RESEARCH Recombinant Baculovirus as a Highly Potent Vector for Gene Therapy of Human Colorectal Carcinoma: Molecular Cloning, Expression, and In Vitro Characterization Arghya Paul • Barbara A. Jardin • Arun Kulamarva • Meenakshi Malhotra • Cynthia B. Elias • Satya Prakash Published online: 9 February 2010 Ó Springer Science+Business Media, LLC 2010 Abstract Present therapeutic strategies for most cancers are restricted mainly to the primary tumors and are also not very effective in controlling metastatic states. Alternatively, gene therapy can be a potential option for treating such cancers. Currently mammalian viral-based cancer gene therapy is the most popular approach, but the efficacy has been shown to be quite low in clinical trials. In this study, for the first time, the insect cell-specific baculovirus Autographa californica multiple nucleopolyhedrovirus (AcMNPV) has been evaluated as a vector for gene delivery to colorectal cancer cells. Experiments involving factorial design were employed to study the individual and combined effects of different parameters such as multiplicity of infection (MOI), viral incubation time and epigenetic factors on transduction efficiency. The results demonstrate that baculovirus gene delivery system holds immense potential for development of a new generation of highly effective virotherapy for colo- rectal, as well as other major carcinomas (breast, pancreas, and brain), and offers significant benefits to traditional ani- mal virus-based vectors with respect to safety concerns. Keywords Colorectal cancer Baculovirus Gene therapy BacMam Transduction Biosafety Therapeutic delivery Introduction Colorectal cancer is the third most commonly diagnosed cancer and the third leading cause of cancer death in both men and women in the United States, with about 148,810 new cases and 49,960 deaths expected in 2008 out of which 72% of cases arise in the colon and about 28% in the rectum [1]. The most common practice to treat such can- cers is surgical resection of the primary tumor. But this often turns out to be inefficient because of the tumor’s progress to metastases after attempted curative resections. Although adjuvant chemotherapy or chemo-radiation had shown some promises, still the long-term survival from colon cancer has remained almost static [2]. Thus, new measures are urgently required. Gene therapy represents a logical approach to address such concerns, which may act as an adjunct or conventional treatment [3–5]. Cancer gene therapy involves the inclusion of foreign therapeutic genes into cancer cells resulting in temporary suppression or complete eradication of the tumor. However, the success of this therapy relies greatly on the efficiency and selectivity at which therapeutic genes are introduced into the tumor cells without causing any adverse side effects. Thus efficient delivery and good biosafety profiles are two very Arun Kulamarva and Meenakshi Malhotra contributed equally to this work. A. Paul B. A. Jardin A. Kulamarva M. Malhotra S. Prakash Biomedical Technology and Cell Therapy Research Laboratory, Department of Biomedical Engineering and Artificial Cells and Organs Research Centre, Faculty of Medicine, McGill University, 3775 University Street, Montreal, QC H3A 2B4, Canada C. B. Elias Process Development, Sanofi Pasteur, Connaught Campus, 1755 Steeles Avenue West, Toronto, ON M2R 3T4, Canada C. B. Elias S. Prakash (&) Centre for Biorecognition and Biosensors, Biomedical Engineering and Artificial Cells and Organs Research Centre, Faculty of Medicine, McGill University, 3775 University Street, Montreal, QC H3A 2B4, Canada e-mail: [email protected] Mol Biotechnol (2010) 45:129–139 DOI 10.1007/s12033-010-9248-7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH
Recombinant Baculovirus as a Highly Potent Vector for GeneTherapy of Human Colorectal Carcinoma: Molecular Cloning,Expression, and In Vitro Characterization
Arghya Paul • Barbara A. Jardin • Arun Kulamarva •
Meenakshi Malhotra • Cynthia B. Elias •
Satya Prakash
Published online: 9 February 2010
� Springer Science+Business Media, LLC 2010
Abstract Present therapeutic strategies for most cancers
are restricted mainly to the primary tumors and are also not
very effective in controlling metastatic states. Alternatively,
gene therapy can be a potential option for treating such
cancers. Currently mammalian viral-based cancer gene
therapy is the most popular approach, but the efficacy has
been shown to be quite low in clinical trials. In this study, for
the first time, the insect cell-specific baculovirus Autographa
californica multiple nucleopolyhedrovirus (AcMNPV) has
been evaluated as a vector for gene delivery to colorectal
cancer cells. Experiments involving factorial design were
employed to study the individual and combined effects of
different parameters such as multiplicity of infection (MOI),
viral incubation time and epigenetic factors on transduction
efficiency. The results demonstrate that baculovirus gene
delivery system holds immense potential for development of
a new generation of highly effective virotherapy for colo-
rectal, as well as other major carcinomas (breast, pancreas,
and brain), and offers significant benefits to traditional ani-
mal virus-based vectors with respect to safety concerns.
Keywords Colorectal cancer � Baculovirus � Gene
therapy � BacMam � Transduction � Biosafety �Therapeutic delivery
Introduction
Colorectal cancer is the third most commonly diagnosed
cancer and the third leading cause of cancer death in both
men and women in the United States, with about 148,810
new cases and 49,960 deaths expected in 2008 out of which
72% of cases arise in the colon and about 28% in the
rectum [1]. The most common practice to treat such can-
cers is surgical resection of the primary tumor. But this
often turns out to be inefficient because of the tumor’s
progress to metastases after attempted curative resections.
Although adjuvant chemotherapy or chemo-radiation had
shown some promises, still the long-term survival from
colon cancer has remained almost static [2]. Thus, new
measures are urgently required. Gene therapy represents a
logical approach to address such concerns, which may act
as an adjunct or conventional treatment [3–5].
Cancer gene therapy involves the inclusion of foreign
therapeutic genes into cancer cells resulting in temporary
suppression or complete eradication of the tumor. However,
the success of this therapy relies greatly on the efficiency and
selectivity at which therapeutic genes are introduced into the
tumor cells without causing any adverse side effects. Thus
efficient delivery and good biosafety profiles are two very
Arun Kulamarva and Meenakshi Malhotra contributed equally
to this work.
A. Paul � B. A. Jardin � A. Kulamarva � M. Malhotra �S. Prakash
Biomedical Technology and Cell Therapy Research Laboratory,
Department of Biomedical Engineering and Artificial Cells
and Organs Research Centre, Faculty of Medicine,
McGill University, 3775 University Street,
Montreal, QC H3A 2B4, Canada
C. B. Elias
Process Development, Sanofi Pasteur, Connaught Campus,
1755 Steeles Avenue West, Toronto, ON M2R 3T4,
Canada
C. B. Elias � S. Prakash (&)
Centre for Biorecognition and Biosensors, Biomedical
Engineering and Artificial Cells and Organs Research Centre,
Faculty of Medicine, McGill University, 3775 University Street,
Montreal, QC H3A 2B4, Canada
e-mail: [email protected]
Mol Biotechnol (2010) 45:129–139
DOI 10.1007/s12033-010-9248-7

important criteria during the selection of a delivery system.
