Journal of Invertebrate Pathology 91 (2006) 19–26 www.elsevier.com/locate/yjipa 0022-2011/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.jip.2005.10.003 Recent and fossil nematode parasites (Nematoda: Mermithidae) of Neotropical ants George Poinar Jr. a,¤ , Jean-Paul Lachaud b , Alfredo Castillo c , Francisco Infante c a Department of Zoology, Oregon State University, Corvallis, OR 97331, USA b Centre de Recherches sur la Cognition Animale (CNRS-UMR 5169), Université Toulouse 3, 118 route de Narbonne, 31062 Toulouse cedex, France c El Colegio de la Frontera Sur (ECOSUR), Carretera Antiguo Aeropuerto, km. 2.5, Tapachula 30700, Chiapas, Mexico Received 3 August 2005; accepted 4 October 2005 Available online 21 November 2005 Abstract The following new recent species of mermithid (Nematoda) parasites of Neotropical ants are described: Meximermis ectatommi n.gen., n. sp. from workers of Ectatomma ruidum Roger in Southern Mexico, Agamomermis cephaloti n. sp. from a worker of Cephalotes minutus (Fab.) collected in Brazil, Agamomermis ecitoni n. sp. from a worker of Eciton burchellii Westwood collected in Venezuela and Agamomer- mis costaricensis n. sp. from a worker of Odontomachus hastatus (Fab.) collected in Costa Rica. In addition, a fossil mermithid parasite of a worker of Linepithema sp. from Dominican amber is described as Heydenius myrmecophila n. sp. These parasites represent the Wrst descriptions of Neotropical ant mermithids and together with previous records listed here, show that ant infection by mermithids in the Neotropics is widespread and has occurred for at least 20–30 million years. 2005 Elsevier Inc. All rights reserved. Keywords: Neotropical ants; Mermithidae; Parasitism; Fossil nematode; Parasitism of ants; Dominican amber 1. Introduction Mermithid parasites of ants have been known for over 250 years (Gould, 1747), yet their life history and eVect on the ant host is poorly known, especially in the tropics. While mermithids are known to deplete the fat body and eventually kill their hosts, one of the more interesting aspects of ant parasitism is that it can produce host interc- astes (Wheeler, 1928). Very little is known about mermithid parasites of Neotropical ants, other than a few observations noticed in pinned specimens that were intercastes (Table 1). The present work describes the Wrst mermithids from Neo- tropical ants, lists the known records of mermithid parasitism of Neotropical ants and shows, with the description of a mer- mithid parasite of a fossil ant in Dominican amber, that ant parasitism by mermithids has a long history in the Neotropics. 2. Materials and methods 2.1. Ectatomma ruidum At the beginning of June 2004, more than 600 E. ruidum workers were collected from 14 nests in a cacao plantation “Rancho San Antonio” (14°557 N; 92°1051W) at 238 m in Tuxtla Chico, Chiapas, Mexico. This plantation con- tained 4500 nests of E. ruidum per hectare. Twenty one of the collected workers had enlarged gasters and were main- tained in the laboratory. Seven of these contained mermith- ids. Three of the seven were preserved as post-parasitic juveniles while the other four were maintained in water until they molted to the adult stage 3 weeks later. 2.2. Other parasitized ants Various workers submitted post-parasitic juvenile mermithids from extant ants. One parasitizing a worker of Cephalotes minutus in Brazil was received from Cesare * Corresponding author. E-mail address: [email protected] (G. Poinar Jr.).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Invertebrate Pathology 91 (2006) 19–26
www.elsevier.com/locate/yjipa
Recent and fossil nematode parasites (Nematoda: Mermithidae) of Neotropical ants
George Poinar Jr. a,¤, Jean-Paul Lachaud b, Alfredo Castillo c, Francisco Infante c
a Department of Zoology, Oregon State University, Corvallis, OR 97331, USAb Centre de Recherches sur la Cognition Animale (CNRS-UMR 5169), Université Toulouse 3, 118 route de Narbonne, 31062 Toulouse cedex, France
c El Colegio de la Frontera Sur (ECOSUR), Carretera Antiguo Aeropuerto, km. 2.5, Tapachula 30700, Chiapas, Mexico
Received 3 August 2005; accepted 4 October 2005Available online 21 November 2005
Abstract
The following new recent species of mermithid (Nematoda) parasites of Neotropical ants are described: Meximermis ectatommi n.gen.,n. sp. from workers of Ectatomma ruidum Roger in Southern Mexico, Agamomermis cephaloti n. sp. from a worker of Cephalotes minutus(Fab.) collected in Brazil, Agamomermis ecitoni n. sp. from a worker of Eciton burchellii Westwood collected in Venezuela and Agamomer-mis costaricensis n. sp. from a worker of Odontomachus hastatus (Fab.) collected in Costa Rica. In addition, a fossil mermithid parasite ofa worker of Linepithema sp. from Dominican amber is described as Heydenius myrmecophila n. sp. These parasites represent the Wrstdescriptions of Neotropical ant mermithids and together with previous records listed here, show that ant infection by mermithids in theNeotropics is widespread and has occurred for at least 20–30 million years. 2005 Elsevier Inc. All rights reserved.
