Recent Advances on Multidisciplinary Research at Rudabánya, Late Miocene (MN9), Hungary: a compendium RAYMOND L. BERNOR 1 , LÁSZLÓ KORDOS 2 , LORENZO ROOK 3 JORDI AGUSTÍ 4 ,PETER ANDREWS 5 , MIRANDA ARMOUR-CHELU 6 , DAVID R. BEGUN 7 , DAVID W. CAMERON 8 , JOHN DAMUTH 9 , GUDRUN DAXNER-HÖCK 10 , LOUIS DE BONIS 11 , OLDRICH FEJFAR 12 , NARDOS FESSAHA 1 , MIKAEL FORTELIUS 13 , JENS FRANZEN 14 , MIHÁLY GASPARIK 15 , ALAN GENTRY 5 , KURT HEISSIG 16 , GABOR HERNYAK 17 , THOMAS KAISER 18 , GEORGE D. KOUFOS 19 , ENDRE KROLOPP 2 , DÉNES JÁNOSSY 15 , MANUEL LLENAS 4 , LUKÁCS MESZÁROS 20 , PAL MÜLLER 2 , PAUL RENNE 21 , ZBYNE ˇ K ROčEK 22 , SEVKET SEN 23 , ROBERT SCOTT 24 , ZBIGNIEW SZYNDLAR 25 , GY . TOPÁL 15 , PETER S. UNGAR 26 , TORSTEN UTESCHER 27 , JAN A. VAN DAM 28 , LARS WERDELIN 29 , REINHARD ZIEGLER 30 KEY WORDS – Mammal Faunas, Biogeography, Palaeoecology, Late Vallesian, Late Miocene, Rudabánya, Hungary ABSTRACT – Rudabánya is a rich late Miocene fossil locality first exploited for its vertebrate remains by Pethö in 1902. The first fossil primate was discovered by the local Chief Mining Geologist, Gabor Hernyák. Professor Miklos Kretzoi made Rudabánya famous in 1969 by publishing a manuscript on the new hominoid primate, Rudapithecus hungaricus, recognized herein as Dryopithecus brancoi. In 1991 L. Kordos and R.L. Bernor initiated a project to undertake new excavations and a detailed systematic study of the vertebrate fauna. This 37 author contribution represents a compendium initial report on these studies. A detailed edited volume will follow this publication. We find that there are 112 vertebrate taxa recorded from Rudabánya, 69 of which are fossil mammals. The Rudabánya fauna outcrops at no less than seven different localities, all believed to be essentially synchronous in age. There are no direct radioisotopic dates from Rudabánya, the lower age is constrained by a single crystal argon date of 11.4 m.y. ± 0.1 m.y., and biochronologic correlations support a latest MN 9 age of ca. 10- 9.7 Ma. Paleogeographically, the Rudabánya fauna developed on the edge of the extensive Pannonian lake, which supported a warm, equable subtropical climate. Zoogeographically, Rudabánya is most closely allied with the late Astaracian (MN8)-early Vallesian (MN9) Spanish verte- brate localities, and particularly Can Llobateres (straddling the MN9/MN10 boundary). These central and western European localities contra- st strikingly with correlative eastern Mediterranean-Southwest Asian localities in their community structure. In particular, Rudabánya and Palaeontographia Italica 89 2002 3-36 tavv. Pisa, 2004 1 College of Medicine, Department of Anatomy, Howard University, 520 W St. NW, Washington D.C., USA - [email protected] 2 Geological Institute of Hungary, H-1143 Budapest, Stefánia u. 14, Hungary - [email protected] 3 Dipartimento di Scienze della Terra and Museo di Storia Naturale (Sezione Geologia e Paleontologia), Università di Firenze, via G. La Pira, 4, 50121 Firenze, Italy - [email protected] 4 Institut de Paleontologia M. Crusafont, C/ Escola Industrial 23, 08201-Sabadell, Spain. 5 Department of Palaeontology, Natural History Museum, London, SW7 5BD, U.K. 6 Virginia Museum of Natural History, 1001 Douglas Ave., Martinsville, Virginia 24112, USA. 7 Department of Anthropology, University of Toronto,Toronto, ON M5S 3G3, Canada 8 Department of Anatomy and Histology, Anderson Stuart Building, The University of Sydney, NSW Australia 2006 9 Department of Ecology, Evolution and Marine Biology, University of California, Santa Barbara, California 93106, USA 10 Department of Geology and Paleontology, Naturhistorisches Museum Wien, Burgring 7, A-1014 Vienna, Austria 11 Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine, Université de Poitiers, 40 av. du Recteur Pineau, 86022 Poitiers, France. 12 Department of Paleontology, Charles University Praha, Albertov 6, CZ-12843 Praha, Czech Republik 13 Departmento of Geology, University of Helsinki, P.O. Box 11, FIN-00014, Finland 14 Forschungsinstitut Senckenberg, Abteilung Paläanthropologie, Senckenberganlage 25, D-60325 Frankfurt am Main; Germany 15 Department of Paleontology, Hungarian Museum of Natural History, Muzeum krt. 16-18, Budapest, Hungary. 16 Universitätsinstitut und Staatssammlung für Paläontologie und Historische Geologie, München, Germany 17 Rudabánya, Rózsavölgy u. 2, Hungary 18 Institut und Museum of Zoology, University of Greifswald, D- 17489 Greifswald, Germany 19 Laboratory of Geology and Palaeontology, Aristotle University of Thesaloniki. 54006 Thessaloniki, Greece. 20 Department of Paleontology, Eötvös Loránd University, Ludovika tér 2, H-1083 Budapest, Hungary 21 Berkeley Geochronology Center, Berkeley, California, USA 22 Department of Palaeobiology and Palaeoecology, Geological Institute, Academy of Sciences, Rozvojová 135, Praha, and Department of Zoology, Charles University, Viničná 7, Praha, Czech Republic 23 Laboratoire de Paléontologie, Muséum National d’Histoire Naturelle, UMR 8569 du CNRS, 8 rue Buffon, 75005 Paris, France 24 Department of Anthropology, University of Texas at Austin, Austin, Texas 78712-1086, USA 25 Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Slawkowska 17, 31-016 Kraków, Poland 26 Department of Anthropology, University of Arkansas, Old Main 330, Fayetteville, Arkansas 72701 USA 27 Geologisches Institut, Universität Bonn, Nussallee 8, D - 53115 Bonn 28 Faculty of Earth Sciences, Utrecht University, PO 80021, 3508TA Utrecht, The Netherlands 29 Department of Palaeozoology, Swedish Museum of Natural History, Box 50007, S-104 05 Stockholm, Sweden 30 Staatliches Museum fhr Naturkunde Stuttgart, Rosenstein 1, D-70191 Stuttgart, Germany.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Recent Advances on Multidisciplinary Research at Rudabánya,Late Miocene (MN9), Hungary: a compendium
RAYMOND L. BERNOR1, LÁSZLÓ KORDOS2, LORENZO ROOK3
JORDI AGUSTÍ4, PETER ANDREWS5, MIRANDA ARMOUR-CHELU6, DAVID R. BEGUN7, DAVID W. CAMERON8,JOHN DAMUTH9, GUDRUN DAXNER-HÖCK10, LOUIS DE BONIS11, OLDRICH FEJFAR12, NARDOS FESSAHA1,
MIKAEL FORTELIUS13, JENS FRANZEN14, MIHÁLY GASPARIK15, ALAN GENTRY5, KURT HEISSIG16, GABOR HERNYAK17,THOMAS KAISER18, GEORGE D. KOUFOS19, ENDRE KROLOPP2, DÉNES JÁNOSSY15, MANUEL LLENAS4,
LUKÁCS MESZÁROS20, PAL MÜLLER2, PAUL RENNE21, ZBYNEK ROčEK22, SEVKET SEN23, ROBERT SCOTT24,ZBIGNIEW SZYNDLAR25, GY. TOPÁL15, PETER S. UNGAR26, TORSTEN UTESCHER27, JAN A. VAN DAM28, LARS WERDELIN29,
REINHARD ZIEGLER30
KEY WORDS – Mammal Faunas, Biogeography, Palaeoecology, Late Vallesian, Late Miocene, Rudabánya, Hungary
ABSTRACT – Rudabánya is a rich late Miocene fossil locality first exploited for its vertebrate remains by Pethö in 1902. The first fossilprimate was discovered by the local Chief Mining Geologist, Gabor Hernyák. Professor Miklos Kretzoi made Rudabánya famous in 1969 bypublishing a manuscript on the new hominoid primate, Rudapithecus hungaricus, recognized herein as Dryopithecus brancoi. In 1991 L.Kordos and R.L. Bernor initiated a project to undertake new excavations and a detailed systematic study of the vertebrate fauna. This 37author contribution represents a compendium initial report on these studies. A detailed edited volume will follow this publication. We findthat there are 112 vertebrate taxa recorded from Rudabánya, 69 of which are fossil mammals. The Rudabánya fauna outcrops at no less thanseven different localities, all believed to be essentially synchronous in age. There are no direct radioisotopic dates from Rudabánya, the lowerage is constrained by a single crystal argon date of 11.4 m.y. ± 0.1 m.y., and biochronologic correlations support a latest MN 9 age of ca. 10-9.7 Ma. Paleogeographically, the Rudabánya fauna developed on the edge of the extensive Pannonian lake, which supported a warm, equablesubtropical climate. Zoogeographically, Rudabánya is most closely allied with the late Astaracian (MN8)-early Vallesian (MN9) Spanish verte-brate localities, and particularly Can Llobateres (straddling the MN9/MN10 boundary). These central and western European localities contra-st strikingly with correlative eastern Mediterranean-Southwest Asian localities in their community structure. In particular, Rudabánya and
Palaeontographia Italica 89 2002 3-36 tavv. Pisa, 2004
1 College of Medicine, Department of Anatomy, Howard University, 520 W St. NW, Washington D.C., USA - [email protected] Geological Institute of Hungary, H-1143 Budapest, Stefánia u. 14, Hungary - [email protected] Dipartimento di Scienze della Terra and Museo di Storia Naturale (Sezione Geologia e Paleontologia), Università di Firenze, via G.
La Pira, 4, 50121 Firenze, Italy - [email protected] Institut de Paleontologia M. Crusafont, C/ Escola Industrial 23, 08201-Sabadell, Spain. 5 Department of Palaeontology, Natural History Museum, London, SW7 5BD, U.K.6 Virginia Museum of Natural History, 1001 Douglas Ave., Martinsville, Virginia 24112, USA.7 Department of Anthropology, University of Toronto,Toronto, ON M5S 3G3, Canada8 Department of Anatomy and Histology, Anderson Stuart Building, The University of Sydney, NSW Australia 20069 Department of Ecology, Evolution and Marine Biology, University of California, Santa Barbara, California 93106, USA10 Department of Geology and Paleontology, Naturhistorisches Museum Wien, Burgring 7, A-1014 Vienna, Austria11 Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine, Université de Poitiers, 40 av. du Recteur Pineau, 86022
Poitiers, France.12 Department of Paleontology, Charles University Praha, Albertov 6, CZ-12843 Praha, Czech Republik13 Departmento of Geology, University of Helsinki, P.O. Box 11, FIN-00014, Finland14 Forschungsinstitut Senckenberg, Abteilung Paläanthropologie, Senckenberganlage 25, D-60325 Frankfurt am Main; Germany15 Department of Paleontology, Hungarian Museum of Natural History, Muzeum krt. 16-18, Budapest, Hungary.16 Universitätsinstitut und Staatssammlung für Paläontologie und Historische Geologie, München, Germany17 Rudabánya, Rózsavölgy u. 2, Hungary18 Institut und Museum of Zoology, University of Greifswald, D- 17489 Greifswald, Germany19 Laboratory of Geology and Palaeontology, Aristotle University of Thesaloniki. 54006 Thessaloniki, Greece.20 Department of Paleontology, Eötvös Loránd University, Ludovika tér 2, H-1083 Budapest, Hungary21 Berkeley Geochronology Center, Berkeley, California, USA22 Department of Palaeobiology and Palaeoecology, Geological Institute, Academy of Sciences, Rozvojová 135, Praha, and
Department of Zoology, Charles University, Viničná 7, Praha, Czech Republic23 Laboratoire de Paléontologie, Muséum National d’Histoire Naturelle, UMR 8569 du CNRS, 8 rue Buffon, 75005 Paris, France24 Department of Anthropology, University of Texas at Austin, Austin, Texas 78712-1086, USA25 Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Slawkowska 17, 31-016 Kraków, Poland26 Department of Anthropology, University of Arkansas, Old Main 330, Fayetteville, Arkansas 72701 USA27 Geologisches Institut, Universität Bonn, Nussallee 8, D - 53115 Bonn 28 Faculty of Earth Sciences, Utrecht University, PO 80021, 3508TA Utrecht, The Netherlands 29 Department of Palaeozoology, Swedish Museum of Natural History, Box 50007, S-104 05 Stockholm, Sweden30 Staatliches Museum fhr Naturkunde Stuttgart, Rosenstein 1, D-70191 Stuttgart, Germany.

4 R.L. BERNOR - L. KORDOS - L. ROOK
I) INTRODUCTION(L. Kordos and R.L. Bernor)
Rudabánya is a very old settlement in NEHungary. It has a long mining history extendingback to the Age of Copper (5000-4000 BP). Fromthe end of the 18th Century, several open-air pitsand an underground shaft were excavated in orderto mine iron. During the 1950’ s extensive miningactivity was pursued and uncovered the soft lateMiocene fossiliferous sediments that cover theTriassic iron-bearing basement rock. The Ruda-bánya mining quarry was closed in the middle ofthe 1980’s (Text-fig. 1).
Discovery of the late Miocene lignitic sedimentsat Rudabánya occured in the last century. The firstknown vertebrate find from Rudabánya, “Mastodonlongirostris”, was collected by Gy. Pethö in 1902.Later, Schréter (1929) described more material,
including “Hipparion”, which was recognized asbeing important for the biochronologic determina-tion of the deposits. After some sporadic lateMiocene vertebrate finds from Rudabánya, the firstprimate fossils were found by the mine’s ChiefGeologist, G. Hernyak in the middle of the 1960’s.Hernyak brought these specimens via A. Tasnadi-Kubacska to Professor M. Kretzoi in Budapest(Geological Institute of Hungary). Kretzoi (inAnonymous, 1967) immediately named this speci-men Rudapithecus hungaricus in a Hungarian dailynewspaper, the Magyar Nemzet. This specimen waslater figured for the first time in a popularHungarian science publication (Tasnadi-Kubacska,A., 1967). Later in 1967, Kretzoi gave a lecture onthe Rudabánya specimen at a syposium on humanevolution (Budapest). In 1969, Kretzoi published abrief description of this same specimen, againunder the nomen of Rudapithecus hungaricus.
Can Llobateres supported diverse lineages of small and medium sized mammals that were dominated by more archaic late early and middleMiocene European faunas. Vallesian localities in Greece and Turkey document an entirely different and progressive “proto-Pikermian” mega-fauna rich with advanced carnivore and ungulate assemblages. Of particular note is the sharp increase in hypsodont ungulates seen in theeastern Mediterranean-southwest Asian region. Finally, we briefly utilize these zoogeographic and paleoecological observations to contrastcurrent competing hypotheses on “European versus African” ancestry of the African ape-human clade.
RIASSUNTO – [Lo stato dell’arte sui risultati delle ricerche multidisciplinari a Rudabánya, Miocene superiore (MN9), Ungheria] -Rudabánya è una miniera di lignite i cui depositi, investigati per la prima volta da Pethö nel 1902, hanno restituito una associazione fauni-stica a vertebrati del Miocene superiore. La prima segnalazione di un primate nell’associazione di Rudabánya si deve a Gabor Hernyák, geo-logo responsabile della miniera. La notorietà di Rudabánya è invece legata al nome del Prof. Miklos Kretzoi, il quale nel 1969 istituì la nuovaspecie di primate ominoide, Rudapithecus hungaricus (qui riconosciuto come sinonimo di Dryopithecus brancoi).Un importante progettodi ricerca internazionale su Rudabánya, che ha visto nuovi scavi ed uno studio dettagliato della associazione faunistica, è iniziato nel 1991,coordinato da L. Kordos e R.L. Bernor. Il presente lavoro (cui contribuiscono 36 autori) rappresenta una prima sintesi di questi undici annidi ricerche, e sarà seguito dalla realizzazione di un volume monografico. L’associazione faunistica di Rudabánya è costituita da 112 specie divertebrati fossili, delle quali 69 sono mammiferi. La fauna proviene da almeno sette differenti affioramenti, tutti nell’area della miniera, etutti correlabili ad uno stesso intervallo cronologico. Pur non essendo possibile effettuare una datazione radiometrica diretta per i sedimentifossiliferi di Rudabánya è tuttavia disponibile, da una vulcanite sottostante i livelli fossiliferi, una età radiometrica (ottenuta con il metodoAr/Ar su cristallo singolo) di 11.4 m.y. ± 0.1 my. che costituisce un vincolo post quem. Questa data è in accordo con la caratterizzazione bio-cronologica della fauna indicante una età a mammiferi tardo MN 9 (circa 10-9.7 Ma). Da un punto di vista paleogeografico Rudabánya sicolloca sulle sponde settentrionali dell’esteso lago Pannonico, in un momento caratterizzato da un clima caldo e subtropicale. Da un punto divista zoogeografico, la fauna di Rudabánya è più vicina alle località Spagnole dell’ Astaraciano superiore (MN8) o del Vallesiano inferiore(MN9), ed in particolar modo alla fauna di Can Llobateres (che si colloca al passaggio tra MN9 e MN10). Si osserva un forte contrasto nellastruttura delle comunità che caratterizzano questo complesso di località dell’Europea centrale ed occidentale, in rapporto alle località coevedel Mediterraneo orientale e del vicino oriente. In particolare, sia a Rudabánya che a Can Llobateres si mantengono varie linee di mammiferidi piccola e media taglia che sono state dominanti nelle associazioni Europee del Miocene inferiore finale e medio. Le località Vallesiane dellaGrecia e della Turchia registrano invece associazioni faunistiche completamente diverse, con faune a grandi mammiferi di tipo “proto-Pikermiano” caratterizzate dalla presenza di forme nuove soprattutto per quanto rigarda carnivori ed ungulati. Di particolare importanza è ilforte incremento nella percentuale di ungulati ipsodonti che si osserva nella associazioni del Mediterraneo orientale e del Sud-Est asiatico.Nella discussione finale vengono utilizzate le evidenze zoogeografiche e paleoecologiche per confrontare le ipotesi oggi in discussione riguardoall’origine “Europea” vs. “Africana” del clado comprendente Antropomorfe africane e uomo.
Text-fig. 1 - Location map of Rudabánya in Hungary; the small box indicates the location of Rudabánya within a paleogeographic sketchof the Pannonian basin (modified from Kordos, 1982), and view of the old lignite mine.

5RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
Later in 1969, Hernyak found yet another Ruda-pithecus mandible from the same locality (Locality1), and the Geological Institute of Hungary quicklyorganized and initiated an excavation at the locali-ty.
Regular excavations were organized by Kretzoiat Locality 1 (“Mastodon Wall”) in 1971, and a sec-ond locality (Locality 2 or “Gibraltar”) was discov-ered in 1972. Locality 1 was mostly lost by slopecollapse in 1973, but the excavation team collectedfurther vertebrate and plant fossils from Locality 3(“The Great Wall”) in 1974. During the last threeyears of field campaign led by Kretzoi, a number ofsignificant accomplishments were realized. In 1976a protective roof was built over Locality 2. Furtherfossil material was collected at this locality andlocalities 4, 5, 6 and 7 were discovered. In 1977,Rudabánya localities 1 and 2 were declared a nat-ural protected area by the Hungarian government.In 1978, Kretzoi collected the last 8 of a total of 74primate remains found up to that point-in-time.
Kretzoi (1974), listed three primate taxa fromRudabánya. In addition to Rudapithecus hungari-cus, two new taxa, Pliopithecus (Anapithecus)hernyaki, and Bodvapithecus altipalatus were recog-nized by Kretzoi in the Rudabánya primate fauna.Finally, Kretzoi (1984) described a new subgenusand species from Rudabánya, Rangwapithecus(Ataxopithecus) serus based on the RUD-71 speci-men. Later in the same paper, he recognized a sec-ond new taxon from Rudabánya, described only asHominoidea indet., based on a single lower molar(RUD-19). In 1976, Kretzoi concluded thatRudapithecus’ phylogenetic relationships were asfollows: “the australopithecines are representing aseparate evolutionary line from the AsianRudapithecus-Pithecanthropus-Homo lineage”.
Kretzoi (1975) and Kretzoi et al. (1976) pub-lished the first results on Rudabánya pollen,macroflora, ostracods, molluscs and vertebratesfrom all localities collected between 1967 and1974. These studies concluded that the age of allRudabánya localities was “Lower Pliocene” (= nowearly late Miocene), and established a new terres-trial mammal age, the “Bodvaium”, intermediatebetween the older “Monacium” and younger“Eppelsheimium”, which is correlated with MN9Kordos (1987b). Between 1979 and 1984, shortexcavation seasons were led by L. Kordos. In 1985Kordos wrote a popular science book on theRudabánya localities. In 1984, Kretzoi placedhominoid materials collected under his direction inthe National Museum of Hungary. These collec-tions remain closed to all other investigators. Onlycasts made by Alan Mann (University of Penn-sylvania, Philadelphia) are currently available forstudy. At the same time, oversight of theRudabánya localities was transferred from thelocal museum to the Geological Institute ofHungary (Budapest) where they are currentlycurated and conserved.
Gabor Hernyák discovered an important femaleskull, RUD-77, at locality 2 in 1985. This was onlythe fourth Miocene hominoid skull known at thetime, the others being the early Miocene Proconsulafricanus skull (Kenya), the late MioceneOreopithecus skull (Baccinello, Italy), the late
Miocene Sivapithecus skull (Pakistan) and the lateMiocene Lufungpithecus skull (China). TheRudabánya skull was initially described by Kordos(1987a), and later revised by Kordos & Begun(1997). Kordos continued excavation at Localities 2and 3 between 1986 and 1989. At Locality 2, theexcavation was mostly in the gray marl, which wasfound to underlie the upper lignite, black clay andblack mud where Anapithecus was found.Anapithecus material found included molar teeth,and in 1988 a partial cranium, RUD-83. At Locality3 more lignite and conglomerate layers were exca-vated, and mostly large mammal bones werefound. Kordos shifted his emphasis at Rudabánya,by concentrating his excavations at Localities 2and 7 from 1989 through 1991. During this period,Kordos discovered that the Locality 7 paleokarsthorizon was deposited on the Rudabánya paleoval-ley floor, and that the lignite deposits were deposit-ed on top of the paleokarst. This period of the exca-vation witnessed major socio-political changes inHungary, and difficulty in accessing funds fromlocal sources.
In the summer of 1991, Kordos invited R.L.Bernor (Washington D.C.) to engage in a multidis-ciplinary research project at Rudabánya Locality 2.With funding from the L.S.B. Leakey Foundationand the National Geographic Society, excavationswere continued by Kordos and Bernor, with M.Armour-Chelu (London) pursuing collection oftaphonomic data both in the field, and the muse-um in 1992. Peter Andrews (London) led the tapho-nomic excavations at Rudabánya 2 in 1993 and1994, while Kordos and Bernor focussed theirefforts on the systematics of the Rudabánya verte-brate faunas. This review has been undertaken incollaboration with a number of specialists whosecontributions follow in the chapters that follow.While the emphasis is on all of the RudabányaLocality 2 fauna, several specialists looked at thevertebrate remains from all the Rudabánya locali-ties as well as the late Miocene Hungarian faunalsuccession (re: Bernor et al., 1999). An edited vol-ume with detailed descriptions of the fauna, itspaleoecologic and taphonomic aspects will followthis contribution in the near future (Bernor &Kordos, Eds.).
Alongside these efforts, L. Kordos and D.R.Begun have made a focussed analysis of theRudabánya primate fauna. In addition, Begun andKordos have conducted three summer field seasons(1997-1999) under the aegis of the “RudabányaField School” offered to college students by theUniversity of Toronto and the Geological Instituteof Hungary.
II) GEOLOGICAL BACKGROUND
GEOLOGY (L. Kordos and G. Hernyak)
The Rudabánya hominoid localities are locatedon the western flank of the northern Carpathianmountains. Locally, the exposed basement rockincludes the Slovakian Karst, the Aggtelek Karst,and the Rudabánya Hill Range. From a structuraland historical viewpoint, the Slovakian and the