Till date adenoviruses have been commonly used for a wide
variety of gene therapy applications; however, its transduc-
tion efficiency in solid tumors has been limited, resulting in
little evidence supporting significant clinical benefits [6–8].
Although researchers are trying to implement replication-
competent oncolytic viruses for cancer gene therapy [9–15],
its self-perpetuating nature in mammalian cells remains a
major concern for safety. Consequently, gene delivery into
tumors although conceptually elegant, has not yet achieved
its expected potential. Under such circumstances, targeting
tumor cells with insect-specific viruses, such as baculovi-
ruses, can become a potentially powerful tool. Since bac-
uloviruses are highly restricted insect viruses, which only
enter but not replicate in mammalian cells, therefore they
work best with priority to the efficacy and safety concerns of
cancer gene therapy. The natural promoters of these viruses
are not active in mammalian cells, as reported, there had been
no detection of expression from the polyhedrin promoter in
mammalian cells like Huh7 cells [16]. On the other hand, it
has shown to deliver genetic materials in vitro and in vivo,
into both dividing and nondividing mammalian cells, which
can be beneficial in case of prostate cancer that propagates at
a slower rate [17]. Earlier, recombinant baculoviruses have
shown to produce functional monoclonal antibody, by
infecting insect cells, which can recognize human colorectal
carcinoma cells [18]. Lyophilized preparation of insect cells
infected with baculovirus producing interferon-beta can
suppress the progressive growth of established orthotopic
tumors of colon cancer cells and occult liver metastasis in
vivo [19]. As known through literature, a combination
treatment of baculovirus gene therapy and chemotherapy
could trigger apoptosis in osteogenic sarcoma cells demon-
strating the potential of baculovirus as a cancer gene therapy
tool [20]. Recently, baculoviruses have also revealed its
potential to suppress gastric cancers in xenografted nude
mice [21].
Due to its growing popularity and potential scope in
cancer gene therapy, we have focused on exploring and
optimizing the combinatorial effects of different transduc-
tion factors using factorial design experiments in order to
enhance the therapeutic effects of baculovirus gene delivery
in treating cancers, particularly colorectal cancers. In this
study, we have investigated and compared the applicability
of baculoviruses as a potent gene therapy agent for some
unexplored, yet common cancers in detail. To perform these
studies, we have constructed recombinant baculoviruses
carrying Monster Green Fluorescent Protein as the reporter
transgenes, driven by the human cytomegalovirus (CMV)
immediate-early promoter. As this insect cell originated
baculovirus can express its transgene in mammalian cells,
driven by the CMV promoter, it is also known as BacMam
virus.
Materials and Methods
Cells and Media
Human colon cancer cells (SW480), breast cancer cells
(SkBr3), neuroblastoma cancer cells (Neuro2a), hepato-
cellular carcinoma cells (HepG2), and pancreatic carcinoma
cells line (PANC-1) were obtained from ATCC. SW480
was maintained in Leibovitz’s L-15 medium, SkBr3 was
maintained in McCoy’s 5A medium, Neuro2a in Eagle’s
Minimum Essential Medium (EMEM), HepG2 in Minimum
Essential Medium Alpha (MEM), and PANC-1 cells were
grown in Modified Eagle’s Medium (DMEM). All the
media were obtained from ATCC and supplemented with
10% Fetal Bovine Serum (FBS; Gibco, Invitrogen). The
cells were routinely maintained as stationary cultures in
75 cm2 cell culture flasks (Corning, NY), incubated at
37 �C in a controlled environment with 5% CO2, except in
case of SW480 where it did not require additional supply of
CO2. Spodoptera frugiperda (Sf9) insect cells (Invitrogen
Life Technologies, Carlsbad, CA) were maintained at 27 �C
in SF900-III SFM (Gibco, Life Technologies, Grand Island,
NY, USA) medium without FBS, using routine procedures
[22]. The Sf9 cells were maintained in exponential growth
phase and subcultured twice per week in 75 cm2 cell culture
flasks with 10 mL as working volume. Larger volumes were
prepared in 125 mL shaker flasks (Erlenmeyer, Corning) as
suspension culture, which were agitated at 115 rpm in a
horizontal incubator shaker with a working volume of
25 mL.
Cloning, Generation, and Quantification
of Recombinant MGFP-BacMam Baculoviruses
(Bac-MGFP)
The phMGFP vector carrying the Monster Green Fluores-
cent Protein (MGFP) and pVL1392 transfer vector, as
depicted in Fig. 1, were procured from Promega (Madison,
WI, USA) and BD Biosciences, respectively. Both the
phMGFP vector, harboring the PCMV promoter with hMGFP
gene, and the pVL1392 vector were digested with BglII and
XbaI restriction enzymes (New England Biolabs, Berverly,
MA). The excised PCMV-hMGFP gene and digested pVL1392
vector were purified and a basic ligation reaction with T4
DNA ligase (Promega) was performed to subclone the PCMV-
hMGFP gene into the pVL1392 transfer vector by directional
cloning method to generate the pVL1392-PCMV-PMGFP
transfer vector. The recombinant transfer vector was
amplified by transformation into the high efficiency DH5 acompetent E. coli (Invitrogen) by heat shock method. The
plasmids were then extracted and purified with QIAprep spin
miniprep kit (Qiagen Sciences, MD).
130 Mol Biotechnol (2010) 45:129–139

To generate the recombinant MGFP baculoviruses,
1 9 106 Sf9 cells were seeded in six well plates. A trans-
fection mixture comprised of 1 lg of the constructed
BacMam transfer vector, linearized baculovirus DNA (BD
Baculogold) and 6 lL of Cellfectin transfection reagent,
diluted in 200 lL media (Invitrogen Life Technologies,
Carlsbad, CA) was prepared. The mixture was incubated
for 45 min at room temperature and then was co-trans-
fected to the seeded cells. After 5 h of incubation at 27 �C,
the transfection media was replaced with Sf900 III media
and cells were further incubated at 27 �C for the recom-
bination event to occur inside the transfected insect cells
between the transfer vector and the linearized viral DNA.
This generates the newly formed active recombinant viru-
ses that start their replication process from within the insect
cells. Eventually the virus buds out of the cell and begins a
secondary round of infection in the nearby cells and so on.
96 h post-transfection (hpi), the cell cultures were col-
lected, centrifuged at 1000g for 10 min and then the
supernatant containing the baculovirus was filtered through
a 0.22-lm membrane and stored at 4 �C (P0 Virus Stock).
The P0 stock was further amplified from time to time by
using routine procedures as described previously [23].
