Coral Reefs(1993) 12:57-64 Coral Reefs Springer-Verlag 1993 Reports Recovery of the coral Montastrea annularis in the Florida Keys after the 1987 Caribbean "bleaching event" William K. Fitt l, Howard J. Spero 2, John Halas 3, Michael W. White 3, and James W. Porter l 1 Department of Ecology, Universityof Georgia, Athens, Georgia 30602 2 Department of Geology, University of California, Davis, California 95616 3 Key Largo National Marine Sanctuary, P.O. Box 1083, Key Largo, Florida 33037 Accepted 3 April 1992 Abstract. Many reef-building corals and other cnidarians lost photosynthetic pigments and symbiotic algae (zo- oxanthellae) during the coral bleaching event in the Caribbean in 1987. The Florida Reef Tract included some of the first documented cases, with widespread bleaching of the massive coral Montastrea annularis beginning in late August. Phototransects at Carysfort Reef showed discoloration of >90% of colonies of this species in March 1988 compared to 0% in July 1986; however no mortality was observed between 1986 and 1988. Samples of corals collected in February and June 1988 had zooxanthellae densities ranging from 0.1 in the most lightly colored corals, to 1.6 x 106 cells/cm 2 in the darker corals. Minimum densities increased to 0.5 x 106 cells/cm 2 by August 1989. Chlorophyll-a content of zooxanthellae and zooxanthellar mitotic indices were significantly higher in corals with lower densities ofzooxanthellae, suggesting that zooxanthellae at low densities may be more nutrient- sufficient than those in unbleached corals. Ash-free dry weight of coral tissue was positively correlated with zooxanthellae density at all sample times and was signifi- cantly lower in June 1988 compared to August 1989. Proteins and lipids per cm 2 were significantly higher in August 1989 than in February or June, 1988. Although recovery of zooxanthellae density and coral pigmentation to normal levels may occur in less than one year, regrowth of tissue biomass and energy stores lost during the period of low symbiont densities may take significantly longer. Introduction Bleaching of symbiotic reef invertebrates refers either to loss of symbiotic dinoflagellates (= zooxanthellae hereafter) and/or.reduced algal pigmentation (Hoegh-Guldberg and Smith 1989; Kleppel et al. 1989; Porter et al. 1989; Glynn and D'Croz 1990). In either case the host appears lighter, mottled or even white in color (Fig. 1). The loss of Correspondence to: W. K. Fitt symbionts translates into loss of translocated photo- synthate to the host (Hoegh-Guldberg and Smith 1989; Porter et al. 1989) with consequent lower tissue biomass (Glynn et al. 1985b; Porter et al. 1989; Szmant and Gass- man 1990). Reduced growth (Porter et al. 1989) and mortality (Glynn 1983) may result. Most coral bleaching events in the Caribbean and South Pacific have been relatively localized, and apparent full recovery ofzooxanthellae densities has occurred with relatively little coral mortality (e.g. Oliver 1985, Gates 1990). However, extensive coral mortality following bleaching in the Pacific was caused by the 1982-3 E1 Nino (Oliver 1985, Harriot 1985, Fisk and Done 1985, Glynn and D'Croz 1990). Reef corals and other symbiotic invertebrates in the Caribbean experienced widespread bleaching during the late summer and fall of 1987 (Williams et al. 1987, Williams and Bunkley-Williams 1988). Our earlier study (Porter et al. 1989) reported reduced densities ofzooxan- thellae and chlorophyll-a per zooxanthellae in bleached Montastrea annularis, as well as reduced tissue biomass, immediately following these events. Stable isotope ratios 0C13/•C 12 of M. annularis skeletons from the Virgin Islands, and photosynthetic measurements, suggested decreased nutritional input from zooxanthellae as well as cessation of deposition of calcium carbonate during the bleaching period. Contrary to the suggestion by Knowlton et al. (1992), the bleached and unbleached colonies used in this previous study were all of "morphotype I" M. annularis, and differences in the data were not due to different ecomorphs of M. annularis. In addition, our study demonstrated more seasonal variation in stable isotopic composition within individual specimens of the same morphotype than Knowlton et al. (1992) showed between putative sibling species of M. annularis. In the present study we document "recovery" of M. annularis in the Florida Keys for 2 years after the 1987 bleaching event, in terms of symbiont densities, chlorophyll-a con- tent of zooxanthellae, and biomass parameters of the coral tissue.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Coral Reefs (1993) 12:57-64 Coral Reefs �9 Springer-Verlag 1993
Reports
Recovery of the coral Montastrea annularis in the Florida Keys after the 1987 Caribbean "bleaching event" William K. Fitt l, Howard J. Spero 2, John Halas 3, Michael W. White 3, and James W. Porter l
1 Department of Ecology, University of Georgia, Athens, Georgia 30602 2 Department of Geology, University of California, Davis, California 95616 3 Key Largo National Marine Sanctuary, P.O. Box 1083, Key Largo, Florida 33037
Accepted 3 April 1992
Abstract. Many reef-building corals and other cnidarians lost photosynthetic pigments and symbiotic algae (zo- oxanthellae) during the coral bleaching event in the Caribbean in 1987. The Florida Reef Tract included some of the first documented cases, with widespread bleaching of the massive coral Montastrea annularis beginning in late August. Phototransects at Carysfort Reef showed discoloration of >90% of colonies of this species in March 1988 compared to 0% in July 1986; however no mortality was observed between 1986 and 1988. Samples of corals collected in February and June 1988 had zooxanthellae densities ranging from 0.1 in the most lightly colored corals, to 1.6 x 106 cells/cm 2 in the darker corals. Minimum densities increased to 0.5 x 106 cells/cm 2 by August 1989. Chlorophyll-a content of zooxanthellae and zooxanthellar mitotic indices were significantly higher in corals with lower densities ofzooxanthellae, suggesting that zooxanthellae at low densities may be more nutrient- sufficient than those in unbleached corals. Ash-free dry weight of coral tissue was positively correlated with zooxanthellae density at all sample times and was signifi- cantly lower in June 1988 compared to August 1989. Proteins and lipids per cm 2 were significantly higher in August 1989 than in February or June, 1988. Although recovery of zooxanthellae density and coral pigmentation to normal levels may occur in less than one year, regrowth of tissue biomass and energy stores lost during the period of low symbiont densities may take significantly longer.