6 R.L. BERNOR - L. KORDOS - L. ROOK
Aggtelek Karst separated from the NW Carpathianrange prior to the Oligocene. The Rudabánya HillRange separated from the other two formationsduring the Oligo-Miocene, by the NE directioncompression of the neighbouring Bûkk andSzendrõ Mountains. By this event, the RudabányaHill moved from the southern foreland to the east-ern margin of the Aggtelek Karst.
The tectonic and sedimentologic history of thearea has been extensively reevaluated during thelast two decades (Balogh, 1982; Grill, 1989; Grill etal., 1984; Szentpétery, 1988; Less, 1998). The origi-nal Mesozoic sediments were mostly calcareous,deposited in three different environmental faciesduring the Triassic and Jurassic periods, namelyfrom N to S: Szilice, Melléte and Torna facies. TheSzilice and Torna facies were deposited on the con-tinental plate, while the Melléte facies was deposit-ed in the mobile continental self of the TethysVardar Ocean. The Szilice facies is Upper Permianto Late Jurassic age. These formations formed theSzilice Nappe system as their uppermost structuralunit. The Melléte facies was deposited during themiddle - late Triassic, and early Jurassic.Deposition was followed by regional metamor-phism. The Torna facies contains only Triassic sedi-ments with later epizonal regional metamorphosis.
The oldest exposed sediments in the Rudabányaregion are of latest Permian age. The underlyingmiddle Permian landscape was covered by evapor-ites during the late Permian and early Triassic. Atthe beginning of the early Triassic, there is a majortransgression, with three developed facies. Themiddle Triassic carbonate-platform type limestonewas divided during the Ladinian into three units:the Szilice facies deposited on the northern conti-nental base, the Melléte facies deposited on thethin continental and oceanic plates, the Tornafacies deposited on the continental base to thesouth. Because the subsidence of this area contin-ued during the middle and late Triassic and in theearly Jurassic, sediments were deposited in a deep-sea environment. Subduction of the Melléte faciescommenced in the middle Jurassic, and formed aseries of nappe systems. Due to subduction, mostof the sediments of the Melléte facies melted andformed granitic and rhyolitic magmatic rocks. Thisvolcanic activity is responsible for the metasomatictype of mineralization typical of the Szilice faciessediments in the Rudabánya Hill Range. The origi-nal nappe system of the Aggtelek-Rudabánya rangewas strongly deformed and reduced in space afterthis event. During the Cretaceous, sediments of thethree nappe systems again became deformed.
From the second half of the Cretaceous, theMesozoic series became elevated and formed akarstic peneplain, and until the end of theOligocene, produced bauxite type soils. The thirdand latest important tectonic event was during thelatest Oligocene and earliest Miocene when a hori-zontal fault produced the compressed range knowntoday as the Rudabánya Hills. This tectonic eventwas part of a larger tectonic mobilism thatinvolved the Bûkk and Szendrõ Mountains, them-selves part of an ancient microplate, which movedin a southwest to northeast trend to their presentlocation due to encroachment by the African Plate.
Late Miocene Paleogeography of Rudabánya(L. Kordos)
The Rudabánya Range was formed in the mid-dle Miocene as a peninsula of the Aggtelek Karst(Kordos, 1982). The late Miocene Pannonian Basindeveloped in an extensional basin behind the com-pressional arch of the Carpathians (Text-fig. 1).Subsidence began during the middle Miocene,forming deep, pelagic basins, separated by reef-bearing ridges. A series of smaller or greaterdepressions are associated with the Alpine chainalong the southern margin of the European Plate,and the largest of these is the Pannonian Basin,which is surrounded by the Carpathians, Alps andDinarids. The uplift of the Carpathian Mountainsbetween 12-11 Ma ago established the PannonianLake system, separating it from the EasternParatethys. The Pannonian lake was completelyfilled in by the terminal Miocene or earliestPliocene, ca. 5 Ma.
Magyar et al. (1999) have recently reconstructedthe paleogeographic evolution of the Pannonianlake recognizing three distinct intervals of develop-ment: an initial, regressive stage that resulted in itsisolation from the Eastern Paratethys; a secondinterval of gradual transgression, and a long thirdinterval of protracted regression, sedimentaryinfilling and progradation.
During the early Sarmatian, the RudabányaRange was 7-8 km in length and 1-2 km wide. Itwas connected to the Aggtelek Karst at its north-ernmost limit. At this time, it was likely covered bySarmatian marine sediments. The Pannonian lakefirst formed circa 12.0 Ma, a time marked by theonset of a regression. This relative sea level dropresulted in isolation of the intra-Carpathian waters(Magyar et al., 1999). Unconformities between themarine Sarmatian and overlying Pannonian lacus-trine environments are common, and especially inthe Borsod Basin adjacent to the Rudabányapeninsula. At this time, the top of the RudabányaRange was eroded by fluvial activity, and tropical-subtropical karst surfaces formed on the base andon the slopes of the valleys. At the base of the inter-peninsular valley-system, terrestrial variegated claywas deposited without any apparent fossil remains.After the separation of the Pannonian lake, thewater table gradually increased, and circa 10.8 Mapenetrated the northern part of the Borsod Basin,flooding the Rudabánya Range and adjacent val-leys. Between 11 and 10 Ma, the valley system wasexposed to a variety of lacustrine and peri-lacus-trine environments including: lake, swamp, wet-land and paleosoil. Rudabánya’s sediments includecyclic layers of gray marl, clay and lignite, totalling8-12 meters in thickness. These sediments, includerich fossiliferous levels that contain plant impres-sions, seeds, molluscs, vertebrate remains and, ofespecial note, catarrhine primates.
The Pannonian lake probably reached its great-est areal extension circa 10-9.7 Ma. The maximumtransgression is likely marked by the uppermostelevation of the lacustrine-swampy sedimentationin the valleys of Rudabánya Peninsula. One oscilla-tion of the water level probably precipitated theresedimentation of Locality 7. Because this sedi-ment was hydraulically resorted, the bone matrix is

7RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
rich and very broken. After its resorting, anothertransgressive lake cycle innundated and buried theLocality 7 bone breccia. According Magyar et al.’s(1999) reconstruction at the ca. 9.0 Ma Congeriapraerhomboidea Biochron, the Pannonian lake sud-denly reduced its areal extent and the lacustrine-swampy sedimentation regime ceased to occur.These sediment types were replaced by fluviatilesands which appear at the top of the RudabányaRange succession covering all local depressionsand forming a plateau that is elevated above thelake sediments.
TAPHONOMY(P. Andrews, M. Armour-Chelu, D. Cameron)
The fossil assemblages from seven stratigraphiclevels at Rudabánya 2 will be described on thebasis of two collections of material. The main col-lection consists of fossils excavated in place (Text-fig. 2). These are analysed for vertical and horizon-tal distribution relative to sedimentological differ-ences, examining their size distribution, directionsof preferred orientation, angles of dip, bone break-age patterns and surface modifications.
The second collection consists of the screening
residues. The fine residues from the screeningprocess have only been sorted in part, but thecoarse residues retained in the 0.8 mm screenshave been sorted into the following constituents:wood, seeds, root, molluscs, large mammal bone,small mammal bone and amphibians. The relativeabundances and distributions of these fractionshave been analysed relative to the meter grid laiddown over the site.
Results indicate several modes of animal boneaccumulation at Rudabánya. Tree trunks and rootsare present in their growth positions in lignites,black muds and massive marls, and the fossil ani-mal bones present show a lack of disturbance.Water transport is indicated for reworked marls,with the animal bones showing evidence of pre-ferred orientation in the direction of water flow.The accumulation of shelly layers and small mam-mal assemblages is associated with pond depositsaccumulated on lake-shore flats. Little evidence ofpredation has been found in any level (neverthelesssee Armour-Chelu & Viranta, 2000), but there isextensive post-depositional modification of the fos-sils from most levels due to highly acid environ-ments. The exception to this is the assemblagefrom the pond deposits (the “black mud”), whichalthough heavily blackened, shows little other evi-dence of post-depositional modification.
III) INVERTEBRATE FAUNAS
TERRESTRIAL AND FRESHWATER MOLLUSCS(E. Krolopp)
The molluscan work reported herein is based onshells collected at Rudabánya during the earlyexcavation years. The mollusc collections are allhoused by the Museum of the HungarianGeological Institute. Pantó (1956: 271) has report-ed 15 mollusc species in his monograph on thegeology of Rudabánya, but his specimens havesince been lost.
The Rudabánya mollusc fauna consists of 44taxa (Table 1). The majority of the species are ter-restrial, but the specimen number of aquatic formsis significantly higher than that of the terrestrialones.
Both bivalve species diversity as well as numberof individuals for those species is very low in theaquatic fauna. Unionids are completely lacking.Given that larvae of these animals live in the skinof fish species for a time as parasites, their absencemay be connected to the absence of fish in theenvironment. The mud was rich in organic materi-al, thus depleted of oxygen, and this may be thereason for the rarity and few specimen of the smallsized Sphaeriidae. All sphaeriid specimens origi-nate from a yellowish clay interdigitated betweentwo lignitic horizons. The low organic content ofthis layer is suggestive of a well aereated environ-ment. Similarly, the stagnant water and low oxygenlevel in other Rudabánya environments mayexplain the low number of Prosobranchiataspecies. Most specimens of this group originatealso from the above mentioned yellowish clay.
The number of species and specimens ofText-fig. 2 - The excavations at Rudabanya site II. Differentviews of 1993 excavation.

8 R.L. BERNOR - L. KORDOS - L. ROOK
Basommatophora is significantly higher than thatof the prosobranchiates. Members of this grouplive mostly on seaweed and may repeatedly climbup to the water surface to breathe atmospheric air.Consequently, they are only moderately influencedby the decomposing organic rich mud and by theoxygen depleted water above it.
The most abundant forms of terrestrial gas-tropods lived on plants that set their roots in water,or on trees adjacent to the shoreline. These formsall require high humidity and live near waterbodies(Carychium and Succinea species). Carychiumspecies are especially abundant. Besides these,Zonitidae, Limacidae and Helicidae, which alsorequire an elevated level of humidity, lived fartherfrom the shores, on areas of open vegetation or ongently overgrown ones (wet meadows and forests).Some forms, having modern relatives living ondrier land, also were found (Pupillacea).
IV) VERTEBRATE FAUNA
AMPHIBIANS (Z. Roček)
Rudabánya’s amphibian fauna includes taxathat were common in Europe from pre-Oligocenetimes (such as Chelotriton , Discoglossidae,Pelbatidae, if the forms recorded are Eopelobates,Palaeobatrachus, Peldytes and Rana). These taxaalso include forms that first appeared after theGrande Coupure (Eocene/Oligocene boundary) andbecame widely distributed throughout Europe dur-ing the Neogene (e.g. Latonia, Discoglossus), whileothers appeared only in the early Miocene andwere widely distributed in eastern Europe, rarelyreaching the western region of the continent (e.g.Mioproteus).
Tailed amphibians are represented byMioproteus caucasicus (Family Proteidae).Mioproteus is the most abundant tailed amphibianat Rudabánya, and this is quite unusual if com-pared with its other known occurrences. Salaman-dridae are represented by two types of Chelotritondifferent in their morphology; both are morpholog-ically similar to the late Oligocene articulatedskeletons of Chelotriton from Rott and Enspel(Germany), but the Rudabánya forms differ insome significant anatomical features (e.g. mor-phology of the maxilla). Since these same morpho-logical differences are seen in the Chelotriton fromGritsev (late Miocene, Ukraine [MN9]), they arebelieved to be of taxonomic importance. AnotherRudabánya salamandrid is represented by frag-mentary maxillae and praemaxillae recalling extantSalamandra, but larger, having tooth crowns withblack tips and processes between tooth bases. Thisform was not yet formally described, and the sameform has also been found at Gritsev. Tiny vertebraemay suggest the presence of still another salaman-drid belonging to the Brachycormus-Triturus com-plex. However, precise taxonomic assignment ofthis form can only be made on cranial elementsthat are not normally found because these bonesbreak during screen washing. Vertebrae and otherpostcranial elements are relatively uniform in thisgroup.
The most common Rudabánya anurans werediscoglossids belonging to the genus Latonia (L.gigantea). In addition, there are several elementsthat also suggest the presence of Bombin, as well asDiscoglossus. Another dominant anuran was apalaeobatrachid, here assigned only to Palaeoba-trachus, although extreme differences in size, asso-ciated with some morphological differences, mayindicate more than a single species. Rudabánya isone of few fossil sites in which Latonia is foundtogether with palaeobatrachids. Besides Latoniaand Palaeobatrachus which were dominant, therewere also Pelobatidae (difficult to distinguish onthe postcranial material only whether Eopelobatesor Pelobates), Pelodytes, Hyla and Rana. Ruda-bánya’s Hyla appears to be the earliest record ofthis genus in Europe.
Size differences of ranid ilia may suggest thatthere were at least two forms of Rana atRudabánya. A rather surprising aspect of theRudabánya herpetofauna is the poor fossil repre-
TABLE 1 - Rudabánya Terrestrial and Freshwater MolluscFaunal List
Sphaerium aff. corneum (L.)Pisidium sp. indet.Pomatias sp. indet.Acicula cf. edlaueri SchlikumBithinia cf. jurinaci BrusinaViviparus sp. indet.Brotia escheri BrongniartCarychiopsis berthae (Halaváts)Carychium sp. indet.Lymnaea cf. palustriformis (Gottsch.)Lymnaea cf. socialis (Gottsch.)Planorbarius aff. corneus (L.)Anisus confusus Soós Anisus cf. septemgyratiformis (Gottsch.)Segmentina cf. filocincta (Sandberger)Succinea sp. indet.Granaria cf. suebleri (Klein)Vertigo callosa (Reuss)Vertigo angustior oecsensis (Halaváts)Vertigo cf. moedlingensis Wenz & EdlauerVertigo sp. indet.Argna oppoliensis (Andrusow)Gastrocopta acuminata (Klein)Gastrocopta fissidens infrapontica WenzGastrocopta cf. nouletiana (Dupuy)Gastrocopta edlaueri (Wenz)Gastrocopta aff. farcimen (Sandberger)Azeca cf. tridentiformis (Gottschieck)Acanthinula cf. anthonini (Michelin)Vallonia sp. indetStrobilops sp. indet.Helicodiscus sp. indet.Discus cf. pleuradrus (Bourg.)Tryptichia sp. indet.Nordsieckia fischeri pontica LuegerSerrulella sp. indet.Vitrea sp. indet.Zonitidae indet.Limacidae indet. ILimacidae indet. IITrichia sp. indetHelicigona sp. indetTropidomphalus cf. doderleini (Brusina)

9RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
sentation of the pelobatid frogs, known only fromseveral ilia, and absence of the Bufonidae,although the earliest record of Bufo in Europe isfrom MN4 of Spain.
SNAKES (Z. Szyndlar)
The fossil snake material from Rudabányadescribed in this short report consists of nearly500, mostly badly damaged, vertebrae belonging tofour or five ophidian taxa. All but one of these ver-tebrae represent the trunk portion of the column.
A single vertebra, characterized by very smallabsolute dimensions (the centrum length less than2 mm) and simplified morphology (lacking neuralspine and haemal keel, with undivided paradi-apophyses), is clearly referable to the InfraorderScolecophidia. The scolecophidian from Ruda-bánya is tentatively considered to be a member ofthe genus Typhlops, the only representative of thefamily Typhlopidae known in Europe and adjacentareas. It should be qualified here however that theidentification of scolecophidian vertebrae is unre-alizable even to the familial level. On the otherhand, a possible presence of the other scolecophid-ian family Leptotyphlopidae, with the genusLeptotyphlops being found today very distant fromCentral Europe, is not highly likely.
There are forty-nine vertebrae that are clearlyreferable to natricine colubrids. These vertebraeare all characterized as having sigmoid shapedhypapophyses throughout the trunk portion of thecolumn, long centra and with parapophysealprocesses projecting anteriorly. Projecting struc-tures are missing in most vertebrae. However, rela-tively high neural spines retained in several of thebones suggest a referral to the extant genus Natrix.
The informal term “Colubrinae” refers to colu-brids devoid of hypapophyses in most of theirtrunk vertebrae. An overwhelming portion of theRudabánya snake material, i.e. approximately 200vertebrae, is referred to a single specificallyunidentified form, named here Colubrinae A. Thevertebrae belonged to relatively large snakes, thecentrum length of the largest one being 6.8 mm.This snake most likely belonged to the living genusColuber, although not resembling extant Europeanmembers of the genus. In some aspects theRudabánya colubrine trunk vertebrae are similarto those of the extinct species C. hungaris fromPolgárdi (re: Venczel, 1994). Six vertebrae, arereferred to a different Colubrinae (Colubrinae B).They are minute and elongate, having strongly flat-tened haemal keels. It cannot be excluded that theyactually represent the posteriormost trunk portionof the column of Colubrinae A. Approximately 200vertebral fragments were too greatly damaged to bedetermined with any degree of accuracy. They aremost likely referable to Colubrinae A.
Fossil remains of the Family Colubridae (s.l.),the most common and diverse snake group inEurope since at least the middle Miocene, are notvery informative unless they can be identified tothe specific level. This is also the case forRudabánya’s colubrid ophidians. Perhaps theRudabánya Colubridae included the same or simi-
lar forms known from other late Miocene localitiesin the region: Coluber hungaricus and Natrix lon-givertebrata (cf. Bachmayer & Szyndlar, 1985;Szyndlar & Zerova, 1992; Venczel, 1994).
The occurrence of Naja romani (FamilyElapidae) was recognized mainly on the basis of asingle, but perfectly preserved relatively large verte-brae; the centrum length is 7.8 mm. Vertebrae ofthe genus Naja can be easily differentiated fromthose of other snakes. They represent a morpholog-ical pattern observed in large colubrines but, con-trary to the latter, they are provided with hypa-pophyses throughout the trunk portion of the col-umn. The Rudabánya vertebra displays a set of fea-tures characteristic of Naja romani (cf. Szyndlar,1991b), the only extinct cobra known from the lateMiocene of Central Europe (Szyndlar & Zerova,1990). The referral of four other vertebral frag-ments to Naja cannot be fully corroborated.
As in most coeval localities in Europe, theRudabánya fauna contained remains of two mem-bers of the genus Vipera; the poor state of preserva-tion of these fossils, however, does not enable theiridentification to the specific level. Members of theViperidae can be easily distinguished from othersnakes bearing hypapophyses throughout their ver-tebral column (natricine colubrids and elapids) onthe basis of posteriorly depressed neural arches,ventrally directed parapophyseal processes, andstraight hypapophyses. Three viperid fragmentarytrunk vertebrae found in Rudabánya are referred toVipera sp. (“aspis” complex) based largely on theirrelatively small size. A single fragmentary trunkvertebra of Vipera, despite its strong damage, isclearly referable to Vipera sp. (“Oriental viper”group), based on its relatively large absolutedimensions.
The most characteristic elements of the ophidi-an assemblage from Rudabánya are the scole-cophidian Typhlops and the elaphid Naja. The pre-sent European range of the genus Typhlops isrestricted to the Balkan Peninsula. The occurrenceof these minute burrowing snakes, present in theregion since at least the early Miocene (Szyndlar,1991a), was also reported from the Ukrainian lateMiocene (Szyndlar & Zerova, 1990, 1992). Beforethe end of the Miocene however, scolecophidiansdisappeared from the area of Central Europe. Thefossil elapid from Rudabánya was classified as theextinct species N. romani. Perfectly preservedremains of this fossil cobra are known from severalEuropean countries from the period between theearly (MN4) through late (MN11) Miocene(Szyndlar & Rage, 1990). In the vicinity ofRudabánya it was reported from Kohfidisch andGritsev (Bachmayer & Szyndlar, 1985; Szyndlar &Zerova, 1990). As did Typhlops, Naja also disap-peared from the Central Paratethys region beforethe end of the Miocene.
The European fossil record suggests that thegeographic range of several snake taxa diminishedduring the latest phase of the Miocene. It seemsthat even before the Messinian Crisis the Europeandistribution of at least Typhlops, Eryx (Boidae), Najaand large Vipera (“Oriental viper” group, Viperidae)became restricted to the peri-Mediterranean coast-line, and to a lesser degree, the Black Sea. Then,

10 R.L. BERNOR - L. KORDOS - L. ROOK
during the course of the Pliocene, all of these taxadisappeared entirely from the westernMediterranean. The extinction of the scolecophidi-an and elapid snakes in Hungary prior to the end ofthe Miocene confirms this supposition. During theterminal Miocene (MN 13), the snake fauna charac-teristic of Rudabánya was replaced in Hungary bythe extant assemblage which was in place by thetime represented by Polgárdi (Venczel, 1994).
AVES (D. Janossy)
The systematic excavations at Rudabánya haveyielded bone fragments representing 12 birdspecies. This material is particularly valuablebecause Rudabánya’s age has been established asbeing late Miocene (MN 9; re: Correlation below). Itis valuable to compare the Rudabánya ornithofau-na with the recently analysed one from Polgárdi(MN 13). The Rudabánya galliforms are completelydifferent from galliforms at Polgárdi. Also,Polgárdi’s only owl is the Barn Owl (Tyto), whilethat at Rudabánya is Strix.
I provide here a short evaluation of both older(reported by Jánossy, 1976, 1977 and by Kretzoi inKretzoi et al., 1976) and newer material collected atRudabánya. Although a great deal of theRudabánya avian material is too fragmentary toidentify beyond the incertae sedis level, severaltaxa can be identified within the avian assemblage.
Family: Anatidae Milne-Edwards (1867) described Anas velox
from the middle Miocene (MN 6) French locality ofSansan. In his revision of the group, Cheneval(1987) mentioned this small form from Germany(Steinheim am Albuch), Bohemia (Dolnice) andRoumanian Dobruja (Credinta). In spite of the factthat the Rudabánya material is fragmentary and ofuncertain determination, it seems distinctly possi-ble that this Anas is well referred to A. aff. velox.There likely is also a larger duck of the genus Anasat Rudabánya; because of the extremely fragmen-tary condition of the remains the determinationcannot be more accurate than Anas sp. (size ofacuta-querquedula).
Family: PhasianidaeBochenszi (1987) revised Miophasianus medius
from Przeworno (Peniborn), which is the same sizeas the Rudabánya form. However, Prezeworno ismuch younger in age (MN 17) and I thus prefer tomaintain Miophasianus cf. medius for theRudabánya pheasant. Palaeortyx aff. grivensis is asmall Galliform bird known from La Grive(France) and Gargano (Italy) (Ballman, 1969a,1976), Rudabánya, Tardosbánya and Sûmeg(Hungary) and Malusteni (Roumania) (Kessler,1984; Jánossy, 1991). Given that the relevantRudabánya material is so fragmentary, I am onlyable to establish that there is another pheasanttaxon intermediate in size between the smallerPalaeortyx aff. grivensis and the larger Miopha-sianus. This intermediate sized taxon is approxi-mately the size of Palaeortyx phasianoides and/orPalaeortyx intermedius.
Family: RallidaeMilne-Edwards (1871) described Miorallus
major from the middle Miocene (MN 6) locality ofSansan. The Rudabánya form agrees well with thistaxon in its size and according to Lambrecht(1933) it agrees in size with Fulica atra. Ballmann(1969b) has noted the occurrence of a rail,“Rallidarum gen. et. sp. indet.” from La Grive (MN7+8) which is chronologically closer in age toRudabánya than is Sansan. He claims that thistaxon is a rail the size of Gallinula chloropus. Giventhe fragmentary nature of the relevant material, lit-tle more can be said about this rail, whose attribu-tion is maintained as ?Miorallus sp..
Family: StrigidaeBallmann (1969a) described a tawny owl, Strix
intermedia, from Wintershof-West (MN3) which ismorphologically quite similar to the extant tawnyowl , Strix aluco. The Rudabánya strigiform bonefragments are referable to Strix aff. brevis (Jánossy,1977). The tarsometatarsus and the length of awell-preserved phalanx 2 digiti 2 support thisassignment. There is yet a smaller owl atRudabánya which I refer to Athene sp.
Family: SylviidaeThere is a Passeriforme bird (Acrocephalus sp.)
at Rudabánya that is the size category and has themorphology of the Great Reed Warbler, Acro-cephalus arundinaceus. Of particular morphologi-cal importance is the Rudabánya taxon’s processusextensiorus (proc. metacarpalis I) and the trochleacarpalis, as well as the reduction of the processusalularis. The morphology is not sufficient for a spe-cific determination. The Rudabánya materialincludes an acrocoracoideum that agrees mostclosely in its morphology to those of the grasshop-per warblers and its allies (Genus Locustella). TheRudabánya material appears to be smaller than thesavis grasshopper warbler (Locustrelle luscin-ioides).
Family: CorvidaeThe presence of corvids at Rudabánya is sup-
ported by a suite of terminal phalanges (claws).The claws in question are characterized as beingthe size of corvids. They also have grooves on theside of the claws that have the length and flatnessseen in corvids. Cranes and geese have similargrooves, but the shape of their bones is much morerobust. Yet, the Rudabánya material is too frag-mentary to identify this taxon beyond the nomenCorvus sp.
Kretzoi (in Kretzoi et al., 1976) characterizedthe Rudabánya avifauna as being indicative of aswamp-forest environment. The modest song-birdfauna composition suggests an environment cov-ered with reeds. A comparison with the avifauna ofPolgárdi (MN13; Jánossy, 1991) exhibits a numberof similarities. The most readily comparable formsare the galliforms, including the small galliform,Palaeortyx cf. grivensis , the intermediate sized P.aff. phasianoides and the somewhat largerMiophasianus cf. medius. Polgárdi has similarsmall and large species. However, the Rudabánya

11RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
and Polgárdi owl faunas are completely differentfrom one another: Strix at Rudabánya versus Tytoat Polgárdi.
INSECTIVORA (R. Ziegler and L. Meszaros)
The Rudabánya insectivore fauna includesspecies belonging to the families Erinaceidae,Metacodontidae, Soricidae, Dimylidae andTalpidae.
The erinaceids include three species: Lantha-notherium sanmigueli, Galerix sp. and Postpa-lerinaceus sp. The remains referred to L. sanmiguelicompare closely in their morphology to the typemandible of Viladecaballs and the more informa-tive specimen of Can Llobateres, the referencelocality of MN 9, with which the Rudabánya faunacorrelates. They also are of the same size as theteeth from Montredon, referred to L. sanmigueli.The remains of L. cf. sanmigueli from Dorn-Dürkheim and Eichkogel roughly fit the presentones in their size. A small Galerix species is repre-sented by an M3 trigonid. The referral of this speci-men to Schizogalerix cannot be excluded. Medium-sized erinaceine dental remains can be referred toa new species of Postpalerinaceus, somewhat small-er and differing in some morphological detailsfrom the type species Postpalerinaceus vireti.
Family Metacodontidae is represented by dentalremains referred to Plesiosorex. These comparemost closely to Plesiosorex styriacus. Given themetric deviation from the type material it will bereferred to Plesiosorex aff. styriacus. The Ruda-bánya Plesiosorex is the latest occurrence of thegenus known to date.
Including four taxa, the soricids are the mostdiverse insectivore family in the Rudabánya fauna.The Dinosorex remains are, in overall size, robust-ness of the mandibular corpus, and in some otherfeatures, indicative of a close relationship toDinosorex pachygnathus, the youngest knownspecies of the genus. Some conspicuous morpho-logical differences make the description of a newspecies necessary. Most of the Rudabánya soricidsare referable to Crusafontina. They are closelyrelated to Crusafontina endemica from CanLlobateres, representing an early member of theevolutionary lineage Rudabánya - Can Llobateres -Kohfidisch. The specimens are referred to Cru-safontina aff. endemica. They are the biostrati-graphically most important soricid fromRudabánya. One tiny mandibular fragment withthree molars in situ is referred to Paenelimnoecusaff. repenningi, in large part because of its biostrati-graphic and geographic proximity to the type local-ity of Kohfidisch, Austria. Paenelimnoecus crouzelifrom Sansan is markedly earlier in time and small-er in size but would fit well morphologically. Onelower jaw fragment bearing M
1can only be allocat-
ed to Soricinae gen. et sp. indet. due to the lack ofa sufficient number of diagnostic characters.
There are also some fossil remains of the familyDimylidae. There are dental remains referable toMetacordylodon schlosseri from Opole that com-pare well in their degree of amblyodonty and
exoedaenodonty, in the strong fusion of the trigo-nid cusps of M
1, and in the reduction of the M
2talonid. However, because of some differenceswith the type specimen, we refer the Rudabányaremains to Metacordylodon aff. schlosseri. Theyrepresent the latest occurrence of this species cur-rently known. The teeth and mandibular frag-ments referred to Plesiodimylus chantrei compareclosely with Pl. chantrei from the type locality LaGrive. This species is known for its long biostrati-graphic range (MN 3/4 to MN 11). A mandibularfragment with an open symphysis is referred toDimylidae gen. et sp. indet. Perhaps it representsan atavistic element in the Metacordylodon sample.Together with Crusafontina, the talpid remainsreferred to a new species of “Archaeodesmana”,represent the dominant insectivores in theRudabánya fauna. However, the most diagnosticelement, the I1 with a bifid apex, is not preserved,allowing no more precise referral than genusArchaeodesmana. Talpa sp. is represented by somehumeri and a jaw fragment. The humeri differ insize and/or morphology from known Talpa speciesthat are either slightly older or younger than theRudabánya sample.
The diverse insectivore fauna indicates a pale-oenvironment that had abundant water andforests. The galericines, represented by numerousLanthanotherium and by one specimen of Galerix,compare with extant S.E. Asian faunas that havehumid forest biotopes with thick undergrowth,often close to water bodies. Crusafontina aff.endemica is morphologically comparable to theextant species Anourosorex squamipes, living in themountainous forests of SE-Asia. The extremelyspecialized dentition of the extinct dimylids, espe-cially of Metacordylodon, suggests a conchiovorousdiet. Some dimylids are believed to have beensemiaquatic in their behavior. The desmanArchaeodesmana, like extant desmans, certainlyhad a semiaquatic mode of life.
CHIROPTERA (G. Topal)
The Rudabánya bat assemblage includes the fol-lowing species: Eptesicus campanensis; Eptesicussp. nov.; cf. Miostrellus risgoviensis.
Up to now, all of the Rudabánya localities sam-ple open-air habitats where bats occur occasional-ly. Rudabánya’s bats are indicative of tropicalmarshy woods where they lived under tree bark, inthe holes of trees, or in the cracks of mountainwalls. Their occurrence in the pond and marshenvironments would have been the coincidence oftheir falling into those habitats. It is worthwhilenoting that Rudabánya’s bat fauna lacks “true”tropical forms such as hipposiderids. However,bones of a few species of the generally distributedvespertilionids have been found. Some of themhave been found to be related to the NorthAmerican fauna. Engesser (1979) has establishedthis same biogeographic connection for Mioceneinsectivores and rodents, while Topál (1989a, b)has demonstrated it for plecotine bats.

12 R.L. BERNOR - L. KORDOS - L. ROOK
PRIMATES
Pliopithecidae or Incertae sedis (L. Kordos)Anapithecus hernyaki was initially reported from
Rudabánya (Hungary, late Miocene, MN9 Zone) byKretzoi (1974) as Pliopithecus hernyaki. Kretzoi(1975) then nominated a new subgenus,Pliopithecus (Anapithecus) hernyaki, which he dis-tinguished from other European pliopithecinesincluding the well-known genera Pliopithecus sensustricto, Epipliopithecus and Plesiopliopithecus.Ginsburg & Mein (1980) raised Anapithecus togenus rank and referred it to the SubfamilyCrouzelinae (of the Pliopithecidae) (also see:Ginsburg, 1964, 1986; Harrison, 1987). Dentalcharacters of Anapithecus have been analysed andreported since by Begun (1989, 1991), discussedand compared further by Harrison (Harrison etal.1991; Harrison in Andrews et al., 1996).
The Rudabánya Anapithecus hernyaki teeth aremorphologically identical to those from Götzen-dorf (Austria; Zapfe, 1989a; Andrews et al., 1996),Salmendingen (Germany; Begun, 1989) and Priay(France; = Pliopithecus priay; Welcomme et al.,1991, formally attributed here to Anapithecushernyaki).
Harrison et al. (1991) gave an emended diagno-sis for the genus Pliopithecus, and later Harrison(in Andrews et al., 1996) listed some key charac-teres of the lower dentition that he erected to dis-tinguish species of Crouzelinae and Pliopithecinae.He further described a number of dental charactersthat are significant for the Pliopithecidae, placingthem in a monophyletic group.
Up until this time, it has never been questionedwhether Anapithecus hernyaki belongs to thePliopithecidae. However, a detailed morphologicalanalysis (Kordos, in progress) reveals thatAnapithecus hernyaki differs from Pliopithecusantiquus (the type of the Pliopithecinae) in manycritical morphological characters of the dentition.The other pliopithecine taxa (Pl. platyodon and Pl.vindobonensis) also exhibit significant differenceswith Anapithecus hernyaki in their tooth morpholo-gy, while “Pliopithecus priensis” is almost identicalin all characters to Anapithecus hernyaki.
The crouzeliines are taxonomically more diversethan the pliopithecines. They differ from the latterin their greater development of cheek tooth shear-ing crests (Köhler et al., 1999). According toKordos’ observations on the Rudabánya assem-blage, Anapithecus hernyaki’s lower dentition dif-fers significantly from both the pliopithecines andcrouzeliines, and is phylogenetically distinct fromthe Pliopithecidae. This result is supported by therecently discovered Anapithecus hernyaki femur atRudabánya that differs significantly from thePliopithecus vindobonensis “Eppelsheim femur”(Kordos & Begun, 1999).
Hominoidea (D. Begun)Begun & Kordos (1993) have assigned the
Rudabánya Dryopithecus to the species D. brancoi.The conclusions that D. brancoi is represented atboth Salmendingen and Rudabánya, and that theSt. Stephan and St. Gaudens samples are both D.fontani, have interesting paleobiogeographic impli-
cations (Begun, 2001). Other probable localities forD. brancoi are Mariathal in Austria and Ebingen,Trochtelfingen, Wissberg, and Melchingen inGermany. Most of the datable localities with D.brancoi are MN 9 in age (Mein, 1986, 1990;Steininger, 1986; Steininger et al., 1990; de Bruijn,et al., 1992; Rögl, et al., 1993; Andrews, et al., 1996).Only Salmendingen is younger and probably attrib-utable to MN 10 (Mein, 1986; 1990; Sen, 1996).This is broadly contemporaneous with D. laietanusand D. crusafonti (Spain). The MN 8 taxon D.fontani has the greatest geographic range, extend-ing from southwestern France to southeasternAustria. MN 9-10 taxa are more geographicallyrestricted, with two species known only fromnortheastern Spain, and a third ranging from theHessen Rhine to the Pannonian Basin.
Dryopithecus fontani is the oldest and mostprimitive species (Begun, 1992). The Vallesianspecies D. crusafonti, D. laietanus and D. brancoihave more elongated molars and premolars, labi-olingually thicker upper incisors, buccolinguallycompressed canines, and little or no expression ofmolar cingula. The mandibles of SpanishDryopithecus tend to be more robust relative todepth and to dental dimensions, and have broadextra molar sulci (Begun, 1992). The Vallesianspecies of Dryopithecus probably represents the sis-ter clade to D. fontani.
Dryopithecus is cladistically a stem great ape(hominid), but there is disagreement on the rela-tionship of Dryopithecus to other hominids.Andrews (1992) and Andrews et al. (1996) considerDryopithecus to be stem great ape. However, sever-al researchers including Andrews et al. (1996), havepresented evidence of shared derived characters ofspecific great apes and Dryopithecus. One hypothe-sis is that Dryopithecus is a member of aSivapithecus/Pongo clade (Moyá-Solá & Köhler,1993). An alternative hypothesis is that Dryopi-thecus is in the clade of the African apes andhumans (Begun, 1994, 1995; Begun & Kordos,1997). Dryopithecus shares with African great apesand Australopithecus a continuous supraorbitaltorus, a shallow sulcus supratoralis, a prominentglabella, an increase in the anteroposterior devel-opment of the frontal bone in the temporal fossa,increased neurocranial length relative to breadth,fused tympanic and articular portions of the tem-poral bone, a deep glenoid fossa, a small articulartubercle, a broad, projecting entoglenoid process, abroad, flat nasal aperture base, a stepped sub-nasalfloor, a biconvex nasoalveolar clivus, and an eth-moidal frontal sinus. There are no derived charac-ters shared between Dryopithecus and any onemember of the African ape/human clade, suggest-ing that Dryopithecus is the sister clade to theAfrican apes and humans as a group. Ourano-pithecus shares many of these characters as well,and is probably closely related to Dryopithecus(Begun, 1995; Begun & Kordos, 1997).
Dryopithecus probably evolved in situ in Europeafter the divergence of the Sivapithecus-Pongoclade. The oldest species, D. fontani, split to formtwo allopatric clades, one in Spain with twospecies, and the other in Germany and CentralEurope with one species. This pattern of species
13RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
distribution along with independant evidence ofecological changes and the apparent extinction ofD. fontani, suggests a vicariant mode of speciation.
While there is no clear evidence of hominids inAfrica before about 6 Ma (Hill & Ward, 1988;Leakey et al, 1996), hominids are numerous andvaried in Eurasia until about this time. This sug-gests the hypothesis that one Eurasian form, prob-ably closely related to the Dryopithecus/Ourano-pithecus clade, migrated to Africa and radiated intothe known lineages of African apes and humans(Begun, 1997).
Sexual dimorphism in Dryopithecus brancoi (D.Cameron)
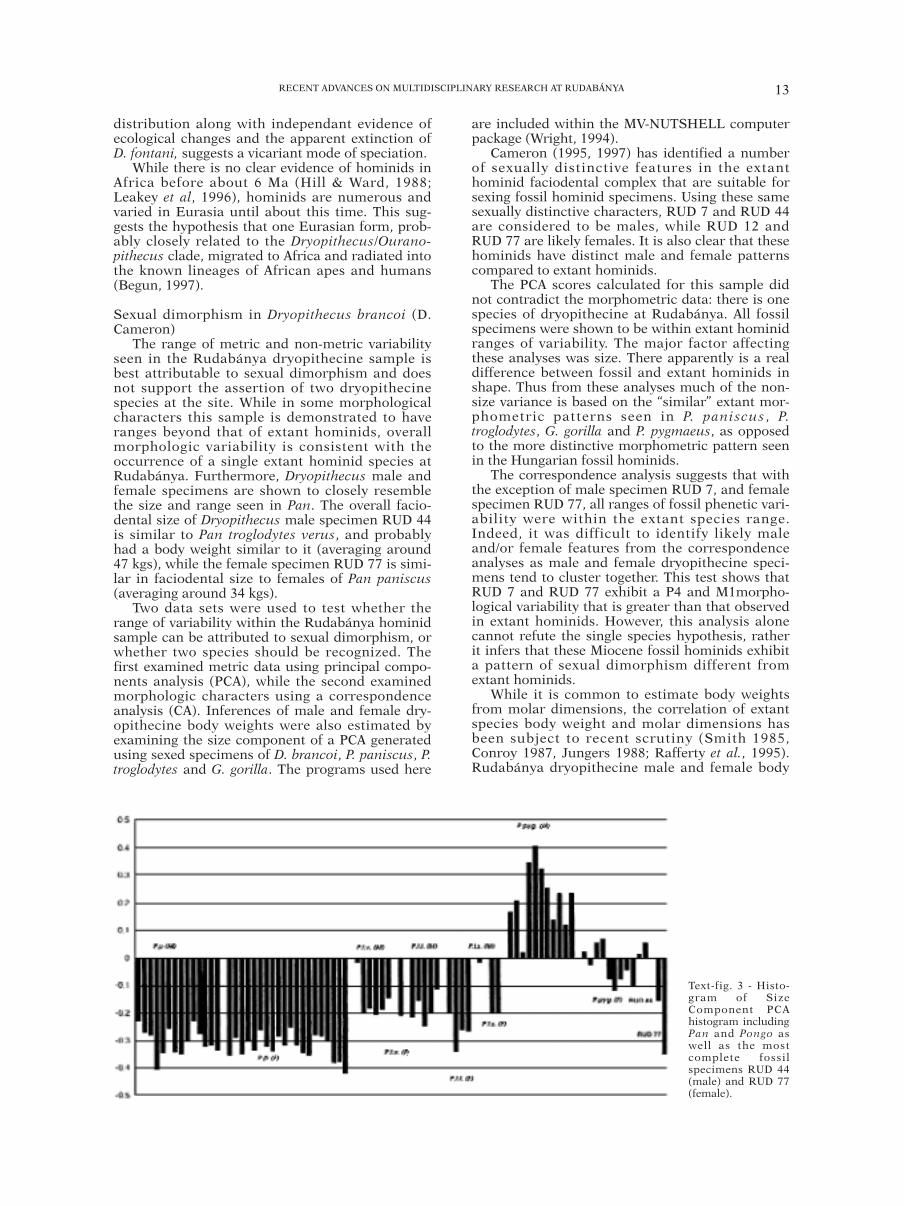
The range of metric and non-metric variabilityseen in the Rudabánya dryopithecine sample isbest attributable to sexual dimorphism and doesnot support the assertion of two dryopithecinespecies at the site. While in some morphologicalcharacters this sample is demonstrated to haveranges beyond that of extant hominids, overallmorphologic variability is consistent with theoccurrence of a single extant hominid species atRudabánya. Furthermore, Dryopithecus male andfemale specimens are shown to closely resemblethe size and range seen in Pan. The overall facio-dental size of Dryopithecus male specimen RUD 44is similar to Pan troglodytes verus, and probablyhad a body weight similar to it (averaging around47 kgs), while the female specimen RUD 77 is simi-lar in faciodental size to females of Pan paniscus(averaging around 34 kgs).
Two data sets were used to test whether therange of variability within the Rudabánya hominidsample can be attributed to sexual dimorphism, orwhether two species should be recognized. Thefirst examined metric data using principal compo-nents analysis (PCA), while the second examinedmorphologic characters using a correspondenceanalysis (CA). Inferences of male and female dry-opithecine body weights were also estimated byexamining the size component of a PCA generatedusing sexed specimens of D. brancoi, P. paniscus, P.troglodytes and G. gorilla. The programs used here
are included within the MV-NUTSHELL computerpackage (Wright, 1994).
Cameron (1995, 1997) has identified a numberof sexually distinctive features in the extanthominid faciodental complex that are suitable forsexing fossil hominid specimens. Using these samesexually distinctive characters, RUD 7 and RUD 44are considered to be males, while RUD 12 andRUD 77 are likely females. It is also clear that thesehominids have distinct male and female patternscompared to extant hominids.
The PCA scores calculated for this sample didnot contradict the morphometric data: there is onespecies of dryopithecine at Rudabánya. All fossilspecimens were shown to be within extant hominidranges of variability. The major factor affectingthese analyses was size. There apparently is a realdifference between fossil and extant hominids inshape. Thus from these analyses much of the non-size variance is based on the “similar” extant mor-phometric patterns seen in P. paniscus , P.troglodytes, G. gorilla and P. pygmaeus, as opposedto the more distinctive morphometric pattern seenin the Hungarian fossil hominids.
The correspondence analysis suggests that withthe exception of male specimen RUD 7, and femalespecimen RUD 77, all ranges of fossil phenetic vari-ability were within the extant species range.Indeed, it was difficult to identify likely maleand/or female features from the correspondenceanalyses as male and female dryopithecine speci-mens tend to cluster together. This test shows thatRUD 7 and RUD 77 exhibit a P4 and M1morpho-logical variability that is greater than that observedin extant hominids. However, this analysis alonecannot refute the single species hypothesis, ratherit infers that these Miocene fossil hominids exhibita pattern of sexual dimorphism different fromextant hominids.
While it is common to estimate body weightsfrom molar dimensions, the correlation of extantspecies body weight and molar dimensions hasbeen subject to recent scrutiny (Smith 1985,Conroy 1987, Jungers 1988; Rafferty et al., 1995).Rudabánya dryopithecine male and female body
Text-fig. 3 - Histo-gram of SizeComponent PCAhistogram includingPan and Pongo aswell as the mostcomplete fossilspecimens RUD 44(male) and RUD 77(female).

R.L. BERNOR - L. KORDOS - L. ROOKweight was estimated using a principal compo-nents analysis (PCA). The use of a PCA was deemedappropriate as it examines the size of a number ofvariables within the one analysis (in this case 15variables were analysed including upper facial,palate and dental morphometrics), rather thanexamining variables individually. Text-fig. 3 is ahistogram of the first component (size) of a PCAincluding Pan and Pongo as well as the most com-plete fossil specimens RUD 44 (male) and RUD 77(female). Pan troglodytes specimens were dividedinto subspecies, so that a greater resolution of bodyweights could be achieved for the fossil specimens.Those specimens that could not be allocated to asubspecies were removed from the data set. Fromthis analysis it is shown that RUD 44 is similar insize to specimens of P. t. versus, while RUD 77 issimilar to males and females of P. paniscus. Fromthe correlation with fossil and extant hominidfaciodental size, male D. brancoi specimens areestimated to have had an average weight of 47 kg,while the female specimen is estimated to have aweight of 34 kg.
Rudabánya fossil primate paleodiet (P. Ungar)At least two ape-like primates are known from
late Miocene deposits in western and centralEurope. Both primitive catarrhines called pliop-ithecids and the hominoid Dryopithecus have beenreported from deposits ranging from Castell deBarbera, Spain to Salmendingen, Germany, toRudabánya, Hungary (e.g., Begun, 1989; Kretzoi,1975; Moyá-Solá et al., 1990). While it is difficult todetermine whether these primates coexisted inboth time and space, it has been argued that theyshow adaptations indicating that they partitionedtheir niches to allow “noncompetitive sympatry”.Following this line of reasoning, Szalay & Delson(1979) suggested that the pliopithecids as a groupwere probably folivores, whereas Dryopithecusspecies were more likely frugivorous. Ginsburg &Mein (1980) further argued that among the pliop-ithecids, the crouzelines were more folivorous thanthe pliopithecines. This latter scenario can be con-sidered consistent with the concept of diet-relatedniche separation, as those pliopithecids found atthe Dryopithecus-bearing localities at Rudabánya,Salmendingen, and Castell de Barbera may all becrouzelines (re: Begun, 1989; Ginsburg, 1986).
I attempt here to reconstruct the diets ofAnapithecus hernyaki and Dryopithecus brancoifrom Rudabánya, through the quantitative studiesof molar tooth shearing crest development andantemortem microscopic tooth wear (i.e.,microwear). These approaches allow comparisonsof the diets of these primates to one another and tothose of other fossil and extant catarrhines. This inturn can provide new insights into the paleoecolo-gy (and perhaps even paleosynecology) of thesecatarrhines. The present results provide no evi-dence for broad trophic differences or concomitantniche separation between Anapithecus andDryopithecus from Rudabánya. Evidence from themicrowear and shearing crest length data both sug-gest that these primates had diets dominated bysoft fruits.
All second molars (M2s) of Rudabánya primates
examined come from collections at the MagyarGeologiai Szolgalat and Naturhistorisches MuseumWien. High-resolution replicas were prepared fol-lowing conventional procedures (re: Ungar, 1996).Only unworn or nearly unworn mandibular secondmolars (M
2’s) were examined for the shearing crest
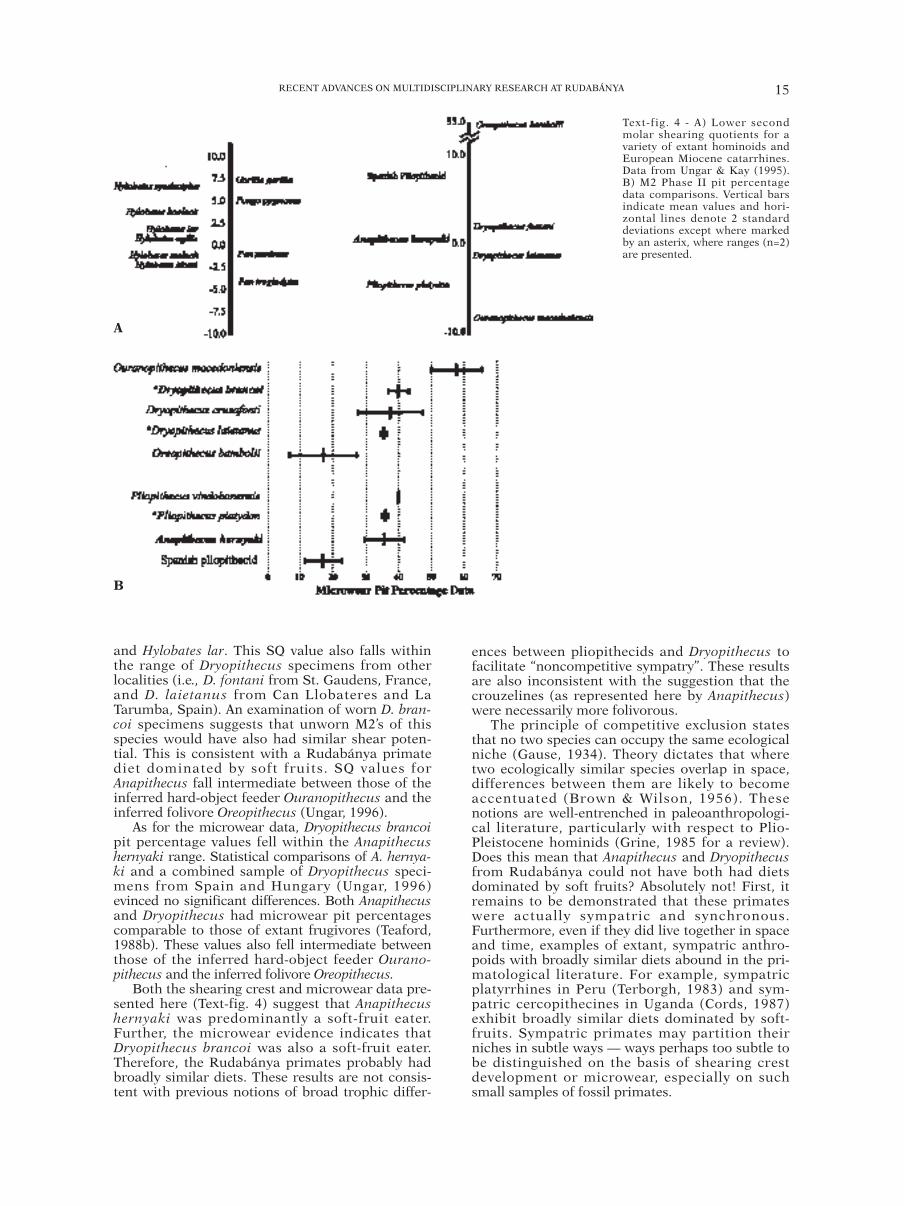
length study (following Kay, 1978). While no avail-able Dryopithecus brancoi specimens met this crite-rion, eight Anapithecus hernyaki teeth could beincluded in this analysis (see Table 2). The lengthsof shearing crests 1-8 and mesiodistal occlusallengths of these specimens were measured and log-transformed (re: Ungar & Kay, 1995). Shearingquotients (SQs) were computed as deviations froma least-squares line regressing summed shearingcrest length over mesiodistal occlusal length for avariety of extant frugivorous hominoids (Text-fig.4). Positive SQ values indicate longer crests thanexpected for an extant frugivorous ape, whereasnegative SQ values indicate shorter crests.
Replicas of all worn M2s were examined in sec-ondary mode by scanning electron microscopy(SEM). Five specimens of Anapithecus hernyaki andtwo Dryopithecus brancoi individuals (see Table 2)preserved antemortem microwear unobscured bytaphonomic or casting artifacts (re: Teaford 1988a).Polaroid photomicrographs of Phase II facets ofthese specimens were taken at 500x magnificationand scanned to computer image files. Dentalmicrowear features were identified usingMicroware 2.1, and pit percentages (percentages ofall features with length-to-width ratios less than4:1) were computed for each specimen. Microweardata were then compared between the Rudabányaprimates, and results were put in the context ofstudies of extant primates and other fossil formsfrom the late Miocene of Europe (Text-fig. 4).
Results for both the shearing crest andmicrowear studies indicate that Anapithecushernyaki had a mean SQ value of 0.60, nearly at thecenter of the range for extant frugivorous apes.This value falls closest to those for Hylobates agilis
TABLE 2 - Shearing Crest and Microwear Pit PercentageData for Available Primate specimens from Rudabánya.
Shearing Crest Study Microwear StudySpecimen M2 Length shearing crests Pit percentages
(mm) (mm)
Anapithecus hernyakiRUD 76 – – 37.3RUD 89 8.5 19.9 43.8RUD 90 – – 29.9RUD 91 8.6 19.6 –RUD 98 8.7 18.3 –RUD 100 9.4 19.6 –RUD 106 8.8 19.5 29.3RUD 108 8.6 20.0 37.5RUD 122 8.8 19.8 –RUD 128 8.2 19.3 –
Dryopithecus brancoiRUD 77 – – 43.0RUD 141 – – 36.6
Tshear is the summed lengths of shearing crests 1-8.
15RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
and Hylobates lar. This SQ value also falls withinthe range of Dryopithecus specimens from otherlocalities (i.e., D. fontani from St. Gaudens, France,and D. laietanus from Can Llobateres and LaTarumba, Spain). An examination of worn D. bran-coi specimens suggests that unworn M2’s of thisspecies would have also had similar shear poten-tial. This is consistent with a Rudabánya primatediet dominated by soft fruits. SQ values forAnapithecus fall intermediate between those of theinferred hard-object feeder Ouranopithecus and theinferred folivore Oreopithecus (Ungar, 1996).
As for the microwear data, Dryopithecus brancoipit percentage values fell within the Anapithecushernyaki range. Statistical comparisons of A. hernya-ki and a combined sample of Dryopithecus speci-mens from Spain and Hungary (Ungar, 1996)evinced no significant differences. Both Anapithecusand Dryopithecus had microwear pit percentagescomparable to those of extant frugivores (Teaford,1988b). These values also fell intermediate betweenthose of the inferred hard-object feeder Ourano-pithecus and the inferred folivore Oreopithecus.
Both the shearing crest and microwear data pre-sented here (Text-fig. 4) suggest that Anapithecushernyaki was predominantly a soft-fruit eater.Further, the microwear evidence indicates thatDryopithecus brancoi was also a soft-fruit eater.Therefore, the Rudabánya primates probably hadbroadly similar diets. These results are not consis-tent with previous notions of broad trophic differ-
ences between pliopithecids and Dryopithecus tofacilitate “noncompetitive sympatry”. These resultsare also inconsistent with the suggestion that thecrouzelines (as represented here by Anapithecus)were necessarily more folivorous.
The principle of competitive exclusion statesthat no two species can occupy the same ecologicalniche (Gause, 1934). Theory dictates that wheretwo ecologically similar species overlap in space,differences between them are likely to becomeaccentuated (Brown & Wilson, 1956). Thesenotions are well-entrenched in paleoanthropologi-cal literature, particularly with respect to Plio-Pleistocene hominids (Grine, 1985 for a review).Does this mean that Anapithecus and Dryopithecusfrom Rudabánya could not have both had dietsdominated by soft fruits? Absolutely not! First, itremains to be demonstrated that these primateswere actually sympatric and synchronous.Furthermore, even if they did live together in spaceand time, examples of extant, sympatric anthro-poids with broadly similar diets abound in the pri-matological literature. For example, sympatricplatyrrhines in Peru (Terborgh, 1983) and sym-patric cercopithecines in Uganda (Cords, 1987)exhibit broadly similar diets dominated by soft-fruits. Sympatric primates may partition theirniches in subtle ways — ways perhaps too subtle tobe distinguished on the basis of shearing crestdevelopment or microwear, especially on suchsmall samples of fossil primates.
A
B
Text-fig. 4 - A) Lower secondmolar shearing quotients for avariety of extant hominoids andEuropean Miocene catarrhines.Data from Ungar & Kay (1995).B) M2 Phase II pit percentagedata comparisons. Vertical barsindicate mean values and hori-zontal lines denote 2 standarddeviations except where markedby an asterix, where ranges (n=2)are presented.

