Receptor for Hyaluronan-Mediated Motility Correlates with Centrosome Abnormalities in Multiple Myeloma and Maintains Mitotic Integrity Christopher A. Maxwell, Jonathan J. Keats, Andrew R. Belch, Linda M. Pilarski, and Tony Reiman Department of Oncology, University of Alberta and Cross Cancer Institute, Edmonton, Alberta, Canada Abstract Elevated expression of receptor for hyaluronan-mediated motility (RHAMM) within ex vivo diagnostic multiple myeloma plasma cells predicts for aggressive disease and patient survival. Here, we investigate the relationship between RHAMM and centrosomal abnormalities within multiple myeloma patient samples. We report that myeloma patient samples contain pervasive structural and numerical centro- somal abnormalities. Structural, but not numerical, centro- somal abnormalities strongly correlate with elevated RHAMM expression. As others have shown that excess pericentriolar material strongly associates with abnormal mitoses, we modeled centrosomal abnormalities with exogenous RHAMM overexpression. RHAMM overexpression in vitro resulted in centrosomal and mitotic defects. To elucidate a mechanism for RHAMM-mediated spindle defects, we further investigated RHAMM mitotic function. RHAMM mitotic localization mir- rors that of targeting protein for Xklp2 (TPX2), and RHAMM interacts with the spindle assembly factors dynein and TPX2. Like TPX2, RHAMM expression is up-regulated during mitosis. Moreover, inhibition of function experiments reveals that RHAMM and TPX2 functions converge to maintain spindle integrity after spindle assembly. We postulate that augmen- tation of RHAMM expression within human cancers, including myeloma, can directly affect centrosomal structure and spindle integrity and potentially modulate apoptotic and cell cycle progression pathways. (Cancer Res 2005; 65(3): 850-60) Introduction The receptor for hyaluronan-mediated motility (RHAMM), first described by Turley (1), has cell surface and intracellular distribution and binds hyaluronan (2), ERK kinase (3) and microtubules (4). RHAMM participates in cell motility, signaling (5), and oncogenic events (6). RHAMM also localizes to the centrosome, the microtubule organizing center, and functions in the maintenance of spindle integrity (7). The COOH terminus of RHAMM is essential for centrosomal localization and bears structural similarity to human kinesin-like protein 2, including conservation of a basic leucine zipper domain (8); this motif is essential for the cell cycle–specific interaction between human kinesin-like protein 2 and dynein, which is mediated through targeting protein for Xklp2 (TPX2; ref. 8). Thus, it is intriguing to hypothesize that RHAMM may interact with dynein and TPX2 and function in spindle assembly and integrity. In multiple myeloma, accumulation of terminally differentiated malignant plasma cells in the bone marrow, despite low plasma cell labeling indices, is suggestive of a clonotypic precursor cell with proliferative potential. RHAMM transcripts are detected in multiple myeloma malignant cells but are weak or absent from normal B cells, bone marrow, or CD138 + plasma cells of control patients (9, 10). MMPCs are characterized by extensive chromo- somal instability and elevated RHAMM expression, which is significantly correlated to increased disease-related events and reduced survival (10). Moreover, the prevalence of cytogenetic abnormalities is greater in high, compared with low, RHAMM expressors (10). RHAMM expression is linked to progression and metastasis of a variety of epithelial tumors, including endometrial, stomach, and breast carcinomas (11–13). The association of RHAMM with centrosomes, cell division, and mitotic integrity may explain the correlation observed between RHAMM expres- sion and the progression of malignancies with extensive chromosomal instability. Centrosomes are composed of two centrioles and an amorphous cloud of pericentriolar material (14). Like RHAMM, many integral components of the pericentriolar material are multiple coiled coil domain proteins that interact with dynein to target centrosomes in a microtubule-independent manner (15). Numerical and/or struc- tural centrosome abnormalities have been reported in a wide range of malignant epithelial tumors with positive correlation to genetic instability and cancer progression (16, 17); to date, however, centrosomal dysregulation has not been investigated within myeloma cells. Centrosomal abnormalities occur early in tumor- igenesis (18) with structural (i.e., excess pericentriolar material), rather than numerical, abnormalities most highly associated with abnormal mitoses (16). Formation of a bipolar spindle is essential to the proper segregation of replicated chromosomes and the maintenance of genetic stability. Spindle assembly can occur through centrosome- dependent and centrosome-independent pathways (19). In some systems, spindle assembly is induced in a Ran-GTP-dependent manner (20, 21). Active Ran, enriched proximal to chromosomes (22), promotes microtubule nucleation through downstream effector proteins, such as TPX2, NuMA, and Aurora A kinase (aurA; refs. 23, 24). TPX2 initiates spindle assembly by nucleating and bundling microtubules (23, 25) and by directly activating aurA in a microtubule-dependent manner (24). The importance of aurA, TPX2, and NuMA has been illustrated in mammalian systems as well (26). Once spindles have been established, their integrity is dependent on the balance of forces generated by microtubule-dependent motors (e.g., dynein). Disruption of this balance, through overexpression or inhibition of structural participants, would be expected to disrupt spindle structure (27). Because RHAMM is associated with the progression of multiple tumors and, within myeloma, elevated RHAMM is associated with Requests for reprints: Tony Reiman, Department of Medical Oncology, Cross Cancer Institute, 11560 University Avenue, Edmonton, Alberta, Canada T6G 1Z2. Phone: 780-432-8513; Fax: 780-432-8888; E-mail: [email protected]. I2005 American Association for Cancer Research. Cancer Res 2005; 65: (3). February 1, 2005 850 www.aacrjournals.org Research Article Research. on February 22, 2016. © 2005 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Receptor for Hyaluronan-Mediated Motility Correlates with
Centrosome Abnormalities in Multiple Myeloma and
Maintains Mitotic Integrity
Christopher A. Maxwell, Jonathan J. Keats, Andrew R. Belch, Linda M. Pilarski,and Tony Reiman
Department of Oncology, University of Alberta and Cross Cancer Institute, Edmonton, Alberta, Canada
Abstract
Elevated expression of receptor for hyaluronan-mediatedmotility (RHAMM) within ex vivo diagnostic multiple myelomaplasma cells predicts for aggressive disease and patientsurvival. Here, we investigate the relationship betweenRHAMM and centrosomal abnormalities within multiplemyeloma patient samples. We report that myeloma patientsamples contain pervasive structural and numerical centro-somal abnormalities. Structural, but not numerical, centro-somal abnormalities strongly correlate with elevated RHAMMexpression. As others have shown that excess pericentriolarmaterial strongly associates with abnormal mitoses, wemodeled centrosomal abnormalities with exogenous RHAMMoverexpression. RHAMM overexpression in vitro resulted incentrosomal and mitotic defects. To elucidate a mechanismfor RHAMM-mediated spindle defects, we further investigatedRHAMM mitotic function. RHAMM mitotic localization mir-rors that of targeting protein for Xklp2 (TPX2), and RHAMMinteracts with the spindle assembly factors dynein and TPX2.Like TPX2, RHAMM expression is up-regulated during mitosis.Moreover, inhibition of function experiments reveals thatRHAMM and TPX2 functions converge to maintain spindleintegrity after spindle assembly. We postulate that augmen-tation of RHAMM expression within human cancers, includingmyeloma, can directly affect centrosomal structure andspindle integrity and potentially modulate apoptotic and cellcycle progression pathways. (Cancer Res 2005; 65(3): 850-60)
Introduction
The receptor for hyaluronan-mediated motility (RHAMM), firstdescribed by Turley (1), has cell surface and intracellulardistribution and binds hyaluronan (2), ERK kinase (3) andmicrotubules (4). RHAMM participates in cell motility, signaling(5), and oncogenic events (6). RHAMM also localizes to thecentrosome, the microtubule organizing center, and functions inthe maintenance of spindle integrity (7). The COOH terminus ofRHAMM is essential for centrosomal localization and bearsstructural similarity to human kinesin-like protein 2, includingconservation of a basic leucine zipper domain (8); this motif isessential for the cell cycle–specific interaction between humankinesin-like protein 2 and dynein, which is mediated throughtargeting protein for Xklp2 (TPX2; ref. 8). Thus, it is intriguing to
hypothesize that RHAMM may interact with dynein and TPX2 andfunction in spindle assembly and integrity.In multiple myeloma, accumulation of terminally differentiated
malignant plasma cells in the bone marrow, despite low plasmacell labeling indices, is suggestive of a clonotypic precursor cellwith proliferative potential. RHAMM transcripts are detected inmultiple myeloma malignant cells but are weak or absent fromnormal B cells, bone marrow, or CD138+ plasma cells of controlpatients (9, 10). MMPCs are characterized by extensive chromo-somal instability and elevated RHAMM expression, which issignificantly correlated to increased disease-related events andreduced survival (10). Moreover, the prevalence of cytogeneticabnormalities is greater in high, compared with low, RHAMMexpressors (10). RHAMM expression is linked to progression andmetastasis of a variety of epithelial tumors, including endometrial,stomach, and breast carcinomas (11–13). The association ofRHAMM with centrosomes, cell division, and mitotic integritymay explain the correlation observed between RHAMM expres-sion and the progression of malignancies with extensivechromosomal instability.Centrosomes are composed of two centrioles and an amorphous
cloud of pericentriolar material (14). Like RHAMM, many integralcomponents of the pericentriolar material are multiple coiled coildomain proteins that interact with dynein to target centrosomes ina microtubule-independent manner (15). Numerical and/or struc-tural centrosome abnormalities have been reported in a wide rangeof malignant epithelial tumors with positive correlation to geneticinstability and cancer progression (16, 17); to date, however,centrosomal dysregulation has not been investigated withinmyeloma cells. Centrosomal abnormalities occur early in tumor-igenesis (18) with structural (i.e., excess pericentriolar material),rather than numerical, abnormalities most highly associated withabnormal mitoses (16).Formation of a bipolar spindle is essential to the proper
segregation of replicated chromosomes and the maintenance ofgenetic stability. Spindle assembly can occur through centrosome-dependent and centrosome-independent pathways (19). In somesystems, spindle assembly is induced in a Ran-GTP-dependentmanner (20, 21). Active Ran, enriched proximal to chromosomes(22), promotes microtubule nucleation through downstreameffector proteins, such as TPX2, NuMA, and Aurora A kinase (aurA;refs. 23, 24). TPX2 initiates spindle assembly by nucleating andbundling microtubules (23, 25) and by directly activating aurA in amicrotubule-dependentmanner (24). The importance of aurA, TPX2,and NuMA has been illustrated in mammalian systems as well (26).Once spindles have been established, their integrity is dependent onthe balance of forces generated by microtubule-dependent motors(e.g., dynein). Disruption of this balance, through overexpression orinhibition of structural participants, would be expected to disruptspindle structure (27).Because RHAMM is associated with the progression of multiple
tumors and, within myeloma, elevated RHAMM is associated with
Requests for reprints: Tony Reiman, Department of Medical Oncology, CrossCancer Institute, 11560 University Avenue, Edmonton, Alberta, Canada T6G 1Z2.Phone: 780-432-8513; Fax: 780-432-8888; E-mail: [email protected].