Introduction
Bleaching of symbiotic reef invertebrates refers either to loss of symbiotic dinoflagellates (= zooxanthellae hereafter) and/or.reduced algal pigmentation (Hoegh-Guldberg and Smith 1989; Kleppel et al. 1989; Porter et al. 1989; Glynn and D'Croz 1990). In either case the host appears lighter, mottled or even white in color (Fig. 1). The loss of
Correspondence to: W. K. Fitt
symbionts translates into loss of translocated photo- synthate to the host (Hoegh-Guldberg and Smith 1989; Porter et al. 1989) with consequent lower tissue biomass (Glynn et al. 1985b; Porter et al. 1989; Szmant and Gass- man 1990). Reduced growth (Porter et al. 1989) and mortality (Glynn 1983) may result.
Most coral bleaching events in the Caribbean and South Pacific have been relatively localized, and apparent full recovery ofzooxanthellae densities has occurred with relatively little coral mortality (e.g. Oliver 1985, Gates 1990). However, extensive coral mortality following bleaching in the Pacific was caused by the 1982-3 E1 Nino (Oliver 1985, Harriot 1985, Fisk and Done 1985, Glynn and D'Croz 1990).
Reef corals and other symbiotic invertebrates in the Caribbean experienced widespread bleaching during the late summer and fall of 1987 (Williams et al. 1987, Williams and Bunkley-Williams 1988). Our earlier study (Porter et al. 1989) reported reduced densities ofzooxan- thellae and chlorophyll-a per zooxanthellae in bleached Montastrea annularis, as well as reduced tissue biomass, immediately following these events. Stable isotope ratios 0C13/•C 12 of M. annularis skeletons from the Virgin Islands, and photosynthetic measurements, suggested decreased nutritional input from zooxanthellae as well as cessation of deposition of calcium carbonate during the bleaching period. Contrary to the suggestion by Knowlton et al. (1992), the bleached and unbleached colonies used in this previous study were all of "morphotype I" M. annularis, and differences in the data were not due to different ecomorphs of M. annularis. In addition, our study demonstrated more seasonal variation in stable isotopic composition within individual specimens of the same morphotype than Knowlton et al. (1992) showed between putative sibling species of M. annularis. In the present study we document "recovery" of M. annularis in the Florida Keys for 2 years after the 1987 bleaching event, in terms of symbiont densities, chlorophyll-a con- tent of zooxanthellae, and biomass parameters of the coral tissue.

58
Materials and methods
Phototransects
Corals on an established transect line at 15 m on Carysfort Reef, Florida (White and Porter 1985) were resurveyed in 1986 and 1988. The number of colonies of M. annularis, their coloration, and the percentage of discolored coral surface were determined from 24 photographic quadrats (0.375 mZ/quadrat), all from the same depth.
removal of lipid. Freshly isolated zooxanthellae were homogenized with a glass tissue grinder in 100~o acetone and placed in a freezer in the dark under nitrogen for 6-24h for chlorophyll analysis. ChlorophyU-a content was determined by the spectrophometric equations of Jeffrey and Humphrey (1975).
Results
Coloration and zooxanthellae densities
Sample collection
Independent small pieces of M. annularis (5-10 cm in diameter and not connected to other sampled colonies) were collected on Carysfort Reef, Florida on February 5, 1988 (n = 18), June 10, 1988 (n = 12) and August 21, 1989 (n = 10). Only morphotype I of M. annularis (Knowlton et al. 1992) was sampled in this study. At each sampling time, divers collected corals of three arbitrary groups: light colored (including specimens that appeared almost white), dark colored, and intermediate colored. No light corals were observed in August 1989. All samples were collected at depths between 12 and 15 m about midday and maintained at ambient temperature in dim light (< 400 IxE m- 2 see- 1) during the daylight hours and in the dark at night in containers of fresh seawater for 4 15 hours, with at least one water change, until processing began.
Biomass determination
Corals were divided into two pieces. Tissue was removed from both pieces with a high-pressure jet of water (Water PikT~). One piece was processed with distilled water; the other with filtered seawater. The resulting freshwater slurry was frozen for biochemical analysis; the seawater slurry was fixed in 5~o formalin for counts of zooxan- thellae. The surface area of all pieces of coral was determined by covering the region with aluminum foil, weighing the foil, and calculating the surface area from a standard curve.
Each slurry was homogenized with a VirTishear homogenizer and the volume of homogenate was recorded. Densities of zooxan- thellae were determined from 6-10 replicate counts of the homo- genized seawater slurry using a haemocytometer. Mitotic indices (MI) and the percentage of zooxanthellae exhibiting a clear division furrow were calculated from replicate counts (n = 6) of zooxanthellae. Determinations of MI for zooxanthellae in corals collected in February 1988 and August 1989 were made on pieces killed in formalin at dawn (about 15 h after collection), while MI determina- tions in June 1988 were from corals maintained in containers of reef seawater after collection and sampled at various times during the day for 24 h immediately after collection. No released zooxanthellae were seen either on the surface of the coral or in microscopic observations of seawater surrounding corals immediately before processing. Replicate aliquots (n = 3) of the slurry were dried at 60 ~ to constant weight, and then combusted at 500 ~ for 24 h to determine ash-free dry weight (AFDW). The rest of the distilled water slurry was lyophiUized and stored at - 70 ~ for biochemical analysis.
A visual survey of 20 r a n d o m l y selected colonies of M. annular~s at 12-15 m, examined 5 months after bleaching was or ig inal ly r epor t ed from Carysfor t Reef in late Augus t 1987, revealed that 19 exhibi ted par t ia l bleaching. Analysis of M. annularis on a pe rmanen t pho to t r ansec t at 15m showed 51 of 56 to ta l colonies (92~o) were d i sco lo red in March of 1988, 7 months after bleaching was first observed, c o m p a r e d to 0Vo d i sco lo ra t ion in the same colonies on the same t ransect in July 1986. A p p r o x i m a t e l y 65~o of the to ta l surface area was affected in M a r c h 1988. D u r i n g a h a p h a z a r d diving survey in June of 1988, only 35~o (n = 40) of the colonies a p p e a r e d d iscolored, and fewer than 10~o of 50 colonies observed in Augus t of 1989 a p p e a r e d d i sco lored or mot t led . In Sep tember 1990, only one co lony out of 100 seen in a visual diving survey was discolored.