16 R.L. BERNOR - L. KORDOS - L. ROOK
LAGOMORPHA (S. Sen)
At Rudabánya lagomorpha are represented onlyby the Ochotonidae Eurolagus sp.. This genus hada geographical distribution typically limited to cen-tral and western Europe. Eurolagus first occurs atRothenstein 1 (MN 5, Germany) and persists,although as a relatively rare element, until the earlyLate Miocene. With only one species recognized,Eurolagus fontannesi, its last occurrence is report-ed in the Spanish locality of Terrassa (MN 10).
RODENTIA
Cricetidae, Sciuridae, Gliridae and Eomyidae(G. Daxner- Höck and O. Fejfar)
Rudabánya rodents belonging to the familiesSciuridae, Cricetidae, Gliridae and Eomyidaeinclude: Sciuridae - Spermophilinus bredai, Blackiamiocaenica, Hylopetes sp., Miopetaurista sp.,Albanensia grimmi; Cricetidae - Eumyarion aff.latior, Democricetodon sp., Kowalskia cf. schaubi,Microtocricetus molassicus; Gliridae - Paragliruluswerenfelsi, Glis aff. minor, Myoglis ucrainicus,Muscardinus hispanicus, Muscardinus aff. valle-siensis, Glirulus lissiensis. Eomyidae -Eomyopscatalaunicus.
This portion of the rodent fauna of Rudabányais characterized by a early Vallesian (MN9) associa-tion with: 1) mainly middle Miocene rodents with2) some late Miocene first appearing taxa and 3)the lack of murids and some other rodents knownto first occur regionally in MN10.
Among cricetids Eumyarion latior, is known tooccur in Central Europe during the middle and lateMiocene, while Microtocricetus molassicus isknown to occur only in early late Miocene horizons(MN 9-10) of western, central and eastern Europe(Wellcomme et al., 1991; Fahlbusch & Mayer, 1975;Kowalski, 1993; Daxner-Höck, 1996; Topachevskii& Scorik, 1992).
Among the Rudabánya flying squirrels and sci-urids, Albanensia grimmi dominates the rodentassemblage in its abundant and well preservedteeth. This species is mainly recorded in centralEurope in MN 9 but rarely occurs in MN 10(Daxner-Höck, 1996). Miopetaurista is thus far veryrare in the late Miocene and is represented inRudabánya by a more progressive form related toMiopetaurista crusafonti from Can Llobateres(MN9). Blackia miocaenica is a species with a verylong stratigraphic duration, and as such is of limit-ed biostratigraphic use. An advanced species ofHylopetes was not known from Rudabánya beforenow, but is known from elsewhere to range at leastfrom MN4 to present. Spermophilinus bredai is avery abundant sciurid in middle Miocene faunas,and as a rule is replaced by Spermophilinus tur-olensis in the late Miocene. The Rudabánya form isone of a few rare occurrences of Spermophilinusbredai known from the late Miocene.
Glirids are documented from Rudabánya, insome cases by relative meagre material. There area few teeth referable to Paraglirulus werenfelsi,Muscardinus hispanicus, Muscardinus aff. valle-siensis and Glirulus lissiensis, and better qualitymaterial of Myoglis ucrainicus and Glis aff. minor.
Although descending from early Miocene glirid lin-eages, four species (M. vallesiensis, M. hispanicus,M. ucrainicus and G. minor intermedius) firstoccurred in MN9. The eomyid Eomyops catalauni-cus is known to occur only in the Vallesian.
Anomalomyidae (L. Kordos)Two species of anomalomyids were reported in
the first faunal list of Rudabánya (Kretzoi et al.,1976): Anomalomys cf gaillardi and Prospalax pet-teri. This material was subsequently studied byFejfar and later published by Rabeder (1985) asAnomalomys gaudryi. On the basis of material col-lected between 1977 and 1987, Kordos (1989)described a new species, Anomalomys rudabanyen-sis in place of the two previous taxa, reviewed byBolliger (1996) and Kowalski (1994). A detailedstudy of central and eastern European NeogeneAnomalomyidae is important from both a phyloge-netic and biostratigaphic point-of-view. Along withcricetids and murids, the anomalomyids are one ofthe most abundantly represented families in thelate Miocene of the Carpathian Basin. TheRudabánya Anomalomyidae belongs to theAnomalomys gaudryi - rudabanyensis - petteri lin-eage. The genus Anomalomys may be derived froma small species of Eumyarion. Early Miocene mem-bers of primitive Anomalomys, A. aliveriensis andA. minor, are early derivatives of the group (deBruijn & Sarac, 1991). The occurrence of A. cf. rud-abanyensis has been reported from outside theCarpathian Basin at Hilleniche, in Germany (MN9; Bolliger, 1996). Some newly discovered and part-ly unpublished Vallesian samples of Anomalomyshave been recovered from Gritsev (Ukraine; possi-bly MN 9) and Götzendorf (Austria; MN 9), andexhibit a transitional morphology betweenAnomalomys rudabanyensis and Allospalax(“Prospalax” or “Allospalax”) petteri (Kordos,unpublished). Consequently, the Allospalax speciesshould be synonymized with Anomalomys.
Castoridae (L. Kordos)The lacustrine and swampy sediments of
Rudabánya contain a large sample of a singlebeaver species, Trogontheium minutum .Trogontherium minutum is smaller in size thanTrogontherium rhenanum from Dorn Dürkheim(Germany; Franzen & Storch, 1975), and is wellknown in the Carpathian Basin between MN9 andMN13. Trogontherium minutum is first recordedfrom Oellingen (MN 3), Germany, and was fre-quent in Europe from MN4 to MN11, becamingrare during MN12 and MN13.
CARNIVORA (L. Werdelin)
Carnivora are taxonomically well represented atRudabánya, although most of the material is frag-mentary in nature and no large samples are avail-able for statistical analysis. Most of the taxa identi-fied herein are known from other deposits that aresimilar in age to Rudabánya, but there may besome new taxa represented among theAmphicyonidae and Mustelidae. All in all, a mini-mum of 17 species-level taxa have been identified,representing 8 carnivoran families: Ursidae,

17RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
Procyonidae, Amphicyonidae, Mustelidae,Viverridae, Nimravidae, Felidae, and Hyaenidae.
As is usual in European deposits of Astaracianand Vallesian age (Werdelin, 1996), the Mustelidaeare by far the most diverse family, with a minimumof 7 species-level taxa. These taxa represent a diver-sity of adaptational types, including small, omnivo-rous Melinae and Mephitinae (Taxodon cf. T.sansaniensis, Melinae gen et sp. indet., Propu-torius) and small hypercarnivorous Mustelinae(e.g., Martes cf. M. filholi, cf. Trochictis sp.). Themost abundant mustelid is, however, the semi-aquatic Lutrinae Paralutra jaegeri. At Rudabánya,this species is accompanied by rare remains ofanother, larger species of Paralutra, previouslyunknown.
The Ursidae are represented by two species,Ursavus primaevus and U. brevirhinus. This associ-ation is interesting in itself, as it is otherwisereported only at Can Llobateres (Spain, MN 9),although this may in part be an artefact of the rela-tive rarity of both of these taxa.
A unique association for Rudabánya is betweenthe two Procyonidae Alopecocyon sp. and Simocyondiaphorus. Of these, the former is generally a mid-dle Miocene taxon and the latter exclusively a lateMiocene one (Werdelin, 1996).
Some of the few remains of large carnivores canbe attributed to a large sized amphicyonid. Thisfamily is relatively rare in European late Miocenedeposits (Viranta, 1996) and the morphologicalcharacteristics of the available material fromRudabánya indicates the presence of a new taxonwith more omnivorous adaptations thanAmphicyon sp.
Another large carnivore represented atRudabánya is the nimravid Sansanosmilus jour-dani, of which only a single definitely referred frag-ment has been identified. Felid sabertooths are,however, conspicuously absent at Rudabánya,unlike in some other penecontemporaneous locali-ties such as Can Llobateres (Spain) andEppelsheim (Germany).
Although sabertooth felids are absent,Rudabánya does include two members of theFelidae, Pseudailurus lorteti and its smaller relativeP. turnauensis. Both of these are relatively wellknown from Astaracian deposits, but are less com-mon in Vallesian ones (Werdelin, 1996). A hyper-carnivorous Viverridae, Semigenetta grandis, is alsorepresented at Rudabánya, and is more abundantin the material than either of the felid species.
The most surprising aspect of the carnivoranassemblage of Rudabánya lies in the dearth ofhyaenas. Only some fragmentary specimens tenta-tively assigned to cf. Thalassictis montadai are pre-sent, in contrast to contemporaneous localities,which generally include relatively abundant mater-ial of one or two species of hyaenid (Werdelin &Solounias, 1996). The explanation for this relativelack of Hyaenidae must lie in local environmentalconditions at Rudabánya.
The carnivoran diversity at Rudabánya ismatched among MN 9 localities only by CanLlobateres in Spain, which records 24 species-leveltaxa (Petter, 1963, 1967; Crusafont Pairó & Kurtén,1976). The carnivoran assemblages of these two
localities are very similar, both in species contentand in trophic structure. The few differences canbe accounted for by the more isolated geographiclocation of Can Llobateres, leading to greaterendemicity, and a somewhat greater environmentalheterogeneity, leading to the presence of derivedspecies characteristic of somewhat more openenvironments than were apparently present atRudabánya.
PROBOSCIDEA (M. Gasparik)
Four proboscidean taxa are known from the lateVallesian (MN 9) vertebrate fauna of Rudabánya:Deinotherium giganteum, Tetralophodon longi-rostris , Tetralophodon cf. longirostris andStegotetrabelodon gigantorostris. These species aretypical proboscideans for the European Vallesian.
Deinotherium giganteum is represented by a dp4and another milk molar fragment in that thedeinothere cheek tooth morphology is rather uni-form between later Miocene species (Gasparik,1993). Up until recently, Deinotherium was notknown from Rudabánya. These new finds are veryimportant because the D. giganteum and T. lon-girostris have been found together from otherEuropean Vallesian localities, such as the“Dinotheriensande” in Germany and from Ko⁄bányain Hungary.
Tetralophodon cf. longirostris has been identifiedon the basis of two lower tusk fragments and someupper and lower molars. Tobien (1986) referred allof the Rudabánya Tetralophodon to this taxon. Itseems however that with the material at hand, onecan separate some Tetralophodon remains from thetypical T. longirostris, but it is not certain whetherthey can be segregated into a different species orsubspecies, or if they fall within a single popula-tion’s range of variability.
Material referred to Tetralophodon longirostris isabundant and includes lower and upper tusks,lower and upper molars, as well as deciduous den-tition. The crown morphology is typical oftetralophodont gomphotheres. The intermediatemolars have four loph(id)s, the last ones (M3 andM
3) have five transverse crests. The development of
the talon(id)s is variable. The pretrite halfcrestsexhibit a trefoil pattern, and on the posterioraspect the conules are less developed or missing.The posttrites have 1-3 internal conelets and unde-veloped conules. In some cases the secondary tre-foil is visible. Cement is lacking.
Material referred to Stegotetrabelodon gigan-torostris includes four upper tusks, lower andupper molars, as well as deciduous dentition. Thistaxon is distinguished from Tetralophodon lon-girostris by its larger size and some small differ-ences in the molar crown morphology. The incisorshave a large diameter. On the molar crowns theconules are slightly more developed than in thecase of T. longirostris. The half loph(id)s show anincipient anancoidy. This is not a true anancoidybecause the main cones (conids) are approximatelyin line with one another, but the pretrite conulesare well developed (the posterior conules on thelower molars and the anterior ones on the uppers)hence it seems that the halfcrests are in slight alter-

18 R.L. BERNOR - L. KORDOS - L. ROOK
nate position. This phenomenon becomes morevisible during the wear of the crests.
PERISSODACTYLA
Equidae, Hipparion s.l. (R.L. Bernor, M. Armour-Chelu, T. Kaiser and R. Scott)
Kretzoi 1983 named a new species of“Hipparion” from Rudabánya based on a completemetatarsal III, “Hipparion” intrans. In a moredetailed description of the hipparion material col-lected solely under the direction of Kretzoi, Bernoret al. (1993a) reported the occurrence of anadvanced member of the Hippotherium primigeniumlineage at Rudabánya. In a subsequent comparisonwith the Götzendorf (MN 9/10, Vienna Basin,Austria) hipparion assemblage, Bernor et al. (1993b)later concurred that the Rudabánya hipparion rep-resented an advanced population of the Hippo-therium primigenium lineage and suggested a lateMN 9 correlation for Rudabánya. A recent analysisof the Dorn Dürkheim (Germany, MN 11) hipparionassemblage by Bernor & Franzen (1997), the Sümeg(Hungary, MN 10) assemblage by Bernor et al.(1999), and a study of the entire Rudabánya hippari-on assemblage collected between 1977 and 1994,have led us to new interpretations of this assem-blage’s systematics, ecological preferences and bio-geographic relationships (Bernor et al., in press).
We still recognize Hippotherium intrans as beinga valid taxon and being the predominate hipparionat Rudabánya. We also recognize that there is ararer, smaller hipparion morph that has beenfound in the Upper Lignite levels of Rudabánya II,the youngest stratigraphic bed in this particularRudabánya section. Because of the rarity of thissmaller hipparion, we have chosen not to recognizea formal species name for it.
Rudabánya Hippotherium intrans is distinctfrom German and Austrian populations ofHippotherium primigenium in its longer metatarsalIII. While there are few fossil hipparion from MN10 localities in the Vienna Basin, material fromProttes would appear to be very similar in its molarocclusal morphology to the Götzendorf and Sümegspecies. These observations suggest that by latestMN 9 time, or Pannonian F equivalent, thePannonian Basin hipparions diverged fromHippotherium primigenium. Bernor & Franzen’s(1997) study of the early Turolian (MN 11) hippari-on assemblage from Dorn Dürkheim (Germany)showed that this population retained conservativecharacters that ally it closely with older MN 9 hip-parion from Eppelsheim and Höwenegg, Germany.Given the current state of knowledge, thePannonian late MN 9 (= Pannonian F) hipparionassemblages would appear to have become vicari-antly separated from the German late MN 9 – MN11 hipparions to the west. The small Sümeg hip-parion appears to be most closely related to thesmall Greek MN 10 form Cremohipparion mace-donicus, and would if that attribution were toprove to be true, be an indication of an openingbiogeographic connection between the centralParatethys and Macedonia (Bernor et al., in press).
In Kaiser’s recent analysis of Hippotheriumintrans mesowear (Bernor et al., in press), he found
that this predominant Rudabánya hipparion had amixed diet including grass and less abrasivebrowse components. This is observed as a deepgrooving across the middle of the maxillary cheek(axis being along the protocone-mesostyle plane)teeth flanked by sharp peaks with the axis runninglabio-lingually across the central point of the pre-fossette and postfossette.
We have used a series of statistical tests to char-acterize Hippotherium intrans postcranial function-al anatomy and ecomorphology. This includesbivariate plots, logarithmic ratio diagrams andprincipal components analysis (Bernor et al., inpress). We have found that Hippotherium intranshad relatively longer metapodials than theHöwenegg sample and as such was adapted formore cursorial behavior. Its closest relatives in thisregard are the larger horses from the Vienna Basinlocality of Gols (MN 10, Austria) and specimensfrom Csákvár (MN 11, Hungary). The estimatedbody mass for Hippotherium intrans based onMAFIV12038 (= 242 kg) is the same as for theHöwenegg MTIII sample (=242 kg). The smallerand rarer horse at Rudabánya, Hipparion sp., maybe related to a smaller MCIII, MAFIV12039, whichhas an estimated body mass of 194 kg.
The Rudabánya hipparions have proven usefulfor finer correlations: Rudabánya 2 correlates withthe latest part of MN 9. This is based on the obser-vation that Hippotherium intrans is advanced overVienna Basin Pannonian D-E populations ofHippotherium primigenium, while not being soadvanced as the Götzendorf hipparion. Rögl et al.(1993) argued for a latest MN 9 (= PannonianStage F) age for Götzendorf based on the absenceof murids there. Bernor et al. (1999) have recentlystudied the equids from Sümeg identifying twotaxa in this fauna: a larger species, Hippotheriumsumegense, and a smaller species, ”Hipparion” sp.small. Of these two, Hippotherium sumegense com-pares closely to the Götzendorf hipparion in its dis-tinct mandibular premolar morphology. Bio-chronologic correlations based on Sümeg’s smallmammals strongly support the fauna’s MN 10 age.Therefore, we can either accept the latest MN 9attribution for Götzendorf based on the absence ofmurids, or accept the occurrence of Eozapus andHippotherium aff. sumegense at Götzendorf as evi-dence of its MN 10 age. In either case, Rudabánya’shipparion is suggestive of a Pannonian F correla-tion and late MN 9 attribution, ca. 10.0 – 9.7 Ma(Rögl & Daxner-Höck, 1996; Steininger et al., 1996;Bernor et al., 1999). By the same argument,Götzendorf and Sümeg are most plausibly correla-tive with Pannonian G, circa 9.7 – 9.0 Ma (sensuRögl & Daxner-Höck, 1996).
Tapiridae (R.L. Bernor)The tapiridae consist of a single fragmentary
lower molar tooth and a fragmentary phalanx fromRudabánya 2. The molar tooth fragment includesthe posterior lophid and distal tooth margin. It wasoriginally referred to the suid, Listriodon. Thisspecimen compares well with other Vallesian sam-ples of Tapirus priscus known from MN 9 ofCentral and Western Europe. This species is associ-ated with warm moist forested environments.

19RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
Chalicotheriidae (L. de Bonis, R.L. Bernor and J.Franzen)
The Rudabánya fauna contains the relativelyrare occurrence of the chalicothere, Chalicotheriumaff. goldfussi. This taxon is common in MN 9 ofCentral and Western Europe and like the tapir, isindicative of warm moist forested environments.This chalicothere is known to occur at bothRudabánya localities 2 and 3, and includes gooddiagnostic maxillary and mandibular cheek toothmaterial. The most complete specimen is an associ-ated partial upper and lower dentition. Our studyof this material has provisionally revealed someunique features of the anterior cheek tooth denti-tion that may or may not prove to distinguish itfrom other members of Chalicotherium goldfussi.Chalicotherium goldfussi is well known fromEppelsheim and Höwenegg, Germany (Zapfe,1989b). Of the two localities, Eppelsheim (ca. 10.5Ma.) is believed to be slightly older than Höwenegg(10.3 Ma; Woodburne et al., 1996); which, in turn,is believed to be older than Rudabánya (ca. 10-9.7Ma; Bernor et al., 1993).
Biogeographically, Rudabánya’s chalicotherewould appear to be a central European form andmay prove to be distinct from western EuropeanVallesian chalicotheres.
Rhinocerotidae (K. Heissig) There are three species of rhinoceroses in the
Rudabánya fauna. Two of them belong to the tribeAceratherini (Hoploaceratherium belvederense andAceratherium incisivum) and one to theRhinocerotini (Lartetotherium sansaniense).
The medium sized, primitive dicerorhine(Rhinocerotini) species cannot be related to one ofthe known Dicerorhinus species of the Miocene. Itshows more similarities to the small, middleMiocene Dicerorhinus steinheimensis than to thewell known large species Dicerorhinus schleierma-cheri from the Vallesian of central and westernEurope. Its dimensions are intermediate betweenboth of these species, and similar to the middleMiocene species Lartetotherium sansaniense. Theobserved differences indicate some phylogenetictrends within the species which suggest their sepa-ration from the typical L. sansaniense asLartetotherium aff. sansaniense. No postcranialremains have been assigned to this species.
The skeletal elements of the two Aceratherinispecies are easily distinguished by their size differ-ence and several very specific characters. TheRudabánya fauna is the first record of the largerspecies, Hoploaceratherium belvederense in the lateVallesian. The other one, the well knownAceratherium incisivum, is smaller and less fre-quent. Both have very large, high crowned malelower incisors in contrast to smaller female ones(not represented in this collection). Stronglycurved incisors with rather distinct wear surfacesare ascribed to Aceratherium incisivum, less curvedincisors with diffuse wear may representHoploaceratherium.
All three rhinoceroses had low crowned teethindicative of browsing, and all three are believed tohave inhabited a wooded or bush habitat. WhereasHoploaceratherium is known from the middle
Miocene, there is no record of Aceratherium inci-sivum earlier than lower Vallesian. The last occur-rence of both aceratherini is not sufficientlyknown. These taxa add no more specific informa-tion about the stratigraphic correlation of the siteother to say that Rudabánya is the youngest knownlocality with Lartetotherium.
ARTIODACTYLA
Suidae (M. Fortelius, R.L. Bernor, N. Fessaha andM. Armour-Chelu)
The Rudabánya suid fauna has been collectedover 18 years time and two taxa are currently rec-ognized from the Rudabánya locality: Propo-tamochoerus palaeochoerus an early suine which isthe most abundant artiodactyl at the site, and ararer form, a previously unrecognized species ofthe primitive tetraconodont, Parachleuastochoerussp. nov.
Kretzoi (in Kretzoi et al., 1976) listed the follow-ing suid taxa from Rudabánya: Korynochoeruspalaeochoerus, Conohyus ferreus (n. sp.) andListriodon splendens. We have found no evidence ofListriodon in the MAFI collections. The suid mater-ial is mostly dental but there are some importantpostcranial remains probably referable to bothspecies.
The Propotamochoerus sample includes over400 specimens allowing some reconstruction of thesequence of tooth eruption and replacement in thistaxon. Our analysis of this sample shows that theRudabánya population of Propotamochoeruspalaeochoerus compares most closely with the typespecimen of the species from Eppelsheim,Germany.
Parachleuastochoerus is possibly represented bytwo morphs, a larger form with relatively broadcheek teeth and a smaller one with relatively nar-row ones. Both morphs are larger than the Spanishspecies Parachleuastochoerus crusafonti (whichalso appears to show a high degree of interpopula-tion variability), and appears to have thinnerenamel, at least on the first molars. We do not rec-ognize Kretzoi’s nomen “Conohyus ferreus” for thismaterial because no type specimen was designatedand there remains uncertainty as to which speci-men/specimens would constitute a valid lectotype.Rather, we will erect a new species, Para-chleuastochoerus n. sp. (to be named in an editedvolume to follow), provisionally restricted to thelarger morph represented by a virtually completeupper and lower dentition. We do not formally rec-ognize the smaller morph as a taxon and defer thisdecision until a larger sample becomes available.
The Rudabánya suid fauna may suggest a bio-geographic disjunction between Spain andHungary sometime during MN 9. The Rudabányasuids are not particularly informative aboutpalaeoecology. Suines are notoriously broad intheir habitat tolerance, and the rather peccary-likeParachleuastochoerus persisted in western Europe(at least) into the drier world that followed themid-Vallesian crisis. However, the extremely highlocal abundance of Propotamochoerus is difficult toreconcile with arid and open habitats.

