2632 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643 DOI: 10.1002/eji.201242532 Recognition of fungal RNA by TLR7 has a nonredundant role in host defense against experimental candidiasis Carmelo Biondo ∗1 , Antonio Malara ∗1 , Alessandro Costa 1 , Giacomo Signorino 1 , Francesco Cardile 1 , Angelina Midiri 1 , Roberta Galbo 1 , Salvatore Papasergi 1 , Maria Domina 1 , Michela Pugliese 2 , Giuseppe Teti 1 , Giuseppe Mancuso 1 and Concetta Beninati 1 1 The Elie Metchnikoff Department, University of Messina, Messina, Italy 2 Department of Veterinary Public Health, University of Messina, Messina, Italy Despite convincing evidence for involvement of members of the Toll-like receptor (TLR) family in fungal recognition, little is known of the functional role of individual TLRs in antifungal defenses. We found here that TLR7 was partially required for the induction of IL-12 (IL-12p70) by Candida albicans or Saccharomyces cerevisiae. Moreover, the IL-12p70 response was completely abrogated in cells from 3d mice, which are unable to mob- ilize TLRs to endosomal compartments, as well as in cells from mice lacking either the TLR adaptor MyD88 or the IRF1 transcription factor. Notably, purified fungal RNA reca- pitulated IL-12p70 induction by whole yeast. Although RNA could also induce moderate TLR7-dependent IL-23 and tumor necrosis factor-alpha (TNF-α) secretion, TLR7 and other endosomal TLRs were redundant for IL-23 or TNF-α induction by whole fungi. Impor- tantly, mice lacking TLR7 or IRF1 were hypersusceptible to systemic C. albicans infection. Our data suggest that IRF1 is downstream of a novel, nonredundant fungal recognition pathway that has RNA as a major target and requires phagosomal recruitment of intracel- lular TLRs. This pathway differs from those involved in IL-23 or TNF-α responses, which we show here to be independent from translocation of intracellular TLRs, phagocytosis, or phagosomal acidification. Keywords: Dendritic cells Cytokines Fungal infections Inflammation Phagocytosis Supporting Information available online Introduction Fungal infections, such as those caused by Candida, Aspergillus, and Cryptococcus spp., are a major public health concern, with Candida albicans representing the most frequent pathogenic species. This yeast often asymptomatically colonizes human mucosal surfaces and is found predominantly in the oral cavity, the gastrointestinal tract, and the vagina [1]. During commen- sal carriage, there is a tenuous balance between the body’s own Correspondence: Dr. Giuseppe Mancuso e-mail: [email protected] defense systems and the remarkable ability of the organism to replicate in vivo. This equilibrium is frequently disrupted by envi- ronmental factors that promote fungal growth or weaken host defenses, leading to localized or systemic diseases [2]. Since the host immune status is the major factor that determines the transi- tion of C. albicans from commensalism to pathogenicity, a better understanding of the mechanisms underlying recognition of and responses to fungi is the key to developing alternative strategies to control these infections. ∗ These authors contributed equally to this work. C 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2632 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643DOI: 10.1002/eji.201242532
Recognition of fungal RNA by TLR7 has a nonredundantrole in host defense against experimental candidiasis
Carmelo Biondo∗1, Antonio Malara∗1, Alessandro Costa1,Giacomo Signorino1, Francesco Cardile1, Angelina Midiri1, RobertaGalbo1, Salvatore Papasergi1, Maria Domina1, Michela Pugliese2,Giuseppe Teti1, Giuseppe Mancuso1 and Concetta Beninati1
1 The Elie Metchnikoff Department, University of Messina, Messina, Italy2 Department of Veterinary Public Health, University of Messina, Messina, Italy
Despite convincing evidence for involvement of members of the Toll-like receptor (TLR)family in fungal recognition, little is known of the functional role of individual TLRs inantifungal defenses. We found here that TLR7 was partially required for the induction ofIL-12 (IL-12p70) by Candida albicans or Saccharomyces cerevisiae. Moreover, the IL-12p70response was completely abrogated in cells from 3d mice, which are unable to mob-ilize TLRs to endosomal compartments, as well as in cells from mice lacking either theTLR adaptor MyD88 or the IRF1 transcription factor. Notably, purified fungal RNA reca-pitulated IL-12p70 induction by whole yeast. Although RNA could also induce moderateTLR7-dependent IL-23 and tumor necrosis factor-alpha (TNF-α) secretion, TLR7 and otherendosomal TLRs were redundant for IL-23 or TNF-α induction by whole fungi. Impor-tantly, mice lacking TLR7 or IRF1 were hypersusceptible to systemic C. albicans infection.Our data suggest that IRF1 is downstream of a novel, nonredundant fungal recognitionpathway that has RNA as a major target and requires phagosomal recruitment of intracel-lular TLRs. This pathway differs from those involved in IL-23 or TNF-α responses, whichwe show here to be independent from translocation of intracellular TLRs, phagocytosis,or phagosomal acidification.
Keywords: Dendritic cells � Cytokines � Fungal infections � Inflammation � Phagocytosis
Supporting Information available online
Introduction
Fungal infections, such as those caused by Candida, Aspergillus,and Cryptococcus spp., are a major public health concern, withCandida albicans representing the most frequent pathogenicspecies. This yeast often asymptomatically colonizes humanmucosal surfaces and is found predominantly in the oral cavity,the gastrointestinal tract, and the vagina [1]. During commen-sal carriage, there is a tenuous balance between the body’s own
Correspondence: Dr. Giuseppe Mancusoe-mail: [email protected]
defense systems and the remarkable ability of the organism toreplicate in vivo. This equilibrium is frequently disrupted by envi-ronmental factors that promote fungal growth or weaken hostdefenses, leading to localized or systemic diseases [2]. Since thehost immune status is the major factor that determines the transi-tion of C. albicans from commensalism to pathogenicity, a betterunderstanding of the mechanisms underlying recognition of andresponses to fungi is the key to developing alternative strategiesto control these infections.
∗These authors contributed equally to this work.
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2012. 42: 2632–2643 Immunity to infection 2633
Anti-fungal defenses are initiated by the activation of germ-line encoded receptors (pathogen recognition receptors (PRRs))after recognition of a relatively small number of highly conservedmicrobial components (pathogen-associated molecular patterns(PAMPs)). By this mechanism, each PRR links the recognition ofa specific PAMP with the selective activation of a defined set oftranscription factors [3]. Considerable progress has been recentlymade in the identification of PRRs and PAMPs involved in anti-fungal defenses [4,5]. Beta-glucan, which is absent in animal cells,but is a major component of the fungal cell wall, is an importantrecognition target [6]. Many PRRs, including dectin-1 [7], scav-enger receptors [8], and complement receptor 3 [9], are capableof binding β-glucan. The signaling cascade triggered by interac-tions between particulate glucan and dectin-1 involves the sequen-tial activation of spleen tyrosine kinase (Syk), CARD9, and ofthe NF-κB and NFAT transcription factors. This pathway leads tophagocytosis, the “respiratory burst”, and cytokine gene induc-tion. The importance of this pathway in anti-fungal host defenseshas been demonstrated in experimental infections [10,11] andis corroborated by the association between increased susceptibil-ity to fungal infection and mutations in human genes encodingfor CARD9 [12]. The Syk/CARD9 pathway is also targeted byother lectin-type PRRs, such as dectin-2, which recognizes cell-wall mannans [13]. Much attention has been devoted to the abil-ity of fungi to activate Toll-like receptors (TLRs) and to the abilityof the latter to cooperate with lectin-type receptors in immuneresponses [14–16]. TLR engagement triggers signaling cascadesinvolving intracellular adaptors, such as MyD88 and TRIF, whichresult in the activation of several transcription factors, includ-ing NF-κB and interferon regulatory factors (IRFs). An importantrole of TLR-mediated recognition in anti-fungal host defenses issuggested by the extreme susceptibility to infection of MyD88-deficient mice [14,17–19]. However, the in vivo role of singleTLRs is uncertain [4,5]. Moreover, the fungal PAMPs responsiblefor TLR stimulation remain largely undefined, although O-linkedmannans and phospholipomannan from C. albicans have been pro-posed as TLR4 [20] and TLR2 [21] ligands, respectively.