F ie ld observa t ions of p igmen ta t i on and color hue were fairly accura te ind ica tors of zooxan the l l ae density. The mos t l ightly p igmented corals col lected in F e b r u a r y 1988 had only slight hints of b r o w n color. N o colonies were comple te ly white, as was observed in the Virgin I s lands (Por te r et al. 1989) or f rom Sep tember t h rough D e c e m b e r 1987 on Carysfor t Reef (Fig. 1). In contras t , mos t of the corals observed in 1989 and 1990 were uni formly da rk - brown, except as ind ica ted above. Zooxan the l l a e densi t ies in corals from each a rb i t ra ry color ca tegory (light, med ium or dark) were var iable and not significantly different (ANOVA, p > 0.05) over the three sampling times (Table 1). However , densit ies of zooxan the l l ae were highest in the "da rk" or n o r m a l - l o o k i n g corals, and lowest in the " l ight" a p p e a r i n g co lonies ( A N O V A , p <0 .05) . These d a t a indicate tha t the relat ive shades of b r o w n observed unde rwa te r were due at least pa r t i a l ly to densit ies of zooxanthel lae .
Table 1. Mean density __+ s.d. (n) of zooxanthellae ( x 106 cells-cm-2) in relation to relative coloration of live coral, n.d. = no data
Biochemical analysis
Lyophillized coral tissue was reconstituted with distilled deionized water to form a stock slurry with a concentration of 10 ~tg AFDW per ml. Protein content of the diluted slurry was analyzed using the method of Lowry et al. (1951). Lipids were extracted from the slurry by standard procedures (Folch et al. 1956). Carbohydrate in the residue was analyzed as described in Dubois et al. (1956) after
Date Coloration "Light . . . . . Medium "b "Dark TM
February 5, 1988 0.27 + 0.20(2) 0.60 + 0.11 (4) 1.08_+ 0.34(4) June 10, 1 9 8 8 0,24+0.20(4) 0.73+0.21(2) 1.12+0.39(5) August 21, 1989 n.d. 0.58+0.21(4) 1.08+0.19(6)
a.b.c significantly different by coloration (ANOVA, F = 12.181, p = 0.002), not significantly different by date (ANOVA, F = 0.155, p = 0.698)

59
Fig. 1. Montastrea annularis (above) and Diploria labyrinthiformis (below) on Grecian Rocks Reef in the Key Largo National Marine Sanctuary during the "bleaching event", October of 1987 (left), and after recovery of normal coloration in August of 1990 (right)
Chlorophyll-a
Chlorophyl l a per zooxanthel la was not significantly different 5 and 10 mon ths post -bleaching (t-test, p > 0.05), but was negatively correlated with zooxanthel lae density (Fig. 2, best fit line through the points: y = - 15.3 L~ + 6.5, r 2 = 0.84). Corals collected after 24 mon ths had a mean of 6.7 ___ 2.9 (n = 6) pg chlorophyl l a per zooxanthel la for
"da rk" appear ing corals and 8.3 _+ 4.3 (n = 4) pg chloro- phyll a per zooxanthel la for "med ium" colored corals (cf. Table l).
Mitotic index
The mitot ic index (MI) of zooxanthel lae peaked at dawn (Fig. 3a). The peak M I of zooxanthel lae f rom corals
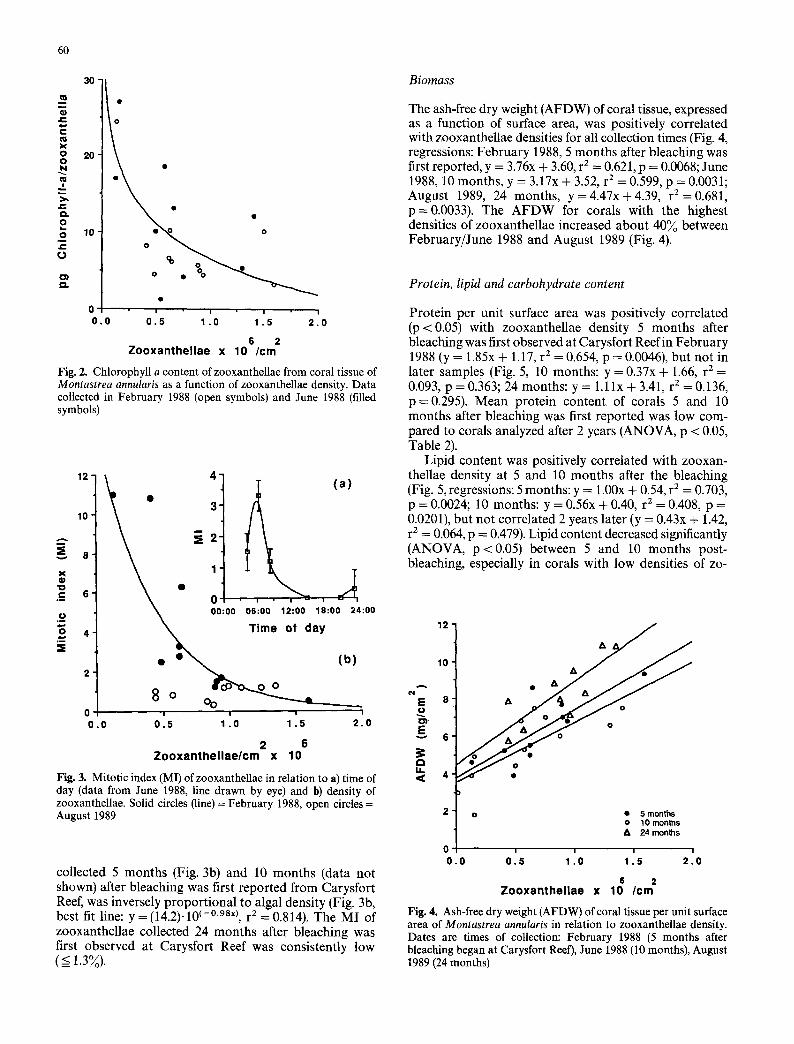
60
30
f0
G) J=: o �9 .i.., C m X o 20 o
~e �9 �9 �9
>,
o 10 ~ �9 o o
r
12.
