ORIGINAL ARTICLE Red blood cells release factors with growth and survival bioactivities for normal and leukemic T cells Ricardo F Antunes 1,2 , Cla ´udia Branda ˜o 1 , Margarida Maia 1 and Fernando A Arosa 1,2,3 Human red blood cells are emerging as a cell type capable to regulate biological processes of neighboring cells. Hereby, we show that human red blood cell conditioned media contains bioactive factors that favor proliferation of normal activated T cells and leukemic Jurkat T cells, and therefore called erythrocyte-derived growth and survival factors. Flow cytometry and electron microscopy in parallel with bioactivity assays revealed that the erythrocyte factors are present in the vesicle-free supernatant, which contains up to 20 different proteins. The erythrocyte factors are thermosensitive and do not contain lipids. Native polyacrylamide gel electrophoresis followed by passive elution and mass spectrometry identification reduced the potential erythrocyte factors to hemoglobin and peroxiredoxin II. Two-dimensional differential gel electrophoresis of the erythrocyte factors revealed the presence of multiple hemoglobin oxy–deoxy states and peroxiredoxin II isoforms differing in their isoelectric point akin to the presence of b-globin chains. Our results show that red blood cells release protein factors with the capacity to sustain T-cell growth and survival. These factors may have an unforeseen role in sustaining malignant cell growth and survival in vivo. Immunology and Cell Biology (2011) 89, 111–121; doi:10.1038/icb.2010.60; published online 4 May 2010 Keywords: cell growth; factor; hemoglobin; red blood cells; survival; T cells Red blood cells (RBC) are emerging as a pleiotropic cell with the capacity to engage in reciprocal cross talk with nearby cells during their lifetime. The mounting evidence indicates that in addition to transporting oxygen and carbon dioxide, RBC also regulate vascular contractility, neutrophil apoptosis, T-cell growth and survival, protei- nase release by fibroblasts and interleukin (IL)-12 release by dendritic cells. 1–5 Recent studies have provided mechanistic insights to explain some of RBC pleiotropy. Thus, although inhibition of IL-12 release by dendritic cells is explained by the interaction of signal regulatory protein a with CD47 present on RBC, 5 vascular contractility is due to the action of nitric oxide release by hemoglobin (Hb). 2 The few studies investigating the mechanisms used by RBC to inhibit T-cell apoptosis and favor proliferation have shown that they are linked with upregu- lation of cytoprotective proteins and downregulation of oxidative stress. 6,7 One reasonable mechanism for activated T cells to increase their cell-cycle progression, while lowering activation-induced cell death to marginal levels, in response to the presence of RBC would be by means of interactions with RBC receptors or with RBC released factors that will trigger signals on T cells involved in cell-cycle and survival pathways. Although RBC were proposed to enhance T- and B-cell responses through CD58–CD2 interactions, 8 we have shown that molecular interactions between GPI-linked receptors present on RBC and their ligands present on T cells are not involved. 9,10 Conversely, it has been reported that RBC release vesicles containing erythrocyte proteins upon changes of the environment, such as pH lowering or ATP and calcium depletion. 11–15 Although RBC-derived vesicles (RBC-ves) are cleared from the circulation, during their lifetime they are putative carriers of bioactivities for other cells. 16,17 Although several studies have shown that dendritic cell-derived vesicles, or exosomes, can activate T cells, 18,19 the possibility that vesicles, or other factors, released by RBC may participate in the enhancement of cell-cycle and survival signaling pathways on activated T cells has never been addressed. In our previous studies, we observed that stimulation of human T cells in vitro in the presence of RBC resulted in a remarkable increase in proliferation regardless of whether the addition of RBC was carried out at the start of the culture or 24h later, suggesting that the growth and survival effects of RBC are exerted on pathways that have already been turned-on. 3 In this context, we have hypothesized that RBC may release factors that promote survival and proliferation of activated, but not quiescent T cells. RESULTS RBC bioactivities are present in conditioned medium We have previously shown that RBC sustain the growth and survival of peripheral blood T cells activated in vitro without the need of accessory cells or physical contact. 6,7,10 Considering that RBC release factors into the extracellular milieu during maturation or upon environmental changes in the culture conditions, 11–15 we wanted to Received 22 December 2009; revised 25 March 2010; accepted 29 March 2010; published online 4 May 2010 1 Lymphocyte Biology Group, IBMC–Instituto de Biologia Molecular e Celular, Porto, Portugal and 2 Instituto de Cie ˆncias Biome ´dicas Abel Salazar, Porto, Portugal 3 Current address: Instituto Superior de Cie ˆncias da Sau ´de Norte (ISCSN), CESPU, Gandra, Portugal. Correspondence: Dr FA Arosa, Centro de Investigac ¸a ˜o em Cie ˆncias da Sau ´de (CICS), Instituto Superior de Cie ˆncias da Sau ´de Norte, Rua Central de Gandra, 1317, Gandra 4585-116, Portugal. E-mail: [email protected] or [email protected] Immunology and Cell Biology (2011) 89, 111–121 & 2011 Australasian Society for Immunology Inc. All rights reserved 0818-9641/11 www.nature.com/icb

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Red blood cells release factors with growth and survivalbioactivities for normal and leukemic T cells
Ricardo F Antunes1,2, Claudia Brandao1, Margarida Maia1 and Fernando A Arosa1,2,3
Human red blood cells are emerging as a cell type capable to regulate biological processes of neighboring cells. Hereby, we
show that human red blood cell conditioned media contains bioactive factors that favor proliferation of normal activated T cells
and leukemic Jurkat T cells, and therefore called erythrocyte-derived growth and survival factors. Flow cytometry and electron
microscopy in parallel with bioactivity assays revealed that the erythrocyte factors are present in the vesicle-free supernatant,
which contains up to 20 different proteins. The erythrocyte factors are thermosensitive and do not contain lipids. Native
polyacrylamide gel electrophoresis followed by passive elution and mass spectrometry identification reduced the potential
erythrocyte factors to hemoglobin and peroxiredoxin II. Two-dimensional differential gel electrophoresis of the erythrocyte factors
revealed the presence of multiple hemoglobin oxy–deoxy states and peroxiredoxin II isoforms differing in their isoelectric point
akin to the presence of b-globin chains. Our results show that red blood cells release protein factors with the capacity to sustain
T-cell growth and survival. These factors may have an unforeseen role in sustaining malignant cell growth and survival in vivo.
Immunology and Cell Biology (2011) 89, 111–121; doi:10.1038/icb.2010.60; published online 4 May 2010
Keywords: cell growth; factor; hemoglobin; red blood cells; survival; T cells
Red blood cells (RBC) are emerging as a pleiotropic cell with thecapacity to engage in reciprocal cross talk with nearby cells duringtheir lifetime. The mounting evidence indicates that in addition totransporting oxygen and carbon dioxide, RBC also regulate vascularcontractility, neutrophil apoptosis, T-cell growth and survival, protei-nase release by fibroblasts and interleukin (IL)-12 release by dendriticcells.1–5 Recent studies have provided mechanistic insights to explainsome of RBC pleiotropy. Thus, although inhibition of IL-12 release bydendritic cells is explained by the interaction of signal regulatoryprotein a with CD47 present on RBC,5 vascular contractility is due tothe action of nitric oxide release by hemoglobin (Hb).2 The few studiesinvestigating the mechanisms used by RBC to inhibit T-cell apoptosisand favor proliferation have shown that they are linked with upregu-lation of cytoprotective proteins and downregulation of oxidativestress.6,7
One reasonable mechanism for activated T cells to increase theircell-cycle progression, while lowering activation-induced cell death tomarginal levels, in response to the presence of RBC would be by meansof interactions with RBC receptors or with RBC released factors thatwill trigger signals on T cells involved in cell-cycle and survivalpathways. Although RBC were proposed to enhance T- and B-cellresponses through CD58–CD2 interactions,8 we have shown thatmolecular interactions between GPI-linked receptors present onRBC and their ligands present on T cells are not involved.9,10
Conversely, it has been reported that RBC release vesicles containing
erythrocyte proteins upon changes of the environment, such as pHlowering or ATP and calcium depletion.11–15 Although RBC-derivedvesicles (RBC-ves) are cleared from the circulation, during theirlifetime they are putative carriers of bioactivities for other cells.16,17
Although several studies have shown that dendritic cell-derivedvesicles, or exosomes, can activate T cells,18,19 the possibility thatvesicles, or other factors, released by RBC may participate in theenhancement of cell-cycle and survival signaling pathways on activatedT cells has never been addressed.
In our previous studies, we observed that stimulation of humanT cells in vitro in the presence of RBC resulted in a remarkable increasein proliferation regardless of whether the addition of RBC was carriedout at the start of the culture or 24 h later, suggesting that the growthand survival effects of RBC are exerted on pathways that have alreadybeen turned-on.3 In this context, we have hypothesized that RBC mayrelease factors that promote survival and proliferation of activated, butnot quiescent T cells.