Anti-fungal defenses crucially rely on the balanced productionof two key cytokines, IL-12p70 and IL-23, which display profounddifferences in the type of responses that they can elicit in cells ofthe innate and adaptive immune system. For example, IL-12p70and IL-23 induce the production of IFN-γ and IL-17, respectively,in T cells. It has been suggested that the production of IL-12p70and IL-23 are reciprocally regulated through the activation orco-activation of various TLRs and lectin-type receptors [4,5].However, little is known of the role of individual TLRs in suchactivities, especially in the context of infection with whole fungi, asopposed to stimulation with purified, nonfungal PRR agonists. Weshow here that TLR7-mediated sensing of fungal RNA leads to theproduction of a number of important cytokines, such as IL-12p70,IL-23, and tumor necrosis factor-alpha (TNF-α). Moreover,TLR7 was required for the induction of IL-12p70, but not IL-23or TNF-α, in the context of whole yeast stimulation. We furthershow that TLR7 cooperates with TLR9 in targeting IRF1, atranscription factor that has not been previously associated with
anti-fungal responses. Notably, lack of TLR7 or IRF1 was asso-ciated with increased susceptibility to experimental C. albicansinfection.
Results
Yeast RNA stimulates the production of IL-12p70,IL-23, and TNF-α
Our previous studies indicated that recognition of yeast RNAresults in the induction of IFN-β [22]. However, it is presentlyunclear whether fungal RNA is also capable of inducing the pro-duction of other primary cytokines, such as IL-12p70, IL-23, andTNF-α, which play a central role in anti-fungal defenses [23–25].Since macrophages and dendritic cells are the major cell types ofthe innate immune system, purified C. albicans RNA was tested forits ability to induce cytokine responses in bone marrow-derivedin vitro-differentiated dendritic cells (BMDCs) or macrophages(BMDMs). The RNA properties were compared with those of well-characterized fungal PAMPs, such as C. albicans DNA and depletedzymosan, a cell wall preparation consisting of particulate β-glucanthat is free of contaminating TLR agonists.
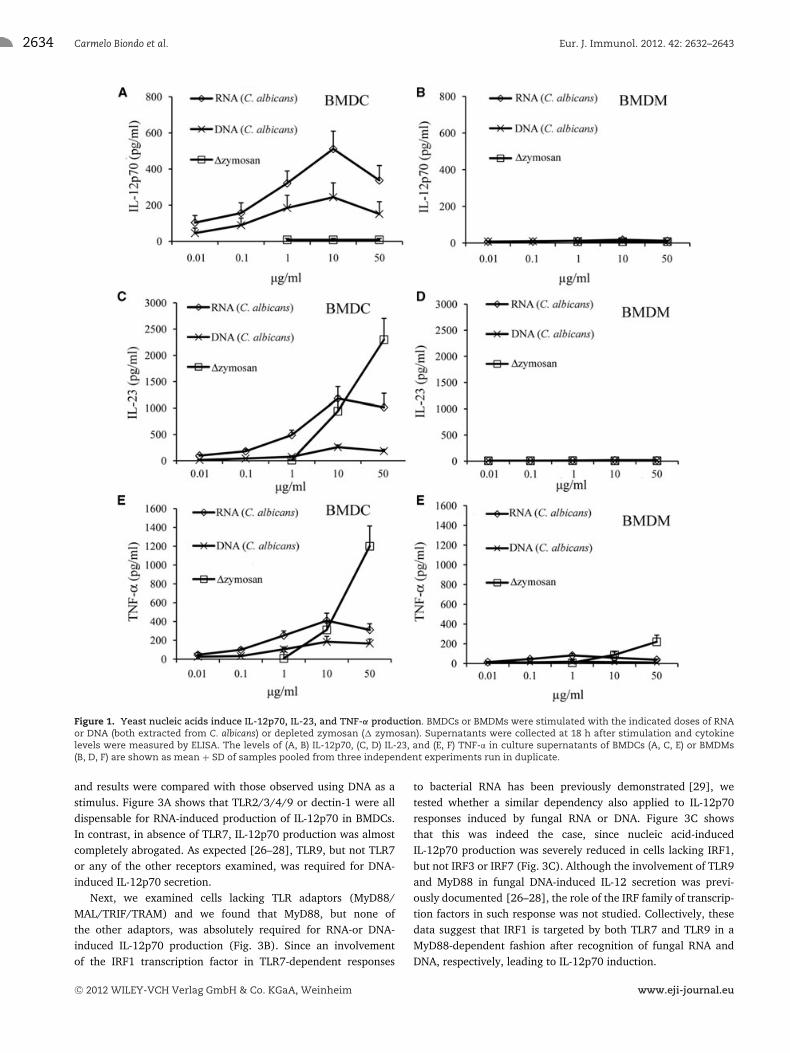
As shown in Figure 1, C. albicans RNA induced significant,dose-dependent elevations in IL-12p70, IL-23, and TNF-α levelsin BMDCs, but not in BMDMs, with an optimal stimulating doseof 10 μg/mL. C. albicans DNA also induced IL-12p70, IL-23, andTNF-α production in BMDCs, although cytokine levels were con-siderably lower than those observed after RNA stimulation. Incontrast, zymosan was totally unable to induce IL-12p70 in eitherBMDMs or BMDCs, although it did induce IL-23 and TNF-α eleva-tions in BMDCs (Fig. 1). Similar results were obtained in parallelexperiments when using, in place of depleted zymosan, depletedcurdlan, which is also a purified β-glucan preparation, or whenusing Saccharomyces cerevisiae RNA in place of C. albicans RNA(data not shown). This first set of data indicates that fungal RNAis able to induce the secretion of IL-12, IL-23, and TNF-α in BMDCs,but not in BMDMs.
To ascertain whether these cytokines were transcriptionallyregulated, we measured mRNA expression in BMDCs at differenttime points after stimulation with C. albicans RNA. As shown inFig. 2, significant elevations of IL-12p40, IL-12p35, IL-23p19, andTNF-α mRNA levels were observed. Such elevations were alreadyevident at 1 h postinfection, peaked at 6 h, and rapidly declinedthereafter. This data indicates that cytokine responses induced byfungal RNA are transcriptionally regulated.
TLR7, MyD88, and IRF1 are required for RNA-inducedIL-12p70 production
Next, it was of interest to identify the signaling pathways respon-sible for RNA-induced cell activation. To this end, we first usedC. albicans RNA to stimulate cells from mice lacking differentTLRs or dectin-1. RNA-induced IL-12p70 release was measured
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
2634 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643
Figure 1. Yeast nucleic acids induce IL-12p70, IL-23, and TNF-α production. BMDCs or BMDMs were stimulated with the indicated doses of RNAor DNA (both extracted from C. albicans) or depleted zymosan (� zymosan). Supernatants were collected at 18 h after stimulation and cytokinelevels were measured by ELISA. The levels of (A, B) IL-12p70, (C, D) IL-23, and (E, F) TNF-α in culture supernatants of BMDCs (A, C, E) or BMDMs(B, D, F) are shown as mean + SD of samples pooled from three independent experiments run in duplicate.
and results were compared with those observed using DNA as astimulus. Figure 3A shows that TLR2/3/4/9 or dectin-1 were alldispensable for RNA-induced production of IL-12p70 in BMDCs.In contrast, in absence of TLR7, IL-12p70 production was almostcompletely abrogated. As expected [26–28], TLR9, but not TLR7or any of the other receptors examined, was required for DNA-induced IL-12p70 secretion.