Keywords: Neotropical ants; Mermithidae; Parasitism; Fossil nematode; Parasitism of ants; Dominican amber
1. Introduction
Mermithid parasites of ants have been known for over250 years (Gould, 1747), yet their life history and eVect onthe ant host is poorly known, especially in the tropics.While mermithids are known to deplete the fat body andeventually kill their hosts, one of the more interestingaspects of ant parasitism is that it can produce host interc-astes (Wheeler, 1928). Very little is known about mermithidparasites of Neotropical ants, other than a few observationsnoticed in pinned specimens that were intercastes (Table 1).
The present work describes the Wrst mermithids from Neo-tropical ants, lists the known records of mermithid parasitismof Neotropical ants and shows, with the description of a mer-mithid parasite of a fossil ant in Dominican amber, that antparasitism by mermithids has a long history in the Neotropics.
* Corresponding author.E-mail address: [email protected] (G. Poinar Jr.).
0022-2011/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.jip.2005.10.003
2. Materials and methods
2.1. Ectatomma ruidum
At the beginning of June 2004, more than 600 E. ruidumworkers were collected from 14 nests in a cacao plantation“Rancho San Antonio” (14°55�7� N; 92°10�51�W) at 238 min Tuxtla Chico, Chiapas, Mexico. This plantation con-tained 4500 nests of E. ruidum per hectare. Twenty one ofthe collected workers had enlarged gasters and were main-tained in the laboratory. Seven of these contained mermith-ids. Three of the seven were preserved as post-parasiticjuveniles while the other four were maintained in wateruntil they molted to the adult stage 3 weeks later.
2.2. Other parasitized ants
Various workers submitted post-parasitic juvenilemermithids from extant ants. One parasitizing a worker ofCephalotes minutus in Brazil was received from Cesare

20 G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26
Baroni Urbani (Wrst mentioned in de Andrade and BaroniUrbani, 1999), another from a worker of Odontomachushastatus in Costa Rica was received from John T. Longinoand a third from a worker of Eciton burchellii in Venezuelawas provided by Stefanie BerghoV.
2.3. Fossil ant parasite
A fossil mermithid, together with its Linepithema sp. anthost, occurred in a piece of amber that weighed 3.0 g, andwas 15 mm long by 7 mm wide by 2 mm deep.
The amber specimen originated from mines in the Cor-dillera Septentrional of the Dominican Republic. Datingof Dominican amber is still controversial with the latestpurposed age of 20–15 mya based on foraminifera(Iturralde-Vincent and MacPhee, 1996) and the earliest as45–30 mya based on coccoliths (Cêpek in Schlee, 1990). Arange of ages for Dominican amber may be likely sincethe amber fossils are associated with turbiditic sandstonesof the Upper Eocene to Lower Miocene Mamey Group(Draper et al., 1994). In addition, Dominican amber is sec-ondarily deposited in sedimentary rocks, which makes adeWnite age determination diYcult (Poinar and Mastalerz,2000). A reconstruction of the Dominican amber forestbased on amber fossils indicated that the climate was sim-ilar to that of a tropical moist forest (Poinar and Poinar,1999).
Observations, drawings, and photographs were madewith a Nikon SMZ steroscopic microscope and a NikonOptiphot microscope with magniWcations up to 1000£. Allmeasurements are in microns unless otherwise noted.
3. Results
3.1. Mermithid from Ectatommi ruidum
Mermithidae Braun 1883Meximermis n. gen.Six head papillae, no lip papillae; broad hypodermal peg
at tip of head; vagina short, L-shaped, with sharp bend justbefore it meets uterus; amphids cup-shaped, with wideopenings, shifted slightly dorsally; spicules paired, separate,greatly curved, J-shaped, 2–3 times body width at cloaca;adult and post-parasitic juvenile cuticles without crossWbers; six hypodermal cords; eggs small; post-parasiticjuvenile with tail appendage.
Etymology: Meximermis refers to the country of originof the parasite.
DiagnosisThe absence of cross Wbers in both the adult and post-par-
asitic juvenile cuticles is a common character in mermithidsthat infect aquatic hosts, but is rare in those parasitizing ter-restrial insects. While some adult terrestrial mermithids lackcuticular cross Wbers, such as members of the spider para-site genus Aranimermis Poinar and Benton (1986) (Poinarand Early, 1990), cross Wbers are normally present in thepost-parasitic juveniles. The new genus Meximermis is mostsimilar to members of the genus Aranimermis and shares sixhypodermal cords, six cephalic papillae arranged in oneplane, cup-shaped amphids located below the cephalicpapillae and two long separate spicules. However, thelength and shape of the vagina is quite diVerent in the twogenera. In Aranimermis, the vagina is Xexed 3–6 times
Table 1Mermithids reported from Neotropical ants
a Also referred to as Camponotus (Myrmothrix) abdominalis Fabr. subsp. stercorarius Forel.b Also referred to as Ectatomma tuberculatum Olivier subsp. punctigerum Emery.c Also referred to as Neoponera villosa Fabr. subsp. inversa F. Smith.d Also referred to as Pheidole opaca Mayr subsp. incrustata Forel.