O | i i i
, 0 0 . 5 1 . 0 1 , 5 2 . 0
6 2 Zooxanthellae x 10 /cm
Fig. 2. Chlorophyll a content of zooxanthellae from coral tissue of Montastrea annularis as a function of zooxanthellae density. Data collected in February 1988 (open symbols) and June 1988 (filled symbols)
1 2 -
1 0
A
a
x
" 0 e- 6
4
0 0 . 0
i 2 -
00:00 08:00 12:00 18:00 :00
Time of day
�9 �9 " ~ (b)
8 0 _ _
i i | i
0 . 5 1 . 0 1 . 5 2 . 0
2 6 Zooxanthellae/cm x 10
Fig. 3. Mitotic index (MI) of zooxanthellae in relation to a) time of day (data from June 1988, line drawn by eye) and b) density of zooxanthellae. Solid circles (line) = February 1988, open circles = August 1989
collected 5 mon ths (Fig. 3b) and 10 mon ths (data not shown) after bleaching was first repor ted f rom Carysfor t Reef, was inversely p ropor t iona l to algal density (Fig. 3b, best fit line: y = (14.2).10 (-~ r 2 = 0.814). The M I of zooxanthel lae collected 24 mon ths after bleaching was first observed at Carysfor t Reef was consistently low ( < 1.3%).
Biomass
The ash-free dry weight (AFDW) of coral tissue, expressed as a function of surface area, was positively correlated with zooxanthellae densities for all collection times (Fig. 4, regressions: Feb rua ry 1988, 5 mon ths after bleaching was first reported, y = 3.76x + 3.60, r 2 = 0.621, p = 0.0068; June 1988, 10 months , y = 3.17x + 3.52, r 2 = 0.599, p = 0.0031; August 1989, 24 months , y = 4.47x + 4.39, r 2 = 0.681, p = 0.0033). The A F D W for corals with the highest densities of zooxanthel lae increased abou t 40~o between Februa ry / June 1988 and August 1989 (Fig. 4).
Protein, lipid and carbohydrate content
Prote in per unit surface area was posit ively correlated (p <0 .05) with zooxanthel lae density 5 mon ths after bleaching was first observed at Carysfor t Reef in Feb rua ry 1988 (y = 1.85x + 1.17, r 2 = 0.654, p = 0.0046), but not in later sample s (Fig. 5, 10 months: y = 0.37x + 1.66, r 2 = 0.093, p = 0.363; 24 months: y = 1.11x + 3.41, r 2 = 0.136, p = 0 . 2 9 5 ) . M e a n prote in content of corals 5 and 10 mon ths after bleaching was first repor ted was low com- pared to corals analyzed after 2 years (ANOVA, p < 0.05, Table 2).
Lipid content was positively correlated with zooxan- thellae density at 5 and 10 mon ths after the bleaching (Fig. 5, regressions: 5 months: y = 1.00x + 0.54, r 2 = 0.703, p = 0.0024; 10 months: y = 0.56x + 0.40, r 2 = 0.408, p = 0.0201), but not corre la ted 2 years later (y = 0.43x + 1.42, r 2 = 0.064, p = 0.479). Lipid content decreased significantly (ANOVA, p <0.05) between 5 and 10 mon ths post- bleaching, especially in corals with low densities of zo-
1 0
E 8 A
E 6 o
u. < 4
2 o �9 5 months o tO months A 24 months
O i i i i
�9 0 0 . 5 1 . 0 1 . 5 2 . 0
6 2 Zooxanthellae x 10 /cm
Fig. 4. Ash-free dry weight (AFDW) of coral tissue per unit surface area of Montastrea annularis in relation to zooxanthellae density. Dates are times of collection: February 1988 (5 months after bleaching began at Carysfort Reef), June 1988 (10 months), August 1989 (24 months)
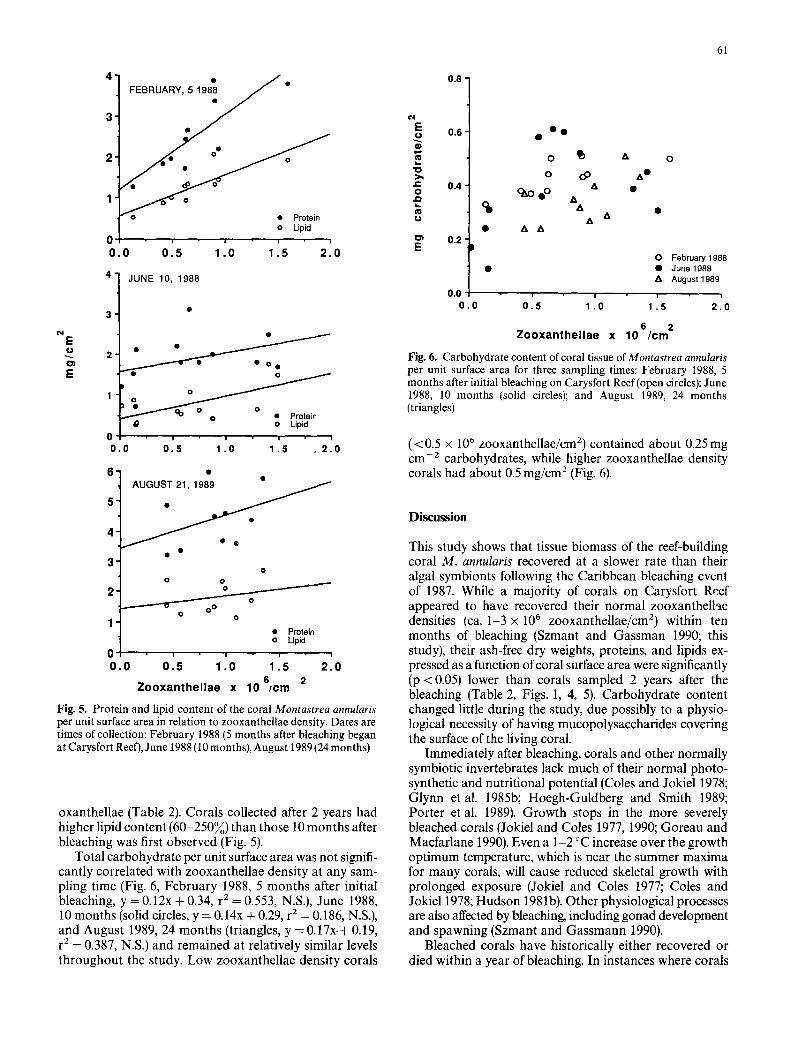
C4 E 0
E
4 -
3 �84
0 0 . 0
FEBRUARY,~ ~ �9
o �9 Protein o Lipid
I I l I
0 . 5 1 . 0 1 . 5 2 , 0
4 -
0 0.0
JUNE 10, 1988
�9 ~ e Oe
o �9 Protein o o
�9 o Lipid
I I I I
0 . 5 1 . 0 1 . 5 . 2 . 0
6 "
5"
4"
3 '
2 '
1
0 O.q
o 0 0 ...--.-
0
0 ~ o o
o o
�9 Protein o Lipid
I i I I
0 . 5 1 . 0 1 . 5 2 . 0 6 2
Z o o x a n t h e l l a e x 10 /cm
Fig. 5. Protein and lipid content of the coral Montastrea annularis per unit surface area in relation to zooxanthellae density. Dates are times of collection: February 1988 (5 months after bleaching began at Carysfort Reef), June 1988 (10 months), August 1989 (24 months)
oxanthellae (Table 2). Corals collected after 2 years had higher lipid content (60-250%) than those 10 months after bleaching was first observed (Fig. 5).
