Recombinant b-1,3-1,4-glucanase from Theobroma cacao impairs Moniliophthora perniciosa mycelial growth Dahyana Santos Britto • Carlos Priminho Pirovani • Bruno Silva Andrade • Tassiara Pereira dos Santos • Cristina Pungartnik • Ju ´lio Cezar M. Cascardo • Fabienne Micheli • Abelmon S. Gesteira Received: 6 August 2012 / Accepted: 2 May 2013 Ó Springer Science+Business Media Dordrecht 2013 Abstract In this work, we identified a gene from Theo- broma cacao L. genome and cDNA libraries, named TcGlu2, that encodes a b-1,3-1,4-glucanase. The TcGlu2 ORF was 720 bp in length and encoded a polypeptide of 239 amino acids with a molecular mass of 25.58 kDa. TcGlu2 contains a conserved domain characteristic of b- 1,3-1,4-glucanases and presented high protein identity with b-1,3-1,4-glucanases from other plant species. Molecular modeling of TcGlu2 showed an active site of 13 amino acids typical of glucanase with b-1,3 and 1,4 action mode. The recombinant cDNA TcGlu2 obtained by heterologous expression in Escherichia coli and whose sequence was confirmed by mass spectrometry, has a molecular mass of about 22 kDa (with His-Tag) and showed antifungal activity against the fungus Moniliophthora perniciosa, causal agent of the witches’ broom disease in cacao. The integrity of the hyphae membranes of M. perniciosa, incubated with protein TcGlu2, was analyzed with propi- dium iodide. After 1 h of incubation, a strong fluorescence emitted by the hyphae indicating the hydrolysis of the membrane by TcGlu2, was observed. To our knowledge, this is the first study of a cacao b-1,3-1,4-glucanase expression in heterologous system and the first analysis showing the antifungal activity of a b-1,3-1,4-glucanase, in particular against M. perniciosa. Keywords Glycosyl hydrolases Witches’ broom Pathogenesis related protein Antifungal activity Abbreviations ORF Open reading frame PB Phosphate buffer PI Propidium iodide Introduction Carbohydrates constitute the major class of organic com- pounds in plant tissues and are involved in numerous physiological processes such as growth, signaling, metab- olism, symbiosis and plant defense [1]. These carbohy- drates, which present large structural and functional diversity, are synthesized, modified and degraded by a great variety of enzymes. The ‘‘carbohydrate-active enzymes’’ are classified based on amino acid sequences and represent 130 different families which can be accessed in the CAZy website (Carbohydrate Active Enzymes database; http://www.cazy.org/;[2]). In particular, b-glu- can degradation in nature is catalyzed by b-glucanases and endoglucanases that depolymerize b-1,3-1,4-D-glucans. These enzymes are organized in four categories: (i) specific This article is dedicated in memory of Ju ´lio Cezar M. Cascardo. D. S. Britto C. P. Pirovani T. P. dos Santos C. Pungartnik J. C. M. Cascardo F. Micheli A. S. Gesteira Centro de Biotecnologia e Gene ´tica, Universidade Estadual de Santa Cruz (UESC), Rodovia Ilhe ´us-Itabuna, km 16, Ilhe ´us, BA 45662-900, Brazil B. S. Andrade Universidade Estadual do Sudoeste da Bahia (UESB), Av. Jose ´ Moreira Sobrinho, Jequie ´, BA 45206-190, Brazil F. Micheli (&) CIRAD-BIOS, UMR AGAP, Avenue Agropolis TA96/03, 34398 Montpellier Cedex 5, France e-mail: [email protected] A. S. Gesteira Embrapa Mandioca e Fruticultura, Caixa Postal 007, Cruz das Almas, BA 44380-000, Brazil 123 Mol Biol Rep DOI 10.1007/s11033-013-2640-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Recombinant b-1,3-1,4-glucanase from Theobroma cacao impairsMoniliophthora perniciosa mycelial growth
Dahyana Santos Britto • Carlos Priminho Pirovani • Bruno Silva Andrade •
Tassiara Pereira dos Santos • Cristina Pungartnik • Julio Cezar M. Cascardo •
Fabienne Micheli • Abelmon S. Gesteira
Received: 6 August 2012 / Accepted: 2 May 2013
� Springer Science+Business Media Dordrecht 2013
Abstract In this work, we identified a gene from Theo-
broma cacao L. genome and cDNA libraries, named
TcGlu2, that encodes a b-1,3-1,4-glucanase. The TcGlu2
ORF was 720 bp in length and encoded a polypeptide of
239 amino acids with a molecular mass of 25.58 kDa.
TcGlu2 contains a conserved domain characteristic of b-
1,3-1,4-glucanases and presented high protein identity with
b-1,3-1,4-glucanases from other plant species. Molecular
modeling of TcGlu2 showed an active site of 13 amino
acids typical of glucanase with b-1,3 and 1,4 action mode.
The recombinant cDNA TcGlu2 obtained by heterologous
expression in Escherichia coli and whose sequence was
confirmed by mass spectrometry, has a molecular mass of
about 22 kDa (with His-Tag) and showed antifungal
activity against the fungus Moniliophthora perniciosa,
causal agent of the witches’ broom disease in cacao. The
integrity of the hyphae membranes of M. perniciosa,
incubated with protein TcGlu2, was analyzed with propi-
dium iodide. After 1 h of incubation, a strong fluorescence
emitted by the hyphae indicating the hydrolysis of the
membrane by TcGlu2, was observed. To our knowledge,
this is the first study of a cacao b-1,3-1,4-glucanase
expression in heterologous system and the first analysis
showing the antifungal activity of a b-1,3-1,4-glucanase, in
particular against M. perniciosa.
Keywords Glycosyl hydrolases � Witches’ broom �Pathogenesis related protein � Antifungal activity
Abbreviations
ORF Open reading frame
PB Phosphate buffer
PI Propidium iodide
Introduction
Carbohydrates constitute the major class of organic com-
pounds in plant tissues and are involved in numerous
physiological processes such as growth, signaling, metab-
olism, symbiosis and plant defense [1]. These carbohy-
drates, which present large structural and functional
diversity, are synthesized, modified and degraded by a
great variety of enzymes. The ‘‘carbohydrate-active
enzymes’’ are classified based on amino acid sequences
and represent 130 different families which can be accessed
in the CAZy website (Carbohydrate Active Enzymes
database; http://www.cazy.org/; [2]). In particular, b-glu-
can degradation in nature is catalyzed by b-glucanases and
endoglucanases that depolymerize b-1,3-1,4-D-glucans.
These enzymes are organized in four categories: (i) specific
This article is dedicated in memory of Julio Cezar M. Cascardo.