Ant species Mermithid Locality Reference
Camponotus atriceps (F. Smith)a Undetermined Mexico Wheeler (1933)Camponotus punctulatus minutior Forel Undetermined Argentina Wheeler (1929)Cephalotes minutus (Fab.) Undetermined Brazil de Andrade and Baroni Urbani (1999)Cephalotes minutus (Fab.) Agamomermis cephaloti Brazil Present studyEciton burchellii Westwood Agamomermis ecitoni Venezuela Present studyEctatomma ruidum Roger Undetermined Trinidad Weber (1946)Ectatomma ruidum Roger Meximermis ectatommi Mexico Present studyEctatomma tuberculatum (Olivier)b Undetermined Panama and Trinidad Wheeler (1930)Linepithema sp. Heydenius myrmecophila Dominican amber Present studyOdontomachus chelifer (Latr.) Undetermined Brazil Emery (1904)Odontomachus hastatus (Fab.) Agamomermis costaricensis Costa Rica Present studyOdontomachus haematodus (L.) Undetermined Costa Rica Emery (1904)Pachycondyla fuscoatra (Roger) Undetermined Bolivia Emery (1904)Pachycondyla inversa (F. Smith)c Undetermined Colombia and Venezuela Emery (1904); Wheeler (1928)Pachycondyla striata (F. Smith) Undetermined Paraguay Strelnikov (1928)Paraponera clavata (Fab.) Undetermined South America Emery (1904)Pheidole absurda Forel Undetermined Costa Rica Emery (1904)Pheidole gouldi Forel Undertermined Guatemala Wheeler (1928)Pheidole cephalica incrustata Foreld Undetermined Guatemala Wheeler (1928)Pheidole susannae atricolor Forel Undetermined Colombia Wheeler (1928)
G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26 21
before meeting the uterus while in Meximermis, it has onlya single bend. Also, the spicules are much longer (three ormore times body width at cloaca) in Aranimermis. Anothergenus similar to Meximermis is Artyukhovskimermis Popov,however members of the latter genus have extremely longspicules, Xask-shaped amphids and cross Wbers in the adultcuticle (Artyukhovsky, 1990). The genus CamponotimermisIpatieva, Pimenova and Muchamedzianova, whose mem-bers parasitize ants in temperate climates, has small eggsand a vagina similar to Meximermis. However, Camponoti-mermis has Xask-shaped amphids and cross Wbers in the
adult cuticle (Artyukhovsky, 1990). Members of the terres-trial genus Oesophagomermis Artyukhovsky also have cup-shaped amphids and long, curved spicules similar to thosein Meximermis, however the adult cuticle has cross Wbersand the eggs are much larger than those of Meximermis(Artyukhovsky, 1990).
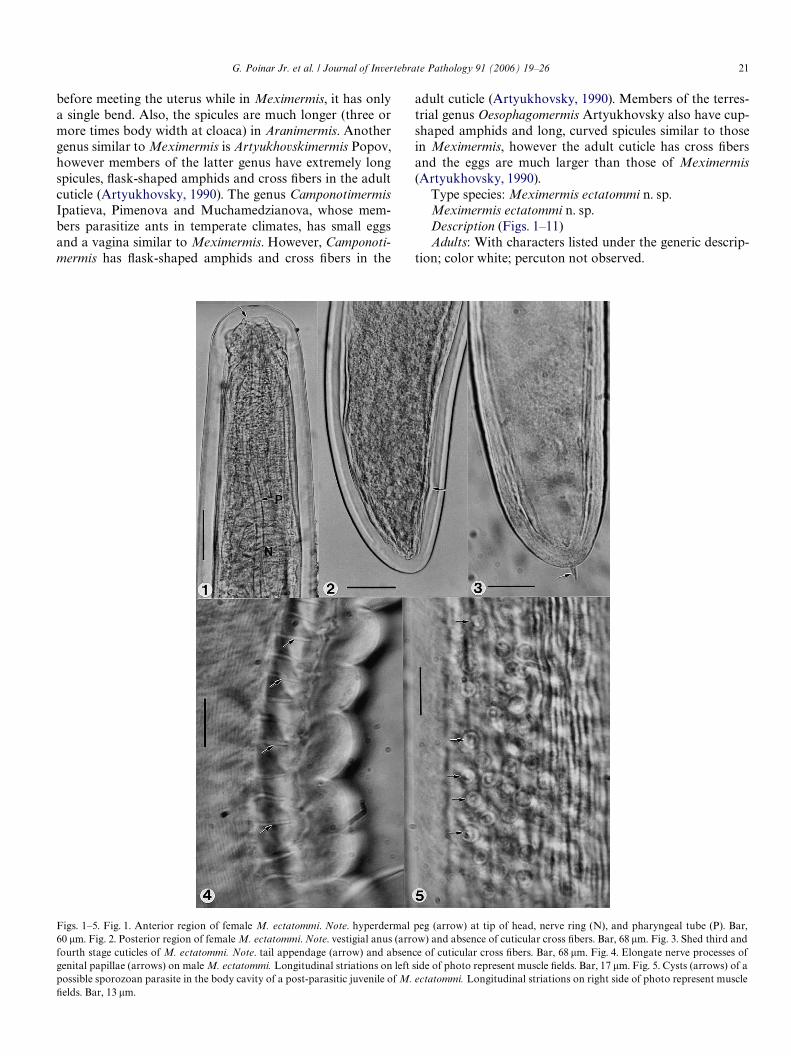
Type species: Meximermis ectatommi n. sp.Meximermis ectatommi n. sp.Description (Figs. 1–11)Adults: With characters listed under the generic descrip-
tion; color white; percuton not observed.