Briefly, Sf9 cells were grown to mid-exponential phase and
diluted to 2 9 106 cells/mL with fresh SF900III medium.
The cultures were infected with P0 virus stock at a multi-
plicity of infection (MOI = plaque forming unit [pfu]/mL)
of 0.1. The supernatant containing the recombinant bacu-
lovirus was harvested 72 hpi by centrifugation. The viral
titre (pfu/mL) of the amplified viral stock was then deter-
mined using the Baculovirus Fast Plax Titer Kit (Novagen,
Madison, WI) according to the manufacturer’s protocol.
Bac-MGFP Transduction of Mammalian Cell Lines
Fluorescent expression levels of transduced cells were
quantified using plate reader Victor3 Multi Label Plate
Counter (Perkin Elmer, USA). Then, 2 9 104 cells were
seeded in each well in a 96 well plate and transduced with
100 lL PBS (no Ca and Mg) containing Bac-MGFP at
MOI ranging from 10 to 1000. After 8 h of incubation of
the cells with viruses at 25 �C, the cells were washed twice
with PBS and replenished with 200 lL of fresh medium
with or without 10 mM sodium butyrate (NaBu). The
relative fluorescence units of each well were measured
24 h post-transduction (hpt). This experiment was done in
triplicates, and the mean was calculated. Another set of
similar experiment was set up to find out the percentage of
cells transduced with Bac-MGFP, 24 hpt. In this set, the
monolayers were washed and the average percentage of
green fluorescent cells per high-power field were quantified
using fluorescence microscope (Nikon Eclipse TE2000-U);
five fields were counted per well and the mean value was
considered as the percentage transduced value.
To detect the fluorescence expression at the transcript
level, the cells were seeded in 6-well plates at 0.5 9
106 cells/well. Prior to transduction, the cell culture med-
ium was replaced by 500 lL of PBS containing Bac-MGFP
virus at varied MOI ranging from 100 to 1000. After 8 h of
viral incubation at 25 �C, the viral solution was removed
and the cells were washed twice with PBS and replenished
either with 2 mL fresh medium or with medium supple-
mented with 10 mM NaBu.
Application of Stat-Ease Design-Expert Software
for Optimization of Transduction Conditions
Optimization of the transduction procedure taking different
factors into account was done using the Design-Expert
software Version 7.1 from Stat-Ease. This software tool
allows the screening of multilevel factorial designs that
helps to look for the critical parameters which may indi-
vidually or synergistically affect a process [24]. In our
study, (i) MOI (ranging from 10 to 1000), (ii) NaBu con-
centration (ranging from 0 to 10 mM), and (iii) viral
incubation time (ranging from 2 to 8 h) were selected as
the three parameters affecting the transduction process
directly or indirectly. For this, SW480 cells were taken and
PVL1392(9639 bp)
phMGFP(4701 bp)
AmpR
PolyhedrinPromoter
MCS(BglII and XbaI)
CMV Promoter
hMGFP (684bp)
AmpR
BglII
XbaI A B
ColE ori
Fig. 1 Schematic
representation of starting
vectors and cloning strategy
for the PCMV-hMGFP/pVL1392
plasmid. The reporter gene
hMGFP was excised along with
the CMV promoter from
phMGFP Vector (a) using BglII
and XbaI restriction enzymes
and inserted in the Multiple
Cloning Site (MCS) of the
baculovirus transfer vector
pVL1392 (b)
Mol Biotechnol (2010) 45:129–139 131

transduced with baculovirus in 96 well plates. Based on
preliminary results, a two level factorial design with 20
experimental runs and four centre points was implemented.
The chosen was percentage of cells expressing GFP. The
statistical analysis of variance (ANOVA) for the responses
(transduction percentage) was generated by the above
mentioned software. Significant factors were established
where p \ 0.01.
Transcriptional Activity Analysis by RT-PCR
For detection of MGFP gene expression, total RNA was
extracted from transduced, mock-transduced and control
cells cultured in 6-well plates using RNA Extraction kit
(Qiagen) according to the manufacturer’s instructions. The
obtained RNA was then reverse transcribed to corresponding
cDNA using the Qiagen’s Reverse Transcription (RT) kit
following the supplied instructions. Further, PCR was per-
formed on the reverse transcribed product using Taq DNA
Polymerase (Invitrogen). To detect specific gene products,
the following forward (FP) and reverse primers (RP) were
used in PCR reactions. For Human Glyceraldehyde-
3-phosphate dehydrogenase (GAPDH) gene (112-bp prod-
uct), the FP was 50-TCAAGGGCATCCTGGGCTAC-30 and
RP was 50-AAGTGGTCGTTGAGGGCAATG-30; for the
MGFP gene (133-bp product) the FP was 50-GATGCAGC
GCAAGACCCTAAAG-30 and the RP was 50-GGTCTTG
AAGTCGCAGCGGTAG-30. Amplifications were carried
out for 25 cycles at 94 �C for 35 s (denaturation), 55 �C for
35 s (annealing), and 72 �C for 25 s (extension).
Results
Generation of Recombinant Baculoviruses
by Co-Transfection in Insect Cells
The pVL1392-PCMV-PMGFP recombinant transfer vector
was constructed by using the two vectors depicted in Fig. 1.
The confirmation of the correct insert in desired orientation
in the transfer vector was done by digesting the vector with
different sets of enzyme combinations and examining the
right band sizes through the agarose gel electrophoresis. The
purified pVL1392-PCMV-PMGFP transfer vector was then
co-transfected with linearized baculovirus DNA in the
insect cells to generate recombinant baculoviruses harboring
PCMV–MGFP genes. After 96 h, P0 viral stock was har-
vested from the culture collection of supernatant following
centrifugation. P0 viral stock was used to infect Sf9 cells in
a T-25 flask at MOI of 0.1 that resulted in a higher titre P1
viral stock, which was further amplified to achieve P2 viral
titre of 2.2 9 109pfu/mL as determined by the viral titration
kit.