D. S. Britto � C. P. Pirovani � T. P. dos Santos � C. Pungartnik �J. C. M. Cascardo � F. Micheli � A. S. Gesteira
Centro de Biotecnologia e Genetica, Universidade Estadual de
Santa Cruz (UESC), Rodovia Ilheus-Itabuna, km 16, Ilheus,
BA 45662-900, Brazil
B. S. Andrade
Universidade Estadual do Sudoeste da Bahia (UESB), Av. Jose
Moreira Sobrinho, Jequie, BA 45206-190, Brazil
F. Micheli (&)
CIRAD-BIOS, UMR AGAP, Avenue Agropolis TA96/03,
34398 Montpellier Cedex 5, France
e-mail: [email protected]
A. S. Gesteira
Embrapa Mandioca e Fruticultura, Caixa Postal 007, Cruz das
Almas, BA 44380-000, Brazil
123
Mol Biol Rep
DOI 10.1007/s11033-013-2640-1

b-1,3-1,4-D-glucanases or true lichenases (EC 3.2.1.73)
that strictly cleave b-1,4-glycosidic linkages adjacent to a
3-O-substituted glucose residue but are inactive against b-
1,4-glucans; (ii) endo-b-1,4-D-glucanases (EC 3.2.1.4),
which hydrolyze b-1,4-glycosidic bonds other than those
targeted by lichenases; (iii) b-1,3(4)-D-glucanases (EC
3.2.1.6) active on b-1,3-1,4-D-glucans and b-1,3-D-glucans;
and (iv) b-1,3-D-glucanases or laminarinase (EC 3.2.1.39)
[2, 3]. The b-1,3-1,4-D-glucanases (lichenases) have been
identified in various microorganisms [4–7] and fungi [8–
10], mainly for industrial applications. However, b-1,3-1,4-
D-glucanases from plants have been little studied, and these
studies focused mainly on plant development [11–13]. In
2009, Akiyama et al. showed that an endo-(1,3;1-4)-b-
glucanase gene from rice (OsEGL2) had its expression
significantly increased in response to methyl jasmonate,
abscissic acid and mechanical wounding [13]. The
mechanical wounding also increased the leaf elongation
rate in rice seedlings in comparison to the control [13]. To
our knowledge, there is only few data about involvement of
b-1,3-1,4-D-glucanases in plant defense or in plant–patho-
gen interactions. Other plant glycosyl hydrolases are well
known to be involved in such responses to pathogens such
as chitinases and b-1,3-glucanases and may be classified as
pathogenesis-related (PR) proteins [1]. Among various
examples, some of them are related to cacao disease
resistance [14, 15]. A PR10 protein of cacao (TcPR-10),
although it does not belong to the glysosyl hydrolase
family, showed in vitro and in vivo antifungal activity
against mono- and dicaryotic mycelium and basidiospores
of M. perniciosa, and against Saccharomyces cerevisiae
[14, 16]. On the other hand, a chitinase gene of cacao
(TcChi1) was used under the control of a modified CaMV-
35S promoter for Agrobacterium-mediated transformation
of cacao somatic embryo cotyledons [15]. The TcChi1
transgenic cacao plants showed an in vivo antifungal
activity against the foliar pathogen Colletotrichum gloeo-
sporioides; fungal growth and leaf necrosis were reduced in
the transgenic plants when compared to controls.
Cacao (Theobroma cacao L.) is an important com-
modity cultivated primarily to provide cacao liquor, butter,
and powder for the chocolate industry and unfortunately
has been frequently the target of several fungal diseases
[17]. For this reason, we focused our attention on a gene of
b-1,3-1,4-glucanase from T. cacao (TcGlu2) as a candidate
for disease control. Here, we report the molecular cloning
of TcGlu2, the heterologous expression of the recombinant
protein and its antifungal activity against Moniliophthora
perniciosa, the causal agent of the witches’ broom disease.
To our knowledge, this is the first study of a cacao glu-
canase expression in heterologous system and the first
analysis showing the antifungal activity of a b-1,3-1,4-
glucanase.
Materials and methods
Identification and analysis of genomic and cDNA
sequences of TcGlu2
The TcGlu2 cDNA was identified from a cDNA library of
susceptible cacao (Catongo variety) inoculated by M.
perniciosa [18] and later the complete sequence was
encountered in the T. cacao genome [19]. The Open
reading frame (ORF) of the nucleotide sequence was
determined using the ORFinder program (Lasergene,
Madison, WI, USA). For function homology analysis, the
sequence was compared with the public sequence database
using BLAST [20]. ClustalW was used for multiple
nucleotide or amino acid sequence alignment [21]. NetPhos
2.0 Server [22] and InterProScan [23] were used for
identification of putative phosphorylation sites and con-
served domains, respectively.
Phylogeny
All b-1,3-1,4-glucanases from T. cacao were obtained at
CocoaGenDB website (http://cocoagendb.cirad.fr/gbrowse/
cgi-bin/gbrowse/theobroma/); TcGlu2 and two more
sequences homologous to this one were found. Phyloge-
netic analyses were performed with the BLAST program
version 2.2.27 [20] using: (i) TcGlu2 and the two other
amino acid sequences of b-1,3-1,4-glucanases from T.
cacao; and (ii) b-1,3-1,4-glucanases from other plant spe-
cies similar to T. cacao glucanases. Amino acid sequences
were aligned using BLOSUM matrix [24] in ClustalW2
(http://www.ebi.ac.uk/Tools/msa/clustalw2/) [25]. The phy-
logenetic Bayesian analyses were performed in MRBA-
YES 3.1.2 [26] using the mixed evolutionary model. Three
independent runs were conducted (each with four chains)
for 2 9 106 generations, sampling every 100 generations.
For this analysis we used one of Brachypodium distachyon
glucanase sequences (XP 0035684871) as outgroup, because
of its more distant evolution relationship with T. cacao.
Molecular modeling of TcGlu2
The structure of the complete ORF of TcGlu2 was built by
Comparative Modeling approach. Initially, TcGlu2 amino
acid sequence was subjected to the Swiss Model Workspace
server, using an automated modeling approach [27, 28] to
find templates that could be used to construct the TcGlu2
model. Additionally, other templates deposited in Protein
Data Bank, were searched. All templates found were sub-
mitted to an alignment with the TcGlu2 sequence using
TCOFFEE [29], to find conserved regions and motifs. Sep-
arately, another TCOFFEE alignment with 1ZM1—a b-1,3-
1,4-D-glucanase of Fibrobacter succinogenes previously
Mol Biol Rep
123

crystallized and considered as a good template for molecular
modeling [30]—was performed in order to find conserved
active site regions between the two sequences. The 3D model
was generated using Swiss Model Workspace [27, 28] in an
alignment mode. Afterwards, the initial model was prepared
by AMBER 11 package [31, 32], using LEAP and SANDER
for structure refinement. The structure was fully minimized
with 500 steps of steepest descent followed by 500 more
steps of conjugate gradient to an RMS gradient of 0.01 kcal/
2.71 A in vacuum. Molecular dynamics simulation of
refined structure was performed in vacuum using f99 force
field at 300 K for 1 ns, restricted to residues 90–145—
according to TcGlu2/1ZM1 alignment. Finally, PRO-
CHECK 3.6 [33] and ANOLEA [34, 35] were used to eval-
uate stereo chemical and energy quality of the final model.