Figs. 1–5. Fig. 1. Anterior region of female M. ectatommi. Note. hyperdermal peg (arrow) at tip of head, nerve ring (N), and pharyngeal tube (P). Bar,60 �m. Fig. 2. Posterior region of female M. ectatommi. Note. vestigial anus (arrow) and absence of cuticular cross Wbers. Bar, 68 �m. Fig. 3. Shed third andfourth stage cuticles of M. ectatommi. Note. tail appendage (arrow) and absence of cuticular cross Wbers. Bar, 68 �m. Fig. 4. Elongate nerve processes ofgenital papillae (arrows) on male M. ectatommi. Longitudinal striations on left side of photo represent muscle Welds. Bar, 17 �m. Fig. 5. Cysts (arrows) of apossible sporozoan parasite in the body cavity of a post-parasitic juvenile of M. ectatommi. Longitudinal striations on right side of photo represent muscleWelds. Bar, 13 �m.

22 G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26
Female: (n D 3) (Figs. 1, 2, 6–9). Length, 87 (68–100) mm.greatest width, 234 (182–299); mouth terminal or shiftedslightly to ventral side; broad hypodermal peg at tip ofhead; pharyngeal tube continues past vulva; amphids cup-shaped, located below lateral cephalic papillae, openingsoval; distance amphids to head, 44 (36–51); length amphid,13 (13–14); width amphid, 17 (14–19); head to nerve ring,270 (247–292); vagina muscular, length vagina, 179 (152–228); vagina with single curve in distal portion; vulva entersvagina at approximately 45° to body wall; % vulva, 58–63;eggs oval, 25 by 20; distance from tail terminus to end oftrophosome, 254 (162–396); tail bluntly pointed, vestigialanus located 61 (53–81) from tail tip.
Male: (n D 1) (Figs. 4 and 10) (head damaged). Length,45 mm; greatest width, 224; length spicules, 423; width spic-ules at proximal end, 11; width spicules at mid-length, 8;spicules paired, separate but closely appressed, especially atapex; tips bluntly pointed, strongly curved upward atapproximately 130°; spicules 2.7 times as long as tail width
at cloaca; with three rows of genital papillae; nerve innerva-tions of genital papillae especially obvious, long and nar-row; 23–25 pre-cloacal papillae in middle and lateral rows;196 post-cloacal papillae in middle row, 61 post-cloacalpapillae in lateral rows; middle row of genital papillaeextends approximately 6 body widths from cloaca; tail withlongitudinal muscle band extending beyond base of spic-ules; tail bluntly pointed; length tail, 253; body width at clo-aca, 153.
Post-parasutic juveniles: (n D 3) (Figs. 3 and 5). Length,44 (38–50) mm; greatest width, 180 (158–208); head trun-cate, without cephalic constriction; tail narrowlyrounded; length tail appendage, 24 (17–36); cuticle inhead region 9 �m wide, in tail region (opposite tailappendage), 28 wide; cuticle lacking cross Wbers; headwith six inconspicuous cephalic papillae; amphids small,posterior to lateral papillae; pharyngeal tube diameter, 9;distance from end of trophosome to head, 577; body cav-ity platelets absent; one specimen with numerous spheres
Figs. 6–11. Fig. 6. Lateral view of head of female M. ectatommi. Bar, 23 �m. Fig. 7. “En face” view of female M. ectatommi. Bar, 20 �m. Fig. 8. Vulvarregion of M. ectatommi. Bar, 38 �m. Fig. 9. Cross section of female M. ectatommi at mid-body. Bar, 49 �m. Fig. 10. Lateral view of male tail ofM. ectatommi. Only middle row of genital papillae shown. Bar, 83 �m. Fig. 11. Lateral view of H. myrmecophila. Note pointed tail. Bar, 84 �m.

G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26 23
approximately 4–6 in diameter (considered a protozoanparasite) in hypodermal cords; shed cuticle double, withfourth stage cuticle thin, contained within thicker thirdstage cuticle.
Type-host: E. ruidum Roger (Hymenoptera: Formici-dae). This diurnal ground-dwelling ectatommine ant ofmedium size (8–9 mm in length) is very common in planta-tions and disturbed habitats as well as in damp forests.Widely distributed from Mexico to northern Brazil (Kuglerand Brown, 1982; Lachaud, 1990), the colonies are small(80–100 individuals on average) but quite numerous inMexican cocoa and coVee plantations, with from 1800 tomore than 11,200 nests per hectare (Levings and Franks,1982; Schatz et al., 1998). Workers typically forage solitar-ily but occasionally exhibit a graduate recruitment (Schatzet al., 1997) and even trail recruitment (Pratt, 1989). Thisspecies is also parasitized by Microdon sp.Xy larvae (Weber,1946) and the eucharitid wasp, Kapala sulcifacies (Howardet al., 2001).
Type-locality: A cacao plantation “Rancho San Anto-nio” (14°55’7” N; 92°10’51”W) at 238 m in Tuxtla Chico,Chiapas, Mexico.
Type-material: (holotype female) T-583t, and allotype(male) T- 584t, deposited in the USDA Nematode Labora-tory, Beltsville, Maryland; Paratypes (free-living femalesand post-parasitic juveniles) deposited in the seniorauthor’s collection.
Etymology: ectatommi refers to the genus of the ant host.
3.2. Mermithids from other recent neotropical ants
Agamomermis Stiles 1903Agamomermis was established as a collective genus for
immature juveniles of mermithid species. If and whenmature worms were obtained, they could be described andassigned to appropriate genera (or new genera) (Poinar andWelch, 1981). Collective genera are recognized in the Codeof Zoological Nomenclature and have been established fornearly 100 years. There are no type species in collectivegenera.