RESULTS
RBC bioactivities are present in conditioned mediumWe have previously shown that RBC sustain the growth and survivalof peripheral blood T cells activated in vitro without the need ofaccessory cells or physical contact.6,7,10 Considering that RBC releasefactors into the extracellular milieu during maturation or uponenvironmental changes in the culture conditions,11–15 we wanted to
Received 22 December 2009; revised 25 March 2010; accepted 29 March 2010; published online 4 May 2010
1Lymphocyte Biology Group, IBMC–Instituto de Biologia Molecular e Celular, Porto, Portugal and 2Instituto de Ciencias Biomedicas Abel Salazar, Porto, Portugal3Current address: Instituto Superior de Ciencias da Saude Norte (ISCSN), CESPU, Gandra, Portugal.Correspondence: Dr FA Arosa, Centro de Investigacao em Ciencias da Saude (CICS), Instituto Superior de Ciencias da Saude Norte, Rua Central de Gandra, 1317,Gandra 4585-116, Portugal.E-mail: [email protected] or [email protected]
Immunology and Cell Biology (2011) 89, 111–121& 2011 Australasian Society for Immunology Inc. All rights reserved 0818-9641/11
www.nature.com/icb
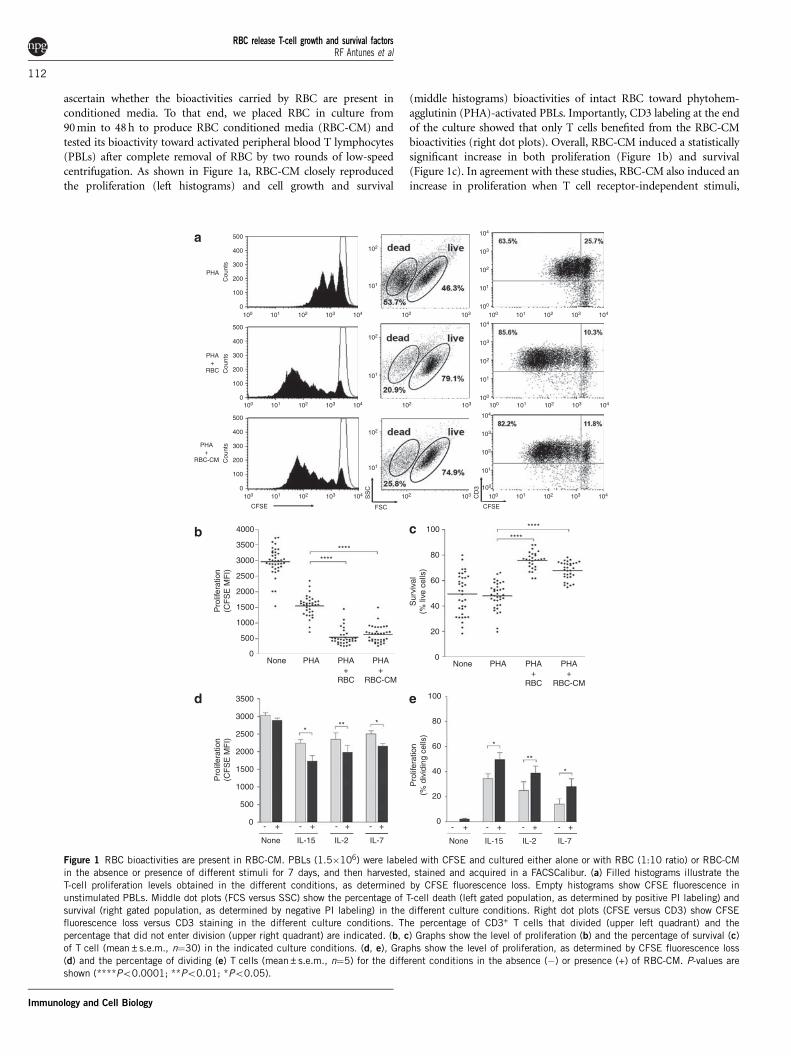
ascertain whether the bioactivities carried by RBC are present inconditioned media. To that end, we placed RBC in culture from90 min to 48 h to produce RBC conditioned media (RBC-CM) andtested its bioactivity toward activated peripheral blood T lymphocytes(PBLs) after complete removal of RBC by two rounds of low-speedcentrifugation. As shown in Figure 1a, RBC-CM closely reproducedthe proliferation (left histograms) and cell growth and survival
(middle histograms) bioactivities of intact RBC toward phytohem-agglutinin (PHA)-activated PBLs. Importantly, CD3 labeling at the endof the culture showed that only T cells benefited from the RBC-CMbioactivities (right dot plots). Overall, RBC-CM induced a statisticallysignificant increase in both proliferation (Figure 1b) and survival(Figure 1c). In agreement with these studies, RBC-CM also induced anincrease in proliferation when T cell receptor-independent stimuli,
PHA
PHA+
RBC
PHA+
RBC-CM
CFSE
********
Pro
lifer
atio
n(C
FS
E M
FI)
4000
3500
3000
2500
2000
1500
1000
500
0
Sur
viva
l(%
live
cel
ls)
100
80
60
40
20
0
Pro
lifer
atio
n(%
div
idin
g ce
lls)
100
80
60
40
20
0
*
- + - + - + - +
None IL-15 IL-2 IL-7
**
*
None PHA PHA+
RBC
PHA+
RBC-CM
********
*** *
Pro
lifer
atio
n(C
FS
E M
FI)
3500
3000
2500
2000
1500
1000
500
0 - + - + - + - +
None IL-15 IL-2 IL-7
None PHA PHA+
RBC
PHA+
RBC-CM
FSC CFSE
500a
b
d e
c
400
300
200
100
0
Cou
nts
500
400
300
200
100
0
Cou
nts
500
400
300
200
100
0
Cou
nts
Figure 1 RBC bioactivities are present in RBC-CM. PBLs (1.5�106) were labeled with CFSE and cultured either alone or with RBC (1:10 ratio) or RBC-CM
in the absence or presence of different stimuli for 7 days, and then harvested, stained and acquired in a FACSCalibur. (a) Filled histograms illustrate the
T-cell proliferation levels obtained in the different conditions, as determined by CFSE fluorescence loss. Empty histograms show CFSE fluorescence in
unstimulated PBLs. Middle dot plots (FCS versus SSC) show the percentage of T-cell death (left gated population, as determined by positive PI labeling) and
survival (right gated population, as determined by negative PI labeling) in the different culture conditions. Right dot plots (CFSE versus CD3) show CFSEfluorescence loss versus CD3 staining in the different culture conditions. The percentage of CD3+ T cells that divided (upper left quadrant) and the
percentage that did not enter division (upper right quadrant) are indicated. (b, c) Graphs show the level of proliferation (b) and the percentage of survival (c)
of T cell (mean±s.e.m., n¼30) in the indicated culture conditions. (d, e), Graphs show the level of proliferation, as determined by CFSE fluorescence loss
(d) and the percentage of dividing (e) T cells (mean±s.e.m., n¼5) for the different conditions in the absence (�) or presence (+) of RBC-CM. P-values are
shown (****Po0.0001; **Po0.01; *Po0.05).
RBC release T-cell growth and survival factorsRF Antunes et al
112
Immunology and Cell Biology
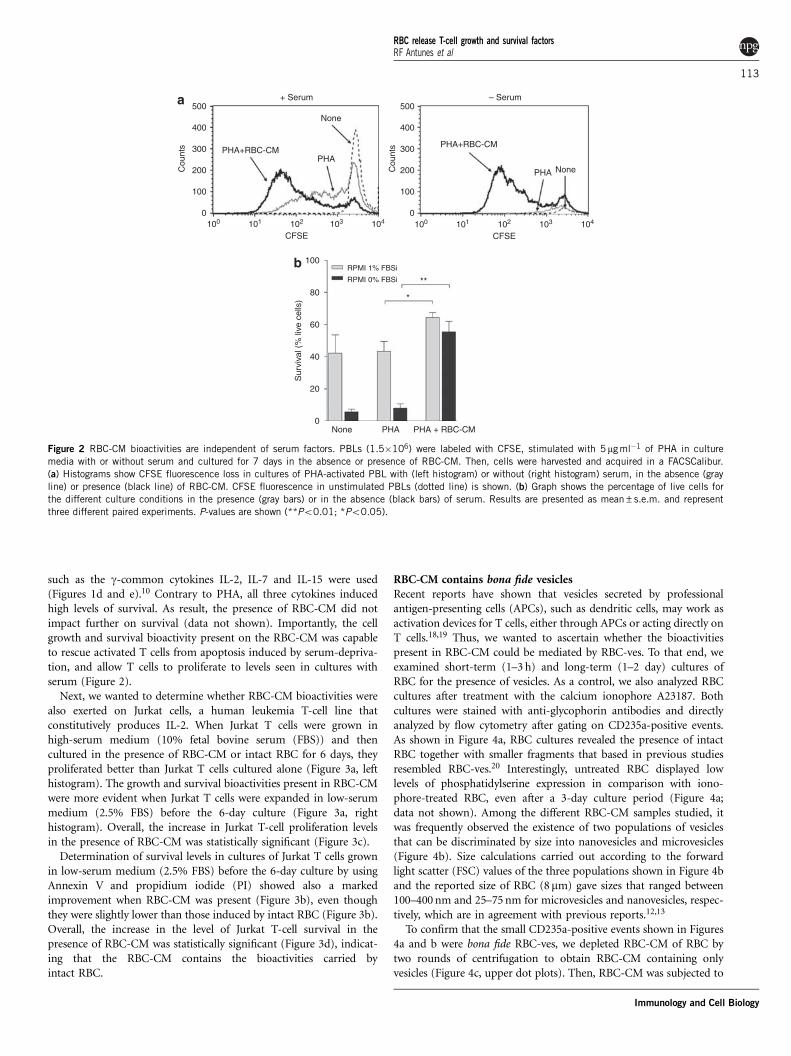
such as the g-common cytokines IL-2, IL-7 and IL-15 were used(Figures 1d and e).10 Contrary to PHA, all three cytokines inducedhigh levels of survival. As result, the presence of RBC-CM did notimpact further on survival (data not shown). Importantly, the cellgrowth and survival bioactivity present on the RBC-CM was capableto rescue activated T cells from apoptosis induced by serum-depriva-tion, and allow T cells to proliferate to levels seen in cultures withserum (Figure 2).