I2005 American Association for Cancer Research.
Cancer Res 2005; 65: (3). February 1, 2005 850 www.aacrjournals.org
Research Article
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

cytogenetic abnormalities, aggressive disease, and shortenedsurvival, we investigated an association between RHAMM andcentrosomal abnormalities and the effects of exogenous RHAMMexpression on centrosomal and spindle structure. We show thatRHAMM gene expression levels correlate with myeloma centro-some volumes and that RHAMM localizes to the centrosome inmyeloma plasma cells to a similar extent as other definedcentrosome proteins. We show that RHAMM overexpressionin vitro induces centrosomal structural abnormalities, similar tothose identified in myeloma cells ex vivo , and abnormal spindlearchitecture. RHAMM interacts with the spindle assembly factorsdynein and TPX2, but not NuMA, and dysregulation of RHAMMexpression affects G2-M transition and spindle integrity. Based onthe work presented here, we postulate that aberrant RHAMMexpression in myeloma and other cancers may lead to errors inchromosomal segregation.
Materials and Methods
Patients and Clinical Data. Bone marrow biopsies from 41 multiple
myeloma patients, 8 monoclonal gammopathy of uncertain significance
(MGUS, a ‘‘premalignant’’ condition that may precede development of
myeloma), and 4 lymphoma patients with uninvolved bone marrow were
identified from the pathology records at the Cross Cancer Institute from
1997 to 2000. Bone marrow mononuclear cells were collected from archived
aspirates from multiple myeloma patients described above (n = 15), with
z40% plasma cell infiltration, for quantitative analysis of RHAMM
expression. Purified plasma cells were collected from an additional 11
multiple myeloma bone marrow aspirates following CD138, or CD38,
selection by fluorescence-activated cell sorting (n = 6 for cytospin analysis of
centrosomal RHAMM) or autoMACS Magnetic Cell Sorter [Miltenyi Biotec,
Auburn, CA; n = 5 for quantitative reverse transcription-PCR (q-RT-PCR)
analysis of RHAMM expression] as described below. All patients consented
to the use of their bone marrow samples.Preparation and Analysis of Multiple Myeloma Bone Marrow Core
Biopsies. Bone marrow core biopsies were fixed, decalcified, and paraffin
embedded. Sections (4 Am) were mounted on sialinized glass slides. For
antigen retrieval, slides were placed in EDTA (1.0 mmol/L, pH 8.0),
preheated to 100jC, and placed in a temperature-controlled microwave
(TTMega) for 10 minutes. Biopsies were sequentially stained with g-tubulin
(1:200), anti-mouse Alexa 594 (1:200), mouse IgG for blocking, and CD138-
FITC (1:20) and mounted with glycerol medium containing 4V,6-diamidino-
2-phenylindole (DAPI) for DNA staining.Image Analysis and Three-dimensional Volume Rendering. Following
immunofluorescence, confocal z-slices were imaged from core biopsies. For
all analysis, confocal slices were imaged at defined zoom (40� objective,
8 zoom), speed, and depth (0.2 Am) and were acquired using a Zeiss
confocal LSM 510 or multiphoton microscope. In general, three confocal
stacks, including at least 30 plasma cells, were collected. Centrosomes were
identified by g-tubulin immunofluorescence, and regions of interest,
containing individual centrosomes, were extracted using Zeiss 510 image
analysis software. Images were analyzed with Imaris 3.2.2 software
(Bitplane AG, Zurich, Switzerland). Centrosomes were manually counted
for at least 80 CD138+ plasma cells per patient; plasma cells with z3
centrosomes were considered abnormal. Abnormal centrosomal structure
was qualitatively assessed using previously published variables (17). For
determination of centrosomal fractions, confocal images were obtained
which included the centrosomal region. Regions of interest were defined
around the centrosome and the whole cell body, and the intensity within
the centrosomal regions of interest was divided by the overall cellular
intensity.q-RT-PCR Assays of Myeloma Bone Marrow Aspirates. Bone marrow
mononuclear cell isolation, CD138 selection, and q-RT-PCR were done as
described previously to measure RHAMMmRNA levels in multiple myeloma
patient bone marrow samples (10). The expression level of each sample was
normalized to the sample with the lowest level of RHAMM expression,
which was set to an expression level of 1.
Antibodies and Plasmids. Staining used a-tubulin (clone B-5-1-2) and
g-tubulin (clone GTU-88, Sigma, St. Louis, MO), h-actin (Sigma), pericentrin
(Covance, Richmond, CA), CD138-FITC (Serotec, Raleigh, NC), CD38-PE
(BD Biosciences, Palo Alto, CA), and CD45-FITC (Beckman Coulter,
Fullerton, CA). The polyclonal RHAMM antibody was produced and
characterized as described (8). A second polyclonal RHAMM antiserum was
kindly provided by V. Assmann (St. Thomas’ Hospital, London, United
Kingdom; ref. 4). TPX2 antiserum was kindly provided by O. Gruss (EMBL,
Heidelberg, Germany; ref. 28). The mouse monoclonal NuMA antibodies
were identified in a monoclonal antibody screen for mitotic chromosome
scaffold proteins (29). CENP-F antiserum was kindly provided by G. Chan
(University of Alberta, Edmonton, Alberta, Canada; ref. 30). Secondary
antibodies were from Molecular Probes (Eugene, OR). GFP-RHAMMFL and
pEGFP-C1 (Invitrogen, Carlsbad, CA) plasmids were prepared as described
previously (7).Cell Culture, Transient Transfection, Immunofluorescence, and
RNA Interference. RPMI 8226 (a myeloma cell line), Raji (a Burkitt’s
lymphoma cell line), and HeLa (a cervical adenocarcinoma line) were
grown as recommended (American Type Culture Collection, Manassas,
VA). Cells were passaged 24 hours before transfection. Suspension cells
were transfected by electroporation (270 mV, 960 AF, 47-53 ms), whereas
HeLa cells were transfected with LipofectAMINE 2000 (Invitrogen)
following the manufacturer’s protocols. At defined time points post-
transfection, cells were fixed and permeabilized in cold methanol and
washed with PBS-0.5% Triton X-100 (Sigma) before immunofluorescence.
For double and triple staining experiments, antibodies were added
sequentially. Cells were washed thrice in PBS-0.5% Tween before and
after incubations. Cells were mounted in 90% glycerol/PBS + DAPI, and
images were acquired using a Zeiss confocal LSM 510 or multiphoton
microscope. Images were processed using MetaMorph Software (Universal
Imaging Corp., Downington, PA) and Photoshop 5.02 software (Adobe
Systems, Inc., Ottawa, Ontario, Canada).
We carried out RNA interference as described (31). Small interfering
RNA targeting human TPX2 (NM_012112) and human RHAMM
(NM_012484) were ordered predesigned using the Cenix Bioscience
algorithm (Ambion, Austin, TX). TPX2 targeted sequence was 5V-
GGAGAUACACAAAACAUAGtt-3V and RHAMM targeted sequence was 5V-
GGUGCUUAUGAUGUUAAAAtt-3V; RHAMM sequence targets all
RHAMMFL, RHAMM�exon 4, and RHAMM�exon 13 isoforms. Control RNA,
targeting luciferase GL2, was ordered from Dharmacon, Inc. (Lafayette,
CO). Oligonucleotides were annealed and transfected with LipofectAMINE
2000 as per manufacturer’s suggested protocols. Briefly, 1.5 AL of 20
mmol/L small interfering RNA complex was incubated with 50 AL Opti-
MEM (Invitrogen) and 1 AL LipofectAMINE 2000 was incubated with 50
AL Opti-MEM. These solutions were mixed and incubated at room
temperature for 20 minutes before being diluted to 2 mL and added to
cells. Cells were isolated for immunoblotting or immunofluorescence 24,
30, and 48 hours post-transfection.
Synchronization, Immunoprecipitation, and Quantitation. Raji and
HeLa cells were synchronized by double thymidine (2 mmol/L, 14-16 hours)
and nocodazole (300 ng/mL, 10-12 hours) block. Unsynchronized popula-
tions were released from plates with 1� trypsin. Both mitotic and
unsynchronized populations were then washed thrice with PBS andlysed
at 5 � 106 to 107 cells/mL in 1% CHAPS plus 10 Ag/mL leupeptin, 10 Ag/mL
antipain, and 1 mmol/L phenylmethylsulfonyl fluoride (all from Sigma). All
immunoprecipitation procedures were done at 4jC as described previously
(7). For quantitation of coprecipitated proteins, post-immunoprecipitation
lysates were collected and analyzed by SDS-PAGE. Protein quantitation
used the Odyssey v1.1 IR imaging system (LI-COR) with detection of
RHAMM Affects Centrosomal and Spindle Integrity
www.aacrjournals.org 851 Cancer Res 2005; 65: (3). February 1, 2005
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
polyclonal sera using IRDye 800–conjugated anti-rabbit IgG (Rockland,
Gilbertsville, PA).