3.2.1. Mermithid from Cephalotes minutusAgamomermis cephaloti n. sp.DescriptionThis is an extremely long and narrow post-parasitic
juvenile female in the process of molting. At this stage, thevulva has not yet formed but there is a partial vagina and itis assumed that the vulva is positioned at the base of thevagina, which would be its normal position. While the headis partially damaged, some features are still discernable.
Post-parasitic juvenile female: (n D 1). Length, 90 mm;head rounded, no cervical constriction; body diameter athead, 94 m, at midbody, 246; diameter of pharyngeal tube 8;cuticle thickness at level of amphid, 7; at midbody, 5; at tail,12; cross Wbers lacking in cuticle; lateral papillae, 6; located38 behind head; amphids short, Xask-shaped, length, 6;amphids located immediately behind lateral cephalic papil-
lae (49 behind head); trophosome begins 60 behind headand extends to 66 from tail tip; diameter of fat globules introphosome, 4–12; body cavity platelets absent; vagina par-tially developed, basal portion straight, 64 in width; head tovagina, 40 mm; vagina to tail, 50 mm; % V D 44; tail nar-rowly rounded, with appendage 20 long.
Host: Cephalotes minutus (Fab) (Hymenoptera: Formici-dae). This little myrmicine ant (3–4 mm in length), commonin both dry and wet Neotropical forests, appears very Xexi-ble in its habitats preferences (Kempf, 1951). Widely dis-tributed from Mexico to Argentina (de Andrade andBaroni Urbani, 1999), colonies nest exclusively in deadstems. Almost nothing is known of its biology.
Etymology: SpeciWc name taken from host genus,Cephalotes.
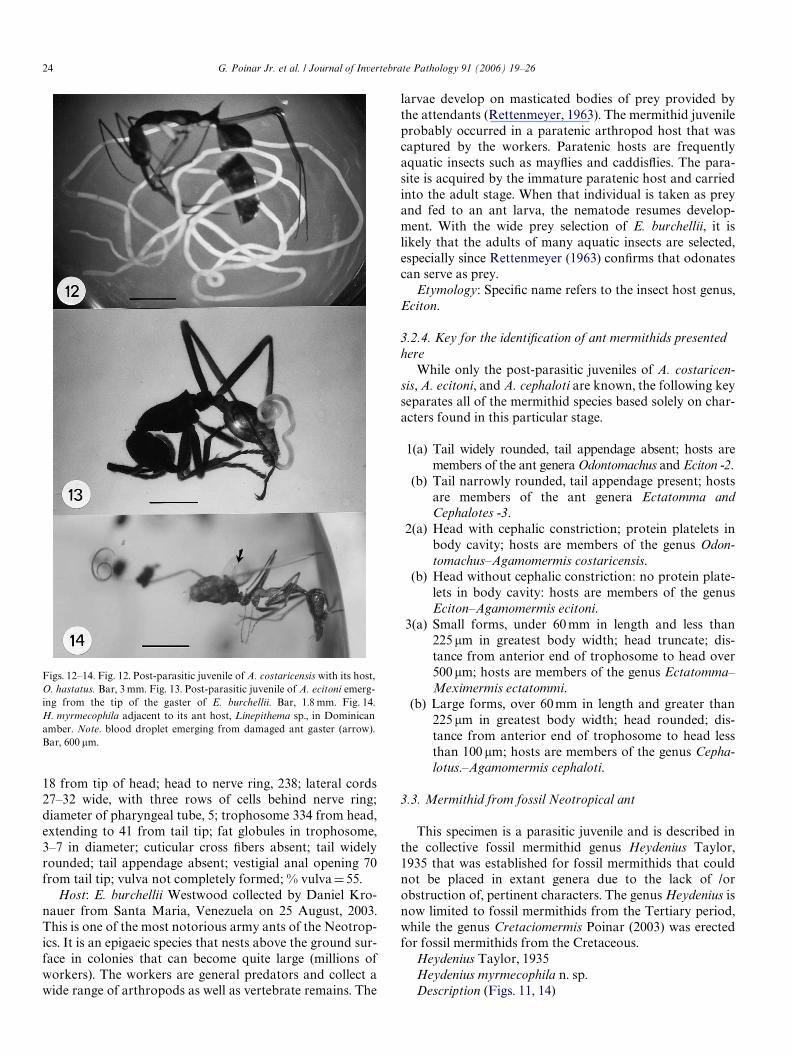
3.2.2. Mermithid from Odontomachus hastatusAgamomermis costaricensis n. sp.Description (Fig. 12)Post-parasitic juvenile: (n D 1). Length, 172 mm; greatest
width, 185; head rounded, with cervical constriction; bodydiameter at head, 127, at midbody, 350; cuticle thickness athead, 14, at midbody, 28, at tail, 25; 6 head papillae located53 from tip of head; head to nerve ring, 247; lateral cords45–50 wide, with three rows of cells behind nerve ring;diameter of pharyngeal tube, 16; trophosome extending to53 from tail tip; fat globules in trophosome, 4–14 in diame-ter; platelets (7–18 in diameter) in body cavity; cuticularcross Wbers absent; tail widely rounded; tail appendageabsent.
Host: Odontomachus hastatus (Fab.) (Hymenoptera:Formicidae). This large and aggressive arboricolous poner-ine ant is provided with a very potent sting. It is mostlyencountered in wet forests where it usually nests in deadbranches, leaf trash gathered in the forks of large trees or inepiphytes. Its range extends from tropical Central to SouthAmerica (Brown, 1976). Its biology is almost unknown butparasitism by an eucharitid parasitoid wasp of the genusKapala has been recorded in Guyana by one of us (JPL,unpublished data).