Next, we examined cells lacking TLR adaptors (MyD88/MAL/TRIF/TRAM) and we found that MyD88, but none ofthe other adaptors, was absolutely required for RNA-or DNA-induced IL-12p70 production (Fig. 3B). Since an involvementof the IRF1 transcription factor in TLR7-dependent responses
to bacterial RNA has been previously demonstrated [29], wetested whether a similar dependency also applied to IL-12p70responses induced by fungal RNA or DNA. Figure 3C showsthat this was indeed the case, since nucleic acid-inducedIL-12p70 production was severely reduced in cells lacking IRF1,but not IRF3 or IRF7 (Fig. 3C). Although the involvement of TLR9and MyD88 in fungal DNA-induced IL-12 secretion was previ-ously documented [26–28], the role of the IRF family of transcrip-tion factors in such response was not studied. Collectively, thesedata suggest that IRF1 is targeted by both TLR7 and TLR9 in aMyD88-dependent fashion after recognition of fungal RNA andDNA, respectively, leading to IL-12p70 induction.
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2012. 42: 2632–2643 Immunity to infection 2635
Figure 2. Kinetics of IL-12p40, IL-12p35, IL-23p19, and TNF-α mRNAexpression in BMDCs stimulated with C. albicans nucleic acids. BMDCsfrom C57BL/6 were stimulated with 10 μg/mL of C. albicans RNA orDNA or CpG B (1 μg/mL). IL-12p40, IL-12p35, IL-23p19, and TNF-α mRNAexpression was measured at the indicated times by RT-PCR. Data arepresented as “fold” increases in mRNA expression relative to uninfectedcells and are from one experiment representative of three.
IRF1 is not involved in RNA-induced TNF-α and IL-23responses
In further studies, we examined signaling requirements for RNA-induced TNF-α and IL-23 production. In these experiments, weused, as a control stimulus, depleted zymosan, which in pre-
vious experiments selectively induced these cytokines, but notIL-12p70 (Fig. 1). TLR7 and MyD88 were essential for the pro-duction of either IL-23 (Supporting Information Fig. 1) or TNF-α(Supporting Information Fig. 2) following RNA stimulation. Incontrast, none of the TLRs or the TLR adaptors examined, includ-ing MyD88, were required for depleted zymosan-induced IL-23 orTNF-α release. Rather, the latter responses were largely dectin-1-dependent (Supporting Information Fig. 1 and 2). Moreover, nei-ther IRF1, IRF3, or IRF7 were required for TNF-α or IL-23 pro-duction in response to RNA, DNA, or zymosan. Thus, IL-23 andTNF-α induction by C. albicans RNA required TLR7 and MyD88,but not IRF1.
Contribution of the TLR7 pathway to cytokineinduction by whole yeast
Since the data presented above indicated that TLR7 was absolutelyrequired for RNA-induced responses, it was of interest to assess therelative contribution of this receptor in the context of whole organ-ism stimulation. Live C. albicans was not used, since it was previ-ously found to produce significant cell toxicity in BMDCs, even atvery low multiplicities of infection or in the presence of high-dosefluconazole [22]. In contrast, the closely related [30] model yeastS. cerevisiae, which is also an opportunistic pathogen [31,32],was devoid of any cell toxicity [22]. After observing thatlive S. cerevisiae potently induced IL-12p70, IL-23, and TNF-αin a dose-dependent fashion, we assessed the signaling require-ments for these responses using BMDCs lacking specific signalingfactors (Fig. 4). Both TLR7 and TLR9, but not dectin-1, were atleast partially required for IL-12p70 responses to whole organ-isms. Moreover, cells lacking the TLR adaptor MyD88 or the tran-scription factor IRF1 were totally unable to produce IL-12p70in response to yeast (Fig. 4). In sharp contrast, IL-23 or TNF-αinduction by S. cerevisiae was independent from TLR7, TLR9,or the IRF1-transcription factor, while largely requiring dectin-1(Fig. 4). Yeast lysates in complex with the cationic lipid car-rier DOTAP recapitulated, in a dose-dependent way, the MyD88-dependent induction of IL-12p70 noted with live S. cerevisiae.Pretreatment of these fungal lysates with RNAse almost completelyabrogated induction of IL-12p70, whereas DNAse treatment wascomparatively less effective and proteinase K treatment was totallyineffective (Supporting Information Fig. 3). Moreover, combinedtreatment with RNase and DNase almost completely suppressedthe IL-12p70-inducing ability of extracts.
Interestingly, IL-23 and TNF-α, induction was partially MyD88-dependent, in agreement with the observation that various TLRagonists can collaborate with dectin-1 agonists in the induc-tion of optimal IL-23 [33] or TNF-α levels [34]. Similar signalingrequirements were found when using heat-killed C. albicans inplace of live S. cerevisiae as a stimulus (Supporting InformationFig. 4), although the latter stimulus was considerably more potentthan killed C. albicans at inducing cytokines.
Collectively, this data suggested that IL-12p70 production inresponse to whole yeast requires a TRL7- and TLR9-initiated
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

2636 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643
Figure 3. Signal requirements for C. albicansnucleic acids-stimulated IL-12p70 in BMDCs.BMDCs lacking various (A) PRRs, (B) TLR adap-tors, or (C) IRFs were stimulated with C. albi-cans RNA or DNA (both at 10 μg/mL). Escherichiacoli LPS (10 ng/mL), CL264 (5 μg/mL) and CpG B(1 μg/mL) were used as controls. Supernatantswere collected at 18 h after stimulation andassayed for IL-12p70 concentration by ELISA.Data are expressed as mean + SD of samplespooled from three independent observations,each conducted on a different animal, run induplicate. *p < 0.05 versus WT BMDCs by one-way analysis of variance and the Student’s-Keuls-Newman test.
pathway involving MyD88 and IRF1. Although stimulation withyeast nucleic acids did result in TLR7/9-dependent TNF-α andIL-23 secretion, these TLRs did not apparently make a significantcontribution to the overall ability of whole fungi to induce thesecytokines.
Role of yeast phagocytosis and TLR translocation toendosomes
Since TLR7 and TLR9 are endosomal receptors, we investigatedwhether IL-12p70 responses were induced by yeast in the absenceof functional UNC93B1, a chaperone protein that mediates thetranslocation of intracellular TLRs (including TLR3/7/8/9) to theendosomal compartment. To this end, we used BMDCs from 3dmice that have a point mutation in a transmembrane domainof UNC93B1, which renders the protein incapable of interactingwith intracellular TLRs [35–37]. TLR7/9 double knock-out micewere also used in these experiments. Notably, IL-12p70 responseswere totally abrogated in the absence of functional UNC93B1 orin cells lacking both TLR7 and TLR9, while neither IL-23 norTNF-α responses were affected (Fig. 5). Similarly, cytochalasin D,an agent that disrupts actin microfilaments and prevents phago-
cytosis, totally abrogated the release of IL-12p70, but not IL-23or TNF-α, by BMDCs after stimulation with S. cerevisiae (Fig. 6).In addition, similar effects were observed after BMDCs treatmentwith bafilomycin A, a drug that prevents phagosomal acidifica-tion. Thus phagocytosis, phagosomal acidification, and TLR7/9translocation to the endosomal compartment were all requiredfor the production of IL-12p70, but not IL-23 or TNF-α in responseto fungal recognition.