Statistical Methods. Data were analyzed using SAS version 8.2 for
Windows (SAS, Inc., Cary, NC). Correlation between centrosomal
variables and continuous variables used Pearson’s correlation coeffi-
cient. Two-group comparisons of centrosome variables used Student’s t
test. Statistical significance was set at P = 0.05 using two-sided
analysis.
Table 1. Ex vivo analysis of centrosomes in MMPC
CD138+ in situ
plasma cells
Patients
examined (n)
Centrosomal volumes (Am3),
mean F SE (range)
Centrosomes per cell,
mean F SE (range)
Abnormal centrosomes (%),
mean F SE (range)
Multiple myeloma 41 0.86 F 0.04* (0.43-1.76) 1.52 F 0.03c (1.2-2.0) 26.4 F 2.11*(4-62)
MGUS 8 0.62 F 0.05 (0.47-0.86) 1.37 F 0.06 (1.2-1.6) 14.1 F 2.19 (5-24.4)
Control 4 0.42 F 0.08 (0.30-0.65) 1.25 F 0.08 (1.1-1.5) 12.3 F 2.15 (6.3-16.3)
NOTE: Bone marrow cores were fixed, decalcified, paraffin embedded, and sectioned. Antigen retrieval preceded double immunofluorescence with
CD138-FITC to highlight plasma cell and g-tubulin to demarcate centrosomes. Centrosomes per cell values and percentage abnormal centrosomes weredetermined by qualitative assessment; experimenters were blinded to core diagnosis at the time of assessment and quantitation. Centrosomal volumes
were determined by three-dimensional rendering of confocal z-stacks.
*P < 0.005 compared with control or MGUS values.cP < 0.05 compared with control or MGUS values.
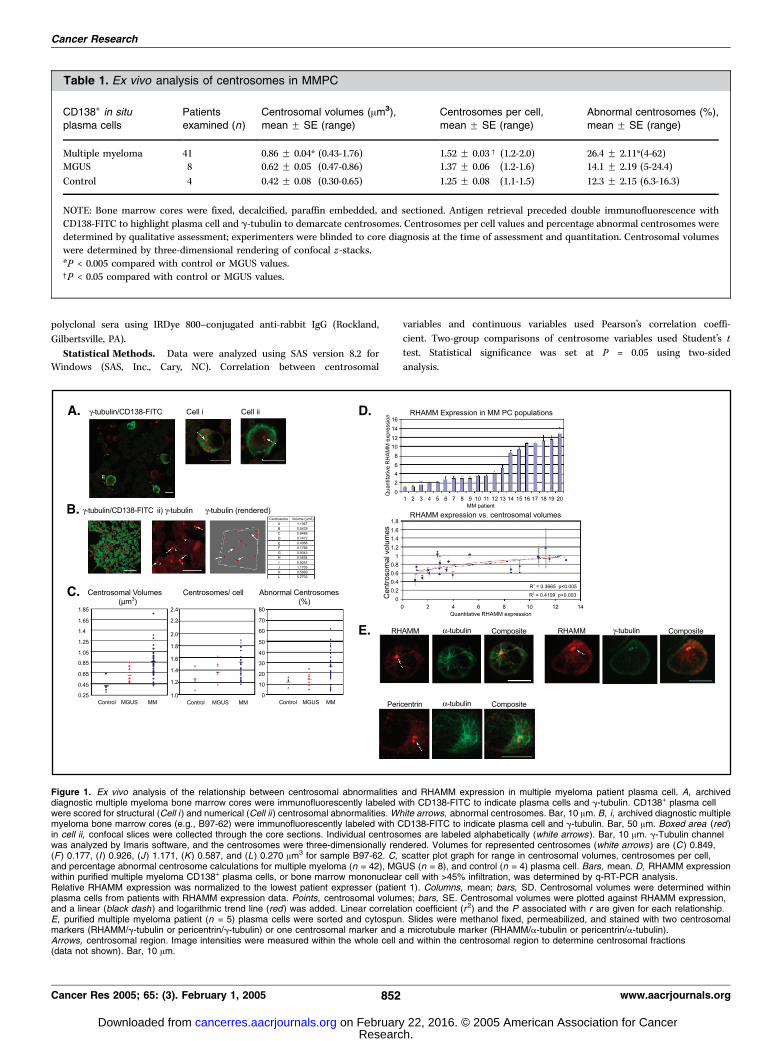
Figure 1. Ex vivo analysis of the relationship between centrosomal abnormalities and RHAMM expression in multiple myeloma patient plasma cell. A, archiveddiagnostic multiple myeloma bone marrow cores were immunofluorescently labeled with CD138-FITC to indicate plasma cells and g-tubulin. CD138+ plasma cellwere scored for structural (Cell i ) and numerical (Cell ii ) centrosomal abnormalities. White arrows, abnormal centrosomes. Bar, 10 Am. B, i, archived diagnostic multiplemyeloma bone marrow cores (e.g., B97-62) were immunofluorescently labeled with CD138-FITC to indicate plasma cell and g-tubulin. Bar, 50 Am. Boxed area (red )in cell ii, confocal slices were collected through the core sections. Individual centrosomes are labeled alphabetically (white arrows ). Bar, 10 Am. g-Tubulin channelwas analyzed by Imaris software, and the centrosomes were three-dimensionally rendered. Volumes for represented centrosomes (white arrows ) are (C) 0.849,(F ) 0.177, (I ) 0.926, (J) 1.171, (K) 0.587, and (L) 0.270 Am3 for sample B97-62. C, scatter plot graph for range in centrosomal volumes, centrosomes per cell,and percentage abnormal centrosome calculations for multiple myeloma (n = 42), MGUS (n = 8), and control (n = 4) plasma cell. Bars, mean. D, RHAMM expressionwithin purified multiple myeloma CD138+ plasma cells, or bone marrow mononuclear cell with >45% infiltration, was determined by q-RT-PCR analysis.Relative RHAMM expression was normalized to the lowest patient expresser (patient 1). Columns, mean; bars, SD. Centrosomal volumes were determined withinplasma cells from patients with RHAMM expression data. Points, centrosomal volumes; bars, SE. Centrosomal volumes were plotted against RHAMM expression,and a linear (black dash ) and logarithmic trend line (red) was added. Linear correlation coefficient (r2) and the P associated with r are given for each relationship.E, purified multiple myeloma patient (n = 5) plasma cells were sorted and cytospun. Slides were methanol fixed, permeabilized, and stained with two centrosomalmarkers (RHAMM/g-tubulin or pericentrin/g-tubulin) or one centrosomal marker and a microtubule marker (RHAMM/a-tubulin or pericentrin/a-tubulin).Arrows, centrosomal region. Image intensities were measured within the whole cell and within the centrosomal region to determine centrosomal fractions(data not shown). Bar, 10 Am.
Cancer Research
Cancer Res 2005; 65: (3). February 1, 2005 852 www.aacrjournals.org
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Results
Centrosome Abnormalities Characterize Myeloma Cells.Myeloma plasma cells are characterized by numerous and complexchromosomal abnormalities (32). Given that centrosomal abnor-malities are often present in chromosomally aberrant malignancies(17), we used multicolor immunofluorescence to examine MMPCfrom archived bone marrow core biopsies for centrosomalabnormalities. Bone marrow cores were examined from multiplemyeloma patients (n = 41), MGUS patients (n = 8), and controlpatients (n = 4) plasma cells. Plasma cells from the morphologicallynormal bone marrow of lymphoma patients were used as controls.These plasma cells would be expected to have normal centrosomes.[If somehow cryptic lymphoma cells were mistaken for plasmacells, then this would be expected to lead to an overestimation ofcontrol plasma cell centrosome abnormalities (33). Such an errorwould only bias the results against the conclusions drawn here.]Experimenters were blinded to sample identity/diagnosis at thetime of analysis and quantitation. Centrosomes in CD138+ plasmacells were visually assessed for qualitative structural and numericalabnormalities (see Table 1, column 5) as described previously (17).To quantitate structural abnormalities within CD138+ cells, plasmacell centrosomal volumes were determined by three-dimensionalrendering of confocal z-stacks labeled with g-tubulin, a definedcentrosomal structural protein (Fig. 1B).Plasma cell from all multiple myeloma samples analyzed showed
numerical, structural, and volumetric abnormalities (Fig. 1A).Centrosome abnormalities, including the mean number ofcentrosomes per cell and the mean total centrosome volume,
were highly correlated with one another in multiple myeloma(Pearson’s r2 coefficient = 0.27; P < 0.001). On average, multiplemyeloma plasma cells showed a significantly greater number ofcentrosomes per cell than MGUS (P < 0.05) and control plasmacells (P < 0.04). Quantitative volumetric assessment of structuralabnormalities revealed a highly significant difference betweenmultiple myeloma and MGUS (P < 0.002) and control plasma cells(P < 0.004). MGUS plasma cells had centrosomal variablesintermediate to those of control plasma cells and multiplemyeloma plasma cells (Table 1); indeed, MGUS plasma cells hadlarger centrosomal volumes when compared with those withincontrol plasma cells.
RHAMM is, among its other functions, a centrosome protein (7).