Total carbohydrate per unit surface area was not signifi- cantly correlated with zooxanthellae density at any sam- pling time (Fig. 6, February 1988, 5 months after initial bleaching, y = 0.12x + 0.34, r e = 0.553, N.S.), June 1988, 10 months (solid circles, y = 0.14x + 0.29, r z = 0.186, N.S.), and August 1989, 24 months (triangles, y = 0.17x.+ 0.19, r 2 = 0.387, N.S.) and remained at relatively similar levels throughout the study. Low zooxanthellae density corals
61
0.8
ot
E o 0.e
" 0 .4 o .Q
~ 0.2 E
%
0 0
o ~ A o
o cP a �9 n �9
C~o �9 a & �9
A & A A
O February 1988 �9 �9 June 1988
A August 1989 0.0 , , ,
O. 0.5 1.0 1.5 2.0
6 2 Z o o x a n t h e l l a e x 10 /cm
Fig. 6. Carbohydrate content of coral tissue ofMontastrea annularis per unit surface area for three sampling times: February 1988, 5 months after initial bleaching on Carysfort Reef (open circles); June 1988, 10 months (solid circles); and August 1989, 24 months (triangles)
(< 0.5 x l0 s zooxanthellae/cm 2) contained about 0.25 mg cm -z carbohydrates, while higher zooxanthellae density corals had about 0.5 mg/cm 2 (Fig. 6).
Discussion
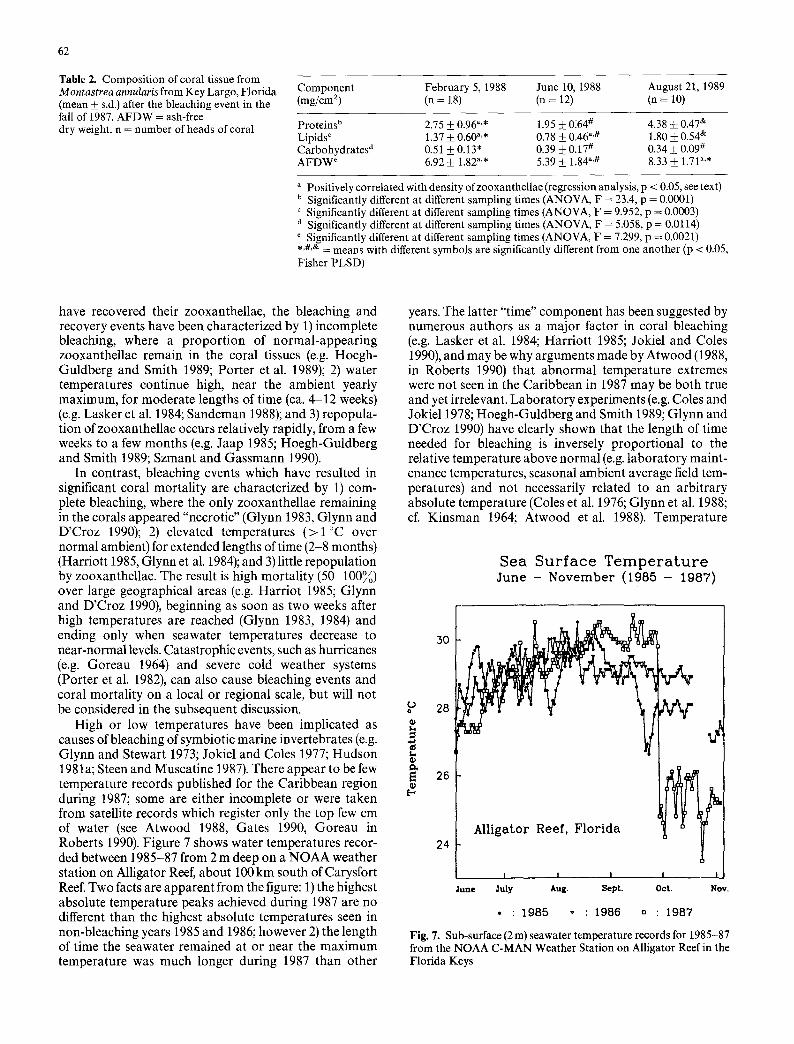
This study shows that tissue biomass of the reef-building coral M. annularis recovered at a slower rate than their algal symbionts following the Caribbean bleaching event of 1987. While a majority of corals on Carysfort Reef appeared to have recovered their normal zooxanthellae densities (ca. 1-3 • 106 zooxanthellae/cm 2) within ten months of bleaching (Szmant and Gassman 1990; this study), their ash-free dry weights, proteins, and lipids ex- pressed as a function of coral surface area were significantly (p < 0.05) lower than corals sampled 2 years after the bleaching (Table 2, Figs. 1, 4, 5). Carbohydrate content changed little during the study, due possibly to a physio- logical necessity of having mucopolysaccharides covering the surface of the living coral.