Role of TLR7, TLR9, UNC93B1, and IRF1 inanti-Candida defenses
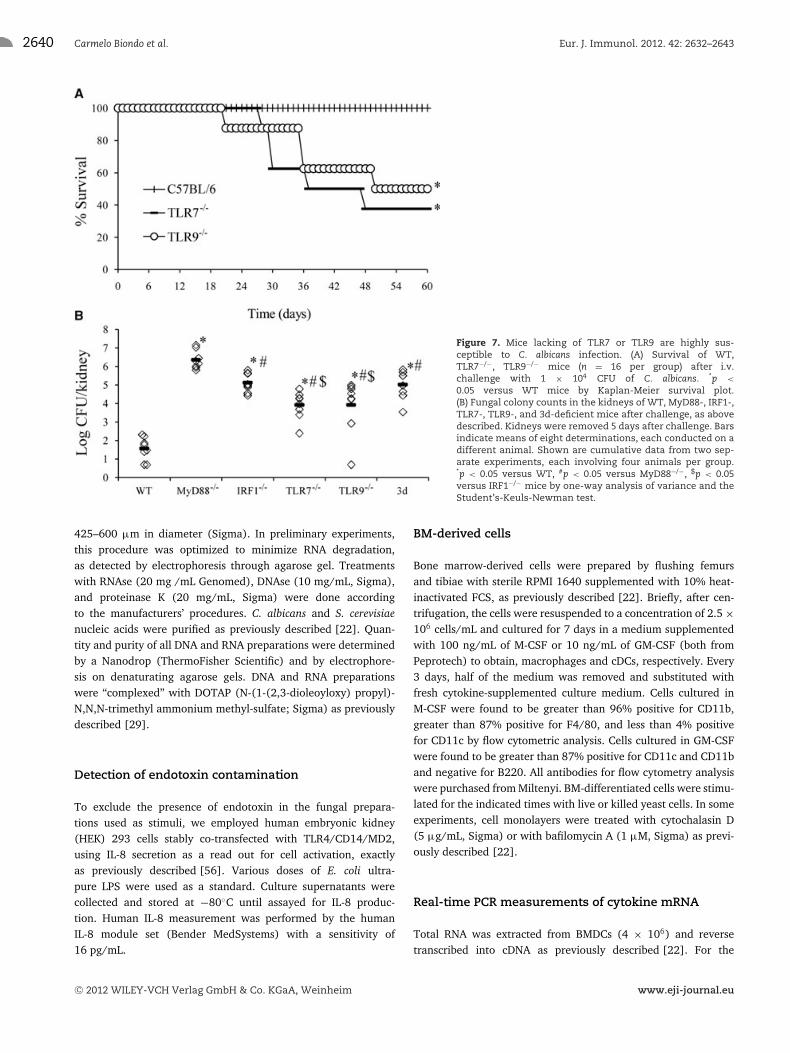
To determine whether signaling through the TLR7 pathway hasa role in host defense against C. albicans, we used an i.v. murinemodel that mimics disseminated C. albicans infection in humans.WT C57BL/6 mice or mice lacking TLR7 or TLR9 were infected i.v.with a low dose (1 × 104 CFU) of C. albicans, a challenge that wasfound to be sublethal for WT mice in preliminary experiments. Sur-vival and morbidity were monitored daily. As shown in Figure 7A,most of the mice lacking either TLR7 or TLR9 succumbed to infec-tion while all WT mice survived. To ascertain whether increasedlethality was associated with a decreased ability of these mice to
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2012. 42: 2632–2643 Immunity to infection 2637
Figure 4. IL-12p70 production in response to S. cerevisiae requires a TRL7- and TLR9-initiated pathway involving MyD88 and IRF1. (A–C) BMDCsfrom WT mice were infected with increasing MOIs (1, 5, and 10) of S. cerevisiae. The concentrations of (A) IL-12p70, (B) IL-23, and (C) TNF-α inculture supernatants were determined by ELISA. BMDCs lacking TLR7, TLR9, dectin-1, MyD88, or IRF1 were stimulated with S. cerevisiae at anMOI of 10. CpG B (1 μg/mL) and CL264 (5 μg/mL) were used as positive controls. The concentrations of (D) IL-12p70, (E) IL-23, and (F) TNF-α inculture supernatants were determined by ELISA. Data are expressed as mean + SD of samples pooled from three independent observations, eachconducted on a different animal, run in duplicate. *p < 0.05 versus WT BMDCs by one-way analysis of variance and the Student’s-Keuls-Newmantest.
control in vivo infection, we measured fungal burden in the kid-ney, the main target of hematogenous C. albicans dissemination, at5 days after infection with the same C. albicans dose (1 × 104 CFU)used in the lethality experiments. In these experiments, we alsotested MyD88−/−, IRF1−/−, and 3d mice in addition of TLR7−/−
and TLR9−/− animals. While low CFU numbers were found in kid-neys of WT mice, fungal burden was significantly increased in micelacking either TLR7 or TLR9 (Fig. 7B). Notably, fungal burden waseven higher in 3d or IRF1−/− mice compared with TLR7−/− orTLR9−/− mice. Mice lacking MyD88 showed the most severe phe-notype of all, with colony counts that were approximately 6 ordersof magnitude higher than those of WT controls. Collectively, thesedata indicated that the TLR7/TLR9/MyD88/IRF1 pathway has anonredundant role in defenses against C. albicans. Moreover, 3dmice (that are unable to mobilize TLR7/9 and other intracellu-lar TLRs to phagosomes) showed a phenotype that was similar tothat of IRF1−/− mice and intermediary between MyD88−/− (highlysusceptible) and TLR9−/− or TLR7−/− (moderately susceptible).
Our results, showing an increased susceptibility of TLR9−/−
mice to C. albicans infection, were apparently in contrast withthose of previous studies showing similar [28,38] or evendecreased [14] susceptibility of TLR9−/− mice in comparison withWT animals. We hypothesized that these discrepancies could berelated to the fact that the cited studies used a higher (1–2 log)challenge doses than the one we used. Therefore, to test thishypothesis, we challenged TLR7- and TLR9- defective mice witha 20-fold higher C. albicans dose than that previously used in theexperiments summarized in Fig. 7. Under these conditions, no dif-
ferences were found in susceptibility to infection between TLR7-or TLR9-deficient mice and WT controls, as measured by kidneycolony counts (Supporting Information Fig. 5). This data indicatethat the effects of TLR7 or TLR9 deficiency on the outcome of theinfection are critically dependent on the challenge dose.