Given the presence of centrosome abnormalities in myeloma and
the clinical significance of elevated RHAMM expression in this
disease (10), we investigated whether elevated expression of
RHAMM in multiple myeloma correlates with centrosome
abnormalities.In Myeloma, RHAMM Expression Correlates with Volumetric
Centrosome Abnormalities. We used q-RT-PCR to investigateRHAMM expression within multiple myeloma patients (n = 20) withpreviously investigated centrosome variables. RNA was obtainedfrom multiple myeloma patient CD138+ plasma cell (n = 5), or bonemarrowmononuclear cell withz40% plasma cell infiltration (n = 15)when purified plasma cells were unavailable, for quantitativeanalysis. Within this multiple myeloma cohort, RHAMM expressionwas normalized to the level of the lowest RHAMM expresser (Fig.1D). As with centrosomal abnormalities, multiple myeloma patients
Table 2. Dysregulation of centrosomal volumes and mitosis with GFP-RHAMMFL overexpression
Transfection RPMI 8226, mean
centrosomal volume (Am3)
HeLa, % of mitotic cells (abnormal, % of total N)
n Prophase Prometaphase Metaphase Anaphase Telophase/
cytokinesis
Total Monopolar/multipolar*
EGFP-C1 0.54 F 0.081c 182 (9.3) 12.1 (0) 9.3 (0.5) 38.5 (6.6) (3.3) 6.6 (1.1) 33.5 (1.1)
Non transfected 0.532 F 0.123c 306 (5.5) 9.8 (0) 6.9 (0) 42.8 (3.3) (2.9) 8.2 (0) 29.1 (1.0)
GFP-RHAMMFL 0.682 F 0.079c,b 153 (49.7) 3.9 (0.7) 15 (3.3) 65.4 (41.2) (16.3) 5.2 (2.0) 13.1 (2.6)
1.039 F 0.262x,k
NOTE: RPMI 8226 cells were transiently transfected with GFP-RHAMMFL using electroporation. Following transfection, live cells were adhered to 1 mg/mLfibronectin, methanol fixed, and stained for gg-tubulin. At least 10 measurements (i.e., confocal stacks and three-dimensional rendering) were acquired
for GFP-RHAMMFL and gg-tubulin signals within transfected and nontransfected cells. HeLa cells were grown on coverslips and transiently transfected
with LipofectAMINE 2000 according to manufacturer’s suggested protocol. Three coverslips were examined for EGFP-C1 and GFP-RHAMMFL
transfections. Transfections resulted in 60-70% transfectants; both transfected (60-70%) and nontransfected (30-40%) populations were analyzed onGFP-RHAMMFL transfected slips. Coverslips were fixed and permeabilized in cold methanol 20 hours post-transfection and immunofluorescently
labeled for aa-tubulin to detect microtubules and DAPI to detect DNA. DNA condensation, alignment and segregation, bipolar spindle formation, and
midzone microtubules were used to differentiate mitotic stages. Binucleated prophase, aberrant/disorganized bipolar spindles, monopolar/tripolar/
tetrapolar/multipolar spindles, tripolar/multipolar anaphase segregation, lagging anaphase chromosomes, telophase chromosomal bridges, and tripolartelophase/cytokinesis daughter cells were scored as abnormal. Number of transfected cells examined are indicated inside parentheses.
*Percentage of total mitotic cells with multipolar or monopolar metaphase spindles.cMeasurement of gg-tubulin centrosomal signal.bP = 0.22 compared with controls.xMeasurement of GFP-RHAMMFL centrosomal signal.kP < 0.05 compared with controls.
RHAMM Affects Centrosomal and Spindle Integrity
www.aacrjournals.org 853 Cancer Res 2005; 65: (3). February 1, 2005
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

varied considerably in the absolute amount of RHAMM expression(Fig. 1D).Centrosome volumes were measured in CD138-FITC-labeled
multiple myeloma plasma cells from multiple myeloma patientsfor which the RHAMM q-RT-PCR expression level was known (n =20). Regression analysis revealed a significant linear (r2 = 0.367;P < 0.005) and logarithmic relationship (r2 = 0.411; P < 0.003)between RHAMM expression and centrosome volumes (Fig. 1D).Centrosome per cell quantitation, however, did not show asignificant linear relationship with RHAMM expression (r2 = 0.189;P = 0.104; data not shown).RHAMM Localizes to Multiple Myeloma Centrosomes to the
Same Extent as Other Centrosome Proteins. Next, we investi-gated the fraction of intracellular RHAMM, which localizes to thecentrosome within MMPC. After CD38 or CD138 fluorescence-activated cell sorting separation, multiple myeloma patient (n = 6)plasma cells were stained with either two centrosomal markers (i.e.,pericentrin, RHAMM, org -tubulin) or one centrosomal marker anda-tubulin to show microtubule nucleation. Image analysis ofconfocal slices containing the centrosomal region revealed similarcentrosomal fractions for pericentrin, RHAMM, and -tubulin (0.10F0.04, 0.08 F 0.04, and 0.06 F 0.03, respectively). Thus, although theintracellular fraction of RHAMM that localizes to interphasemyeloma centrosomes is low, it is comparable with that of otherdefined components of the centrosome. As expected, intracellularRHAMM, but not pericentrin or g-tubulin, also colocalized withmicrotubules (Fig. 1E).
RHAMM Overexpression In vitro Affects Centrosome Sizeand Structure. To investigate a potential cause-effect relationship,we next examined whether overexpression of RHAMM in vitrocould amplify centrosome volumes in a myeloma cell line. RPMI8226 cells were transiently transfected with EGFP-C1 empty vectoror GFP-RHAMMFL, and centrosomal structure was examined 16 to20 hours later. Previous analysis indicates that transient transfec-tion results in f5-fold overexpression of exogenous RHAMM (7).Centrosomal volumes were determined with indirectimmunofluorescence targeting g-tubulin or GFP-RHAMMFL
fluorescence (Fig. 2A). g-Tubulin fluorescence within (a) cellstransfected with EGFP-C1 and (b) cells that failed to express GFP-RHAMMFL (internal control) was used as a control for pericen-triolar material volume determination (Fig. 2A). The meancentrosomal volumes for negative control samples were 0.541 F0.081 and 0.532 F 0.123 m3, respectively (Table 2). Overexpressionof GFP-RHAMMFL resulted in mean pericentriolar materialvolumes of 1.039 F 0.262 m3 and a trend toward larger -tubulinvolumes (0.682 F 0.079 m3; Table 2). Thus, RHAMM overexpressionadds a significant amount of excess pericentriolar material at thecentrosome.RHAMM Overexpression Compromises Mitotic Integrity.
Structural centrosomal abnormalities, such as those that correlatewith RHAMM expression in myeloma (Fig. 1D), have been stronglyrelated to abnormal mitoses in human cancer (16). Thus, weanalyzed the effect of RHAMM overexpression on mitotic integrityin vitro .
Figure 2. RHAMM overexpression affects centrosomalstructure and mitotic integrity. A, RPMI 8226 cellswere transiently transfected with GFP-RHAMMFL,or EGFP-C1, and stained with g-tubulin. Confocal z -stackswere collected through individual cells and regionsof interest, containing the g-tubulin signal, were exportedto Imaris for three-dimensional rendering. Bar, 10 Am.Inset boxes, 2.5 � 2.5 Am extracted regions aroundthe centrosome. A median (of 2 pixels) filter was appliedto extracted regions. Effective zoom is 4� original image.Bar, 1 Am. B, HeLa cells were transiently transfectedwith GFP-RHAMMFL, or EGFP-C1, and stained witha-tubulin (data not shown). DNA was visualized with DAPI.Both transfected and nontransfected cells were scoredwithin GFP-RHAMMFL transfected populations. Cellswere examined for normal DNA alignment and segregationas well as for symmetrical, organized bipolar spindles(GFP-RHAMMFL and a-tubulin signals). Individualmetaphase cells were scored as normal, disorganizedspindles, unaligned chromosomes, monopolar,tripolar, tetrapolar, or multipolar spindles. Cells scoredas unaligned chromosomes had phenotypicallynormal spindles. Bar, 10 Am.