The final structure of TcGlu2 and the crystallographic
structure of 1ZM1 were structurally aligned by Pymol
1.5.0.4 (The PyMOL Molecular Graphics System, 2012)
using the script fitting.py, and restricted to the amino acids
fragments 90–145 in the TcGlu2 sequence, and 58–148 in the
1ZM1 sequence. The active site of TcGlu2 was estimated
using superimposition and alignment data, and observing
which 1ZM1 catalytic amino acids best fit with the target
model amino acids.
Expression of recombinant TcGlu2
The TcGlu2 cDNA was amplified by PCR using the TcGlu-F
(50-GGCGGGATCCATATGTCAGGTCCGCAGTGC-30)and TcGlu-R (50-GGCGGTCGACTCAGGGTTTAGCTTT
TAAG-30) primers. The PCR product was cloned into the
NdeI and SalI sites of the plasmid pET28a (Novagen�),
and the resulting in frame fusion plasmid was transformed
into Escherichia coli strain Rosetta (DE3). Overexpression
of the TcGlu2 tagged with six histine residues at the
N-terminus was induced by 1 mM of isopropyl-b-D-thiog-
alactoside at 37 �C. To establish the kinetics of TcGlu2
induction, bacteria were collected and protein content
analyzed 1, 2 and 3 h after induction. Larger quantities of
TcGlu2 that are needed for protein purification were
obtained 4 h after induction. For recombinant protein
purification, bacteria were centrifuged and washed once in
equilibration buffer [50 mM phosphate buffer (PB),
300 mM NaCl, pH 7.4], suspended in lysis buffer
(50 mM PB, 300 mM NaCl, 0.1 mg/plate lysozyme, pH
7.4) and kept at room temperature for 1 h. The sample was
kept in ice and sonicated (Gex Ultrasonic processor 130,
130W; 8 pulses of 30 s each, 75 % output, 30 s intervals),
and the resulting lysate was centrifuged for 20 min at
11,000g. The supernatant containing soluble proteins was
loaded onto Talon resin metal affinity column (Clontech�
Laboratories), eluted with 150 mM imidazole and dialyzed
against 500 ml of 50 mM PB, pH 7.0. The purified TcGlu2
protein was digested with trypsin (25 ng/ll) at 37 �C for
12 h according to the manufacturer’s instructions (Pro-
mega). The resulting tryptic digests were vacuum con-
centrated (Concentrator 5301, Eppendorf), desalted using a
pre-Symmetry column (Waters, Mildford, MA, USA) C18
(5, 180 lm in inner diameter 9 20 mm long), and then
fractioned by C18 reverse phase chromatography column
(100 mm 9 100 lm, 1.7 lm particles) on the nanoAcquity
UPLC (WATERS) for 50 min under 0.6 ll min-1 aceto-
nitrile flux. The following gradient was used: 1 % for
1 min, 1–50 % in 40 min, 50–85 % in 5 min, 85 % for
2 min, 85–1 % in 1 min, 1 % for 2 min. Afterwards, the
peptides were deionized at 3,000 V and split on positive
mode with minimum relative intensity of 10 counts on
Micromass ESI-Q-TOF (WATERS). Spectra were ana-
lyzed using the ProteinLynx Global Server 4.2 (WATERS)
and compared with the NCBI database (http://www.ncbi.
nlm.nih.gov/BLAST/).
M. perniciosa growth conditions
M. perniciosa strain ALF553 cultures (CCBM000257,
UEFS, Feira de Santana, Brazil) were grown and toxicity
tests were performed on M. perniciosa pseudo-colonies as
previously described [36]. Dikaryotic cultures were grown
in CPD (2 % glucose, 2 % peptone) in liquid media,
without agitation at 25 �C, for 5–7 days. Agar 2 % was
added when solid media were used.
Antifungal activity of TcGlu2
TcGlu2 (6, 12 and 32 lg/plate) or PB (control) or purified
protein extract from E. coli expressing pET28a without insert
was spread onto 20 ml of CPD agar plates (control). After-
wards, 1 ml of dikaryotic M. perniciosa broken hyphae [36]
was spread one the same plates which were further incubated
for 7 days at 25 �C. M. perniciosa survival was defined as the
percentage of grown pseudo-colonies (treated/PB con-
trol 9 100). The experiment was made in triplicate.
Membrane viability analysis
To check for viability and membrane integrity, fungal
hyphae were incubated with both TcGlu2 (8 lg/ll) and
2 lM of propidium iodide (PI) at 25 �C. After incubation
intervals ranging from 30 min to 1 h, PI-labeled, hyphae
were washed twice in 0.05 mM PB and then observed
under fluorescence microscope DMRA2 (Leica�) attached
with PI filter. The following negative controls were used:
(i) assay with TcGlu2 boiled for 1 h at 100 �C; (ii) assay in
presence of NaN3 (inhibitor of metabolism); and (iii) assay
at 4 �C (inhibition of protein transport). Images were
captured using 409 and 1009 objectives under bright field
Mol Biol Rep
123

A
B
Mol Biol Rep
123

as well as under fluorescent filter using the IM50 software
(Leica�).
Results
Sequence analysis
The sequence TcGlu2 was identified as a probable b-1,3-
1,4-glucanase with an ORF of 720 nucleotides encoding a
protein of 239 amino acids residues. The protein contains a
domain conserved between b-1,3-1,4-glucanases (from
amino acid 5–238) and 11 putative phosphorylation sites
on serines (S17, S86, S115, S119, S160, S176 and S221) and
tyrosine (Y69, Y222 and Y237) (Fig. 1a). The complete
protein has a calculated molecular mass of 25.58 kDa and
an isoelectric point of 5.85. The cDNA obtained from
cDNA libraries already published [18] presents an incom-
plete ORF of 582 nucleotides encoding a protein of 194
amino acids residues (Fig. 1a, filled triangle). The corre-
sponding predicted protein, without the His-Tag, has a
calculated molecular mass of 20.61 kDa (22.2 kDa with
the His-Tag) and an isoelectric point of 6.82. The amino
acid alignment of TcGlu2 with different members of the
glycosylated hydrolase family (b-1,3-1,4 glucanases)
indicated high similarity between the sequences (Fig. 1b).
Phylogeny of T. cacao glucanases
The Bayesian consensus phylogram positioned the glu-
canases found in T. cacao genome in relation to those
found in other plant families (Fig. 2). T. cacao glucanase
(g009650) and TcGlu2 (g022720) formed a polyphyletic
clade with glucanases of Ricinus communis and Glycine
max. T. cacao (g022730) presented itself externally over
the tree, forming a polyphyletic clade with R. communis
glucanase and with a group of R. communis (XP 0025167911)
and G. max (XP 0035428951) glucanases.