Discussion
The identification of receptors and signal transduction pathwaysinvolved in immune responses to fungi is essential to understandthe mechanisms underlying the development of mycoses and todevise alternative strategies to control these difficult to treat infec-tions. Recent studies have focused on responses induced by cell-wall components through C-type lectin receptors, such as dectin-1and dectin-2 [8,13,39]. Less is known of TLRs involved in fungalsensing and of their functional importance during in vivo infec-tion. We show here the existence of a TLR7/TLR9/MyD88/IRF1-dependent fungal recognition pathway that led to the produc-tion of IL-12p70. This pathway required a receptor (TLR7), achaperone protein (UNC93B1), and a transcription factor (IRF1)that have not been previously studied in the context of immuneresponses to fungi. We found that TLR7, UNC93B1, and IRF1had nonredundant roles in host resistance against C. albicans,as shown by increased susceptibility to infection of geneticallydefective animals. Increased susceptibility was at least partially aconsequence of impaired innate, rather than adaptative, defenses,since it was already evident early during infection. Moreover, inthe systemic candidiasis model we used, host defenses are largely
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

2638 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643
Figure 5. IL-12p70 production in response to S. cerevisiae requiresTLR7/9 traslocation to the endosomal compartments. BMDCs lacking3d or from TLR7/9 double ko mice were stimulated with S. cerevisiae atan MOI of 10. CpG B (1 μg/mL) was used as a positive control. The levelsof (A) IL-12p70, (B) IL-23, and (C) TNF-α in culture supernatants weredetermined by ELISA. Data are expressed as mean + SD of samplespooled from three independent observations, each conducted on a dif-ferent animal, run in duplicate. *p < 0.05 versus WT BMDCs by one-wayanalysis of variance and the Student’s-Keuls-Newman test.
independent from the adaptative immune system [40–42]. TheIRF1 transcription factor was previously shown to be downstreamof MyD88 and to upregulate, after TLR engagement, a distinctivegroup of genes, including IFN-β, IL-12p35, and inducible nitricoxide synthase [43,44]. Accordingly, we found that IL-12p70, butnot TNF-α or IL-23, production was markedly impaired in IRF1-deficient cells after stimulation with whole yeast. Therefore, thehypersusceptibility of IRF1-deficient mice to C. albicans infectionmay be linked to defective production of IL-12p70 and IFN-β, sinceboth of these factors have been previously linked to host defensesin systemic candidiasis models [22,45]. Moreover, since IRF1 has
an essential role in polarizing the T-cell response toward a Th1type [46], it will be important, in future studies, to examine theeffects of the TLR7/9-IRF1 axis in T-cell differentiation duringcandidosis. Collectively, our data indicate that IRF1 is an essentialtranscription factor not only in anti-bacterial [29,47], but also inanti-fungal host defenses.
Two considerations indicate that RNA is the ligand recognizedby TLR7 in BMDCs. In the first place, TLR7 is strictly RNA spe-cific and single stranded RNA is its only natural agonist [29,48].In the second place, the ability of whole yeast to induce TLR7-dependent IL-12p70 secretion could be recapitulated here byyeast RNA, which was, in this activity, more potent thanfungal DNA. Our data confirm and extend those of a previ-ous report showing that yeast RNA was capable of stimulatingDCs for increased IL-12 production [49]. Although the involve-ment of TLR7 in recognition of single-stranded RNA viruseshas been traditionally recognized [48], its role in host defensesagainst bacterial [29] and protozoan [50] organisms has beenonly recently demonstrated. We now show that TLR7 is a criticalinnate immune receptor involved in recognition and host resis-tance to a fungal infection. TLR9, in addition to TLR7, may partic-ipate in triggering anti-fungal responses, as suggested by partiallydecreased IL-12p70 production in TLR9−/− BMDCs after stimu-lation with whole yeasts and total abrogation of this responsein TLR7/9 double KO mice. Moreover, purified DNA was ableto activate a TLR9- and IRF1-dependent pathway leading toIL-12p70 induction. In summary, our data suggest that TLR7 andTLR9 collaborate in a fungal recognition mechanism that tar-gets nucleic acids (RNA and DNA, respectively) and activatesa common, MyD88- and IRF1-dependent, pathway. Activationof this pathway was absolutely dependent on phagocytosis andphagosomal acidification, both of which are known requirementsfor TLR9- and TLR7-mediated recognition. An additional fea-ture of the TLR7/9-dependent responses described here is theircell-type specificity. Indeed, BMDC, but not BMDM, mountedrobust cytokine responses to yeast nucleic acids. The reasons forthese differences are presently unclear, but they may relate todifferential TLR or IRF1 expression or to differential STAT1 phos-phorylation in response to nucleic acid stimulation [51].
Our data are only apparently in contrast with previous reportsindicating that TLR9-defective mice display similar [28,38] oreven increased [14] resistance to C. albicans. Differences betweenour data and those of others were unequivocally linked, in thepresent study, to the different doses used for challenge. In fact,increased susceptibility to C. albicans infection in the absence ofTLR7 or TLR9 was observed only using a low challenge dose.When we challenged mice with the high doses used in the studiescited above, no effect of TLR7 or TLR9 deficiency was observed.Our data are in agreement with the notion that lack of specifichost factors has different and even opposite effects on the out-come of experimental infection depending on the challenge dose,the associated severity of infection, and risk of death [19,52,53].Thus, it appears that the contribution of TLR7 or TLR9 tohost defenses against C. albicans can be evidenced only underexperimental conditions associated with mild, sublethal infection.
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2012. 42: 2632–2643 Immunity to infection 2639
Figure 6. Yeast induced IL-12p70, but not IL-23or TNF-α, production is dependent on phagocy-tosis and phagosomal acidification. Cytokine lev-els in culture supernatants of BMDCs treated withcytochalasin D (5 μg/mL) or bafilomycin A (1 μM)and stimulated with S. cerevisiae at an MOI of 10 weredetermined by ELISA. Data are expressed as mean +SD of samples pooled from three independent obser-vations, each conducted on a different animal, runin duplicate. *p < 0.05 relative to untreated controlby one-way analysis of variance and the Student’s-Keuls-Newman test.
The use of low rather than high challenge doses seems logical,since under most natural circumstances, the immune system isexposed to low numbers of microbial cells in the initial stages ofinfection. Moreover, overwhelming infection is often associatedwith the deleterious release of pathophysiological mediators bythe host and/or of immunosuppressive products by the pathogen,both of which may obscure the contribution of individual immunefactors [19,52–54].
Collectively, our data indicate the presence of at least twodifferent cellular mechanisms underlying fungal recognition thatlead to the production of two different sets of defense factors.The first mechanism, underlying the production of IL-23 andTNF-α, relies predominantly on the detection of cell-wall struc-tures by receptors located on the host cell surface, such as dectin-1.This mechanism does not necessarily require phagocytosis and islargely independent from TLR or TRL adaptors. The second mech-anism depends on progression of the organisms along the classi-cal phagolysosomal route and, likely, on the release of nucleicacids from the fungal cell envelope. DNA and RNA are thendetected by, respectively, TLR7 and TLR9 and trigger MyD88- andIRF1-dependent responses. Interestingly, our data indicating thatactin polymerization and phagocytosis are not required for dectin-1-dependent cytokine induction (e.g. for S. cerevisiae-inducedTNF-α secretion) are in agreement with a recent report show-ing that dectin-1 can be activated by β-glucan immobilized on anonphagocytosable surface (such as a culture plate), occurs priorto initiation of phagocytic cup formation and is not dependent onactin dynamics [55].
Altogether our data, pointing to the importance in anti-fungaldefenses of the latter pathway, may be useful to better understandthe strategies used by C. albicans to evade the innate immunesystem and to devise alternative therapeutic strategies.
Materials and methods
Mice
Knock-out mice were originally obtained from T. Taniguchi(IRF1−/−, IRF3−/−, and IRF7−/−), and S. Akira (TLR2−/−, TL3−/−,
TLR4−/−, TLR7−/−, TLR9−/−, MyD88−/−, Mal−/−, TRAM−/− andTRIF−/−) as previously described [29]. Dectin 1−/−, 3d andTLR7/9 double Ko (TLR7−/−/TLR9−/−) mice were providedby, respectively, G. Brown [11], B. Beutler [35] and S. Bauer.C57BL/6 WT mice, used as controls, were purchased from CharlesRiver Laboratories (Calco, Italy). The mice were housed and bredunder pathogen-free conditions in the animal facilities of the ElieMetchnikoff Department, University of Messina.