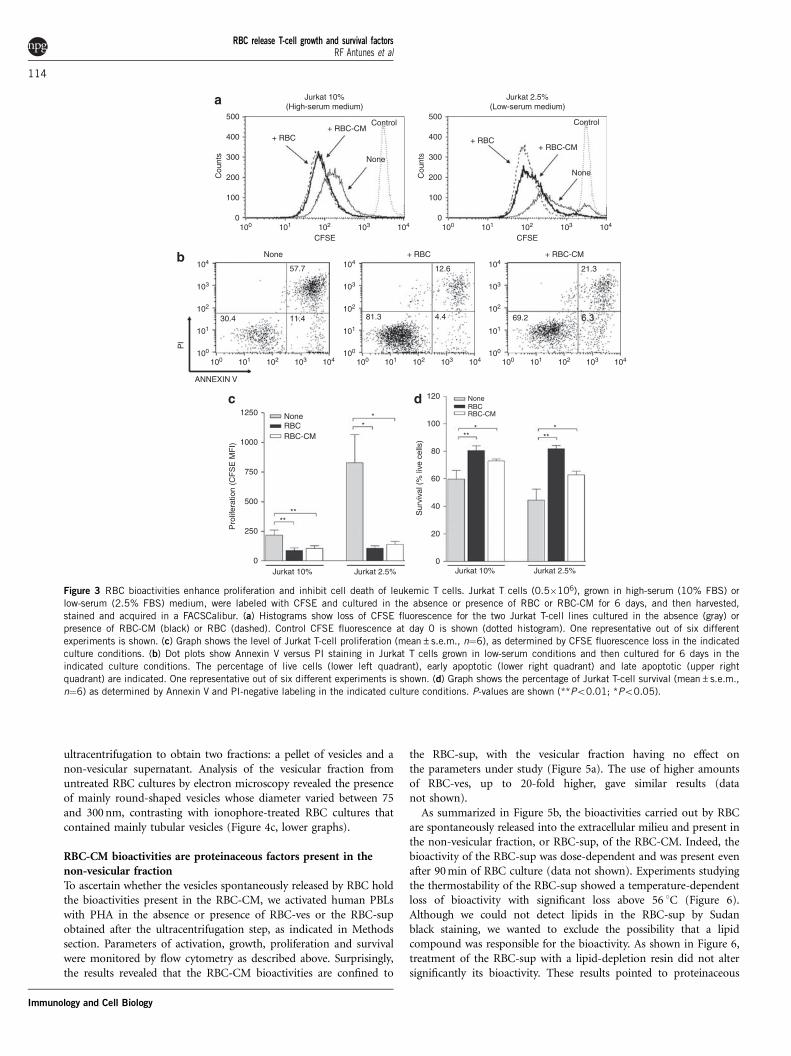
Next, we wanted to determine whether RBC-CM bioactivities werealso exerted on Jurkat cells, a human leukemia T-cell line thatconstitutively produces IL-2. When Jurkat T cells were grown inhigh-serum medium (10% fetal bovine serum (FBS)) and thencultured in the presence of RBC-CM or intact RBC for 6 days, theyproliferated better than Jurkat T cells cultured alone (Figure 3a, lefthistogram). The growth and survival bioactivities present in RBC-CMwere more evident when Jurkat T cells were expanded in low-serummedium (2.5% FBS) before the 6-day culture (Figure 3a, righthistogram). Overall, the increase in Jurkat T-cell proliferation levelsin the presence of RBC-CM was statistically significant (Figure 3c).
Determination of survival levels in cultures of Jurkat T cells grownin low-serum medium (2.5% FBS) before the 6-day culture by usingAnnexin V and propidium iodide (PI) showed also a markedimprovement when RBC-CM was present (Figure 3b), even thoughthey were slightly lower than those induced by intact RBC (Figure 3b).Overall, the increase in the level of Jurkat T-cell survival in thepresence of RBC-CM was statistically significant (Figure 3d), indicat-ing that the RBC-CM contains the bioactivities carried byintact RBC.
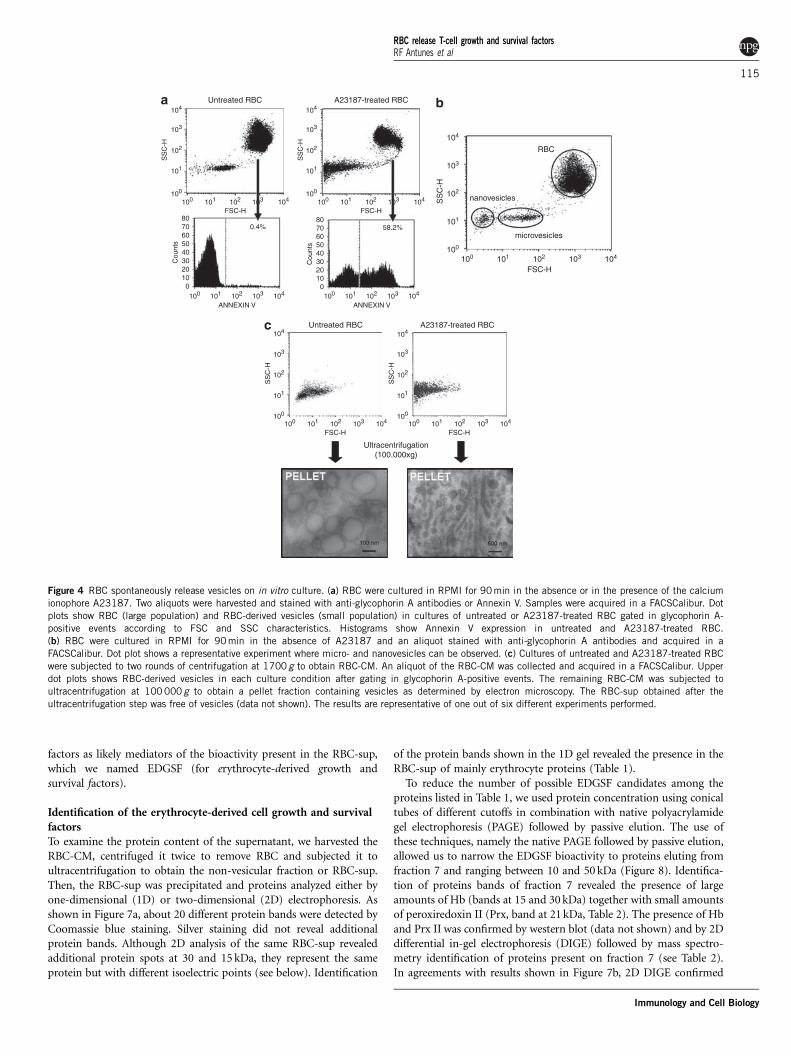
RBC-CM contains bona fide vesiclesRecent reports have shown that vesicles secreted by professionalantigen-presenting cells (APCs), such as dendritic cells, may work asactivation devices for T cells, either through APCs or acting directly onT cells.18,19 Thus, we wanted to ascertain whether the bioactivitiespresent in RBC-CM could be mediated by RBC-ves. To that end, weexamined short-term (1–3 h) and long-term (1–2 day) cultures ofRBC for the presence of vesicles. As a control, we also analyzed RBCcultures after treatment with the calcium ionophore A23187. Bothcultures were stained with anti-glycophorin antibodies and directlyanalyzed by flow cytometry after gating on CD235a-positive events.As shown in Figure 4a, RBC cultures revealed the presence of intactRBC together with smaller fragments that based in previous studiesresembled RBC-ves.20 Interestingly, untreated RBC displayed lowlevels of phosphatidylserine expression in comparison with iono-phore-treated RBC, even after a 3-day culture period (Figure 4a;data not shown). Among the different RBC-CM samples studied, itwas frequently observed the existence of two populations of vesiclesthat can be discriminated by size into nanovesicles and microvesicles(Figure 4b). Size calculations carried out according to the forwardlight scatter (FSC) values of the three populations shown in Figure 4band the reported size of RBC (8mm) gave sizes that ranged between100–400 nm and 25–75 nm for microvesicles and nanovesicles, respec-tively, which are in agreement with previous reports.12,13
To confirm that the small CD235a-positive events shown in Figures4a and b were bona fide RBC-ves, we depleted RBC-CM of RBC bytwo rounds of centrifugation to obtain RBC-CM containing onlyvesicles (Figure 4c, upper dot plots). Then, RBC-CM was subjected to
PHA+RBC-CM
Cou
nts
Cou
nts PHA+RBC-CM
None
None
+ Serum – Serum
PHA
PHA
500a
b
400
300
200
100
0
500
400
300
200
100
0
CFSE
RPMI 1% FBSi
RPMI 0% FBSi
*
**
Sur
viva
l (%
live
cel
ls)
100
80
60
40
20
0None PHA PHA + RBC-CM
100 101 102 103 104
CFSE100 101 102 103 104
Figure 2 RBC-CM bioactivities are independent of serum factors. PBLs (1.5�106) were labeled with CFSE, stimulated with 5mg ml�1 of PHA in culture
media with or without serum and cultured for 7 days in the absence or presence of RBC-CM. Then, cells were harvested and acquired in a FACSCalibur.
(a) Histograms show CFSE fluorescence loss in cultures of PHA-activated PBL with (left histogram) or without (right histogram) serum, in the absence (gray
line) or presence (black line) of RBC-CM. CFSE fluorescence in unstimulated PBLs (dotted line) is shown. (b) Graph shows the percentage of live cells for
the different culture conditions in the presence (gray bars) or in the absence (black bars) of serum. Results are presented as mean±s.e.m. and represent
three different paired experiments. P-values are shown (**Po0.01; *Po0.05).
RBC release T-cell growth and survival factorsRF Antunes et al
113
Immunology and Cell Biology
ultracentrifugation to obtain two fractions: a pellet of vesicles and anon-vesicular supernatant. Analysis of the vesicular fraction fromuntreated RBC cultures by electron microscopy revealed the presenceof mainly round-shaped vesicles whose diameter varied between 75and 300 nm, contrasting with ionophore-treated RBC cultures thatcontained mainly tubular vesicles (Figure 4c, lower graphs).