Combinatorial Effects of Viral Dose, Viral Incubation
Time, and NaBu Concentration on Transduction
Efficiency: Factorial Matrix Design Experiments
The full factorial design was implemented to identify the
parameters having significant effects on increasing the
transduction efficiency. The factors considered were MOI,
viral incubation time and NaBu concentration at two levels
in a factorial design. The response factor studied was on
the transduction efficiency of SW480 colorectal cancer
cells. The three factors and their corresponding ranges are
mentioned in Table 1. Results under the condition of 8 h
viral incubation are shown in Fig. 2a. The two-dimensional
graph demonstrates that the MOI and NaBu concentration
both have significant positive effects on the transduction
percentage (response). Also their combined effect had a
greater impact in boosting the response compared to each
of them individually. This is clear from the graph (Fig. 2a)
where at MOI of around 750 and NaBu concentration of
5 mM, the transduction efficiency was around 70%,
whereas in absence of NaBu the efficiency dropped down
to approximately 50%. Graph of Fig. 2b depicts the pattern
of increment of the response level with increasing incu-
bation time and NaBu concentration under constant MOI of
1,000. The pattern suggests that increasing the incubation
time can have significant positive effect on the transduc-
tion, as it allows more viral particles to enter the host cell,
which in turn allows more expression of the transgene. The
transduction can be further boosted by addition of epige-
netic factor NaBu. This is evident from the graph, where
with a constant incubation time of 8 h, transduction effi-
ciency of around 73% can be increased to 85% in presence
of NaBu. Figure 2c explains that keeping the NaBu con-
centration constant, there can also be an increase in
transduction efficiency with increasing MOI and incubation
time. The two parameters are also dependant on each other
for maximum transduction. Like in the graph (Fig. 2c), a
low MOI (257.5) with less incubation time of 2 h gives us
low efficiency of approximately 37%, which can be
increased to around 49% with an increase in viral incuba-
tion time to 5 or 8 h. On the other hand, an increase in MOI
from 257.5 to 505 increases the transduction efficiency
from the range of 37–49% at constant incubation time of
Table 1 Factors used for the factorial design experiment and their
ranges
Factors Name Units Factors level range
Low Mean High
A MOI pfu/cell 10 505 1000
B NaBu mM 0 5 10
C Incubation time hour 2 5 8
132 Mol Biotechnol (2010) 45:129–139
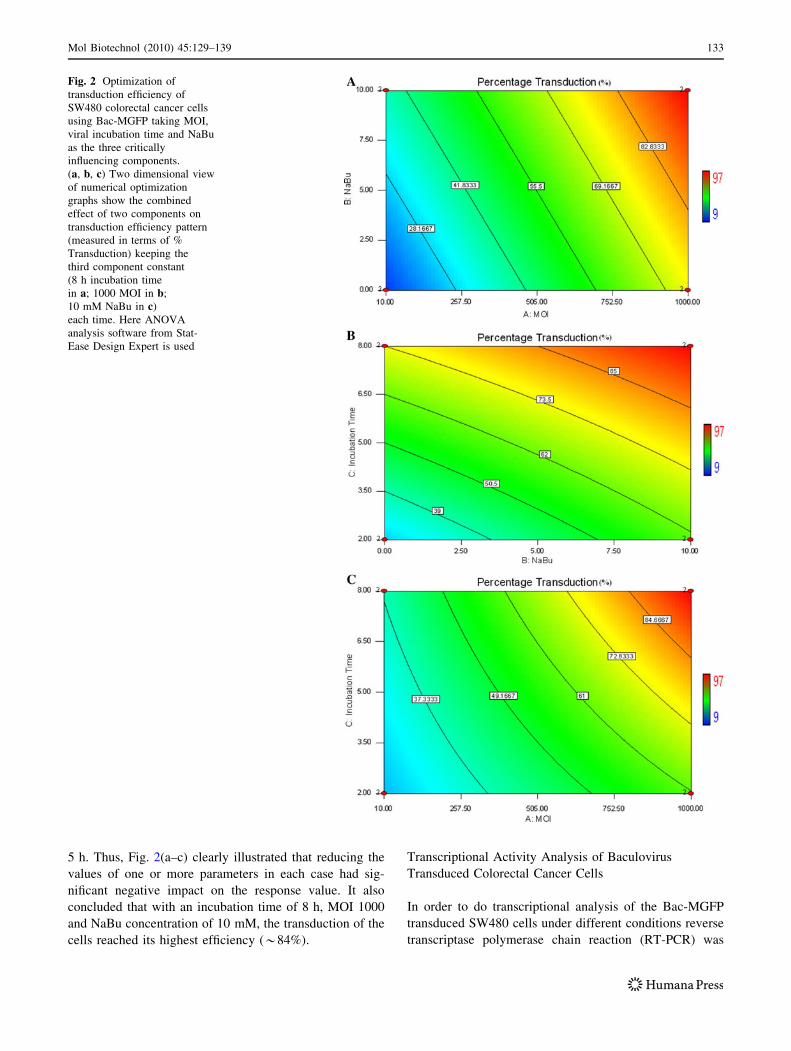
5 h. Thus, Fig. 2(a–c) clearly illustrated that reducing the
values of one or more parameters in each case had sig-
nificant negative impact on the response value. It also
concluded that with an incubation time of 8 h, MOI 1000
and NaBu concentration of 10 mM, the transduction of the
cells reached its highest efficiency (*84%).
Transcriptional Activity Analysis of Baculovirus
Transduced Colorectal Cancer Cells
In order to do transcriptional analysis of the Bac-MGFP
transduced SW480 cells under different conditions reverse
transcriptase polymerase chain reaction (RT-PCR) was
Fig. 2 Optimization of
transduction efficiency of
SW480 colorectal cancer cells
using Bac-MGFP taking MOI,
viral incubation time and NaBu
as the three critically
influencing components.
(a, b, c) Two dimensional view
of numerical optimization
graphs show the combined
effect of two components on
transduction efficiency pattern
(measured in terms of %
Transduction) keeping the
third component constant
(8 h incubation time
in a; 1000 MOI in b;
10 mM NaBu in c)
each time. Here ANOVA
analysis software from Stat-
Ease Design Expert is used
Mol Biotechnol (2010) 45:129–139 133

performed. At 24 hpt, the transcription of MGFP gene was
done using RT-PCR. The 133 bp fragment was only found
in the sample of transduced SW480 cells indicating the
successful transcription of MGFP gene in cells (Fig. 3).
Further evaluation of the expression levels in SW480 cells
was determined by comparing the band intensities in
transduced cells, which revealed that with increase in
amount of MOI treatment, the amount of MGFP transcript
also increases. It also shows that addition of NaBu further
complements the increase in transcript levels. These data
were further reconfirmed by the fluorescence photographs
of the corresponding cells (Fig. 3a). The results were in
accordance with what we have obtained with protein level
expression as shown in Fig. 3a and b for colon cells.
Time Course Profile of Transgene Expression
in Transduced Colorectal Cancer Cells and Its
Dependence on Viral Incubation Time
In order to determine the duration of gene expression
mediated by Bac-MGFP, the SW480 cells were transduced
with an MOI of 1000 in a 96-well plate. Moreover, to find
out the dependence of gene expression on viral incubation
time the SW480 cells were transduced with Bac-MGFP at
varying incubation time (2, 5, and 8 h). The fluorescein
plate readings of each well were taken from time to time
(12 h, day 1, day 3, day 6, day 9, day 12, day 15, day 18,
day 21) using plate reader. The obtained average fluores-
cein values of each well were then normalized to that of the
wells having 8 h of viral incubation 24 hpt. The results as
represented in Fig. 4a shows a rapid high expression in the
cells on day 1, with higher incubation time giving the
higher expression (25% for 2 h; 57% for 5 h; 100% for
8 h). The expression gradually reduces with time and by
day 12 it almost ceases for the cells with viral incubation of
2 h. But with incubations of 5 and 8 h, the normalized
expression at day 12 still remained much higher to 10%
and 23% approximately. Thus the cells with 8 h incubation
time showed better normalized gene expression as well as
better persistence of the transgene with time, as studied in
21 days. These analyses were further reconfirmed by the
fluorescence photographs of the cells (Fig. 4b).