Etymology: SpeciWc name refers to country of origin,Costa Rica.
3.2.3. Mermithid from Eciton burchelliiAgamomermis ecitoni n. sp.While this specimen has a double cuticle, vestigial vulvar
opening and well-formed cephalic papillae and amphidstypical of post-parasitic juveniles, further development mayhave occurred if the ant host had not been killed. This spe-cies is small in comparison to the other mermithidsdescribed here, which may reXect the restricted gaster sizeof the host (where development normally occurs).
Description (Fig. 13)Post-parasitic juvenile female: (n D 1). Length, 16 mm;
greatest width, 239; head rounded, without cervical con-striction; body diameter at head, 56; cuticle thickness athead, 5; at midbody, 18; at tail, 10; 6 head papillae located
24 G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26
18 from tip of head; head to nerve ring, 238; lateral cords27–32 wide, with three rows of cells behind nerve ring;diameter of pharyngeal tube, 5; trophosome 334 from head,extending to 41 from tail tip; fat globules in trophosome,3–7 in diameter; cuticular cross Wbers absent; tail widelyrounded; tail appendage absent; vestigial anal opening 70from tail tip; vulva not completely formed; % vulva D 55.
Host: E. burchellii Westwood collected by Daniel Kro-nauer from Santa Maria, Venezuela on 25 August, 2003.This is one of the most notorious army ants of the Neotrop-ics. It is an epigaeic species that nests above the ground sur-face in colonies that can become quite large (millions ofworkers). The workers are general predators and collect awide range of arthropods as well as vertebrate remains. The
Figs. 12–14. Fig. 12. Post-parasitic juvenile of A. costaricensis with its host,O. hastatus. Bar, 3 mm. Fig. 13. Post-parasitic juvenile of A. ecitoni emerg-ing from the tip of the gaster of E. burchellii. Bar, 1.8 mm. Fig. 14.H. myrmecophila adjacent to its ant host, Linepithema sp., in Dominicanamber. Note. blood droplet emerging from damaged ant gaster (arrow).Bar, 600 �m.
larvae develop on masticated bodies of prey provided bythe attendants (Rettenmeyer, 1963). The mermithid juvenileprobably occurred in a paratenic arthropod host that wascaptured by the workers. Paratenic hosts are frequentlyaquatic insects such as mayXies and caddisXies. The para-site is acquired by the immature paratenic host and carriedinto the adult stage. When that individual is taken as preyand fed to an ant larva, the nematode resumes develop-ment. With the wide prey selection of E. burchellii, it islikely that the adults of many aquatic insects are selected,especially since Rettenmeyer (1963) conWrms that odonatescan serve as prey.
Etymology: SpeciWc name refers to the insect host genus,Eciton.
3.2.4. Key for the identiWcation of ant mermithids presented here
While only the post-parasitic juveniles of A. costaricen-sis, A. ecitoni, and A. cephaloti are known, the following keyseparates all of the mermithid species based solely on char-acters found in this particular stage.
1(a) Tail widely rounded, tail appendage absent; hosts aremembers of the ant genera Odontomachus and Eciton -2.
(b) Tail narrowly rounded, tail appendage present; hostsare members of the ant genera Ectatomma andCephalotes -3.
2(a) Head with cephalic constriction; protein platelets inbody cavity; hosts are members of the genus Odon-tomachus–Agamomermis costaricensis.
(b) Head without cephalic constriction: no protein plate-lets in body cavity: hosts are members of the genusEciton–Agamomermis ecitoni.
3(a) Small forms, under 60 mm in length and less than225 �m in greatest body width; head truncate; dis-tance from anterior end of trophosome to head over500 �m; hosts are members of the genus Ectatomma–Meximermis ectatommi.
(b) Large forms, over 60 mm in length and greater than225 �m in greatest body width; head rounded; dis-tance from anterior end of trophosome to head lessthan 100 �m; hosts are members of the genus Cepha-lotus.–Agamomermis cephaloti.
3.3. Mermithid from fossil Neotropical ant
This specimen is a parasitic juvenile and is described inthe collective fossil mermithid genus Heydenius Taylor,1935 that was established for fossil mermithids that couldnot be placed in extant genera due to the lack of /orobstruction of, pertinent characters. The genus Heydenius isnow limited to fossil mermithids from the Tertiary period,while the genus Cretaciomermis Poinar (2003) was erectedfor fossil mermithids from the Cretaceous.
Heydenius Taylor, 1935Heydenius myrmecophila n. sp.Description (Figs. 11, 14)

G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26 25
Parasitic juvenile (n D 1): Body completely opaque, inter-nal structures not visible; length, 1.9 mm; greatest width, 56;head truncate; body width at head, 14; attenuated pointedtail.
Specimen: Parasitic juvenile in Dominican amber depos-ited in the Poinar amber collection (Accession No. N-3-65)maintained at Oregon State University.
Host: Linepithema sp. (Hymenoptera: Formicidae).While very few extant Dolichoderinae are represented inthe modern ant fauna of the West Indies, fossil representa-tives are abundance in Dominican amber, especially of thegenera Azteca and Linepithema (Wilson, 1985).