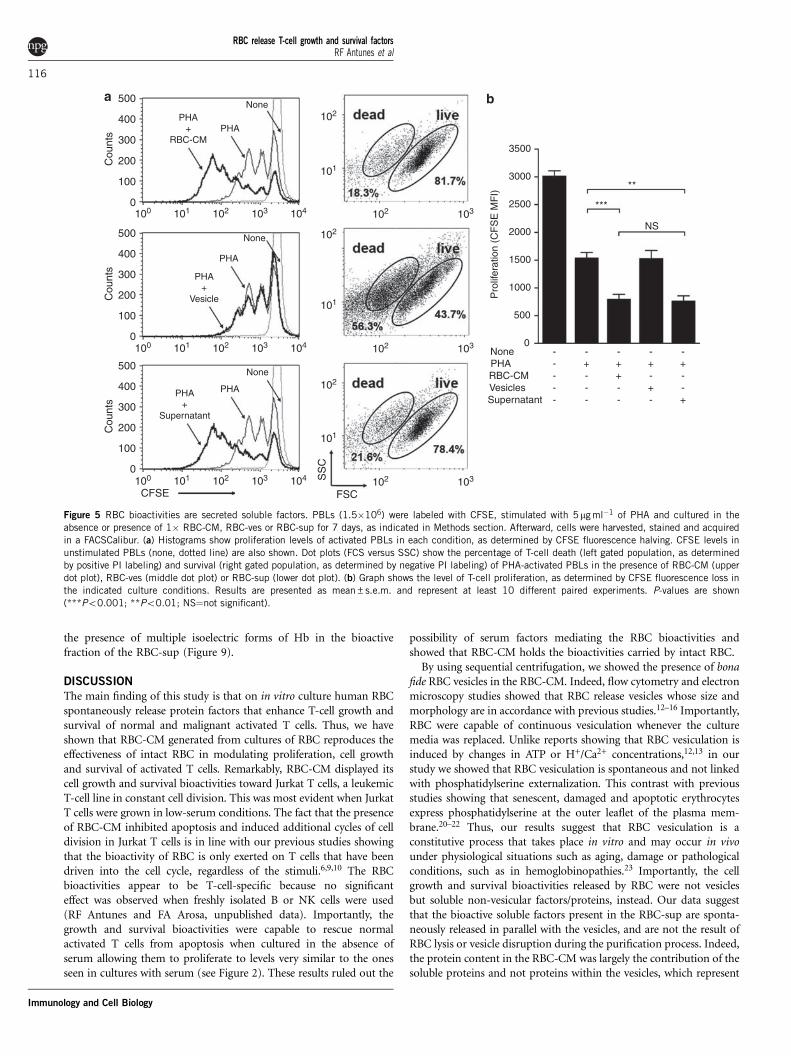
RBC-CM bioactivities are proteinaceous factors present in thenon-vesicular fractionTo ascertain whether the vesicles spontaneously released by RBC holdthe bioactivities present in the RBC-CM, we activated human PBLswith PHA in the absence or presence of RBC-ves or the RBC-supobtained after the ultracentrifugation step, as indicated in Methodssection. Parameters of activation, growth, proliferation and survivalwere monitored by flow cytometry as described above. Surprisingly,the results revealed that the RBC-CM bioactivities are confined to
the RBC-sup, with the vesicular fraction having no effect onthe parameters under study (Figure 5a). The use of higher amountsof RBC-ves, up to 20-fold higher, gave similar results (datanot shown).
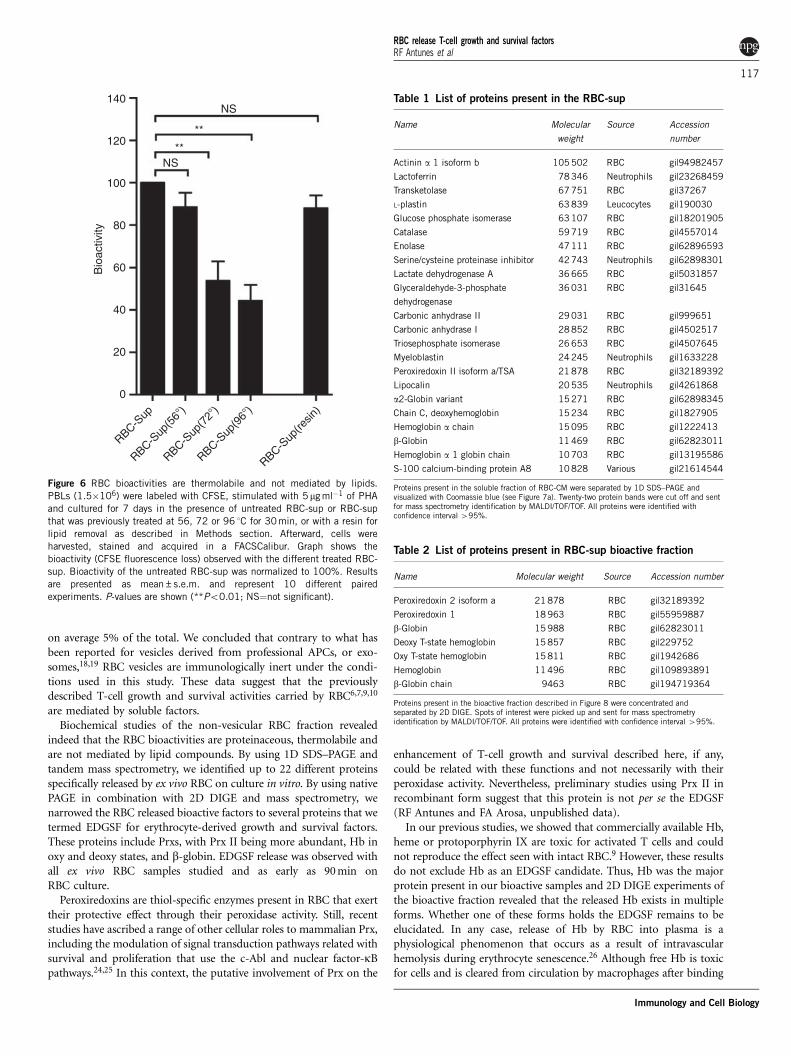
As summarized in Figure 5b, the bioactivities carried out by RBCare spontaneously released into the extracellular milieu and present inthe non-vesicular fraction, or RBC-sup, of the RBC-CM. Indeed, thebioactivity of the RBC-sup was dose-dependent and was present evenafter 90 min of RBC culture (data not shown). Experiments studyingthe thermostability of the RBC-sup showed a temperature-dependentloss of bioactivity with significant loss above 56 1C (Figure 6).Although we could not detect lipids in the RBC-sup by Sudanblack staining, we wanted to exclude the possibility that a lipidcompound was responsible for the bioactivity. As shown in Figure 6,treatment of the RBC-sup with a lipid-depletion resin did not altersignificantly its bioactivity. These results pointed to proteinaceous
Jurkat 10%(High-serum medium)
+ RBC
+ RBC
+ RBC
+ RBC-CM
+ RBC-CM
+ RBC-CM
Control Control
None
None
None
500
a
b
c d
400
300
200
100
0
None
* *** **
Sur
viva
l (%
live
cel
ls)
120
100
80
60
40
20
0
**
Pro
lifer
atio
n (C
FS
E M
FI)
****
NoneRBCRBC-CM
RBCRBC-CM1250
1000
750
500
250
0Jurkat 10% Jurkat 10%Jurkat 2.5% Jurkat 2.5%
ANNEXIN V
Pl
30.4
57.7
11.4 81.3
12.6
4.4 69.2
21.3
CFSE100 101 102 103 104
100 101 102 103 104 100 101 102 103 104 100 101 102 103 104
CFSE100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
Cou
nts
500
400
300
200
100
0
Cou
nts
Jurkat 2.5%(Low-serum medium)
Figure 3 RBC bioactivities enhance proliferation and inhibit cell death of leukemic T cells. Jurkat T cells (0.5�106), grown in high-serum (10% FBS) or
low-serum (2.5% FBS) medium, were labeled with CFSE and cultured in the absence or presence of RBC or RBC-CM for 6 days, and then harvested,
stained and acquired in a FACSCalibur. (a) Histograms show loss of CFSE fluorescence for the two Jurkat T-cell lines cultured in the absence (gray) or
presence of RBC-CM (black) or RBC (dashed). Control CFSE fluorescence at day 0 is shown (dotted histogram). One representative out of six different
experiments is shown. (c) Graph shows the level of Jurkat T-cell proliferation (mean±s.e.m., n¼6), as determined by CFSE fluorescence loss in the indicated
culture conditions. (b) Dot plots show Annexin V versus PI staining in Jurkat T cells grown in low-serum conditions and then cultured for 6 days in the
indicated culture conditions. The percentage of live cells (lower left quadrant), early apoptotic (lower right quadrant) and late apoptotic (upper right
quadrant) are indicated. One representative out of six different experiments is shown. (d) Graph shows the percentage of Jurkat T-cell survival (mean±s.e.m.,
n¼6) as determined by Annexin V and PI-negative labeling in the indicated culture conditions. P-values are shown (**Po0.01; *Po0.05).
RBC release T-cell growth and survival factorsRF Antunes et al
114
Immunology and Cell Biology
factors as likely mediators of the bioactivity present in the RBC-sup,which we named EDGSF (for erythrocyte-derived growth andsurvival factors).
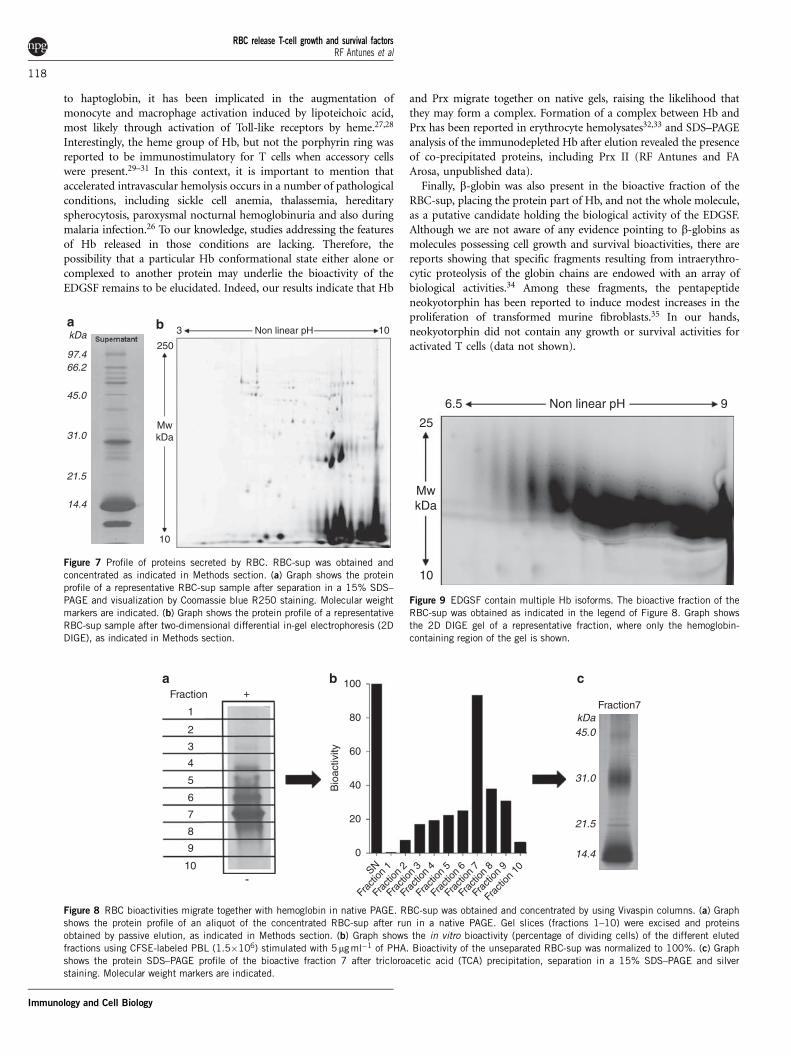
Identification of the erythrocyte-derived cell growth and survivalfactorsTo examine the protein content of the supernatant, we harvested theRBC-CM, centrifuged it twice to remove RBC and subjected it toultracentrifugation to obtain the non-vesicular fraction or RBC-sup.Then, the RBC-sup was precipitated and proteins analyzed either byone-dimensional (1D) or two-dimensional (2D) electrophoresis. Asshown in Figure 7a, about 20 different protein bands were detected byCoomassie blue staining. Silver staining did not reveal additionalprotein bands. Although 2D analysis of the same RBC-sup revealedadditional protein spots at 30 and 15 kDa, they represent the sameprotein but with different isoelectric points (see below). Identification
of the protein bands shown in the 1D gel revealed the presence in theRBC-sup of mainly erythrocyte proteins (Table 1).