Fig. 3 a Transduction of Bac-MGFP on SW480 colorectal cancer
cells. The cells were transduced with Bac-MGFP for 8 h in PBS at
25 �C with and without NaBu treatment at an MOI of 50 and 100.
Transduced cells were photographed 24 hpt with fluorescence
microscope. Fluorescence images in the absence (top) and presence
(bottom) of phase contrast illumination are shown. b RT-PCR
analysis of MGFP gene expression 24 hpt with Bac-MGFP in Sw480.
The cells were given a viral incubation time of 8 h with different MOI
(50, 100, 500, and 1000).The cells treated with NaBu are compared
with nontreated cells. Controls include no template control for PCR
(NTC), no reverse transcriptase control for reverse transcription
reaction (-RT) and mock-infected cell control (Mock) for transduction
procedure. Amplification of housekeeping GAPDH gene serves an
internal control for the efficiency of reverse transcription reaction.
Molecular weight (Mw) markers are low range DNA ladder and the
sizes of the amplicon are indicated with arrows
134 Mol Biotechnol (2010) 45:129–139

Baculovirus-Mediated Superior Gene Transfer
to Colorectal Cancer Cells in Comparison to Breast,
Pancreas, and Neuronal Carcinomas
To explore the possibility of using baculovirus-mediated
gene delivery for a wide range of cancer cells, cell lines
from breast, pancreas, and neuronal tumor origins were
taken for study, apart from colorectal. The efficiency of
transduction, as evaluated by number of green fluorescent
cells, was determined 24 hpt and presented in Fig. 5
(in presence of NaBu) and Fig. 6 (in absence of NaBu).
The data show that the baculovirus vectors transduced
A
1
1
Nor
mal
ised
GF
P e
xpre
ssio
n (%
)
B
0
20
40
60
80
100
120
0
Day1 (5h)
3
8h
)
6 9Days po
5h
Day1 (8h)
9 12ost transductio
2h
D
15on
Day6 (8h)
18
Day
21
18 (8h)
Fig. 4 a Effect of viral
incubation time (in h) over time-
course profile of transgene
expression in SW480 cells. The
average expressed fluorescein
values were normalized to that
of cells with 1 day post
transduction, having 8 h of viral
incubation at 25 �C, and are
represented as normalized mean
expression in percentage value.
The cells were transduced with
Bac-MGFP for various
incubation periods. The data
represent the mean of three
independent culture
experiments ± Standard
Deviation (SD). b Transduced
cells with different incubation
times (denoted in brackets as 5
and 8 h) were photographed at
different time points(day 1, day
6, day 18) with fluorescence
microscope. Fluorescence
images in the absence (top) and
presence (bottom) of phase
contrast illumination are shown
Fig. 5 Quantification of
fluorescent cells (percentage
of total) with relation to MOI
(treated with 10 mM NaBu),
24 h post-transduction. SW480,
SkBr3, Neuro2a, and Panc1
cells were transduced with Bac-
MGFP at MOI of 100, 250, and
500 with viral incubation time
of 8 h at 25 �C. The cells were
then replenished with media
containing 10 mM NaBu. Data
represent the mean ± SD
Mol Biotechnol (2010) 45:129–139 135

SW480 cells with much higher efficiency than other cells.
At an MOI of 500, the baculovirus transduced 90 ± 2.5%
[standard deviation (SD), n = 3] and 37 ± 4.1% (SD,
n = 3) of the SW480 and Neuro2a cells, respectively, in
NaBu-treated cells. At MOI of 500, SkBr3 and Panc1 cells
show an efficiency of 40 ± 3.2% (SD, n = 3) and 31 ±
3.6% (SD, n = 3), respectively, in presence of NaBu.
SW480 cells showed almost equal transduction efficiency
with that of HepG2 cells, taken as a positive control
[25, 26]. The transduction efficiency appeared to have a
direct relationship with MOI in all the cell lines. To further
investigate, we checked the fluorescein expression of the
cells using plate reader 24 hpt as analyzed in Fig. 7. The
fluorescein expression level in SW480 cells treated with
10 mM NaBu was almost doubled as compared to NaBu
untreated cells. SW480 cells had three fold higher
expression levels in comparison to both Neuro2a and
SkBr3.
Discussions
In this report, we evaluated the application of baculovirus
gene therapy in diverse types of cancer (Table 2), where
the colorectal cancer cells showed highest transduction
efficiency in terms of GFP positive cells as well as its high
expression level as detected by fluorescein reading in the
plate reader. But leukemic cells like HL-60 (ATCC)
showed poor transduction with baculovirus vector (data not
shown). The study also focuses on investigating the critical
parameters that have individual as well as combinatorial
effects on the transduction level in the colorectal cancer
cells. The data show that the MOI, viral incubation time
and NaBu concentration have significant effects as mea-
sured by transduction efficiency and transgene expression
levels in the cells. We concluded that with an incubation
time of 8 h, MOI 1000 and NaBu concentration of 10 mM,
the transduction of the colorectal cancer cells reached its
highest efficiency. This data were also supported at the
transcript level as evaluated by RT-PCR in colon cancer
cells. The increase in viral incubation time also increased
the retention of transgene expression in transduced cells.
Similar observations were also noticed with other cancer
cells of different origins, which showed marked response in
transgene expression level and transduction efficiency with
varying viral dosage and treatment with NaBu.
The disparity in GFP expression among various cancer
cells was because each cell line has its unique ability to
Fig. 6 Quantification of
fluorescent cells (percentage
of total) with relation to MOI
(without treatment of 10 mM
NaBu), 24 hpt. SW480, SkBr3,
Neuro2a, and Panc1 cells were
transduced with Bac-MGFP at
MOI of 100, 250, and 500 with
viral incubation time of 8 h at
25 �C. The cells were then
replenished with media. Data
represent the mean ± SD
Fig. 7 Quantification of
fluorescent cells (fluoresccein
expression) with and without
10 mM NaBu treatment, 24 hpt.
SW480, SkBr3, Neuro2a, and
Panc1 cells were transduced
with Bac-MGFP at MOI of 500
with 8 h viral incubation time at
25 �C. The cells were then
replenished with media with or
without having NaBu in it. The
data represent the means ± SD
of triplicate samples
136 Mol Biotechnol (2010) 45:129–139

drive the CMV promoter to express the reporter gene.
