turkey Symposium, 3, 141–158.Sillen, A., & Kavanagh, M. (1982). Strontium and paleodietary research: a review. Physical Anthropology, 25,
67–69.Speller, C. F., Kemp, B. M., Wyatt, S. D., Monroe, C., Lipe, W. D., Amdt, U. M., & Yang, D. Y. (2010).
Ancient mitochondrial DNA analysis reveals complexity of indigenous North American turkey domes-tication. Proceedings of the National Academy of Science, 107, 2807–2812.
Spicer, R. L. (1959).Wild turkey in New Mexico: An evaluation of habitat development, bulletin no. 10. SantaFe: New Mexico Department of Game and Fish.
Swannack, J. D., Jr. (1969). Big juniper house, Mesa Verde national park, Colorado. Archaeological researchseries No. 7-C. Washington: U.S. Department of the Interior.
Swetnam, T. W., & Betancourt, J. A. (1998). Mesoscale disturbance and ecological response to decadal-scaleclimate variability in the American Southwest. Journal of Climate, 11, 3128–3147.
Taber, R. D., & Dasmann, R. F. (1958). The black-tailed deer of the chaparral: Its life history andmanagement in the north coast range of California (California department of fish and game bulletinNo. 8). Sacramento: California Department of Fish and Game.
Thornton et al. (2012). Earliest Mexican Turkeys (Meleagris gallopavo) in the Maya region: implications forpre-Hispanic animal trade and the timing of turkey domestication. PLoS ONE, 7(8): e42630; doi:10.1371/journal.pone.0042630.
Tierson, W. C., Mattfeld, G. F., Sage, R. W., & Behrend, D. F. (1985). Seasonal movements and home range ofwhite-tailed deer in the Adirondacks. The Journal of Wildlife Management, 49(3), 760–769.
Turkowski, F. J. (1975). Dietary adaptability of the desert cottontail. The Journal of Wildlife Management, 39,748–756.
Vorhies, C. T. (1945). Water requirements of desert animals in the Southwest. University of Arizona, Collegeof Agriculture, Agricultural Experiment Station, Technical Bulletin, 107, 485–525.
Vorhies, C. T., & Taylor, W. P. (1933). The life histories and ecology of jack rabbits, Lepus alleni and Lepuscalifornicus spp., in relation to grazing in Arizona. University of Arizona, College of Agriculture,Agricultural Experiment Station Technical Bulletin, 49, 471–587.
Wagner, D. M., Drickamer, L. C., Krpata, D. M., Allender, C. J., Van Pelt, W. E. C., & Keim, P. (2006).Persistence of Gunnison’s prairie dog colonies in Arizona, USA. Biological Conservation, 130, 331–339.
Grimstead et al.
Author's personal copy

Walker, E. A. (1951). Wild turkey studies in the divide areas of the Edwards plateau (Texas game, fish andoyster commission, federal Aid report, series 6). Austin: Texas Game, Fish and Oyster Commission.
Wallmo, O. C. (1981). Distribution and habits. In O. C. Wallmo (Ed.), Mule and black-tailed deer of NorthAmerica (pp. 1–26). Lincoln: Wildlife Management Institute and University of Nebraska Press.
Whitaker, J. O. (1996). National Audubon society field guide to North American mammals. New York: Knopf.Wigley, T. B., Sweeney, J. M., Garner, M. E., & Melchiors, M. A. (1986). Wild turkey home ranges in the
Ouachita Mountains. The Journal of Wildlife Management, 50, 540–544.Zwickel, F., Jones, G., & Brent, H. (1953). Movement of Columbian black-tailed deer in the Willapa hills area,
Washington. The Murrelt, 34, 41–46.
87Sr/86Sr indicate turkey domestication at Chaco Canyon, NM, USA
Author's personal copy
Related Documents