Ethics statement
All studies were performed in agreement with the EuropeanUnion guidelines of animal care and were approved by the EthicsCommittee of the Metchnikoff Department of the University ofMessina (CESA) and by the relevant national authority (IstitutoSuperiore di Sanita).
Fungal strains and reagents
C. albicans (ATCC 90028) was purchased from the American TypeCulture Collection. S. cerevisiae strain A11 was isolated in theclinical mycology laboratory of the Elie Metchnikoff Department,University of Messina [22]. For in vivo and in vitro experiments,these two strains were grown in a chemically defined mediumas previously described [22]. CFU numbers used in each experi-ment were determined after plating on Sabouraud dextrose agar(Difco Laboratories). Heat killed C. albicans strains were preparedas previously described [22], followed by washing with PBS andresuspension to the original volume. Depleted zymosan (i.e. hotalkali-treated zymosan, which is devoid of TLR-dependent stim-ulating properties) and control stimuli (poly I:C, Escherichia coliultrapure LPS, CpG B, CL264) were purchased from InvivoGen.Curdlan was purchased from Wako Pure Chemicals and detoxi-fied using cold NaOH treatment [22].
Yeast lysates and purification of C. albicans nucleicacids
Fungal cell extracts were obtained by vortexing of C. albicans(grown in the mid-log phase) in the presence of glass beads
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
2640 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643
Figure 7. Mice lacking of TLR7 or TLR9 are highly sus-ceptible to C. albicans infection. (A) Survival of WT,TLR7−/−, TLR9−/− mice (n = 16 per group) after i.v.challenge with 1 × 104 CFU of C. albicans. *p <
0.05 versus WT mice by Kaplan-Meier survival plot.(B) Fungal colony counts in the kidneys of WT, MyD88-, IRF1-,TLR7-, TLR9-, and 3d-deficient mice after challenge, as abovedescribed. Kidneys were removed 5 days after challenge. Barsindicate means of eight determinations, each conducted on adifferent animal. Shown are cumulative data from two sep-arate experiments, each involving four animals per group.*p < 0.05 versus WT, #p < 0.05 versus MyD88−/−, $p < 0.05versus IRF1−/− mice by one-way analysis of variance and theStudent’s-Keuls-Newman test.
425–600 μm in diameter (Sigma). In preliminary experiments,this procedure was optimized to minimize RNA degradation,as detected by electrophoresis through agarose gel. Treatmentswith RNAse (20 mg /mL Genomed), DNAse (10 mg/mL, Sigma),and proteinase K (20 mg/mL, Sigma) were done accordingto the manufacturers’ procedures. C. albicans and S. cerevisiaenucleic acids were purified as previously described [22]. Quan-tity and purity of all DNA and RNA preparations were determinedby a Nanodrop (ThermoFisher Scientific) and by electrophore-sis on denaturating agarose gels. DNA and RNA preparationswere ‘‘complexed’’ with DOTAP (N-(1-(2,3-dioleoyloxy) propyl)-N,N,N-trimethyl ammonium methyl-sulfate; Sigma) as previouslydescribed [29].
Detection of endotoxin contamination
To exclude the presence of endotoxin in the fungal prepara-tions used as stimuli, we employed human embryonic kidney(HEK) 293 cells stably co-transfected with TLR4/CD14/MD2,using IL-8 secretion as a read out for cell activation, exactlyas previously described [56]. Various doses of E. coli ultra-pure LPS were used as a standard. Culture supernatants werecollected and stored at −80◦C until assayed for IL-8 produc-tion. Human IL-8 measurement was performed by the humanIL-8 module set (Bender MedSystems) with a sensitivity of16 pg/mL.
BM-derived cells
Bone marrow-derived cells were prepared by flushing femursand tibiae with sterile RPMI 1640 supplemented with 10% heat-inactivated FCS, as previously described [22]. Briefly, after cen-trifugation, the cells were resuspended to a concentration of 2.5 ×106 cells/mL and cultured for 7 days in a medium supplementedwith 100 ng/mL of M-CSF or 10 ng/mL of GM-CSF (both fromPeprotech) to obtain, macrophages and cDCs, respectively. Every3 days, half of the medium was removed and substituted withfresh cytokine-supplemented culture medium. Cells cultured inM-CSF were found to be greater than 96% positive for CD11b,greater than 87% positive for F4/80, and less than 4% positivefor CD11c by flow cytometric analysis. Cells cultured in GM-CSFwere found to be greater than 87% positive for CD11c and CD11band negative for B220. All antibodies for flow cytometry analysiswere purchased from Miltenyi. BM-differentiated cells were stimu-lated for the indicated times with live or killed yeast cells. In someexperiments, cell monolayers were treated with cytochalasin D(5 μg/mL, Sigma) or with bafilomycin A (1 μM, Sigma) as previ-ously described [22].
Real-time PCR measurements of cytokine mRNA
Total RNA was extracted from BMDCs (4 × 106) and reversetranscribed into cDNA as previously described [22]. For the
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2012. 42: 2632–2643 Immunity to infection 2641
quantification of IL-12p35, IL-12p40, IL-23p19, and TNF-α mRNA,real-time quantitative RT-PCR assays were conducted, in dupli-cate, with an Applied Biosystems 7500 (Applied Biosystems) asdescribed [22]. Primers and TaqMan MGB probes for the abovecytokines were purchased from Applied Biosystems. PCR condi-tions were as follows: 95◦C, 10 min; (95◦C, 15 s; 60◦C, 1 min) ×40 cycles. Gene expression was measured by the comparative CTmethod (��CT) as previously described [22].
Cytokine measurements
IL-12p70, IL-23, and TNF-α concentrations were measured in thesupernatants of bone marrow-derived cultures using the mouseIL-12p70 Quantikine Immunoassay, IL-23 Quantikine Immunoas-say, and Duo-set TNF-α ELISA (all purchased from R&D Systems).The lower detection limit of these was 16 pg/mL for all assays.Samples below the detection levels were assigned a theoreticalvalue of one-half the detection limit.
Murine infection models
WT and knockout mice were infected i.v., via the lateral tail vein,with 1 × 104 CFU C. albicans ATCC strain 90028 as previouslydescribed [22]. Mice were observed twice daily for signs of diseaseand lethality. The kidney fungal burden was determined exactlyas previously described [22].
Data expression and statistical significance
Cytokine levels and log CFU were expressed as mean ± standarddeviation of the mean (SD) of several determinations, each con-ducted on a different animal in an independent experiment. Dif-ferences in cytokine levels and organ CFUs were assessed by one-way analysis of variance and the Student’s-Keuls-Newman test.Survival data were analyzed with Kaplan–Meier survival plots fol-lowed by the log rank test (JMP Software; SAS Institute, Cary, NC)on an Apple Macintosh computer. When p values of < 0.05 wereobtained, differences were considered statistically significant.
Acknowledgements: We thank S. Akira, G. Brown, B. Beut-ler, S. Bauer, and T. Taniguchi for providing KO mice. Thiswork was partially supported by the Programma OperativoNazionale PON01 00117/8 from the Ministero dell’Istruzione,dell’Universita e della Ricerca of Italy and by Progetti di Ricercad’Ateneo A.013.BIO200809 and A.013.MAN200809 from the Uni-versity of Messina granted to CBi and GM, respectively.
Conflict of interest: The authors declare no financial or commer-cial conflict of interest
References
1 Pfaller, M. A. and Diekema, D. J., Epidemiology of invasive candidiasis: a
persistent public health problem. Clin. Microbiol. Rev. 2007. 20: 133–163.
2 Calderone, R. A., Candida and candidiasis. ASM Press, Washington D.C.,
2002. 451 pp.