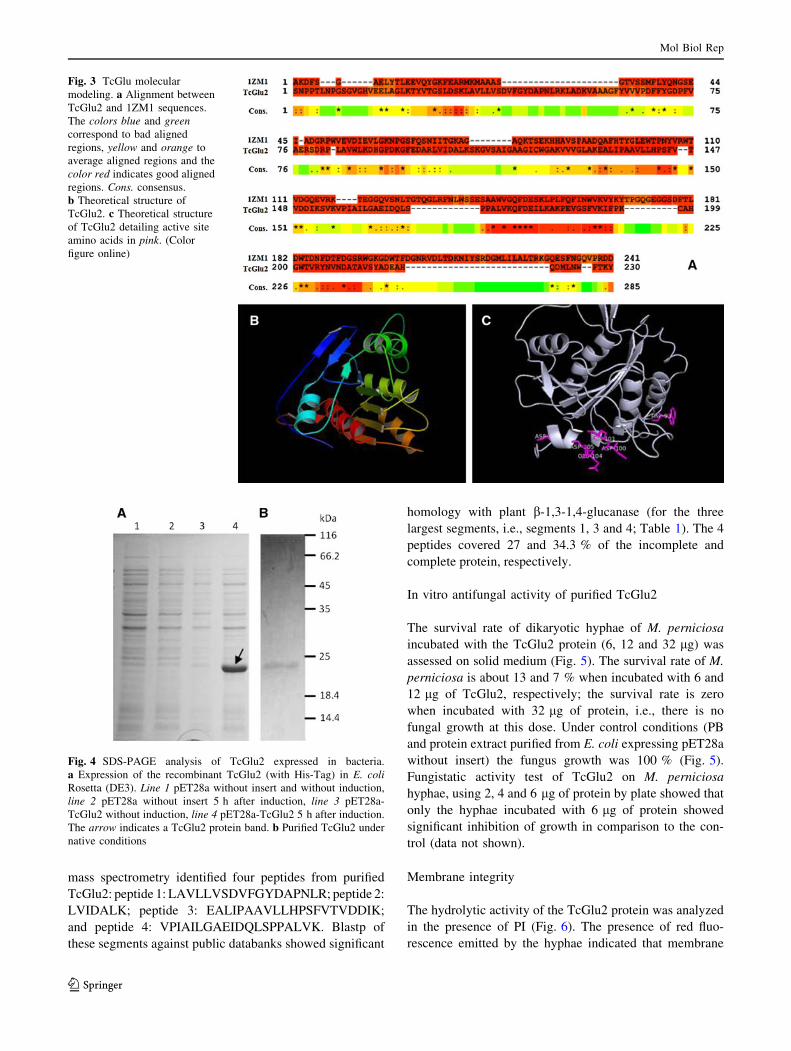
TcGlu2 structure
TcGlu2 is a single b-1,3-1,4-D-glucanase chain presenting
7 a-helices, 10 b-sheets and 4 salt bridges (Fig. 3b). After
TCOFFEE alignment with 1ZM1 sequence, a high level of
identity between both sequences was found, in particular
with the active site region of 1ZM1 (Fig. 3a). The active
site of TcGlu2 is composed of 13 amino acids (Trp93,
Leu94, Lys95, Asp96, His97, Gly98, Pro99, Asp100, Lys101,
Gly102, Phe103, Glu104, Asp105), a space of 2831 A, and
presents a-helices and loop structures (Figs. 3c, 1a).
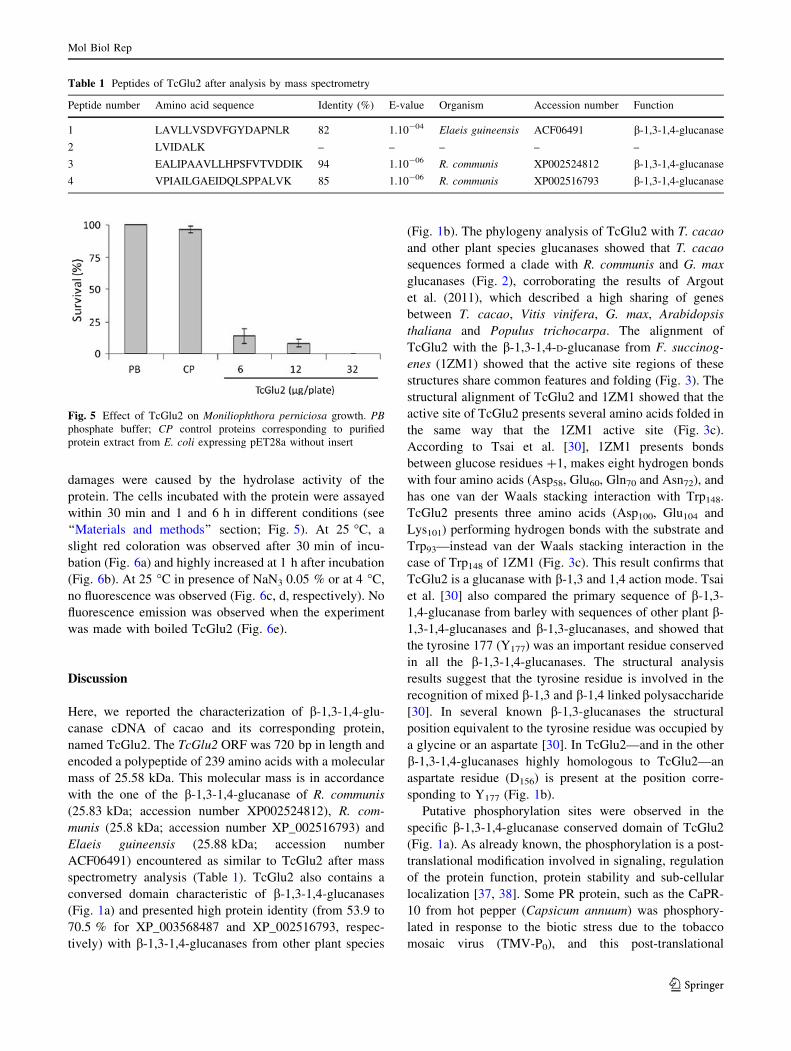
Expression analysis of recombinant protein TcGlu2
and mass spectrometry sequence validation
The TcGlu2 cDNA was successfully cloned in pET28a
plasmid and expressed in E. coli Rosetta (DE3) after 1 h of
induction. The highest yield of the recombinant protein was
obtained after 5 h of induction (Fig. 4a, line 4), whereas no
visible band was observed in the control (pET28a without
insert) (Fig. 4a, line 2). The soluble enzyme purified from the
cytosolic fraction of lysed cells showed a molecular mass of
25 kDa—with His-Tag (Fig. 4b). Trypsin digestion and
Fig. 2 Bayesian phylogenetic analysis, using amino acid data.