Etymology: From the Greek “myrmekus” for ant.DiagnosisThe shape of this specimen closely resembles that of
developing mermithids in extant insect hosts. The pointedtail is characteristic of early developmental stages ofmermithids as shown in the 1.9 mm long, 3-day old para-sitic juvenile of the midge parasite, Hydromermis conop-ophaga (Poinar, 1975) and the 1.4 mm long, 6-day-oldparasitic juvenile of the beetle parasite, Filipjevimermisleipsandra Poinar and Welch (Poinar, 1968). Developingnematodes sometimes leave their hosts under stressfulconditions, such as being trapped in resin, especially whenthe body wall of the host has been damaged and providesan escape route. In the present case, a droplet of hemo-lymph (containing several hemocytes) is adjacent to awound in the gaster of the ant (Fig. 14, arrow). The nema-tode probably emerged from this opening. A similar situa-tion occurred with two parasitic juveniles of Heydeniusdominicus Poinar (1984) that emerged from their mos-quito host in Dominican amber.
4. Discussion
No life cycle of any mermithid parasite of Neotropicalants is known. The small eggs and muscular vagina ofM. ectatmmi are typical of mermithids that have an indirectlife cycle. In this type of development, the ant (developmen-tal host) becomes infected by eating paratenic hosts con-taining infective stages of the mermithid. Paratenic hosts ofmermithids are usually small aquatic insects, which ingestmermithid eggs by chance while feeding on the bottomdebris. After hatching in the gut of the paratenic host, themermithid juvenile penetrates the gut wall and enters a rest-ing stage in the body cavity. Infection of the ant occurs inthe larval stage when the mermithid is transferred, alongwith tissues of the paratenic host, to the developing antlarva. The mermithid juvenile then enters the body cavity ofthe ant larva and initiates development. The nematode canemerge from the larva, pupa or be carried into the adultstage of the ant. Upon emergence from the ant, the post-parasitic juvenile enters an aquatic or moist habitat tomature, mate, and oviposit.
The European ant mermithid, Pheromermis villosaKaiser, has an indirect type of life cycle (Kaiser, 1986). It ispossible that all of the other mermithids described here
have similar indirect life cycles, including the fossil species.However, we cannot rule out a direct life cycle, where pre-parasitic mermithids emerging from eggs infect the antdirectly (Poinar, 1968).
The cysts in the body of the post-parasitic juvenile of M.ectatommi are considered to represent stages of a sporo-zoan parasite. Similar cysts have been reported from thebody cavity of the New Zealand spider mermithid, Arani-mermis gigantea (Poinar and Early, 1990) and from severalfree-living nematodes (Micoletzky, 1925). Such infectionsare known to be fatal to free-living nematodes under cer-tain conditions (Poinar and Hess, 1988).
All of the mermithids reported by Strelnikov (1928);Wheeler (1928, 1929, 1930, 1933) and Weber (1946) fromNeotropical ants (Table 1) were recognized from morpho-logical modiWcations of the ants brought about by the para-sites. Dissections then revealed the presence of thenematodes, however it appears that none of these nema-todes were removed from the body cavity of their hosts andstudied. It is diYcult to say whether the apparent rarity ofNeotropical ant mermithids reXects their true numbers orwhether they are more abundant, but have been unnoticedor unreported. Yet, it is obvious from the specimen in 20–30 year old Dominican amber that mermithid parasites ofNeotropical ants have had a long period of co-evolution,possibly as long as those of Palearctic ants, which extendsback at least 40 million years (Poinar, 2002).
Acknowledgments
The authors thank José Antonio López-Méndez andAldo Alejandro de la Mora Rodríguez for assistance in col-lecting ants in Chiapas, Mexico, Cesare Baroni Urbani foridentifying the Dominican amber ant and supplying themermithid from Cephalotes, Stefanie BerghoV for submit-ting the mermithid-infected specimen of Eciton, John Long-ino for the nematode parasitized Odontomachus, Alex E.Brown for assistance in obtaining the Dominican amberfossil and Luc Passera for his help in providing additionalreferences to Neotropical ans parasitized by mermithids.
References
de Andrade, M.L., Baroni Urbani, C., 1999. Diversity and adaptation inthe ant genus Cephalotes, past and present (Hymenoptera: Formici-dae). Stuttgarter Beitr. Naturkunde Serie B Nr. 271, 1–889.
Artyukhovsky, A.K., 1990. Soil Mermithids; Systematics, Biology andUtilization. Voronezh University publication, Voronezh (inRussian).
Brown Jr., W.L., 1976. Contributions toward a reclassiWcation of theFormicidae. Part VI. Ponerinae, tribe Ponerini, subtribe Odontomac-hiti. Section A. Introduction, subtribal characters. Genus Odontoma-chus. Studia Entomol. 19, 67–171.
Draper, G., Mann, P., Lewis, J.F., 1994. Hispaniola. In: Donovan, S., Jack-son, T.A. (Eds.), Caribbean Geology: An Introduction, Jamaica. TheUniversity of the West Indies Publishers’ Association, Kingston, pp.129–150.
Emery, C., 1904. Zur Kenntniss des Polymorphismus der Ameisen. Zoolog.Jahrb. Supplement-Band 7, 587–610.

26 G. Poinar Jr. et al. / Journal of Invertebrate Pathology 91 (2006) 19–26
Gould, W., 1747. An Account of English Ants. A. Millar, London.Howard, R.W., Pérez-Lachaud, G., Lachaud, J.-P., 2001. Cuticular hydro-
carbons of Kapala sulcifacies (Hymenoptera: Eucharitidae) and itshost, the ponerine ant Ectatomma ruidum (Hymenoptera: Formicidae).Ann. Entomol. Soc. America 94, 707–716.