To reduce the number of possible EDGSF candidates among theproteins listed in Table 1, we used protein concentration using conicaltubes of different cutoffs in combination with native polyacrylamidegel electrophoresis (PAGE) followed by passive elution. The use ofthese techniques, namely the native PAGE followed by passive elution,allowed us to narrow the EDGSF bioactivity to proteins eluting fromfraction 7 and ranging between 10 and 50 kDa (Figure 8). Identifica-tion of proteins bands of fraction 7 revealed the presence of largeamounts of Hb (bands at 15 and 30 kDa) together with small amountsof peroxiredoxin II (Prx, band at 21 kDa, Table 2). The presence of Hband Prx II was confirmed by western blot (data not shown) and by 2Ddifferential in-gel electrophoresis (DIGE) followed by mass spectro-metry identification of proteins present on fraction 7 (see Table 2).In agreements with results shown in Figure 7b, 2D DIGE confirmed
c Untreated RBC
Ultracentrifugation(100.000xg)
A23187-treated RBC
SS
C-H
104
103
102
101
100
SS
C-H
104
103
102
101
100
FSC-H104103102101100
FSC-H104103102101100
Untreated RBCa bA23187-treated RBC
RBC
nanovesicles
microvesicles
FSC-H
0.4% 58.2%
ANNEXIN V
SS
C-H
104
103
102
101
100
SS
C-H
104
103
102
101
100
SS
C-H
104
103
102
101
100
104103102101100
104103102101100
ANNEXIN V104103102101100
FSC-H104103102101100
FSC-H104103102101100
Cou
nts
8070605040302010
0
Cou
nts
8070605040302010
0
Figure 4 RBC spontaneously release vesicles on in vitro culture. (a) RBC were cultured in RPMI for 90 min in the absence or in the presence of the calcium
ionophore A23187. Two aliquots were harvested and stained with anti-glycophorin A antibodies or Annexin V. Samples were acquired in a FACSCalibur. Dot
plots show RBC (large population) and RBC-derived vesicles (small population) in cultures of untreated or A23187-treated RBC gated in glycophorin A-
positive events according to FSC and SSC characteristics. Histograms show Annexin V expression in untreated and A23187-treated RBC.
(b) RBC were cultured in RPMI for 90 min in the absence of A23187 and an aliquot stained with anti-glycophorin A antibodies and acquired in a
FACSCalibur. Dot plot shows a representative experiment where micro- and nanovesicles can be observed. (c) Cultures of untreated and A23187-treated RBC
were subjected to two rounds of centrifugation at 1700 g to obtain RBC-CM. An aliquot of the RBC-CM was collected and acquired in a FACSCalibur. Upper
dot plots shows RBC-derived vesicles in each culture condition after gating in glycophorin A-positive events. The remaining RBC-CM was subjected to
ultracentrifugation at 100 000 g to obtain a pellet fraction containing vesicles as determined by electron microscopy. The RBC-sup obtained after the
ultracentrifugation step was free of vesicles (data not shown). The results are representative of one out of six different experiments performed.
RBC release T-cell growth and survival factorsRF Antunes et al
115
Immunology and Cell Biology
the presence of multiple isoelectric forms of Hb in the bioactivefraction of the RBC-sup (Figure 9).
DISCUSSION
The main finding of this study is that on in vitro culture human RBCspontaneously release protein factors that enhance T-cell growth andsurvival of normal and malignant activated T cells. Thus, we haveshown that RBC-CM generated from cultures of RBC reproduces theeffectiveness of intact RBC in modulating proliferation, cell growthand survival of activated T cells. Remarkably, RBC-CM displayed itscell growth and survival bioactivities toward Jurkat T cells, a leukemicT-cell line in constant cell division. This was most evident when JurkatT cells were grown in low-serum conditions. The fact that the presenceof RBC-CM inhibited apoptosis and induced additional cycles of celldivision in Jurkat T cells is in line with our previous studies showingthat the bioactivity of RBC is only exerted on T cells that have beendriven into the cell cycle, regardless of the stimuli.6,9,10 The RBCbioactivities appear to be T-cell-specific because no significanteffect was observed when freshly isolated B or NK cells were used(RF Antunes and FA Arosa, unpublished data). Importantly, thegrowth and survival bioactivities were capable to rescue normalactivated T cells from apoptosis when cultured in the absence ofserum allowing them to proliferate to levels very similar to the onesseen in cultures with serum (see Figure 2). These results ruled out the
possibility of serum factors mediating the RBC bioactivities andshowed that RBC-CM holds the bioactivities carried by intact RBC.
By using sequential centrifugation, we showed the presence of bonafide RBC vesicles in the RBC-CM. Indeed, flow cytometry and electronmicroscopy studies showed that RBC release vesicles whose size andmorphology are in accordance with previous studies.12–16 Importantly,RBC were capable of continuous vesiculation whenever the culturemedia was replaced. Unlike reports showing that RBC vesiculation isinduced by changes in ATP or H+/Ca2+ concentrations,12,13 in ourstudy we showed that RBC vesiculation is spontaneous and not linkedwith phosphatidylserine externalization. This contrast with previousstudies showing that senescent, damaged and apoptotic erythrocytesexpress phosphatidylserine at the outer leaflet of the plasma mem-brane.20–22 Thus, our results suggest that RBC vesiculation is aconstitutive process that takes place in vitro and may occur in vivounder physiological situations such as aging, damage or pathologicalconditions, such as in hemoglobinopathies.23 Importantly, the cellgrowth and survival bioactivities released by RBC were not vesiclesbut soluble non-vesicular factors/proteins, instead. Our data suggestthat the bioactive soluble factors present in the RBC-sup are sponta-neously released in parallel with the vesicles, and are not the result ofRBC lysis or vesicle disruption during the purification process. Indeed,the protein content in the RBC-CM was largely the contribution of thesoluble proteins and not proteins within the vesicles, which represent
PHA+
RBC-CM
PHA+
Vesicle
PHA+
Supernatant
CFSE FSC
SS
C
PHA
PHA
PHA
None
None
None
**
***
NS
Pro
lifer
atio
n (C
FS
E M
FI)
3500
3000
2500
2000
1500
1000
500
0None -
-
--
-
-+
--
-
-+
-+
-
-+
+-
-
-+
--
+
PHARBC-CMVesiclesSupernatant
102
101
102
101
102
101
102 103
102 103
102 103
500
400
300
200
100
0100 101 102 103 104
100 101 102 103 104
100 101 102 103 104
Cou
nts
500
400
300
200
100
0
Cou
nts
500
400
300
200
100
0
Cou
nts
Figure 5 RBC bioactivities are secreted soluble factors. PBLs (1.5�106) were labeled with CFSE, stimulated with 5mg ml�1 of PHA and cultured in the
absence or presence of 1� RBC-CM, RBC-ves or RBC-sup for 7 days, as indicated in Methods section. Afterward, cells were harvested, stained and acquired
in a FACSCalibur. (a) Histograms show proliferation levels of activated PBLs in each condition, as determined by CFSE fluorescence halving. CFSE levels in
unstimulated PBLs (none, dotted line) are also shown. Dot plots (FCS versus SSC) show the percentage of T-cell death (left gated population, as determinedby positive PI labeling) and survival (right gated population, as determined by negative PI labeling) of PHA-activated PBLs in the presence of RBC-CM (upper
dot plot), RBC-ves (middle dot plot) or RBC-sup (lower dot plot). (b) Graph shows the level of T-cell proliferation, as determined by CFSE fluorescence loss in
the indicated culture conditions. Results are presented as mean±s.e.m. and represent at least 10 different paired experiments. P-values are shown
(***Po0.001; **Po0.01; NS¼not significant).
RBC release T-cell growth and survival factorsRF Antunes et al
116
Immunology and Cell Biology
on average 5% of the total. We concluded that contrary to what hasbeen reported for vesicles derived from professional APCs, or exo-somes,18,19 RBC vesicles are immunologically inert under the condi-tions used in this study. These data suggest that the previouslydescribed T-cell growth and survival activities carried by RBC6,7,9,10
are mediated by soluble factors.Biochemical studies of the non-vesicular RBC fraction revealed
indeed that the RBC bioactivities are proteinaceous, thermolabile andare not mediated by lipid compounds. By using 1D SDS–PAGE andtandem mass spectrometry, we identified up to 22 different proteinsspecifically released by ex vivo RBC on culture in vitro. By using nativePAGE in combination with 2D DIGE and mass spectrometry, wenarrowed the RBC released bioactive factors to several proteins that wetermed EDGSF for erythrocyte-derived growth and survival factors.These proteins include Prxs, with Prx II being more abundant, Hb inoxy and deoxy states, and b-globin. EDGSF release was observed withall ex vivo RBC samples studied and as early as 90 min onRBC culture.