Bayesian consensus phylogram of T. cacao b-1,3-1,4-glucanases
and glucanases from other plant species
Fig. 1 Nucleotide and amino acid sequences of TcGlu2. a Nucleotide
and deduced amino acid sequences of TcGlu2. The asterisk represents
the ORF termination codon. Putative phosphorylation sites are squared
on the amino acid sequence. The conserved domain between b-1,3-1,4-
glucanases, is underlined. Putative active site is shaded in grey. The
filled triangle represents the beginning of the lost region in TcGlu2
cDNA. b Comparison of predicted amino acid sequence of TcGlu2 with
those of Vitis vinifera b-1,3-1,4-D-glucanase (XP_002275697), R.communis b-1,3-1,4-D-glucanase (XP_002524812), R. communis b-
1,3-1,4-D-glucanase (XP_002516793), G. max endo-1,3-1,4-b-D-glu-
canase (XP_003550071), Arabidopsis thaliana b-1,3-1,4-D-glucanase
(AAM61180), E. guineensis endo-1,3-1,4-b-D-glucanase (ACF06491),
Brachypodium distachyon endo-1,3-1,4-b-D-glucanase (XP_00356
8483), Brachypodium distachyon endo-1,3-1,4-b-D-glucanase
(XP_003568487). Gaps introduced to get the best alignment are
indicated by dashes, (asterisks) represents identical amino acids
between all sequences, (dot) and (colon) represent conserved substitu-
tions and semi-conserved substitutions, respectively. Identity percent
between TcGlu2 and the other sequences is indicated at the right of each
sequence end. Conserved aspartate residues potentially involved in
recognition and/or cleavage of a specific substrate are shaded in grey
b
Mol Biol Rep
123
mass spectrometry identified four peptides from purified
TcGlu2: peptide 1: LAVLLVSDVFGYDAPNLR; peptide 2:
LVIDALK; peptide 3: EALIPAAVLLHPSFVTVDDIK;
and peptide 4: VPIAILGAEIDQLSPPALVK. Blastp of
these segments against public databanks showed significant
homology with plant b-1,3-1,4-glucanase (for the three
largest segments, i.e., segments 1, 3 and 4; Table 1). The 4
peptides covered 27 and 34.3 % of the incomplete and
complete protein, respectively.
In vitro antifungal activity of purified TcGlu2
The survival rate of dikaryotic hyphae of M. perniciosa
incubated with the TcGlu2 protein (6, 12 and 32 lg) was
assessed on solid medium (Fig. 5). The survival rate of M.
perniciosa is about 13 and 7 % when incubated with 6 and
12 lg of TcGlu2, respectively; the survival rate is zero
when incubated with 32 lg of protein, i.e., there is no
fungal growth at this dose. Under control conditions (PB
and protein extract purified from E. coli expressing pET28a
without insert) the fungus growth was 100 % (Fig. 5).
Fungistatic activity test of TcGlu2 on M. perniciosa
hyphae, using 2, 4 and 6 lg of protein by plate showed that
only the hyphae incubated with 6 lg of protein showed
significant inhibition of growth in comparison to the con-
trol (data not shown).
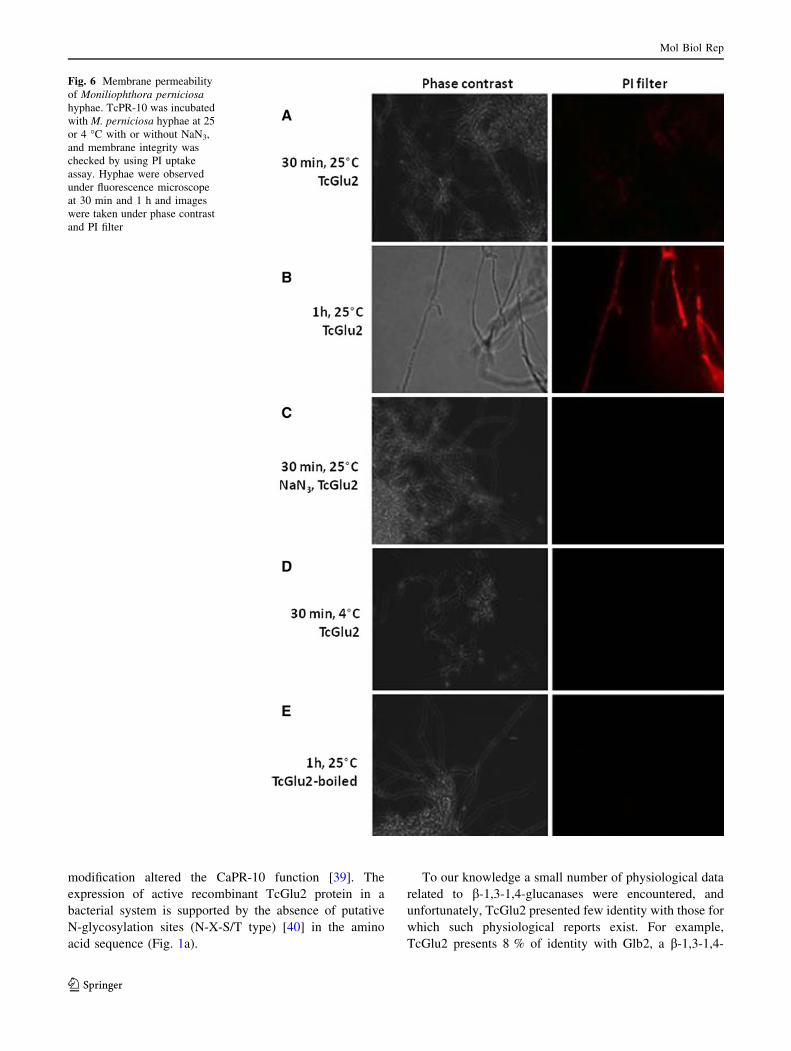
Membrane integrity
The hydrolytic activity of the TcGlu2 protein was analyzed
in the presence of PI (Fig. 6). The presence of red fluo-
rescence emitted by the hyphae indicated that membrane
Fig. 3 TcGlu molecular
modeling. a Alignment between
TcGlu2 and 1ZM1 sequences.
The colors blue and greencorrespond to bad aligned
regions, yellow and orange to
average aligned regions and the
color red indicates good aligned
regions. Cons. consensus.
b Theoretical structure of
TcGlu2. c Theoretical structure
of TcGlu2 detailing active site
amino acids in pink. (Color
figure online)
Fig. 4 SDS-PAGE analysis of TcGlu2 expressed in bacteria.
a Expression of the recombinant TcGlu2 (with His-Tag) in E. coliRosetta (DE3). Line 1 pET28a without insert and without induction,
line 2 pET28a without insert 5 h after induction, line 3 pET28a-
TcGlu2 without induction, line 4 pET28a-TcGlu2 5 h after induction.
The arrow indicates a TcGlu2 protein band. b Purified TcGlu2 under
native conditions
Mol Biol Rep
123
damages were caused by the hydrolase activity of the
protein. The cells incubated with the protein were assayed
within 30 min and 1 and 6 h in different conditions (see
‘‘Materials and methods’’ section; Fig. 5). At 25 �C, a
slight red coloration was observed after 30 min of incu-
bation (Fig. 6a) and highly increased at 1 h after incubation
(Fig. 6b). At 25 �C in presence of NaN3 0.05 % or at 4 �C,
no fluorescence was observed (Fig. 6c, d, respectively). No
fluorescence emission was observed when the experiment
was made with boiled TcGlu2 (Fig. 6e).