Immediately after bleaching, corals and other normally symbiotic invertebrates lack much of their normal photo- synthetic and nutritional potential (Coles and Jokiel 1978; Glynn et al. 1985b; Hoegh-Guldberg and Smith 1989; Porter et al. 1989). Growth stops in the more severely bleached corals (Jokiel and Coles 1977, 1990; Goreau and Macfarlane 1990). Even a 1-2 ~ increase over the growth optimum temperature, which is near the summer maxima for many corals, will cause reduced skeletal growth with prolonged exposure (Jokiel and Coles 1977; Coles and Jokiel 1978; Hudson 1981b). Other physiological processes are also affected by bleaching, including gonad development and spawning (Szmant and Gassmann 1990).
Bleached corals have historically either recovered or died within a year of bleaching. In instances where corals
62
Table 2. Composition of coral tissue from Montastrea annularis from Key Largo, Florida (mean 4- s.d.) after the bleaching event in the fall of 1987. AFDW = ash-free dry weight, n = number of heads of coral
Component February 5, 1988 June 10, 1988 August 21, 1989 (mg/cm 2) (n = 18) (n = 12) (n = 10)
Proteins b 2.75 + 0.96 a'* 1.95 • 0:64 # 4.38 + 0.47 e" Lipids c 1.37 +_ 0.60 a'* 0.78 • 0.46 "'# 1.80 _+ 0.54 & Carbohydrates d 0.51 4- 0.13" 0.39 • 0.17 # 0.34 4- 0.09 # AFDW ~ 6.92 • 1.82"'* 5.39 • 1.84 "'# 8.33 4-_ 1.71a'*
a Positively correlated with density ofzooxanthellae (regression analysis, p < 0.05, see text) b Significantly different at different sampling times (ANOVA, F = 23.4, p = 0.0001) c Significantly different at different sampling times (ANOVA, F = 9.952, p = 0.0003) d Significantly different at different sampling times (ANOVA, F = 5.058, p = 0.0114) e Significantly different at different sampling times (ANOVA, F = 7.299, p = 0.0021) .,#,e, = means with different symbols are significantly different from one another (p < 0.05, Fisher PLSD)
have recovered their zooxanthellae, the bleaching and recovery events have been characterized by 1) incomplete bleaching, where a propor t ion of normal-appear ing zooxanthellae remain in the coral tissues (e.g. Hoegh- Guldberg and Smith 1989; Porter et al. 1989); 2) water temperatures continue high, near the ambient yearly maximum, for moderate lengths of time (ca. 4-12 weeks) (e.g. Lasker et al. 1984; Sandeman 1988); and 3) repopula- tion of zooxanthellae occurs relatively rapidly, from a few weeks to a few months (e.g. Jaap 1985; Hoegh-Guldberg and Smith 1989; Szmant and Gassmann 1990).
In contrast, bleaching events which have resulted in significant coral mortality are characterized by 1) com- plete bleaching, where the only zooxanthellae remaining in the corals appeared "necrotic" (Glynn 1983, Glynn and D'Croz 1990); 2) elevated temperatures ( > I ~ over normal ambient) for extended lengths of time (2-8 months) (Harriott 1985, Glynn et at. 1984); and 3) little repopulation by zooxanthellae. The result is high mortality (50-100%) over large geographical areas (e.g. Harriot 1985; Glynn and D'Croz 1990), beginning as soon as two weeks after high temperatures are reached (Glynn 1983, 1984) and ending only when seawater temperatures decrease to near-normal levels. Catastrophic events, such as hurricanes (e.g. Goreau 1964) and severe cold weather systems (Porter et al. 1982), can also cause bleaching events and coral mortality on a local or regional scale, but will not be considered in the subsequent discussion.
High or low temperatures have been implicated as causes of bleaching of symbiotic marine invertebrates (e.g. Glynn and Stewart 1973; Jokiel and Coles 1977; Hudson 1981a; Steen and Muscatine 1987). There appear to be few temperature records published for the Caribbean region during 1987; some are either incomplete or were taken from satellite records which register only the top few cm of water (see Atwood 1988, Gates 1990, Goreau in Roberts 1990). Figure 7 shows water temperatures recor- ded between 1985-87 from 2 m deep on a NOAA weather station on Alligator Reef, about 100 km south of Carysfort Reef. Two facts are apparent from the figure: 1) the highest absolute temperature peaks achieved during 1987 are no different than the highest absolute temperatures seen in non-bleaching years 1985 and 1986; however 2) the length of time the seawater remained at or near the maximum temperature was much longer during 1987 than other
years. The latter "time" component has been suggested by numerous authors as a major factor in coral bleaching (e.g. Lasker et al. 1984; Harr iot t 1985; Jokiel and Coles 1990), and may be why arguments made by Atwood (1988, in Roberts 1990) that abnormal temperature extremes were not seen in the Caribbean in 1987 may be both true and yet irrelevant. Labora tory experiments (e.g. Coles and Jokiel 1978; Hoegh-Guldberg and Smith 1989; Glynn and D 'Croz 1990) have clearly shown that the length of time needed for bleaching is inversely proport ional to the relative temperature above normal (e.g. laboratory maint- enance temperatures, seasonal ambient average field tem- peratures) and not necessarily related to an arbitrary absolute temperature (Coles et al. 1976; Glynn et al. 1988; cf. Kinsman 1964; Atwood et al. 1988). Temperature
Sea Surface Tempera tu re J u n e - N o v e m b e r (1985 - 1987)
3O
2 8
9 L
2 6 [...,
24 Al l iga tor Reef, F lo r ida
June
I I I
July Aug. Sept.