20 R.L. BERNOR - L. KORDOS - L. ROOK
Ruminants (A. Gentry)The Rudabánya ruminants are represented by
five species and four families including oneTragulidae (Dorcatherium naui), one Cervidae(Lucentia aff. pierensis), one Moschidae(Micromeryx flourensianus) and two Bovidae(Miotragocerus sp. and Bovidae, smaller sp.).
A Rudabánya right upper molar in early wear(1973/15), agrees with Dorcatherium naui, firstdescribed from Eppelsheim, Germany. It is moreadvanced than upper molars of middle MioceneDorcatherium (Fahlbusch, 1985; pl. 2) in that itslingual cusps are more fully crescentic, the outerwalls more nearly vertical, the labial rib of themetacone reduced, the enamel thinner, themesostyle more prominent, the parastyle project-ing less forwards, and the cingula weaker andsmoother surfaced. A tragulid left astragalus(1988/50), could be conspecific with the uppermolar.
Lucentia aff. pierensis is the most commonruminant at Rudabánya, with over 100 identifiedspecimens. Antlers are about the size of the mod-ern European Capreolus capreolus, although theteeth are perhaps bigger. The pedicle is long andset at a low angle above the horizontal plane. Thepedicle and antler diverge a little from their part-ner of the other side, and the antler then curvesslightly upwards and forwards in side view andeventually inwards again. The first tine is short andsmall, inserted quite high above the rose and oftenonly slightly angled on the distal beam. There is noadditional tine above the first tine. The cheek teethare low crowned and have rugose enamel. Upperteeth have lingual cingula. Upper molars havebasal pillars (entostyles), the styles are quite bulky,the paracone ribs are strong and somewhat inflect-ed forwardly, but metacone ribs are weak orabsent. The posterior crest of the protocone isbifurcated. On most lower molars there is aPalaeomeryx-fold. On M
3the hypoconulid lobe has
a central fossette. On P4
the lingual end of themetaconid crest has twin flanges directed anterior-ly and posteriorly. Fairly large and robust uppercanines are still present. The Rudabánya deerappears to be a little changed descendant ofEppelsheim Euprox dicranocerus (larger size andhigher first tine) or more probably a contempora-neous close relative evolving towards Lucentiapierensis.
Micromeryx flourensianus is known by morethan 60 specimens from Rudabánya. The tinyMicromeryx flourensianus, type species of thismoschid genus, is mainly a middle Miocenespecies, but has been reported as late as theVallesian of Spain (Morales & Soria, 1981) and hasbeen reported from China. It had enlarged maleupper canines and is thought to have been withoutantlers. The P
4’s usually show fusion of paraconid
with metaconid forming a closed wall along muchof the lingual side of the tooth. Thenius (1950) haspointed to the dangers of confusing the teeth withthose of Lagomeryx. Shoulder height in Micromeryxwas only about 40 cm. Judged from a reeconstruct-ed skeleton in Basel (Heizmann, 1976; fig.7), thelegs were longer than in modern Muntiacus.Habitual body conformation may have been like
that of the modern African neotragine antelopeNeotragus moschatus (hind legs flexed under thelowered rear of the body; see Kingdon, 1982:192-194; Estes, 1991: 42), and unlike a modernCapreolus capreolus.
Boselaphine bovids are known from the middleMiocene onwards. The Rudabánya boselaphine isknown by more than 70 specimens. It is assignedto Miotragocerus sp. on the evidence of a distal endof a right horn core, 1992/66, about 90 mm long.The tip itself is missing and the top of the anteriorkeel is just visible at the base of the fragment. Thetermination of the anterior keel some way belowthe tip of the horn core creates a demarcationbetween the main proximal part of the horn coreand a distal part often of markedly smaller antero-posterior diameter. This condition allows 1992/66to be identified as a boselaphine. Most probablythis boselaphine is Miotragocerus pannoniae, butfor the present it is better to label it only asMiotragocerus sp.
Fourteen tooth specimens belong to a Bovidaespecies smaller than the middle Miocene taxonTethytragus koehleri of Pașalar or Eotragus clavatus(Gervais 1850 - in Gervais 1848-1852) of westernEurope. The teeth are slightly hypsodont.
With a tragulid, two cervoids and two bovids,the ruminant fauna has a composition familiar incentral European localities of late middle or lateMiocene age. No giraffid material has yet been dis-covered at Rudabánya. The strength of the cervoidrepresentation and the presence of Dorcatheriummust indicate a less open, more moist or moretemperate habitat than in the classical Turolianfaunas further to the south and east in Europe andneighbouring parts of Asia. Compared with pre-sent-day Muntiacus, Micromeryx had longer legsand longer distal elements in relation to humerusand femur, but it might still have lived in habitatswith thickets if the African neotragine antelopeNeotragus moschatus (also with longer legs and rel-atively longer distal elements than Muntiacus)were a satisfactory modern analogue (Kingdon,1982:194). Köhler (1993:51) suggested closedrather than open habitats for Micromeryx.
The teeth of the small bovid agree with lateMiocene species of Gazella, but their generalisedmorphology and degree of hypsodonty do not allowa safe attribution even to the Antilopini. Thisdiminishes any need to infer an open habitat.
Every clue given by the ruminants about thetemporal correlation of Rudabánya suggests anearly late Miocene and even an early Vallesiandate. The Dorcatherium supports a late Mioceneage. The Lucentia has antlers with long pediclesand only one bifurcation. It is rare to find fossilantlers with preserved second bifurcations beforeMN12, although earlier incomplete antlers mayhave been referred to species or genera known tohave second bifurcations at later time levels.Species lacking a second bifurcation persistedinto the Pliocene, although pedicles did becomeshorter. Micromeryx flourensianus mainly occursin the middle Miocene, but is known to have sur-vived into the Vallesian of Spain. On what isknown of it, the Miotragocerus would fit a pre-Turolian date.

21RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
V) AGE AND ZOOGEOGRAPHY
FAUNAL LIST AND CORRELATIONS (R.L. Bernor, L.Kordos, P. Müller and P. Renne)
Rudabánya II has yielded an extraordinarilydiverse vertebrate fauna: 112 species (re: Table 3)including 1 species of fish, 18 species of amphib-ians, 13 species of reptiles, 11 species of birds and69 species of mammals. Our attempts to directlydate the locality using magnetostratigraphy failedto yield a discernable magnetic signal. Yet, we canconstrain the lower limit using the stratigraphicallydistantly underlying radioisotopic determination ofthe “Upper Sarmatian Tuff” which has been datedby the potassium-argon method as being 11.5 Ma ±0.5 Ma (Balogh, 1984) and 11.4 Ma ± 0.1 m.y. bysingle crystal argon (P.Renne, unpublished).
Rudabánya’s age depends on biochronologiccorrelations. The fauna includes an interestingmixture of archaic holdover taxa alongside MN 9immigrants. The retention of archaic taxa in thefauna is clearly due to the subtropical forest condi-tions nurtured by warm equable lakeside climates.
The fish, amphibian, reptile and avian faunasare of little use for correlation. The exceptions arethe proteid amphibian Mioproteus sp. which firstoccurs in Central Europe during the late Miocene(Rudabánya likely being an early occurrence),Typhlops sp. that last occurs in MN 12?, and Najaromani which last occurs in MN 11.
The insectivore taxa include Galerix sp.,Plesiosorex aff. styriacus and Talpa sp. which arearchaic and have very long chronologic ranges.Lanthanotherium sanmigueli, Postpalerinaceus sp.,Crusafontia aff. endemica and Archeodesmana n. sp.are advanced forms that support an MN 9 correla-tion. The erinaceid Postpalerinaceus n. sp. does nototherwise have a first occurrence earlier than MN10, and suggests the possibility of a later MN 9 cor-relation.
The rodents include a number of taxa that sup-port an MN 9 age, including: Democricetodon n.sp., Microtocricetus molassicus, Kowalskia cf.schaubi, Albanensia grimmi, Miopetaurista sp.,Eomyops catalaunicus, Trogonotherium minutumand Anomalomys rudabanyensis and Myoglisucrainicus. The rodents further biochronologicallycharacterize the Rudabánya fauna as follows: 1)mainly middle Miocene rodents (Eumyarion latior,Spermophilinus bredai, Paraglirulus werenfelsi,Myoglis meini)associated with 2) late Miocene firstappearing taxa (Microtocricetus molassicus ,Albanensia grimmi, Glirulus lissiensis, Muscardinusaff. vallesiensis, Myoglis ucrainicus, Kowalskia cf.schaubi) but without 3) Progonomys, Parapodemus,modern Kowalskia-species and Pliopetaurista . Theassemblage is characteristic for an early Vallesian(MN9) age. It should be further remarked that thelack of Megacricetodon in this assemblage furthersupports Rudabánya’s MN 9 age. The most signifi-cant change in Central Paratethys rodent faunastook place in MN10, a period when middleMiocene species were almost completely replaced(Daxner-Höck, 1996: 6-7).
The carnivore fauna has many taxa that firstoccur in the early and middle Miocene of Central
and Western Europe. Taxa that would appear to belimited to a MN 9 distribution include the viverridSemigenetta grandis and the large new species ofAmphicyon. Many of these archaic taxa carry overinto the Turolian making them unuseful for corre-lation.
The cooccurrence of Deinotherium giganteumand Tetralophodon cf. longirostris is characteristicfor MN 9 in Central Europe. The equid,Hippotherium intrans Kretzoi is an advanced mem-ber of the Hippotherium lineage (re: Bernor et al.,1993a; Bernor et al., 1996; Bernor & Armour-Chelu,1999) correlative with Pannonian F, upper MN 9,ca. 10-9.7 Ma. Hippotherium intrans is perhaps themost biochronologically discriminating species inthe assemblage. The rare occurrence ofHippotherium small sp. from the Upper Lignitezone may herald a younger age for that horizonthan the remainder of the Rudabánya II fauna. Thetapir, Tapirus cf. priscus, and chalicothere,Chalicotherium aff. goldfussi, are a characteristicassociation for MN 9.
The rhinoceroses Hoploaceratherium belved-erense and Aceratherium incisivum are also MN 9biochrons. Aceratherium incisivum is an MN 9immigrant into Central Europe;.Lartetotherium aff.sansaniensis is an archaic form and its occurrenceat Rudabánya is believed to be the latest known inEurope.
Of the Artiodactyla, the suids are the most use-ful for biochronologic correlations. The suine,Propotamochoerus palaeochoerus, occurs in CentralEurope during MN 8 and MN 9; it has a very late(and very rare) occurrence at Maramena Greeceduring MN13/14 (Hellmund, 1995). The tetracon-odont Parachleuastochoerus n. sp. is known onlyfrom Rudabánya, and its closest related forms arefrom MN 8 and MN 9 of Spain. The ruminantsmostly have extended geochronologic ranges. Thecervid Lucentia aff. pierensis has its earliest knownoccurrence at Rudabánya, and the bovidMiotragocerus sp. (similar to M. pannoniae) is typi-cal for MN 9 in the Central Paratethys.
A synthesis of the biochronologic bases forRudabánya II’s age strongly supports a late MN 9,Pannonian F correlation, ca. 10-9.7 Ma.
PALAEOZOOGEOGRAPHY OF THE RUDABÁNYAFAUNA (R.L. Bernor and L. Rook)
The Rudabánya fauna accumulated in late MN9, near the shoreline of the Pannonian lake. Thefauna is now known to have a unique combinationof primitive early and middle Miocene vertebratesmixed with relatively few first occurring MN 9taxa.
Text-fig. 5 is a Genus Faunal Resemblance Index(GFRI’s) histogram between Rudabánya and 15other European, west Asian and African lateMiddle Miocene – Late Miocene vertebrate locali-ties. The dataset used for this study was derivedfrom a data file downloaded from the NOW data-base (M. Fortelius, coordinator), available online atthe website: www.helsinki.fi/science/now/. As inBernor et al. (2001) we calculate both Dice andSimpson GFRI’s. Dice’s index is the one most com-monly used (neo-zoological) faunal resemblance

22 R.L. BERNOR - L. KORDOS - L. ROOK
TABLE 3 - Rudabánya vertebrate faunal list
VERTEBRATE TAXA FAD LAD ECOLOGYPisces Pices
Pices indet.Amphibia
CaudataProteidae
Mioproteus caucasicus MN5 MN16 lacustrine environmentsSalamandridae
Chelotriton sp., type I MP12 MN9 lacustrine environmentsChelotriton sp., type IISalamandridae new form MN 4 MN9 lacustrine or wet environmentsSalamandridae indet.Triturus sp. L. Mio. Recent lacustrine environmentscf. Triturus sp.
AnuraDiscoglossidae
Latonia gigantea MN4 MN15 wet environmentsBombina sp. MN1 Recent wet environmentscf. Discoglossus sp. ? MP30 RecentDiscoglossidae inc. sedis Recent wet environmentscf. Discoglossidae
PalaeobatrachidaePalaeobatrachus sp. ? MP6 MN16 lacustrine environmentscf. Palaeobatrachus sp.
PelobatidaePelobatidae indet. MP7 Recent possibly sandy environments
PelodytidaePelodytes sp. MP16 Recent moderately wet environments
HylidaeHyla sp. MN4 Recent
Ranidae Rana sp. ? MP17 Recent wet environments
ReptiliaTrionyx sp.Geomyda sp.Testudo cf. kalksburgensisTestudo sp. ITestudo sp. IIScolecophidia indet.
TyphlophidaeTyphlops sp. ? MN12? tiny burrowing snake
?Leptotyphlopidae gen. and sp. indet.Colubridae
Natrix sp.Colubridae Indet. AColubridae Indet. B
ElapidaeNaja romani MN4 MN11 cobraSerpentes indet.
AvesAnatidae
Anas aff. velox aquatic [duck]Anas sp. aquatic [duck]
PhasianidaeMiophasianus aff. medius bird fauna typical of lake forest envirnomentPalaeortyx aff. grievensis bird fauna typical of lake forest envirnomentPalaeortyx cf. phasianoides/ bird fauna typical of lake forest envirnoment
intermediusRallidae
?Miorallus sp. bird fauna typical of lake forest envirnomentStrigidae
Strix cf. brevis bird fauna typical of lake forest envirnomentAthene sp. bird fauna typical of lake forest envirnoment
SylviidaeAcrocephalus sp. bird fauna typical of lake forest envirnomentLocustella sp. bird fauna typical of lake forest envirnoment
CorvidaeCorvus sp. bird fauna typical of lake forest envirnoment
MammaliaInsectivora generally idicative of water and forestErinaceidae
Lanthanotherium sanmigueli MN 9 MN 11 compares with SE Asian tropical faunas

23RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
Galerix sp. compares with SE Asian tropical faunasPostpalerinaceus n. sp. MN 9 MN 9
MetacodontidaePlesiosorex aff. styriacus MN 4
SoricidaeDinosorex n. sp. MN 7/8 MN 9Crusafontia aff. endemica MN9 MN9 extant ecomorph in montain forests of SE AsiaPaenelimnoceus aff. repenningi MN 6 MN 10
DimylidaeMetacordylodon aff. schlosseri MN 5 MN 9 likely semiaquaticPlesiodimylus chantrei MN4 MN 11
TalpidaeArcheodesmana n. sp. MN 9 MN 9 certainly semiaquaticTalpa sp. MN 2 Recent
ChiropteraVespertilionidae
Eptesicus campanensis MN 6 no "true" tropical forms, but tropical marshy woodsEptesicus n. sp. MN 9 no "true" tropical forms, but tropical marshy woodscf. Miotrellus risgoviensis MN 6 no "true" tropical forms, but tropical marshy woods
Primates?Pliopithecidae
Anapithecus hernyaki MN 7/8 MN 9 subtropical forest, arboreal, suspensory, ate soft fruitsHominidae
Dryopithecus brancoi MN 7/8 MN 9 subtropical forest, arboreal, suspensory, ate soft fruitsRodentiaCricetidae
Eumyarion aff. latior MN 4 MN 9 ground of the forest-open country (elevations)Democricetodon n. sp. MN 9 MN 9 ground of the forest-open country (elevations)Microtocricetus molassicus MN 9 MN 11 riaprian environment; folivor (reed vegetation)Kowalskia cf. schaubi MN 9 MN 9 ground of the forest-open country (elevations)
SciuridaeAlbanensia grimmi MN 9 MN 9 arboricol, frugivor, canopy of the forestMiopetaurista sp. MN 7/8 MN 9 arboricol, frugivor, canopy of the forestHylopetes sp. MN 4 present arboricol, frugivor, canopy of the forestBlackia cf. miocaenica MN 1 MN 11 arboricol, frugivor, canopy of the forestSpermophilinus bredai MN 5 MN 10 probably fossorial, soft ground of the forest
GliridaeParaglirus werenfelsi MN 7/8 MN 9 arboricol, frugivor/granivor, canopy of the forestGlis sp. (cf. minor Kowalski) MN 9 MN 9 arboricol, frugivor/granivor, canopy of the forestMyoglis ucrainicus MN 9 MN 9 arboricol, frugivor/granivor, canopy of the forestMuscardinus hispanicus MN 7/8 MN 10 arboricol, frugivor/granivor, canopy of the forestMuscardinus aff. vallesiensis MN 9 MN 9 arboricol, frugivor/granivor, canopy of the forestGlirulus lissiensis MN 4 MN 14 arboricol, frugivor/granivor, canopy of the forest
EomyidaeEomyops catalaunicus MN 9 MN 12 arboricol, frugivor/granivor, canopy of the forest
CastoridaeTrogonotherium minutum MN 4 L. Plio. aquatic
AnomalomyidaeAnomalomys rudabanyensis MN 9 MN 9 probably fossorial, soft ground of the forest
LagomorphaOchotonidae
Eurolagus sp. MN 5 MN 10CarnivoraViverridae
Semigenetta grandis MN 9 MN 9 hypercarnivorous, subtropical forestMustelidae
Taxodon cf. sansaniensis MN 5 MN 9 terrestrial, omnivoreMelinae gen. et sp. indet. terrestrial, omnivoreProputorius sp. MN 5 MN 12 semiaquatic, terrestrial, invertebrate-eaterMartes cf. filholi terrestrial cf. Trochitis sp.Paralutra jaegeri MN 5 MN 9 aquaticParalutra n. sp. aquatic
UrsidaeUrsavus primaevus MN 6 MN 11 terrestrial, omnivore, predom. meat-eaterUrsavus brevirhinus MN 4 MN 11 terrestrial, omnivore, predom. meat-eater
ProcyonidaeAlopecocyon sp. MN 4 MN 13 terrestrial, omnivoreSimocyon diaphorus MN 9 MN 9 terrestrial, carnivore, meat and bone eater
AmphicyonidaeAmphicyon n. sp. MN9 terrestrial, omnivore/scavenger
NimravidaeSansanosmilus jourdani MN 6 MN 9 terrestrial, carnivore, meat-only

24 R.L. BERNOR - L. KORDOS - L. ROOK
indices, while Simpson’s index has long been usedby paleontologists and additionally adjusts for dif-ferences in sample sizes, which is an issue in thisanalysis.
The Rudabánya fauna exhibits its strongestresemblance with the Spanish MN 9 localities ofCan Llobaters and Can Ponsic I. Its next strongestresemblance is with the late MN 8 locality ofCastell de Barberá (Spain), the MN 9 locality ofEppelsheim (Germany) and the MN 11 locality ofDorn Dürkheim (Germany). The strong resem-blance of this suite of latest MN 8 – MN 11 westernand central European localities suggests a strongprovinciality for this geographic region (Bernor,1978, 1983, 1984). In contrast, this set of centraland western European faunas has relatively lowresemblance to the remaining 10 localities. A strik-ing contrast is revealed between Rudabánya andthe Spanish and German MN 8-MN 9 localities onone hand, and the Sinap Hominid Zone (HZ) onthe other. The Sinap Hominid Zone (Sinap HZ)fauna of Anatolia is closely similar in age to theRudabánya fauna, but is 100% different from it atthe genus level. The closely equivalent-agedMacedonian localities yelding the fossil ApeOuranopithecus (Ouranopithecus Faunal Zone) (re:Greece below) are likewise strongly dissimilar toRudabánya in its faunal content. The remainder ofthe faunal sample has little similarity at the genuslevel to Rudabánya: the Oreopithecus Fauna Zone
(OFZ in Bernor et al., 2001) faunas were highlyendemic during the MN11-12 interval; Baynunah,Lothagam, Sahabi, Venta del Moro and Italian MN13 localities are substantially younger and the lat-ter reflect the strong faunal turnover that occurredin Europe during the Turolian age, and mostspecifically the medial (MN12) and late (MN13)Turolian. Interestingly, Pikermi (MN 11/12) andMaramena (MN 13) have slightly elevated GFRI’sin comparison to Rudabánya, probably because oftheir more forested character.
Text-fig. 6 presents a pie-diagram of the mam-malian family composition of the Rudabánya, CanLlobateres, Sinap HZ and Ouranopithecus FaunalZone assemblages (Ouranopithecus FZ): the firstthree sites are late MN 9 age, while the last is MN10 age. Rudabánya and Can Llobateres are similarin their higher diversity of mammalian families(Rudabánya with 34 families and Can Llobatereswith 36 families), while the Sinap HZ (22 mam-malian families) and Ouranopithecus FZ (13 fami-lies) have much reduced faunas. Faunal composi-tion at Rudabánya and Can Llobateres suggests thepresence of warm equable forested environments,and in respect to the presence of: Amphicyonidae,Castoridae, Cervidae, Chalicotheriidae, Deino-theriidae, Erinaceidae, Eomyidae, Gliridae,Hominidae, diverse Mustelidae, ?Pliopithecidae(Anapithecus hernyaki at Rudabánya only), lowcrowned Rhinocerotidae, Sciuridae, Soricidae,
FelidaePseudaelurus turnauensis MN 4 MN 11 terrestrial, carnivore, meat-onlyPseudaelurus lorteti MN 4 MN 9 terrestrial, carnivore, meat-only
Hyaenidaecf. Thalassictis montadai MN 7/8 MN 9 terrestrial open country, carnivore, meat and bone eater
Proboscidea all brachydont; subtropical woodland environmentsDeinotheriidae
Deinotherium giganteum MN 6 L. Plio. brachydont browserGomphotheriidae
Tetralophodon longirostris MN 7/8 L. Plio. brachydont browser/grazerTetralophodon cf. longirostris MN 7/8 L. Plio. brachydont browser/grazer"Stegotetrabelodon" gigantorostris MN 9 brachydont browser/(grazer?)
PerissodactylaEquidae
Hippotherium intrans MN 9 running form, mixed graze/browse, deer ecomorphHippotherium small sp. MN 9 likely more grass eating than H. intrans
TapiridaeTapirus cf. priscus MN 9 MN 11 brachydont, browser, subtropical woodland environments
ChalicotheriidaeChalicotherium aff. goldfussi MN 9 MN 12 brachydont, browser, subtropical woodland environments
Rhinocerotidae All brachydont; subtropical woodland environmentsHoploaceratherium belvederense MN 9 MN 10 brachydont browserAceratherium incisivum MN 6 L. Plio. brachydont browser; wide habitat toleranceLartetotherium aff. sansaniensis MN 4 MN 9 brachydont browser
ArtiodactylaSuidae
Propotamochoerus palaeochoerus MN 7/8 MN 14 omnivoreParachleuastochoerus n. sp. MN9 ? Hard object frugivore, omnivory
TragulidaeDorcatherium naui MN 4 MN 13 browser, subtropical woodland environments
CervidaeLucentia aff. pierensis MN 9 MN 11 browser, subtropical woodland environments
MoschidaeMicromeryx flourensianus MN 2 MN 11 browser, subtropical woodland environments
BovidaeMiotragocerus sp. MN9 MN 15 browser, subtropical woodland environmentsBovidae, smaller sp.

25RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
Talpidae, Tapiridae, Tragulidae, primitive Ursidaeand Viverridae. The Sinap HZ and OuranopithecusFZ have an early “Pikermian type” open countrywoodland chronofauna (re: Bernor, 1983, 1984;Bernor et al., 1996; Solounias et al., 1999; Forteliuset al., in press), characterized by a strong reductionof Vallesian forest elements and enhancement ofdiverse Bovidae, Felidae, Giraffidae, Hyaenidaeand Percrocutidae.
VI) PALAEOCLIMATE ESTIMATES FROMBIOTIC PROXIES
(J. Damuth, J.A. van Dam and T. Utescher)
Quantitative reconstruction of past climaticconditions based on faunal and floral associationsis still fairly imprecise under our current state ofknowledge. However, it is possible to provide forRudabánya independent estimates of mean annualprecipitation from large mammals, small mam-mals, and fossil plant remains, respectively. Fossilfloras also permit reconstruction of a wider rangeof climatic parameters. The estimates we are ableto offer are consistent with each other and with theinterpretation of forested subtropical habitats atRudabánya. Note that the reconstructed rainfallvalues are estimates of the realized amount ofmoisture experienced by the plant and animalcommunities in one specific locality. Rudabánya
probably represents an unusually wet local envi-ronment as a result of its physical setting, so thesepalaeoprecipitation estimates do not necessarilycorrespond to average regional values of true pre-cipitation.
The mean value of ungulate herbivore tooth-crown height (hypsodonty) can be used to estimatemean annual precipitation (e.g. Fortelius et al.,2002), and gives a value for Rudabánya of 1190mm/yr. Unfortunately, there is much scatter in theregression (95% confidence interval is 250–5070mm). In addition, when virtually all species arebrachydont (as at Rudabánya) the mean hypsodon-ty statistic is at a minimum and cannot furtherresolve differences in precipitation. An alternatemethod (PMH; Damuth et al., 2002) does not havethis statistical limitation and shows a stronger rela-tionship, but the PMH value depends upon bothmean hypsodonty and species richness, whichintroduces a complication. Species richness forMiocene mammalian faunas of Rudabánya’s ageand older is often considerably higher than in mod-ern faunas from comparable habitats (Janis et al.,2000, 2002); with 15 ungulate browsers Rudabányais unusually rich. This currently poorly-understoodphenomenon causes PMH to yield, for midMiocene and earlier faunas in North America,unrealistic estimates that are about twice thosethat are estimated from contemporaneous fossilfloras. PMH estimates rainfall for Rudabánya as
Text-fig. 5 - Genus Faunal Resemblance Index (GFRI’s) between Rudabánya and 15 other European, West Asian and African late MiddleMiocene – late Miocene vertebrate localities (re: Bernor et al., 2001 for further information on methods and dataset).
26 R.L. BERNOR - L. KORDOS - L. ROOK
1800mm/yr. Using the North American data for arough calibration, this would represent the expect-ed approximate doubling of the proper estimatecaused by the high species richness. Thus, thelarge-mammal data suggest a most probable rangeof 900–1200 mm in annual precipitation forRudabánya, although there is still considerableuncertainty about this estimate.
A new method for estimating past rainfall levelson the basis of small mammal community struc-ture has recently been developed and applied to theNeogene of Europe (van Dam, 2001; van Dam, tobe submitted). The method involves a multipleregression equation based on recent small mam-mal communities using the proportions of arborealand insectivorous species as independent variables.The size of the 95% confidence intervals for thepredicted mean annual rainfall levels in the indi-vidual localities ranges between 350 and 400mm/year (one-sided). In addition to mean levels,the amount of rainfall in the driest month could bebest estimated using the proportion of arborealspecies only. The confidence intervals around thisestimate average about ± 20 mm.
In Rudabánya both the percentages arborealand insectivorous species are very high: 41% and34%, respectively (12 and 10 out of 29, with aerial,aquatic, and semiaquatic forms excluded). This
results in a mean annual precipitation 1235 mm/yr,which is the highest value for the entire data set ofalmost 250 Neogene small mammal faunas stud-ied. The small mammal results support the exis-tence of an important regional component in theRudabánya precipitation regime, because time-equivalent small mammal faunas generally showhigh values as well: Götzendorf (Austria), Jujurieux(France), Grtsev (Ukraine), and Can Llobateres(NE Spain) all have levels between 1000 and 1200mm/year. The estimated value for precipitation inthe driest month is 84 mm. This value might beslightly too high, although the large numbers ofarboreal small mammals certainly indicate thepresence of forest and the absence of a significantdry season.
These results are supported by the analyses ofthe paleobotanical record (Kretzoi et al., 1974)using the coexistence approach (Mosbrugger andUtescher, 1997). The method employs tolerances ofthe Nearest Living Relatives known for a fossilflora with respect to various climate parameters todetermine intervals in which most of the taxa cancoexist. The significance of the results obtainedhighly depends on the taxa diversity of the floraanalyzed. In case of Rudabánya, 30 extant planttaxa contribute with climate data. With maximumpercentages of coexisting taxa between 86.7 and
Text-fig. 6 - Pie-diagram of the mammalian family composition of the following assemblages: a) Rudabánya; b) Can Llobateres; c) SinapHominid Zone; d) Ouranopithecus Faunal Zone.

27RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
100% (mean: 92.2%) the resulting climate intervalsare considered as reliable. The climate data aresummarized in table 4. The mean annual tempera-ture ranges between 15.6 and 15.7°C, with a warmmonth mean temperature of around 26°C, and acold month mean temperature of ca. 5°C. Withmean annual precipitation rates between 897 and1297 mm and a mean annual range of precipitationbetween 86 and 89 mm, a moderate seasonality inprecipitation is obvious from the data. Additionally,wet climate conditions during the warm season areindicated by mean precipitation rates of thewarmest month between 83 and 89 mm.Summarizing up these results, the Rudabánya floragrew in a warm temperate, permanently humid cli-mate with warm summers and mild winters (CfaKöppen-type climate; Köppen, 1931).
As stated above, the Rudabánya flora mostprobably existed in a very wet habitat. This is alsoobvious from the floral composition. Approxi-mately 60% of the extant reference taxa of the fos-sil assemblage presently occur in the alluvial wet-land vegetation, additionally, numerous waterplants have been recorded. Thus, precipitationpotentially was not as important as limiting factorfor the plant community, a fact that could bias thereconstruction of precipitation data using the coex-istence approach. However, the mean annual pre-cipitation calculated for the Rudabánya flora isconsistent with data obtained from neighboring,about time-equivalent localities. The precipitationmap for the circum-alpine realm in an early LateMiocene time slice based on the analyses of 28megafloras shows rates of over 1000 mm all overthe Eastern Paratethys (Bruch et al., in press).
VII) PALEOECOLOGY(R.L. Bernor, M. Fortelius and L. Rook)
The Rudabánya fauna has a subtropical aspectindicative of equable climates (Tables 3, 4). Basingon Kretzoi et al. (1976) data, Damuth et al. (thispaper) characterized the Rudabánya flora as typi-cal of warm temperate, permanently humid cli-mate with warm summers and mild winters.
Andrews et al. (1997) stressed close communityecology comparisons between Rudabánya and theearly middle Miocene (MN 6) localities of Pașalar(Turkey) and Sansan (France). In support of theirpaleoecologic characterization, Andrews et al.(1997) cite the common occurrence of semiarbore-al species, abundant browsing herbivores and com-mon frugivores and insectivores. As stated above,the Rudabánya fauna includes an interesting mix-ture of archaic European taxa commingled withbroader Eurasian immigrant taxa. Table 3 summa-rizes this research group’s current understandingof the age and ecology of the fauna, which we sum-marize further below.
The Amphibia all suggest the existence of per-sistent wet conditions at Rudabánya. The snakeshave a distinctly subtropical character highlightedby the occurrence of a cobra (Naja romani) andtaxa all of which became extinct in Central Europebefore the end of the Miocene. The avian faunaincludes many aquatic forms and is typical of a
lake-forest environment.The mammals are particularly useful for paleoe-
cologic reconstruction. Within the insectivores arefound strong indications of subtropical environ-ments such as those that exist today in S.E. Asia.Specifically, the erinaceids Lanthanotherium san-migueli and Galerix sp. closely comparable withinsectivores living in S.E. Asia today. The soricidCrusafontia aff. endemica is most like extantAnurosorex squamipes living today in montaneforests of S.E. Asia. The dimylid insectivoreMetacordylodon aff. schlosseri was likely semiaquat-ic, while the talpid Archeodesmana n. sp. was cer-tainly semiaquatic. Clearly, the Rudabánya paleo-lake margin afforded year round warm andequable conditions to maintain this diverse insecti-vore fauna.
The rodent fauna has a number of species thatclearly persist from the middle Miocene of centraland western Europe. The diverse sciruids, gliridsand eomyids suggest forested conditions. The cas-torid, Trogonotherium minutum was an obligatoryaquatic form.
The Carnivora include several lineages that con-tinue from earlier Miocene times of central andwestern Europe, and include: Semigenetta ,Proputorius, Ursavus (U. primaevus and U. brevirhi-nis), Amphicyon, Sansanosmius and Pseudailurus(P. turuensis and P. lorteti). The high diversity ofmustelids, with which cooccur a viverrid, primitiveursids, nimravid and felids are characteristic ofsubtropical environments typical of western andcentral European middle Miocene localities. Thediversity of smaller body size Carnivora, especiallymustelids, is also typical of extant tropical environ-ments. The low diversity of hyaenids is atypical forlate Miocene Eurasian faunas and is yet anotherindication of the relatively closed forested condi-tions that prevailed at Rudabánya.
The ungulates further support the interpretationof subtropical forests at Rudabánya. The pro-boscidean species were all browsers. Deinotheriumgiganteum and Tetralophodon cf. longirostris consti-tute a characteristic probocidean element for MN 9of central Europe.
TABLE 4 - Rudabánya paleoclimatic values.
N taxa with climate data 30,0MAT_L [°C] 15,6MAT_R [°C] 15,7MAT_% coexisting taxa 93,3CMM_L [°C] 5,0CMM_R [°C] 5,1CMM_% coexisting taxa 90,0WMM_L [°C] 25,6WMM_R [°C] 26,3WMM_% coexisting taxa 90,0MAP_L [mm] 897,0MAP_R [mm] 1297,0MAP_% coexisting taxa 93,3MMPWM_L [mm] 83,0MMPWM_R [mm] 89,0MMPWM_% coexiting taxa 100,0MARP_L [mm] 86,0MARP_R [mm] 89,0MARP_% coexisting taxa 86,7

28 R.L. BERNOR - L. KORDOS - L. ROOK
The analysis of Hippotherium primigenium veri-fies its mixed browse/graze dietary behavior. Thistaxon, along with the smaller hipparion and thehyaenid cf. Thalassictis montadai support thehypothesis that more open country mesophytichabitats were found distal to the lake margin suchas has been characterized for MN 9 of the ViennaBasin (Bernor et al., 1988). The remaining perisso-dactyls – Tapirus cf. priscus, Chalicotherium aff.goldfussi , Hoploaceratherium belvederense,Aceratherium incisivum and Lartetotherium aff.sansaniensis – all were brachydont browsers andare characteristic taxa of central European sub-tropical/ warm temperate Miocene faunas.
The suids Propotamochoeurs palaeochoerus andParachleuastochoerus are also characteristic of cen-tral and western European MN 8-9 faunas; the for-mer was an omnivore and the latter likely a mixedhard-object frugivore and omnivore. Neither ofthese suids have been found in the open countryPikermian faunas that characterize the laterMiocene of Hungary.
The ruminants are not diverse, but include aspecies of tragulid, a cervid, a moschid and twobovids. All ruminant species were browsers andcharacteristically are found in central Europeansubtropical forest settings.
The prevailing habitat type at Rudabánya wassubtropical forest that prevailed under equable cli-matic conditions. If seasonality existed, it was mildand experienced summer rainfall. It is evident,however, that some taxa were adapted to moreopen country mixed woodland and grassland.Hippotherium intrans was a mixed feeder (grassand browse). The proboscideans, rhinos andhyaenid also probably inhabited more open wood-lands distal to the lake margin forests.
Bernor et al. (2001) presented a simple contrastof several late middle Miocene – latest Miocenelocalities from the standpoint of ungulate crownheight. In Text-fig. 7 we provide a contrast betweenRudabánya, Can Llobateres, Sinap HZ andOuranopithecus FZ ungulate crown heights. The 3-part subdivision: brachydont whereby M2 crownlength is greater than its crown height; mesodontwhereby M2 crown length is roughly the same ascrown height; hypsodont whereby M2 crownheight is 2X + that of crown length.
This analysis reveals that Rudabánya and CanLlobateres are similar in their predominance ofbrachydont forms and low percentage of hyp-sodont forms. In fact, the only hypsodont taxa atRudabánya are the two hipparion species; a small-er form known from one or two specimens, and theremainder of the collection being referable toHippotherium intrans. As demonstrated by Bernoret al. (in press), Hippotherium intrans was a mixedfeeder with a substantial amount of browse in itsdiet. The Sinap HZ and Ouranopithecus FZ show adramatic drop in the percentage of low-crownedungulates. Both have a substantially larger percent-age of hypsodont and mesodont forms in their fau-nas than either Rudabánya or Can Llobateres: 57%hypsodont and mesodont forms at Sinap HZ and51% forms in the Ouranopithecus FZ (Text-fig. 7).
VIII) LATE MIOCENE EUROPEAN HOMINIDEVOLUTION
LATE MIOCENE PERIMEDITERRANEAN HOMINID(s.l.) EVOLUTION
The hominid fossil record of Spain (J. Agustí and M.Llenas)
The late Aragonian-Vallesian record of higherprimates in the Vallès-Penedès Basin is one of themost abundant and complete from westernEurasia. The first anthropoid record in the Vallès-Penedès Basin occurs at Sant Quirze, in levels oflate Aragonian age (Megacricetodon ibericus Zoneof Agustí & Moyá-Solá, 1991), well before thebegining of the Vallesian (Garcés et al., 1997). Thefaunal assemblage from Sant Quirze is very similarto that of other late Aragonian European localitiessuch as La Grive M and includes the cervidStehlinoceros elegantulus (Azanza & Menéndez,1990). The presence of the flying squirrel and thelarge castorids indicates the existence of humid,forest conditions during this time (Mein, 1970,Aldana, 1992). Castell de Barberà is similar to SantQuirze in its abundance of wet, forest indicatorssuch as tragulids, castorids, flying-squirrels anddormice. This humid character is similar to thatfound in some early Vallesian localities such as CanLlobateres. In contrast, the late Astaracian localityof Lower Hostalets has a more open character. Ingeneral, and despite the presence of Dryopithecus,the latest Aragonian levels are characterized by theabsence of forest or humid climate indicators, suchas castorids or flying-squirrels, and by low diversi-ties of dormice rodents.
The beginning of the Early Vallesian in theVallès-Penedès (Megacricetodon ibericus-HipparionZone of Agustí & Moyá-Solá , 1991) is character-ized by the presence of the first Hippotherium rep-resentatives (Hippotherium primigenium catalan-icum), but the rest of the association is composedof the same elements as the latest Aragonian witheven more accentuated dry conditions. No homi-noid remains have yet been found in these levels.In contrast, localities of late early Vallesian ageincluded in the Cricetulodon Zone (Can Ponsic,Santiga, Can Llobateres 1) are characterized by avery different faunal association, and againincludes large castorids, flying-squirrels, tragulids,and tapirids. After an earliest Vallesian “gap”, thehominoid record suddenly increases in the lateearly Vallesian with the occurrence of Dryopithecusin the localities of Can Ponsic and Can Llobateres1. These faunal assemblages reflect a recovery ofmore forested, humid conditions comparable withthose present in the late middle and early lateAragonian. This is also the time when hominoids(Dryopithecus laietanus) attain their highest levelsof representation. The boundary between the earlyand late Vallesian is clearly defined in the Vallès-Penedès Basin by the “Mid-Vallesian Crisis” (Agustí& Moyá-Solá, 1990). This event involved the sud-den disappearence of most of the humid elementscharacterizing early Vallesian localities such as CanLlobateres 1 (rhinocerotids, suids, cervids) andreplacement by eastern immigrants (dry-adaptedsuids like Schizochoerus, large sivatherine giraffids

29RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
and boselaphine bovids). Initially, this extinctionevent did not affect the occurrence of Dryopithecus.Rare, but well preserved finds of Dryopithecus stilloccur in fact at the beginning of the late Vallesian(MN 10), just after the Mid-Vallesian Crisis (Moyá-Solá & Köhler, 1993, 1996; Agustí et al.,1996), but itis absent from younger deposits. The environmen-tal change involved in the Mid-Vallesian Crisisseemed to strongly affect Dryopithecus, but afterthe exit of these hominoids, the pliopithecines,absent from the rich early Vallesian localities suchas Can Ponsic 1 and Can Llobateres 1, reappearedagain shortly before final extinction.
The hominid fossil record of Greece (L. de Bonis andG. Koufos)
There are very few Greek Vallesian macromam-mal faunas. An exception is the suite of macro-mammal localities in central Macedonia (AxiosValley and Chalkidiki), where the hominidOuranopithecus macedoniensis is found. These fau-nas have a macromammal fauna very differentfrom those of central and western Europe, andexhisted under more seasonal, open country envi-ronmental conditions. The localities of the lowerAxios Valley are found in the Nea MessimvriaFormation which includes the lower part of thelate Miocene succession (Koufos, 1980; de Bonis etal., 1988, 1998; de Bonis & Koufos, 1999). Recentmagnetostratigraphic studies of the Axios Valleydeposits suggest an age of ~9.6 Ma for the
Xirochori-1 fauna and ~9.3 Ma for faunas of Ravinde la Pluie and Ravin des Zouaves-1 (Sen et al.,2000). Ravin de la Pluie, Ravin des Zouaves-1,Xirochori-1 and Pentalophos-1 includes faunal ele-ments different from those of the typical Vallesianof NW Europe, making biochronological compar-isons difficult. Nevertheless, such fauna is best cor-related with the late Vallesian, MN 10 (de Bonis etal., 1990; de Bonis & Koufos, 1999). A possiblyyounger locality is that of Nikiti 1, in the area ofthe Chalkidiki Peninsula (Koufos et al., 1991). Thissite has yielded remains of Ouranopithecus mace-doniensis (Koufos, 1993, 1995) in association witha fauna showing similarities with those of AxiosValley, but with some taxa, namely Microstonyx ery-manthius, believed to be a local first occurrence.Although Nikiti was originally attributed to theearly Turolian (MN 11; Koufos, 1993), at present itis interpreted by us as representing a latestVallesian assemblage (de Bonis & Koufos, 1999).Using all biochronological data for Nikiti-1, thecomparison of that fauna with those of the Axiosvalley (Ravin de la Pluie and Ravin des Zouaves-5),and their known magnetostratigraphic ages, sug-gests a latest Vallesian age attribution or an agebetween about 9.3-8.7 Ma for Nikiti-1 (Sen et al.,2000; Koufos, 2000). Ouranopithecus macedonien-sis appears to have out-survived western and cen-tral European Dryopithecus species. Ouranopi-thecus macedoniensis is currently understood notto have occurred in any Greek localities of Turolian
Text-fig. 7 - Ungulate crown height contrast between a) Rudabánya; b) Can Llobateres; c) Sinap Hominid Zone; d) OuranopithecusFaunal Zone. Brachidont, Mesodont, Hypsodont.

30 R.L. BERNOR - L. KORDOS - L. ROOK
age, among which only the colobine Mesopithecusis found.
The hominid fossil record of Turkey (Sinap) (R.L.Bernor and M. Fortelius)
At Sinap (Turkey), recent field work has yieldeda precise magnetostratigraphic framework for acritical interval of the late Miocene, which includesa dense chronologic and mammalian biostrati-graphic record across the middle – late Mioceneboundary (MN 8/ MN 9) and across the MN 9/ MN10 boundary. Ankarapithecus meteai occurs withinVallesian age horizons and is roughly similar inage, or slightly younger than the Rudabánya 2locality. Kappelman et al. (1996:92) have reportedan age of 9.74 Ma for the Locality 12 occurrence ofAnkarapithecus meteai. This age correlates closelythen with ages projected for both the CanLlobateres and Rudabánya hominoid occurrences(see also Lunka et al., 1999). The associated Sinapmammalian fauna is, like the Ouranopithecus FZ,an open country fauna thought to be of the proto-Pikermi chronofauna type. An edited volume onthe fauna, its geological and paleoecological con-texts, will elaborate further on Sinap’s significance(Fortelius et al., in press).
The hominid fossil record of Italy (L. Rook)Late Miocene paleogeography of the central
Mediterranean area had a complex history that ledto the development of endemic land mammal bio-provinces within Italy (Kotsakis et al., 1997). One ofthese, the Tusco-Sardinian paleobioprovince, is doc-umented by the late Miocene Baccinello faunal suc-cession made famous by the abundant sample of theenigmatic hominoid Oreopithecus bambolii (cf.Bernor et al., 2001). Biochronologic correlations ofthe Oreopithecus – bearing faunas has been prob-lematic because of their endemic character. Recentgeochronologic work has secured a single crystalargon date of 7.55 +/- 0.03 Ma for a horizon at theV1/V2 boundary (Rook et al., 2000). This has pro-vided a basis for a more robust estimate of theBaccinello V0-V2 (Oreopithecus-bearing) horizons asca. 9.5-6.5 Ma (Rook et al., 2000; Bernor et al., 2001).
The apparent extinction of Oreopithecus circa6.5 Ma is significantly later than that of otherEuropean Miocene hominoids, all presumablyextinct by the late Vallesian (Bernor et al., 1996;Agusti et al., 1996, 1999). Both Ouranopithecusmacedoniensis and non-cercopithecid euca-tarrhines (such as Pliopithecus) occur rarely inlocalities post-dating the mid-Vallesian crisis(Andrews & Bernor, 1999; de Bonis et al., 1999).The distribution pattern of fossil reef-building z-corals (Esteban, 1996; Rosen, 1999) would suggestthat the eastern Mediterranean (terrestrial) envi-ronments (and especially that of the northTyrrhenian) experienced warmer and possibly wet-ter conditions, and persisted longer than elsewherein Europe. Terminal Miocene paleogeographicchanges in the Tyrrhenian area were a consequenceof intensive Messinian tectonism which itself pro-duced the Apennine mountain chain, and as wellthe termination of the Tusco-Sardinian paleobio-province and, evidently, the consequent extinctionof the Oreopithecus bambolii insular fauna.
ANCESTRY OF THE AFRICAN APE/HUMAN CLADE?(L. Rook and R.L. Bernor)
European-S.W. Asian hominoid primates of thelater Miocene have a patchy distribution in timeand space, but nevertheless show considerable evo-lutionary diversity. These hominoids have a variedrepertoire of morphologies, locomotor and dietaryadaptations, and an evolutionary history covering atime span of more than 6 million years. This diver-sity has led to a variety of hypotheses concerningtheir phylogenetic relationships and adaptations(re: Andrews and Bernor, 1999). We briefly discusshere some competing hypotheses concerning theorigin of the African ape-human clade.
The European hominoid with the greatest tem-poral span and geographical range is Dryopithecus.It was a large-bodied arboreal primate with belowbranch arm swinging capability adapted for softobject frugivory (Moyá Solá & Köhler, 1996; Kay &Ungar, 1997). It is a stem great ape (hominid), butthere is disagreement on the relationship ofDryopithecus to other hominids (Andrews, 1992;Andrews et al., 1996; Andrews & Bernor, 1999).Moyá Solá & Köhler (1993) have advanced thehypothesis that Dryopithecus is a member of theSivapithecus/Pongo clade. Begun (1994, 1995,2002) and Begun & Kordos (1997) have advancedthe hypothesis that Dryopithecus is a member ofthe African ape/human clade. Köhler et al. (2001)have countered this hypothesis by arguing thatthere are no clear synapomorphies betweenDryopithecus and the African ape-human clade.According to them, Dryopithecus is then a primi-tive, generalized, hominoid.
Oreopithecus bambolii is in many aspects a verypeculiar hominoid with a typical hominoid post-cranial skeleton, a very specialized dentition, andan unusual cranial morphology. Craniodentalanatomy has always played a particularly impor-tant role in discussions of the phylogenetic rela-tionships of this fossil primate (re: Alba et al., 2001,for an exhaustive review). Oreopithecus is currentlybroadly accepted as a hominoid belonging to thegreat ape and human clade (the Hominidae s.l.)(Moyá Solá & Köhler, 1997; Harrison & Rook,1997). Oreopithecus and Dryopithecus are uniquelysimilar Eurasian apes in their postcranial anatomyand intermembral proportions. This observationhas led Moyá Solá & Köhler (1997), Harrison &Rook (1997), Andrews & Bernor (1999) and Alba etal. (2001) to suggest that they are sister taxa. Thederived craniodental and postcranial structure ofOreopithecus developed under insular conditions(Moyá Solá & Köhler, 1997; Alba et al., 2001). Thecheek tooth dentition has been found to be homo-plasious with peccaries (Alba et al., 2001), while thelower limb is structurally convergent on hominids(s.s.) in having evolved a degree of habitualbipedalism (Köhler & Moyá Solá, 1997; Rook et al.,1999). The hand, with precision grip, is also likethat of African hominids (s.s.) (Moyá Solá et al.,1999b).
The European faunal affinities of the Maremmaregion (Rook et al., 1996; Rook et al. 2000; Bernoret al. 2001) argue in favor of European origin forOreopithecus. Supporting this evidence is the fact

31RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
that: a) Dryopithecus had a very broad geographicrange (Spain to Georgia, Caucasus - Gabunia et al.,2001) and long temporal range (ca. 12 – 8 m.y.;Andrews et al., 1996; Andrews & Bernor, 1999); b)that possibly there is only an insignificant temporalgap between the last appearance of Dryopithecus inCan Llobateres (about 9.5 Ma; Agustí et al., 1996)and the first occurrence of insular faunas in theMaremma region (Bernor et al., 2001).
Ouranopithecus (= Graecopithecus of Andrews etal., 1996) is a large hominoid, the size of a femalegorilla, adapted to extreme hard object frugivory,and known only from some late Vallesian localities(MN 10) of Greece. De Bonis & Koufos (1997,1999) and de Bonis et al. (1998), have recentlyidentified a set of craniodental characters inOuranopithecus which they argue are synapomor-phies with African hominins, Australopithecus andHomo. Further they suggest a direct phylogeneticrelationship between Ouranopithecus and theAustralopithecus/Homo clade. On the contrary,Begun and Kordos (1997) have argued thatOuranopithecus is related most closely toDryopithecus. However, critical postcranials areentirely unknown in Ouranopithecus. The relation-ship between these two taxa solely on the basis ofcraniodental adaptations is still ambiguous. In fact,Dryopithecus has been established as be a softobject frugivore with below branch suspensoryadaptations, while Ouranopithecus, whose locomo-tor character is unknown, was adapted to hardobject frugivory (Andrews & Bernor, 1999).
Begun (1997) and Begun & Kordos (1997) haveargued that the Dryopithecus / Ouranopithecusclade is effectively ancestral to the African ape-human clade, and thus a Eurasian origin for thelatter. We are of the opinion that one cannot ruleout that the cranio-dental morphology ofOuranopithecus is an example of convergent evolu-tion. Both Ouranopithecus and Australopithecuswere adapted to hard-object frugivory as a conse-quence to a likely increased terrestrial adaptation(Andrews & Bernor, 1999).
The adaptive radiation of Eurasian Miocenehominoid was the consequence of one or more dis-persal events from Africa that took place some-time(s) during the middle Miocene (Andrews et al.,1996; Andrews & Bernor, 1999; Moyá Solá et al.,1999a; but see Heizmann & Begun, 2001). Therecan be no doubt that habitats differed, even strik-ingly, between Dryopithecus and penecontempora-neous and slightly younger Ankarapithecus (SinapHZ, Turkey) and slightly younger Ouranopithecus(Macedonia, Greece). The mammalian faunas ofRudabánya and Sinap are demonstrably 100% dif-ferent from one another at least at the genericlevel. The Rudabánya fauna is associated with lake-side swampy subtropical environments whereasSinap HZ, and the Ouranopithecus FZ are faunasof open country woodlands. Sinap, at the veryleast, was apparently even more open country thanthe Turolian fauna of Pikermi, Greece.
Does the available paleontological/paleoecologi-cal information support the dispersal ofDryopithecus from Europe into Africa? Do theysupport the dispersal of an Ouranopithecus-likeform into Africa? Or does the available evidence
support better an intra-African origin for theAfrican ape-human clade? Which one of these sev-eral possibilities is the more probable? The “out ofEurope” hypothesis is strongly favoured by someauthors (Begun & Kordos, 1997; Begun, 1997,2002; Stewart & Disotell, 1998) on the basis of a“hominoid vacuum” in Africa between 12.5 and 6m.y. These authors further support this hypothesiswith cladistic analyses that represent Dryopithecusand Ouranopithecus as sister-taxa of the Africanape-human clade. In fact, the macromammal fossilrecord in Africa is still relatively poor for this timeinterval. Nevertheless, the absence of hominoid evi-dence within the relatively scant African fossilrecord is not necessarily demonstrative of theirabsence (Andrews & Bernor, 1999; Moyá Solá et al.,1999a).
Dryopithecus, with its dependence on equable,subtropical/warm temperate forests, below branchlocomotion and soft-object frugivory, is not a verylikely candidate for a biogeographic extensionacross the more open country (“Pikermian”) envi-ronments that existed in southwest Asia and NorthAfrica during most of the late Miocene interval.Latest Miocene environments were probably evenmore seasonal and dry in these regions.Ouranopithecus also does not appear to be a likelycandidate for ancestry of the African ape-humanclade because its thick enamel and extreme adapta-tion to hard object frugivory is advanced comparedto an Ethiopian bipedal hominid, Ardipithecusramidus (White et al., 1994), that is known to havehad thin enamel and lived within a forested con-text. In fact, the 9 m.y. old hominoid Ourano-pithecus macedoniensis had a masticatory appara-tus that is more closely comparable functionally tomid-Pliocene - early Pleistocene hominids than isthat of Ardipithecus. There is effectively a 5+ m.y.hiatus between the craniofacial morphology ofOuranopithecus and that of the australopithecineswith which it is homoplasious.
IX) CONCLUSIONS(R. L. Bernor and L. Kordos)
Rudabánya is a very diverse MN 9, ca. 10.-9.7m.y., vertebrate fauna. It provides one of the mostdetailed “windows” into the evolution, biogeogra-phy and paleoecology of late Miocene terrestrialecosystems in Europe. It is clear that theRudabánya fauna was largely endemic to centralEurope, having some biogeographic connections towest and southwest Europe. At the same time,there was a virtually complete disjunction of theRudabánya fauna from the mammalian faunas ofthe eastern Mediterranean and southwest Asia.Rudabánya has, and will continue to stimulate agreat deal of interest from the vertebrate paleontol-ogy community because of its extraordinary fossilprimate collection. It will, of course, continue to bea pivotal fauna for testing the “out of Eurasia” ver-sus the “within Africa” hypotheses for the origin ofthe African ape-human clade. We hope that thesucceeding monographic treatment of theRudabánya fauna will shed more light on theseactively competing hypotheses.

32 R.L. BERNOR - L. KORDOS - L. ROOK
ACKNOWLEGMENTS
The Multidisiplinary Research Project at Rudabánya directedby Professors Raymond L. Bernor and Laszlo Kordos was fundedby the National Geographic Society and LSB Leakey Foundation.We gratefully acknowledge their generous support throughout thecourse of this project. Professor Lorenzo Rook offered invaluableassistance in assembling this compendium and in developing newzoogeographic and paleoecologic interpretations of theRudabánya fauna with Bernor during his stay in the Laboratoryof Evolutionary Biology, Howard University. He wishes toacknowledge support for this work by a CNR-NATO OutreachFellowship. Bernor and Kordos thank all contributors to thiscompendium, the staff of the Geological Institute of Hungary, andthe people of Rudabánya for their assistance throughout ouryears of work there. We dedicate this contribution to ProfessorMiklos Kretzoi (Budapest) and Mr. Gabor Hernyák (Rudabánya).Without their considerable contribution and commitment toRudabánya, this work could not be realized.
We suggest that whomever reads and cites this work,acknowledge the individual authors by the section they haveauthored. As an example:
SYNDLAR, S., 2003, Snakes: In Bernor R.L. et al., RecentAdvances on Multidisciplinary Research at Rudabánya, LateMiocene (MN9), Hungary: a compendium. PalaeontographiaItalica, 89: 1-34, Pisa.
LITERATURE CITED
AGUSTÍ J., GARCÉS M. & LLENAS M. (1999): Mammal turnoverand global climatic change in the late Miocene terrestrialrecord of the Vallés Penedés basin (NE Spain). In AGUSTÍ J.,ROOK L. & ANDREWS P. (eds.): The Evolution of NeogeneTerrestrial Ecosystems in Europe, 389-403, CambridgeUniversity Press.
AGUSTÍ J., KÖHLER M., MOYÁ SOLÁ S., CABRERA L., GARCÉS M. &PARÉS J.M. (1996): Can Llobateres: the pattern and timing ofthe Vallesian hominoid radiation reconsidered. J. Hum.Evol., 31, 143-155.
AGUSTÍ J., & MOYÁ-SOLÀ S. (1990): Mammal extinctions in theVallesian (Upper Miocene). Lecture Notes in Earth Science,30, 425-432.
AGUSTÍ J., & MOYÁ-SOLÀ S. (19919: Spanish Neogene Mammalsuccession and its bearing on the continental biochronology.Newslett. Stratigr., 25 (2), 91-114.
ALBA, D.M., MOYÁ SOLÁ S., KÖHLER M. & ROOK L. (2001):Heterochrony and the cranial anatomy of Oreopithecus:some cladistic fallacies and the significance of developmen-tal constraints in phylogenetic analysis. In BONIS L. DE,KOUFOS G. & ANDREWS P. (eds): Phylogeny of the NeogeneHominoid Primates of Eurasia, 284-315, CambridgeUniversity Press.
ALDANA E. (1992): Los Castoridae (Rodentia, Mammalia) delNeógeno de Cataluña (España). Trab. Mus. Geol. Barcelona,2, 99-141.
ANDREWS P. (1992): Evolution and environment in theHominoidea. Nature, 360, 641-646.
ANDREWS P. BEGUN D.R. & ZYLSTRA M. (1997): Interrelationshipsbetween functional anatomy and paleoenvironments inMiocene hominoids. In BEGUN D.R., WARD C.V. & ROSE M.D.(eds.): Function, Phylogeny and Fossils: Miocene HominoidOrigins and Adaptations, 29-58, Plenum Press, New York.
ANDREWS P. & BERNOR R.L. (1999): Vicariance biogeography andpaleoecology of Eurasian Miocene hominoid primates. InAGUSTÍ J., ROOK L. & ANDREWS P. (eds.): The Evolution ofNeogene Terrestrial Ecosystems in Europe, 445-478,Cambridge University Press.
ANDREWS P. HARRISON T., DELSON E., BERNOR R.L. & MARTIN L.(1996): Distribution and biochronology of European andSouthwest Asian Miocene Catarrhines. In BERNOR R.L.,FAHLBUSCH V. & MITTMANN H.-W. (eds): The Evolution ofWestern Eurasian Neogene Mammal Faunas, 168-295,Columbia University Press, New York.
ANONYMOUS (1967): Páratlanul értékes óslelet Rudabányán.Magyar Nemzet, 1 October 1967.
ARMOUR-CHELU M. & VIRANTA S. (2000): Carnivore modificationto Rudabánya bones. Carolinea, 58, 93-102.
AZANZA B. & MENÉNDEZ E. (1990): Los ciervos fósiles delNeógeno español. Paleont. y Evol., 23, 47-52.
BACHMAYER F. & SZYNDLAR Z. (1985): Ophidians (Reptilia:Serpentes) from the Kohfidisch fissures of Burgenland,Austria. Ann. Naturhist. Mus. Wien., 87A, 79-100.
BALLMAN P. (1969a): Die Vögel aus der altburdigalenSpaltenfullung von Wintershof (West) bei Eichstatt inBayern. Zittel, 1: 5-60.
BALLMAN P. (1969b): Les Oiseaux miocénes de la Grive-Saint-Alban (Isére). Géobios, 2, 157-204.
BALLMAN P. (1976): Fossil Voegel aus dem Neogen der HabinselGargano (Italien); zweiter Teil. Scripta Geologica, 38, 1-59.
BALOGH K. (1982): A Rudabányai-hegység problémái. Földt.Kut., 25(2), 55-60.
BALOGH K. (1984): Methods and results of K/Ar method inHungary. Unpublished Ph.D. Thesis, Kossuth University,Debrecen (Hungary).
BEGUN D.R. (1989): A Large Pliopithecine Molar from Germanyand Some Notes on the Pliopithecinae. Folia Primatologica,52, 156-166.
BEGUN D.R. (1992): Dryopithecus crusafonti sp. nov., a newMiocene hominid species fron Can Ponsic (NortheasternSpain). Am. J. Phys. Anthropol., 87, 291-310.
BEGUN D.R. (1994): Relations among the great apes andhumans: New interpretations based on the fossil great apeDryopithecus. Yarbook. Phys. Anthropol., 37, 11-63.
BEGUN D.R. (1995): Late Miocene European orang-utans, gorillas,humans, or none of the above. J. Hum. Evol., 29, 169-180.
BEGUN D.R. (1997): A Eurasian Origin of the Hominidae. Am. J.Phys. Anthropol., Supplement 24, 73-74.
BEGUN D.R. ( 2001): African and Eurasian Miocene hominoidsand the origin of Hominidae. In BONIS L. DE, KOUFOS G. &ANDREWS P. (eds): Phylogeny of the Neogene HominoidPrimates of Eurasia, 231-253, Cambridge University Press.
BEGUN D.R. (2002): European Hominoids. In HARTWIG W.G.(ed.): The Primate Fossil Record, 339-368, CambridgeUniversity Press.
BEGUN D.R. & KORDOS L. (1993): Revision of Dryopithecus bran-coi SCHLOSSER 1901 based on the fossil hominoid materialfrom Rudabánya. J. Hum. Evol., 25, 271-286.
BEGUN D.R. & KORDOS L. (1997): Phyletic affinities and func-tional convergence in Dryopithecus and other Miocene andliving hominoids. In BEGUN D.R., WARD C.V. & ROSE M.D.(eds.): Function, Phylogeny and Fossils: Miocene HominoidOrigins and Adaptations, 291-316, Plenum Press.
BERNOR R.L. (1978): The mammalian systematics, biostratigra-phy and biochronology of Maragheh and its importance forunderstanding Late Miocene Hominoid zoogeography andevolution. Ph.D. dissertation, University of California, LosAngeles, 314 pp.
BERNOR R.L. (1983): Geochronology and zoogeographic rela-tionships of Miocene Hominoidea. In CIOCHON R.L. &CORRUCCINI R.S. (eds): New interpretation of ape andHuman ancestry, 21-64, Plenum Press.
BERNOR R.L. (1984): A zoogeographic theather and biochrono-logic play: the time/biofacies phenomena of Eurasian andAfrican Miocene mammal provincies. Paléobiol. Cont., 14,121-142.
BERNOR R.L. & ARMOUR-CHELU M. (1999): Toward anEvolutionary History of African Hipparionine Horses. InBROMMAGE T. & SCHRENK F. (eds): African Biogeography,Climate Change and Early Hominid Evolution, 189-215,Oxford University Press.
BERNOR R.L., ARMOUR-CHELU M., KAISER T. & SCOTT R. (inpress): An evaluation of the late MN9 (Late Miocene,Vallesian Age): Systematic Background, FunctionalAnatomy and Paleoecology. Coloquios de Paleontología.
BERNOR R.L., FORTELIUS M. & ROOK L. (2001): EvolutionaryBiogeography and Paleoecology of the “Oreopithecus bam-bolii Faunal Zone” (late Miocene, Tusco-SardinianProvince). Boll. Soc. Paleont. Ital., 40(2), 139-148.
BERNOR R.L. & FRANZEN J. (1997): The equids (Mammalia,Perissodactyla) from the late Miocene (Early Turolian) ofDorn- Dürkheim 1 (Germany, Rheinhessen). CourierForschungsinstitut Senckenberg, 197, 117-186.
BERNOR R.L., KAISER T.M., KORDOS L. & SCOTT R.S. (1999):Stratigraphic context, systematic position and paleoecologyof Hippotherium sumegense KRETZOI, 1984 from MN 10(Late Vallesian of the Pannonian Basin). Bayer. Mitteil.Staats. Paläont. und Hist. Geol., 39, 1-35.
BERNOR R.L. & KORDOS L. (in preparation): The Late MioceneHominoid Locality of Rudabánya, Hungary. ColumbiaUniversity Press.
BERNOR R.L., KOVAR-EDER J.D., LIPSCOMB D., RÖGL F., SEN S. &

33RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
TOBIEN H. (1988): Systematics, stratigraphic and paleoenvi-ronmental contexts of first-appearing Hipparion in theVienna Basin, Austria. J. Vert. Paleont., 8(4), 427-452.
BERNOR R.L., KRETZOI M., MITTMANN H.-W. & TOBIEN H. (1993a):A preliminary systematic assessment of the Rudabánya hip-parions. Bayer. Mitteil. Staats. Paläont. und Hist. Geol., 33, 1-20.
BERNOR R.L., MITTMANN H.-W. & RÖGL F. (1993b): TheGötzendorf hipparion. Ann. Naturhistor. Mus., Wien, 95A,101-120.
BERNOR R.L., SOLOUNIAS N., SWISHER C.C. III & VAN COUVERING
J.A. (1996): The correlation of three classical “Pikermian”mammal faunas – Maragheh, Samos, and Pikermi – with theEuropean MN unit system. In BERNOR R.L., FAHLBUSCH V. &MITTMANN H.-W. (eds): The Evolution of Western EurasianNeogene Mammal Faunas, 137-154, Columbia UniversityPress.
BOCHENSZI Z. (1987): Miophasianus medius (Milne-Edwards 1869)from Przeworno (SW-Poland) and some general remarks onthe genus Miophasianus. Acta Zool. Cracow., 30, 1-80.
BOLLIGER T. (1996): A current understanding about theAnomalomyidae (Rodentia). Reflections on stratigraphy,paleobiogeography, and evolution. In BERNOR R.L.,FAHLBUSCH V. & MITTMANN H.-W. (eds): The Evolution ofWestern Eurasian Neogene Mammal Faunas, 235-245,Columbia University Press.
BONIS L. DE, BOUVRAIN G., GERAADS D. & KOUFOS G.D. (1990):New hominid skull material from the late Miocene ofMacedonia in Northern Greece. Nature, 345, 712-714.
BONIS L. DE, BOUVRAIN G. & KOUFOS G.D. (1988): Late Miocenemammal localities of the lower Axios valley (Macedonia,Greece) and their stratigraphical significance. Modern Geol.,13, 141-147.
BONIS L. DE & KOUFOS G. (1997): The phylogenetic and function-al implications of Ouranopithecus macedoniensis. In BEGUN
D.R., WARD C.V. & ROSE M.D. (eds.): Function, Phylogenyand Fossils: Miocene Hominoid Origins and Adaptations,317-326, Plenum Press.
BONIS L. DE & KOUFOS G. (1999): The Miocene land mammalsuccession in Greece. In AGUSTÍ J., ROOK L. & ANDREWS P.(eds.): The Evolution of Neogene Terrestrial Ecosystems inEurope, 197-229, Cambridge University Press.
BONIS L. DE, KOUFOS G., GUY F., PEIGNÉ S. & SILVESTROU I.(1998): Nouveausx restes du primate hominoïdeOuranopithecus dans les dépôt du Miocène supérieur demacédonie (Grèce). C.R. Acad. Sci., Paris, 327, 141-146.
BROWN W.L. & WILSON E.O. (1956): Character displacement.Sys. Zool., 5, 49-64.
BRUCH A. A., UTESCHER T., MOSBRUGGER V., GABRIELYAN I. &IVANOV D.A. (in press) Late Miocene climate in the circum-Alpine realm. A quantitative palaeobotanical analysis.Palaeogeography, Palaeoclimatology, Palaeoecology
BRUIJN H. DE, DAAMS R., DAXNER-HÖCK G., FAHLBUSCH V.,GINSBURG L,. MEIN P. & MORALES J. (1992): Report of theRCMNS working group on fossil mammals, Reisenburg,1990. Newsl. Stratigr., 26, 65-118.
BRUIJN H. DE & SARAC G. (1991): Early Miocene rodent faunasfrom the eastern Mediterranen are Part I. The genusEumyarion. Proc. Konik. Ned. Akad. V. Wetensch., 94(1), 1-36.
CAMERON D.W. (1995): The systematics of the European Miocenefaciodental Fossils ascribed to the Family Hominidae:Aspects of Anatomical Variability, Taxonomy and Phylogeny.Unpublished PhD. Dissertation, Dept. Archaeology andAnthropology, Australian National University. 790 pp.
CAMERON D.W. (1997): Sexual dimorphic features within extanthominid faciodental skeletal anatomy and testing the singlespecies hypothesis. Z. Morph. Anthropol., 81, 253-288.
CHENEVAL J. (1987): Les Anatidae (Aves, Anseriformes) duMiocéne de France. Revision systématique et évolution. Doc.Lab. Géol. Lyon, 99, 137-156.
CONROY G.C. (1987): Problems of body-weight estimation in fos-sil primates. Int. J. Primatol., 8, 115-137.
CORDS M. (1987): Forest guenons and patas monkeys: male-malecompetition in one-male groups. In SMUTS B.B., CHENEY
D.L., SEYFARTH R.M., WRANGHAM R.W. & STRUHSAKER T.T.(eds.): Primate Societies, 98-111, University of ChicagoPress.
CRUSAFONT PAIRÓ M. & KURTÉN B. (1976): Bears and bear-dogsfrom the Vallesian of the Vallés-Penedés Basin, Spain. ActaZoologica Fennica, 144, 1-29.
DAM J. A. VAN (2001) Paleoprecipitation predictions using small
mammal dietary and locomotory adaptations: new methodswith an application to the Messinian. Workshop ESFProgramme Environments and Ecosystem Dynamics of theEurasian Neogene (EEDEN). 15-17 November, Sabadell,Abstract volume.
DAMUTH J.D., FORTELIUS M., ANDREWS P., BADGLEY C., HADLEY E.A., HIXON S., JANIS C., MADDEN R. H., REED K., SMITH F. A.,THEODOR J., VAN DAM J. A., VAN VALKENBURGH B., WERDELIN
L. (2002) Reconstructing mean annual precipitation basedon mammalian dental morphology and local species rich-ness. Journal of Vertebrate Paleontology, 22 (suppl), 48A.
DAXNER- HÖCK G. (1996): Faunenwandel im Obermiozän undKorrelation der MN-”Zonen” mit den Biozonen des Pannonsder Zentralen Paratethys. Beit. zur Paläont., 21, 1-9.
ENGESSER B. (1979): Relationships of some insectivores androdents from the Miocene of North America and Europe.Bulletin of Carnagie Museum of Natural History, 14, 1-68.
ESTEBAN M. (1996): An overview of Miocene reefs fromMediterranean areas: general trends and facies models.Concepts in Sedimentology and Paleontology, 5, 3-531.
ESTES R.D. (1991): The behavior guide to African mammals.611pp., University of Cicago Press.
FAHLBUSCH V. (1985): Säugetierreste (Dorcatherium, Steneofiber)aus der miozänen Braunkohle von Wackersdorf/Oberpfalz.Bayer. Mitteil. Staats. Paläont. und Hist. Geol., 25, 81-94.
FAHLBUSCH V. & MAYR H. (1975): Microtoide Cricetiden(Mammalia, Rodentia) aus der Oberen Süßwasser-MolasseBayerns. Paläont. Zeit., 49, 78-39.
FORTELIUS M., ERONEN J., JERNVALL J., LIPING LIU, PUSHKINA D.,RINNE J., TESAKOV A., VISLOBOKOVA I., ZHAOQUN ZHANG &LIPING ZHOU (2002) Fossil mammals resolve regional pat-terns of Eurasian climate change over 20 million years.Evolutionary Ecology Research, 4, 1005-1016
FORTELIUS M., MADE J. VAN DER & BERNOR R.L. (1996) Middleand late Miocene Suoidea of Central Europe and the easternMediterranean: evolution, biogeography, and paleoecology.In BERNOR R.L., FAHLBUSCH V. & MITTMANN H.-W. (eds): TheEvolution of Western Eurasian Neogene Mammal Faunas,348-377, Columbia University Press.
FORTELIUS M., KAPPELMAN J., SEN, S. & BERNOR R.L. (in press):The Miocene Sinap Formation in central Turkey. ColumbiaUniversity Press.
FRANZEN J.L. & STORCH G. (1975): Die unterpliozäne (turolische)Wirbeltierfauna von Dorn-Dürkheim, Rheinhessen (SW-Deutschland).1. Entdeckung, Geologie, Mammalia:Carnivora, Proboscidea, Rodentia. Grabungsergebnisse1972-1973. Senck. Leth., 56(4/5), 233-303.
GABUNIA L., GABASHVILI E., VEKUA A. & LORDKIPANIDZE D. (2001):The late Miocene hominoid from Georgia. In BONIS L. DE,KOUFOS G. & ANDREWS P. (eds): Phylogeny of the NeogeneHominoid Primates of Eurasia, 316-325, CambridgeUniversity Press.
GARCÉS M., CABRERA L., AGUSTÍ J. & PARÉS J.M. (1997): OldWorld first appearence datum of “Hipparion” horses: lateMiocene large mammal dispersal and global events.Geology, 25, 19-22.
GASPARIK M. (1993): Deinotheres (Proboscidea, Mammalia) ofHungary. Annales Historico-Naturales Musei NationalisHungarici, 85, 3-17.
GAUSE G.F. (1934): The Struggle for Existence. Williams andWilkins, Baltimore.
GERVAIS F.L.P. (1848-1852): Zoologie et PaléontologieFrançaises. Bertrand, Paris.
GINSBURG L. (1964): Nouvelle découverte de Pliopitheque dansles faluns helvétiens de l`Anjou. Bull. Mus. natn. Hist. Nat.,Paris, 274D, 3345-3347.
GINSBURG L. (1986): Chronology of the European Pliopithecids.In ELSE J.G. & LEE P.C. (eds.): Primate evolution, Vol. 1, 47-57, Cambridge University Press
GINSBURG L. & MEIN P. (1980): Crouzelia rhodanica, nouvelleespece de Primate catarhinien, et essai sur la position systé-matique des Pliopithecidae. Bull. Mus. natn. Hist. Nat.,Paris, 4e sér., 1980, section C, 2, 57-85.
GRILL J. (1989): Az Aggtelek-Rudabányai-hegység szerkezetfe-jlõdése. Földt. Int. Évi Jel., 1987, 411-432.
GRILL J., KOVÁCS S., LESS GY., RÉTI ZS., RÓTH L. & SZENTPÉTERY I.(1984): Az Aggtelek-Rudabányai-hegység földtani felépítéseés fejlõdéstörténete. Földt. Kut., 27(4), 49-56.
GRINE F.E. (1985): Was interspecific competition a motive forcein early hominid evolution?. In VRBA E.S. (ed.): Species andSpeciation, 143-152, Transvaal Museum Monograph No. 4,Pretoria.