Peroxiredoxins are thiol-specific enzymes present in RBC that exerttheir protective effect through their peroxidase activity. Still, recentstudies have ascribed a range of other cellular roles to mammalian Prx,including the modulation of signal transduction pathways related withsurvival and proliferation that use the c-Abl and nuclear factor-kBpathways.24,25 In this context, the putative involvement of Prx on the
enhancement of T-cell growth and survival described here, if any,could be related with these functions and not necessarily with theirperoxidase activity. Nevertheless, preliminary studies using Prx II inrecombinant form suggest that this protein is not per se the EDGSF(RF Antunes and FA Arosa, unpublished data).
In our previous studies, we showed that commercially available Hb,heme or protoporphyrin IX are toxic for activated T cells and couldnot reproduce the effect seen with intact RBC.9 However, these resultsdo not exclude Hb as an EDGSF candidate. Thus, Hb was the majorprotein present in our bioactive samples and 2D DIGE experiments ofthe bioactive fraction revealed that the released Hb exists in multipleforms. Whether one of these forms holds the EDGSF remains to beelucidated. In any case, release of Hb by RBC into plasma is aphysiological phenomenon that occurs as a result of intravascularhemolysis during erythrocyte senescence.26 Although free Hb is toxicfor cells and is cleared from circulation by macrophages after binding
NS
NS
**
**
Bio
activ
ity140
120
100
80
60
40
20
0
RBC-Sup
RBC-Sup
(56°
)
RBC-Sup
(72°
)
RBC-Sup
(96°
)
RBC-Sup
(resin
)
Figure 6 RBC bioactivities are thermolabile and not mediated by lipids.
PBLs (1.5�106) were labeled with CFSE, stimulated with 5mg ml�1 of PHA
and cultured for 7 days in the presence of untreated RBC-sup or RBC-sup
that was previously treated at 56, 72 or 96 1C for 30min, or with a resin for
lipid removal as described in Methods section. Afterward, cells were
harvested, stained and acquired in a FACSCalibur. Graph shows the
bioactivity (CFSE fluorescence loss) observed with the different treated RBC-
sup. Bioactivity of the untreated RBC-sup was normalized to 100%. Results
are presented as mean±s.e.m. and represent 10 different paired
experiments. P-values are shown (**Po0.01; NS¼not significant).
Table 1 List of proteins present in the RBC-sup
Name Molecular
weight
Source Accession
number
Actinin a 1 isoform b 105 502 RBC gi|94982457
Lactoferrin 78 346 Neutrophils gi|23268459
Transketolase 67 751 RBC gi|37267
L-plastin 63 839 Leucocytes gi|190030
Glucose phosphate isomerase 63 107 RBC gi|18201905
Catalase 59 719 RBC gi|4557014
Enolase 47 111 RBC gi|62896593
Serine/cysteine proteinase inhibitor 42 743 Neutrophils gi|62898301
Lactate dehydrogenase A 36 665 RBC gi|5031857
Glyceraldehyde-3-phosphate
dehydrogenase
36 031 RBC gi|31645
Carbonic anhydrase II 29 031 RBC gi|999651
Carbonic anhydrase I 28 852 RBC gi|4502517
Triosephosphate isomerase 26 653 RBC gi|4507645
Myeloblastin 24 245 Neutrophils gi|1633228
Peroxiredoxin II isoform a/TSA 21 878 RBC gi|32189392
Lipocalin 20 535 Neutrophils gi|4261868
a2-Globin variant 15 271 RBC gi|62898345
Chain C, deoxyhemoglobin 15 234 RBC gi|1827905
Hemoglobin a chain 15 095 RBC gi|1222413
b-Globin 11 469 RBC gi|62823011
Hemoglobin a 1 globin chain 10 703 RBC gi|13195586
S-100 calcium-binding protein A8 10 828 Various gi|21614544
Proteins present in the soluble fraction of RBC-CM were separated by 1D SDS–PAGE andvisualized with Coomassie blue (see Figure 7a). Twenty-two protein bands were cut off and sentfor mass spectrometry identification by MALDI/TOF/TOF. All proteins were identified withconfidence interval 495%.
Table 2 List of proteins present in RBC-sup bioactive fraction
Name Molecular weight Source Accession number
Peroxiredoxin 2 isoform a 21 878 RBC gi|32189392
Peroxiredoxin 1 18 963 RBC gi|55959887
b-Globin 15 988 RBC gi|62823011
Deoxy T-state hemoglobin 15 857 RBC gi|229752
Oxy T-state hemoglobin 15 811 RBC gi|1942686
Hemoglobin 11 496 RBC gi|109893891
b-Globin chain 9463 RBC gi|194719364
Proteins present in the bioactive fraction described in Figure 8 were concentrated andseparated by 2D DIGE. Spots of interest were picked up and sent for mass spectrometryidentification by MALDI/TOF/TOF. All proteins were identified with confidence interval 495%.
RBC release T-cell growth and survival factorsRF Antunes et al
117
Immunology and Cell Biology
to haptoglobin, it has been implicated in the augmentation ofmonocyte and macrophage activation induced by lipoteichoic acid,most likely through activation of Toll-like receptors by heme.27,28
Interestingly, the heme group of Hb, but not the porphyrin ring wasreported to be immunostimulatory for T cells when accessory cellswere present.29–31 In this context, it is important to mention thataccelerated intravascular hemolysis occurs in a number of pathologicalconditions, including sickle cell anemia, thalassemia, hereditaryspherocytosis, paroxysmal nocturnal hemoglobinuria and also duringmalaria infection.26 To our knowledge, studies addressing the featuresof Hb released in those conditions are lacking. Therefore, thepossibility that a particular Hb conformational state either alone orcomplexed to another protein may underlie the bioactivity of theEDGSF remains to be elucidated. Indeed, our results indicate that Hb
and Prx migrate together on native gels, raising the likelihood thatthey may form a complex. Formation of a complex between Hb andPrx has been reported in erythrocyte hemolysates32,33 and SDS–PAGEanalysis of the immunodepleted Hb after elution revealed the presenceof co-precipitated proteins, including Prx II (RF Antunes and FAArosa, unpublished data).
Finally, b-globin was also present in the bioactive fraction of theRBC-sup, placing the protein part of Hb, and not the whole molecule,as a putative candidate holding the biological activity of the EDGSF.Although we are not aware of any evidence pointing to b-globins asmolecules possessing cell growth and survival bioactivities, there arereports showing that specific fragments resulting from intraerythro-cytic proteolysis of the globin chains are endowed with an array ofbiological activities.34 Among these fragments, the pentapeptideneokyotorphin has been reported to induce modest increases in theproliferation of transformed murine fibroblasts.35 In our hands,neokyotorphin did not contain any growth or survival activities foractivated T cells (data not shown).
kDa250
3 Non linear pH 10
MwkDa
10
97.466.2
45.0
31.0
21.5
14.4
Figure 7 Profile of proteins secreted by RBC. RBC-sup was obtained and
concentrated as indicated in Methods section. (a) Graph shows the protein
profile of a representative RBC-sup sample after separation in a 15% SDS–
PAGE and visualization by Coomassie blue R250 staining. Molecular weight
markers are indicated. (b) Graph shows the protein profile of a representative
RBC-sup sample after two-dimensional differential in-gel electrophoresis (2D
DIGE), as indicated in Methods section.
Fractiona b c
+
-
1
2
Fraction7kDa45.0
31.0
21.5
14.4
Bio
activ
ity
100
80
60
40
20
0
SN
Frac
tion
1
Frac
tion
2
Frac
tion
3
Frac
tion
4
Frac
tion
5
Frac
tion
6
Frac
tion
7
Frac
tion
8
Frac
tion
9
Frac
tion
10
3
4
5
6
7
8
9
10
Figure 8 RBC bioactivities migrate together with hemoglobin in native PAGE. RBC-sup was obtained and concentrated by using Vivaspin columns. (a) Graph
shows the protein profile of an aliquot of the concentrated RBC-sup after run in a native PAGE. Gel slices (fractions 1–10) were excised and proteins
obtained by passive elution, as indicated in Methods section. (b) Graph shows the in vitro bioactivity (percentage of dividing cells) of the different eluted
fractions using CFSE-labeled PBL (1.5�106) stimulated with 5mg ml�1 of PHA. Bioactivity of the unseparated RBC-sup was normalized to 100%. (c) Graph
shows the protein SDS–PAGE profile of the bioactive fraction 7 after tricloroacetic acid (TCA) precipitation, separation in a 15% SDS–PAGE and silver
staining. Molecular weight markers are indicated.
6.5
25
MwkDa
10
Non linear pH 9
Figure 9 EDGSF contain multiple Hb isoforms. The bioactive fraction of the
RBC-sup was obtained as indicated in the legend of Figure 8. Graph shows
the 2D DIGE gel of a representative fraction, where only the hemoglobin-
containing region of the gel is shown.
RBC release T-cell growth and survival factorsRF Antunes et al
118
Immunology and Cell Biology

Overall, these studies have unveiled that RBC release novel factor(s)involved in the regulation of growth, proliferation and survival ofdividing T cells, placing RBC as a novel regulatory cell. In the contextof recent proteomic studies showing that some of the RBC releasedproteins identified in this study are present in plasma of patients withcancer,36,37 it is tempting to speculate that the EDGSF might have anunforeseen role in sustaining cell growth and survival of leukemiacells, and other tumor cells, in vivo.
METHODSReagents and mAbsPHA-P (from Phaseolus vulgaris), antibiotic-antimycotic solution (APS),
bovine serum albumin, PI, ionomycin (from Streptomyces conglobatus), phor-
bol dibutyrate, calcium ionophore A23187 and Sudan black B were from
Sigma-Aldrich (Madrid, Spain). Lymphoprep was from Nycomed (Oslo, Nor-
way). RPMI 1640 GlutaMAX, Hanks Balanced Salt Solution and inactivated
FBS (FBSi) were obtained from Gibco (Paisley, Scotland). Human rIL-2, rIL-7
and rIL-15 were obtained from R&D Systems (Minneapolis, MN, USA).