Moreover, the uncoating efficiency of the baculovirus and
its genomic DNA movement into the host cell nucleus is
specific and unique to each cell-type. The colorectal cells
showed a rapid expression pattern with high-transduction
efficiencies which gradually diminished with time in a viral
incubation time dependant manner. The reason for this
transient expression could be attributed to the lack of nat-
ural mechanism of baculovirus to integrate with the host
genome, which is an inherent nature of most mammalian
viruses. Hence it does not exhibit long-term expression,
which arises out of genomic integration of the viral genome
into the host chromosome and its subsequent transfer to the
cell progeny. This is an important characteristic of the
insect cell-originating baculovirus with respect to its bio-
safety nature. Whereas most of the common mammalian
viral vectors have the probability to randomly integrate into
the host’s crucial genomic regions, thereby resulting in
unexpected tumerogenesis. Such transient expression can
be a limiting factor for cancer gene therapy, this problem
can be handled by using a hybrid baculovirus-adeno
associated viral (AAV) vector that harbors a gene cassette
flanked by AAV-inverted terminal repeats for prolonged
expression of transgene in host cells [27]. Moreover,
administration of multiple viral dosages can be another
solution to overcome the obstacle of transient expression.
Transduced cells exhibited marked increase in expres-
sion level with increase in viral dosage (MOI) and/or with
increase in viral exposure time. This was due to the
increase in MOI, wherein the virus to cell ratio increases
and thus in turn increases the chance of more number of
viral particles getting inside the cell and being transcribed.
Increasing the viral incubation time to more than 8 h did not
show any increase in transgene expression. This observa-
tion was understood as 8 h of incubation was optimal for
cellular uptake of viruses. Another reason could be that
extended viral exposure ([8 h) may pose a detrimental
effect on cells due to cytotoxicity. In order to detect the
false effects caused by down regulation of reporter gene
expression, induced by histone deacetylation condition of
the transduced cells expression, we performed studies at
both transcript and protein levels, in the presence and
absence of an epigenetic factor, NaBu. The most remark-
able effect of butyrate exhibited that epigenetic regulations
play an important role in influencing the transgene expres-
sion pattern. This chemical regulates histone deacetylation
by controlling the activation of histone deacetylase
enzymes [28]. It creates a histone hyperacetylation condi-
tion within the cells inducing a more relaxed chromatin
conformation conferring proper binding of transcription
factors to the promoter with consequential upregulation of
transgene expressions. That is why we noticed a marked
increase in transgene expression in all the transduced cell
lines in the presence of NaBu. Thus, we infer that epigenetic
phenomena, such as acetylation or compaction of chromatin
play an important role in baculovirus-mediated transgene
expression system. Proper chromatin state of the baculovi-
rus genome is thus crucial for efficient gene expression.
Despite its promising roles, baculovirus has still to over-
come some major obstacles, the lack of specific tumor tar-
geting being one of them. This has also been one of the
major reasons why the first generation nonreplicative ade-
noviruses showed low efficacy and disappointing clinical
trial data for cancer therapy [29]. Modifying the viral
envelope proteins to display tumor homing peptide ligands
on the baculovirus surface can be one way to increase
baculovirus transduction efficiency and circumvent the
problem [30].
There are several reports that have demonstrated the
broad range of cellular susceptibility of baculoviruses
toward human [31–34], porcine [35], rabbit [36], rodent
[37], avian [38], and fish [39] origins. This wide range of
permissive cell lines has encouraged researchers to extend
its applications toward cancer therapy. Current curative
strategies for most of the cancers are restricted to the pri-
mary tumor and early stages which are mainly treated by
surgical resection. As the effect of conventional treatments
like chemotherapy or radiotherapy to control metastatic
diseases are not sustained, therefore, gene therapy repre-
sents an alternative modality to treat cancer. Many safety
issues are under consideration with present mammalian
virus-based gene therapy trials [40]. On the contrary, insect
cell specific baculovirus plays an important role at this
juncture as a potential biologically safe gene delivery
Table 2 Baculovirus as a potential gene delivery vehicle for trans-
ducing different cancer cells
Origin Cell Line Gene therapy
Pancreas Panc-1
Neuronal Neuro2a
Colon/colorectal SW480
Breast SkBr3
Leukemic HL-60
Liver HepG2
Mol Biotechnol (2010) 45:129–139 137

vehicle. Fusing decay accelerating factor with baculovirus
gp64 gene under the control of polyhedrin promoter [41]
are shown to produce complement resistant baculoviruses.
This had actually resolved the major problem of in vivo
inactivation of baculovirus by the serum complements.
Cobra venom factor and anti-C5 antibody treatment as C
inhibitor [42] also shows to enhance transduction. Thus
baculovirus can emerge as an attractive gene delivery
system for cancer treatments [17, 43, 44].
Through this work, our data suggest for the first time
that baculovirus vectors could be used efficiently for
colorectal cancers. The future directions for this would be
the generation of recombinant baculovirus that can express
tumor suppressor genes like ATOH1 and APC for treating
colon cancers [45, 46], BRCA1 to treat breast cancers [47],
DPC4 for pancreatic cancer [48], p53 to treat hepatocar-
cinoma [49], and FKN to treat neuroblastoma [50]. Recent
concepts of targeting the colon cancer stem cells or using
suicide transgenes to induce antitumor effect are also some
of the promising alternative approaches where recombinant
baculovirus delivery system can be beneficial [27, 51].
Success of these studies will greatly improve the clinical
perspective of baculovirus-mediated gene therapy for can-
cer. Recombinant baculoviruses thus offers a potential
alternative to currently used adenoviral vectors for cancer
gene therapy. The increasing number of unique and specific
molecular targets for different cancer cells may prompt us
in future to design individualized baculoviruses for treating
respective cancer types with high efficacy. Such baculo-
viruses will not only make the delivery of genes restricted
and target specific, but will also increase the transduction
efficiency due to negligible loss to other nonspecific body
parts. This can further be boosted by the synergistic effect
of combination therapies comprising of radio-, chemo-, and
virotherapies.
Acknowledgments We gratefully acknowledge the assistance
received from the Natural Sciences and Engineering Research
Council of Canada (NSERC) to Dr. S. Prakash. A.P. acknowledges
financial support from Fonds de la recherche en sante du Quebec
(FRSQ), Quebec, Canada. A.K. acknowledges the Alexander Graham
Bell Post Graduate Scholarship—Doctoral from NSERC. M.M.
acknowledges the support of the McGill Faculty of Medicine Internal
Scholarship. The authors would like to thank B. Boulay, R. Tran, and
J. Montes from National Research Council Canada (NRC-BRI) for
their technical advice and assistance.