Discussion
Here, we reported the characterization of b-1,3-1,4-glu-
canase cDNA of cacao and its corresponding protein,
named TcGlu2. The TcGlu2 ORF was 720 bp in length and
encoded a polypeptide of 239 amino acids with a molecular
mass of 25.58 kDa. This molecular mass is in accordance
with the one of the b-1,3-1,4-glucanase of R. communis
(25.83 kDa; accession number XP002524812), R. com-
munis (25.8 kDa; accession number XP_002516793) and
Elaeis guineensis (25.88 kDa; accession number
ACF06491) encountered as similar to TcGlu2 after mass
spectrometry analysis (Table 1). TcGlu2 also contains a
conversed domain characteristic of b-1,3-1,4-glucanases
(Fig. 1a) and presented high protein identity (from 53.9 to
70.5 % for XP_003568487 and XP_002516793, respec-
tively) with b-1,3-1,4-glucanases from other plant species
(Fig. 1b). The phylogeny analysis of TcGlu2 with T. cacao
and other plant species glucanases showed that T. cacao
sequences formed a clade with R. communis and G. max
glucanases (Fig. 2), corroborating the results of Argout
et al. (2011), which described a high sharing of genes
between T. cacao, Vitis vinifera, G. max, Arabidopsis
thaliana and Populus trichocarpa. The alignment of
TcGlu2 with the b-1,3-1,4-D-glucanase from F. succinog-
enes (1ZM1) showed that the active site regions of these
structures share common features and folding (Fig. 3). The
structural alignment of TcGlu2 and 1ZM1 showed that the
active site of TcGlu2 presents several amino acids folded in
the same way that the 1ZM1 active site (Fig. 3c).
According to Tsai et al. [30], 1ZM1 presents bonds
between glucose residues ?1, makes eight hydrogen bonds
with four amino acids (Asp58, Glu60, Gln70 and Asn72), and
has one van der Waals stacking interaction with Trp148.
TcGlu2 presents three amino acids (Asp100, Glu104 and
Lys101) performing hydrogen bonds with the substrate and
Trp93—instead van der Waals stacking interaction in the
case of Trp148 of 1ZM1 (Fig. 3c). This result confirms that
TcGlu2 is a glucanase with b-1,3 and 1,4 action mode. Tsai
et al. [30] also compared the primary sequence of b-1,3-
1,4-glucanase from barley with sequences of other plant b-
1,3-1,4-glucanases and b-1,3-glucanases, and showed that
the tyrosine 177 (Y177) was an important residue conserved
in all the b-1,3-1,4-glucanases. The structural analysis
results suggest that the tyrosine residue is involved in the
recognition of mixed b-1,3 and b-1,4 linked polysaccharide
[30]. In several known b-1,3-glucanases the structural
position equivalent to the tyrosine residue was occupied by
a glycine or an aspartate [30]. In TcGlu2—and in the other
b-1,3-1,4-glucanases highly homologous to TcGlu2—an
aspartate residue (D156) is present at the position corre-
sponding to Y177 (Fig. 1b).
Putative phosphorylation sites were observed in the
specific b-1,3-1,4-glucanase conserved domain of TcGlu2
(Fig. 1a). As already known, the phosphorylation is a post-
translational modification involved in signaling, regulation
of the protein function, protein stability and sub-cellular
localization [37, 38]. Some PR protein, such as the CaPR-
10 from hot pepper (Capsicum annuum) was phosphory-
lated in response to the biotic stress due to the tobacco
mosaic virus (TMV-P0), and this post-translational
Table 1 Peptides of TcGlu2 after analysis by mass spectrometry
Peptide number Amino acid sequence Identity (%) E-value Organism Accession number Function
1 LAVLLVSDVFGYDAPNLR 82 1.10-04 Elaeis guineensis ACF06491 b-1,3-1,4-glucanase
2 LVIDALK – – – – –
3 EALIPAAVLLHPSFVTVDDIK 94 1.10-06 R. communis XP002524812 b-1,3-1,4-glucanase
4 VPIAILGAEIDQLSPPALVK 85 1.10-06 R. communis XP002516793 b-1,3-1,4-glucanase
Fig. 5 Effect of TcGlu2 on Moniliophthora perniciosa growth. PBphosphate buffer; CP control proteins corresponding to purified
protein extract from E. coli expressing pET28a without insert
Mol Biol Rep
123
modification altered the CaPR-10 function [39]. The
expression of active recombinant TcGlu2 protein in a
bacterial system is supported by the absence of putative
N-glycosylation sites (N-X-S/T type) [40] in the amino
acid sequence (Fig. 1a).
To our knowledge a small number of physiological data
related to b-1,3-1,4-glucanases were encountered, and
unfortunately, TcGlu2 presented few identity with those for
which such physiological reports exist. For example,
TcGlu2 presents 8 % of identity with Glb2, a b-1,3-1,4-
Fig. 6 Membrane permeability
of Moniliophthora perniciosahyphae. TcPR-10 was incubated
with M. perniciosa hyphae at 25
or 4 �C with or without NaN3,
and membrane integrity was
checked by using PI uptake
assay. Hyphae were observed
under fluorescence microscope
at 30 min and 1 h and images
were taken under phase contrast
and PI filter
Mol Biol Rep
123

glucanase isoenzyme II from Hordeum vulgare (accession
number AAA32962; [41]), 8 % of identity with the b-1,4-
glucanase (TcGlu1) from T. cacao (accession number
AY487173 [42]), and 15.5 and 13 % of identity with
OsEGL1 and OsEGL2 b-1,3-1,4-glucanases from rice,
respectively (accession numbers AAV37460 and
BAB85436, respectively; [13]) (data not shown). Glb2, as
other barley glucanases [3, 11] is possibly involved in cell
wall organization during seed germination [41]. In cacao,
the expression of TcGlu1 was induced in leaves after
treatment by the Necrosis and Ethylene inducing Protein 1
and after infection by the fungus Phytophthora megakarya,
and may be associated with senescence process [42]. In
rice, the OsEGL1 gene was expressed in response to
methyl jasmonate, abscissic acid, ethephron and mechani-
cal wounding [13].
Recombinant incomplete TcGlu2 inhibited the M. per-
niciosa mycelium growth (Fig. 5) suggesting an antifungal/
antimicrobial activity of this enzyme. However, in the lit-
erature, antimicrobial activity was mainly observed in b-1,3-
glucanases. For example, Ji and Kuc [43] showed that a b-
1,3-glucanase from cucumber inhibited spore germination
and mycelium growth of the necrotrophic fungus Colleto-
trichum lagenarium, and Sela-Buurlage et al. [44] showed
that the b-1,3-glucanase from tobacco was active against the
fungus Fusarium solani, resulting in lysis of the hyphal tips
and growth inhibition. In both works, there was a synergistic
activity of the b-1,3-glucanase studied and a chitinase from
the same organism (cucumber vs tobacco) [43, 44].