5- (and -6)-carboxyfluorescein diacetate succinimidyl ester (CFSE) and Annex-
in V-Alexa488 were purchased from Molecular Probes (Amsterdam, the
Netherlands). Anti-human glycophorin A PE-conjugated (clone AME-1,
CD235a-PE) was from ImmunoTools (Friesoythe, Germany). Anti-human
CD3-PE, CD4-APC, CD8-PE-Cy5 and rabbit anti-mouse-fluorochrome con-
jugated antibodies were from Dako (Glostrup, Denmark). Anti-human Hb
(clone GTX77484) was from GeneTex Inc. (Irvine, CA, USA). Cleanascite was
from Biotech Support Group (North Brunswick, NJ, USA). Molecular weight
rainbow markers were from Amersham Biosciences (Uppsala, Sweden).
CellsFresh peripheral blood mononuclear cells were obtained from buffy coats
after centrifugation over Lymphoprep. Peripheral blood mononuclear cells
were washed twice with Hanks Balanced Salt Solution and contaminating
RBC lysed in red cell lysis solution (10 mM Tris, 150 mM NH4Cl, pH 7.4)
for 10 min at 37 1C. RBC were collected from the pellet region after Lympho-
prep centrifugation, washed twice with Hanks Balanced Salt Solution and
diluted 1:10 in RPMI supplemented with 1% APS solution and stored
at 4 1C until use. RBC purity was assessed by flow cytometry and CD235a
labeling that revealed 499.5% CD235a+. Partially purified PBLs were obtained
after culture of peripheral blood mononuclear cells overnight in RPMI
supplemented with 1% APS solution and 5% FBSi. The recovered
non-adherent cell suspensions were routinely 485% CD3+ T cells and are
referred to as PBLs. In addition, the leukemic T-cell line Jurkat (clone E 6.1)
was used and maintained in two different culture conditions: a high-serum
medium (RPMI supplemented with 1% APS solution and 10% FBSi) and a
low-serum medium (RPMI supplemented with 1% APS solution and 2.5%
FBSi). In both conditions, culture medium was changed every 3 days. To
remove dead cells in the low-serum cultures, we centrifuged Jurkat T cells over
Lymphoprep (800 g, 12 min), collected live cells from the interface and after
washing twice with 1� phosphate-buffered saline (PBS) placed them again
in culture.
Production and isolation of RBC-CM, RBC-sup and RBC-vesTo obtain RBC-CM, we cultured RBC (60�106 per ml) in six-well plates in an
incubator at 37 1C, 5% CO2 and 99% humidity for 48 h (thereafter designated
by RBC-CM) in RPMI media supplemented with 1% APS solution. RBC-CM
was obtained after two sequential centrifugations at 1700 g for 10 min to
remove RBC. The absence of RBC as well as the presence of vesicles in the
RBC-CM was confirmed by flow cytometry after staining with CD235a, a
monoclonal antibody against glycophorin A. Vesicles revealed 99% CD235a+.
Annexin V staining of RBC using Ca2+-based staining buffer (see below) was
performed to evaluate the RBC apoptotic state in culture. Alternatively, and as a
control, RBC-CM was produced after treating RBC cultures with 5mM of
A23187, a Ca2+ ionophore, in RPMI media supplemented with 1 mM CaCl2.
A23187 treatment was stopped by addition of 5 mM EDTA. RBC-ves were
isolated from RBC-CM, previously depleted of RBC, after ultracentrifugation at
100 000�g for 2 h, at 4 1C in a Sorvall Ultra Pro80 centrifuge with a T1270
angular rotor. After centrifugation the non-vesicular supernatant (RBC-sup)
and the pellet containing the vesicles (RBC-ves) were collected, quantitated
using the BCA protein assay kit (Pierce, Rockford, IL, USA) and either used
immediately for bioactivity assays in vitro or frozen at �70 1C for further
studies. On average, the RBC-sup and the RBC-ves produced by 60�106 RBC
contained approximately 20 and 1mg ml�1 of protein, respectively, which agrees
with the amount of protein in the bulk RBC-CM (that ranges between 18 and
22mg ml�1). Absence of vesicles in the RBC-sup was confirmed by flow
cytometry.
CFSE labelingTen million PBLs were labeled with CFSE at a final concentration of 5mM for
10 min at 37 1C, with occasional mixing. Then, cells were washed twice with
PBS/20% FBSi and resuspended in culture media. Analysis of cells immediately
after CFSE labeling indicated a labeling efficiency higher than 99%.
RBC-sup treatmentsRBC-sup was routinely concentrated using Vivaspin 20 ultrafiltration spin
columns with a 3-kDa molecular weight cutoff (Sartorius, Goettingen,
Germany) and quantitated. In the in vitro assays, the concentrated RBC-sup
was diluted to 1� to maintain the original concentration of the RBC-CM. For
thermostability studies, we boiled concentrated RBC-sup at 56, 72 or 96 1C for
30 min in a heat block. The RBC-sup was centrifuged at 11 000 g before the
in vitro bioactivity assays. For lipid depletion, we added Cleanascite to the RBC-
sup in a ratio 1:4 and incubated the mixture first in a rotator at room
temperature for 10 min, followed by a further incubation at 4 1C for 30 min,
following manufacturer’s instructions. Then, the mixture was centrifuged to
remove the resin and the RBC-sup collected and concentrated as indicated
above before the in vitro bioactivity assays. Differential concentration of RBC-
sup proteins was carried out by using Vivaspin 20 ultrafiltration spin columns
with 30 and 50 molecular weight cutoff (Sartorius). Recovery of RBC-sup from
the upper and lower fraction after centrifugation allowed obtaining four
different fractions that were subsequently assayed for bioactivity in vitro.
Culture conditionsPBLs (1.5�106) and Jurkat T cells (0.5�106) were cultured in six-well plates, in
a final volume of 5 ml for up to 7 days in an incubator at 37 1C, 5% CO2 and
99% humidity. PBLs were either left unstimulated or stimulated with PHA-P
(5mg ml�1) and either IL-2, IL-7 or IL-15, all at 10 ng ml�1. Jurkat T cells were
left unstimulated. PBLs and Jurkat T cells were cultured in the absence or
presence of autologous RBC at a PBL/RBC ratio of 1:10 and 1:100, respectively,
or in the presence of 1� RBC-CM, RBC-ves or RBC-sup. Culture media was
RPMI 1640 and 1% FBSi. In some experiments, PBLs were cultured in the
absence of serum in RMPI 1640 GlutaMAX supplemented with 1% APS
solution. At the end of the culture, PBLs were harvested, washed and acquired
in a FACSCalibur (Becton Dickinson, Mountain View, CA, USA). For each
sample, 50 000 events were acquired using FSC/side light scatter (SSC)
characteristics and analyzed using CellQuest or FlowJo software (Becton
Dickinson, Mountain View, CA, USA).
Flow cytometry determinationsCell stainings were normally performed at 4 1C for 30 min in 1� PBS or
staining buffer (PBS, 0.2% bovine serum albumin, 0.1% NaN3) in 96-well
round-bottom plates (Greiner Bio One, Frickenhausen, Germany). In cultures
that received RBC, erythrocytes were first lysed with red cell lysis solution.
Irrelevant mAbs were used as negative controls to define background staining.
T-cell death and survival were determined by two methods: (1) a decrease in
cell size according to FSC/SSC parameters and (2) double Annexin V and PI
staining, for 15 min at room temperature in the dark using Ca2+-based staining
buffer (10 mM HEPES/140 mM NaCl/2.5 mM CaCl2). T-cell activation and
proliferation were studied by two methods: (1) determination of blast trans-
formation according to FCS/SSC parameters and (2) CFSE fluorescence loss.
Rounds of cell divisions were determined by sequential halving of CFSE-
fluorescence intensity.
RBC release T-cell growth and survival factorsRF Antunes et al
119
Immunology and Cell Biology

Transmission electron microscopyRBC-ves were fixed in a solution of 1.25% glutaraldehyde/4% paraformalde-
hyde and post-fixed in osmium 1% in sodium cacodylate. After washing with
H2O, we fixed the pellets in uranyl acetate 1% and placed them overnight in
glutaraldehyde 2.5%. After dehydration, we embedded the samples in EPON.
Thin sections were mounted on grids, stained with lead citrate and post-stained
with uranyl acetate. Images were acquired with a Zeiss 902A (Oberkochen,
West Germany).
SDS–PAGE, native PAGE and passive elutionThe RBC-sup was precipitated with 10% tricloroacetic acid for 30 min on ice,
centrifuged at 11 000 g for 15 min and then washed in cold acetone overnight at
�20 1C. Samples were centrifuged again at 11 000 g for 15 min and resuspended
in sample buffer (25% Tris (pH 6.8), 10% SDS, 0.5% bromophenol blue, 10%
glycerol), heated at 96 1C for 10 min and quantitated by the BCA protein assay
kit (Pierce). Aliquots corresponding to 20–25mg of protein were resolved in
15% SDS–PAGE under reducing conditions. Gels were stained with Coomassie
brilliant blue R250 (Sigma-Aldrich) to visualize protein bands. In some
experiments, gels were also stained using silver staining. For native PAGE
studies, we diluted aliquots of the precipitated RBC-sup in sample buffer (0.5 M
Tris (pH 8.0), 0.1 M DTT, 50% saccharose/10� bromophenol blue) and
resolved them by 6% native PAGE in a 4 1C room. For visualization of proteins,
we stained gels with Coomassie blue solution or silver staining. For bioactivity
assays, we excised native gel fractions, washed them with 1� PBS and placed
them in eppendorfs containing 1 ml of 1� PBS. Proteins were allowed to
passively elute out of the gel slices for 24 h at 4 1C. Afterward, gel slices were
removed and eluted proteins concentrated using Vivaspin 500 spin columns
with 3 molecular weight cutoff (Sartorius). Concentrated proteins were washed
several times on Vivaspin 500 spin columns using RPMI media supplemented
with 1% APS and stored at �70 1C.