�9 : 1 9 8 5 ', : 1 9 8 6
g
# rj
I I
O c t . Nov.
o : 1987
F i g . 7 . Sub-surface (2 m) seawater temperature records for 1985-87 from the NOAA C-MAN Weather Station on Alligator Reef in the Florida Keys

elevations 4 ~ or higher than the average ambient are followed by immediate bleaching and 90-95% mortal- ity of corals (Jokiel and Coles 1977; Coles et al. 1976; Neudecker 1981), whereas 1-2 ~ increases will produce moderate bleaching with low or no mortality only after prolonged exposure (Coles and Jokiel 1978; Hudson 1981a; Gates 1990; Cook et al. 1990).
Other factors have been suggested as important in bleaching, such as calm weather (e.g. Jaap 1979, Williams and Bunkley-Williams 1988) and irradiance (Coles and Jokiel 1978). Corals bleach mainly on their upper surfaces first, possibly due to exposure to UV light (e.g. Jokiel and York 1982; Siebeck 1988; Lesser et al. 1990). However, UV light is rapidly attenuated with depth (e.g. Fleischmann 1989) and reef corals have concentrations of UV-blocking compounds inversely proportional to depth (Dunlap et al. 1986), suggesting other factors associated with visible light may have a role in bleaching.
Zooxanthellae from corals collected during "peak" bleaching periods contain significantly less chlorophyll a per cell than controls at the same depth and light regime (Porter et al. 1989; Kleppel et al. 1989). However, "recover- ing" corals typically have the same or higher concentra- tions of chlorophyll a per cell than controls (Fig. 2, Szmant and Gassman 1990; Clay Cook pers. com.). Increased concentrations of chlorophyll a and higher MI of zooxanthellae are also characteristics of zooxanthellae exposed to high levels of dissolved nitrogen (ammonia and nitrate) and phosphorus (Hoegh-Guldberg and Smith 1989; Muscatine et al. 1989). Our interpretation is that zooxanthellae remaining after the bleaching event lie in a nutrient-rich intracellular habitat and grow relatively faster than zooxanthellae in corals with higher densities of zooxanthellae. The large range in the chlorophyll data collected from "recovering" corals (Porter et al. 1989; Hayes and Bush 1990; Szmant and Gassmann 1990, this study) may therefore be explained by the variation in both the extent of bleaching of individual coral heads and subsequent zooxanthellae repopnlation rates. There is no evidence that zooxanthellae "repopulate" tissues by infec- ting polyps from seawater, but this mode is certainly possible (Fitt 1984). The evidence presented here, Glynn's (1984) observations of eventual death of bleached corals that had no healthy zooxanthellae remaining, and nu- merous observations of rapid recovery of bleached corals containing zooxanthellae, argue for regrowth of zooxan- thellae primarily from algae remaining in the host after bleaching. Obviously long-term or repeated bleaching may leave corals with few or no zooxanthellae available to repopulate the host tissue and no tissue reserves, resulting in mortality after subsequent bleachings (Savina 1991; Porter and Meier 1992).
Acknowled#ements. We wish to thank the staff of the National Marine Sanctuary program in Washington D.C. and Key Largo for the permits and logistic support necessary to conduct this research. Barry Fitt, Doug Leeper, Ouida Meier, and Yaunlin Zhang helped in various stages of the collection and processing of the specimens. Ian Koblick and the staff of the Marine Resources Development Foundation in Key Largo kindly provided laboratory and other logistic support. Drs. Clay Cook, Clive Wilkinson, Terry Hughes and 2 anonymous reviewers helped to improve the manuscript. This
63
work was supported by NSF (OCE-88-05761 to JWP and WKF; OCE 9203327 to WKF), ONR (N0001492J1734 to WKF), and grants from the MacArthur Foundation through the Florida Institute of Oceanography (JWP), the National Parks Service (JWP), and.the Marine Sanctuary Program of the NOAA.
References
Atwood DK, Sylvester JC, Corredor JE, Morell JM, Mendez A, Nodal WJ, Huss BE Foltz C (1988) Seasurface temperature anomalies for the Caribbean, Gulf of Mexico, Florida Reef Track and the Bahamas considered in light of the 1987 regional coral bleaching event. Proc Assoc Is Mar Lab Carib 21:47
Coles SL, Jokiel PL (1978) Synergistic effects of temperature, salinity and light on the hermatypic coral Montipora verrucosa. Mar Biol 49:187-195
Coles SL, Jokiel PL, Lewis CR (1976) Thermal tolerance in fropical versus subtropical Pacific reef corals. Pac Sci 30:159-166
Cook CB (1990) Elevated temperatures and bleaching on a high latitude coral reef: the 1988 Bermuda event. Coral Reefs 9:45-49
Dubois M, Gilles KA, Hamilton JK, Revers PA, Smith F (1956) Colorometric method for the determination of sugars and related substances. Anal Chem 28:350-356
Dunlap WC, Chalker BE, Oliver JK (1986) Bathymetric adaptations of reef-building corals at Davies Reef, Great Barrier Reef, Australia. III. UV-B absorbing compounds. J Exp Mar Biol Ecol 104:239-248
Fisk DA, Done TJ (1985) Taxonomic and bathymetric patterns of bleaching in corals, Myrmidon Reef (Queensland). Proc 5th Int Coral Reef Symp 6:149-154
Fitt WK (1984) The role of chemosensory behavior of Symbiodinium microadriaticum, intermediate hosts, and host behavior in the infection of coelenterates and molluscs with zooxanthellae. Mar Biol 81:9-17
Fleischmann EM (1989) The measurement and penetration of ultraviolet radiation into tropical marine water. Limnol Ocean- ogr 34:1623-1629
Folch J, Lees M, Sloane-Stanley GH (1956) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497-509
Gates RD (1990) Seawater temperature and sublethal coral bleaching in Jamaica. Coral Reefs 8:193 197
Glynn PW (1983) Extensive "bleaching" and death of reef corals on the Pacific coast of Panama. Environ Conserv 10:149-154