According to Jach et al., [45] co-overexpression of genes
coding for b-1,3-glucanase and chitinase from barley in
tobacco plants led to a significant increase of plant defense
when infected by the fungus Rhizoctonia solani in compar-
ison to the non-transformed plants. Similar results were
observed by introducing a maize b-1,3-glucanase (M-GLU)
into tomato and by submitting the transformed plant to
inoculation with Alternaria solani spores (causal agent of the
early blight disease); compared to control (non-transformed)
plants, the transgenic lines carrying M-GLU showed
enhanced resistance to early blight disease [46]. According
to the sequence structure of TcGlu2 (D instead of Y residue
as the position 156 of the substrate recognition site) and to the
its functional characteristics as antifungal molecule, this
glucanase presents more similarities with b-1,3-glucanase
than with the b-1,3-1,4-glucanase family to which it belongs,
but on the other hand, the structure of the active site of
TcGlu2 suggests a b-1,3 and 1,4 glucanase activity. It is
interesting to note that the 198–239 region of the protein is
not necessary for the antifungal activity. Moreover, to our
knowledge, this is the first study of a cacao b-1,3-1,4-glu-
canase expression in heterologous system and the first
analysis showing the antifungal activity of a b-1,3-1,4-glu-
canase, in particular against M. perniciosa. The use of PI,
known to be a marker of cell viability, revealed that the
antifungal activity of TcGlu2 may be related to cell mem-
brane damages (Fig. 6). Because the PI fluorescence was
observed only 1 h after TcGlu2 application (Fig. 6b), it may
be suggested that the destruction of the membrane was a
consequence of substrate-specific recognition of TcGlu2 to
polymers of the fungus hyphae, and that TcGlu2 may have
acted by permeabilization or by direct destruction of the
fungus membrane, as observed for other antifungal proteins
in plants [43]. Furthermore, according to the data presented
here, TcGlu2 may be a good candidate to increase T. cacao
resistance to the fungus M. perniciosa by a similar approach
as the one developed for a class I chitinase gene [15].
Acknowledgments This research was supported by the Conselho
Nacional de Desenvolvimento Cientıfico e Tecnologico (CNPq,
Brazil) and the International Foundation for Science (IFS). The work
of DSB and TPS was supported by the Fundacao de Amparo a Pes-
quisa da Bahia (FAPESB, Brazil). We thank Dr. Alan Pomella (Grupo
Farroupilha, Brazil) for kindly providing the M. perniciosa strain and
Dr. Claudia Fortes Ferreira (Embrapa, Cruz das Almas-BA, Brazil)
for critical reading of the manuscript.
References
1. Minic Z (2008) Physiological roles of plant glycoside hydrolases.
Planta 227:723–740
2. Cantarel BL, Coutinho PM, Rancurel C, Bernard T, Lombard V,
Henrissat B (2009) The Carbohydrate-Active EnZymes database
(CAZy): an expert resource for glycogenomics. Nucleic Acids
Res 37:D233–D238
3. Hrmova M, Fincher GB (2001) Structure-function relationships
of b-D-glucan endo- and exohydrolases from higher plants. Plant
Mol Biol 47:73–91
4. P Antoni (2000) Bacterial 1,3-1,4-b-glucanases: structure, func-
tion and protein engineering. Biochim Biophys Acta 1543:
361–382
5. Nakatani Y, Lamont IL, Cutfield JF (2010) Discovery and char-
acterization of a distinctive Eex-1,3/1,4-b-glucanase from the
marine bacterium Pseudoalteromonas sp. strain BB1. Appl
Environ Microbiol 76:6760–6768
6. Niu D, Zhou X-x, Yuan T-y, Lin Z-w, Ruan H, Li W-f (2010)
Effect of the C-terminal domains and terminal residues of cata-
lytic domain on enzymatic activity and thermostability of li-
chenase from Clostridium thermocellum. Biotechnol Lett 32:
963–967
7. Huang H, Yang P, Luo H, Tang H, Shao N, Yuan T, Wang Y, Bai
Y, Yao B (2008) High-level expression of a truncated 1,3-1,4-b-
D-glucanase from Fibrobacter succinogene in Pichia pastoris by
optimization of codons and fermentation. Appl Microbiol Bio-
technol 78:95–103
8. Takeda T, Takahashi M, Nakanishi-Masuno T, Nakano Y, Saitoh
H, Hirabuchi A, Fujisawa S, Terauchi R (2010) Characterization
of endo-1,3-1,4-b-glucanases in GH family 12 from Magnaportheoryzae. Appl Microbiol Biotechnol 88:1113–1123
9. Yang S, Qiaojuan Y, Jiang Z, Fan G, Wang L (2008) Biochemical
characterization of a novel thermostable b-1,3–1,4-glucanase
(lichenase) from Paecilomyces thermophila. J Agric Food Chem
56:5345–5351
10. Celestino K, Cunha R, Felix C (2006) Characterization of a beta-
glucanase produced by Rhizopus microsporus var. microsporus,
Mol Biol Rep
123

and its potential for application in the brewing industry. BMC
Biochem 7:23
11. Slakeski N, Fincher GB (1992) Developmental regulation of
(1 ? 3,1 ? 4)-b-glucanase gene expression in barley : tissue-
specific expression of individual isoenzymes. Plant Physiol
99:1226–1231
12. Takeda H, Sugahara T, Kotake T, Nakagawa N, Sakurai N (2010)
Sugar treatment inhibits IAA-induced expression of endo-1,3:1,4-
b-glucanase EI transcripts in barley coleoptile segments. Physiol
Plant 139:413–420
13. Akiyama T, Jin S, Yoshida M, Hoshino T, Opassiri R, Ketudat
Cairns JR (2009) Expression of an endo-(1,3;1,4)-b-glucanase in
response to wounding, methyl jasmonate, abscisic acid and eth-
ephon in rice seedlings. J Plant Physiol 166:1814–1825
14. Pungartnik C, da Silva AC, de Melo SA, Gramacho KP, de
Mattos Cascardo JC, Brendel M, Micheli F, da Silva Gesteira A
(2009) High-affinity copper transport and Snq2 export permease
of Saccharomyces cerevisiae modulate cytotoxicity of PR-10
from Theobroma cacao. Mol Plant Microbe Interact 22:39–51
15. Maximova S, Marelli J-P, Young A, Pishak S, Verica J, Guiltinan
M (2006) Over-expression of a cacao class I chitinase gene in
Theobroma cacao L. enhances resistance against the pathogen
Colletotrichum gloeosporioides. Planta 224:740–749
16. Menezes SP, dos Santos JL, Cardoso THS, Pirovani CP, Micheli
F, Noronha FSM, Alves AC, Faria AMC, da Silva Gesteira A
(2012) Evaluation of the allergenicity potential of TcPR-10
protein from Theobroma cacao. PLoS ONE 7:e37969
17. Micheli F, Guiltinan M, Gramacho KP, Wilkinson MJ, Figueira
AVdO, Cascardo JCdM, Maximova S, Lanaud C (2010) Func-
tional genomics of cacao. In: Jean-Claude K, Michel D (eds)
Advances in botanical research, Chapt 3. Academic Press, Lon-
don, pp 119–177
18. Gesteira AS, Micheli F, Carels N, Da Silva A, Gramacho K,
Schuster I, Macedo J, Pereira G, Cascardo J (2007) Comparative
analysis of expressed genes from cacao meristems infected by
Moniliophthora perniciosa. Ann Bot 100:129–140
19. Argout X, Salse J, Aury J-M, Guiltinan MJ, Droc G, Gouzy J,
Allegre M, Chaparro C, Legavre T, Maximova SN, Abrouk M,
Murat F, Fouet O, Poulain J, Ruiz M, Roguet Y, Rodier-Goud M,
Barbosa-Neto JF, Sabot F, Kudrna D, Ammiraju JSS, Schuster
SC, Carlson JE, Sallet E, Schiex T, Dievart A, Kramer M, Gelley
L, Shi Z, Berard A, Viot C, Boccara M, Risterucci AM, Guignon
V, Sabau X, Axtell MJ, Ma Z, Zhang Y, Brown S, Bourge M,
Golser W, Song X, Clement D, Rivallan R, Tahi M, Akaza JM,
Pitollat B, Gramacho K, D’Hont A, Brunel D, Infante D, Kebe I,
Costet P, Wing R, McCombie WR, Guiderdoni E, Quetier F,
Panaud O, Wincker P, Bocs S, Lanaud C (2011) The genome of
Theobroma cacao. Nat Genet 43:101–108
20. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller
W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucleic Acids
Res 25:3389–3402
21. Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence align-
ment through sequence weighting, position-specific gap penalties
and weight matrix choice. Nucleic Acids Res 22:4673–4680
22. Blom N, Gammeltoft S, Brunak S (1999) Sequence and structure-
based prediction of eukaryotic protein phosphorylation sites.