2D DIGE and mass spectrometry analysis2D-DIGE was performed by Applied Biomics (Hayward, CA, USA). Briefly,
aliquots of concentrated RBC-sup proteins were covalently linked to CyDye
and run on first dimension isoelectric focusing, followed by a second dimen-
sion SDS–PAGE. Image analysis was performed using DeCyder software, and
spots of interest were picked and analyzed by mass spectrometry (MALDI/
TOF/TOF). Further protein identification was performed after 1D electrophor-
esis. Briefly, aliquots of concentrated RBC-sup proteins were run on 15% SDS–
PAGE followed by Coomassie brilliant blue R250 (Sigma-Aldrich) and protein
bands excised. Protein identification (MALDI/TOF/TOF) was performed by
Alphalyse (Odense, Denmark).
Statistical analysisStatistical analysis was performed using Excel or GraphPad Prism 5 software
GraphPad Software (San Diego, CA, USA). Student’s t-test was used to evaluate
the significance of the differences between group means. Statistical significance
was defined as Po0.05.
CONFLICT OF INTERESTThe authors declare no conflict of interest.
ACKNOWLEDGEMENTSWe thank Instituto Portugues do Sangue (Porto, Portugal) for providing buffy
coats used in this study. We also thank Jorge Azevedo and Pedro Pereira for
helpful comments during initial part of this work. We are indebted to Frederico
Silva, Rui Fernandes and Paula Sampaio for expert technical assistance. This
work was funded by Grant APCL2006-30.1.AP/MJ from Associacao Portuguesa
Contra a Leucemia (APCL, Portugal). RFA was funded by a PhD fellowship
from Fundacao para a Ciencia e a Tecnologia (SFRH/BD/24524/2005).
1 Pawloski JR, Hess DT, Stamler JS. Export by red blood cells of nitric oxide bioactivity.Nature 2001; 409: 622–626.
2 Aoshiba K, Nakajima Y, Yasui S, Tamaoki J, Nagai A. Red blood cells inhibit apoptosisof human neutrophils. Blood 1999; 93: 4006–4010.
3 Arosa FA, Pereira CF, Fonseca AM. Red blood cells as modulators of T cell growth andsurvival. Curr Pharm Des 2004; 10: 191–201.
4 Fredriksson K, Liu XD, Lundahl J, Klominek J, Rennard SI, Skold CM. Red blood cellsincrease secretion of matrix metalloproteinases from human lung fibroblasts in vitro.Am J Physiol Lung Cell Mol Physiol 2006; 290: L326–L333.
5 Schakel K, von Kietzell M, Hansel A, Ebling A, Schulze L, Haase M et al. Human 6-sulfo LacNAc-expressing dendritic cells are principal producers of early interleukin-12and are controlled by erythrocytes. Immunity 2006; 24: 767–777.
6 Fonseca AM, Porto G, Uchida K, Arosa FA. Red blood cells inhibit activation-inducedcell death and oxidative stress in human peripheral blood T lymphocytes. Blood 2001;97: 3152–3160.
7 Fonseca AM, Pereira CF, Porto G, Arosa FA. Red blood cells upregulate cytoprotectiveproteins and the labile iron pool in dividing human T cells despite a reduction inoxidative stress. Free Radic Biol Med 2003; 35: 1404–1416.
8 Virella G, Rugeles MT, Hyman B, La Via M, Goust JM, Frankis M et al. The interaction ofCD2 with its LFA-3 ligand expressed by autologous erythrocytes results in enhancementof B cell responses. Cell Immunol 1988; 116: 308–319.
9 Fonseca AM, Pereira CF, Porto G, Arosa FA. Red blood cells promote survival and cellcycle progression of human peripheral blood T cells independently of CD58/LFA-3 andheme compounds. Cell Immunol 2003; 224: 17–28.
10 Antunes RF, Brandao C, Carvalho G, Girao C, Arosa FA. Red blood cells carry out T cellgrowth and survival bioactivities that are sensitive to cyclosporine A. Cell Mol Life Sci2009; 66: 3387–3398.
11 Gros M, Vrhovec S, Brumen M, Svetina S, Zeks B. Low pH induced shape changesand vesiculation of human erythrocytes. Gen Physiol Biophys 1996; 15:145–163.
12 Allan D, Thomas P. Ca2+-induced biochemical changes in human erythrocytes andtheir relation to microvesiculation. Biochem J 1981; 198: 433–440.
13 Salzer U, Hinterdorfer P, Hunger U, Borken C, Prohaska R. Ca(++)-dependent vesiclerelease from erythrocytes involves stomatin-specific lipid rafts, synexin (annexin VII),and sorcin. Blood 2002; 99: 2569–2577.
14 Turner EJ, Jarvis HG, Chetty MC, Landon G, Rowley PS, Ho MM et al. ATP-dependentvesiculation in red cell membranes from different hereditary stomatocytosis variants. BrJ Haematol 2003; 120: 894–902.
15 Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation duringreticulocyte maturation. Association of plasma membrane activities with releasedvesicles (exosomes). J Biol Chem 1987; 262: 9412–9420.
16 Willekens FL, Werre JM, Kruijt JK, Roerdinkholder-Stoelwinder B, Groenen-Dopp YA,van den Bos AG et al. Liver Kupffer cells rapidly remove red blood cell-derivedvesicles from the circulation by scavenger receptors. Blood 2005; 105:2141–2145.
17 Denzer K, Kleijmeer MJ, Heijnen HF, Stoorvogel W, Geuze HJ. Exosome: from internalvesicle of the multivesicular body to intercellular signaling device. J Cell Sci 2000; 113
(Pt 19): 3365–3374.18 Thery C, Duban L, Segura E, Veron P, Lantz O, Amigorena S. Indirect activation of
naive CD4+ T cells by dendritic cell-derived exosomes. Nat Immunol 2002; 3:1156–1162.
19 Kovar M, Boyman O, Shen X, Hwang I, Kohler R, Sprent J. Direct stimulation of T cellsby membrane vesicles from antigen-presenting cells. Proc Natl Acad Sci USA 2006;103: 11671–11676.
20 Bevers EM, Wiedmer T, Comfurius P, Shattil SJ, Weiss HJ, Zwaal RF et al. DefectiveCa(2+)-induced microvesiculation and deficient expression of procoagulant activity inerythrocytes from a patient with a bleeding disorder: a study of the red blood cells ofScott syndrome. Blood 1992; 79: 380–388.
21 Connor J, Pak CC, Schroit AJ. Exposure of phosphatidylserine in the outer leaflet ofhuman red blood cells. Relationship to cell density, cell age, and clearance bymononuclear cells. J Biol Chem 1994; 269: 2399–2404.
22 Boas FE, Forman L, Beutler E. Phosphatidylserine exposure and red cell viabilityin red cell aging and in hemolytic anemia. Proc Natl Acad Sci USA 1998; 95:3077–3081.
23 Westerman M, Pizzey A, Hirschman J, Cerino M, Weil-Weiner Y, Ramotar Pet al. Microvesicles in haemoglobinopathies offer insights into mechanisms ofhypercoagulability, haemolysis and the effects of therapy. Br J Haematol 2008; 142:126–135.
24 Butterfield LH, Merino A, Golub SH, Shau H. From cytoprotection to tumor suppres-sion: the multifactorial role of peroxiredoxins. Antioxid Redox Signal 1999; 1:385–402.
25 Wood ZA, Schroder E, Robin Harris J, Poole LB. Structure, mechanism and regulationof peroxiredoxins. Trends Biochem Sci 2003; 28: 32–40.
26 Bunn HF, Forget BG. Hemoglobin: Molecular, Genetic and Clinical Aspects. WBSaunders: Philadelphia, PA, 1986.
27 Hasty DL, Meron-Sudai S, Cox KH, Nagorna T, Ruiz-Bustos E, Losi E et al. Monocyteand macrophage activation by lipoteichoic acid is independent of alanine and ispotentiated by hemoglobin. J Immunol 2006; 176: 5567–5576.
28 Figueiredo RT, Fernandez PL, Mourao-Sa DS, Porto BN, Dutra FF, Alves LS et al.Characterization of heme as activator of Toll-like receptor 4. J Biol Chem 2007; 282:20221–20229.
29 Stenzel KH, Rubin AL, Novogrodsky A. Mitogenic and co-mitogenic properties ofhemin. J Immunol 1981; 127: 2469–2473.
30 Novogrodsky A, Suthanthiran M, Stenzel KH. Immune stimulatory properties ofmetalloporphyrins. J Immunol 1989; 143: 3981–3987.
RBC release T-cell growth and survival factorsRF Antunes et al
120
Immunology and Cell Biology

31 Lander HM, Levine DM, Novogrodsky A. Haemin enhancement of glucosetransport in human lymphocytes: stimulation of protein tyrosine phosphataseand activation of p56lck tyrosine kinase. Biochem J 1993; 291(Pt 1):281–287.
32 Stuhlmeier KM, Kao JJ, Wallbrandt P, Lindberg M, Hammarstrom B, Broell H et al.Antioxidant protein 2 prevents methemoglobin formation in erythrocyte hemolysates.Eur J Biochem 2003; 270: 334–341.
33 Babusiak M, Man P, Sutak R, Petrak J, Vyoral D. Identification of heme binding proteincomplexes in murine erythroleukemic cells: study by a novel two-dimensional nativeseparation—liquid chromatography and electrophoresis. Proteomics 2005; 5:340–350.
34 Filippova MM, Khachin DP, Sazonova OV, Blishchenko E, Iatskin ON, Nazimov IV et al.Fragments of functional proteins in the primary culture of human erythrocytes. BioorgKhim 2008; 34: 160–170.
35 Blishchenko EY, Mernenko OA, Yatskin ON, Ziganshin RH, Philippova MM, Karelin AAet al. Neokyotorphin and neokyotorphin (1–4): secretion by erythrocytes and regulationof tumor cell growth. FEBS Lett 1997; 414: 125–128.
36 Chatterji B, Borlak J. Serum proteomics of lung adenocarcinomas induced by targetedoverexpression of c-raf in alveolar epithelium identifies candidate biomarkers. Proteo-mics 2007; 7: 3980–3991.
37 Deng R, Lu Z, Chen Y, Zhou L, Lu X. Plasma proteomic analysis of pancreatic cancer by2-dimensional gel electrophoresis. Pancreas 2007; 34: 310–317.
RBC release T-cell growth and survival factorsRF Antunes et al
121
Immunology and Cell Biology
Related Documents