Glynn PW (1984) Widespread coral mortality and the 1982/83 El Nino warming event. Environ Conserv 11:133-146
Glynn PW, Stewart RH (1973) Distribution of coral reefs in the Pearl Islands (Gulf of Panama) in relation to thermal conditions. Limnol Oceanogr 18:367-379
Glynn QW, Perez M, Gilchrist SL (1985) Lipid decline in stressed corals and their crustacean symbionts. Biol Bull 168:276-284
Glynn PW, Cortes J, Guzman HM, Richmond RH (1988) E1 Nino (1982-83) associated coral mortality and relationship to sea surface temperature deviations in the tropical eastern Pacific. Proc. 6th Int Coral Reef Symp 3:237-243
Glynn PW, D'Croz L (1990) Experimental evidence for high tem- perature stress as the cause of El Nino-coincident coral mortality. Coral Reefs 8:181-191
Goreau TF (1964) Mass expulsion of zooxanthellae from Jamaican reef communities after Hurricane Flora. Science 145:383-386
Goreau T J, Macfarlane AH (1990) Reduced growth rate of Montastrea annularis following the 1987-88 coral-bleaching event. Coral Reefs 8:211-215
Harriott VJ (1985) Mortality rates of scleractinian corals before and during a mass bleaching event. Mar Ecol 21:81-88
Hayes RL, Bush PG (1990) Microscopic observations of recovery in the reef-building scleratinian coral, Montastrea annularis, after bleaching on a Cayman reef. Coral Reefs 8:203-209
Hoegh-Guldberg O, Smith GJ (1989) The effect of sudden changes

64
in temperature, light and salinity on the population density and export of zooxanthellae from the reef corals Stylophora pistillata Esper and Seriatopora hystrix Dana. J Exp Mar Biol Ecol 129:279-303
Hudson HJ (1981a) Response of Montastrea annularis to environ- mental change in the Florida Keys. Proc 4th Int Coral Reef Syrup 2: 233 -240
Hudson H J. (1981b) Growth rates in Montastrea annularis: a record of environmental change in Key Largo Coral Reef Marine Sanctuary, Florida. Bull Mar Sci 31:444-459
Jaap WC (1979) Observations on zooxanthellae expulsion at Middle Sambo Reef, Florida Keys. Bull Mar Sci 29:414-422
Jaap WC (1985) An epidemic zooxanthellae expulsion during 1983 in the Lower Florida Keys coral reefs: hyperthermic etiology. Proc 5th Int Coral Reef Syrup 6:143-148
Jeffrey SW, Humphrey GF (1975) New spectrophotometric equations for determining chlorophylls a, b, cl and c2 in higher plants, algae and natural phytoplankton. Biochem Physiol Planz 167: 191-194
Jokiel PL, Coles SL (1977) Effects of temperature on the mortality and growth of Hawaiian reef corals. Mar Biol 43:201-208
Jokiel PL, York RHJr (1982) Solar ultraviolet photobiology of the reef coral Pocillopora damicornis and symbiotic zooxanthellae. Bull Mar Sci 32:301-315
Jokiel PL, Coles SL (1990) Response of Hawaiian and other Indo-Pacific reef corals to elevated temperature. Coral Reefs 8:155-162
Kleppel GS, Dodge RE, Reese CJ (1989) Changes in pigmentation associated with the bleaching of stony corals. Limnol Oceanogr 34:1331-1335
Kinsman DJ (1964) Reef coral tolerance of high temperatures and salinities. Nature 202:1280-1282
Knowlton N, Weil E, Weight LA, Guzman HM (1992) Sibling species in Montastrea annularis, coral bleaching, and the coral climate record. Science 255: 330-333
Lasker RL, Peters EC, Coffroth MA (1984) Bleaching of reef coelenterates in the San Blas Islands, Panama. Coral Reefs 3:183-190
Lesser MP, Stochaj WR, Tapley DW Shick JM (1990) Bleaching in coral reef anthozoans: effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 8:225-232
Lowry OH, Rosenborough HJ, Farr AL, Randall RJ (1951) Protein measurement with folin phenol reagent. J Biol Chem 193:265-275
Muscatine L, Falkowski PG, Dubinsky Z, Cook PA, McCloskey LR (1989) The effect of external nutrient resources on the population dynamics of zooxanthellae in a reef coral. Proc R Soc Lond 236:311-324
Neudecker S (1981) Growth and survival of scleractinian corals exposed to thermal effluents at Guam. Proc 4th Int Coral Reef Symp 1 : 173-180
Oliver J (1985) Recurrent seasonal bleaching and mortality of corals on the Great Barrier Reef. Proc. 5th Int Coral Reef Symp 6: 201-206
Porter JW, Battey JF, Smith GJ (1982) Perturbation and change in coral reef communities. Proc Natl Acad Aci 79:1678-1681
Porter JW, Fitt WK, Spero HJ, Rogers CS, White MW (1989) Bleaching in reef corals: physiological and stable isotopic re- sponses. Proc Natl Acad Sci 86:9342-9346
Porter JW, Meier O (1992) Quantification of loss and change in Floridian reef coral population. Amer. Zool. 32: 625-640
Roberts L (1990) Warm waters, bleached corals. Science 249:213 Sandeman IM (1988) Coral bleaching at Discovery Bay Jamaica: a
possible mechanism for temperature related bleaching. Proc Assoc Is Mar Lab Carib 21:50
Savina LA (1991) Naturally occurring and laboratory induced bleaching in two Caribbean coral species. Am Zool. 31:48A
Siebeck O (1988) Experimental investigation of UV tolerance in hermatypic corals (Scleractinia). Mar Ecol Prog Ser 43:95-103
Steen GR, Muscatine L (1987) Low temperature evokes rapid exocytosis of symbiotic algae by a sea anemone. Biol Bull 172:246-263
Szmant AM, Gassman NJ (1990) The effects of prolonged "bleaching" on the tissue biomass and reproduction of the reef coral Montasrea annularis. Coral Reefs 8:217-224
White MW, Porter JW (1985) The establishment and monitoring Of two permanent photograph transects in Looe Key and:Key Largo National Marine Sanctuaries (Florida Keys). Proc 5th Int Coral Reef Symp 6:531-537
Williams EH, Goenaga C, Vincente V (1987) Mass bleaching on Atlantic coral reefs. Science 237:877-878
Williams EH, Bunkley-WiUiams L (1988) Circumtropical coral reef bleaching in 1987-88. Proc 6th Coral Reef Symp 3:313-318
Related Documents