References
1. Jemal, A., Siegel, R., Ward, E., Hao, Y. P., Xu, J. Q., Murray, T.,
et al. (2008). Cancer statistics, 2008. Ca—A Cancer Journal forClinicians, 58, 71–96.
2. Midgley, R., & Kerr, D. (1999). Colorectal cancer. Lancet, 353,
391–399.
3. Birkenkamp-Demtroder, K., Christensen, L. L., Olesen, S. H.,
Frederiksen, C. M., Laiho, P., Aaltonen, L. A., et al. (2002). Gene
expression in colorectal cancer. Cancer Research, 62, 4352–
4363.
4. Palmer, D. H., Chen, M. J., & Kerr, D. J. (2002). Gene therapy for
colorectal cancer. British Medical Bulletin, 64, 201–225.
5. Yamamoto, T. Y., Okuda, J. O., Toyoda, M. T., & Tanigawa, N.
T. (2002). Gene therapy for colon cancer using survivin antisense
expressing replication-incompetent adenoviral vectors. Interna-tional Journal of Cancer, (Suppl 13), 125.
6. Hemminki, A. (2002). From molecular changes to customised
therapy. European Journal of Cancer, 38, 333–338.
7. Kanerva, A., & Hemminki, A. (2004). Modified adenoviruses for
cancer gene therapy. International Journal of Cancer, 110, 475–
480.
8. Chung-Faye, G. A., Chen, M. J., Green, N. K., Burton, A.,
Anderson, D., Mautner, V., et al. (2001). In vivo gene therapy for
colon cancer using adenovirus-mediated, transfer of the fusion
gene cytosine deaminase and uracil phosphoribosyltransferase.
Gene Therapy, 8, 1547–1554.
9. Bazan-Peregrino, M., Sainson, R., Carlisle, R., Waters, R., Harris,
A. L., Hernandez-Alcoceba, R., et al. (2008). Virotherapy
expressing regulatory angiogenic gene therapy in treatment of
breast cancer. Human Gene Therapy, 19, 415.
10. Cafferata, E. G., Maccio, D. R., Lopez, M. V., Viale, D. L.,
Carbone, C., Mazzolini, G., et al. (2009). A novel A33 promoter-
based conditionally replicative adenovirus suppresses tumor
growth and eradicates hepatic metastases in human colon cancer
models. Clinical Cancer Research, 15, 3037–3049.
11. Huang, Q., Ji, X., Zhang, J., Cheng, L., Wei, F., Li, H., et al.
(2009). Oncolytic adenovirus delivering herpes simplex virus
thymidine kinase suicide gene reduces the growth of human
retinoblastoma in an in vivo mouse model. Experimental EyeResearch, 89, 193–199.
12. Rognoni, E., Widmaier, M., Haczek, C., Mantwill, K., Holz-
muller, R., Gansbacher, B., et al. (2009). Adenovirus-based
virotherapy enabled by cellular YB-1 expression in vitro and in
vivo. Cancer Gene Therapy, 16, 753–763.
13. Wang, G., Li, G., Liu, H., Yang, C., Yang, X., Jin, J., et al.
(2009). E1B 55-kDa deleted, Ad5/F35 fiber chimeric adenovirus,
a potential oncolytic agent for B-lymphocytic malignancies.
Journal of Gene Medicine, 11, 477–485.
14. Ying, B., Toth, K., Spencer, J. F., Meyer, J., Tollefson, A. E.,
Patra, D., et al. (2009). INGN 007, an oncolytic adenovirus
vector, replicates in Syrian hamsters but not mice: comparison of
biodistribution studies. Cancer Gene Therapy, 16, 625–637.
15. Koyama, F., Sawada, H., Hirao, T., Fujii, H., Hamada, H., &
Nakano, H. (2000). Combined suicide gene therapy for human
colon cancer cells using adenovirus-mediated transfer of Esche-
richia coli cytosine deaminase gene and Escherichia coli uracil
phosphoribosyltransferase gene with 5-fluorocytosine. CancerGene Therapy, 7, 1015–1022.
16. Hofmann, C., Sandig, V., Jennings, G., Rudolph, M., Schlag, P.,
& Strauss, M. (1995). Efficient gene-transfer into human hepa-
tocytes by baculovirus vectors. Proceedings of the NationalAcademy of Sciences of the United States of America, 92, 10099–
10103.
17. Stanbridge, L. J., Dussupt, V., & Maitland, N. J. (2003). Bac-
uloviruses as Vectors for Gene Therapy against Human Prostate
Cancer. Journal of Biomedicine and Biotechnology, 2003, 79–91.
18. Nesbit, M., Fu, Z. F., Mcdonaldsmith, J., Steplewski, Z., &
Curtis, P. J. (1992). Production of a functional monoclonal-anti-
body recognizing human colorectal-carcinoma cells from a bac-
ulovirus expression system. Journal of Immunological Methods,151, 201–208.
138 Mol Biotechnol (2010) 45:129–139

19. Ozawa, S., Lu, W. X., Bucana, C. D., Kanayama, H. O., Shino-
hara, H., Fidler, I. J., et al. (2003). Regression of primary murine
colon cancer and occult liver metastasis by intralesional injection
of lyophilized preparation of insect cells producing murine
interferon-beta. International Journal of Oncology, 22, 977–984.
20. Song, S. U., & Boyce, F. M. (2001). Combination treatment for
osteosarcoma with baculoviral vector mediated gene therapy (p53)
and chemotherapy (adriamycin). Experimental and MolecularMedicine, 33, 46–53.
21. Huang, W., Tian, X. L., Wu, Y. L., Zhong, J., Yu, L. F., Hu, S. P.,
et al. (2008). Suppression of gastric cancer growth by baculovirus
vector-mediated transfer of normal epithelial cell specific-1 gene.
World Journal of Gastroenterology, 14, 5810–5815.
22. Mirzaei, M., Xu, Y., Elias, C. B., & Prakash, S. (2009). Nonviral
production of human interleukin-7 in spodoptera frugiperda
insect cells as a soluble recombinant protein. Journal of Bio-medicine and Biotechnology, 2009, 8 pp. doi:10.1155/2009/
637942.
23. Jardin, B. A., Montes, J., Lanthier, S., Tran, R., & Elias, C.
(2007). High cell density fed batch and perfusion processes for
stable non-viral expression of secreted alkaline phosphatase
(SEAP) using insect cells: comparison to a batch Sf-9-BEV
system. Biotechnology and Bioengineering, 97, 332–345.
24. Jardin, B. A., Zhao, Y., Selvaraj, M., Montes, J., Tran, R.,
Prakash, S., et al. (2008). Expression of SEAP (secreted alkaline
phosphatase) by baculovirus mediated transduction of HEK 293
cells in a hollow fiber bioreactor system. Journal of Biotechnol-ogy, 135, 272–280.