34 R.L. BERNOR - L. KORDOS - L. ROOK
HARRISON T. (1987): The phylogenetic relationships of the earlycatarrhine primates: A review of the current evidence. J.Hum. Evol., 16, 41-80.
HARRISON T., DELSON E. & GUAN JIAN (1991): A new species ofPliopithecus from the middle Miocene of China and itsimplications for early catarrhine zoogeography. J. Hum.Evol., 21, 329-361.
HARRISON T. & ROOK L. (1997): Enigmatic anthropoid or misun-derstood ape: the phylogenetic status of Oreopithecus bam-bolii reconsidered. In BEGUN D.R., WARD C.V. & ROSE M.D.(eds.): Function, Phylogeny and Fossils: Miocene HominoidOrigins and Adaptations, 327-362, Plenum Press.
HEIZMANN E.P.J. (1976): Die palaeontologische Erforschung desSteinheimer Beckens. In Meteorkrater Steinheimer Becken,Steinheim an Albuch, 29-45, Bürgermeisteramt.
HEIZMANN E.P.J. & BEGUN D. (2001): The oldest Eurasian homi-noid J. Hum. Evol., 41, 463-481.
HELLMUND M. (1995): Suidae (Artiodactyla, Mammalia). Münch.Geowiss., Abh. 28, 143-156.
HILL A. & WARD S. (1988): Origin of the Hominidae: the recordof African large hominoid evolution between 14 my and 4my. Yearbook Phys. Anthropol., 32, 48-83.
JANIS C. M., DAMUTH J. & THEODOR J. M. (2000) Miocene ungu-lates and terrestrial primary productivity: Where have allthe browsers gone? Proceedings of the National Academy ofScience, USA, 97, 7899-7904.
JANIS C. M., DAMUTH J. & THEODOR J. M. (2002) The origins andevolution of the North American grassland biome: the storyfrom the hoofed mammals. Palaeogeography, Palaeo-climatology, Palaeoecology, 177, 183-198.
JÁNOSSY D. (1976): Plio-Pleistocene bird remains from theCarpathian Basin. II. Galliformes, Phasianidae. Aquila, 83,29-42.
JÁNOSSY D. (1977): Plio-Pleistocene bird remains from theCarpathian Basin. III. Strigiformes, Falconiformes,Caprimulgiformes, Apodiformes. Aquila, 84, 9-36.
JÁNOSSY D. (1991): Late Miocene bird remains from Polgárdi(W-Hungary). Aquila, 89, 13-35.
JUNGERS W.L. (1988): New estimates of body size in australop-ithecines. In GRINE F. (ed.): Evolutionary History of the“Robust” Australopithecines, 115-125, Aldine de Gruyter,New York.
KAY R.F. (1978): Molar structure and diet in extantCercopithecidae. In BUTLER P.M. & JOYSEY K.A. (eds.):Development, Function, and Evolution of Teeth, 309-339,Academic Press, New York.
KAY R.F. & UNGAR P.S. (1997): Dental evidence for diet in someMiocene catarrhines with comments on the effects of phy-logeny on the interpretation of adaptation. In BEGUN D.R.,WARD C.V. & ROSE M.D. (eds.): Function, Phylogeny andFossils: Miocene Hominoid Origins and Adaptations, 131-151, Plenum Press, New York.
KESSLER E. (1984): On some bird remains from the Pliocene ofMalusteni in the Laboratory of Paleontology-University ofBucharest. 75 Years Laboratory Paleontology, Spec. Vol., 287-292.
KINGDON J. (1982): East African Mammals, Vol.III, C (Bovids),393 pp., Academic Press, London.
KÖHLER M. (1993): Skeleton and habitat of Recent and fossilruminants. Münch. Geowiss., Abh. A, 25, 1-88.
KÖHLER M. & MOYÁ SOLÁ S. (1997): Ape-like or hominid-like?The positional behaviour of Oreopithecus bambolii reconsid-ered. Proc. Natl. Acad. Sci. USA, 94, 11747-11750.
KÖHLER M., MOYÁ SOLÁ S. & ALBA D.M. (2001): Eursian homi-noid evolution in the light of recent Dryopithecus findings.In BONIS L. DE, KOUFOS G. & ANDREWS P. (eds): Phylogeny ofthe Neogene Hominoid Primates of Eurasia, 192-212,Cambridge University Press.
KÖHLER M., MOYÁ SOLÁ S. & ANDREWS, P. (1999): OrderPrimates. In RÖSSNER G.E. & HEISSIG K. (eds.): The MioceneLand Mammals of Europe, 91-104, Verlag Dr. FriedrichPfeil, Munchen.
KÖPPEN W. (1931) Grundriß der Klimakunde. DeGruyter, Berlin,Leipzig.
KORDOS L. (1982): The prehominid locality of Rudabánya (NEHungary) and its neighbourhood: A palaeogeographicreconstruction. Mag. All. Foldt. Intez. Evi. Jel., 1980, 395-406.
KORDOS L. (1985): Az els_ ötvemillió év. _sök Rudabányán.Gondolat, Budapest.
KORDOS L. (1987a): Description and reconstruction of the skull
of Rudapithecushungaricus KRETZOI (Mammalia). Annls.Hist. Nat. Mus. natn. Hung., 79, 77-88.
KORDOS L. (1987b): Neogene Vertebrate Biostratigraphy inHungary. Ann. Inst. Geol. Publ. Hung., 70, 393-396.
KORDOS L. (1989): Anomalomyidae (Mammalia, Rodentia)remains from the Neogene of Hungary. Földt. Int. Évi Jel.,1987, 293-311.
KORDOS L. & BEGUN D.R. (1997): A New Reconstruction of RUD77, a Partial Cranium of Dryopithecus brancoi fromRudabánya, Hungary. Am. J. Phys. Anthropol., 103, 277-294.
KORDOS L. & BEGUN D.R. (1999): Femora of Anapithecus fromRudabánya. Am. J. Phys. Anthropol., 108(28), 173.
KOTSAKIS T., BARISONE G. & ROOK L. (1997): Mammalianbiochronology in an insular domain: the Italian Tertiary fau-nas. Mém. Trav. E.P.H.E. Inst. Montpellier, 21, 431-441.
KOUFOS G.D. (1993): A mandible of Ouranopithecus macedonien-sis (Hominidae, Primates) from the late Miocene ofMacedonia. Am. J. Phys. Anthropol., 91: 225-234.
KOUFOS G.D. (1995) The late Miocene percrocutas of Macedonia(Greece). Palaeovert., 24, 67-84.
KOUFOS G.D. (2000): The hipparions of the late Miocene locality“Nikiti 1”, Chalkidiki, Macedonia, Greece. RevuePalaeobiologique, 19(1), 47-77.
KOUFOS G.D., SYRIDES G.E., KOLIADIMOUS K.K. & KOSTOPOULOS
D.S. (1991): Un nouveau gisement de Vertébrés avec homi-noide dans le Miocene supérieur de Macédoine (Grèce). C.R. Acad. Sci. Paris, ser. II, 313, 691-696.
KOWALSKI K. (1993): Microtocricetus molassicus FAHLBUSCH &MAYR 1975 (Rodentia, Mammalia) from the Miocene ofBelchatòv (Poland). Act zooL. Cracov., 36, 251-258.
KOWALSKI K. (1994): Evolution of Anomalomys GAILLARD 1900(Rodentia, Mammalia) in the Miocene of Poland. Acta Zool.Cracov., 37, 163-176.
KRETZOI M. (1969): Geschichte der Primaten und hominisation.Symp. Biol. Hung., 9: 3-11.
KRETZOI M. (1974): Towards Hominization. AntropológiaiKözlemények, 18, 119-126.
KRETZOI M. (1975): New ramapithecines and Pliopithecus fromthe lower Pliocene of Rudabánya in north-eastern Hungary.Nature, 257, 578-581.
KRETZOI M. (1976). Die Hominisation und dieAustralopithecinen. Anthrop.Közl., 20, 3-11.
KRETZOI M. (1984): New Hominoid Form from Rudabánya.Anthrop. Közl., 28, 1-96.
KRETZOI M., KROLOPP E., LÓRINCZ H. & PÁLFALVY I. (1976): Flora,fauna und stratigraphische lage der unterpannonischenPrahominiden-fundstelle von Rudabánya (NO-Ungarn).Földt. Int. Évi Jel., 1974, 365-394.
LAMBRECHT K. (1933): Handbuch der Palaornithologie, 124 pp.,Borntrager, Berlin.
LEAKEY M.G., FEIBEL C.S., BERNOR R.L. HARRIS J.M., CERLING T.E.,STEWART K.M., STORRS G.W., WALKER A., WERDELIN L. &WINKLER A.J. (1996): Lothagam: a record of faunal change inthe Late Miocene of East Africa. J. Vert. Paleont., 16, 556-570.
LESS G. (1998): Földtani felépítés. In BAROSS G. (ed.): Szerk. “AzAggteleki Nemzeti Park”, 26-66, Mezõgazda Kiadó,Budapest.
LUNKKA J.P., FORTELIUS M., KAPPELMAN J. & SEN S. (1999):Chronology and mammalian faunas of the Miocene SinapFormation, Turkey. In AGUSTÍ J., ROOK L. & ANDREWS P.(eds.): The Evolution of Neogene Terrestrial Ecosystems inEurope, 238-264, Cambridge University Press.
MAGYAR I., GEARY D. H. & MÜLLER P. (1999): Paleogeographicevolution of the Late Miocene Lake Pannon in CentralEurope. Palaeogeography, Palaeoclimatology, Palaeoecology,147, 151-167.
MEIN P. (1970): Les Sciuropteres (Mammalia, Rodentia)néogènes d´Europe occidentale. Géobios, 3, 7-77.
MEIN P. (1986): Chronological succession of hominoids in theEuropean Neogene. In ELSE J.G. & LEE P.C. (eds.): PrimateEvolution, 59-70, Cambridge University Press.
MEIN P. (1990): Updating of the MN zones. In LINDSAY E.H.FAHLBUSCH V. & MEIN P. (eds.): European Neogene MammalChronology, 73-90, Plenum Press, New York.
MILNE-EDWARDS A. (1867): Recherches anatomiques et paléon-tologiques pour servir á l`histoire des oiseaux fossiles de laFrance. Vol. I, 474pp., Masson, Paris.
MILNE-EDWARDS A. (1871): Recherches anatomiques et paléon-tologiques pour servir á l`histoire des oiseaux fossiles de laFrance. Vol. II, 632pp., Masson-Paris.
MORALES J. & SORIA D. (1981): Los artiodactilos de Los Valles deFuentiduena (Segovia). Estud. Geol., 37, 477-501.

35RECENT ADVANCES ON MULTIDISCIPLINARY RESEARCH AT RUDABÁNYA
MOSBRUGGER V. & UTESCHER T. (1997) The coexistence approach- a method for quantitative reconstructions of Tertiary ter-restrial palaeoclimate data using plant fossils. Palaeo-geography, Palaeoclimatology, Palaeoecology, 134, 61-86.
MOYÁ-SOLÀ S. & KÖHLER M. (1993): Recent discoveries ofDryopithecus shed new light on evolution of great apes.Nature, 365, 543-545.
MOYÁ-SOLÀ S. & KÖHLER M. (1996): A Dryopithecus skeleton andthe origins of great-ape locomotion. Nature, 379, 156-159.
MOYÁ-SOLÀ S. & KÖHLER M. (1997): The phylogenetic relation-ships of Oreopithecus bambolii GERVAIS, 1872. C. R. Acad.Sci., Paris, ser. II, 324, 141-148.
MOYÁ-SOLÀ S. & KÖHLER M. & ALBA D.M. (1999a): Primate evo-lution - in or out of Africa (comments). Curr. Biol., 9(15),547-548.
MOYÁ-SOLÀ S. & KÖHLER M. & ROOK L. (1999b): Evidence ofhominid-like precision grip capabilities in the hand of theEuropean Miocene ape Oreopithecus. Proc. Natl. Acad. Sci.USA, 96, 313-317.
MOYÁ-SOLÀ S., PONS MOYÁ J. & KÖHLER M. (1990): Primatescatarrinos (Mammalia) del Neogeno de la peninsula Iberica.Paleont. y Evol., 23, 41-45.
PANTÓ G. (1956): A rudabányai vasércvonulat földtani felépítése(Constitutuóion géologique de la chaîne de mineral de fer deRudabánya). A Magyar Állami Földtani Intézet Évkönyve,44(1), 330-490.
PETTER G. (1963): Contribution à l’Étude des mustélidés desbassins néogènes du Vallès-Pénédès et de Calatayud-Teruel.Mém. Soc. Géol. France, N.S., 97, 1-44.
PETTER G. (1967): Mustélidés nouveaux du Vallésien deCatalogne. Ann. Paléont., 53, 93-113.
RABEDER G. (1985): Die Saugetiere des Pannonien:Chronostratigraphie und Neostratotypen. M6, Pannonien,440-463.
RAFFERTY K.L., WALKER A., RUFF C.B., ROSE M.D. & ANDREWS P.J.(1995): Postcranial estimates of body weight in Proconsul,with a note on the distal tibia of P. major from Napak,Uganda. Am. J. Phys. Anthropol., 97, 391-402.
RÖGL F., ZAPFE H., BERNOR R.L., BRZOBOHATY H., DAXNER-HÖCK
G., DRAXLER I., FEJFAR O., GAUDANT J., HERMANN P., RABEDER
G., SCHULTZ O. & ZETTLER R. (1993): Die PrimatenfundstelleGötzendorf an der Leitha (Obermiozän des Wiener Beckens,Niederösterreich). Jharbuch Geologische B. undesanstalt,Wien, A. 136(2), 503-526.
RÖGL F. & DAXNER-HÖCK G. (1996): Late Miocene Paratethyscorrelations. In BERNOR R.L., FAHLBUSCH V. & MITTMANN H.-W. (eds): The Evolution of Western Eurasian NeogeneMammal Faunas, 47-55, Columbia University Press, NewYork.
ROOK L., BONDIOLI L., KÖHLER M., MOYÁ SOLÁ S. & MACCHIARELLI
R. (1999): Oreopithecus was a bipedal ape after all: evidencefrom the iliac cancellous architecture. Proc. Natl. Acad. Sci.USA, 96, 8795-8799.
ROOK L., HARRISON T. & ENGESSER B. (1996): The taxonomic sta-tus and biochronological implications of new finds ofOreopithecus from Baccinello (Tuscany, Italy). J. Hum. Evol.,30, 3-27.
ROOK L., RENNE P., BENVENUTI M. & PAPINI M. (2000):Geochronology of Oreopithecus - bearing succession atBaccinello (Italy) and the extinction pattern of EuropeanMiocene hominoids. J. Hum. Evol., 39, 577-582
ROSEN B. (1999): Palaeoclimatic implications of the energyhypothesis from Neogene corals of the Mediterraneanregion. In AGUSTÍ J., ROOK L. & ANDREWS P. (eds.): TheEvolution of Neogene Terrestrial Ecosystems in Europe,301-319, Cambridge University Press.
SCHAUB S. & ZAPFE H. (1953): Die Fauna der miozanenSpaltenfüllung von Neudorf an der March (CSR). Öst. Akad.Wiss. math.-naturw., Xl., 162(3), 181-215.
SCHRÉTER Z. (1929): A borsod-hevesi szén- és lignitterületekbányaföldtani leírása. Földt. Int. Alk. Kiadv., Budapest.
SEN S. (1996): Late Miocene Hystricidae in Europe andAnatolia. In BERNOR R.L., FAHLBUSCH V. & MITTMANN H.-W.(eds): The Evolution of Western Eurasian Neogene MammalFaunas, 264-265, Columbia University Press, New York.
SEN S., KOUFOS G.D., KONDOPOULOU D. & BONIS L. DE (2000):Magnetostratigraphy of the late Miocene continentaldeposits of the lower Axios valley, Macedonia, Greece. InKOUFOS G.D. & IOAKIM E. (eds): Mediterranean Neogenecyclostratigraphy in marine-continental deposits, Bull. Geol.Soc. Greece, special publication, 9, 197-206.
SMITH R.J. (1985): The present as a key to the past: Body weight
of Miocene hominoids as a test of allometric methods forpalaeontological inference. In JUNGERS W.L. (ed.): Size andScaling in Primate Biology, 437-448, Plenum Press, NewYork.
SOLOUNIAS N., PLAVCAN J.M., QUADE J. & WITMER L. (1999): Thepaleoecology of the Pikermian biome and the savanna mythIn AGUSTÍ J., ROOK L. & ANDREWS P. (eds.): The Evolution ofNeogene Terrestrial Ecosystems in Europe, 436-453.Cambridge University Press.
STEININGER F.F. (1986): Dating the Paratethys Miocene homi-noid record. In ELSE J.G. & LEE P.C. (eds.): PrimateEvolution, 71-84, Cambridge University Press.
STEININGER F.F., BERNOR R.L. & FAHLBUSCH V. (1990): EuropeanNeogene marine/continental chronologic correlations InLINDSAY E.H. FAHLBUSCH V. & MEIN P. (eds.): EuropeanNeogene Mammal Chronology, 15-46, Plenum Press, NewYork.
STEININGER F.F., BERGGREN W.A., KENT D.V., BERNOR R.L., SEN S.& AGUSTÍ J. (1996): Circum-Mediterranean Neogene(Miocene and Pliocene) marine-continental chronologic cor-relations of European mammal units. In BERNOR R.L.,FAHLBUSCH V. & MITTMANN H.-W. (eds): The Evolution ofWestern Eurasian Neogene Mammal Faunas, 7-46,Columbia University Press.
STEWART C.B. & DISOTELL T.R. (1998): Primate evolution - in andout of Africa. Curr. Biol., 8(16), 582-588.
SZALAY F. S. & DELSON E. (1979): Evolutionary History of thePrimates. Academic Press, New York.
SZENTPÉTERY I. (1988): A Rudabányai-hegység és környezeténekoligocén, alsó-miocén képzõdményei. Földt. Int. Évi Jel.,1986, 121-128.
SZYNDLAR Z. (1991a): A review of Neogene and Quaternarysnakes of Central and Eastern Europe. Part I.Scolecophidia, Boidae, Colubrinae. Estud. Geol., 47(1-2),103-126.
SZYNDLAR Z. (1991b): A review of Neogene and Quaternarysnakes of Central and Eastern Europe. Part II. Natricinae,Elapidae, Viperidae. Estud. Geol., 47(3-4), 237-266.
SZYNDLAR Z. & RAGE J.C. (1990): West Palearctic cobras of thegenus Naja (Serpentes: Elapidae): interrelationships amongextinct and extant species. Amphibia-Reptilia, 11(4), 385-400.
SZYNDLAR Z. & ZEROVA G.A. (1990): Neogene cobras of the genusNaja (Serpentes: Elapidae) of East Europe. Ann. Natur. Mus.Wien, 91A, 53-61.
SZYNDLAR Z. & ZEROVA G.A. (1992): Miocene snake fauna fromCherevichnoie (Ukraine, USSR), with description of a newspecies of Vipera. Neu. Jahrb. Geol. Paläo., Abh. 184(1), 87-99.
TASNÁDI-KUBACSKA A. (1967): A Rudapithecus hungaricus: A rud-abányai ósmajom. Élet és Tudomány, 22, 2083-2085.
TEAFORD M.F. (1988): Scanning electron microscope diagnosisof wear patterns versus artifacts on fossil teeth. Scan.Microsc., 2, 1167-1175.
TEAFORD M.F. (1988): A review of dental microwear and diet inmodern mammals. Scan. Microsc., 2, 1149-1166.
TERBORGH J. (1983): Five New World Primates: A Study inComparative Ecology. Princeton University Press.
THENIUS E. (1950): Die tertiären Lagomeryciden und Cervidender Steiermark. Sitz. Oesterr. Akad. Wiss. Math.-Naturwiss.Kl., Wien, I, 159, 219-254.
TOBIEN H. (1986): Wissenschaftliches Bericht über dieMastodonten aus der obermiozänen FossilfundstelleRudabánya (NE-Ungarn). unpublished manuscript.
TOPACHEVSKI V.A. & SCORIK A.F. (1992): Neogene and PleistoceneCricetidae of early evolutionary stage of south easternEurope. Acad. Sci. Ukr. Inst. Zoo., 1992, 1-242.
TOPÁL G. (1989a): Tertiary and Early Quaternary remains ofCorynorhinus and Plecotus from Hungary (Mammalia,Chiroptera). Vertebrata Hungarica, 23, 33-55.
TOPÁL G. (1989b): New Tertiary plecotines from Hungary(Mammalia, Chiroptera). In HANÁK V., HORACEK I. & GAISLER
J. (eds.): European Bat Research, 77-86, Charles UniversityPress, Praha.
UNGAR P.S. (1996): Dental microwear of European Miocenecatarrhines: evidence for diets and tooth use. J. Hum. Evol.,31, 335-366.
UNGAR P.S. & KAY R.F. (1995): The dietary adaptations ofEuropean Miocene catarrhines. Proc. Natl. Acad. Sci. USA,92, 5479-5481.
VENCZEL M. (1994): Late Miocene snakes from Polgárdi(Hungary). Acta Zool.Cracov., 37(1), 1-29.

36 R.L. BERNOR - L. KORDOS - L. ROOK
VIRANTA S. (1996): European Miocene Amphicyonidae: taxono-my, systematics and ecology. Acta Zool. Fenn., 204, 1-61.
WELCOMME J.-L., AGUILAR J.-P. & GINSBURG L. (1991): Discoveryof a new Pliopithecus (Primates, Mammalia) associated withrodents in the Upper Miocene Priay Sands (Ain, France),and remarks on the paleogeography of the Bresse area dur-ing the Vallesian. C.R. Acad. Sci. Paris, 313, Sér II, 723-729.
WERDELIN L. (1996): Carnivores, exclusive of Hyaenidae, fromthe later Miocene of Europe and western Asia. In BERNOR
R.L., FAHLBUSCH V. & MITTMANN H.-W. (eds): The Evolutionof Western Eurasian Neogene Mammal Faunas, 271-289,Columbia University Press.
WERDELIN L. & SOLOUNIAS N. (1996): The evolutionary history ofhyenas in Europe and western Asia during the Miocene. InBERNOR R.L., FAHLBUSCH V. & MITTMANN H.-W. (eds): TheEvolution of Western Eurasian Neogene Mammal Faunas,290-306, Columbia University Press.
WHITE T.D., SUWA G. & ASFAW B. (1994): Australopithecusramidus, a new species of early hominid from Aramis,Ethiopia. Nature, 371, 306-312.
WOODBURNE M.O., BERNOR R.L. & SWISHER C.S. III (1996): Anappraisal of the stratigraphic and phylogenetic bases for the“Hipparion” Datum in the Old World In BERNOR R.L.,FAHLBUSCH V. & MITTMANN H.-W. (eds): The Evolution ofWestern Eurasian Neogene Mammal Faunas, 124-136,Columbia University Press.
WRIGHT R.V.S. (1994): MV-NUTSHELL: Dept. Anthropology,University of Sydney.
ZAPFE H. (1979): Chalicotherium grande aus der MiozänenSpaltenfüllung von Neudorf an der March (D_vinská NováVes), Tscheschoslowakei. Neu. Denk. Naturhist. Mus., Wien.,156, 1-282.
ZAPFE H. (1989a): Pongidenzahne (Primates) aus dem Pontienvon Götzendorf, Niederösterreich. Sitzungsberichte ÖsterrAkad. Wissensch., 197, 423-450.
ZAPFE H. (1989b): Chalicotherium goldfussi KAUP aus demVallesium vom Höwenegg im Hegau (Südwestdeutschland).Andrias, 6, 117-128.
Related Documents