J Mol Biol 294:1351–1362
23. Quevillon E, Silventoinen V, Pillai S, Harte N, Mulder N, Ap-
weiler R, Lopez R (2005) InterProScan: protein domains identi-
fier. Nucleic Acids Res 33:W116–W120
24. Henikoff S, Henikoff JG (1992) Amino acid substitution matrices
from protein blocks. Proc Natl Acad Sci USA 89:10915–10919
25. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan
PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R,
Thompson JD, Gibson TJ, Higgins DG (2007) Clustal W and
Clustal X version 2.0. Bioinformatics 23:2947–2948
26. Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phy-
logenetic inference under mixed models. Bioinformatics 19:
1572–1574
27. Arnold K, Bordoli L, Kopp J, Schwede T (2006) The SWISS-
MODEL workspace: a web-based environment for protein
structure homology modelling. Bioinformatics 22:195–201
28. Bordoli L, Kiefer F, Arnold K, Benkert P, Battey J, Schwede T
(2008) Protein structure homology modeling using SWISS-
MODEL workspace. Nat Protocols 4:1–13
29. Notredame C, Higgins DG, Heringa J (2000) T-coffee: a novel
method for fast and accurate multiple sequence alignment. J Mol
Biol 302:205–217
30. Tsai L-C, Chen Y-N, Shyur L-F (2008) Structural modeling of
glucanase–substrate complexes suggests a conserved tyrosine is
involved in carbohydrate recognition in plant 1,3-1,4-b-D-glu-
canases. J Comput Aided Mol Des 22:915–923
31. Weiner SJ, Kollman PA, Case DA, Singh UC, Ghio C, Alagona
G, Profeta S, Weiner P (1984) A new force field for molecular
mechanical simulation of nucleic acids and proteins. J Am Chem
Soc 106:765–784
32. Weiner SJ, Kollman PA, Nguyen DT, Case DA (1986) An all
atom force field for simulations of proteins and nucleic acids.
J Comput Chem 7:230–252
33. Laskowski RA, MacArthur MW, Moss DS, Thornton JM (1993)
PROCHECK: a program to check the stereochemical quality of
protein structures. J Appl Crystallogr 26:283–291
34. Melo F, Feytmans E (1997) Novel knowledge-based mean force
potential at atomic level. J Mol Biol 267:207–222
35. Melo F, Feytmans E (1998) Assessing protein structures
with a non-local atomic interaction energy. J Mol Biol 277:
1141–1152
36. Filho DF, Pungartnik C, Cascardo JCM, Brendel M (2006)
Broken hyphae of the basidiomycete Crinipellis perniciosa allow
quantitative assay of toxicity. Curr Microbiol 52:407–412
37. Durek P, Schmidt R, Heazlewood JL, Jones A, MacLean D,
Nagel A, Kersten B, Schulze WX (2010) PhosPhAt: the Ara-bidopsis thaliana phosphorylation site database. An update.
Nucleic Acids Res 38:D828–D834
38. Lin J, Xie Z, Zhu H, Qian J (2010) Understanding protein
phosphorylation on a systems level. Brief Funct Genomics
9:32–42
39. Park C-J, Kim K-J, Shin R, Park JM, Shin Y-C, Paek K-H (2004)
Pathogenesis-related protein 10 isolated from hot pepper func-
tions as a ribonuclease in an antiviral pathway. Plant J
37:186–198
40. Song W, Henquet MGL, Mentink RA, van Dijk AJ, Cordewener
JHG, Bosch D, America AHP, van der Krol AR (2011) N-gly-
coproteomics in plants: perspectives and challenges. J Proteomics
74:1463–1474
41. Wolf N (1991) Complete nucleotide sequence of a Hordeumvulgare gene encoding (1 ? 3, 1 ? 4)-b-glucanase isoenzyme
II. Plant Physiol 96:1382–1384
42. Bailey B, Bae H, Strem M, Antunezdemayolo G, Guiltinan M,
Verica J, Maximova S, Bowers J (2005) Developmental expres-
sion of stress response genes in leaves and their response to Nep1
treatment and a compatible infection by Phytophthora megak-arya. Plant Physiol Biochem 43:611–622
43. Ji C, Kuc J (1996) Antifungal activity of cucumber beta-1,3-
glucanase and chitinase. Physiol Mol Plant Pathol 49:
257–265
44. Sela-Buurlage MB, Ponstein AS, Bres-Vloemans SA, Melchers
LS, van den Elzen PJM, Cornelissen BJC (1993) Only specific
tobacco (Nicotiana tabacum) chitinases and [beta]-1,3-glucanases
exhibit antifungal activity. Plant Physiol 101:857–863
Mol Biol Rep
123

45. Jach G, Gornhardt B, Mundy J, Logemann J, Pinsdorf E, Leah R,
Schell J, Maas C (1995) Enhanced quantitative resistance against
fungal disease by combinatorial expression of different barley
antifungal proteins in transgenic tobacco. Plant J 8:97–109
46. Schaefer S, Gasic K, Cammue B, Broekaert W, van Damme E,
Peumans W, Korban S (2005) Enhanced resistance to early blight
in transgenic tomato lines expressing heterologous plant defense
genes. Planta 222:858–866
Mol Biol Rep
123
Related Documents