25. Bilello, J. P., Delaney, W. E., Boyce, F. M., & Isom, H. C.
(2001). Transient disruption of intercellular junctions enables
baculovirus entry into nondividing hepatocytes. Journal ofVirology, 75, 9857–9871.
26. Boyce, F. M., & Bucher, N. L. R. (1996). Baculovirus-mediated
gene transfer into mammalian cells. Proceedings of the NationalAcademy of Sciences of the United States of America, 93, 2348–
2352.
27. Wang, C. Y., & Wang, S. (2005). Adeno-associated virus
inverted terminal repeats improve neuronal transgene expression
mediated by baculoviral vectors in rat brain. Human GeneTherapy, 16, 1219–1226.
28. Marks, P. A., Rifkind, R. A., Richon, V. M., Breslow, R., Miller,
T., & Kelly, W. K. (2001). Histone deacetylases and cancer:
causes and therapies. Nature Reviews Cancer, 1, 194–202.
29. Rein, D. T., Breidenbach, M., & Curiel, D. T. (2006). Current
developments in adenovirus-based cancer gene therapy. FutureOncology, 2, 137–143.
30. Makela, A. R., Matilainen, H., White, D. J., Ruoslahti, E., &
Oker-Blom, C. (2006). Enhanced baculovirus-mediated trans-
duction of human cancer cells by tumor-homing peptides. Jour-nal of Virology, 80, 6603–6611.
31. Kronschnabl, M., Marschall, M., & Stamminger, T. (2002).
Efficient and tightly regulated expression systems for the human
cytomegalovirus major transactivator protein IE2p86 in permis-
sive cells. Virus Research, 83, 89–102.
32. Ma, L., Tamarina, N., Wang, Y., Kuznetsov, A., Patel, N.,
Kending, C., et al. (2000). Baculovirus-mediated gene transfer
into pancreatic islet cells. Diabetes, 49, 1986–1991.
33. Zeng, J., Du, J., Zhao, Y., Palanisamy, N., & Wang, S. (2007).
Baculoviral vector-mediated transient and stable transgene
expression in human embryonic stem cells. Stem Cells, 25, 1055–
1061.
34. Chuang, C. K., Wong, T. H., Hwang, S. M., Chang, Y. H., Chen,
G. Y., Chiu, Y. C., et al. (2009). Baculovirus transduction of
mesenchymal stem cells: in vitro responses and in vivo immune
responses after cell transplantation. Molecular Therapy, 17, 889–
896.
35. Grassi, G., Kohn, H., Dapas, B., Farra, R., Platz, J., Engel, S.,
et al. (2006). Comparison between recombinant baculo- and
adenoviral-vectors as transfer system in cardiovascular cells.
Archives of Virology, 151, 255–271.
36. Nakamichi, K., Matsumoto, Y., Tohya, Y., & Otsuka, H. (2002).
Induction of apoptosis in rabbit kidney cell under high-level
expression of bovine herpesvirus 1 U(s) ORF8 product. Intervi-rology, 45, 85–93.
37. Cheng, T., Xu, C. Y., Wang, Y. B., Chen, M., Wu, T., Zhang, J.,
et al. (2004). A rapid and efficient method to express target genes
in mammalian cells by baculovirus. World Journal of Gastro-enterology, 10, 1612–1618.
38. Ping, W., Ge, J., Li, S., Zhou, H., Wang, K., Feng, Y., et al.
(2006). Baculovirus-mediated gene expression in chicken pri-
mary cells. Avian Diseases, 50, 59–63.
39. Leisy, D. J., Lewis, T. D., Leong, J. A., & Rohrmann, G. F.
(2003). Transduction of cultured fish cells with recombinant
baculoviruses. Journal of General Virology, 84, 1173–1178.
40. Senior, K. (2000). Gene therapy: a rocky start to the new mil-
lennium. Molecular Medicine Today, 6, 93.
41. Huser, A., Rudolph, M., & Hofmann, C. (2001). Incorporation of
decay-accelerating factor into the baculovirus envelope generates
complement-resistant gene transfer vectors. Nature Biotechnol-ogy, 19, 451–455.
42. Hofmann, C., & Strauss, M. (1998). Baculovirus-mediated gene
transfer in the presence of human serum or blood facilitated by
inhibition of the complement system. Gene Therapy, 5, 531–536.
43. Clay, W. C., Condreay, J. P., Moore, L. B., Weaver, S. L.,
Watson, M. A., Kost, T. A., et al. (2003). Recombinant baculo-
viruses used to study estrogen receptor function in human oste-
osarcoma cells. Assay Drug Development Technologies, 1, 801–
810.
44. Song, J., Liang, C.-y., & Chen, X.-w. (2007). Baculovirus-med-
iated expression of p35 confers resistance to apoptosis in human
embryo kidney 293 cells. Virologica Sinica, 22, 389–396.
45. Bossuyt, W., Kazanjian, A., De Geest, N., Van Kelst, S., De
Hertogh, G., Geboes, K., et al. (2009). Atonal homolog 1 is a
tumor suppressor gene. Plos Biology, 7, 311–326.
46. Kerr, D. (2003). Clinical development of gene therapy for colo-
rectal cancer. Nature Reviews Cancer, 3, 615–622.
47. Obermiller, P. S., Tait, D. L., & Holt, J. T. (2000). Gene therapy
for carcinoma of the breast: therapeutic genetic correction strat-
egies. Breast Cancer Research, 2, 28–31.
48. Liu, F. (2001). SMAD4/DPC4 and pancreatic cancer survival—
Commentary re: M. Tascilar et al., The SMAD4 protein and
prognosis of pancreatic ductal adenocarcinoma. Clinical Cancer
Research, 7: 4115–4121, 2001. Clinical Cancer Research, 7,
3853–3856.
49. Vollmer, C. M., Ribas, A., Butterfield, L. H., Dissette, V. B.,
Andrews, K. J., Eilber, F. C., et al. (1999). p53 selective and
nonselective replication of an E1B-deleted adenovirus in hepa-
tocellular carcinoma. Cancer Research, 59, 4369–4374.
50. Zeng, Y., Jiang, J., Huebener, N., Wenkel, J., Gaedicke, G.,
Xiang, R., et al. (2005). Fractalkine gene therapy for neuroblas-
toma is more effective in combination with targeted IL-2. CancerLetters, 228, 187–193.
51. Todaro, M., Alea, M. P., Di Stefano, A. B., Cammareri, P.,
Vermeulen, L., Lovino, F., et al. (2007). Colon cancer stem cells
dictate tumor growth and resist cell death by production of
interleukin-4. Cell Stem Cell, 1, 389–402.
Mol Biotechnol (2010) 45:129–139 139
Related Documents