3 Medzhitov, R., Recognition of microorganisms and activation of the
immune response. Nature 2007. 449: 819–826.
4 Willment, J. A. and Brown, G. D., C-type lectin receptors in antifungal
immunity. Trends Microbiol. 2008. 16: 27–32.
5 Levitz, S. M. and Specht, C. A., Innate recognition of fungal cell walls.
Plos Pathogens 2010. 6: 1000758.
6 Bowman, S. M. and Free, S. J., The structure and synthesis of the fungal
cell wall. Bioessays. 2006. 28: 799–808.
7 Brown, G. D. and Gordon, S., Immune recognition. A new receptor for
beta-glucans. Nature 2001. 413: 36–37.
8 Means, T. K., Mylonakis, E., Tampakakis, E., Colvin, R. A., Seung, E.,
Puckett, L., Tai, M. F. et al., Evolutionarily conserved recognition and
innate immunity to fungal pathogens by the scavenger receptors SCARF1
and CD36. J. Exp. Med. 2009. 206: 637–653.
9 Gelderman, K. A., Lam, S., Sier, C. F. and Gorter, A. Cross-linking
tumor cells with effector cells via CD55 with a bispecific mAb induces
beta-glucan-dependent CR3-dependent cellular cytotoxicity. Eur. J.
Immunol. 2006. 36: 977–984.
10 Gross, O., Gewies, A., Finger, K., Schafer, M., Sparwasser, T., Peschel, C.,
Forster, I. et al., Card9 controls a non-TLR signalling pathway for innate
antifungal immunity. Nature 2006. 442: 651–656.
11 Taylor, P. R., Tsoni, S. V., Willment, J. A., Dennehy, K. M., Rosas, M.,
Findon, H., Haynes, K. et al., Dectin-1 is required for beta-glucan recog-
nition and control of fungal infection. Nat. Immunol. 2007. 8: 31–38.
12 Glocker, E. O., Hennigs, A., Nabavi, M., Schaffer, A. A., Woellner, C.,
Salzer, U., Pfeifer, D. et al., A homozygous CARD9 mutation in a family
with susceptibility to fungal infections. N. Engl. J. Med. 2009. 361: 1727–
1735.
13 Saijo, S., Ikeda, S., Yamabe, K., Kakuta, S., Ishigame, H., Akitsu, A.,
Fujikado, N. et al., Dectin-2 recognition of alpha-mannans and induction
of Th17 cell differentiation is essential for host defense against Candida
albicans. Immunity 2010. 32: 681–691.
14 Bellocchio, S., Montagnoli, C., Bozza, S., Graziano, R., Rossi, G., Mambula,
S., Vecchi, A. et al., The contribution of the Toll-like/IL-1 receptor super-
family to innate and adaptive immunity to fungal pathogens in vivo.
J. Immunol. 2004. 172: 3059–3069.
15 Romani, L., Immunity to fungal infections. Nat. Rev. Immunol. 2004. 11:
275–288.
16 Takeda, K. and Akira, S., Microbial recognition by Toll-like receptors.
J. Dermatol. Sci. 2004. 34: 73–82.
17 Marr, K. A., Balajee, S. A., Hawn, T. R., Ozinsky, A., Pham, U., Akira, S.,
Aderem, A., et al., Differential role of MyD88 in macrophage-mediated
responses to opportunistic fungal pathogens. Infect. Immun. 2003. 71:
5280–5286.
18 Yauch, L. E., Mansour, M. K., Shoham, S., Rottman, J. B. and Levitz, S. M.,
Involvement of CD14, Toll-like receptors 2 and 4, and MyD88 in the host
response to the fungal pathogen Cryptococcus neoformans in vivo. Infect.
Immun. 2004. 72: 5373–5382.
19 Biondo, C., Midiri, A., Messina, L., Tomasello, F., Garufi, G., Catania, M.
R., Bombaci, M. et al., MyD88 and TLR2, but not TLR4, are required for
host defense against Cryptococcus neoformans. Eur. J. Immunol. 2005. 35:
870–878.
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

2642 Carmelo Biondo et al. Eur. J. Immunol. 2012. 42: 2632–2643
20 Netea, M. G., Gow, N. A., Munro, C. A., Bates, S., Collins, C., Ferwerda,
G., Hobson, R. P. et al., Immune sensing of Candida albicans requires
cooperative recognition of mannans and glucans by lectin and Toll-like
receptors. J. Clin. Invest. 2006. 116: 1642–1650.
21 Jouault, T., Ibata-Ombetta, S., Takeuchi, O., Trinel, P. A., Sacchetti, P.,
Lefebvre, P., Akira, S. et al., Candida albicans phospholipomannan is
sensed through toll-like receptors. J. Infect. Dis. 2003. 188: 165–172.
22 Biondo, C., Signorino, G., Costa, A., Midiri, A., Gerace, E., Galbo, R.,
Bellantoni, A. et al., Recognition of yeast nucleic acids triggers a host-
protective type I interferon response. Eur. J. Immunol. 2011. 41: 1969–
1979.
23 Herring, A. C., Falkowski, N. R., Chen, G. H., McDonald, R. A., Toews, G.
B. and Huffnagle, G. B., Transient neutralization of tumor necrosis factor
alpha can produce a chronic fungal infection in an immunocompetent
host: potential role of immature dendritic cells. Infect. Immun. 2005. 73:
39–49.
24 Ashman, R. B., Vijayan, D. and Wells, C. A., IL-12 and related cytokines:
function and regulatory implications in Candida albicans infection. Clin.
Dev. Immunol. 2011. 686597.
25 Wei, X. Q., Rogers, H., Lewis, M. A. and Williams, D. W., The role of
the IL-12 cytokine family in directing T-cell responses in oral candidosis.
Clin. Dev. Immunol. 2011. 697340.
26 Nakamura, K., Miyazato, A., Xiao, G., Hatta, M., Inden, K., Aoyagi, T.,
Shiratori, K. et al., Deoxynucleic acids from Cryptococcus neoformans acti-
vate myeloid dendritic cells via a TLR9-dependent pathway. J. Immunol.
2008. 180: 4067–4074.
27 Ramirez-Ortiz, Z. G., Specht, C. A., Wang, J. P., Lee, C. K., Bartholomeu,
D. C., Gazzinelli, R. T. and Levitz, S. M., Toll-like receptor 9-dependent
immune activation by unmethylated CpG motifs in Aspergillus fumigatus
DNA. Infect. Immun. 2008. 76: 2123–2129.
28 Miyazato, A., Nakamura, K., Yamamoto, N., Mora-Montes, H. M.,
Tanaka, M., Abe, Y., Tanno, D. et al., Toll-like receptor 9-dependent
activation of myeloid dendritic cells by deoxynucleic acids from Candida
albicans. Infect. Immun. 2009. 77: 3056–3064.
29 Mancuso, G., Gambuzza, M., Midiri, A., Biondo, C., Papasergi, S., Akira,
S., Teti, G. et al., Bacterial recognition by TLR7 in the lysosomes of con-
ventional dendritic cells. Nat. Immunol. 2009. 10: 587–594.
30 Lott, T. J., Kuykendall, R. J. and Reiss, E., Nucleotide sequence analysis
of the 5.8S rDNA and adjacent ITS2 region of Candida albicans and related
species. Yeast. 1993. 9: 1199–1206.
31 Murphy, A. and Kavanagh, K., Emergence of Saccharomyces cerevisiae as a
human pathogen: implications for biotechnology. Enzyme Microbiol. Tech-
nol. 1999. 25: 551–557.
32 Diezmann, S. and Dietrich, F. S., Saccharomyces cerevisiae: population
divergence and resistance to oxidative stress in clinical, domesticated
and wild isolates. PLoS One 2009. 4: e5317.