Iturralde-Vincent, M., MacPhee, R.D.E., 1996. Age and Paleogeographicorigin of Dominican amber. Science 273, 1850–1852.
Kaiser, H., 1986. Uber Wechselbeziehungen zwischen Nematoden (Mermi-thidae) und Ameisen. Zoolog. Anzeig. 217, 156–177.
Kempf, W.W., 1951. A taxonomic study on the ant tribe Cephalotini(Hymenoptera: Formicidae). Rev. Entomol. (Rio de Janeiro) 22, 1–244.
Kugler, C., Brown, W.L., 1982. Revisionary and other studies on the antgenus Ectatomma, including the descriptions of two new species.Search Agric. 24, 1–7.
Lachaud, J.-P., 1990. Foraging activity and diet in some Neotropical pon-erine ants. I. Ectatomma ruidum Roger (Hymenoptera, Formicidae).Folia Entomol. Mexicana 78, 241–256.
Levings, S.C., Franks, N.R., 1982. Patterns of nest dispersion in a tropicalground ant community. Ecology 63, 338–344.
Micoletzky, H., 1925. Die Freilebenden Süsswasser-und MoornematodenDanemarks. Mém. l’Académie Royale Sci. Lettres Dänemark 10, 1–254.
Poinar Jr., G.O., 1975. Entomogenous Nematodes. Brill, Leiden.Poinar Jr., G.O., 1968. Parasitic development of Filipjevimermis leipsandra
Poinar and Welch in Diabrotica u. undecimpunctata (Chrysomelidae).Proc. Helminthol. Soc. Washington 35, 161–169.
Poinar Jr., G.O., 1984. Heydenius dominicus n. sp. (Nematoda: Mermithi-dae), a fossil parasite from the Dominican Republic. J. Nematol. 16,371–375.
Poinar Jr., G.O., 2002. First fossil record of nematode parasitism of ants; a40 million year tale. Parasitology 125, 457–459.
Poinar Jr., G.O., Welch, H.E., 1981. Parasites of invertebrates in the terres-trial environment. In: Slusarski, W. (Ed.), Review of Advances in Para-sitology. Polish Academy of Sciences, Warsaw, pp. 947–954.
Poinar Jr., G.O., Benton Jr., C.L.B., 1986. Aranimermis aptispicula n.g.,n.sp. (Mermithidae:Nematoda), a parasite of spiders (Arachnida:Araneida). System. Parasitol. 8, 56–61.
Poinar Jr., G.O., Hess, R., 1988. Protozoan Infections. In: Poinar, Jr., G.O.,Jansson, H-B. (Eds.), Diseases of Nematodes, vol. 1. CRC Press, BocaRaton, pp. 103–131.
Poinar Jr., G.O., Early, J.W., 1990. Aranimermis giganteus n. sp. (Mermithi-dae: Nematoda), a parasite of New Zealand mygalomorph spiders(Araneae: Arachnida). Rev. Nematol. 13, 403–410.
Poinar Jr., G.O., Poinar, R., 1999. The Amber Forest: A Reconstruction ofa Vanished World. Princeton University Press, Princeton.
Poinar Jr., G.O., Mastalerz, M., 2000. Taphonomy of fossilized resins: deter-mining the biostratinomy of amber. Acta Geolog. Hispanica 35, 171–182.
Pratt, S.C., 1989. Recruitment and other communication behavior in theponerine ant Ectatomma ruidum. Ethology 81, 313–331.
Rettenmeyer, C.W., 1963. Behavioral studies of Army ants. UniversityKansas Sci. Bull. 44, 281–465.
Schatz, B., Lachaud, J.-P., Beugnon, G., 1997. Graded recruitment andhunting strategies linked to prey weight and size in the ponerine antEctatomma ruidum. Behav. Ecol. Sociobiol. 40, 337–349.
Schatz, B., Lachaud, J.-P., Fourcassié, V., Beugnon, G., 1998. Densité et dis-tribution des nids chez la fourmi Ectatomma ruidum Roger (Hymenop-tera; Formicidae; Ponerinae). Actes Colloq. Insectes Soc. 11, 103–107.
Schlee, D., 1990. Das Bernstein-Kabinett. Stuttgart. Beitrage Naturk. (C),No. 28, 100 pp.
Strelnikov, T.D., 1928. Mermis parasitism in Pachycondyla striata. Psyche35, 199–200.
Weber, N.A., 1946. Two common ponerine ants of possible economic sig-niWcance, Ectatomma tuberculatum (Olivier) and E. ruidum Roger.Proc. Entomol. Soc. Washington 48, 1–16.
Wheeler, W.M., 1928. Mermis parasitism and intercastes among ants. J.Experiment. Zool. 50, 165–237.
Wheeler, W.M., 1929. A Camponotus mermithergate from Argentina. Psy-che 36, 102–106.
Wheeler, W.M., 1930. Two mermithergates of Ectatomma. Psyche 37,48–54.
Wheeler, W.M., 1933. Mermis parasitism in some Australian and Mexicanants. Psyche 40, 20–31.
Wilson, E.O., 1985. Ants of the Dominican amber (Hymenoptera: Form-icidae). 3. The subfamily Dolichoderinae. Psyche 92, 17–37.
Related Documents