Cancer Research
Cancer Res 2005; 65: (3). February 1, 2005 854 www.aacrjournals.org
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
RHAMM mitotic localization has been documented inmultiple different human cell models (34, 4, 7) and the mitoticfunction of RHAMM is conserved in Xenopus (35). Given theconservation of mitotic localization and function, transiently
transfected HeLa cells were examined as they represent a standardin vitro system within which mitoses, and aberrant mitoses, can beanalyzed and compared. EGFP-C1 transfected cells and neighboring,nontransfected cells (internal control) were used as negative
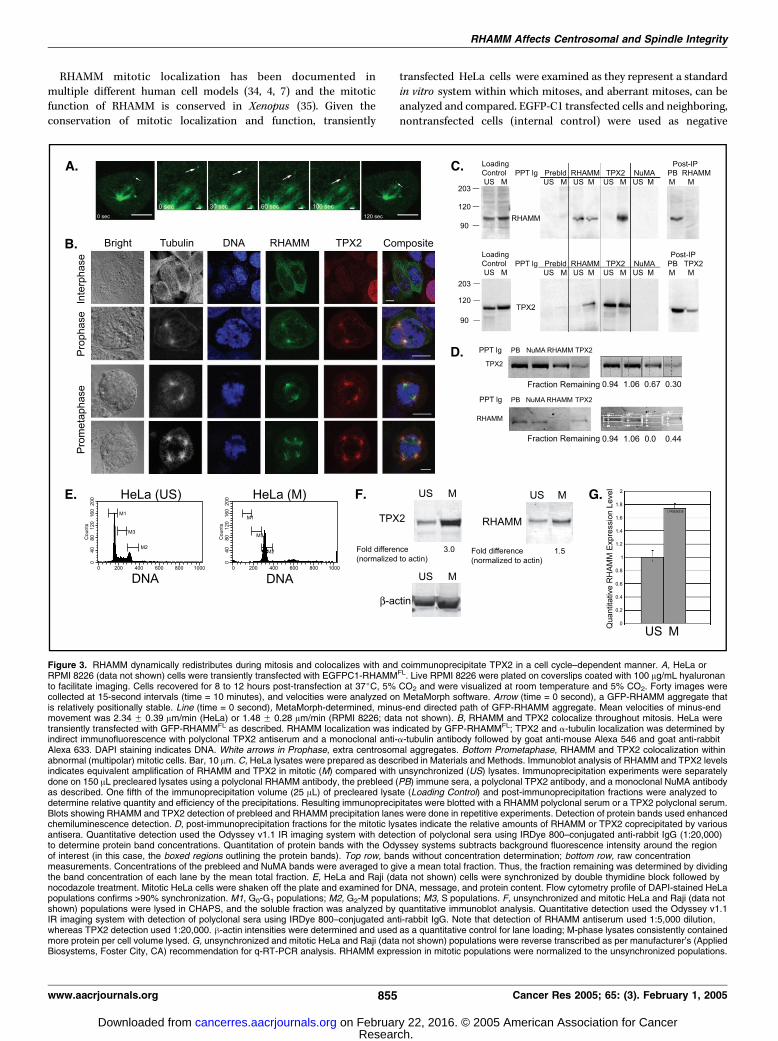
Figure 3. RHAMM dynamically redistributes during mitosis and colocalizes with and coimmunoprecipitate TPX2 in a cell cycle–dependent manner. A, HeLa orRPMI 8226 (data not shown) cells were transiently transfected with EGFPC1-RHAMMFL. Live RPMI 8226 were plated on coverslips coated with 100 Ag/mL hyaluronanto facilitate imaging. Cells recovered for 8 to 12 hours post-transfection at 37jC, 5% CO2 and were visualized at room temperature and 5% CO2. Forty images werecollected at 15-second intervals (time = 10 minutes), and velocities were analyzed on MetaMorph software. Arrow (time = 0 second), a GFP-RHAMM aggregate thatis relatively positionally stable. Line (time = 0 second), MetaMorph-determined, minus-end directed path of GFP-RHAMM aggregate. Mean velocities of minus-endmovement was 2.34 F 0.39 Am/min (HeLa) or 1.48 F 0.28 Am/min (RPMI 8226; data not shown). B, RHAMM and TPX2 colocalize throughout mitosis. HeLa weretransiently transfected with GFP-RHAMMFL as described. RHAMM localization was indicated by GFP-RHAMMFL; TPX2 and a-tubulin localization was determined byindirect immunofluorescence with polyclonal TPX2 antiserum and a monoclonal anti-a-tubulin antibody followed by goat anti-mouse Alexa 546 and goat anti-rabbitAlexa 633. DAPI staining indicates DNA. White arrows in Prophase, extra centrosomal aggregates. Bottom Prometaphase, RHAMM and TPX2 colocalization withinabnormal (multipolar) mitotic cells. Bar, 10 Am. C, HeLa lysates were prepared as described in Materials and Methods. Immunoblot analysis of RHAMM and TPX2 levelsindicates equivalent amplification of RHAMM and TPX2 in mitotic (M) compared with unsynchronized (US) lysates. Immunoprecipitation experiments were separatelydone on 150 AL precleared lysates using a polyclonal RHAMM antibody, the prebleed (PB) immune sera, a polyclonal TPX2 antibody, and a monoclonal NuMA antibodyas described. One fifth of the immunoprecipitation volume (25 AL) of precleared lysate (Loading Control) and post-immunoprecipitation fractions were analyzed todetermine relative quantity and efficiency of the precipitations. Resulting immunoprecipitates were blotted with a RHAMM polyclonal serum or a TPX2 polyclonal serum.Blots showing RHAMM and TPX2 detection of prebleed and RHAMM precipitation lanes were done in repetitive experiments. Detection of protein bands used enhancedchemiluminescence detection. D, post-immunoprecipitation fractions for the mitotic lysates indicate the relative amounts of RHAMM or TPX2 coprecipitated by variousantisera. Quantitative detection used the Odyssey v1.1 IR imaging system with detection of polyclonal sera using IRDye 800–conjugated anti-rabbit IgG (1:20,000)to determine protein band concentrations. Quantitation of protein bands with the Odyssey systems subtracts background fluorescence intensity around the regionof interest (in this case, the boxed regions outlining the protein bands). Top row, bands without concentration determination; bottom row, raw concentrationmeasurements. Concentrations of the prebleed and NuMA bands were averaged to give a mean total fraction. Thus, the fraction remaining was determined by dividingthe band concentration of each lane by the mean total fraction. E, HeLa and Raji (data not shown) cells were synchronized by double thymidine block followed bynocodazole treatment. Mitotic HeLa cells were shaken off the plate and examined for DNA, message, and protein content. Flow cytometry profile of DAPI-stained HeLapopulations confirms >90% synchronization. M1, G0-G1 populations; M2, G2-M populations; M3, S populations. F, unsynchronized and mitotic HeLa and Raji (data notshown) populations were lysed in CHAPS, and the soluble fraction was analyzed by quantitative immunoblot analysis. Quantitative detection used the Odyssey v1.1IR imaging system with detection of polyclonal sera using IRDye 800–conjugated anti-rabbit IgG. Note that detection of RHAMM antiserum used 1:5,000 dilution,whereas TPX2 detection used 1:20,000. h-actin intensities were determined and used as a quantitative control for lane loading; M-phase lysates consistently containedmore protein per cell volume lysed. G, unsynchronized and mitotic HeLa and Raji (data not shown) populations were reverse transcribed as per manufacturer’s (AppliedBiosystems, Foster City, CA) recommendation for q-RT-PCR analysis. RHAMM expression in mitotic populations were normalized to the unsynchronized populations.
RHAMM Affects Centrosomal and Spindle Integrity
www.aacrjournals.org 855 Cancer Res 2005; 65: (3). February 1, 2005
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

controls for spindle abnormalities induced by GFP-RHAMMFL
overexpression.Similar to observations in RPMI 8226 (7), GFP-RHAMMFL
transfection results in a metaphase block with dramatically
fewer transfected cells observed in prophase (3.92% versus 12.1%
and 9.8%) and telophase/cytokinesis (13.1% versus 33.5% and
29.1%) compared with EGFP-C1 and nontransfected controls
(Table 2). Despite the high transfection efficiency (60-70%),
fewer GFP-RHAMMFL transfected cells were mitotic (n = 153)
than nontransfected internal control cells (n = 306), suggesting
a significant decrease in viable mitotic cells dependent on
GFP-RHAMMFL overexpression (Table 2).To further investigate the effects of RHAMM overexpression on
the dysregulation of mitosis, the number of abnormal mitotic
cells, at various stages, was quantitated. DNA condensation,
alignment, and segregation as well as the structure and number of
spindle poles were analyzed to differentiate between normal and
abnormal mitotic cells. The criteria for abnormal metaphase and
anaphase was similar to that reported for Aurora A–inhibited
S2 cells (36). GFP-RHAMMFL overexpression induced multiple
abnormalities at various stages, with the most frequent being
disorganized/long metaphase spindle fibers (18.3%; data not
shown) and monopolar/tripolar/tetrapolar/multipolar metaphase
spindles (16.3%; Fig. 2B); the frequency of disorganized spindle
phenotypes (0% and 0.5%; data not shown) and mono/multipolarspindles (2.9% and 3.3%) was significantly reduced withinnontransfected and EGFP-C1 transfected control populations,respectively (Table 2). Few (n = 28, 18.3%) mitotic, GFP-RHAMMFL
overexpressing cells were observed post-metaphase (Table 2).Interestingly, 7 of 28 (25%) transfected cells showed laggingchromosomes, multipolar segregation, or chromosome bridges(Table 2). These post-metaphase phenotypes were present in 3 of114 (2.6%, 1.0% of total) and 4 of 73 (5.5%, 1.1% of total) of non-transfected and EGFP-C1 transfected controls.Given the evidence from clinical myeloma samples and in vitro
transfection studies that link RHAMM to centrosome and mitotic
errors, we sought to elucidate some of the functional aspects of
RHAMM at the mitotic spindle that might be operative in
producing these errors. The centrosome targeting domain of
RHAMM bears significant homology to the BZIP motif of Xklp2,
which interacts with dynein and TPX2 to target minus-end
microtubules (7, 8). We therefore focused our efforts on the
interaction of RHAMM with dynein and TPX2.RHAMM Functionally Interacts with Dynein in Mitosis.
Previous investigation of RHAMM function revealed that a subset
of RHAMM colocalized with and coprecipitated dynein and vice
versa (7). To extend on these findings, we investigated the
dynamics of GFP-RHAMMFL in transiently transfected live
HeLa and RPMI 8226 cells at room temperature (22-25jC). Inaddition to the mitotic spindle association, we observed
aggregates of GFP-RHAMMFL that would load to the spindle
pole with vectoral movement (Fig. 3A). On average, GFP-
RHAMMFL cells contained 13.25 aggregates, with 17% demon-
strating movement during the time course. We observed poleward
movement of RHAMMFL-GFP aggregates with velocities [2.34 F0.39 (HeLa, n = 7) and 1.48 F 0.28 (8226, n = 5) m/min; Fig. 3A]
similar to those observed for minus-end directed movement of
NuMA-GFP by cytoplasmic dynein (1.0 F 0.3 to 2.6 F 1.0 m/min;
ref. 37). Although singular, larger aggregates were observed within
EGFP transfected cells, these aggregates did not transit during the
experiment (t = 15 minutes). These observations, in combination
with our previous coprecipitation data (7), show that RHAMM
associates with the dynein motor complex to localize to the
spindle pole within live cells.RHAMM Colocalizes with and Coimmunoprecipitates TPX2
and Vice Versa in a Cell Cycle–Dependent Manner. We next
investigated whether RHAMM colocalizes with TPX2 during
mitosis. During interphase, GFP-RHAMMFL colocalizes with micro-
tubules and centrosomes, whereas TPX2 is nuclear (Fig. 3B). At
prophase, however, both RHAMM and TPX2 redistribute to the
separating centrosomes (Fig. 3B). At prometaphase and throughout
metaphase, both proteins localize to the poles, and along the arms,
of phenotypically normal as well as multipolar, mitotic spindles
(Fig. 3B). Both proteins localize to the spindle midzone during
anaphase and concentrate at the midzone during telophase (data
not shown).To further investigate a putative RHAMM-TPX2 association,
HeLa cells were synchronized through double thymidine/
nocodazole block to provide mitotic extracts. Mitotic and
unsynchronized extracts were separately immunoprecipitated
with prebleed serum and sera targeting RHAMM, TPX2, and
NuMA. NuMA/dynein complexes play an integral role in spindle
assembly and focusing (37) and thus NuMA served as an
excellent control for the specificity of a putative RHAMM-TPX2
interaction. Lysis of equivalent numbers of cells from mitotic
and unsynchronized populations resulted in amplification of
both RHAMM and TPX2 levels in the mitotic extracts (Fig. 3C).