33 Dennehy, K. M., Willment, J. A., Williams, D. L. and Brown, G. D., Recip-
rocal regulation of IL-23 and IL-12 following co-activation of Dectin-1 and
TLR signaling pathways. Eur. J. Immunol. 2009. 39: 1379–1386.
34 Dennehy, K. M., Ferwerda, G., Faro-Trindade, I., Pyz, E., Willment, J. A.,
Taylor, P. R., Kerrigan, A. et al., Syk kinase is required for collaborative
cytokine production induced through Dectin-1 and Toll-like receptors.
Eur. J. Immunol. 2008. 38: 500–506.
35 Tabeta, K., Hoebe, K., Janssen, E. M., Du, X., Georgel, P., Crozat, K., Mudd,
S. et al., The Unc93b1 mutation 3d disrupts exogenous antigen presen-
tation and signaling via Toll-like receptors 3, 7 and 9. Nat. Immunol. 2006.
7: 156–164.
36 Brinkmann, M. M., Spooner, E., Hoebe, K., Beutler, B., Ploegh, H. L. and
Kim, Y. M., The interaction between the ER membrane protein UNC93B
and TLR3, 7, and 9 is crucial for TLR signaling. J. Cell Biol. 2008. 177: 265–
275.
37 Kim, Y. M., Brinkmann, M. M., Paquet, M. E. and Ploegh, H. L., UNC93B1
delivers nucleotide-sensing toll-like receptors to endolysosomes. Nature
2008. 452: 234–238.
38 Van de Veerdonk, F. L., Netea, M. G., Jansen, T. J., Jacobs, L., Verschueren,
I., van der Meer, J. W. and Kullberg, B. J., Redundant role of TLR9 for anti-
Candida host defense. Immunobiology 2008. 213: 613–620.
39 Dennehy, K. M. and Brown, G. D., The role of the beta-glucan recep-
tor Dectin-1 in control of fungal infection. J. Leukoc. Biol. 2007. 82: 253–
258.
40 Cutler, J. E., Acute systemic candidiasis in normal and congenitally
thymicdeficient (nude) mice. J. Reticuloendothel. Soc. 1976. 19: 121–124.
41 Mahanty, S., Greenfield, R. A., Joyce, W. A. and Kincade, P. W., Inocu-
lation candidiasis in a murine model of severe combined immunodefi-
ciency syndrome. Infect. Immun. 1988. 56: 3162–3166.
42 Greenblatt, M. B., Aliprantis, A., Hu, B. and Glimcher, L. H., Calcineurin
regulates innate antifungal immunity in neutrophils. J. Exp. Med. 2010.
207: 923–931.
43 Tamura, T., Yanai, H., Savitsky, D. and Taniguchi, T., The IRF family
transcription factors in immunity and oncogenesis. Annu. Rev. Immunol.
2008. 26: 535–584.
44 Schmitz, F., Heit, A., Guggemoos, S., Krug, A., Mages, J., Schiemann,
M., Adler, H. et al., Interferon-regulatory-factor 1 controls Toll-like
receptor 9-mediated IFN-beta production in myeloid dendritic cells. Eur.
J. Immunol. 2007. 37: 315–327.
45 Zelante, T., De Luca, A., Bonifazi, P., Montagnoli, C., Bozza, S., Moretti,
S. and Belladonna, M. L. et al., IL-23 and the Th17 pathway promote
inflammation and impair antifungal immune resistance. Eur. J. Immunol.
2007. 37: 2695–2706.
46 Kano, S., Sato, K., Morishita, Y., Vollstedt, S., Kim, S., Bishop, K., Honda,
K. et al., The contribution of transcription factor IRF1 to the interferon-
gamma-interleukin 12 signaling axis and TH1 versus TH-17 differentia-
tion of CD4+ T cells. Nat. Immunol. 2008. 9: 34–41.
47 Lohoff, M. and Mak, T. W., Roles of interferon-regulatory factors in T-
helper-cell differentiation. Nat. Rev. Immunol. 2005. 5: 125–135.
48 Diebold, S. S., Kaisho, T., Hemmi, H., Akira, S. and Reis Sousa, C., Innate
antiviral responses by means of TLR7-mediated recognition of single-
stranded RNA. Science 2004. 303: 1529–1531.
49 Bacci, A., Montagnoli, C., Perruccio, K., Bozza, S., Gaziano, R., Pitzurra,
L., Velardi, A. et al., Dendritic cells pulsed with fungal RNA induce pro-
tective immunity to Candida albicans in hematopoietic transplantation.
J. Immunol. 2002. 168: 2904–2913.
50 Caetano, B. C., Carmo, B. B., Melo, M. B., Cerny, A., dos Santos, S. L.,
Bartholomeu, D. C., Golenbock, D. T. et al., Requirement of UNC93B1
reveals a critical role for TLR7 in host resistance to primary infection
with Trypanosoma cruzi. J. Immunol. 2011. 187: 1903–1911.
51 Schroder, K., Spille, M., Pilz, A., Lattin, J., Bode, K. A., Irvine, K. M., Bur-
rows, A. D. et al., Differential effects of CpG DNA on IFN-beta induc-
tion and STAT1 activation in murine macrophages versus dendritic
cells: alternatively activated STAT1 negatively regulates TLR signaling
in macrophages. J. Immunol. 2007. 179: 3495–3503.
52 Mancuso, G., Midiri, A., Beninati, C., Biondo, C., Galbo, R., Akira, S., Hen-
neke, P. et al., Dual role of TLR2 and myeloid differentiation factor 88 in a
mouse model of invasive group B streptococcal disease. J. Immunol. 2004.
172: 6324–6329.
53 Biondo, C., Midiri, A., Gambuzza, M., Gerace, E., Falduto, M., Galbo, R.,
Bellantoni, A. et al., IFNalpha/beta signaling is required for polarization
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2012. 42: 2632–2643 Immunity to infection 2643
of cytokine responses toward a protective type 1 pattern during experi-
mental cryptococcosis. J. Immunol. 2008. 181: 566–573.
54 Eichacker, P. Q., Parent, C., Kalil, A., Esposito, C., Cui, X., Banks, S. M.,
Gerstenberger, E. P. et al., Risk and the efficacy of antiinflammatory
agents: retrospective and confirmatory studies of sepsis. Am. J. Respir.
Crit. Care Med. 2002. 166: 1197–1205.
55 Goodridge, H. S., Reyes, C. N., Becker, C. A., Katsumoto, T. R., Ma, J., Wolf,
A. J., Bose, N. et al., Activation of the innate immune receptor Dectin-1
upon formation of a ‘phagocytic synapse’. Nature 2011. 472: 471–475.
56 Mancuso, G., Midiri, A., Biondo, C., Beninati, C., Gambuzza, M., Macrı,
D., Bellantoni, A. et al., Bacteroides fragilis-derived lipopolysaccharide pro-
duces cell activation and lethal toxicity via TLR4. Infect. Immun. 2005. 73:
5620–5627.
Abbreviations: BMDC: bone marrow-derived in vitro-differentiated den-
dritic cell · BMDM: bone marrow-derived in vitro-differentiated den-
dritic macrophage · IRF: interferon regulatory factor
Full correspondence: Dr. Giuseppe Mancuso, Torre Biologica IIp,Policlinico, Via Consolare Valeria, 1, 98125 Messina, ItalyFax: +39-090-221-3312e-mail: [email protected]
Received: 14/3/2012Revised: 6/6/2012Accepted: 27/6/2012Accepted article online: 10/7/2012
C© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Related Documents