These results are consistent with previous reports demonstrating
cell cycle regulation of TPX2 (26) and RHAMM (ref. 38; see Fig.
3F and G). The high efficiency of immunoprecipitation was
confirmed by examination of post-immunoprecipitation lysates,
with 70% to 100% of mitotic TPX2 and RHAMM being
precipitated (Fig. 3C). During mitosis, RHAMM antibodies
coprecipitated a significant amount of TPX2 and vice versa.
This reciprocal coprecipitation was not obtained within unsyn-
chronized lysates. To quantitate the level of coprecipitation, IR
detection of protein was used to determine the levels of
RHAMM and TPX2 remaining in the post-immunoprecipitation
lysates (Fig. 3D). Unlike protein detection based on enzymatic
amplification (enhanced chemiluminescence, Amersham Bio-
sciences, Sunnyvale, CA), IR labeling results in linear detection
of protein and, as a consequence, results in differential detection
of RHAMM and TPX2 bands. Thus, the level of mitotic RHAMM
seems greater with enhanced chemiluminescence detection.Neither prebleed nor NuMA precipitation resulted in asignificant loss of RHAMM or TPX2 protein levels in the post-immunoprecipitation fractions. The concentrations of the bandsresulting from prebleed and NuMA precipitations were averagedto give an expected post-precipitation fraction. The fractionremaining was determined by dividing the band concentration ofeach lane by the expected post-precipitation fraction. Immuno-precipitation with RHAMM antibodies resulted in a depletion of36% and 100% of the cellular TPX2 and RHAMM in mitoticlysates, respectively. Within mitotic lysates, TPX2 precipitationresulted in a 70% and 56% loss of TPX2 and RHAMM,respectively (Fig. 3D). These results show that a significantpopulation of RHAMM is associated with TPX2 within HeLamitotic cells and vice versa.
Cancer Research
Cancer Res 2005; 65: (3). February 1, 2005 856 www.aacrjournals.org
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
RHAMM, Like TPX2, Is Regulated during the Cell Cycle. Asmentioned, RHAMM and TPX2 are subject to cell cycle regulation(26, 38) although the level of RHAMM amplification during mitosishas not been reported. HeLa, and Raji (data not shown), lines weresynchronized during G2-M with a double thymidine/nocodazoleblock. The efficiency of these treatments was confirmed by flowcytometry analysis of DAPI staining within the synchronizedpopulations (Fig. 3E). Cell lysates were prepared through equivalent
CHAPS lysis of HeLa (5 � 106/mL) and Raji (20 � 106/mL)synchronized populations. Quantitative immunoblot analysis wasdone by normalizing the RHAMMor TPX2 level to the correspondingh-actin levels. RHAMM protein levels, like TPX2, are cell cycleregulated (Fig. 3F) with 1.5 to 2.0 times greater expression in G2-Mversus unsynchronized Raji and HeLa lysates. q-RT-PCR revealed a1.43 to 1.75 times amplification of RHAMM message in G2-M Rajiand HeLa populations (Fig. 3G).
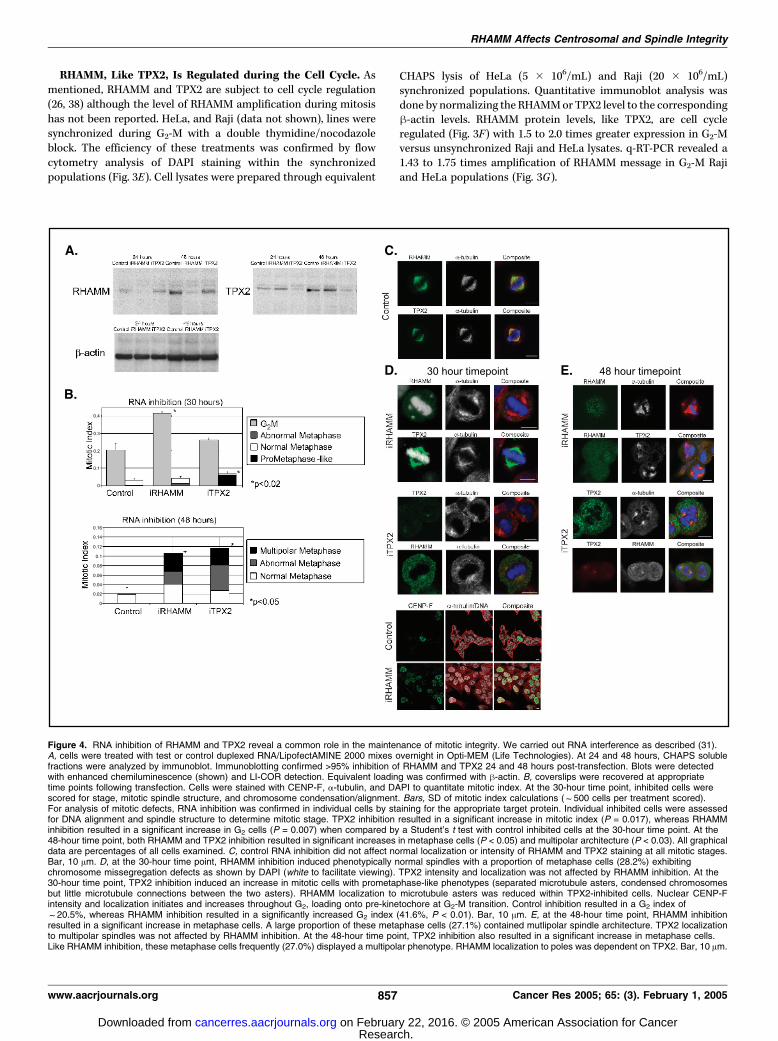
Figure 4. RNA inhibition of RHAMM and TPX2 reveal a common role in the maintenance of mitotic integrity. We carried out RNA interference as described (31).A, cells were treated with test or control duplexed RNA/LipofectAMINE 2000 mixes overnight in Opti-MEM (Life Technologies). At 24 and 48 hours, CHAPS solublefractions were analyzed by immunoblot. Immunoblotting confirmed >95% inhibition of RHAMM and TPX2 24 and 48 hours post-transfection. Blots were detectedwith enhanced chemiluminescence (shown) and LI-COR detection. Equivalent loading was confirmed with h-actin. B, coverslips were recovered at appropriatetime points following transfection. Cells were stained with CENP-F, a-tubulin, and DAPI to quantitate mitotic index. At the 30-hour time point, inhibited cells werescored for stage, mitotic spindle structure, and chromosome condensation/alignment. Bars, SD of mitotic index calculations (f500 cells per treatment scored).For analysis of mitotic defects, RNA inhibition was confirmed in individual cells by staining for the appropriate target protein. Individual inhibited cells were assessedfor DNA alignment and spindle structure to determine mitotic stage. TPX2 inhibition resulted in a significant increase in mitotic index (P = 0.017), whereas RHAMMinhibition resulted in a significant increase in G2 cells (P = 0.007) when compared by a Student’s t test with control inhibited cells at the 30-hour time point. At the48-hour time point, both RHAMM and TPX2 inhibition resulted in significant increases in metaphase cells (P < 0.05) and multipolar architecture (P < 0.03). All graphicaldata are percentages of all cells examined. C, control RNA inhibition did not affect normal localization or intensity of RHAMM and TPX2 staining at all mitotic stages.Bar, 10 Am. D, at the 30-hour time point, RHAMM inhibition induced phenotypically normal spindles with a proportion of metaphase cells (28.2%) exhibitingchromosome missegregation defects as shown by DAPI (white to facilitate viewing). TPX2 intensity and localization was not affected by RHAMM inhibition. At the30-hour time point, TPX2 inhibition induced an increase in mitotic cells with prometaphase-like phenotypes (separated microtubule asters, condensed chromosomesbut little microtubule connections between the two asters). RHAMM localization to microtubule asters was reduced within TPX2-inhibited cells. Nuclear CENP-Fintensity and localization initiates and increases throughout G2, loading onto pre-kinetochore at G2-M transition. Control inhibition resulted in a G2 index off20.5%, whereas RHAMM inhibition resulted in a significantly increased G2 index (41.6%, P < 0.01). Bar, 10 Am. E, at the 48-hour time point, RHAMM inhibitionresulted in a significant increase in metaphase cells. A large proportion of these metaphase cells (27.1%) contained mutlipolar spindle architecture. TPX2 localizationto multipolar spindles was not affected by RHAMM inhibition. At the 48-hour time point, TPX2 inhibition also resulted in a significant increase in metaphase cells.Like RHAMM inhibition, these metaphase cells frequently (27.0%) displayed a multipolar phenotype. RHAMM localization to poles was dependent on TPX2. Bar, 10 Am.
RHAMM Affects Centrosomal and Spindle Integrity
www.aacrjournals.org 857 Cancer Res 2005; 65: (3). February 1, 2005
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Inhibition of RHAMM or TPX2 Function Results in SimilarlyAberrant Mitoses. As significant amounts of RHAMM and TPX2interact during mitosis, we hypothesized that RNA inhibition ofRHAMMmay induce similar phenotypes to that of TPX2 inhibition.RNA inhibition resulted in sustained loss of >95% of cellularRHAMM and TPX2 protein 24 to 48 hours post-transfection (Fig.4A). Cells were lysed and examined by immunoblot 24 and 48hours post-transfection and compared with control-treated cells(GL2). Treatments were examined in triplicate. Inhibited andcontrol cells were then fixed at defined time points and examinedfor mitotic index using the marker CENP-F and a-tubulin. CENP-Fexpression, and localization, varies during the cell cycle, demon-strating increasing nuclear staining during G2 and accumulation onkinetochores early in G2-M transition (39). Mitotic index wasscored for an average of 493 cells per treatment at 30 and 48 hourspost-transfection.At 30 hours, inhibition of TPX2 resulted in an increased mitotic
index (7.0% versus 3.3%; P < 0.02) and the suppression of spindleformation (Fig. 4C). As described by Gruss et al. (26), prometa-phase-like phenotypes, characterized by chromosome condensa-tion, aster separation, and decreased microtubule polymerization,were observed in 82.1% of TPX2-inhibited mitotic cells (Fig. 4D).Within these cells, RHAMM fluorescence was not significantlylocalized to the separated microtubule asters, suggesting a role forTPX2 in the spindle pole targeting of RHAMM (Fig. 4D). Very fewmitotic cells (2.4%) were observed post-metaphase, which stronglysupports the hypothesis that TPX2 is important in the initiation ofspindle assembly (Fig. 4D). However, as TPX2-inhibited cells clearlyproceed through mitosis, albeit at later time points and withsignificantly aberrant spindles (see Fig. 4E), we conclude that it isnot essential for mitotic assembly. We confirm previous work (27)and report that at later time points TPX2 inhibition induces amitotic block with f30.9% of TPX2-inhibited metaphase cellscontaining multipolar spindle phenotypes. Within this populationof cells, we detected reduced amounts of TPX2 at the spindle poles.In the presence of reduced TPX2, RHAMM localized to the poles(Fig. 4E). However, a proportion of TPX2-inhibited cells lackedpolymerized spindles similar to that described above; within thispopulation, RHAMM was not localized to microtubule asters.At 30 hours, RHAMM inhibition resulted in a significant
increase in G2 cells (41.6% versus 20.5%; P < 0.01) characterizedby CENP-F-positive nuclei (Fig. 4B and D). Although bipolarspindles formed by the 30-hour time point, approximately onethird of the metaphase cells had chromosomal missegregationdefects (see Fig. 4D). The increased G2 population suggests thatRHAMM inhibition results in a G2-M stall before the initiation ofprophase. At the 48-hour time point, RHAMM inhibition resultedin an accumulation of mitotic cells in metaphase (see Fig. 4B andE). A large proportion of these metaphase cells (36.8%) containedmultipolar architecture, confirming our previous microinjectionobservations (7). RHAMM inhibition also resulted in a populationof mitotic cells, designated abnormal metaphase, with smallspindles or no spindles (similar to that described for TPX2inhibition). Within the RHAMM-inhibited multipolar populations,TPX2 localized to the mitotic spindle poles (Fig. 4E). At 48 hours,both RHAMM and TPX2 inhibition resulted in dramatic increasesin apoptotic cells and few G1-S cells, demarcated by CENP-F-negative nuclei (data not shown). These results support a role forRHAMM and TPX2 in the maintenance of mitotic integrity,consistent with the conclusions drawn previously for TPX2 (27).Temporal examination of RHAMM and TPX2 inhibition reveal
that these proteins perform distinct roles early in mitoticassembly; inhibition of RHAMM delays mitosis at the G2-Mboundary, whereas TPX2 inhibition delays spindle assembly afternuclear envelope breakdown. These differences in early mitoticfunction are to be expected between proteins that localize to thecentrosome and nucleus during interphase. However, thefunctions of these proteins converge during metaphase. In theabsence of RHAMM (or TPX2), mitotic spindles stall andfragment during metaphase; this phenomenon implicates bothRHAMM and TPX2 as structural components of the spindle thatcross-link microtubules and maintain spindle integrity as variousmicrotubule-dependent motors exert force on spindle micro-tubules.
Discussion
Elevated expression of centrosomal proteins, including pericen-trin and aurA, have been associated with centrosomal abnormal-ities in vivo , and exogenous overexpression of these gene productscan induce centrosomal and chromosomal abnormalities (17, 40).Here, we investigate the relationship among RHAMM, centrosomalabnormalities, and mitotic stability. Elevated RHAMM expressioncharacterizes the malignant multiple myeloma cell (9) andcorrelates with poor prognosis, cytogenetic abnormalities (10),and centrosomal abnormalities within multiple myeloma plasmacell ex vivo . Elevated RHAMM significantly correlates withcentrosomal structural abnormalities as quantitatively determinedby centrosomal volume measurements but not numerical abnor-malities. These data are consistent with RHAMM contributing topericentriolar material structure but not affecting centriolarreplication. Cells with pericentriolar excess are strongly associatedwith abnormal mitoses (16). Accordingly, exogenous GFP-RHAMMFL localizes to centrosomes, increasing their size and theamount of g-tubulin present, and induces aberrant mitoticdivision. Live cell imaging confirms previous coprecipitation dataand suggests that RHAMM targets spindle poles through aninteraction with dynein.RHAMM coprecipitates a significant amount of cellular TPX2
in a cell cycle–dependent manner. RNA inhibition experimentsshow that at early time points TPX2 plays an important, butnonessential, role in spindle assembly, whereas RHAMM functionsin the progression through G2. Interestingly, addition of a solubleRHAMM variant, lacking the NH2-terminal microtubule bindingdomain, also dysregulates G2-M progression (41), whereas over-expression of a RHAMM COOH deletion variant inhibits mitoticprogression (7). After spindle assembly, RHAMM and TPX2converge to play essential roles in maintaining spindle integrity.These data are consistent with a model in which a subset ofintracellular RHAMM contributes to centrosomal structure duringinterphase and functions in early G2-M progression events. Duringspindle assembly, RHAMM interacts with a large fraction of thecellular pool of TPX2, which may facilitate an interaction betweenRHAMM and the dynein motor complex. This ternary RHAMM-TPX2-dynein complex participates in the maintenance of spindleintegrity. Depletion of either TPX2 or RHAMM results in animbalance of motor forces and subsequent spindle fragmentation.Conversely, overexpression of RHAMM results in an oppositeimbalance of force cumulating in disorganized or multipolarspindles and an inability to appropriately align and segregateDNA. Similarly, TPX2 overexpression in Xenopus extracts resultsin unbalanced monopolar spindles (42). Although multipolar
Cancer Research
Cancer Res 2005; 65: (3). February 1, 2005 858 www.aacrjournals.org
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

phenotypes are likely catastrophic, disorganized or fragmentedspindles may induce more subtle losses of genetic material (seeFig. 4D), or impede mitotic progression, and potentially result inaneuploid progeny. The progression of such defects throughmitosis is likely dependent on simultaneous disruption of thep53-dependent cell cycle checkpoint (43). A direct link amongRHAMM overexpression, oncogenesis, and apoptosis may bemediated through TPX2 and aurA.Like RHAMM, aurA localizes to interphase centrosomes,
duplicating prophase centrosomes, and the mitotic spindle pole(7, 44); aurA localization and activation at spindle poles is alsodependent on the action of TPX2 (28). Here, we describe a cellcycle–dependent interaction between significant mitotic fractionsof RHAMM and TPX2, suggesting that RHAMM may affect TPX2-mediated aurA activation. AurA is an essential determinant ofG2-M progression that regulates, and is regulated by, the activitiesof p53 (44), ras-GTPase-activating protein (45), BRCA1 (46), andapoptotic pathways (47). Overexpression of RHAMM, throughan interaction/sequestration of TPX2, may disrupt these path-ways and induce/permit aberrant, or abortive, mitoses with con-sequent centrosome amplification and chromosomal instability.Alternatively, overexpression of RHAMM, or splice variants(i.e., RHAMM�exon4), may augment spindle forces leading to spindlefragmentation and chromosomal segregation defects.Depending on reagents, methodologies, cell models, and culture
conditions, intracellular RHAMM has multiple documentedsubcellular localizations (i.e., interactions with cytoskeletal com-ponents, centrosomes, mitochondria, nuclei, and podosomes)
and functions, including participation in hyaluronan-mediatedmotility, signaling events, and cytoskeletal interactions (34, 4, 7).However, multiple groups, with multiple different reagents, havedocumented RHAMM at the mitotic spindle within mammalianandXenopus systems (4, 34, 7, 35). These observations, alongwith theelevated expression of RHAMM in proliferative tissues like the testis(48) and its cell cycle–dependent regulation, imply a conserved rolefor this protein during cell division. Interestingly, evidence ismounting for the accumulation of intracellular hyaluronan duringmitosis (49), and RHAMM-hyaluronan interactions participate inthe accumulation of hyaluronan within the nucleus during motility(50). Thus, the possibility exists for intercompartmental cross-talkinvolving RHAMM-hyaluronan interactions outside and within thecell during division. However, the relevance of intracellularhyaluronan accumulation, and its effect(s) on RHAMM or otherB(X)7B proteins, is currently undefined.
Acknowledgments
Received 4/22/2004; revised 10/12/2004; accepted 11/22/2004.Grant support: Canadian Institutes of Health Research (L.M. Pilarski, T. Reiman,
and A.R. Belch); National Cancer Institute grant CA80963 (L.M. Pilarski and A.R.Belch); Canada Research in Biomedical Nanotechnology Chairs Program (L.M.Pilarski); and Natural Sciences and Engineering Research Council of Canadastudentship, Alberta Heritage Foundation for Medical Research studentship, andDepartment of Oncology Ph.D. endowed studentship (C.A. Maxwell).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Laith Dabbagh and Sam Johnson for their expert assistance.
References1. Turley EA. Purification of a hyaluronate-bindingprotein fraction that modifies cell social behavior.Biochem Biophys Res Commun 1982;108:1016–24.
2. Yang B, Zhang L, Turley EA. Identification of twohyaluronan-binding domains in the hyaluronan recep-tor RHAMM. J Biol Chem 1993;268:8617–23.
3. Zhang S, Chang MC, Zylka D, Turley S, Harrison R,Turley EA. The hyaluronan receptor RHAMM regulatesextracellular-regulated kinase. J Biol Chem 1998;273:11342–8.
4. Assmann V, Jenkinson D, Marshall JF, Hart IR. Theintracellular hyaluronan receptor RHAMM/IHABPinteracts with microtubules and actin filaments. J CellSci 1999;112:3943–54.
5. Turley EA, Noble PW, Bourguignon LY. Signalingproperties of hyaluronan receptors. J Biol Chem 2002;277:4589–92.
6. Hall CL, Yang B, Yang X, et al. Overexpression of thehyaluronan receptor RHAMM is transforming and is alsorequired for H-ras transformation. Cell 1995;82:19–26.
7. Maxwell CA, Keats JJ, Crainie M, et al. RHAMM is acentrosomal protein that interacts with dynein andmaintains spindle pole stability. Mol Biol Cell 2003;14:2262–76.
8. Wittmann T, Boleti H, Antony C, Karsenti E, Vernos I.Localization of the kinesin-like protein Xklp2 to spindlepoles requires a leucine zipper, a microtubule-associatedprotein, and dynein. J Cell Biol 1998;143:673–85.
9. Crainie M, Belch AR, Mant MJ, Pilarski LM. Over-expression of the receptor for hyaluronan-mediatedmotility (RHAMM) characterizes the malignant clone inmultiple myeloma: identification of three distinctRHAMM variants. Blood 1999;93:1684–96.
10. Maxwell CA, Rasmussen E, Zhan F, et al. RHAMMexpression and isoform balance predict aggressivedisease and poor survival in multiple myeloma. Blood2004;104:1151–8.
11. Rein DT, Roehrig K, Schondorf T, et al. Expressionof the hyaluronan receptor RHAMM in endometrial
carcinomas suggests a role in tumour progressionand metastasis. J Cancer Res Clin Oncol 2003;129:161–4.
12. Li H, Guo L, Li JW, Liu N, Qi R, Liu J. Expression ofhyaluronan receptors CD44 and RHAMM in stomachcancers: relevance with tumor progression. Int J Oncol2000;17:927–32.
13. Wang C, Thor AD, Moore DH, et al. The over-expression of RHAMM, a hyaluronan-binding proteinthat regulates ras signaling, correlates with over-expression of mitogen-activated protein kinase and isa significant parameter in breast cancer progression.Clin Cancer Res 1998;4:567–76.
14. Kellogg DR, Moritz M, Alberts BM. The centrosomeand cellular organization. Annu Rev Biochem 1994;63:639–74.
15. Salisbury JL. Centrosomes: coiled-coils organize thecell center. Curr Biol 2003;13:R88–90.
16. Lingle WL, Salisbury JL. Altered centrosome struc-ture is associated with abnormal mitoses in humanbreast tumors. Am J Pathol 1999;155:1941–51.
17. Pihan GA, Purohit A, Wallace J, et al. Centrosomedefects and genetic instability in malignant tumors.Cancer Res 1998;58:3974–85.
18. Pihan GA, Wallace J, Zhou Y, Doxsey SJ. Centrosomeabnormalities and chromosome instability occurtogether in pre-invasive carcinomas. Cancer Res 2003;63:1398–404.
19. Varmark H. Functional role of centrosomes inspindle assembly and organization. J Cell Biochem2004;91:904–14.
20. arazo-Salas RE, Guarguaglini G, Gruss OJ, Segref A,Karsenti E, Mattaj IW. Generation of GTP-bound Ranby RCC1 is required for chromatin-induced mitoticspindle formation. Nature 1999;400:178–81.
21. Kalab P, Pu RT, Dasso M. The ran GTPaseregulates mitotic spindle assembly. Curr Biol 1999;9:481–4.
22. Kalab P, Weis K, Heald R. Visualization of a Ran-GTPgradient in interphase and mitotic Xenopus eggextracts. Science 2002;295:2452–6.
23. Gruss OJ, Carazo-Salas RE, Schatz CA, et al. Raninduces spindle assembly by reversing the inhibitoryeffect of importina onTPX2 activity. Cell 2001;104:83–93.
24. Tsai MY, Wiese C, Cao K, et al. A Ran signallingpathway mediated by the mitotic kinase Aurora A inspindle assembly. Nat Cell Biol 2003;5:242–8.
25. Schatz CA, Santarella R, Hoenger A, et al. Importina-regulated nucleation of microtubules by TPX2.EMBO J 2003;22:2060–70.
26. Gruss OJ, Wittmann M, Yokoyama H, et al. Chromo-some-induced microtubule assembly mediated by TPX2is required for spindle formation in HeLa cells. Nat CellBiol 2002;4:871–9.
27. Garrett S, Auer K, Compton DA, Kapoor TM. hTPX2is required for normal spindle morphology andcentrosome integrity during vertebrate cell division.Curr Biol 2002;12:2055–9.
28. Kufer TA, Sillje HH, Korner R, Gruss OJ, Meraldi P,Nigg EA. Human TPX2 is required for targeting Aurora-Akinase to the spindle. J Cell Biol 2002;158:617–23.
29. Compton DA, Yen TJ, Cleveland DW. Identificationof novel centromere/kinetochore-associated proteinsusing monoclonal antibodies generated against humanmitotic chromosome scaffolds. J Cell Biol 1991;112:1083–97.
30. Chan GK, Jablonski SA, Sudakin V, Hittle JC, Yen TJ.Human BUBR1 is a mitotic checkpoint kinasethat monitors CENP-E functions at kinetochoresand binds the cyclosome/APC. J Cell Biol 1999;146:941–54.
31. Elbashir SM, Harborth J, Lendeckel W, Yalcin A,Weber K, Tuschl T. Duplexes of 21-nucleotide RNAsmediate RNA interference in cultured mammalian cells.Nature 2001;411:494–8.
32. Fonseca R, Barlogie B, Bataille R, et al. Genetics andcytogenetics of multiple myeloma: a workshop report.Cancer Res 2004;64:1546–58.
33. Kramer A, Schweizer S, Neben K, et al. Centrosomeaberrations as a possible mechanism for chromosomalinstability in non-Hodgkin’s lymphoma. Leukemia2003;17:2207–13.
RHAMM Affects Centrosomal and Spindle Integrity
www.aacrjournals.org 859 Cancer Res 2005; 65: (3). February 1, 2005
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

34. Turley EA, Harrison R. RHAMM, a member of thehyaladherins. In: Hascall VC, Yanagashita M, editors.Science of hyaluronan today, vol. 11; 1999. Availablefrom: http://www.glycoforum.gr.jp/science/hyaluronan/hyaluronanE.html.
35. Groen AC, Coughlin ML, Mitchison TJ, Ohi R.Organization of spindle pole b7y the Xenopus laevishuman homologue RHAMM [abstract L108]. AmericanSociety for Cell Biology Annual Meeting; 2002.
36. Giet R, McLean D, Descamps S, et al. DrosophilaAurora A kinase is required to localize D-TACC tocentrosomes and to regulate astral microtubules. J CellBiol 2002;156:437–51.
37. Merdes A, Heald R, Samejima K, Earnshaw WC,Cleveland DW. Formation of spindle poles by dynein/dynactin-dependent transport of NuMA. J Cell Biol2000;149:851–62.
38. Cho RJ, Huang M, Campbell MJ, et al. Transcriptionalregulation and function during the human cell cycle.Nat Genet 2001;27:48–54.
39. Liao H, Winkfein RJ, Mack G, Rattner JB, Yen TJ.CENP-F is a protein of the nuclear matrix that assembles
onto kinetochores at late G2 and is rapidly degradedafter mitosis. J Cell Biol 1995;130:507–18.
40. Zhou H, Kuang J, Zhong L, et al. Tumour amplifiedkinase STK15/BTAK induces centrosome amplification,aneuploidy and transformation. Nat Genet 1998;20:189–93.
41. Mohapatra S, Yang X, Wright JA, Turley EA, Green-
berg AH. Soluble hyaluronan receptor RHAMM induces
mitotic arrest by suppressing Cdc2 and cyclin B1
expression. J Exp Med 1996;183:1663–8.42. Wittmann T, Wilm M, Karsenti E, Vernos I. TPX2, Anovel Xenopus MAP involved in spindle pole organiza-tion. J Cell Biol 2000;149:1405–18.
43. Raff JW. Centrosomes and cancer: lessons from aTACC. Trends Cell Biol 2002;12:222–5.
44. Katayama H, Sasai K, Kawai H, et al. Phosphorylation
by aurora kinase A induces Mdm2-mediated destabili-
zation and inhibition of p53. Nat Genet 2004;
36:55–62.
45. Gigoux V, L’Hoste S, Raynaud F, Camonis J, Garbay C.Identification of Aurora kinases as RasGAP Src
homology 3 domain-binding proteins. J Biol Chem2002;277:23742–6.
46. Ouchi M, Fujiuchi N, Sasai K, et al. BRCA1phosphorylation by Aurora-A in the regulation of G2
to M transition. J Biol Chem 2004;279:19643–8.
47. Anand S, Penrhyn-Lowe S, Venkitaraman AR. AURO-RA-A amplification overrides the mitotic spindleassembly checkpoint, inducing resistance to Taxol.Cancer Cell 2003;3:51–62.
48. Greiner J, Ringhoffer M, Taniguchi M, et al. Receptorfor hyaluronan acid-mediated motility (RHAMM) is anew immunogenic leukemia-associated antigen inacute and chronic myeloid leukemia. Exp Hematol2002;30:1029–35.
49. Hascall VC, Majors AK, De La Motte CA, et al.Intracellular hyaluronan: a new frontier for inflamma-tion? Biochim Biophys Acta 2004;1673:3–12.
50. Collis L, Hall C, Lange L, Ziebell M, Prestwich R,Turley EA. Rapid hyaluronan uptake is associatedwith enhanced motility: implications for an intracellu-lar mode of action. FEBS Lett 1998;440:444–9.
Cancer Research
Cancer Res 2005; 65: (3). February 1, 2005 860 www.aacrjournals.org
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

2005;65:850-860. Cancer Res Christopher A. Maxwell, Jonathan J. Keats, Andrew R. Belch, et al. Maintains Mitotic IntegrityCentrosome Abnormalities in Multiple Myeloma and Receptor for Hyaluronan-Mediated Motility Correlates with
Updated version
http://cancerres.aacrjournals.org/content/65/3/850
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/65/3/850.full.html#ref-list-1
This article cites 48 articles, 24 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/65/3/850.full.html#related-urls
This article has been cited by 20 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 22, 2016. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
Related Documents











